
A Theoretical Model of centrosome functioning
184
1 MARCO REGOLINI A THEORETICAL MODEL OF CENTROSOME FUNTIONING
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A Theoretical Model of centrosome functioning

1
MARCO REGOLINI
A THEORETICAL MODEL OF
CENTROSOME FUNTIONING

2
Short Forms
MC: Mother Centriole DC: Daughter Centriole MT: MicroTubule
MTOC: MicroTubule Organizing Center γ-TuRC: γ-Tubulin Ring Complex
PCM: Peri Centriolar Material RNP: RiboNucleo Protein
The centrosome at a glance
The requirements of a spherical reference system based on two orthogonal
protractors/goniometers show a surprising correspondence with the evidence
emerging from numerous experimental studies on centrioles and centrosomes: the
centrosome, as the main microtubule organizing center and because of the 9-fold
symmetry of its centrioles, their (transient) orthogonal arrangement and, above all,
their circumferential polarity (non-equivalence of spokes and triplets), may play the
role of a biological discrete and noise resistant interface, composed of two orthogonal
protractors, that recognizes and decodes morphogenetic instructions, or, more
generally, topogenic molecular targeting signals (frequently present at the N-terminus
of newly synthesized proteins or in the 3’UTR of mRNAs) and translates them by
delivering each targeted molecular complex (polarity and adhesion factors,
transmembrane receptors, mRNAs) into its expected 3D real location in the cell: like
an interface or a wiring device, the centrosome connects each targeting sequence with
the corresponding correctly-oriented microtubule: in this way morphogenetic
geometric (DNA coded) instructions are translated by the centrosome into actual
locations in cells, tissues and organs. Centrosome molecular geometry and
architecture imply its function: through the centrosome and its aster of robust
microtubules DNA can draw, build and “label” the intrinsic 3D grid line of the cell.
Centrosome, aster and primary cilium (its basal body is a centriole of the centrosome)
constitute the “hardware” of an interactive cross-talking system that manages

3
geometrical communication inside the cell and between cells and establishes in
tissues coordinated and shared cell polarity. Targeting sequences and related receptors
constitute the “software”. In addition, the centrosome (the most chiral and
enantiomorphous cell structure) plays a geometric key role in left-right patterning:
mother centriole circumferential polarity, if reversely oriented, constitutes a likely
base for bilateral symmetry and symmetry breaking (asymmetry establishment).
The centrosome, discovered by Edouard Van Beneden in 1883 and studied by
Theodor Boveri few years later, is an organelle of animal cells that plays the role of
MTOC (MicroTubule Organizing Center), controls the cell-cycle and takes part to
mitosis; however it is not the only MTOC of eukaryotic cells: fungi and plants, that
do not possess centrosomes, use other structures and pathways to organize
microtubules (MTs). The centrosome, composed of dozens of highly conserved
proteins (“if conserved, they are important”) plays an important role in mitosis but it
is not indispensable and therefore not essential for cell division. So, a first question
arises: such a sophisticated and accurately conserved organelle must play any
important role, but which?
Centrosomes are made up of two orthogonal (during S, G2 and M phases of the cell
cycle) centrioles, disposed like the capital letter “L”: an axial ”Mother” (see later)
Centriole (MC) and an eccentric “Daughter” Centriole (DC:) embedded in an
(apparently) amorphous protein matrix named PeriCentriolar Material (PCM),
responsible for anchoring and nucleation of MTs: an “aster” of non-intersecting
robust MTs irradiates radially from the centrosome to the cell cortex, like, from the
central square of a city, many streets irradiate toward (and connect with) the
periphery. When centrioles disengage (interphase) the PCM persists around the MC.
MTs are nucleated by γ-TuRCs (γ-Tubulin Ring Complexes), displaced on the
centrosome surface and supported by protein scaffolds; the centrifugal direction of
each MT is the consequence of the orientation and inclination of its own γ-TuRC:
therefore the PCM is not an amorphous matrix but a well ordered frame of proteins

4
able to orientate γ-TuRCs and their scaffolds (or docking platforms). Centrioles are
roughly cylindrical structures composed of nine blades of three parallel MTs
(“triplets”; some exceptions: the fruit fly Drosophila melanogaster has “doublets”, he
worm Caenorhabditis elegans “singlets”) arranged in a cylindrical or, rather,
prismatic barrel. All the properties of a biological spherical reference system small
organizer tool, based on two orthogonal protractors, are clearly showed, aren’t they?
…and a glance in a cell
In a cell there are approximately 1010
protein molecules (ten billions; Earth
population: ~7x109), subdivided into about ten thousand (10
4) different types. A cell
contains also millions of ribosomes, thousands of mitochondria, thousands and
thousands of surface receptors… Even more complex is the extracellular matrix that
surrounds cells, with dozens of different proteins and glycans (each type having many
different isoforms) showing astonishingly precise dispositions and orientations (the
cornea, for example: orthogonal layers, each one made up of parallel fibres). To
prevent an uncontrollable senseless chaos, eukaryotic cells have developed their inner
order: different membranes create compartments and the cell cortex too is
compartmentalized. “A living cell is not an aggregate of molecules but an organized
pattern, structured in space and in time… Some conceptual issues in the genesis of
spatial architecture: how molecules find their proper location in cell space, the
origins of supramolecular order, the role of the genes, cell morphology, the continuity
of cells, and the inheritance of order. The discussion is framed around a hierarchy of
physiological processes that bridge the gap between nanometer-sized molecules and
cells three to six orders of magnitude larger. Stepping stones include molecular self-
organization, directional physiology, spatial markers, gradients, fields, and physical
forces. The knowledge at hand leads to an unconventional interpretation of biological
order. I have come to think of cells as self-organized systems composed of genetically
specified elements plus heritable structures. The smallest self that can be fairly said

5
to organize itself is the whole cell. If structure, form, and function are ever to be
computed from data at a lower level, the starting point will be not the genome, but a
spatially organized system of molecules. This conclusion invites us to reconsider our
understanding of what genes do, what organisms are, and how living systems could
have arisen on the early Earth” (Harold, 2005, in ” Molecules into Cells: Specifying
Spatial Architecture”). According to this point of view, an MTOC able to irradiate,
from its oriented γ-TuRCs, an aster of conveniently directed MTs, not overlapping
nor intersecting, each one distinguishable and labeled by a specific receptor that only
recognizes (and interacts with) the targeting sequence intended for itself and thus able
to univocally wire the cell, would be able to ensure the establishment of a controlled
global map firstly in cells (a “cell wide web” or a “cell grid line”) and then
reproduced in tissues and organs (controlled disposition and orientation of extra-
cellular matrix fibers): then a first quick glance at centrosome functioning is
necessary to look for the possible correlation between development (morphogenesis)
and centrosome, already evident in early zygote cleavage. In effect as we will see
better later, Azimzadeh and colleagues, in “Centrosome loss in the evolution of
planarians” (2012), write: "We hypothesize that centrosome loss occurred
concomitantly with the loss of the spiral cleavage and oriented cell divisions in the
ancestor of planarians and schistosomes… A significant difference can be found in
the mode of embryonic cleavage however. Macrostomum [whose cells do have
centrosomes] retained the ancestral spiral cleavage, also found in annelids and
molluscs, which relies on a stereotypical pattern of cell division orientation. In
contrast, planarian and schistosomes embryos undergo divergent modes of
embryonic cleavage, which apparently do not involve oriented cell divisions”
(Azimzadeh et al., 2012). Cleavage has its characteristic pattern in each Clade and
Taxon: it is radial in Amphibians, spiral in Molluscans, bilateral in Ascidians,
syncytial in flies, discoidal in birds, rotational in Mammals.
“The embryogenesis of freshwater planarians is equally intriguing: cleavage of the
fertilized egg was described as ‘anarchic’. No overt gastrulation or epiboly has been

6
described.” (Sànchez Alvarado).
In mice, first embryonic divisions, described as irregularly rotational and
asynchronous, occur along unpredictable cleavage planes; in this stage, geometry of
cell arrangement is of little use; inner mass cell compactation does not follow any
architectural design and blastomeres lack centrioles that will appear only at the
blastocyst stage, when an evident embryo’s architecture emerges (orderly stereotyped
spatial disposition of cells).
A last number: Humans have ~1014
cells: ten thousand billion of (very complicated)
cells, organized in highly ordered tissues and organs. In each living organism there is
an absolute ”anarchy”: no Chief-cell, no Command-organ. Nonetheless living
organisms are the most ordered existing system. What is their architectural secret?
Shape and development: two main actors in morphogenesis
The aim and purpose of Metazoa development is to realize large organs and
organisms (able to run, fly, swim) making use of huge numbers of small dividing and
replicating units (the cells): the addition of thousands of little error in the 3D
displacement of the two new cells with respect to the position of the mother cell
would compromise (it is clearly incompatible with) the realization of high quality
architecture. What is the key to success in development and growth?
The biological mechanisms involved in tissue and organ development are highly
directional: division plane orientations, cell movements and convergent extension
take place according to precise directions; internal and external forces (osmotic
pressure, tension of extra-cellular fibers) stretch and bend cylindrical structures
according to the angle between cell axes and extra-cellular fiber directions; adhesions
between cells are not random but accurately localized and continuously remodeled
(differential adhesion); gradients of morphogens are formed using directionally
selective transport. Cells must be orientated and “informed” about their physical

7
spatial position and about directions: cells must know the real physical location of
“up”, “down”, “front”, “rear” and these points of reference must be shared with the
neighboring cells; plants, like compasses and GPSs, use extrinsic point of references
or cues: not so in Metazoa which must generate their own (shared) points of
reference. “Centriole duplication is part of the mechanism by which the cytoskeleton
of the daughter cell is patterned upon that of the mother” (Harold, 2005). This is
astonishingly evident in the organization of Drosophila imaginal discs and in their
process of extroflection; topology of imaginal disc primordia (composed of a few
cells) corresponds to that of developed limbs; the leg imaginal disc is firstly divided
along two orthogonal axes (dictated by Wingless, Decapentaplegic and Hedgehog
gradients) into different domains; only a small number of cells, at the intersection of
these domains, form the distal pole tip of the forming leg; in imaginal disc, cells
arrange themselves in 3D concentric rings corresponding to the region of the adult
leg: the most outer ring corresponds to the most proximal leg region (coxa), the
bounding inner ring corresponds to the trochanter, followed by other internal rings
relative to the femur, tibia, tarsal segments, while the innermost central disc
corresponds to the claws (the most distal region); the position of the distal pole tip is
near to the center of the disc but a little off-centered; the disc is also lightly
asymmetric, oval or ellipsoidal; also each ring or annulus shows its own
characteristic 3D off-centering and oval shape (each elliptic ring having its own
eccentricity and position of major and minor axes and of focal points), like an
irregular odd archery target; off-centering and ellipsoidal ring shapes are bilaterally
symmetric in both left and right imaginal discs; in this way the future 3D tilt of the
proximal-distal axis of each region of the forming limb is determined: its orientation
in respect to the three axes of the body and adjacent limb regions is the consequence
of the controlled 3D off-centering and eccentricity of each annulus (designed by the
carefully forecast 3D disposition of cells) that orient and drive the corresponding
extroflection tilt; the accurate 3D “off-centering” and eccentricity seems to be
radially and centrifugally transmitted from the central few cells outwards to annuli,

8
which are subdivided in some sectors like a dart board (clones or compartments),
controlled, step by step, by genetic programs carried out by cells able to manage their
own geometry.
The Drosophila mutants defective for the centriolar DSas-4 protein (Basto et al.,
2006) develop up to the adult stage, and, after using up maternal provisions of DSas-
4, are almost completely lacking in centrioles, at least in the brain, starting from the
third larval instar; the images of the adult mutant fly (see on the Internet the
comparative images of wild type vs. mutant in Basto’s free article “Fly without
Centrioles”) show an individual with monstrous deformities: the shape, the tilt and
the anomalous curvature of the wings certainly impede flight, just as the abnormal
angle between coxa and body cannot allow walking movements; what is the link
between morphogenesis (literally “shape-creation”) and centrosome? Metazoan cells
behave as if they had at their disposal an intrinsic (i.e. without external cues)
biological tool (the centrosome?), conceivable as roughly resembling and comparable
to a 3D-compass or 3D-GPS navigator (which, on the contrary, are driven by
external cues), able to grasp coded spatial indications (topogenic sequences) and
translate them to find and reach the intended directions and locations. Indeed, cell,
tissue and organ topology need, and cannot help but needing, such a specifically
dedicated “instrument”: effectively in migrating cells (in vivo and in vitro) the
centrosome reorients the cytoskeleton to reach the desired targets.
In multicellular Eukaryotes, a single fertilized oocyte (zygote) develops into an adult
organism (composed of billions of cells) which shows the characteristic shape of the
species: how is the correct species specific shape achieved? How are cells guided to
occupy their forecast position in the complex architectural plan of each organism?
Cell differentiation cannot explain how a particular shape is acquired: in effect
structures made up of the same type of differentiated cells show completely different
forms, as we can observe in skeletal stratified muscles, all composed of the same
differentiated type of cells and tissues (fibers of muscular tissue derived from
identical myoblasts, and connective tissue containing fibroblasts) but each one having

