
Activity of antioxidant enzymes in response to cadmium in Crotalaria juncea
10
Plant and Soil 239: 123–132, 2002. © 2002 Kluwer Academic Publishers. Printed in the Netherlands. 123 Activity of antioxidant enzymes in response to cadmium in Crotalaria juncea G.J.G. Pereira 1 , S.M.G. Molina 2 , P.J. Lea 3 & R.A. Azevedo 2,4 1 Departamento de Gen´ etica e Evolução, Universidade Estadual de Campinas, Campinas CEP 13083-970, SP, Brasil; 2 Departamento de Gen´ etica, Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Av. P ´ adua Dias, 11, Piracicaba CEP 13400-970, SP, Brasil; 3 Department of Biological Sciences, Lancaster University, Lancaster LA1 4YQ, UK; 4 Corresponding author ∗ Received 23 May 2001. Accepted in revised form 13 November 2001 Key words: cadmium, catalase, Crotalaria juncea, glutathione reductase, heavy metals, superoxide dismutase Abstract The effects of the heavy metal Cadmium (Cd) on the growth and the activities of the antioxidant enzymes, catalase (CAT, EC 1.11.1.6), superoxide dismutase (SOD, EC 1.15.1.1) and glutathione reductase (GR, EC 1.6.4.2) have been investigated in Crotalaria juncea seedlings. Concentrations above 0.2 mM CdCl 2 were shown to inhibit strongly the growth of roots and shoots. Cd was shown to accumulate to very high concentrations in the roots, whilst in the leaves, the maximum concentration obtained following treatment with 2 mM CdCl 2 , was only 6% of that determined in the roots. Although CAT activity did not exhibit any major variation in the roots following CdCl 2 treatment, 2 mM CdCl 2 induced a 6-fold increase in activity in the leaves when compared to the untreated control. Non-denaturing PAGE gels stained for SOD activity revealed four isoenzymes, two Mn-SOD and two Cu/Zn- SOD. The results observed for SOD were different of those observed for CAT activity, since in both, leaves and roots, no significant changes in the total activity or of the four isoenzymes were observed following the treatment with CdCl 2 . GR activity exhibited a similar pattern of that of CAT activity. The concentration of 2 mM CdCl 2 induced a small increase in activity in the roots after 48 h of exposure, whereas in leaves a 7-fold increase in GR activity was detected after 48 hr exposure to 2 mM CdCl 2 . The results suggest that in C. juncea the reactive oxygen species (ROS) induced by Cd, are metabolised by CAT in the peroxisomes. In the case of GR activity, the increase observed in the leaves suggest that GR is also playing a role in the detoxification of Cd-induced ROS possibly via the glutathione-ascorbate cycle. Abbreviations: Cd – cadmium; CAT – catalase; GR – glutathione reductase; ROS – reactive oxygen species; SOD – superoxide dismutase Introduction The heavy metal cadmium (Cd) is not an essential nu- trient for plants and it is normally toxic, with the clear exception of certain hyperaccumulator plant species (Schickler and Caspi, 1999). Cd can accumulate in the top soil in areas of agricultural land which have been subjected to sewage sludge application (Lombi et al., 2000) and the concentrations can be very high (100–6000 mg Kg −1 dry weight) in areas subjected ∗ FAX No: 19-3433-6706. E-mail: [email protected] to mining (Lombi et al., 2000). Within the cell sys- tem, the concentration of Cd can be decreased, thus avoiding direct damage to the plant tissue by a group of peptides termed phytochelatins, which may bind Cd and other heavy metals (Bruns et al., 2001; Grill et al., 1987). Phytochelatins are structurally related to glutathione and the sulphur atom of cysteine is able to bind Cd (Harada et al., 2001; Höfgen et al., 2001). In general, plant growth and the net photosynthetic rate in the presence of Cd have been shown to be sig- nificantly reduced, even at low concentrations of the metal (Boussama et al., 1999a; Khudsar et al., 2001),
-
Upload
kurniatransport -
Category
Documents
-
view
0 -
download
0
Transcript of Activity of antioxidant enzymes in response to cadmium in Crotalaria juncea

Plant and Soil 239: 123–132, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
123
Activity of antioxidant enzymes in response to cadmium in Crotalariajuncea
G.J.G. Pereira1, S.M.G. Molina2, P.J. Lea3 & R.A. Azevedo2,4
1Departamento de Genetica e Evolução, Universidade Estadual de Campinas, Campinas CEP 13083-970, SP,Brasil; 2Departamento de Genetica, Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo,Av. Padua Dias, 11, Piracicaba CEP 13400-970, SP, Brasil; 3Department of Biological Sciences, LancasterUniversity, Lancaster LA1 4YQ, UK; 4Corresponding author∗
Received 23 May 2001. Accepted in revised form 13 November 2001
Key words: cadmium, catalase, Crotalaria juncea, glutathione reductase, heavy metals, superoxide dismutase
Abstract
The effects of the heavy metal Cadmium (Cd) on the growth and the activities of the antioxidant enzymes, catalase(CAT, EC 1.11.1.6), superoxide dismutase (SOD, EC 1.15.1.1) and glutathione reductase (GR, EC 1.6.4.2) havebeen investigated in Crotalaria juncea seedlings. Concentrations above 0.2 mM CdCl2 were shown to inhibitstrongly the growth of roots and shoots. Cd was shown to accumulate to very high concentrations in the roots,whilst in the leaves, the maximum concentration obtained following treatment with 2 mM CdCl2, was only 6% ofthat determined in the roots. Although CAT activity did not exhibit any major variation in the roots following CdCl2treatment, 2 mM CdCl2 induced a 6-fold increase in activity in the leaves when compared to the untreated control.Non-denaturing PAGE gels stained for SOD activity revealed four isoenzymes, two Mn-SOD and two Cu/Zn-SOD. The results observed for SOD were different of those observed for CAT activity, since in both, leaves androots, no significant changes in the total activity or of the four isoenzymes were observed following the treatmentwith CdCl2. GR activity exhibited a similar pattern of that of CAT activity. The concentration of 2 mM CdCl2induced a small increase in activity in the roots after 48 h of exposure, whereas in leaves a 7-fold increase in GRactivity was detected after 48 hr exposure to 2 mM CdCl2. The results suggest that in C. juncea the reactive oxygenspecies (ROS) induced by Cd, are metabolised by CAT in the peroxisomes. In the case of GR activity, the increaseobserved in the leaves suggest that GR is also playing a role in the detoxification of Cd-induced ROS possibly viathe glutathione-ascorbate cycle.Abbreviations: Cd – cadmium; CAT – catalase; GR – glutathione reductase; ROS – reactive oxygen species; SOD– superoxide dismutase
Introduction
The heavy metal cadmium (Cd) is not an essential nu-trient for plants and it is normally toxic, with the clearexception of certain hyperaccumulator plant species(Schickler and Caspi, 1999). Cd can accumulate inthe top soil in areas of agricultural land which havebeen subjected to sewage sludge application (Lombiet al., 2000) and the concentrations can be very high(100–6000 mg Kg−1 dry weight) in areas subjected
∗ FAX No: 19-3433-6706.E-mail: [email protected]
to mining (Lombi et al., 2000). Within the cell sys-tem, the concentration of Cd can be decreased, thusavoiding direct damage to the plant tissue by a groupof peptides termed phytochelatins, which may bindCd and other heavy metals (Bruns et al., 2001; Grillet al., 1987). Phytochelatins are structurally related toglutathione and the sulphur atom of cysteine is able tobind Cd (Harada et al., 2001; Höfgen et al., 2001).In general, plant growth and the net photosyntheticrate in the presence of Cd have been shown to be sig-nificantly reduced, even at low concentrations of themetal (Boussama et al., 1999a; Khudsar et al., 2001),

124
whereas leaf senescence is accelerated (Siedlecka andKrupa, 1999). Moreover, Cd can also inhibit the activ-ity of several groups of enzymes such as those ofthe photosynthetic Calvin cycle (Stevens et al., 1997),nitrogen metabolism (Boussama et al., 1999a,b; Ku-mar and Dubey, 1999), sugar metabolism (Verma andDubey, 2001), and sulphate assimilation (Lee andLeustek, 1999). The concentrations of betaine, putres-cine and spermine have been shown to be differentlyaffected by growth of plants in high concentrations ofCd and other heavy metals, suggesting a protectionagainst metal-induced oxidative stress by polyamines(Bergmann et al., 2001; Groppa et al., 2001). Cdhas also been shown to induce lipid peroxidation andchlorophyll breakdown in plants (Somashekaraiah etal., 1992), indicating the production of reactive oxy-gen species (ROS). Further evidence of ROS gener-ation induced by Cd has been obtained through thedetection of new isoenzymes of peroxidases (Tahlil etal., 1999; Van Assche and Clijsters, 1990).
