
Age-associated changes in lymphoid and antigen-presenting cell functions in mice immunized with...
15
Mechanisms of Ageing and Development 118 (2000) 71–85 Age-related alterations in inflammatory response during experimental autoimmune prostatitis Gabriel Moro ´ n, Belkys Maletto, Miguel Orsilles, Mirtha Depiante-Depaoli, Marı ´a Cristina Pistoresi-Palencia * Departamento de Bioquı ´mica Clı ´nica, Facultad de Ciencias Quı ´micas, Uni6ersidad Nacional de Co ´rdoba., 5000 Co ´rdoba, Argentina Received 28 April 2000; accepted 5 July 2000 Abstract Experimental autoimmune prostatitis (EAP) is an experimental model of autoimmune disease, developed in Wistar rats against prostatic components. The 12-and 18-month-old rats with EAP show a higher cellular autoimmune response and lower humoral autoimmune response compared to 3-month-old rats. The analysis of NO · and O 2 - production by peritoneal exudate cells (PECs) resulted in a higher NO · and O 2 - production in EAP rats at all ages, compared to control animals. PECs from 12- and 18-month-old rats produced more NO · and less O 2 - than PECs from 3-month-old rats. However, lipopolysacharide (LPS) did not stimulate PECs from aged rats for NO · production as much as in 3-month-old rats and thus, turning out in a lower index of LPS-stimulation of PECs from aged rats, compared to 3-month-old rats. Furthermore, the mast cells number in prostates of EAP rats, especially the number of degranulated cells, was higher than in control animals, but no significant differences were found between 3- and 12-month-old control rats. In conclusion, these results show that aging affects differentially the inflammation mediators during EAP. © 2000 Published by Elsevier Science Ireland Ltd. Keywords: Aging; Mast cells; Nitric oxide; Prostatitis; Superoxide anion www.elsevier.com/locate/mechagedev * Corresponding author. Tel.: +54-3514334174; fax: +54-3514334187. E-mail address: [email protected] (M.C. Pistoresi-Palencia). 0047-6374/00/$ - see front matter © 2000 Published by Elsevier Science Ireland Ltd. PII:S0047-6374(00)00159-7
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Age-associated changes in lymphoid and antigen-presenting cell functions in mice immunized with...

Mechanisms of Ageing and Development
118 (2000) 71–85
Age-related alterations in inflammatoryresponse during experimental autoimmune
prostatitis
Gabriel Moron, Belkys Maletto, Miguel Orsilles,Mirtha Depiante-Depaoli, Marıa Cristina Pistoresi-Palencia *
Departamento de Bioquımica Clınica, Facultad de Ciencias Quımicas,Uni6ersidad Nacional de Cordoba., 5000 Cordoba, Argentina
Received 28 April 2000; accepted 5 July 2000
Abstract
Experimental autoimmune prostatitis (EAP) is an experimental model of autoimmunedisease, developed in Wistar rats against prostatic components. The 12-and 18-month-oldrats with EAP show a higher cellular autoimmune response and lower humoral autoimmuneresponse compared to 3-month-old rats. The analysis of NO· and O2
− production byperitoneal exudate cells (PECs) resulted in a higher NO· and O2
− production in EAP rats atall ages, compared to control animals. PECs from 12- and 18-month-old rats produced moreNO· and less O2
− than PECs from 3-month-old rats. However, lipopolysacharide (LPS) didnot stimulate PECs from aged rats for NO· production as much as in 3-month-old rats andthus, turning out in a lower index of LPS-stimulation of PECs from aged rats, compared to3-month-old rats. Furthermore, the mast cells number in prostates of EAP rats, especiallythe number of degranulated cells, was higher than in control animals, but no significantdifferences were found between 3- and 12-month-old control rats. In conclusion, these resultsshow that aging affects differentially the inflammation mediators during EAP. © 2000Published by Elsevier Science Ireland Ltd.
Keywords: Aging; Mast cells; Nitric oxide; Prostatitis; Superoxide anion
www.elsevier.com/locate/mechagedev
* Corresponding author. Tel.: +54-3514334174; fax: +54-3514334187.E-mail address: [email protected] (M.C. Pistoresi-Palencia).
0047-6374/00/$ - see front matter © 2000 Published by Elsevier Science Ireland Ltd.
PII: S0047 -6374 (00 )00159 -7

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8572
1. Introduction
Experimental autoimmune prostatitis (EAP) is an experimental model of autoim-mune disease developed in Wistar rats against male sex accessory glands (RAG,which in the rat consist of prostate, seminal vesicles and coagulating glands). Thisdisease is induced by i.d. injection with a chemically modified extract of RAG,(MRAG) in complete Freund’s adjuvant (CFA) (Pacheco-Rupil et al., 1981;Galmarini et al., 1988). In 3-month-old rats, EAP is characterized by the presenceof specific cellular and humoral response against RAG and mononuclear cellsinfiltration in the prostate. Besides, the disease can be passively induced in normalsyngeneic rats by adoptive transfer of spleen T cells from immune rats and it canbe specifically suppressed with low doses of the RAG autoantigens (Depiante-De-paoli et al., 1984; Ferro et al., 1990).