9
its own characteristic conformation: pectoralis major is quite different from deltoid,
platysma is completely different from trapezius, and so on; can the difference of
origin and insertion dictate the shape of each muscle? The different shapes of muscles
appear to be due to the layers of connective tissue which surround it and its fibers:
endomysium and muscle fibers are almost identical in every muscle, whereas, in each
muscle, fascia, epymisium and perimysium have their own characteristic form
(spindle shaped, cylindrical, flattened) dictated by the orientation of extracellular
fibres; these connective layers are made up of collagen (and many other components)
fiber whose orientation is carefully controlled by fibroblasts. Similar considerations
can be made about bones, articulations and cartilages: same type of differentiated
cells and tissues, but very different forms. What dictates shapes and forms?
The shape of organs and organisms is the consequence of the spatial disposition of
cells and orientation of extra cellular fibers: our hands and feet (except wrists and
ankles) have very similar (if not the same) anatomical and histological structures
(confirmed by phylogenetics and embryology), nevertheless they have clearly
different shapes because of the diverse spatial disposition of the cells (osteocytes
above all) and the different geometry of the extra cellular fibers. In our skin, the
microscopic arrangement of the cells in sebaceous and sweat glands is different: each
type of gland possesses not only its own type of differentiated cells, but also its own
particular histological disposition of cells, easily recognizable through a simple
microscope; there are millions of these glands, often very close, and always and
--------------------------------------------------------------------------------------------------------------------------------
Fig. 1 Centriole and centrosome cycle.
On each line, on the left the respective position of centrioles is depicted: the MC has distal appendages, the
DC has distal ribs, the PCM is a light halo. On the right the position of the centrosome in a dividing cell is
schematized (N: nucleus; astral MTs are outlined only in anaphase); in non dividing (G0) non motile cells the
MC is the basal body of the primary cilium.
A) Phase G1: MC and DC are disengaged.
B) Phase S:
i) the DC begins to mature close to the MC proximal end;
ii) the DC, now close to the MC distal end, loses its distal ribs;

10
iii) the DC acquires the distal appendages (now immature and short). It becomes a “new” MC; the
old MC orientates the new MC (“0°” mark coordination);
iv) from both MCs (old and new) a new procentriole-DC (cartwheel) arises; the diplosome
(MC+DC) containing the old MC maintains its orientation and is pulled by astral MTs towards
the forecast next spindle pole; the diplosome containing the new MC is displaced by astral MTs
(originating from the old MC) and pulled toward the forecast location of the new spindle pole.
C) Phase G2: procentrioles grow, becoming adult new DCs (distal ribs).
D) Prophase: both MCs form their own PCM.
E) Metaphase: both centrosomes are definitely positioned; the distal appendages of the new DC grow.
F) Anaphase: both MCs are completely formed (identical distal appendages) and their PCM is also
completely built. From both PCMs, astral MTs wire and map cell membranes of the two new cells.
G) Telophase: disengagement starts (only one cell is represented).
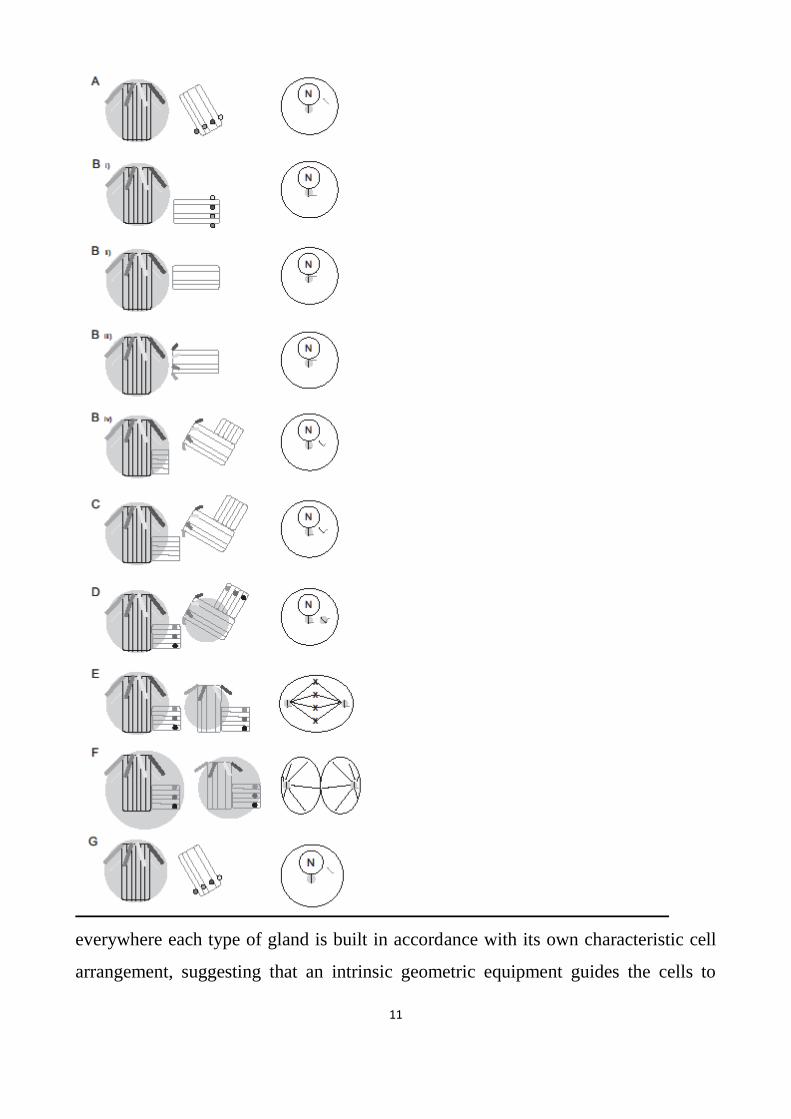
11
everywhere each type of gland is built in accordance with its own characteristic cell
arrangement, suggesting that an intrinsic geometric equipment guides the cells to

12
reach their correct position, or that a geometrical mechanism subdivides the cell
cortex and membrane in many different and non-equivalent compartments
(geometrically controlled to displace differentially adhesion complexes) in order to
adhere in a typical stereotyped fashion to neighboring cells (differential adhesion).
As every polyhedron has its characteristic shape with faces arranged in a fixed
position, each one with its own particular form, orientation and extension, anatomy
and histology show that cells, tissues and organs (and each microscopic structure like
the neprhon or the Corti’s organ) have their characteristic conformation and
topological arrangement; how do cells, tissues, organs and organisms achieve their
“geometrical-polyhedral” macro- and microscopic shape?
There are two main actors (hot topics) in morphogenesis: growth anisotropy and
heredity of forms.
As is known, anisotropy means that, given a material (a developing embryo in this
case), a physical quantity (growth rate, here) takes directionally dependent values.
When growth rate is rigorously isotropic (equal in every direction) spherical shapes
are formed; directional, but unpredictable, differences of growth rate (random
anisotropy) generate amorphous structures (neoplastic tissues are an example); to
realize well defined forms, anisotropy of growth rate must be attentively projected
and forecast: every direction shows its characteristic growth rate; so, growth is
orderly anisotropic from the earliest stages of zygote cleavage and becomes highly
anisotropic during organogenesis, resulting in structures that develop and grow in
predetermined directions; cell populations are correctly guided into fixed forecast 3D
positions, following programs characteristic of each species: cleavage, as we have
seen, can be radial, spiral, bilateral, syncytial, discoidal or rotational.
DNA: the first actor

13
"Heredity of forms" means that the zygote’s genome contains and transmits to the
offspring all the necessary programs to control growth, capable of precisely
indicating the geometry, or, rather, the topology (accurately forecast, memorized and
stored in DNA) which growth must respect: fertilized eggs only generate organisms
that acquire the characteristic shape of the species (in the few crosses that occur
successfully, morphological characteristics are blended). The famous transfer of a
nucleus from a mammary gland cell of a Finn Dorset sheep (DNA donor) into the
cytoplasm of an enucleated oocyte of a Scottish Blakface sheep (DNA host) produced
Dolly, which showed the morphological features of a Finn Dorset sheep (the donor of
DNA). In snails, only one maternal gene is responsible for left- or right-handed spiral
cleavage. DNA (or the nucleus, if we want to include epigenetic, histonic and non
histonic, nucleo-proteins), the first actor in morphogenesis, controls growth and
development of organisms: it provides coded (and stored) “directional signals” that
guide cells to reach and occupy their forecast positions: thus the genome strictly rules
the orientation of cell movements, differential cell-cell adhesions and division planes;
in addition it masters the directional organization of fibers in the extra cellular matrix.
To better explain this concept, let’s think about a dividing cell: the respective position
of the two daughter cells can be dictated by the orientation of the division plane
(which is a geometrical process) or by a topological disposition of adhesion factors
on the cell membrane (which is a geometrical process too: topological disposition of
polarity and adhesion factors on a controlled grid line): both processes require that
the cell has been previously mapped and that a tool does exist, able to manage any
cell reference system (cross-talking interactively with DNA, whose language is made
of polynucleotides). Comparative anatomy and molecular phylogenetics indicate that
the homology of structures, organs and tissues is founded on the homology of
genomes: comparisons of whole genomes show high similarity levels (homology of
sequences) that fit in with anatomical and morphological features; morphological
taxonomy corresponds to homology of sequences. Therefore DNA contains heritable
morphogenetic (coded) guidelines that, by means of molecular targeting signals, drive

14
and orientate populations of cells, imposing and dictating the correct direction that
cells must follow to reach their proper topological targets/locations and to correctly
assemble the extra cellular matrix: indeed the final result we can see in every adult
living being of every species is the correct stereotypical achievement of the forecast
species specific form, with every cell at its precise location (e.g. Bowman’s capsule
and loop of Henle in the nephron or Corti’s, Deiters’, Koelliker’s cells in the
cochlea) and extracellular fibers with organ or tissue characteristic orientation
(cornea, hairs, nails, beaks, claws). Then, how can DNA manage and supervise
developmental processes without a geometrical tool capable of recognizing,
understanding and translating (like an interface) its genetic topogenic instructions?
“The genetic instructions often include information pertinent to the localization of the
product. Targeting sequences direct proteins to the plasma membrane, nucleus,
mitochondria, or lysosomes. Certain proteins and mRNAs are transported
individually to particular locations in cell space, and this localization depends on
having an appropriate sequence. Transport vesicles recognize specific target
membranes, such as the Golgi, vacuole, or plasma membrane, with the aid of SNARE
proteins. But there is much more to growth and division than manufacturing the
parts. A rod-shaped cell must also elongate with constant diameter, construct an
efficient apparatus to partition its chromosomes, locate its midpoint, lay down a
septum, and undergo fission. In eukaryotic cells, targeted vesicle fusion requires, in
addition to the SNAREs, both a delivery system and a secretory apparatus” (Harold,
2005). For identifying precise locations in the cell cortex (apical, basal, anterior,
posterior) and reaching them, firstly the cortex must be “mapped” or “wired” (i.e.
subdivided in many non-equivalent compartments, orderly disposed on a virtual grid
line, somehow identifiable and recognizable through any kind of label), then the cell
must have at its disposal all the instruments necessary to manage this “map”: a tool is
necessary that can “understand” molecular geometric and directional coded signals
and “translate” them finding and physically reaching the intended cortical position.
“Structural and genetic studies suggest that asymmetry of the centriole (basal body)

15
plays a critical determining role in organizing the internal organization of algal cells,
through the attachment of microtubule rootlets and other large fiber systems to
specific sets of microtubule triplets on the centriole…To understand cell organization,
it will be critical to understand how the different triplets of the centriole come to have
distinct molecular identities” (Marshall, 2012).
Whatever the molecular signals are (polynucleotides or polypeptides), how are they
recognized, decoded and translated, identifying with precision the desired direction,
location and orientation?
Centrosome: the second actor
Centrosome and morphogenesis are somehow correlated. Without the centrosome,
planarian and schistosome embryos cannot retain the ancestral spiral cleavage: they
undergo divergent modes of cleavage, without performing oriented cell divisions,
and, in addition, gastrulation fails.
Pathology shows that mutations in several loci coding for centriolar proteins cause
several morphogenetic disorders, from Protists to humans: Chlamydomonas bld
mutant does not have flagella, uni has only one flagellum, vfl shows multiple ectopic
flagella: they all present abnormalities in the cytoskeleton, location of the nucleus and
formation of the mitotic furrow; in humans, ciliopathies are associated with some
morphological disorders: renal and hepatic malformations, polydactyly, Meckel-
Gruber and Bardet-Biedel syndrome, etc. Cilia are assembled by basal
bodies/centrioles: what is the role of centrioles and centrosomes in development?
What is the link between the genes involved in morphogenesis and the architectural
organization of cells? Whatever the signal is (whether it is a cue to differentiate or a
factor to proliferate or a stimulus to move, etc.) the last step of each morphogenetic
process (beyond differentiation) is necessarily topologic, geometric and directional,
in order to drive cells and tissues toward the forecast architectural position they must