ROS such as superoxide (O2−), hydroxyl radicals
(OH.) and hydrogen peroxide (H2O2) are producednaturally during cell metabolism in processes such asphotosynthesis, photorespiration (Foyer and Noctor,2000), fatty acid oxidation and senescence (Vitóriaet al., 2001). ROS can also be induced by externalstresses such as exposure to gaseous pollutants, herb-icides, pathogen attack (Grant and Loacke, 2000),extreme temperatures and drought (Noctor and Foyer,1998).
Due to these large number of processes that pro-duce ROS, plants have evolved a series of detoxific-ation reactions, which are located in distinct cell or-ganelles. H2O2 formed during photorespiration by theconversion of glycolate into glyoxylate (Kendall et al.,1983) can be decomposed into H2O and O2 within theperoxisome, by catalase (CAT, EC 1.11.1.6), whichdoes not require an additional substrate (Scandalios,1994). The scavenging of H2O2 in other cell com-partments depends on distinct peroxidases that use areduced substrate for activity. Plant glutathione per-oxidases (EC 1.11.1.9) which can degrade H2O2 havealso been recently reported (Mullineaux et al., 2000).Ascorbate peroxidases (EC 1.11.1.11) use ascorbateas a reducing substrate and have been shown to belocalized in several cell compartments such as chloro-plasts, the cytosol and microbodies (Corpas et al.,2001; De Leonardis et al., 2000; Noctor and Foyer,1998). Glutathione reductase (GR, EC 1.6.4.2) canbe located in the chloroplast, cytoplasm and mito-chondria of higher plants (Stevens et al., 1997) and is
involved in the breakdown of H2O2 via the ascorbate-glutathione cycle (Noctor and Foyer, 1998), exhibitingchanges in activity following the application of envir-onmental stresses (Azevedo et al., 1998; Pilon–Smitset al., 2000).
The dismutation of superoxide into H2O2 and O2by superoxide dismutases (SOD, EC 1.15.1.1) hasbeen extensively studied (Bowler et al., 1992). Threeclasses of SOD have been observed in plants, cytoso-lic and chloroplastic Cu-Zn/SODs, a chloroplasticFe/SOD and a mitochondrial Mn/SOD, of which theformer two have been shown to be regulated by oxid-ative stress (Azevedo et al., 1998; Kliebenstein et al.,1998).
Recent reports on the responses of plant antioxid-ant enzymes to Cd and other metals, have indicatedseveral distinct patterns, which varied according tothe species and tissue analysed (Cardoso et al., 2002;Cuypers et al., 2000; Ferreira et al., 2002; Fornazier etal., 2002; Gallego et al., 1999; Hegedus et al., 2001;Schickler and Caspi, 1999; Vitória et al., 2001). Theaim of this study was to investigate the changes inactivities of the antioxidant enzymes, CAT, SOD andGR in the leaves and roots of C. juncea, a legumeplant species used as a compost (green fertilizer), inresponse to increasing concentrations of Cd.
Materials and methods
Plant material
Seeds of Crotalaria juncea were obtained from theseed bank of the Instituto Agronômico de Campinas,Campinas, Brazil.
Growth and Cd exposure
Seeds of C. juncea were germinated in a growth cham-ber under controlled environmental conditions (60–80% relative humidity, 25–27 ◦C temperature and a16/8 h light/dark photoperiod at 350 µmol m−2 s−1)for 7-d. At the end of this period, seedlings of ap-proximately the same size were selected and grownfor a further 96 h period in 1.5 l pots containingHoagland’s nutrient solution to which had been addedtwo CdCl2 concentrations (0.2 and 2 mM), plus theuntreated control. Leaf and root samples were harves-ted periodically during the experiment, leaf and rootgrowth was recorded and the samples were frozen inliquid nitrogen and stored at −80 ◦C, prior to enzymeextraction.

125
The experimental design was a randomized com-plete block with treatments arranged as individualpots, with five plants per pot, each replicated 4 times.Data were statistically analysed within each experi-ment and the results expressed as mean (± S.E.) offour independent replicates for antioxidant enzymesactivity measurements.
Cd accumulation
The accumulation of Cd in C. juncea leaves and rootswas analysed as described by Vitória et al. (2001) in ainductively coupled plasma mass spectrometer (ICP-MS).
Enzyme extraction
Enzyme extractions were carried out at 4 ◦C. Frozenplant tissues were ground to a fine powder in liquidnitrogen in a pestle and mortar and extracted at a ratio1:3 (w/v) fresh weight to extraction buffer (100 mMpotassium phosphate buffer [pH 7.5] containing 1 mMEDTA, 3 mM DTT and 5% [w/v] insoluble PVPP).The homogenate was centrifuged at 10 000 × g for30 min and the supernatant kept stored in separate ali-quots at −80 ◦C to be used for CAT, GR and SODanalyses. Protein was estimated by the method ofBradford (1976), using BSA as a standard.