One of the most important age-related manifestations in both humans andlaboratory animals is a gradual decline in immune effectiveness. All the changesreported during aging have been associated with an increased incidence of infec-tions, malignant diseases and autoimmune disorders (Burns and Goodwin, 1997).In our study, the word aging has been used as a biological phenomenon observedin an individual organism from the time of fertilization to death. The function ofthe immune system peaks at around puberty and gradually declines thereafter withadvancing age. The ability of both T and B cells to respond to various forms ofstimulus change with increasing age. However, T cell functions seem to be moresensitive to the process of aging than any other type of immune cells (Pawelec et al.,1999).
In previous studies we have found that 12-month-old rats, showing alteredimmune parameters corresponding to aged animals, have a higher cellular autoim-mune response and lower humoral autoimmune response to RAG than 3-month-old rats (Moron et al., 1998). In the peritoneal cavity of 12-month-old rats, we havedescribed a deficit of peritoneal cells expressing MHC class II-IE+ molecules.These cells have been involved in the induction of specific suppression of theautoimmune response in EAP. Since in these animals it is more difficult to inducesuppression of the autoimmune response EAP (Pistoresi-Palencia et al., 1991, 1994),it shows that antigen presenting cells, which are found in peritoneal cells, areplaying a role in the development of the higher autoimmune response. Besides itsrole in the induction and modulation of the immune response, the macrophageshave many effector functions, such as the production of reactive oxygen intermedi-ates (ROI) (Babior, 1984) and reactive nitrogen intermediates (RNI) (Xie andNathan, 1994). One of the most studied ROI in aged animals is the superoxideanion (O2
−). It has been found that the macrophages of aged rats show an impairedO2
− production (Hayakawa et al., 1995; Alvarez et al., 1996), which is translatedinto a diminished resistance to infection (Edwards et al., 1992). RNI include nitricoxide (NO·), and the stable decomposition products: nitrite (NO2
−) and nitrate(NO3
−) (Iyengar et al., 1987). Controversial reports have shown that macrophagesfrom old animals have either decreased (Alvarez et al., 1996; Kissin et al., 1997) orincreased (Chen et al., 1996) NO· secretion. We have previously shown that

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 73
peritoneal exudate cells (PECs) from 3-month-old rats with EAP released signifi-cantly more ROI and RNI than cells from control rats (Orsilles et al., 1993; Orsillesand Depiante-Depaoli, 1995).
During an inflammatory process, included autoimmune diseases, the participa-tion of mast cell is a transcendental event (Dines and Powell, 1997; Metcalfe et al.,1997). It has been observed that 3-month-old rats immunized with RAG incorpo-rated into liposomes (developing also EAP) presented mast cell infiltration inprostate (Rivero et al., 1995). However, there is almost no information whethermast cells are affected by aging.
In this study, we studied the autoimmune response of 18-month-old rats andcompared O2
− and NO· release in 3-, 12- and 18-month-old rats with EAP, tocorrelate the effector capacities of peritoneal cells during the autoimmune responsein aged rats. Furthermore, we carry out a quantitative study about the number andactivation status of mast cells during EAP and during aging.
2. Materials and methods
2.1. Animals and antigen
Three, 12- and 18-month-old male Wistar rats were used. A saline extract ofRAG was prepared as described in Moron et al., 1998, MRAG was prepared bycoupling the RAG saline extract with diazonium derivatives of arsanilic andsulfanilic acid following an earlier procedure (Galmarini et al., 1988). All thereagents were purchased from Sigma Chemical Co., St. Louis, USA, unlessotherwise specified.
2.2. Experimental design
Groups of 3-, 12- and 18-month-old rats were i.d. isoimmunized with 5 mg/0.5 mlof MRAG in CFA. Control animals of each age were divided in two groups: onegroup was injected with CFA and the other group remained without treatment(control). On day 20 all groups were challenged with RAG for DTH, and bloodwas obtained from tail vein puncture for measurement of anti RAG antibody levels.On day 21, the peritoneal cavity of the animals was opened and the peritonealexudate cells (PECs) were collected with 20 to 30 ml of phenol red free-Hank’sbalanced salt solution (HBSS). The cell suspension was washed twice in HBSS anddiluted to obtain an appropriate concentration. PECs suspensions were essentiallymacrophages (90%), as determined by examination of cells with May Grunwald–Giemsa stain and non-specific esterase stain. At the same day, prostates of 3- and12-month-old rats were obtained and fixed in 4% p-formaldehyde, aclared in xylol,embedded in wax paraffin and sections (6 mm thick) were stained with ToluidineBlue for mast cell count.
All these experiments were performed twice and a representative result of eachexperiment is hereby depicted.

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8574
2.3. Assays
2.3.1. DTH responseFootpad testing was accomplished by injecting 0.1 mg of RAG dissolved in 0.1
ml of 0.15 M NaCl into the right footpad, and an equal volume of 0.15 M NaClinto the left footpad. Swelling was observed and measured with a micrometer 24 hafter injection, and the difference of swelling was calculated. A difference betweenboth footpads of 4×10−1 mm or higher was considered to be a positive footpad test.