16
occupy: forms and shapes are not the consequence of the kind or type of cells
(differentiation), on the contrary they are determined by the spatial disposition of
cells (geometrical architecture): in the Corti’s organ, the different type of cells is not
surprising and is the consequence of cell differentiation, but the precise orientation of
the basilar, tectorial and Reissner’s membranes is astonishing and cannot be
explained by cell differentiation. Let’s consider Drosophila gastrulation: no Gurken
signal from the oocyte to ventral follicle cells allows these cells produce Pipe; then a
long cascade of gene activities is triggered (gastrulation-defective, snake, easter,
spätzle, toll, tube, pelle, cactus, dorsal) till Dorsal protein (that is found everywhere
in the syncytial blastoderm) enters the nucleus only in ventral cells; this cascade of
gene activation (together many other – zerknüllt, tolloid, decapentaplegic, rhomboid,
twist, snail, fgf8 – that are activated or repressed depending on the level of
stimulating factors) can explain the different fates (differentiaton) of cells (dorsal-
ectoderm, lateral-ectoderm, neurogenic ventrolateral ectoderm, ventral-mesoderm),
but not movements: how ventral midline cells modify their cortical adhesion
junctions, and actin cytoskeleton to invaginate (with bilaterally symmetric
movements) at the central furrow and form the ventral tube? Morphogenetic signals
are molecules (sequences of nucleotides or amino-acids) that impose directions to
already polarized cells (receptors for Planar Polarity factors are not random
distributed on the cell membrane, but occupy defined locations): then, there must be a
noise-resistant structure, capable of precisely recognizing and identifying geometric
topological signals, decoding, interpreting and translating them to find and finally
reach the desired locations to drive cell movements. "The preceding interphase aster
centers and orients a pair of centrosomes prior to nuclear envelope breakdown, and
the spindle assembles between these prepositioned centrosomes…Current models for
cleavage plane determination propose that metaphase spindles are positioned and
oriented by interactions of their astral microtubules with the cellular cortex, followed
by cleavage in the plane of the metaphase plate. In early frog and fish embryos,
where cells are unusually large, astral microtubules in metaphase are too short to

17
position and orient the spindle. Rather, the preceding interphase aster centers and
orients a pair of centrosomes prior to nuclear envelope breakdown, and the spindle
assembles between these prepositioned centrosomes" (Wühr et al., 2010).
Centrosomes are pulled up to their forecast position by astral (labeled and
distinguishable) MTs that ensure one-to-one correspondence between centrosomal
(labeled and distinguishable) and cortical compartments.
To drive cell movements, DNA (the operator) needs a “steering wheel”, a rudder that
transforms (translates) its orders into correctly oriented motions. This implies the
existence of a tool able to organize a three-dimensional reference system, the second
actor in morphogenesis, made up of real cellular structures; evidently, the classical
Cartesian reference system with three axes crossing the cytoplasm, does not exist
(this assertion is true for Metazoa, whereas in Plant parallel MT rings under the cell
wall design a real grid line, like a globe with designed parallel but without meridians,
something similar to a “cylindrical Cartesian system”); on the contrary, however, a
spherical reference system organizer does exist, which requires a structure, as small
as is desired, composed of two “protractors/goniometers” orthogonal to each other
and capable of generating oriented rays: this is the centrosome, with its two
orthogonal centrioles, built with a 9-fold symmetry and capable of assembling robust
microtubules (MTs) in oriented directions. May we suspect that differently directed
MTs are also differently labeled by specific receptors to play a useful role in mapping
and labeling cells? Indeed cells equipped with a non-labeled cytoskeleton seem more
confusing than cells without a cytoskeleton: imagine a bus, a railway or a subway
station with platforms lacking numbered information panels or an airport whose gates
have no numerals nor displays. In effect an MTOC that irradiates an aster of identical
undistinguishable MTs where motor proteins (kinesins and dyneins) walk up and
down would be a useless and chaotic obstacle for molecule diffusion. On the
contrary, through the centrosome, DNA builds and manages a real cell “subway
network”, made up of a real central station with numbered distinguishable platforms
(the centrosome ant its oriented and labelled γ-TuRC scaffolds), rails (MTs), trains

18
(kinesins and dyneins) and different suburban stations (cortical compartments).
About cleavage and centrosome
“When centrioles are experimentally ablated, spindles drift within the cell”
(Azimzadeh and Marshall, 2010).
“Some characteristics of mouse early development could be linked to the absence of
centrioles: (1) the absence of regular cleavage planes during early development; (2)
the absence of any detectable axis of polarity within the blastomeres before the 8-cell
stage; (3) the random position of the spindle relative to the axis of polarity within the
blastomere during the fourth cleavage. Centrioles may act to keep PeriCentriolar
Material components in a precise position throughout the cell cycle and so be useful
in the control of the position of the axis of polarity and division. This may become
more important in differentiated cells, such as those found in the outer layer of the
blastocyst” (Gueth-Hallonet et al., 1993).
Let’s read again what Azimzadeh et colleagues write in “Centrosome loss in the
evolution of planarians” (2012): "The absence of centrioles in dividing cells implies
that planarians do not use the pathway for centriole duplication that underlies
centrosome reproduction in other animals, but only assemble centrioles de novo
during the differentiation of ciliated cells...We hypothesize that centrosome loss
occurred concomitantly with the loss of the spiral cleavage and oriented cell
divisions in the ancestor of planarians and schistosomes….It is remarkable that the
loss of such a conserved organelle as the centrosome occurred within not-parasitic
flatworms, as cellular and developmental processes appear largely conserved
between these species. A significant difference can be found in the mode of embryonic
cleavage however. Macrostomum retained the ancestral spiral cleavage, also found in
annelids and molluscs, which relies on a stereotypical pattern of cell division
orientation. In contrast, planarian and schistosomes embryos undergo divergent

19
modes of embryonic cleavage, which apparently do not involve oriented cell
divisions”.
About spiral cleavage, Rabinowitz and Lander (2010) write:
“Spiralling embryos are found in a large group of invertebrate phyla but are largely
uncharacterized at a molecular level. These embryos are thought to be particularly
reliant on autonomous cues for patterning, and thus represent potentially useful
models for understanding asymmetric cell division. The series of asymmetric
divisions that produce the micromere quartets are particularly important for
patterning because they subdivide the animal-vegetal axis into tiers of cells with
different developmental potentials. In the embryo of the snail Ilyanassa, the IoLR5
RNA [Ilianassa obsoleta Long R5 RNA] is specifically segregated to the first quartet
cells during the third cleavage. Here, we show that this RNA, and later the protein,
are maintained in the 1q(121) cells and their descendents throughout development.
Some IoLR5-expressing cells become internalized and join the developing cerebral
ganglia. Knockdown of IoLR5 protein results in loss of the larval eyes, which
normally develop in association with these ganglia. Segregation of this RNA to the
first quartet cells does not occur if centrosomal localization is bypassed. We show
that the specific inheritance of the RNA by the first quartet cells is driven by a
discrete RNA sequence in the 3' UTR that is necessary and sufficient for localization
and segregation, and that localization of another RNA to the first quartet is mediated
by a similar element. These results demonstrate that micromere quartet identity, a
hallmark of the ancient spiralian developmental program, is controlled in part by
specific RNA localization motifs.”.
“One of the major types of mRNA transport in Drosophila embryos is the movement
of mRNAs involved in developmental pattern formation to the apical centrosome”
(Blower, 2013). It is well known (Scott, 2014) that different mRNAs are
asymmetrically distributed in Ilyanassa blastomeres by the centrosomes:
Decapentaplegic mRNA is delivered by the centrosomes to the macromeres whereas
micromeres receive by the centrosomes Long R5 RNA: the supposition that the

20
centrosome is equipped with different receptors to recognize targeting sequences is
plausible. We have just already seen (Wühr et al., 2010) that centrosomes, in the large
frog blastomeres, are positioned in such a way (through distinguished, oriented and
labelled MTs) that they look like trolley buses correctly connected to the two
polarized electric wires, orientated and properly front-rear positioned in respect to the
“electric city-skeleton” made of aerial-suspended wires: during their controlled
movements, guided and routed by a labelled MT cytoskeleton, they take and delivery
mRNAs to the same destination they are reaching.
"It came as a surprise to all of us that planarians could get rid of centrosomes
without affecting their regenerative potential, says Howard Hughes Medical Institute
and Stowers investigator Alejandro Sánchez Alvarado, Ph.D. It suggests to us that the
evolutionary pressure to maintain centrosomes may have very little to do with cell
division itself. There may be another function for centrosomes that is still
obscured…The fact that centrioles were retained in this organism while centrosomes
were lost, really speaks to the idea that centrioles evolved primarily for making cilia,
and not for their mitotic functions, says senior author Wallace Marshall“.
“Although the orienting cytoskeletal element is not yet known, this model
[intracellular origin of Left-Right patterning] provides a comprehensive, quantitative
synthesis of all the molecular and biophysical steps leading from Left-Right
orientation within single cells to asymmetric gene expression in the early embryo,
and it does not depend on ciliary motion. This scheme also parallels a model of
dorsal-ventral patterning, where an embryo-wide pattern arises from early
intracellular movement of Wnt containing particles by kinesin motor proteins along
oriented microtubule tracks” (Levin and Mercola, 2007).
“The centrosome has evolved in multicellular organisms from the basal
body/axoneme of the unicellular ancestor. It plays a major role in organizing the
microtubule cytoskeleton in animal cells. During interphase, the centrosome
organizes an astral array of microtubules that participate in fundamental cellular
functions such as intracellular trafficking, cell motility, cell adhesion and cell

21
polarity” (Azimzadeh and Bornens, 2007).
“There may be another function for centrosomes that is still obscured…” (Sánchez
Alvarado, 2012).
Another function for the centrosome: what is it?
”Is it possible to confirm this idea that the circumferential, morphological, structural
and molecular asymmetry of centrioles can be inferred from Mammals ciliated
epithelia? While the circumferential anisotropy of centrioles cannot be ascertained
within the centrosome, its existence can be inferred from the properties they express
during ciliogenesis, be it the formation of a primary cilium or of bona fide 9+2 cilia
in ciliated epithelia, some of which at least derive directly from the centrioles. As in
Ciliates and flagellates, these basal bodies nucleate appendages of various molecular
compositions (basal foot, striated rootlets, alarm sheets, etc, which anchor the basal
body to the membrane and to the cytoskeleton) and these nucleations arise at specific
sites of the basal body cylinder; in particular, the basal foot is located on triplets 5
and 6 corresponding to the side of the effective stroke of the cilium. What is
remarkable is that basal feet develop before the basal bodies reach their membrane
site and before they acquire their functional orientation“ (Beisson and Jerka-
Dziadosz, 1999).
Indeed the centrosome has a structure able to perform a different (unique and
peculiar) function, a logistic task: it looks like a “network switch”, or a “switching
hub”, the well-known computer networking multi-port device that receives
destination-addressed messages or frames from different equipment which it is
connected to (through one-to-one cables) and delivers each message only to the
(labelled) device which the message is addressed to: an “one-to-one” wired
connection links the switch to each device, each device is “labelled” and
distinguishable (private address) and each data is also labelled and addressed to only
one device. The previous considerations on the centrosome create the impression that
it plays the role of a molecular interface (see Fig. 4 and 5 on page 45 - 46) that
receives molecular complexes labelled by targeting sequences (ligands), “reads”