Enzyme assays
CAT activity was assayed spectrophotometrically at25 ◦C as described by Chaparro–Giraldo et al. (2000)using an assay mixture comprising 1 ml 100 mM po-tassium phosphate buffer (pH 7.5) and 2.5 µl H2O2(30% solution) prepared immediately before use. Thereaction was started by the addition of 15 µl of plantextract and activity measured by monitoring the de-gradation of H2O2 at 240 nm over 1 min, against aplant extract-free blank.
GR activity was assayed spectrophotometricallyat 30 ◦C as described by Barata et al. (2000) ina reaction mixture consisting of 3 ml 100 mM po-tassium phosphate buffer (pH 7.5) containing 1 mM5,5′′-dithiobis(2-nitrobenzoic acid), 1 mM oxidizedglutathione and 0.1 mM NADPH. The reaction wasinitiated by the addition of 50 µl of plant extract. Therate of reduction of oxidized glutathione was followedby monitoring the increase in absorbance at 412 nmover 2 min.
CAT and SOD activity staining
Electrophoresis was carried out under native conditionin 5% and 8% polyacrylamide mini-gels for CAT andSOD activity staining, respectively. Electrophoresisrunning conditions were as described by Vitória et al.(2001) and buffers and gels were prepared as describedby Laemmli (1970) lacking SDS. Equal amounts ofprotein (40 µg) were loaded on to each lane.
CAT activity was also assayed in native PAGE gelsas described by Woodbury et al. (1971). Gels were in-cubated in 0.003% H2O2 for 10 min and developed ina 1% (w/v) FeCl3 and 1% (w/v) K3Fe(CN6) solutionfor 10 min.
SOD activity was determined on native PAGE gelsas described by Beauchamp and Fridovich (1971) andmodified by Azevedo et al. (1998). The gels wererinsed in distilled-deionized water and incubated inthe dark for 30 min at room temperature in an assaymixture containing 50 mM potassium phosphate buffer(pH 7.8), 1 mM EDTA, 0.05 mM riboflavin, 0.1 mMnitroblue tetrazolium and 0.3% (v/v) N,N,N′′,N′′-tetramethylethylenediamine (TEMED). At the endof this period, the gels were rinsed with distilled-deionized water and placed in distilled-deionized wa-ter and exposed on a light box for 5–10 min at roomtemperature until the development of colourless bandsof SOD activity in a purple-stained gel was visible.The reaction was stopped by transferring the gels to6% (v/v) acetic acid. Bovine SOD (Sigma ChemicalCo.) was used as a positive control of SOD activ-ity in all gels. SOD isoenzymes were visualized andclassified as described previously by Azevedo et al.(1998) by incubating gels for 20 min in solutions con-taining 2 mM KCN and 5 mM H2O2, separately andthen incubating in the SOD reaction mixture as above.The pre-treatment of the gels in H2O2 and KCN be-fore SOD staining allowed the classification of theSOD into Cu/Zn-SOD, Fe-SOD or Mn-SOD isoen-zymes. Mn-SOD is resistant to both inhibitors, theFe-SOD isoenzymes is resistant to KCN and inhibitedby H2O2, and Cu/Zn-SOD is inhibited by both inhib-itors. The concentration of protein loaded onto the gelwas increased to 150 µg in order to identify any minorSOD isoenzymes.
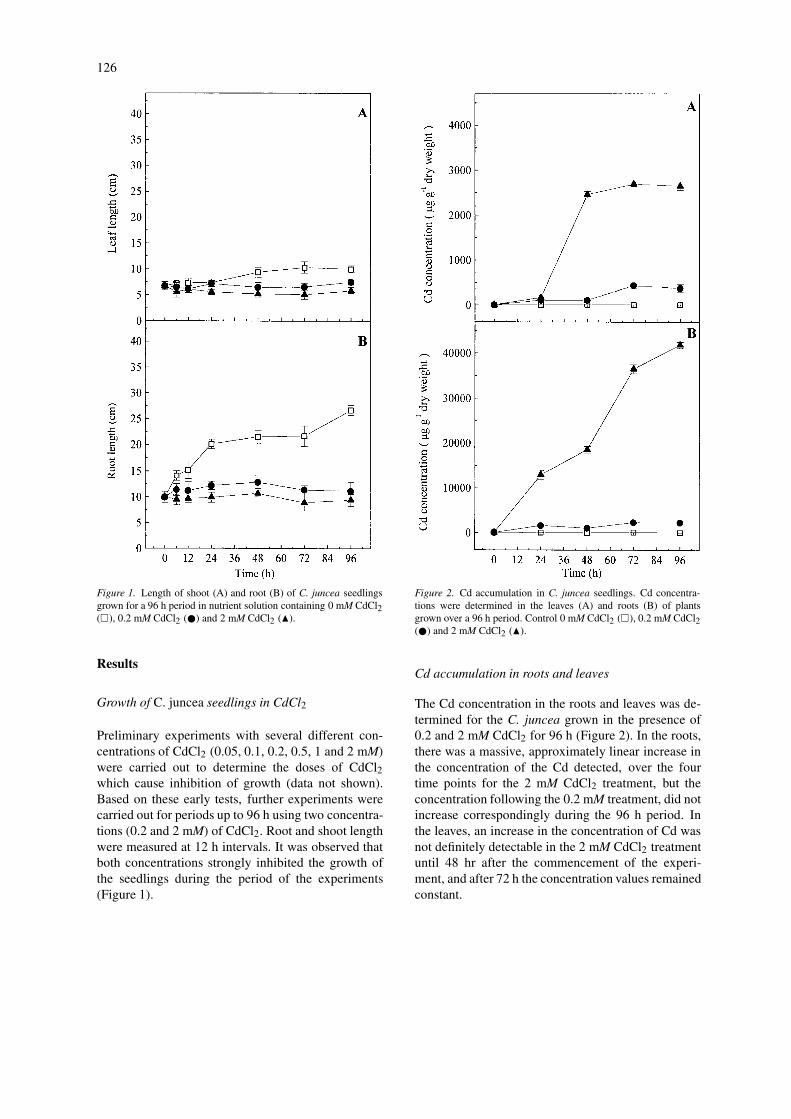
126
Figure 1. Length of shoot (A) and root (B) of C. juncea seedlingsgrown for a 96 h period in nutrient solution containing 0 mM CdCl2(�), 0.2 mM CdCl2 (�) and 2 mM CdCl2 (�).
Results
Growth of C. juncea seedlings in CdCl2
Preliminary experiments with several different con-centrations of CdCl2 (0.05, 0.1, 0.2, 0.5, 1 and 2 mM)were carried out to determine the doses of CdCl2which cause inhibition of growth (data not shown).Based on these early tests, further experiments werecarried out for periods up to 96 h using two concentra-tions (0.2 and 2 mM) of CdCl2. Root and shoot lengthwere measured at 12 h intervals. It was observed thatboth concentrations strongly inhibited the growth ofthe seedlings during the period of the experiments(Figure 1).