2.3.2. Antibody assaysThe measure of specific IgG autoantibodies to RAG by ELISA test was
performed as described (Moron et al., 1998). Briefly, polystyrene plates (Nunc A/S,Rocklide, Denmark) were coated with RAG (10 mg/well) in carbonate buffer, pH9.6. After blocking with gelatin-PBS and later incubation with the plasma samples,plates were incubated with peroxidase-labeled mouse anti-rat IgG antibody. Plateswere read on a Bio Rad model 450 microplate reader at 492 nm after incubationwith H2O2 and o-phenylendiamine. The mean of absorbance of duplicates was usedfor statistical analysis.
2.3.3. Assay of NO · productionPECs (1×106/ml) were incubated for 24 h at 37°C 5% CO2 in HBSS medium
containing L-arginine (1 mM) with or without lipopolysacharide (LPS, 500 ng/ml).Simultaneous experiments were performed with cells incubated in medium containingthe arginine analogous, L-NG-monomethyl arginine (L-NMMA, 1mM). After 24 hof incubation at 37°C in 5% CO2 and humidified conditions, the supernatants wereremoved and analyzed for NO. measurement. Nitric oxide production was deter-mined on cell-free supernatant through the measuring of NO2
− concentration by aspectrophotometric assay based on the Griess reaction. Briefly, 1 ml aliquots weremixed with an equal volume of Griess reagent (1% sulfanilamide, 0.1% naphtylendi-amine dihydrochloride, 2% H3PO4) and incubated at room temperature for 10 min.The absorbance at 546 nm was measured spectrophotometrically. The NO2
−
concentration was calculated from a standard curve and expressed as mM/106 cellsfrom determinations corrected for the background levels of NO2
− in the mediumwithout L-arginine.
The stimulation index of NO2− production by PECs in the presence of LPS was
calculated, for each animal, according to:
Production of NO in presence of LPSProduction of NO without LPS
2.3.4. Assay for O2− production
Superoxide anion was measured by the reduction of ferricytochrome c21 withslight modifications. PECs (1×106 cells) were mixed with 1.5 ml of 80 mMferricytochrome c solution (type III) in phenol red free-HBSS with or withoutstimulant (PMA, 3 mg) and incubated for 90 min at 37°C. Control tubes containing

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 75
cytochrome c solution and PMA, but not PECs, were included. After completion ofthe incubation, the reaction was stopped by returning the tubes to an ice-bath andthe contents of the tubes were promptly centrifuged at 800×g for 10 min. Thespecificity of cytochrome reduction was controlled by inclusion in the same samplesof 100 units/ml of superoxide dismutase (SOD) (type I, 3000 units/mg). The opticaldensity of supernatants was measured at 550 nm against controls consisting ofcytochrome c solution from tubes without PECs. The O2
− level was determined bycalculating the difference between the samples without SOD and those with SODand the concentration of cytochrome c reduced by using the formula E550 nm=2.1×104 M−1 cm−1. The O2
− release was expressed as nmoles of O2−/106 cells/min.
2.3.5. Mast cell countMast cells were evaluated in prostate of control and MRAG/CFA immunized
animals by counting out on sections of lateral and ventral prostatic lobules stainedwith Toluidin Blue. Resting and activated mast cells were differentiated by theabsence or presence, respectively, of metachromatic granules outside the mast cellsin the surrounding connective tissue. The counting was expressed as mast cellsnumber/mm2. A reticulated ocular lens and a 40X objective lens were employed tomeasure the area of observed tissue. Each field of counting corresponded to an areaof 0.0576 mm2. At least 20 and 40 fields of lateral and ventral prostatic lobules,respectively, and two to three different sections of prostate of each animal wereevaluated. All counting was performed by one of the authors (G. M.) in a maskedfashion.
2.4. Statistical analysis
The results are expressed as mean9standard deviation of the mean (SD) or asmean9standard error of the mean (SEM), as indicated. In all experiments signifi-cant differences between groups were calculated using non-parametric Mann–Whit-ney U test.
3. Results
3.1. Autoimmune response in 18-month-old rats
Cellular immune response against RAG was tested by DTH in 3-, 12- and18-month-old rats on day 20 after immunization. The DTH response on day 21after immunization was higher in 12- and 18-month-old rats than in 3-month-oldrats, but there were no differences between 12- and 18-month-old rats (Fig. 1A).
The determination of plasmatic levels of IgG against RAG was performed byELISA in plasma of 3-, 12- and 18-month-old rats on day 21 after immunization.The levels of specific IgG against RAG in 18-month-old rats were significantlylower than in 3- and 12-month-old rats. Although the levels of IgG anti RAG werediminished in 12-month-old rats compared to 3-month-old rats, no statisticallysignificant differences were found (Fig. 1B).

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8576
3.2. NO · production
The capacity of macrophages to secrete NO· was accomplished using peritonealexudate cells, which provide an appropriate number of macrophages for the study,without need for further purification steps that could lead to anomalousmacrophage activation. The results are shown in Fig. 2. After 24 h of culture, PECs
Fig. 1. Autoimmune response in aged rats. (A) DTH response against RAG. The 3-, 12- and18-month-old rats were injected with CFA or immunized with MRAG/CFA. Twenty days afterimmunization, the DTH assay was performed. The results are expressed as mean9SD. *, PB0.001versus 3-month-old rats. (B) Plasmatic levels of IgG specific against RAG: 3-, 12- and 18- month-old ratswere injected with CFA or immunized with MRAG/CFA. Twenty one days after immunization, theELISA assay was performed. The results are expressed as mean9SD. *, PB0.02 versus 3-month-oldrats and **, PB0.03 versus 12-month-old rats.