22
labels (signal-receptor interaction and capturing) through its γ-TuRCs (non-
equivalent) receptors and, by kinesins (motor proteins) that move along the
corresponding oriented MTs, delivers each molecular complex (transmembrane
receptors, polarity and adhesion factors, mRNAs above all) to where they are
destined. Thus it can be the cellular instrument used by DNA to manage cell, tissue
and organ topology. "Basal bodies and centrioles display structural and functional
polarities that play an important role in the spatial arrangement of the cytoskeleton
and hence the polarity of the cell" (Geimer and Melkonian, 2004). This is the key
idea: in Metazoa both mother’s and daughter’s centriole polarities (structural and
functional i.e. inclination and non-equivalence) are transmitted to the aster of
centrosomal MTs so that the aster is made of non-equivalent MTs (each MT is
distinguishable because of its own orientation and molecular labelling of its γ-TuRC:
one direction, one label) ; in Ciliates (Paramecium) centriole triplets are different and
distinguishable and acquire a precise and coordinated orientation with the complex
cytoskeleton: the triplet N° 9 links to the “postciliary ribs”, the triplet N° 4 is attached
to the “transverse ribbon”, the triplets N° 5-6-7 are connected to the “kinetodesmal
fibers”; similarly the (non-equivalent!) pins of a microchip are connected to precise
conductive tracks of a printed circuit board; as in Ciliates the centrioles organize the
“cyto”-skeleton, in Metazoa the two orthogonal centrioles of the centrosome organize
the architecture of the “PCM”-skeleton and confer to the centrosomal γ-TuRCs (or
their scaffolds) molecular complexes (labels) that on the one hand identify and
distinguish each MT with correspondence to its centrosomal geometrical location and
orientation, on the other hand allow an univocal interaction between targeting
sequences intended for cortical locations reachable through the oriented MT they
label. “It seems that in Protists and in Metazoa the triplets of basal bodies are not-
equivalent” (Beisson and Jerka-Dziadosz, 1999).
Plants and animal anatomy

23
Fixed in the ground, plants control their anisotropic growth by extrinsic reference
systems (gravity and light); animals, on the other hand, need an intrinsic self-made
reference system (something like an oriented cell grid line) to manage their geometry:
plants do not have orthogonal centrioles, animals do. Then it seems that the
centrosome is the intrinsic reference system tool of animals.
Plants, centrosome-free, have developed simple anatomical and histological
structures, with cylindrical or laminar arrangement: beautiful but anatomically
simple, repeated a large number of times. “The overall structural organization of
plants is generally simpler than that of animals. For instance, plants have only four
broad types of cells, which in mature plants form four basic classes of
tissues…organized into just four main organ systems” (Lodish 2012). In contrast
animals, centrosome equipped, have developed anatomical forms that are particularly
varied and complex (3D rather than 2D laminar arrangements) whose architecture
implies the existence of a right and proper geometric tool: the peacock’s livery, the
shells of crustaceans; animals show a high architectural accuracy and precision also
at the tissue level: kidney cortex and medulla or spongy bone osteons and trabeculae;
the same holds true for organs: skeleton and heart. In Vertebrates the shape of
structures that perform complex functions is astonishing: the curvature of cornea, lens
and retina strictly meets the need for projecting and focusing images; the inner ear -
labyrinth and cochlea- in Mammals and Birds has a shape perfectly suited to measure
the different vector-components of acceleration and analyze the frequency of
acoustical signals (the basilar membrane of the Corti’s organ performs, in a sense, a
“biological Fast Fourier Transform”).
Previous in-depth studies have investigated “what” centrosomes might be and
“what” might be their task: now the question is “how” centrosomes work.
A possible model of centrosome and centrioles function in cell and

24
tissue topology: theoretical foundation
How cells control geometry of tissues and organs
“Several principles of construction of a microscopically small device for locating the
directions of signal sources in microscopic dimensions: it appears that the simplest
and smallest device that is compatible with the scrambling influence of thermal
fluctuations as are demonstrated by Brownian motion is a pair of cylinders oriented
at right angles to each other.” (Albrecht-Buehler, 1981).
“The centrosome has evolved in multicellular organisms from the basal
body/axoneme of the unicellular ancestor. It plays a major role in organizing the
microtubule cytoskeleton in animal cells. During interphase, the centrosome
organizes an astral array of microtubules that participate in fundamental cellular
functions such as intracellular trafficking, cell motility, cell adhesion and cell
polarity” (Azimzadeh and Bornens, 2007).
“A new centriole normally arises in a definite spatial relationship to an existing one
and at right angles to it” (Harold, 2005).
“It seems that in Protists and in Metazoa the triplets of basal bodies are not-
equivalent” (Beisson and Jerka-Dziadosz, 1999).
In order to coordinate these highly anisotropic, polarized and oriented functions, the
centrosome must have a suitable geometric structure, able to manage cell polarity and
decode “geometric” messages transmitted by DNA, to whose control all the previous
functions are subject.
The orthogonal arrangement of the centrioles in the centrosome suggests that this
organelle is the cell “reference system” organizer: a spherical-system builder based
on two orthogonal protractors/goniometers, one to manage geometry in the “x y”
plane, the second to manage geometry in an orthogonal plane containing the “z” axis.

25
Centrosome geometry and architecture must necessarily imply its function: as a
technical designer firstly squares a sheet, similarly, during the last period of each cell
division, through the centrosome, DNA firstly maps and wires the non-polarized and
homogeneous cell cortex: so, DNA can build the intrinsic 3D map of the cell,
transforming a (DNA-coded) “virtual” grid line in a real “actual” cellular grid (the
“cell wide web”) with intrinsic points of reference which dictate and orientate the
position of membrane polarity factors; DNA uses the centrosome to polarize the
whole cellular cortex and membrane in order to assume the control and the mastery
of the cellular and extracellular environment. Please pay attention to the expressions
“through the centrosome, DNA firstly maps” and “DNA uses the centrosome”: the
centrosome is only a sophisticated instrument in the hands of (controlled by) DNA,
then it does not work autonomously, but is strictly directed and managed by DNA.
Then we must look for a plausible molecular “hardware” (geometrical PCM
structure) and a corresponding molecular “software” (targeting sequences and
centrosomal receptors) able to perform such functions.
What are the requirements of a spherical reference system organizer made up of two
orthogonal protractors? How may the centrosome play the role of an interface,
composed by two orthogonal goniometers, making use of the 9-fold symmetry of
centrioles, their orthogonal arrangement and their circumferential polarity?
How must a “biological protractor/goniometer” be organized? Basic
characteristic of the protractors.
The difference between a simple ring and a protractor/goniometer (or a clock dial)

26
consists in the strictly ordered circumferential polarity and asymmetry of the
graduated protractor: like the dial of a clock is subdivided into an ordered row of
equidistant sectors and segments equipped with diverse (non-equivalent) marks,
similarly a protractor has different equidistant numerals, sequenced, in an ordered
fashion; if a homogenous ring is rotated about its axis, an observer will not perceive
any difference: the ring will appear always the same, identical and indistinguishable
from the original arrangement; on the contrary the observer will realize whether a
protractor (or a watch dial) has been rotated and will also estimate the angle of
rotation; “circumferential asymmetry” (or polarity) means that a disc or a circle are
made up of several different elements (arcs, sectors, segments) which are
distinguishable because of own individual label/marks, and then non-equivalent. One
mark (“0°” on the protractor, “12” on the clock) must be considered the beginning
(here it will be named the “0°” mark): it is fundamental for building and neatly
assembling the ordered sequence of the other (non-equivalent) marks, and essential
for orienting the protractor. A clock dial, marked by different and distinguishable
“Braille” characters and oriented in the space (horizontal, with the numeral “12” as
usually opposite to the user) allows a blind or a visually-impaired person to precisely
find a definite sector of the plane. Let us consider a protractor or a clock dial with
only nine equidistant, different, distinguishable Braille marks like the nine equidistant
triplets of the centriole wall that, at this moment of our analysis, we suppose to be
non-equivalent: why? So a sophisticated architecture, together with 9-fold symmetry,
costly and difficult to realize, would be meaningless and useless if their 9 components
were undistinguishable: in transverse sections, a centriole looks like a gear of a pump
or a shaft of a rotor or a drum of a turbine, but neither drills nor propellers can be
useful inside the cytoplasm and cells are neither mixers nor centrifuges; therefore we
now admit, but later we will face this argument, that the “non-equivalence” of the
nine centriolar triplets is an “informational non-equivalence” and this appears a
plausible, impressive and convincing reason (centriole works like a biological
informational protractor) to explain the centriolar and centrosomal complicated and

27
conserved architecture. Coming back to the “Braille” protractor, the plane containing
the protractor is divided into nine corresponding, ordered, labeled, identifiable (and
then non-equivalent) sectors like a dart board. After orienting the protractor (the “0°”
mark, like the numeral “12” on a clock dial, in the farthest segment opposite to the
user) a blind (or a visually-impaired) person that receives a vocal signal (a number),
through his fingers matches the signal with the corresponding Braille character and
individuates the sector of the plane corresponding to the signal: a signal (intended for
a desired location and containing the corresponding coded information) has been
“translated” into its real location on a plane through the interaction between the signal
and the corresponding Braille character operated by the blind person’s fingers. The
blind (or the visually-impaired) and the Braille clock dial are the elements of a set
which symbolizes the biological-protractor/centriole.
Left-Right (a first quick glance)
By reverting (clockwise > counterclockwise) the order of the sequence of the Braille
characters, the protractor becomes symmetric in respect to the original one. If the
blind (or visual impaired) has a “reverse” protractor (horizontal and orientated like
the original one, with the “0°” mark always in the same position, in front of him or,
better, opposite to him or, rather, on the farthest sector in respect to him, like “12” on
a clock dial), receiving the same vocal signal as before, matches the signal with the
corresponding Braille character but individuates a sector of the plane which is
symmetric, compared to that he would find through a non-reverse Braille protractor:
he “translates” the same coded signal into a symmetric location. By only a simple
reversion of the sequence of the marks, it is possible to realize bilateral symmetry,
without changing an enormous number of topogenic instructions; this is particularly
interesting from an evolutionary point of view: one single change in place of
thousands of changes is much more likely. Left-Right question will be deeply

28
analyzed later.
DNA, through the centrosome, generates ex novo a 3D oriented environment in
a previously homogenous cell
A “Braille marked” protractor is not used to measure, it is a translator of address-
signals into their corresponding locations in the plane; two such orthogonal
protractors perform the same identical task in 3D; in a cell, such instrument is an
interface, a biological geometric tool that receives molecular coded signals (input:
targeting or topogenic sequences), each one intended for a particular sector,
recognizes them through their tertiary 3D structure, matches each one with the
corresponding mark (ligand-receptor interaction) and returns (or indicates) the spatial
position (output) of the desired locations: any location can be easily reached through
an oriented ray arising from the selected mark. This comparison reveals other
important features of centrosome functioning; for a blind person, the space of an
empty room is homogeneous, without any point of reference, everywhere identical
and then “meaningless”; through an oriented 9-graduated “Braille” protractor, the
same space acquires nine intrinsic, autonomous (relative to the blind) and useful
points of reference (without external cues): it is no longer homogeneous but
subdivided into nine different and recognizable sectors and becomes a “meaningful”
space, a useful room, a detailed and usable environment. Similarly DNA, through the
centrosome, generates ex novo a 3D (autonomously oriented) labeled environment, a
grid line in a previously homogenous cell, transforming and converting it in a
mapped, wired, “webbed” and polarized in detail cell: an “useless” cell cortex
becomes an “useful and usable” space. This is the reason we have chosen the
expression “informational non-equivalence” for the different triplets. Through the
centrosome, a “blind” DNA becomes a “sighted” DNA: through the centrosome
DNA, so to say, turns on the light in the cell.