Figure 2. Cd accumulation in C. juncea seedlings. Cd concentra-tions were determined in the leaves (A) and roots (B) of plantsgrown over a 96 h period. Control 0 mM CdCl2 (�), 0.2 mM CdCl2(�) and 2 mM CdCl2 (�).
Cd accumulation in roots and leaves
The Cd concentration in the roots and leaves was de-termined for the C. juncea grown in the presence of0.2 and 2 mM CdCl2 for 96 h (Figure 2). In the roots,there was a massive, approximately linear increase inthe concentration of the Cd detected, over the fourtime points for the 2 mM CdCl2 treatment, but theconcentration following the 0.2 mM treatment, did notincrease correspondingly during the 96 h period. Inthe leaves, an increase in the concentration of Cd wasnot definitely detectable in the 2 mM CdCl2 treatmentuntil 48 hr after the commencement of the experi-ment, and after 72 h the concentration values remainedconstant.
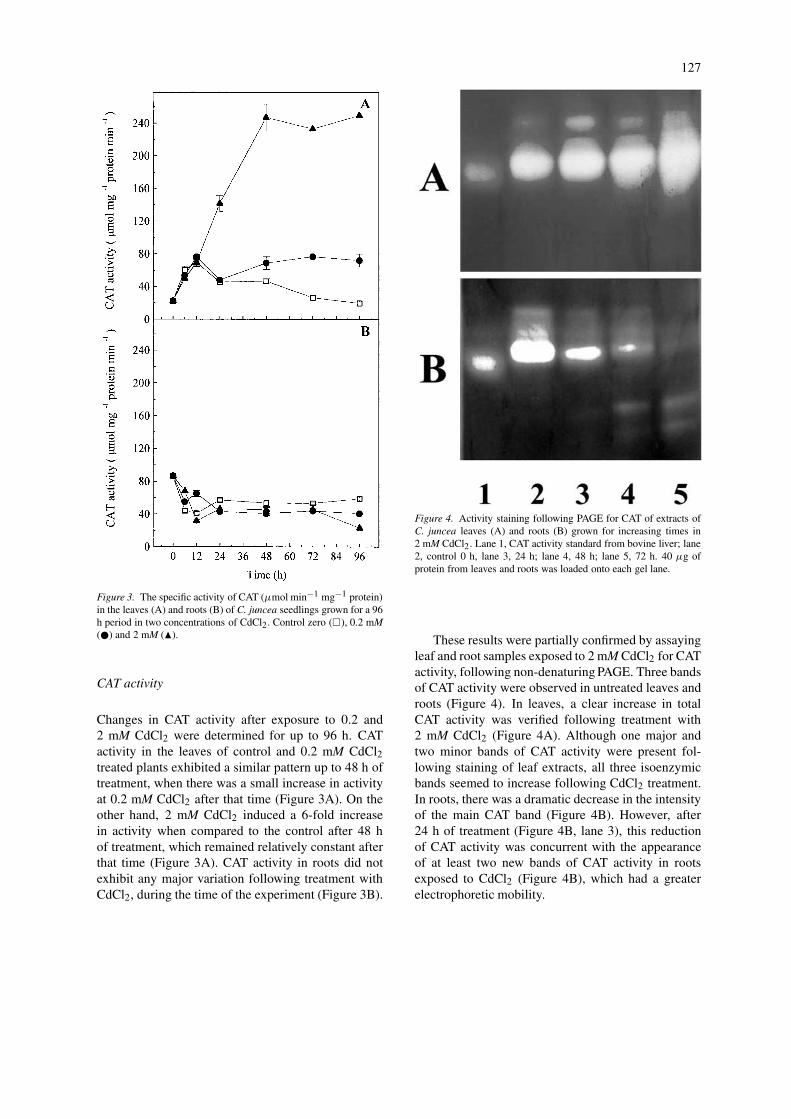
127
Figure 3. The specific activity of CAT (µmol min−1 mg−1 protein)in the leaves (A) and roots (B) of C. juncea seedlings grown for a 96h period in two concentrations of CdCl2. Control zero (�), 0.2 mM(�) and 2 mM (�).
CAT activity
Changes in CAT activity after exposure to 0.2 and2 mM CdCl2 were determined for up to 96 h. CATactivity in the leaves of control and 0.2 mM CdCl2treated plants exhibited a similar pattern up to 48 h oftreatment, when there was a small increase in activityat 0.2 mM CdCl2 after that time (Figure 3A). On theother hand, 2 mM CdCl2 induced a 6-fold increasein activity when compared to the control after 48 hof treatment, which remained relatively constant afterthat time (Figure 3A). CAT activity in roots did notexhibit any major variation following treatment withCdCl2, during the time of the experiment (Figure 3B).
Figure 4. Activity staining following PAGE for CAT of extracts ofC. juncea leaves (A) and roots (B) grown for increasing times in2 mM CdCl2. Lane 1, CAT activity standard from bovine liver; lane2, control 0 h, lane 3, 24 h; lane 4, 48 h; lane 5, 72 h. 40 µg ofprotein from leaves and roots was loaded onto each gel lane.
These results were partially confirmed by assayingleaf and root samples exposed to 2 mM CdCl2 for CATactivity, following non-denaturing PAGE. Three bandsof CAT activity were observed in untreated leaves androots (Figure 4). In leaves, a clear increase in totalCAT activity was verified following treatment with2 mM CdCl2 (Figure 4A). Although one major andtwo minor bands of CAT activity were present fol-lowing staining of leaf extracts, all three isoenzymicbands seemed to increase following CdCl2 treatment.In roots, there was a dramatic decrease in the intensityof the main CAT band (Figure 4B). However, after24 h of treatment (Figure 4B, lane 3), this reductionof CAT activity was concurrent with the appearanceof at least two new bands of CAT activity in rootsexposed to CdCl2 (Figure 4B), which had a greaterelectrophoretic mobility.