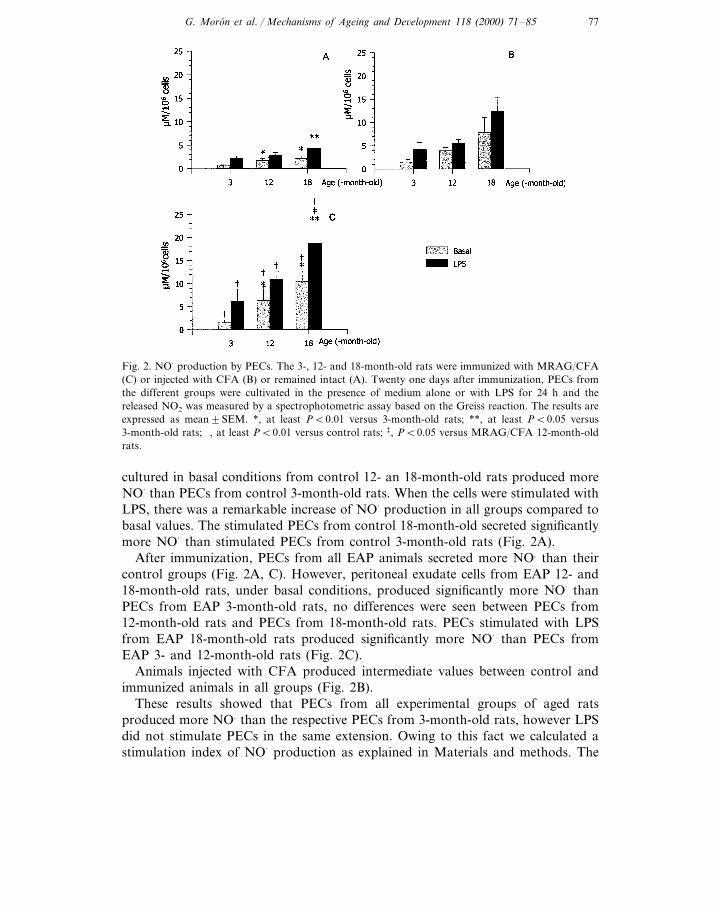
G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 77
Fig. 2. NO· production by PECs. The 3-, 12- and 18-month-old rats were immunized with MRAG/CFA(C) or injected with CFA (B) or remained intact (A). Twenty one days after immunization, PECs fromthe different groups were cultivated in the presence of medium alone or with LPS for 24 h and thereleased NO2 was measured by a spectrophotometric assay based on the Greiss reaction. The results areexpressed as mean9SEM. *, at least PB0.01 versus 3-month-old rats; **, at least PB0.05 versus3-month-old rats; †, at least PB0.01 versus control rats; ‡, PB0.05 versus MRAG/CFA 12-month-oldrats.
cultured in basal conditions from control 12- an 18-month-old rats produced moreNO· than PECs from control 3-month-old rats. When the cells were stimulated withLPS, there was a remarkable increase of NO· production in all groups compared tobasal values. The stimulated PECs from control 18-month-old secreted significantlymore NO· than stimulated PECs from control 3-month-old rats (Fig. 2A).
After immunization, PECs from all EAP animals secreted more NO. than theircontrol groups (Fig. 2A, C). However, peritoneal exudate cells from EAP 12- and18-month-old rats, under basal conditions, produced significantly more NO· thanPECs from EAP 3-month-old rats, no differences were seen between PECs from12-month-old rats and PECs from 18-month-old rats. PECs stimulated with LPSfrom EAP 18-month-old rats produced significantly more NO· than PECs fromEAP 3- and 12-month-old rats (Fig. 2C).
Animals injected with CFA produced intermediate values between control andimmunized animals in all groups (Fig. 2B).
These results showed that PECs from all experimental groups of aged ratsproduced more NO. than the respective PECs from 3-month-old rats, however LPSdid not stimulate PECs in the same extension. Owing to this fact we calculated astimulation index of NO· production as explained in Materials and methods. The

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8578
calculation revealed that NO· production stimulation indexes of PECs from 12- and18-month-old rats were lower than indexes of PECs from 3-month-old rats in allexperimental groups (Table 1).
3.3. Production of O2−
PECs from control 12- and 18-month-old rats had a lower O2− production
compared to PECs from control 3-month-old rats, in both basal and PMAstimulated conditions (Fig. 3A). Immunization with MRAG/CFA results in asignificant increase in O2
− releasing in PECs of all ages, in both basal and PMAstimulated conditions; however, PECs of 12- and 18-month-old rats producedsignificantly less O2
− than PECs of 3-month-old rats (Fig. 3C).Animals injected with CFA produced intermediate values between control and
immunized animals in all ages (Fig. 3B).When a stimulation index of O2
− was calculated, no statistically differences wereobserved (data not shown).