29
Orientation of the first protractor
Speaking about the symmetric “Braille” protractor, a lot of expressions have been
used to explain its position and orientation in respect to the user, in order to underline
how delicate the issue of orientation is; in effect protractors/goniometers must be
oriented: in a globe, one protractor is “horizontal” (equatorial), the other one is
“vertical” (meridian) and passes through the North and South poles. The first
protractor, arranged on the equatorial plane, lies on the “x y” plane of the spherical
reference system and its axis coincides with the “z” axis of the system: it is
responsible for indicating the longitude ( coordinate). The “0°” mark is used to
orient the protractor: on the globe it coincides, by convention, with the meridian
passing through Greenwich; on the “x y” plane it coincides with the “x” axis. Its nine
marks indicate nine meridian (or vertical) wedges. In a metazoan cell, as we have
already seen, DNA polarizes the cell cortex, often in the absence of any cue: there is
no extrinsic point of reference as in other systems (plants, compasses, GPSs), but
each species uses its own mode to polarize the cell: the “0°” mark can coincide with
the entry of the sperm or be positioned randomly, like a technical designer is free to
square a square sheet choosing randomly which side will be the “top” side.
Orientation of the second protractor
The second protractor, responsible for the latitude (θ coordinate), is vertical,

30
orthogonal to the first (Fig. 2 at page 35); it is possible to define its “top” and its
“bottom”: as in a clock on a tower the mark “12” is on the vertical axis and always at
the top, the diameter crossing the “0°” mark is vertical, parallel to the “z” axis (the
axis of the first protractor) and the “0°” mark is positioned at the top, aligned with (in
front of, facing) the “0°” mark of the first protractor. In a classic spherical reference
system, θ takes values from 0° to 180°, and then it is convenient to consider the
second protractor divided, by the “vertical” diameter crossing its “0°” mark, into two
halves (two facing symmetric hemi-protractors) the “right” one showing on its round
external border four marks (+40°; +80°; +120°; +160°) clockwise ordered starting
from the “0°” mark, and the “left” one showing the same four marks, but
counterclockwise ordered (-40°; -80°; -120°; -160°). So, these eight marks (four
“right” and four corresponding “left”) are symmetrically positioned relative to the
“0°” and its vertical axis : they divide the space into five parallel “horizontal” sectors
(two polar caps and three parallel disks): each “horizontal” sector (cap or disk) is
subdivided into nine parts by the first protractor while each meridian “vertical”
wedge is subdivided into five parts by the second protractor. It is not necessary that
both goniometer centres coincide. As on a globe, longitude covers the entire
circumference (2π radians or 360°; 9 meridian 40° wedges) whereas latitude covers
(symmetrically) only half circumference (π radians or 180°: π/2 or 90° North and π/2
or 90° South; 2 caps and 3 parallel disks: Fig. 3 at page 37). Note the eccentricity of
the two protractors: on a globe the protractors have the same center and so it is in the
canonical spherical reference system, but, in order to model the centrosome, we must
follow the eccentric disposition of the two centrioles. This “two-
protractors/goniometers-instrument” is sufficient to subdivide the 3D space into 45
pyramids (Fig. 3 at page 37) with the apex at the centre: each pyramid is identifiable
by its own longitude and latitude ( and θ coordinates, corresponding to the
protractors’ marks): each base faces and subtends a vertex solid angle of 4π /45
steradians, then its extension (4π r2/45), in a cell with a diameter of 10 μm (radius: 5
μm; surface: 4π r2 approximately 314 μm
2) corresponds to a cell cortex extension of

31
about 7 μm 2
(a circle with a diameter of 3 μm, or a square with a side of 2.6 μm).
These dimensions together with the physical properties of the MTs (bending-
resistance and rigidity) give an idea about the interesting order of magnitude of the
noise-resistance of this system and of its precision, much better than that of chemical
gradients: 45 cell cortex compartments (or rather poles) are much more than six poles
(anterior, posterior, dorsal, ventral, left and right) and assure a fine tuned polarity.
Fig. 2 Centrosome theoretical geometrical model: a spherical reference system composed of
two orthogonal protractors/goniometers.
A: frontal view of two orthogonal protractors/goniometers, subdivided into nine sectors, which
represents the two orthogonal centrioles: the first (horizontal) represents the base of the MC,
arranged on the equatorial “x y“ plane; its “0° ” mark is used to orient the protractor/centriole; the
second, the DC (vertical, orthogonal to the first), is closer to the reader: both “0°“ marks coincide; it
is convenient to consider the second protractor divided, by its “vertical” diameter crossing the “0°”
mark, into two halves (two opposite symmetrical hemi-protractors).
B: schematic lateral view of the proximal end of both centrioles (during S, G2) to show the
respective position of the above two sections.
An input-output spherical reference system tool based on two orthogonal
protractors/goniometers

32
A spherical reference system tool based on and built by two orthogonal
protractors/goniometers with nine notches, which is what the centrioles precisely are,
in order to recognize (input) coded geometric signals (often at the N-terminus of the
newly synthesized proteins or in the 3’UTR of mRNAs) and translate (output) them
by delivering each targeted complex into the desired final location, must possess: 1)
different marks (non-equivalent centriolar triplets); 2) a constant and ordered
sequence of marks; 3) a start mark; 4) a controlled orientation.
A centrosome will possess all these requirements if its two orthogonal centrioles,
built with 9-fold symmetry, organize the PCM in order to orient 45 scaffolds that
sustain γ-TuRCs (longitude and latitude inclination), and transfer to the PCM their
circumferential non-equivalence, labeling (in coordinated accordance with
neighboring cells) its 45 scaffolds with molecular receptors capable of recognizing
exclusively the targeting sequences corresponding to their orientation: the assembly
of robust MTs with corresponding oriented directions will be the last step.
As it will be better discussed later, Mennella (2012) found that “by using SIM and
STORM subdiffraction-resolution microscopies to visualize proteins critical for
centrosome maturation, we demonstrate that the PCM is organized into two main
structural domains: a layer juxtaposed to the centriole wall, and proteins extending
later away from the centriole organized in a matrix. Analysis of Pericentrin-like
protein reveals that its carboxy terminus is positioned at the centriole wall, it radiates
outwards into the matrix and is organized in clusters having quasi-nine-fold
symmetry. By RNA-mediated interference, we show that Pericentrin-like protein
fibrils are required for interphase recruitment and proper mitotic assembly of the
PCM matrix”. Centriole geometry is then well defined: through the assembly of
molecular complexes with centrifugal radial direction, centrioles are really able to
organize scaffolds/docking platforms for γ-TuRCs, transmitting their circumferential
polarity (non-equivalence of 9-fold rotational polarity is, for the moment, only
supposed): each scaffold (and its γ-TuRCs) assumes a double inclination imparted by
two orthogonal centrioles (longitude and latitude) to acquire its own orientation,

33
parallel to the local centrosome surface tangent plane and receives from centrioles
also the marks (molecular receptors) corresponding to its orientation. The surface of
the centrosome is compartmentalized into oriented and distinguishable (targeted or
labelled) docking platforms for γ-TuRCs (Fig. 3 on this page, 4 on page 43 and 5 on
page 44).
-------------------------------------------------------------------------------------------------------
Fig. 3 Centrosome theoretical geometrical model: discrete subdivision of the centrosome surface.
Nine meridians and four parallels subdivide the centrosome surface into 45 small areas (scaffolds for -
TuRC, which include SAS-4/CPAP, CNN, Asl and Pericentrin), each oriented in correspondence to its
position: their inclination is the result of the addition of two inclinations, one imposed by the MC (longitude)
and the other by the DC (latitude). As on a globe, longitude covers the entire circumference (2π; 9 different
meridians or 9 different meridian 40° wedges) while latitude covers (symmetrically) only half circumference
(π; 2 caps and 3 parallel discs).
-------------------------------------------------------------------------------------------------------
The centrosome-based wiring system could also function in the opposite way. As
before observed, according to this hypothesis, DNA acquires a complete command of
development geometry through the centrosome (and its role of geometrical interface).
Signals from other cells (gap junctions, adhesion molecules, signalling factors,
morphogens) in addition to their intrinsic signaling property can take interesting
information about their provenance, particularly useful, for instance, on the midline;

34
inwards carried by MT-linked dyneins, the direction of their provenance can be
interpreted by the spherically polarized centrosome and immediately used by DNA
which is thus able to integrate external inputs, cues and data (type of signalling
molecule, concentration level, provenance direction) with own genetic programs: a
very important interactively cross-talking between DNA, inner and outer
environments can be carried out by the centrosome and, as we will examine above,
the primary cilium, also connected to the centrosomal MC which constitutes its basal
body. Then, different cells (nearby or far away) can communicate interactively to
build complex organs and tissues (many experiments on competition between
genetically engineered clones in Drosophila wing illustrate this concept). Indeed,
something of the kind happens in the intraflagellar system of transport: Rosenbaum
and colleagues (1969) observed that, in Chlamydomonas (a bi-flagellated unicellular
green alga) following partial surgical ablation of one flagellum, the other is
immediately reduced in length, then re-growth occurs simultaneously in both flagella
until they reach the normal length. Evidently, the basal body organizes the formation
of the axoneme, and, in addition, it is able to decipher and decode information
coming from the periphery of the axoneme (Engel et al., 2012). “Our results reveal a
mechanism that orchestrates both the centriole-to-basal body transition and
subsequent cilia assembly through miRNA-mediated post-transcriptional regulation”
(Cao et al., 2012). The film by Ritter and Griffiths (2012), in which the centrosome of
cytotoxic T lymphocytes (CTL) “senses” the area of the cell membrane where the T
cell has recognized a tumor cell, is significant: this contact must be absolutely
strongly maintained, so the T cell response is the rotation, coordinated and directed
by the centrosome, of nucleus, cytoskeleton and cytoplasm in order to carry the killer
machinery close to the point of cell-cell contact. “The centrosome determines where
secretion occurs by contacting the plasma membrane at the point where the T cell
recognizes the tumor cell…Cytotoxic T lymphocytes are immune cells that destroy the
structural integrity of virus-infected or cancer-causing cells by releasing toxic
granules into them. At the point of contact between the CTL and its target cell an

35
immunological synapse is generated, through which the cells interact. Inside cells the
centrosome, where the cell skeletal elements are made, is responsible for moving
granules to the right place. Brought to the synapse in a CTL the centrosome ensures
that the toxic granules will hit their intended target when the immune cells dock onto
diseased cells. When the centrosome fails to move to the synapse, docking is
unsuccessful, and diseased cells cannot be discarded” (Griffiths and Wathne, 2012).
In conclusion, the finding of Mennella and colleagues, the evidence from Beisson and
Jerka-Dziadosz taken together with centrosome role in transferring different mRNAs
into different cells in Ilyanassa, indicates that our words “two orthogonal centrioles,
built with 9-fold symmetry, organize the PCM and transfer to it their circumferential
non-equivalence (γ-TuRC receptors) in order to orient 45 scaffolds that sustain γ-
TuRCs” are not off track; we are on the right rails.
This is “in nuce” (as ancient Romans said, that is “in embryo”, an expression which is
particularly apt and sounds good in this context) the geometrical model of
centrosome functioning. Now this embryonic and immature (“zygotic”) idea must
develop and mature into an adult presentable proposal. Until now we have conducted
an inductive reasoning, making some generalizations from theoretical and
experimental observations. From now on we will conduct a deductive analysis,
formulating hypotheses strongly founded on experimental evidences and testing the
possibilities to reach logical conclusions. To do that, it is necessary to ascertain more
deeply the theoretical and biophysical properties of a centrosome-based reference
system, starting from empiric data (experimental evidences and findings: from facts
to hypotheses and theories) .
The centrosome
The centrosome (Greek: κέντρον, center, and σωμα, body; “central body” because of

36
its recurring position near the center of the cell) is a sphere made of two orthogonal
centrioles embedded in a proteinaceous matrix (the PCM) from which many radially
(spherically, centrifugally) directed MTs originate (the “aster”).
Centrioles and basal bodies (at the base of cilia and flagella) are the same organelle:
the centriole of the spermatocyte becomes the basal body of the sperm, which, after
fertilization, is again a centriole in the zygote; in non-dividing cells, the MC becomes
the basal body of the non-motile primary cilium. Centrioles are cylinders (or, rather,
prisms, although the orientation of their faces is tilted: each face rotates inward
(about 55°) around its longitudinal edge); depending on the species, centrioles have a
height of 150 to 500 nm and a base diameter of 100-200 nm; their wall is made up of
nine longitudinal bundles (the faces of the prism), each consisting of three parallel
MTs that form the “triplets” (one MT, named “A”, is a complete cylindrical MT, the
other two, named “B” and “C”, parallel to the first one, are incomplete, in transverse
sections appearing like a capital letter C); inside, the structure of the proximal portion
of a new arising centriole has the appearance of a cartwheel, with a central hub and
nine spokes, each one radially directed towards the A-MT of a triplet.
The centrosome, found only in Metazoa and in multicellular algae but not in higher
plants and most fungi, is a sphere of electron-dense material inside which there are
two centrioles (orthogonal to each other during S phase and mitosis). The centrosome
shows many peculiar and unique characteristics: it is the only organelle that exists in
a single copy per cell, together with the nucleus and the primary cilium whose basal
body, in non-dividing cells, is (surprise surprise) the MC of the centrosome itself; it is
the only organelle that does not have a membrane: however the PCM (Peri Centriolar
Material), seemingly amorphous, is strongly organized in cylindrical layers and its
components do not diffuse into the cytoplasm, although some components show
remarkable turnover and high spatiotemporal variability; it is in contact through its
microtubules with each other organelle and each cytoplasmic and cortical location.
Up to now, more than 200 different molecular complexes, highly conserved from
Protists to Mammals, have been identified in centrioles and centrosomes. A recurring