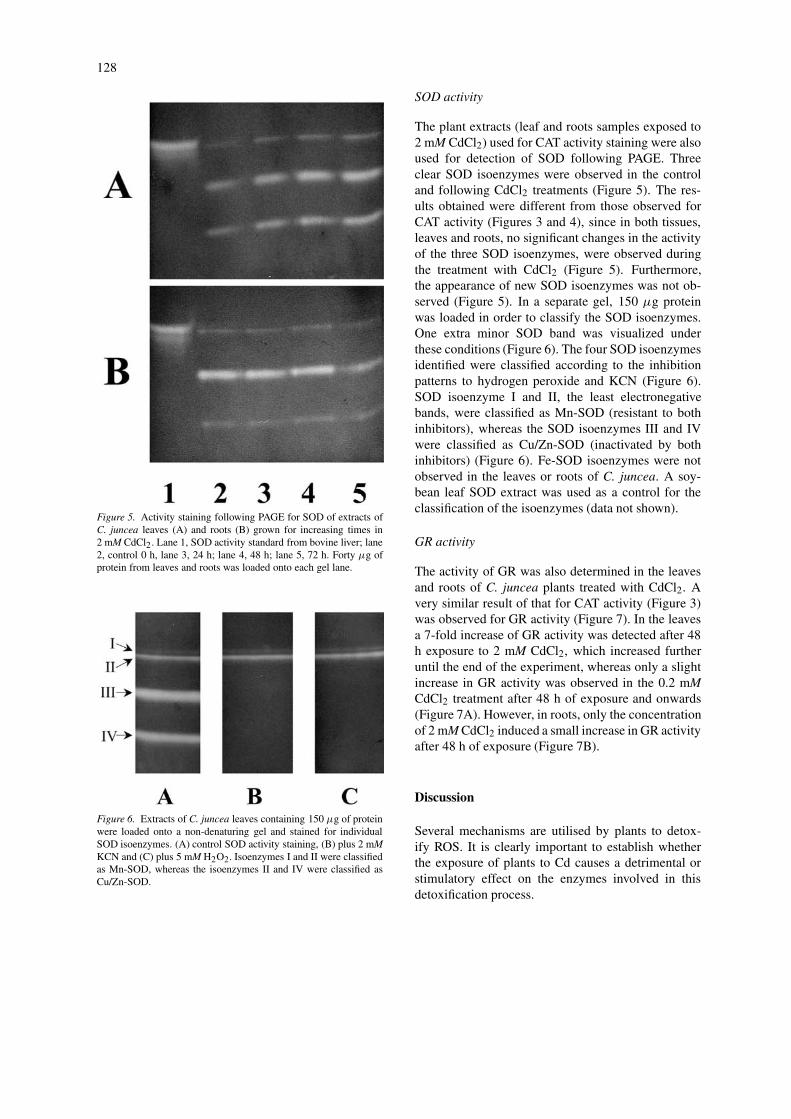
128
Figure 5. Activity staining following PAGE for SOD of extracts ofC. juncea leaves (A) and roots (B) grown for increasing times in2 mM CdCl2. Lane 1, SOD activity standard from bovine liver; lane2, control 0 h, lane 3, 24 h; lane 4, 48 h; lane 5, 72 h. Forty µg ofprotein from leaves and roots was loaded onto each gel lane.
Figure 6. Extracts of C. juncea leaves containing 150 µg of proteinwere loaded onto a non-denaturing gel and stained for individualSOD isoenzymes. (A) control SOD activity staining, (B) plus 2 mMKCN and (C) plus 5 mM H2O2. Isoenzymes I and II were classifiedas Mn-SOD, whereas the isoenzymes II and IV were classified asCu/Zn-SOD.
SOD activity
The plant extracts (leaf and roots samples exposed to2 mM CdCl2) used for CAT activity staining were alsoused for detection of SOD following PAGE. Threeclear SOD isoenzymes were observed in the controland following CdCl2 treatments (Figure 5). The res-ults obtained were different from those observed forCAT activity (Figures 3 and 4), since in both tissues,leaves and roots, no significant changes in the activityof the three SOD isoenzymes, were observed duringthe treatment with CdCl2 (Figure 5). Furthermore,the appearance of new SOD isoenzymes was not ob-served (Figure 5). In a separate gel, 150 µg proteinwas loaded in order to classify the SOD isoenzymes.One extra minor SOD band was visualized underthese conditions (Figure 6). The four SOD isoenzymesidentified were classified according to the inhibitionpatterns to hydrogen peroxide and KCN (Figure 6).SOD isoenzyme I and II, the least electronegativebands, were classified as Mn-SOD (resistant to bothinhibitors), whereas the SOD isoenzymes III and IVwere classified as Cu/Zn-SOD (inactivated by bothinhibitors) (Figure 6). Fe-SOD isoenzymes were notobserved in the leaves or roots of C. juncea. A soy-bean leaf SOD extract was used as a control for theclassification of the isoenzymes (data not shown).
GR activity
The activity of GR was also determined in the leavesand roots of C. juncea plants treated with CdCl2. Avery similar result of that for CAT activity (Figure 3)was observed for GR activity (Figure 7). In the leavesa 7-fold increase of GR activity was detected after 48h exposure to 2 mM CdCl2, which increased furtheruntil the end of the experiment, whereas only a slightincrease in GR activity was observed in the 0.2 mMCdCl2 treatment after 48 h of exposure and onwards(Figure 7A). However, in roots, only the concentrationof 2 mM CdCl2 induced a small increase in GR activityafter 48 h of exposure (Figure 7B).
Discussion
Several mechanisms are utilised by plants to detox-ify ROS. It is clearly important to establish whetherthe exposure of plants to Cd causes a detrimental orstimulatory effect on the enzymes involved in thisdetoxification process.
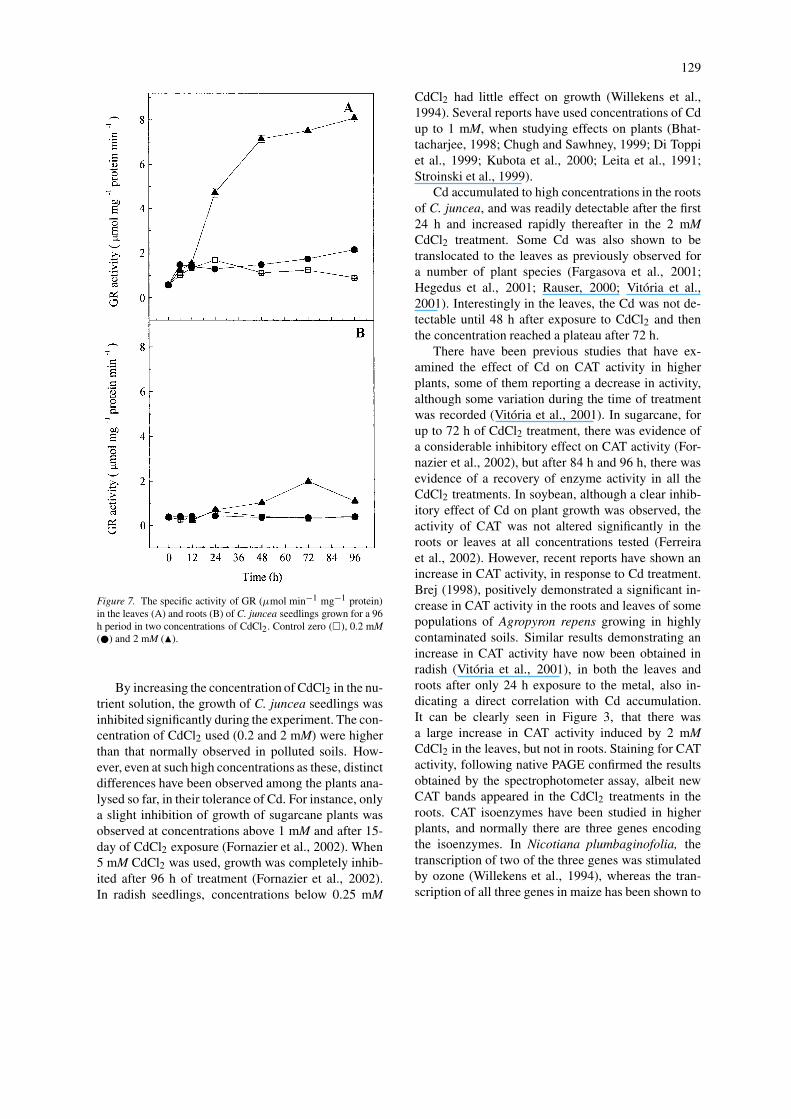
129
Figure 7. The specific activity of GR (µmol min−1 mg−1 protein)in the leaves (A) and roots (B) of C. juncea seedlings grown for a 96h period in two concentrations of CdCl2. Control zero (�), 0.2 mM(�) and 2 mM (�).