3.4. Mast cell infiltration in prostate
Another component of inflammatory processes is the mast cell. To evaluate itspossible participation in the autoimmune response during EAP and whether itcould be affected by aging, the mast cells number was counted in sections of thelobules of prostate of 3- and 12-month-old rats stained with Toluidine blue on day21 after immunization with MRAG. The results are shown in Table 2 and Table 3.We found that mast cells in lateral and ventral prostatic lobules are present in theconnective tissue of prostate from control animals mostly as resting cells (approxi-mately 80% of total mast cells number). In control animals of both ages, the mastcells total number was significantly higher in the ventral lobule than in the lateralone (PB0.05). After immunization, the mast cells number incremented remarkably,especially the number of degranulated cells, thus approximately 50–60% of mastcells were considered activated in both lobules of 3- and 12-month-old rats.However, no statistically significative differences were seen between 3- and 12-month-old control rats or between 3- and 12-month-old immunized rats, in any ofboth prostatic lobules.
Table 1Stimulation index of NO− production by PECs by LPSa
MRAG/CFAControl CFA
3-month-old 5.191.43.891.33.390.51.890.3*b12-month-old 1.590.1**b 1.790.2**b
1.890.3***b1.990.52.490.4*b18-month-old
a Control, animals without treatment; CFA, animals injected with CFA; MRAG/CFA, animalsimmunized with MRAG/CFA. The determination of NO· was performed 21 days after immunization.
b The values are expressed as mean9SEM. *, PB0.03; **, PB0.01 and ***, PB0.02 versus3-month-old animals of their respective groups.

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 79
Tab
le2
Mas
tce
llsnu
mbe
rin
vent
ral
lobu
leof
pros
tate
ofco
ntro
lan
dM
RA
G/C
FA
imm
uniz
ed3-
and
12-m
onth
-old
rats
a
MR
AG
/CF
AC
ontr
ol
Mas
tce
lls/m
m2
Mas
tce
lls/m
m2
Not
degr
anul
ated
bD
egra
nula
tedc
Tot
ald
Not
%D
egra
nula
tede
Deg
ranu
late
dcT
otal
d%
Deg
ranu
late
de
degr
anul
ated
b
10.99
5.3*
f21
.79
6.9*
f489
14*f
10.29
2.3
1.79
0.1
179
410
.89
2.5
3m
8.59
311
.59
3.7*
*f27
.29
5.0*
*f429
7**f
12m
9.89
82.
69
0.8
12.49
2.5
259
1015
.69
2.5*
*f
aC
ontr
ol,
anim
als
wit
hout
trea
tmen
t;M
RA
G/C
FA
,an
imal
sim
mun
ized
wit
hM
RA
G/C
FA
.T
hem
easu
reof
pros
tati
car
eaw
aspe
rfor
med
inse
ctio
nsof
pros
tate
stai
ned
wit
hT
olui
dine
Blu
e,us
ing
are
ticu
late
ocul
arle
ns(a
rea
mea
sure
dw
ith
a40
Xob
ject
ive
lens
=0.
0576
mm
2).
bN
otde
gran
ulat
edm
ast
cells
.c
Deg
ranu
late
dm
ast
cells
.d
Deg
ranu
late
dan
dno
tde
gran
ulat
edm
ast
cells
.e
%of
degr
anul
ated
=pe
rcen
tage
ofde
gran
ulat
edm
ast
cells
wit
hre
gard
toth
em
ast
cells
tota
lnu
mbe
r.fT
here
sult
sar
eex
pres
sed
asm
ean9
SD(n
=fo
urto
six
anim
als
per
grou
p).
*,PB
0.02
vers
usco
ntro
l3-
mon
th-o
ldra
ts**
,PB
0.03
vers
usco
ntro
l12
-mon
th-o
ldra
ts.

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8580
Tab
le3
Mas
tce
llnu
mbe
rin
late
ral
lobu
leof
pros
tate
ofco
ntro
lan
dM
RA
G/C
FA
imm
uniz
ed3-
and
12-m
onth
-old
rats
a
MR
AG
/CF
AC
ontr
ol
Mas
tce
lls/m
m2
Mas
tce
lls/m
m2
Not
degr
anul
ated
bD
egra
nula
tedc
Tot
ald
Not
%D
egra
nula
tede
Deg
ranu
late
dcT
otal
d%
Deg
ranu
late
de
degr
anul
ated
b
13.59
6.9*
f23
.29
11.7
*f599
13*f
3.09
1.6
0.99
0.6
199
99.
79
7.1*
f3
m2.
69
1.0
7.49
4.8*
*f16
.29
10.9
**f
519
11**
f12
m4.
59
3.5
1.19
1.3
5.79
4.6
239
148.
69
7.7
aC
ontr
ol,
anim
als
wit
hout
trea
tmen
t;M
RA
G/C
FA
,an
imal
sim
mun
ized
wit
hM
RA
G/C
FA
.T
hem
easu
reof
pros
tati
car
eaw
aspe
rfor
med
inse
ctio
nsof
pros
tate
stai
ned
wit
hT
olui
dine
Blu
e,us
ing
are
ticu
late
ocul
arle
ns(a
rea
mea
sure
dw
ith
a40
Xob
ject
ive
lens
=0.