37
question: why are so many different (and highly conserved) proteins orderly arranged
in coaxial cylinders around both centrioles in the PCM that compose the centrosome?
When a cell enters the S-phase, the two centrioles separate (disengagement) and,
orthogonally to each of them, a new one is assembled; the two new centrosomes,
each containing an old centriole, named “mother”, and a younger, newly assembled,
centriole, the “daughter”, participate (without being strictly indispensable) in mitosis
and form the mitotic spindle, of which they constitute the poles; each centrosome will
be inherited by one of the two daughters (sisters) cells. This unique semi-conservative
duplication is reminiscent of something similar to DNA, but it has been clearly and
definitively shown that centrioles and centrosome do not contain DNA and their
duplication does not occur by copying a template: on the contrary two different
pathways for centriole duplication have been described, one requiring a preexisting
MC that works like a platform to control the new DC assembly, and a de novo (“ex
novo”, to me, is a more correct Latin form, but it is rarely used in biological papers)
assembly pathway, turned off when a MC is present (with few exceptions, like
multiciliated cells). Such unusual and unique duplication modality (in which the self-
assembly capability of many macromolecular centriolar complexes is particularly
important) might allow the centrioles of the daughter (sister) cells to acquire the same
circumferential ordered polarity (positioning of the “0°” mark) and orientation of
their mother: so, from the zygote on, every cell could have its polarity coordinated
with the global polarity of the whole tissue and its architectural project.
The centrioles inside the centrosome are quite different: the older one, the ”mother”,
has nine external radial distal (“distal” is toward the centrosome surface, “proximal”
toward the centrosome center) and nine sub-distal appendages, while the younger one
(the “daughter”) has nine different small distal ribs (structural difference); only the
MC can form a “primary cilium” (functional difference); the orthogonality between
MC and DC is asymmetric: the daughter’s longitudinal axis, if prolonged, crosses the
other, so that the base of the DC faces the lateral surface of the MC’s proximal end,
but not vice-versa so that only the MC has a central pivotal position like a hub

38
(geometric difference: eccentricity). The centrosome is polarized: the “L” shape of
the two centrioles allows a proximal-distal axis to be identified (it coincides with the
MC axis), with a distal pole characterized by the described MC appendages, and a
second orthogonal axis along the DC. During centriole duplication, one spoke of the
“cartwheel” of the DC is parallel to the MC axis.
After mitosis, the MC, in non dividing cells, remains fixed, anchored to the cell
membrane by numerous MTs, positioned at the base of the primary cilium (of which
it is the basal body), whereas, during interphase in dividing cells it is often linked to
the nucleus; the DC is much more free in the cytoplasm.
The centrosome is the main cytoskeletal organizing center: from its PCM numerous
robust MTs emerge, centrifugally directed towards the cell cortex, assembled by rings
containing -tubulin (-TuRC, composed of -TuSCs: -Tubulin Small Complexes)
anchored through the participation of several proteins (SAS-4 and pericentrin, above
all) which form scaffolds or docking platforms. MTs are also associated with several
proteins: kinesins and dyneins are two families of motor proteins that utilize MTs as
rail tracks to carry organelles and macromolecules respectively outwards and
inwards; MAPs (Microtubules Associated Proteins) are a family of tissue-specific
proteins with different activity: MT (de)stabilization, linkage to the cytoskeleton and
association to the cell membrane.
MTs are long tubes whose walls are made up of polarized heterodimers of α and β
tubulin, arranged in 13 polarized longitudinal parallel filaments, with a ”minus” end
fixed (in vivo) on a -TuRC and a growing “plus” end, centrifugally outwards
directed: in vivo, α-β tubulin heterodimers are added to the “plus” growing end; what
is the function of the -TuRC-linked “minus” end? The -TuRC is clearly an
anchoring base and an oriented platform that dictates the direction of the MT.
The centrosome and its centrioles are likely the most sophisticated biological
structures: nevertheless still now the centrosome is a “character in search of an
Author” to use the title of the comedy by Pirandello. What the centrosome’s role is
and how it functions, it is still unknown.

39
-------------------------------------------------------------------------------------------------------
Fig. 4 Centrosome theoretical geometrical model: orientation and labeling of -TuRCs.
There is an absolute univocal and unambiguous one-to-one correspondence between targeting signal, -
TuRC receptor, -TuRC orientation, MT direction, cell cortex compartment. Signal: undulating line
(A) linked to a “twisted-chain” protein; the signal, also named in the text “geometric address” or “ targeting
sequence”, is encoded in DNA and, after being transcribed or translated, is contained and displayed in the
polypeptide or in the polyribonucleotide (twisted tube) destined for a pre-established cortical localization and
recognized by the -TuRC receptor (undulating line at the base of the hemi-circle structure, the scaffold of
the -TuRC disk on the scaffold); different -TuRC receptors (B,C) do not fit in with the “undulating”
line/signal: they mark -TuRCs having orientations corresponding to different directions; orientation of the
-TuRC itself, from which an MT (D arrow) originates, with a precise direction, points to the desired cell
cortex location (E) (thus implicitly encoded in each signal/address). A kinesin carrier (wheel) transports the
complex centrifugally. -TuRC orientation, MT direction and cortical compartments are the molecular
“hardware” of the system whereas signals and -TuRC receptors represent the molecular “software”.

40
Fig. 5 Centrosome theoretical geometrical model: functioning.
Small ellipses represent γ-TuRCs on the centrosome (large sphere): each one is identified by its own private
receptor (capital letters: A, B, C) which recognizes only the corresponding targeting sequence. Each γ-TuRC
has the orientation of the plane which, in that point, is tangent to the centrosome “spherical” surface. MTs
(arrows) are nucleated with directions imposed by the orientation of the corresponding γ-TuRC: like
orientation, like direction (one “discrete” orientation, one “discrete” direction). So, a molecular complex
(twisted tube) through its “geometric” targeting sequence, recognizes and links exclusively the receptor (A or
B or C) that marks that γ-TuRC which has the correct orientation to nucleate a microtubule directed to the
desired (corresponding: A or B, or C) destination, reached through a kinesin carrier (wheel): one targeting
sequence, one γ-TuRC receptor, one cortical compartment: one-to-one univocal correspondence.
-------------------------------------------------------------------------------------------------------
“The strange case of ”… the centrosome cycle
Like Dr. Jekyll, centrioles have a double identity: daughter in a cell cycle, mother in
the next. In Ciliates new centrioles arise using a pre-existing centriole as a platform:
instead in Metazoa, firstly the DC changes identity and matures into a new MC, then

41
from both MCs (old and new) two new DCs arise. The “old” MC, after
disengagement, during S phase assembles a new DC orthogonally, whereas the “old”
Daughter, after moving from the proximal end of its “old” Mother toward the distal
end, disengages from its “old” Mother, though maintaining a connection with it, is
transformed in a new MC (losing its ribs and acquiring the distal and subdistal
appendages characteristic of the MC) and, only after its maturation from Mother to
Daughter, it too assembles a new DC. The two new centrosomes, each one containing
two orthogonal centrioles, one old and one just newly formed, also maintain a strict
connection during S phase and prophase. This is a very strange case and, perhaps,
unique: other cellular organelles, structures and components, before cell division, are
divided or produced de (ex) novo or built copying a template, but each one maintains
its identity, its role and its function; DNA filaments are separated by DNA
helicase/polymerase but the old filament 3’-5’ remains in the new cell a 3’-5’ filament
(leading strand), and so does the other 5’-3’ filament (lagging strand), each one
conserving its role and its leading or lagging identity. As we have seen, centrioles,
that can be formed de (ex) novo, normally are assembled by using only the MC as a
platform and the new centriole arises always at right angle in respect to the MC: why
do Metazoa form a new centrosome through such a complicated, unique and
expensive process? (Fig. 1 on page 13). Embryo’s development is a very risky
process, above all for those species that use external fertilization, because of
predators: cleavage divisions have to occur very fast: 37,000 divisions in 43hours in
frogs, 50,000 divisions in 12 hours in flies, one every ten minutes; nonetheless this
unique, complicated and expensive mode of centrosome duplication has been chosen
also by those Metazoa that use external fertilization. Why? “If conserved, it is
important”: an excellent reason and an important explanation must exist. A last
observation about the eccentricity and uniqueness of centriole duplication: the MC of
a cell, after division, is inherited by only one of the two new sister/daughter cells,
which, at its turn, will transmit this same MC to one of its daughter cells: so, in a
clone of cells (derived from the same common ancestor cell) the MC of the ancestor

42
survives (like a long-lived imperishable Phoenix) in only one cell of the group; the
age of each cell is the age of its MC: isn’t it very strange? In a sense it is possible to
affirm that every cell division is asymmetric: one cell remains a mother cell (that one
which conserves its old MC) whereas the other one is a real daughter cell with a new
MC: after each division there are not two sister cells, but a mother and a daughter
cell.
No centrosome? No gastrulation
“It is not birth, marriage or death, but gastrulation, which is truly the most important
time of your life” (Wolpert, 1986).
It is my personal conviction that this process of DC maturation (from Daughter to
Mother centriole) is evidence of an evolutionary event of exceptional importance: the
passage from already complex single-celled beings (many Protists show astonishing
complex shapes) or 2D multi-celled organisms to a complete 3D architecture (i.e. not
only simply laminar or tubular/cylindrical structures); this passage has deeply
involved centrioles and centrosomes. Multi-cellular organisms need coordinated and
common 3D polarity; gastrulation furnishes a clear example of 3D organization and
illustrates the fundamental difference of 3D architecture and dynamics in respect to
2D laminar processes: gastrulation requires a fine polarization of cell (differential
adhesion) and, above all an orderly orientation of the extracellular matrix fibers
which must be orientated according to common and shared directions: gastrulation
goes in parallel with the appearance of mesenchyme and mesoderm (and their
organized fibers), and, above all, with a well formed centrosome: “The
embryogenesis of freshwater planarians is equally intriguing: cleavage of the
fertilized egg was described as ‘anarchic’ by early developmental biologists. No overt
gastrulation or epiboly has been described in these embryos, yet they manage to
develop anteroposterior and dorsoventral axes without difficulty.” (Sànchez

43
Alvarado).
No centrosome? No gastrulation.
Lacking centrosomes, Planaria not only have lost the ancestral cleavage pattern, but
also the capability of performing gastrulation.
Shared and coordinated orientation of metazoan centrosomes
From facts and evidences (just now examined) to new hypotheses and theories.
“Centriole duplication is part of the mechanism by which the cytoskeleton of the
Daughter cell is patterned upon that of the mother” (Harold). The MC, before
disengagement, likely transmits to its old DC the information of orientation, and
physically orients its old DC in respect to the cytoskeleton; effectively also in Ciliates
the process of centriole duplication occurs at right angle and utilizes a pre-existing
centriole as a platform to orientate the arising centriole polarity in order to insert it
correctly in the complex cytoskeleton, something like a new trolley-bus (whose two
sprung trolley poles must be correctly connected to the two polarized electric wires)
is orientated and correctly (front-rear) positioned in respect to the “electric city-
skeleton” made of aerial-suspended wires; so the cells in a tissue become co-
ordinately polarized by co-ordinately oriented centrosomes: in Metazoa the
centrosome is the “intrinsic” (no external cues) reference system; plants, fixed in the
ground, use an “extrinsic” reference system (light and gravity), just as a compass uses
(extrinsic) Earth magnetism and a GPS utilizes (extrinsic) satellites: as an “extrinsic”
reference system is common to each receiver, similarly an “intrinsic” reference
system must be the same (identically oriented) in each cell; multicellular organisms
must possess a mechanism to transmit, share and co-ordinate their inside points of
reference (as we have seen, centrosomes increase cell polarity up to 45 poles) and this
function is performed through the orientation imprinted by the MC to its “old” DC
before disengagement, so that two co-oriented MCs build two co-oriented

44
centrosomes before cell division: so, all the cells of a tissue have the same points of
reference to correctly build complex 3D organs. This unique behaviour of
centrosomes supports the idea that their main role is the translation of the virtual
(DNA coded) cell geometry into an actual real cell wiring system. When a new
procentriole arises (orthogonally and near the MC) the cartwheel is formed: 9-fold
symmetry and chiral (Guichard, 2013) non-equivalence of triplets (fundamental, in
the geometrical model of centrosome functioning, for left-right patterning) are
established co-ordinately and corresponding with those of the mother centriole:
"Whereas the cartwheel is thought to nucleate the nine fold rotational symmetry of
the microtubular triplets the acorn [see at page 51] might play an equally important
role imposing rotational asymmetry on the microtubular triplets, perhaps leading to
the asymmetric assembly of basal-body-associated fibers and hence cellular
asymmetry in general" (Geimer and Melkonian (2004). [Looking for “centriole
architecture” through any web search engine, it is possible (and convenient) to have
an updated idea on this continuously developing matter]. So, in tissues, all the
centrosomes maintain a common, shared and coordinated orientation to operate as
molecular geometric 3D interfaces that translate morphogenetic signals into precise
locations in the cell and in the whole tissue (extra cellular matrix fibers): their
biochemical structure is the “hardware” and their “software” consists in “targeting
sequences” and “labelled” γ-TuRC receptors; unfortunately, the comparison between
computers and living organisms ends here: in living self-replicating cells, each newly
formed geometric tool (centriole and centrosome) must “tuned” every cell division.
As we have already seen, through the centrosome, DNA organizes the cell cortex by
fine-tuning its polarity (45 cortical non-equivalent compartments) that must be
transmitted (and shared) from cell to cell.
About the non-equivalence of the triplets in centriole/basal bodies in Ciliates