By increasing the concentration of CdCl2 in the nu-trient solution, the growth of C. juncea seedlings wasinhibited significantly during the experiment. The con-centration of CdCl2 used (0.2 and 2 mM) were higherthan that normally observed in polluted soils. How-ever, even at such high concentrations as these, distinctdifferences have been observed among the plants ana-lysed so far, in their tolerance of Cd. For instance, onlya slight inhibition of growth of sugarcane plants wasobserved at concentrations above 1 mM and after 15-day of CdCl2 exposure (Fornazier et al., 2002). When5 mM CdCl2 was used, growth was completely inhib-ited after 96 h of treatment (Fornazier et al., 2002).In radish seedlings, concentrations below 0.25 mM
CdCl2 had little effect on growth (Willekens et al.,1994). Several reports have used concentrations of Cdup to 1 mM, when studying effects on plants (Bhat-tacharjee, 1998; Chugh and Sawhney, 1999; Di Toppiet al., 1999; Kubota et al., 2000; Leita et al., 1991;Stroinski et al., 1999).
Cd accumulated to high concentrations in the rootsof C. juncea, and was readily detectable after the first24 h and increased rapidly thereafter in the 2 mMCdCl2 treatment. Some Cd was also shown to betranslocated to the leaves as previously observed fora number of plant species (Fargasova et al., 2001;Hegedus et al., 2001; Rauser, 2000; Vitória et al.,2001). Interestingly in the leaves, the Cd was not de-tectable until 48 h after exposure to CdCl2 and thenthe concentration reached a plateau after 72 h.
There have been previous studies that have ex-amined the effect of Cd on CAT activity in higherplants, some of them reporting a decrease in activity,although some variation during the time of treatmentwas recorded (Vitória et al., 2001). In sugarcane, forup to 72 h of CdCl2 treatment, there was evidence ofa considerable inhibitory effect on CAT activity (For-nazier et al., 2002), but after 84 h and 96 h, there wasevidence of a recovery of enzyme activity in all theCdCl2 treatments. In soybean, although a clear inhib-itory effect of Cd on plant growth was observed, theactivity of CAT was not altered significantly in theroots or leaves at all concentrations tested (Ferreiraet al., 2002). However, recent reports have shown anincrease in CAT activity, in response to Cd treatment.Brej (1998), positively demonstrated a significant in-crease in CAT activity in the roots and leaves of somepopulations of Agropyron repens growing in highlycontaminated soils. Similar results demonstrating anincrease in CAT activity have now been obtained inradish (Vitória et al., 2001), in both the leaves androots after only 24 h exposure to the metal, also in-dicating a direct correlation with Cd accumulation.It can be clearly seen in Figure 3, that there wasa large increase in CAT activity induced by 2 mMCdCl2 in the leaves, but not in roots. Staining for CATactivity, following native PAGE confirmed the resultsobtained by the spectrophotometer assay, albeit newCAT bands appeared in the CdCl2 treatments in theroots. CAT isoenzymes have been studied in higherplants, and normally there are three genes encodingthe isoenzymes. In Nicotiana plumbaginofolia, thetranscription of two of the three genes was stimulatedby ozone (Willekens et al., 1994), whereas the tran-scription of all three genes in maize has been shown to

130
be increased by the addition of hydrogen peroxide, al-though the time course of the response varied betweenthe genes (Polidoros and Scandalios, 1999).
The major function of CAT activity in leaves isto metabolise the peroxide liberated in the peroxi-some during photorespiration and the activity of theenzyme is greatly reduced when plants are grown inelevated atmospheres of CO2 (Azevedo et al., 1998).The increase in CAT activity in the leaves of C. junceadetected following CdCl2 treatment supports the hy-pothesis that Cd is causing the formation of peroxidewhich is then metabolised by CAT. Although no signi-ficant changes in total CAT activity following CdCl2treatment were detected in the roots (Figure 3B), itis clear from Figure 4B, that there has been a largechange in isoenzymic complement. The reduction inactivity of the predominant band would suggest thatthere is an effect of CdCl2, with the induction of newisoenzymic forms caused by the presence of ROS.However, it is not possible to speculate as to thesubcellular localisation of the CAT isoenzymes.
Three major SOD isoenzyme bands were observedfollowing the staining of native PAGE gels for enzymeactivity in the leaves and roots (Figure 5), however, aextra SOD isoenzyme band was visible when largeramounts of protein were loaded onto the gel (Fig-ure 6). The use of KCN and H2O2 in inhibition studies,indicated that the SOD isoenzymes I and II are Mn-SOD, while isoenzymes III and V were classified asCu/Zn-SODs (Figure 6). Contrary to the results ob-tained for CAT activity, SOD activity was unalteredin leaves or roots, at both CdCl2 concentrations andduring the time that the seedlings were exposed tothe metal. Previous research examining the effects ofCd on SOD activity in plants has also produced var-ied results as indicated for CAT activity, although thegreat majority have shown a decrease or no changein SOD activity (Ferreira et al., 2002; Fornazier etal., 2002). On the other hand, in pea (Dalurzo et al.,1997), Alyssum species (Schickler and Caspi, 1999)and radish (Vitória et al., 2001), increases in SODactivity have been observed in response to Cd expos-ure. In the case of radish (Vitória et al., 2001) SODisoenzymes were identified and separated on nativePAGE in an attempt to identify specific alteration inresponse to CdCl2, however a general increase for allisoenzymes was observed. Different oxidative stresseshave been shown previously to increase both the activ-ity and expression of genes encoding SOD (Bowleret al., 1992; Willekens et al., 1994). The results ob-tained suggest that, although Cd may be generating
an oxidative stress, there is not a sufficient increase insuperoxide to induce a change in SOD activity.