0576
mm
2).
bN
otde
gran
ulat
edm
ast
cells
.c
Deg
ranu
late
dm
ast
cells
.d
Deg
ranu
late
dan
dno
tde
gran
ulat
edm
ast
cells
.e
%of
degr
anul
ated
=pe
rcen
tage
ofde
gran
ulat
edm
ast
cells
wit
hre
gard
toth
em
ast
cells
tota
lnu
mbe
r.fT
here
sult
sar
eex
pres
sed
asm
ean9
SD(n
=fo
urto
six
anim
als
per
grou
p).*
,PB
0.01
vers
usco
ntro
l3-m
onth
-old
rats
**,a
tle
ast
PB
0.02
vers
usco
ntro
l12
-mon
th-o
ldra
ts.

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 81
Fig. 3. Superoxide anion production by PECs. The 3-, 12- and 18-month-old rats were immunized withMRAG/CFA (C) or injected with CFA (B) or remained intact (A). Twenty one days after immunization,PECs from the different groups were cultivated in the presence of medium alone or with PMA for 1 hand O2
− production was measured by the reduction of ferricytochrome c21. The results are expressed asmean9SEM. *, at least PB0.05 versus 3-month-old rats; **, at least PB0.01 versus 3-month-old rats;†, at least PB0.01 versus control rats; ‡, PB0.05 versus MRAG/CFA 12-month-old rats.
4. Discussion
Aging is accompanied by changes in the behavior of immune system. In thiswork, we want to extend our previous findings, studying the autoimmune responseagainst RAG in 18-month-old animals and how aging affects the ability of immunesystem to trigger an inflammatory response during EAP. For the first objective wemeasured the DTH response and the IgG levels against prostatic components andfor the second one, we quantified the nitric oxide and superoxide anion generation(NO· and O2
−, respectively) and the capacity of mast cells to invade prostate as wellas their activation levels in aged rats.
The values of DTH and antibodies that we have previously observed altered inmiddle aged rats, was seen even more affected in 18-month-old rats, showing anincreased cellular and a diminisshed humoral autoimmune response.
PECs from control, CFA and EAP 12- and 18-month-old rats produced moreNO· and less O2
− than PECs of respectives 3-month-old rats. Nevertheless, despite

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8582
PECs of all groups were sensitive to LPS stimulus, the action of LPS did not hasthe same extension at all ages. PECs of 12- and 18-month-old rats were lessstimulated by LPS than those from 3-month-old rats. This could has to twopossible reasons: either PECs of aged rats reached their maximum activity withLPS, and/or NO. secretion without LPS stimulation is already enhanced in agedanimals, so the relationships between activated/basal NO· levels resulted dimin-ished. The same phenomenon was not observed with O2
− generation because thecapacity of PECs to be stimulated by PMA seems not to be affected by aging andtherefore remained highly sensitive to PMA.
At the same time that immunization of Wistar rats with MRAG/CFA resulted inDTH response and autoantibodies against RAG (Pacheco-Rupil et al., 1981;Galmarini et al., 1988; Moron et al., 1998) there was an activation of peritonealmacrophages with a raise of NO· and O2
− generation in young and aged rats. Nitricoxide and O2
− secretion have been related to damage mechanisms during EAP(Orsilles and Depiante-Depaoli, 1995, 1998) as well as in other models of autoim-mune disease such as insulin-dependent diabetes, murine SLE in MRL-lpr/lpr mice,rheumatoid arthritis, etc. (Nomikos et al., 1998; McCartney-Francis et al., 1993;Weinberg et al., 1994). However, now is under discussion the possible roles of NO·
production by immune cells; some authors have proposed that NO· wouled has andimmunoregulatory role rather than dangerous role in some autoimmune diseases(Bogdan, 1998). Therefore, the role of NO· in EAP, specially in old rats, could bealso associated to an attempt of immune cells to control the development of EAP.
The generation of both metabolites is dependent on the activation level ofmacrophages, mediated by the action of pro-inflammatory cytokines, such as INFg
and TNFa (Hayakawa et al., 1995). These cytokines are involved in the shift ofimmune response to Th1 response, characterized by the presence of DTH response,production of IgG2b and IgG2c (Mosmann and Sad, 1996), activation of cytotoxiccells and development of inflammatory infiltration. All these parameters have beenobserved during EAP (Moron et al., 1998; Casas-Ingaramo et al., 1991), suggestingthat autoimmune response during EAP is biased to Th1-like response and it isexacerbated in aged rats (Moron et al., 1998; Maccioni et al., 1996). Furthermore,in mice, NO· secretion has been associated with a Th1-like response (Munder et al.,1998). Therefore, the increased values of NO· in animals with EAP seem to confirmthe fact that EAP is a disease driven by Th1 cells.
However, O2− shows an age-related diminution. Superoxide anion reacts with
NO· to form peroxynitrite anion and, therefore, NO· is capable of outcompetingsuperoxide dismutase for O2
−. This catabolic pathway of O2− destruction as well as
defects in the superoxide dismutase/catalase pathway could be possible explanationsto these results, associated with possible age-related alterations in the synthesissteps of O2
− as depletion of NADPH levels and impairments in its productionpathway (Alvarez et al., 1996).