45
“Among flagellates, the appendages are also varied, biochemically and
morphologically; even microtubule appendages may have highly complex shapes as,
for example, in Physarum or in Ochromonas” (Beisson and Jerka-Dziadosz, 1999).
Non-equivalence of the triplets, their molecular-structural individuality capable of
distinguishing each other, is the main question to face.
Facts, above all: many studies on Protists have demonstrated that the nine triplets of
their centrioles/basal-bodies are different (not-equivalent) and arranged in an ordered
sequence: circumferential polarity in the arrangement of basal body triplets is
accorded with the disposition of the cytoskeleton. Ciliates contain only centrioles,
each at the base of a cilium or flagellum: they do not have centrosomes with two
orthogonal centrioles; orthogonality appears only during the assembly of a new
centriole, which arises and grows perpendicularly to a pre-existing one. The
biflagellate unicellular green alga Chlamydomonas reinhardtii has an ordered
location of its organelles: cilia, oral apparatus, nucleus, chloroplasts, pyrenoid,
eyespot, excretory vacuoles; this organization depends on the disposition of different
cytoskeletal fibers (four cruciform rootlets: two thick, made up of 4 MTs, and two
thin, made up of 2 MTs); the cell appears clearly polarized: an Apical-Basal axis from
the cilia to the pyrenoid, a Dorsal-Ventral axis orthogonal to the first, from the
nucleus to the oral apparatus; also a transverse axis of asymmetry (orthogonal to the
other two axes) is established because of the asymmetric position of the eyespot. In
Protists, centrioles organize the cytoskeleton (Feldman et al., 2007): structural
anomalies in centrioles cause disorders in the cytoskeleton (cruciform fibers lose their
normal composition or their orthogonal disposition). Chlamydomonas has two apical
flagella, whose movements are coordinated during planar 2D strokes, and during
conical-helical 3D rotations. The axoneme of its flagella is composed by the
canonical nine MTs doublets/blades (cilia and flagella have “doublets”, not triplets);
these blades are not equivalent: electron microscopy has allowed each one of the nine
doublets to be distinguished, each one showing its own morphological characteristic,
that identifies itself from the other, and its own fixed location relatively to the others;

46
the two central MTs of the axoneme are also different. Electron microscopy (Hoops
and Witman, 1983; Bui et al., 2009) has highlighted that the circumferential
asymmetry of the axoneme corresponds to (and is likely built by) to an even more
marked circumferential asymmetry of the basal body/centriole: it is possible to
distinguish each one of its nine triplets and their orderly sequenced arrangement,
determine the 180° rotation of one basal body compared to the second and observe
the connection between each triplet and particular fibers of the cytoskeleton: the
striated fibers of the “distal connector”, tie (or rather “connect”) and fasten the
triplets 9-1-2 of both basal bodies; the thick cruciform fibers are attached to the
triplets N° 3 and 4, the thin ones are linked to the triplet N° 8; the newly-forming
procentriol is always in front of the triplet N° 9. Geimer and Melkonian (2004) have
described an “acorn-like” structure in the inner distal part of the basal body, adhering
in a highly asymmetric manner to the triplets 2-1-9-8-7, and another structure, shaped
like the capital letter “V” in contact with the triplets 9, 5 and 4: "whereas the
cartwheel is thought to nucleate the nine fold rotational symmetry of the microtubular
triplets the acorn might play an equally important role imposing rotational
asymmetry on the microtubular triplets, perhaps leading to the asymmetric assembly
of basal-body-associated fibers and hence cellular asymmetry in general".
Thus, the process through which centrioles are built appears composed of two
different stages: first the 9-fold symmetry is firstly established (cartwheel) then
rotational polarity is imposed (acorn). By the way, there is a question with regard to
the cartwheel, which in Metazoa is formed only in the daughter centriole during its
building (“In vertebrate centrosomes, a cartwheel structure is present at the base of
procentrioles but is no longer seen in daughter and mother centrioles” Azimzadeh
and Marshall, 2010): if it were only a structural component of the centriole, it would
be present not only in the proximal portion of the nascent centriole and, in addition,
could not be absent in mature daughter and mother centrioles. It is likely the
cartwheel has, apart from its transient structural role in probasalbodies, other tasks:
establishment of the 9-fold symmetry and, perhaps, the arrangement of the molecular

47
platform for generating the non-equivalence of the 9-fold rotational symmetry; this
idea is supported by several measurements of the tilt angle of inward rotation of the
centriolar triplets blades (see “The centrosome” at page 40) that increases from the
proximal to the distal end: only the proximal region of the daughter centriole (where
the cartwheel is formed) is responsible for assembling the PCM around the DC, as we
will see later.
The shape of any structure is the consequence of its molecular composition: in C.
reinhardtii the polypeptide VFL1 coded by the gene vfl1 (Variable number of
FLagella), binds only to the triplet N° 1 (Silflow et al., 2001) confirming the
biochemical nature of the circumferential asymmetry.
Another Protist, Paramecium, has similar characteristics (Beisson and Jerka-
Dziasdosz, 1999): the high number of basal bodies (and cilia: about 4,000) is
accompanied by their ability to self-organize and to connect with each other in about
70 regular rows, always with the same orientation of the triplets (in order to beat with
coordination and synchronism), just using their circumferential asymmetry and
structural differences between the triplets; each new basal body arises at right angle
from an old one, then straightens up and rotates to acquire a precise and coordinated
orientation with its complex cytoskeleton: the triplet N° 9 links to the “postciliary
ribs”, the triplet N° 4 is attached to the “transverse ribbon”, the triplets N° 5-6-7 are
connected to the “kinetodesmal fibers”. This role of basal bodies in organizing the
arrangement of cortex and cytoskeleton agrees with the movements of the DC
described in many Metazoa (cortical cytasters: Salinas-Saavedra and Vargas, 2011)
and in Mammals (Piel et al., 2000), where it seems to organize in detail specific
peripheral cell sectors. Interventions of the centrioles/basal bodies in a complicated
cortical organization have also been described in Trypanosoma (Lacomble et al.,
2010), in which a characteristic rotation of the new DC around the MC has been
observed.
Chlamydomonas basal-bodies are also capable of regulating the length of the two
flagella: Rosenbaum and colleagues (1969) observed that, following partial surgical

48
ablation of one flagellum, the other is immediately reduced in length, then re-growth
occurs simultaneously in both flagella until they reach the normal length: basal-
bodies organize axonemal components, attentively regulating the activity of kinesins
and dyneins through a highly sophisticated mechanism, the Intra Flagellar Transport.
Summarizing, in protistan centrioles, built with 9-fold symmetry, each triplet shows
its own morphology, its fixed position and linkage to the cytoskeleton and even a
molecular individuality has been proved for the triplet N° 1; furthermore, centrioles
are univocally oriented in the cytoplasm, in respect to the cytoskeleton: all of the
theoretical requirements of a “biological” protractor are satisfied. And in Metazoa?
V.I.P. Very Important Proceedings
Two studies have opened a new scenario for the centrosome: (1) the first (Dodgson,
2013) faces the problem of cell cortex compartmentalization that allows cells to
achieve a more detailed polarity (indeed Anterior-Posterior and Dorsal-Ventral axes
are too few to build complex organs); interestingly, the compartmentalization of the
centrosome surface (45 small areae) quantitatively corresponds to the cortical
compartmentalization shown by this study (one-to-one correspondence between
centrosomal and cortical compartments); (2) the second (Blower, 2013) highlights the
role of the centrosome in binding and delivering mRNAs into defined cytoplasmic
localization.
These works introduce the idea of a centrosome that operates like a mail-sorting
machine: zip-codes (alpha-numeric characters not able to be recognised by the optical
devices of the machine) are firstly read by a scanner and transformed into barcodes
(stripes of vertical bars printed onto the envelopes) able to trigger the devices that
control the entrance in the correct lane. DNA (in exons, or in introns or in other part
of the sequences that flank a gene) stores the code for the cortical position where an
mRNA or a protein (transmembrane receptors, polarity and adhesion factors above

49
all) must be located: this code is transcribed into RNA (3’ UTR, miRNAs?), which
can match and accompany mRNAs and proteins, or can be translated into the protein
itself (N-terminus targeting sequences/ligands) so that the final 3D shape of each
code is easily recognized by -TuRC receptors (see later); for instance nanos and
bicoid mRNAs are localized in the fly oocyte cytoplasm through MT transport,
driven by their 3’UTR sequence. Let’s then read these findings:
1) “The polarity machinery of fission yeast is arranged into cortical clusters that are
dynamic and persistent…This suggests that the interaction between different polarity
factor species may be an actively regulated feature, and not simply occur as a default
when polarity factors reside within a same general domain at the cell cortex. Future
work should aim to clarify how the various polarity factors are targeted to different
clusters. This may be accomplished through post-translational modification of certain
polarity factors through competitive binding interactions. Thus, cortical
compartmentalization of proteins into discrete clustered domains may be a general
feature exploited by cells to control spatiotemporal signaling events with high
fidelity…Cortical compartmentalization of proteins into discrete clustered domains
may be a general feature exploited by cells to control spatiotemporal signalling
events with high fidelity. As we observe similar polarity factor clusters in
Saccharomyces cerevisiae and Caenorhabditis elegans cells, the clustered
organization of polarity factors within cortical subdomains and regulated cluster
segregation may be key regulatory features across many organisms. In the case of cell
polarity, cortical nodes and their spatial segregation could contribute to robust ‘on’
or ‘off ’ switching of polarity at the right place and time in cells, following intra- or
extracellular signals…This suggested that localized microtubule delivery (of polarity
factor Tea1) and/or oligomerization could suffice for the formation and maintenance
of cortical nodes of both Tea1 and its interactor Mod5…A quantitation of node
number in Tea1-GFP, Tea3-GFP, Tea4-GFP and GFP-Mod5 cells showed that a
typical cell end consistently organizes 20–30 nodes of each protein dispersed along
the cell cortex” (Dodgson et al.).

50
2) “Localization of mRNAs to specific destinations within a cell or an embryo is
important for local control of protein expression. mRNA localization is well-known to
function in very large and polarized cells such as neurons, and to facilitate embryonic
patterning during early development. However, recent genome-wide studies have
revealed that mRNA localization is more widely utilized than previously thought to
control gene expression. Not only can transcripts be localized asymmetrically within
the cytoplasm, they are often also localized to symmetrically-distributed organelles.
Recent genetic, cytological, and biochemical studies have begun to provide molecular
insight into how cells select RNAs for transport, move them to specific destinations,
and control their translation…One of the major types of mRNA transport in
Drosophila embryos is the movement of mRNAs involved in developmental pattern
formation to the apical centrosome. Movement of these mRNAs occurs in a dynein-
dependent manner and requires Egl and BicD for transport. However, little was
known about the mechanism of RNA anchoring at the centrosome... Microtubules are
used for the majority of long-distance transport within cells. Microtubules are
important for the localization of the majority of known localized mRNAs (with the
notable exception of localized mRNAs in Saccharomyces cerevisiae, which require
actin-based transport) through the action of various motor proteins. However, several
studies suggest that microtubules are not just a highway for transporting mRNAs, but
can also serve a destination for some localized transcripts. Purification of
microtubules from a variety of organisms (sea urchin, frog, human) has demonstrated
that hundreds of mRNAs co-purify with microtubules. The majority of microtubule-
localized mRNAs code for proteins that appear to function in cell cycle and
microtubule-related processes. Our group recently tested this idea directly by
knocking down the mRNAs for 10 uncharacterized microtubule-localized transcripts
in HeLa cells. We found that the majority of these mRNAs coded for proteins required
for proper microtubule organization during both interphase and mitosis. Work in
Xenopus oocytes has also found that both spindle localization and translational
control of a subset of mRNAs are important for completion of both meiotic divisions,