In a similar manner to CAT, changes in GR activitywere observed in response to the Cd induced stressin C. juncea seedlings. Such changes were clearlymore pronounced in the leaves following treatmentwith 2 mM CdCl2, but less so at 0.2 mM CdCl2 (Fig-ure 7). Several reports have detected increases in GRactivity in plants following exposure to Cd (Chaoui etal., 1997; Ferreira et al., 2002; Fornazier et al., 2002;Schickler and Caspi, 1999; Stroinski et al., 1999;Vitória et al., 2001), although the responses have oftenbeen dose dependent and variable over time. Oxidat-ive stress has been shown to induce increases in GRactivity (Stevens et al., 1997). Furthermore, it has beendemonstrated that the activity of ascorbate peroxidasealso increases following exposure to Cd (Chaoui etal., 1997; Shaw, 1995). Previous reports have shownthat Cd accumulates mainly in the roots of plantswhen compared to leaves and stems (Hart et al., 1998;Rauser, 2000; Vitória et al., 2001). The slight increasein GR activity in the roots suggests that this enzymemay be responding to Cd exposure and possible ac-cumulation of the metal in this tissue, by maintainingglutathione in the reduced form to be incorporated intophytochelatins (Cobbett, 2000). However, the signific-ant increase in GR activity in the leaves suggest thatthis enzyme plays a role in the detoxification of Cd-induced-ROS possibly via the glutathione-ascorbatecycle. These results for GR activity are differentfrom those recently reported for Phaseolus vulgaris(Chaoui et al., 1997), potato (Stroinski et al., 1999),radish (Vitória et al., 2001), soybean (Ferreira et al.,2002) and sugarcane (Fornazier et al., 2002) seed-lings, which when exposed to CdCl2, consistentlyexhibited significant increases in GR activity in theroots, although such increases were dependent uponCd concentration and the time of exposure. The resultssuggest that in C. juncea the reactive oxygen species(ROS) induced by Cd, are metabolised by CAT in theperoxisomes. In the case of GR activity, the increaseobserved in the leaves suggest that GR is also playing arole in the detoxification of Cd-induced ROS possiblyvia the glutathione-ascorbate cycle.
Acknowledgements
This work was financed by Fundação de Apoio àPesquisa no Estado de São Paulo (FAPESP) grant no.99/06429-0 and by the British Council. R. A. A. re-

131
ceived a research fellowship from Conselho Nacionalde Desenvolvimento Científico e Tecnológico (CNPq).G. J. G. P. received a Ph.D. scholarship from FAPESP.
References
Azevedo R A, Alas R M, Smith R J and Lea P J 1998 Response ofantioxidant enzymes to transfer from elevated carbon dioxide toair and ozone fumigation, in the leaves and roots of wild-type anda catalase-deficient mutant of barley. Physiol. Plant. 104, 280–292.
Barata R M, Chapparro A, Chabregas S M, Gonzalez R, Labate CA, Azevedo R A, Sarath G, Lea P J and Silva–Filho M C 2000Targeting of the soybean leghemoglobin to tobacco chloroplasts:Effects on aerobic metabolism in transgenic plants. Plant Sci.155, 193–202.
Beauchamp C H and Fridovich I 1971 Superoxide dismutase: Im-proved assays and an assay applicable to acrylamide gels. Anal.Biochem. 44, 276–287.
Bergmann H, Machelett B, Lippmann B and Friedrich Y 2001Influence of heavy metals on the accumulation of trimethylgly-cine, putrescine and spermine in food plants. Amino Acids 20,325–329.
Bhattacharjee S 1998 Membrane lipid peroxidation, free radicalscavengers and ethylene evolution in Amaranthus as affected bylead and cadmium. Biol. Plant. 40, 131–135.
Boussama N, Ouariti O and Ghorbal M H 1999a Changes in growthand nitrogen assimilation in barley seedlings under cadmiumstress. J. Plant Nutr. 22, 731–752.
Boussama N, Ouariti O, Suzuki A and Ghorbal M H 1999b Cd-stress on nitrogen assimilation. J. Plant Physiol. 155, 310–317.
Bowler C, Van Montagu M and Inzé D 1992 Superoxide-dismutaseand stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol.43, 83–116.
Bradford M M 1976 A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal. Biochem. 72, 248–254.
Brej T 1998 Heavy metal tolerance in Agropyron repens (L.) P.Bauv. populations from Legnica copper smelter area, LowerSilesia. Acta Soc. Pol. 67, 325–333.
Bruns I, Sutter K, Menge S, Neumann D and Krauss G J 2001 Cad-mium lets increase the glutathione pool in bryophytes. J. PlantPhysiol. 158, 79–89.
Cardoso P F, Molina S M G, Pereira G J G, Vitória A P and AzevedoR A 2002 Response of rice inbred lines to cadmium exposure. J.Plant Nutr. 25, 927–944.
Chaoui A, Mazhoudi S, Ghorbal M H and El Ferjani E 1997 Cad-mium and zinc induction of lipid peroxidation and effects onantioxidant enzyme activities in bean (Phaseolus vulgaris L).Plant Sci. 127, 139–147.
Chaparro–Giraldo A, Barata R M, Chabregas S M, Azevedo R A andSilva–Filho M C 2000 Soybean leghemoglobin targeted to potatochloroplasts influences growth and development of transgenicplants. Plant Cell Rep. 19, 961–965.
Chugh L K and Sawhney S K 1999 Effect of cadmium on activitiesof some enzymes of glycolysis and pentose phosphate pathwayin pea. Biol. Plant. 42, 401–407.
Cobbett C S 2000 Phytochelatins and their roles in heavy metaldetoxification. Plant Physiol. 123, 825–832.
Corpas F J, Barroso J B and del Rio L A 2001 Peroxisomesas a source of reactive oxygen species and nitric oxide signalmolecules in plant cells. Trends Plant Sci. 6, 145–150.
Cuypers A, Vangronsved J and Cliisters H 2000 Biphasic effect ofcopper on the ascorbate-glutathione pathway in primary leavesof Phaseolus vulgaris seedlings during early stages of metalassimilation. Physiol. Plant. 110, 512–517.
Dalurzo H C, Sandalio L M, Gomez M and Del Rio L A 1997 Cad-mium infiltration of detached pea leaves: Effect on its activatedoxygen metabolism. Phyton. 37, 59–64.
De Leonardis S, Dipierro N and Dipierro S 2000 Purification andcharacterization of an ascorbate peroxidase from potato tubermitochondria. Plant Physiol. Biochem. 38, 773–779.
Di Toppi L S, Lambardi M, Pecchioni N, Pazzagli L, Durante Mand Gabbrielli R 1999 Effects of cadmium stress on hairy rootsof Daucus carota. J. Plant Physiol. 154, 385–393.
Fargasova A 2001 Phytotoxic effects of Cd, Zn, Pb, Cu and Fe onSinapsis alba L. seedlings and their accumulation in roots andshoots. Biol. Plant. 44, 471–473.