Another participant of the inflammatory response is the mast cell. The remark-able diversity of mediators derived from mast cells can convert these cells in verysignificant participants of inflammatory responses, such as autoimmune diseases.Thirty days after immunization (at the same time when the infiltration of mononu-

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 83
clear cells in prostate reached its maximum (Moron et al., 1998)), it was observeda significant increment of the number of mast cells, especially of degranulated mastcells (between 5 to 15 folds increment), which represented around 50% of total mastcells. As we established previously (Moron et al., 1998), the main site of mononuclearinfiltration is the lateral lobule of prostate. In this work, we have found that althoughin control animals the number of mast cells in ventral lobule is significantly higherthan in lateral lobule at both ages, the number of mast cells in both prostatic lobulesof immunized 3- and 12-month-old rats was almost the same, suggesting that mastcells are particularly engaged in the inflammatory response in the lateral lobule, witha strong correlation with the infiltration of lymphocytes and macrophages. Mast cellswould facilitate, through degranulation and releasing of their mediators, the arrivaland maintenance of infiltrating cells in prostate. However, no statistically significantdifferences in the number and percentages of degranulation of mast cells wereobserved between 3- and 12-month-old rats, in either control or immunized groups.There is very scarce bibliography regarding mast cell during aging. Majeed, 1994, hasfound that in mice and rats there are no variations in the number of mast cell withage. Some years ago, Czuwaj et al., 1992 reported that releasing of histamine by mastcell from aged rats is diminished, but there are no further reports on this issue. Thepresent study suggests that, as in other models of autoimmune disease (Dines andPowell, 1997; Metcalfe et al., 1997), mast cells can play an important role in therecruiting of mononuclear cells toward the prostate, in particular to the lateral lobule,as well as in the capacity of promoting injury. Because of the capacity of mast cellsto invade the prostate and get activated seems not to decay with aging, mast cellscould be particularly important in aged rats.
In conclusion, according to the intermediate analyzed, the way that aged ratsrespond during EAP is different from the way that 3-month-old rats do. Nitric oxidereleasing as well as mast cell infiltration are more relevant than O2
− releasing in agedrats during EAP. These results indicate that the aging does not affect the inflamma-tion mechanisms of immune system in the same extension and direction.
Acknowledgements
This work was supported by grants from Consejo Nacional de InvestigacionesCientıficas y Tecnicas (CONICET), Consejo de Investigaciones Cientıficas y Tecno-logicas de la Provincia de Cordoba (CONICOR) and Secretarıa de Ciencia y Tecnicade la Universidad Nacional de Cordoba (SECyT). G. M. is a recipient of a fellowshipof CONICET. M.C. P.P. is a career member of CONICET. M. D.D. has recentlydied; this work is dedicated to her memory.
References
Alvarez, A., Machado, A., Sobrino, F., Santa Marıa, C., 1996. Nitric oxide and superoxide anionproduction decrease with age in resident and activated rat peritoneal macrophages. Cell Immunol.169, 152–155.

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–8584
Babior, B.M., 1984. Oxidants from phagocytes: agents of defense and destruction. Blood 64, 959–966.Bogdan, C.H., 1998. The multiplex function of nitric oxide in (auto)immunity. J. Exp. Med. 187,
1361–1365.Burns, E.A., Goodwin, J.S., 1997. Immunodeficiency of ageing. Drugs Aging 11, 374–397.Casas-Ingaramo, A., Depiante-Depaoli, M., Pacheco-Rupil, B., 1991. Activation of cytotoxic cells by
syngeneic prostate antigens in experimental autoimmune vesiculo-prostatitis. Autoimmunity 9,151–157.
Czuwaj, M., Bartosz, G., Wyczolkowska, J., 1992. Anaphylactic histamine release and membranefluidity of mast cells from young and aged rats. Agents Actions, Special Conference Issue,C283–C286.
Chen, L.C., Pace, J.L., Russell, S.W., Morrison, D.C., 1996. Altered regulation of inducible nitric oxidesynthase expression in macrophages from senescent mice. Infect. Immun. 64, 4288–4298.
Depiante-Depaoli, M., Pacheco-Rupil, B., Britos, S., Casas, A., 1984. Experimental autoimmunedamage to rat males accessory glands: I: transfer of autoimmune response by spleen cells. Am. J.Reprod. Immunol. 5, 9–11.
Dines, K.C., Powell, C.H., 1997. Mast cell interactions with the nervous system: relationship tomechanisms of disease. J. Neuropathol. Exp. Neurol. 56, 627–640.
Edwards III, C.K., Ghiasuddin, S.M., Yunger, L.M., et al., 1992. In vivo administration of recombinantgrowth hormone or gamma interferon activates macrophages: enhanced resistance to experimentalSalmonella typhimurium infection is correlated with generation of reactive oxygen intermediates.Infect. Immun. 60, 2514–2515.
Ferro, M.E., Romero-Piffiguer, M., Riera, C.M., 1990. Antigen-induced inhibition to rat male accessoryglands: distinct role of I-A and I-E positive peritoneal cells. Eur. J. Immunol. 20, 1007–1012.
Galmarini, M., Ferro, M.E., Riera, C.M., 1988. Delayed hypersensitivity and lesions following isoimmu-nization with modified rat male accessory glands: kinetics of induction. J. Reprod. Immunol. 13,147–157.
Hayakawa, H., Sato, A., Yagi, T., Uchiyama, H., Ide, K., Nakano, M., 1995. Superoxide generation byalveolar macrophages from aged rats: improvent by in vitro treatment with INFg. Mech. AgeingDev. 80, 199–211.
Iyengar, R., Stuehr, D.J., Marletta, M.A., 1987. Macrophages synthesis of nitrite, nitrate and N-ni-trosamines: precursors and role of the respiratory burst. Proc. Natl. Acad. Sci. USA 84, 6369–6373.
Kissin, E., Tomasi, M., McCartney-Francis, N., Gibbs, C.L., Smith, P.D., 1997. Age-related decline inmurine macrophage production of nitric oxide. J. Infect. Dis. 175, 1004–1007.
Maccioni, M., Rivero, V., Riera, C.M., 1996. Autoantibodies against rat prostate antigens. Associationof specific IgG2b and IgG2c with the DTH response. J. Autoimmun. 9, 485–491.
Majeed, S.K., 1994. Mast cell distribution in rats. Arzneimittelforschung 44, 370–374.McCartney-Francis, N., Allen, J.B., Mizel, D.E., et al., 1993. Suppression of arthritis by an inhibitor of
nitric oxide synthase. J. Exp. Med. 178, 749–754.Metcalfe, D.D., Baram, D., Mekori, Y.A., 1997. Mast cells. Physiol. Rev. 77, 1033–1079.Moron, G., Maletto, B., Ropolo, A., Pistoresi-Palencia, M.C., 1998. Effect of aging on experimental
autoimmune prostatitis: differential kinetics of development. Clin. Immunol. Immunopathol. 87,256–265.
Mosmann, T.R., Sad, S., 1996. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol.Today 17, 138–146.
Munder, M., Eichmann, K., Modolell, M., 1998. Alternative metabolic states in murine macrophagesreflected by the nitric oxide synthase/arginase balance: competitive regulation by CD4+ T cellscorrelates with Th1/Th2 phenotype. J. Immunol. 160, 5354–5374.
Nomikos, I.N., Wang, Y., Lafferty, K.J., 1998. Involvement of O2 radicals in ‘autoimmune’ diabetes.Immunol. Cell Biol. 67, 85–87.
Orsilles, M.A., Pacheco-Rupil, B.N., Depiante-Depaoli, M., 1993. Experimental autoimmune prostatitis(EAP): enhanced release of reactive oxygen intermediates (ROI) in peritoneal macrophages. Autoim-munity 16, 201–207.

G. Moron et al. / Mechanisms of Ageing and De6elopment 118 (2000) 71–85 85
Orsilles, M.A., Depiante-Depaoli, M., 1995. Production of reactive nitrogen intermediates (RNI) byperitoneal macrophages from rats with experimental autoimmune prostatitis (EAP). Inflammation19, 415–422.
Orsilles, M.A., Depiante-Depaoli, M., 1998. Oxidative stress-related parameters in prostate of rats withexperimental autoimmune prostatitis. Prostate 34, 270–274.
Pacheco-Rupil, B., Depiante-Depaoli, M., Romero, M., Romero, O., Yantorno, C., 1981. Experimentalautoimmune damage to rat male accessory glands (MAG). Am. J. Reprod. Immunol. 1, 255–261.
Pawelec, G., Effros, R.B., Caruso, C., Remarque, E., Barnett, Y., Solana, R., 1999. T cells and aging.Front Biosci. 4, d216–d269.
Pistoresi-Palencia, M.C., Romero-Piffiguer, M., Ferro, M.E., 1991. Effect of aging on the autoimmuneresponse to rat male accesory glands: deficit of I-E+ peritoneal cells capable of inducing suppres-sion. Eur. J. Immunol. 21, 1685–1689.
Pistoresi-Palencia, M.C., Romero-Piffiguer, M., Moron, G., Ferro, M.E., 1994. Effect of aging onautoimmune response to rat male accesory glands: young, but not aged, antigen presenting cellsefficiently induce suppression in aged rats. Mech. Ageing Dev. 76, 33–41.
Rivero, V.E., Iribarren, P., Riera, C.M., 1995. Mast cells in accessory glands of experimentally inducedprostatitis in male rats. Clin. Immunol. Immunopathol. 74, 236–242.
Weinberg, J.B., Granger, D.L., Pisetsky, D.S., et al., 1994. The role of nitric oxide in the pathogenesisof murine autoimmune disease: increased nitric oxide production and nitric oxide synthase expressionin MRL-lpr/lpr mice, and reduction of spontaneus glomerulonephritis and arthritis by orallyadministered NG-monomethyl-L-Arginine. J. Exp. Med. 179, 651–660.
Xie, Q., Nathan, C., 1994. The high-output nitric oxide pathway: role and regulation. J. Leuk. Biol. 56,576–582.
.




