Ferreira R R, Fornazier R F, Vitória A P, Lea P J and Azevedo R A2002 Changes in antioxidant enzyme activities in soybean undercadmium stress. J. Plant Nutr. 25, 327–342.
Fornazier R F, Ferreira R R, Vitória A P, Molina S M G, Lea P J andAzevedo R A 2002 Effects of cadmium on antioxidant enzymeactivities in sugar cane. Biol. Plant., 41, 91–97.
Foyer C H and Noctor G 2000 Oxygen processing in photosynthesis:regulation and signalling. New Phytol. 146, 359–388.
Gallego S M, Benavides M P and Tomaro M L 1999 Effectof cadmium ions on antioxidant defence system in sunflowercotyledons. Biol. Plant. 42, 49–55.
Grant J J and Loake G J 2000 Role of reactive oxygen intermediatesand cognate redox signalling in disease resistance. Plant Physiol.124, 21–29.
Groppa M D, Tomaro M L and Benavides M P 2001 Polyamines asprotectors against cadmium or copper-induced oxidative damagein sunflower leaf discs. Plant Sci. 161, 481–488.
Harada E, Choi Y E, Tsuchisaka A, Obata H and Sano H 2001Transgenic tobacco plants expressing a rice cysteine synthasegene are tolerant to toxic levels of cadmium. J. Plant Physiol.158, 655–661.
Hart J J, Welch R M, Norvell W A, Sullivan L A and Kochian LV 1998 Characterization of cadmium binding, uptake and trans-location in intact seedlings of bread and durum wheat cultivars.Plant Physiol. 116, 1413–1420.
Hegedus A, Erdei S and Horvath G 2001 Comparative studiesof H2O2 detoxifying enzymes in green and greening barleyseedlings under cadmium stress. Plant Sci. 160, 1085–1093.
Höfgen R, Kreft O, Willmitzer L and Hesse H 2001 Manipulationof thiol contents in plants. Amino Acids 20, 291–299.
Kendall A C, Keys A J, Turner J C, Lea P J and Miflin B J 1983The isolation and characterization of a catalase-deficient mutantof barley (Hordeum vulgare L.). Planta 159, 505–511.
Khudsar T, Mahmooduzzafar and Iqbal M 2001 Cadmium-inducedchanges in leaf epidermes, photosynthetic rate and pigmentconcentration in Cajanus cajan. Biol. Plant. 44, 59–64.
Kliebenstein P J, Monde R A and Last R L 1998 Superoxide dis-mutase in Arabidopsis: an eclectic enzyme family with disparateregulation and protein localisation. Plant Physiol. 118, 637–650.
Kubota H, Sato K, Yamada T and Maitani T 2000 Phytochelatinhomologs induced in hairy roots of horseradish. Phytochemistry53, 239–245.
Kumar R G and Dubey R S 1999 Glutamine synthetase isoformsfrom rice seedlings: Effects of stress on enzyme activity and theprotective roles of osmolytes. J. Plant Physiol. 155, 118–121.
Laemmli U K 1970 Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680–685.

132
Lee S M and Leustek T 1999 The effect of cadmium on sulphateassimilation enzymes in Brassica juncea. Plant Sci. 141, 201–207.
Leita L, Contin M and Maggioni A 1991 Distribution of cadmiumand induced Cd binding proteins in roots, stems and leaves ofPhaseolus vulgaris. Plant Sci. 77, 139–147.
Mullineaux P, Ball L, Escobar C, Karpinska B, Creissen G and Kar-pinski S 2000 Are diverse signalling pathways in the regulationof Arabidopsis antioxidant defence gene expression in responseto excess excitation energy. Phil. Trans. Roy. Soc. Lond. B. 355,1531–1540.
Noctor G and Foyer C H 1998 Ascorbate and glutathione: Keep-ing active oxygen under control. Annu. Rev. Plant Physiol. PlantMol. Biol. 49, 249–279.
Pilon–Smits E A H, Zhu Y L, Sears T and Terry N 2000 Over-expression of glutathione reductase in Brassica juncea: Effectson cadmium accumulation and tolerance. Physiol. Plant. 110,455–460.
Polidoros A N and Scandalios J G 1999 Role of hydrogen per-oxide and different classes of antioxidants in the regulation ofcatalase and glutathione S-transferase gene expression in maize(Zea mays L.). Physiol. Plant. 106, 112–120.
Rauser W E 2000 Roots of maize seedlings retain most of theircadmium through two complexes. J. Plant Physiol. 156, 545–551.
Scandalios J G 1994 Regulation and properties of plant catalases. InCauses of Photo-oxidative Stress and Amelioration of DefenseSystems in Plants. Eds C H Foyer and P M Mullineaux. pp 275–316. CRC Press, Boca Raton, FL.
Schickler H and Caspi H 1999 Response of antioxidative enzymesto nickel and cadmium stress in hyperaccumulator plants of thegenus Alyssum. Physiol. Plant. 105, 39–44.
Shaw B P 1995 Effects of mercury and cadmium on the activitiesof antioxidative enzymes in the seedlings of Phaseolus aureus.Biol. Plant. 37, 587–596.
Siedlecka A and Krupa Z 1999 Cd/Fe interaction in higher plants –its consequences for the photosynthetic apparatus. Photosynthet-ica 36, 321–331.
Somashekaraiah B V, Padmaja K and Prasad A R K 1992 Phyto-toxicity of cadmium ions on germinating seedlings of mungbean (Phaseoulus vulgaris) – involvement of lipid peroxides inchlorophyll degradation. Physiol. Plant. 85, 85–89.
Stevens R G, Creissen G P and Mullineaux P M 1997 Cloning andcharacterisation of a cytosolic glutathione reductase cDNA frompea (Pisum sativum L.) and its expression in response to stress.Plant Mol. Biol. 35, 641–654.
Stroinski A, Kubis J and Zielezinska M 1999 Effect of cadmium onglutathione reductase in potato tubers. Acta Physiol. Plant. 21,201–207.
Van Assche F and Clijsters H 1990 Effects of metals on enzymeactivity in plants. Plant Cell Environ. 13, 195–206.
Verma S and Dubey R S 2001 Effect of cadmium on soluble sugarsand enzymes of their metabolism in rice. Biol. Plant. 44, 117–123.
Vitória A P, Lea P J and Azevedo R A 2001 Antioxidant enzymesresponses to cadmium in radish tissues. Phytochemistry 57, 710–710.
Willekens H, Langebartels C, Tire D, Van Montagu M, Inzé D andVan Camp W 1994 Differential expression of catalase genes inNicotiana plumbaginifolia (L.). Proc. Natl Acad. Sci. USA 91,10450–10454.
Woodbury W, Spencer A K and Stahmann M A 1971 An improvedprocedure using ferricyanide for detecting catalase isoenzymes.Anal. Biochem. 44, 301–305.
Section editor: A.J.M. Baker




