51
highlighting the link between mRNA localization and translational control. In
addition to containing mRNAs that control the microtubule cytoskeleton,
microtubules also contain a number of transcripts that appear to be translationally
repressed and are likely to be passive cargo on the spindle rather than active
participants in controlling microtubule-related events. This idea is consistent with
recent work looking at RNA localization in early cell divisions of the snail Ilyanassa
obsoleta. In this work, the authors used RNA FISH to examine several mRNAs
important for early development. Some of these transcripts show an unequal
distribution during cell division by selectively associating with one of the two
centrosomes of a dividing blastomere. This suggests that unequal segregation of
important developmental mRNAs by the mitotic spindle could be an important
mechanism for the establishment of early patterning asymmetries. While many
mRNAs are known to localize to meiotic and mitotic microtubules little is known
about the sequences or proteins involved in this process” (Blower).
Facts and evidences, these, from which hypotheses can be formulated: Dodgson
(control his schematic in his article, free on the Internet) models the process of
cortical compartmentalization in such a manner that agrees completely with our
images (Fig. 4 and 5). From RNA localization to centrosome and its alternative
differential delivering it is plausible to formulate the hypothesis of the geometrical
role of the centrosome as a biological interface.
It is convenient, now, to review other evidence.
“While centrioles are dispensable for spindle assembly, they are more important for
spindle positioning. When centrioles are experimentally ablated, spindles drift within
the cell” (Azimzadeh and Marshall, 2010).
In Drosophila embryos all syncytial mitotic divisions are not randomly oriented, but
strictly controlled by the centrosomes that remain linked to their own nuclei (if
centrosomes are experimentally damaged, development stops immediately): the first
4 divisions generate nuclei that remain radially equidistant from the center and form a
sphere; then, during cycles 4-6, the division planes are oriented in such a manner that

52
nuclei distance themselves along the AP axis, transforming the sphere into an
ellipsoid whose marginal nuclei are symmetrically equidistant from the cortex.
During the stages 7-10 a characteristic symmetric migration towards the cortex
occurs; Baker and colleagues (1993) have proposed that "cortical migration is driven
by microtubule-dependent forces that repel adjacent nuclei". A network of
interdigitating microtubules (like the polar MTs that in metaphase connect the two
opposite spindle poles, overlapping their oppositely directed “+” ends) forms between
yolk cells centrosomes (not-migrating) and peripheral centrosomes (migrating): this
geometric network of MTs carries nuclei to the cortex while their final ordered
position appears to be due to astral MT population (unlikely polar MTs, astral MTs
with reversely directed polarity do not overlap: boundary MTs of different asters go
parallel with their plus ends pointing in the same direction). "In Drosophila the
movements of nuclei to the embryo cortex are mediated by forces acting on the
centrosomes rather than on the nucleus itself. Asters are presumably the main target
of such forces. It is then conceivable that MTs, nucleated on either side of the
centrosome or which display different characteristics, are nucleated under the
influence of opposite sides of the centriolar shaft, just as different appendages arise
from basal bodies. The situation in S. cerevisiae gives some support to the idea: the
spindle pole body, functional equivalent of the centrosome, displays a marked
structural and functional bipolarity with an intranuclear spindle and an aster of
cytoplasmic microtubules. Like in Metazoa, defective astral microtubules lead to
defective nuclear positioning and defective budding. The biochemical and
physiological differences between the two microtubule arrays are already well
documented. Different gamma-tubulin binding complexes interacting with the inner
or outer plate respectively, are involved in the nucleation of the two microtubule
arrays” (Beisson and Jerka-Dziadosz, 1999). The ordered separation of nuclei, the
controlled asymmetry of their arrangement, in anterior-posterior direction, and their
division into two cortical halves (Right and Left) have been highlighted in a study
based on the differential spatio-temporal beginning and speed of mitosis N° 14,

53
defining domains in which cells start to divide simultaneously (Foe, 1989); 25 left
and 25 right domains with different form and extent are evident: "all the domains
described also occur as pairs. Whether paired or not, every domain is bilaterally
symmetric." A clear midline is evident both dorsally and ventrally; domains show an
interesting link between centrosomal and cleavage geometry (very surprising are the
images by Foe, 1989, in: “Mitotic domains reveal early commitment of cells in
Drosophila embryos”, freely available on the Internet). Strikingly, domains appear
arranged according to the theoretical model of the centrosome in longitudinal
wedges, one on the midline, four “left” or “Occidental, Western” (so to say, if one
prefers geographical terms) and four “right” or “Oriental, Eastern”, intersecting two
polar caps and three parallel rings; it appears that the cortex of the embryo is mapped
and compartmentalized by the zygote centrosome: both maps (theoretical centrosome
compartmentalization showed in Fig. 3 on page 37 and fruit fly embryo cortex at 14th
mitosis) coincide; as we have seen, through the centrosome DNA “invents a
spherically organized space”, creates a 3D design upon an equal, homogenous,
equivalent and nameless cortical surface and draws a labeled grid line: a featureless
and unidentified cell cortex, mapped and wired by the centrosome aster, acquires a
useful labeled compartmentalization: it becomes polarized in detail.
From these studies an extraordinary and surprising correspondence emerges with the
theoretical geometrical model of centrosome functioning, about both the centrosome
“hardware” (spherical compartmentalization and one-to-one correspondence between
centrosomal and cortical compartments, and therefore 3D inclination of γ-TuRC
scaffolds) and the centrosome “software” (different centrosomal receptors, differently
located on the centrosome to bind different ligands for differently delivering them in
the cytoplasm).
Morphogenesis and thermal fluctuations in the cell environment

54
“We hypothesize that centrosome loss occurred concomitantly with the loss of the
spiral cleavage and oriented cell divisions in the ancestor of planarians and
schistosomes….It is remarkable that the loss of such a conserved organelle as the
centrosome occurred within not-parasitic flatworms, as cellular and developmental
processes appear largely conserved between these species. A significant difference
can be found in the mode of embryonic cleavage however. Macrostomum retained the
ancestral spiral cleavage, also found in annelids and molluscs, which relies on a
stereotypical pattern of cell division orientation. In contrast, planarian and
schistosomes embryos undergo divergent modes of embryonic cleavage, which
apparently do not involve oriented cell divisions” (Azimzadeh et al., 2012). As we
have seen on page 46, centrosome loss occurred concomitantly with the loss of
gastrulation too. To create the correct “stereotypical pattern of cell division
orientation”, evidently the species-specific instructions for positioning the spindle
poles in each cell division are encoded and stored in DNA: in effect we have already
seen that only one gene is responsible for left- or right-handed spiralian cleavage. The
centrosome receives any recognizable signal from DNA and, through its MTs,
nucleated by properly oriented γ-TuRCs, positions the poles of the spindle in such a
way that cleavage is, as forecast by DNA, radial, spiral, bilateral, syncytial, discoidal
or rotational. During cleavage, the size of blastomeres can vary in a continuous
fashion (starvation, abundance of nutrients, temperature) but the angle between two
subsequent division planes is precisely and discretely fixed, always constant for each
species and not continuously distributed in an interval of values: to give an example,
the angle between the planes of two successive early meridian divisions in sea urchin
embryos (from 2 to 4 cells, and continuing to 60 cells) does not vary around a main
value, it always has the same identical precise value, whatever the size is of the
zygote; after the surgical separation of the two first blastomeres of a sea urchin or a
frog embryo, two complete, but smaller (about half-size) embryos develop,
nonetheless the plane of each cleavage division occurs following and maintaining the
same precise stereotyped pattern (orientation), characteristic of the species: size

55
changes, directions do not, like vectors; and so it is during the first stages of organ
development. It seems that metazoan cells possess a “discrete” instrument able to
precisely read angle values from DNA instructions (specie specific, then genetic,
pattern of cleavage division orientations) and realize 3D, bilaterally symmetric when
necessary, angles (spindle pole positioning) having the programmed value: in
addition, angle amplitude appears to be chosen in a limited number of discrete values;
consider for example left and right semicircular canals and their striking directional
precision: the macroscopic result (high spatial precision of anatomical structures)
descends from the addition of many and many microscopic arrangement of cells,
developmental microprograms or modules which organize the assembly of a few
cells with an extraordinary level of precision (as in cleavage); the addition of many
different modules is necessarily affected and conditioned by different factors (e.g.
diverse cell size due to random localized abundance/deficiency of nutrients). Indeed
about the word “precision” a comment is necessary: 45 cell cortex compartments are
much more than 10 thousand dots per inch (dpi) that is a very high graphic precision;
however the surface of each compartment (a square with a side of 2.6 μm) is
enormous if compared to the dimensions of a protein (the average diameter size is
about 3-6 nm); so in metazoan organisms, composed of thousands billion cells (1014
),
we can see high precision of shapes but not absolute identity; Metazoa utilizes that
level of precision which necessity and sufficiency require to achieve high quality
stereotyped reproducibility, but not identity.
Let’s come back to the work of Dodgson and colleagues: “The establishment of
domains of polarity factors occupies and confers different identity and function to
restricted areas of the cell cortex”: like the centrosome surface, the cell cortex too
appears to be compartmentalized into non-equivalent distinguishable areas.
...“Complexes are delivered to cell ends by growing microtubule tips”: one-to-one
correspondence between corresponding centrosome and cortical compartments is
assured by MTs originated from oriented centrosomal γ-TuRCs.
...“Intriguingly, nodes did not appear to correspond to similarly distributed lipid

56
microdomains...,Thus, oligomerization can trigger cluster formation and enable
cortical enrichment of polarity factors in cells”: the large dimension of the cortical
compartments with respect to the protein diameters is compensated for by self-
oligomerization (self-assembly of homo- and hetero-monomers).
...“What could be the physiological advantages of spatially confining polarity
regulators to nodes? One possibility is that nodes could provide a means for cells to
control polarity at the cortex in space or in time, by regulating or preventing polarity
factor interaction, or by segregating certain node populations from others....In
summary, the polarity machinery of fission yeast is arranged into cortical clusters
that are dynamic and persistent, and may be formed by oligomerization (direct or
mediated by an interacting partner). Many polarity factors can be found in
neighbouring but separate cluster populations, with this spatial segregation
potentially key to their function. Although the clusters are dynamic, they are also
persistent and appear relatively immobile, and it is not yet clear what the functional
significance of that persistence/immobility might be. One possibility is that it may
allow polarity factors to maintain a stable domain of localization and function.
Alternatively, it may ensure that certain polarity factor clusters only interact with
others at specific locations and times within the cell, as exemplified by Tea1 and
Tea3. This suggests that the interaction between different polarity factor species may
be an actively regulated feature, and not simply occur as a default when polarity
factors reside within a same general domain at the cell cortex”. Fine tuned polarity is
absolutely necessary in Metazoa: an accurate and careful control of polarity at the
cortex in space and in time is the most important characteristic of differential
adhesions, indispensable in convergence and extension, epiboly, invagination.
The “software”

57
The idea about the spatio-temporal dynamics of cortical compartments can explain
another property of the centrosome: we must not think of cells like globes mapped
once and for all: on the contrary the centrosome (which really is compartmentalized
once and for all), in different cells and in different phases of the cell cycle and life,
can assume different (controlled, never random) positions and, through it, DNA can
“design” a newly oriented cortical grid line (e.g. in migrating cells that respond to
differently localized chemoattractants); when the centrosome is near the center of the
cell, one of its 45 compartments nucleates an MT directed toward a particular cortex
area (e.g. that one where one spindle pole must be positioned in order to dispose the
division plane with a desired angle with respect to the proximal-distal axis of a
developing limb); differently, when the centrosome is near the cell membrane, for
reaching the same cortical area, another γ-TuRC must be chosen, that one which, in
this different configuration, is properly oriented. The centrosome can change its own
orientation (not randomly but under control) the orientation of the cytoskeleton and
the global polarity of the whole cortex (like in migrating cells) but cells always know
their orientation and the position of the centrosome; in DNA the cell map and its
relative virtual grid line, that DNA “physically and really” draws in the cell through
the centrosome, is stored and coded: DNA chooses, step by step, the most convenient
orientation; on this data, during evolution, new geometrical steps (targeting sequences
calculated in accord with the position of the centrosome) have been added to
developmental programs in order to build more and more complex organs and
organisms. Step by step, stage by stage, new targeting sequences, appropriate to the
spatio-temporal position of the centrosome, have been added (“approved” by
purifying selection) to developmental programs in such a way that cells are correctly
guided and positioned to realize very complex organs. Targeting sequences are not
“absolute” (like North, West, etc.) but relative to the centrosome (known) position
and cortex compartmentalization (known) orientation: to reach an area with a given
angle respectively to the Dorsal-Ventral axis and a given θ angle respective to the
plane containing the Dorsal-Ventral and Anterior-Posterior axes, in a developing

58
embryo, different targeting sequences are chosen (and have been selected during
evolution) from time to time, depending on the position of the centrosome and, as a
consequence, on the γ-TuRC (labelled once and for all by its receptors) that is
properly oriented to nucleate an MT correctly directed. Something similar occurs in
GPS devices: when we turn, the map on the screen rotates in order to show the actual
position of “forwards” or “straight on”; our “straight on” direction is not “absolute”
and always the same and does not point towards a constant direction, but its
orientation depends on the position and orientation of our car. Topogenic sequences
do not indicate a fixed position of the cell cortex, but one fixed centrosome
compartment; the choice of one particular topogenic sequence is relative to the
centrosome location.
The language used by metazoan systems for their morphogenetic programs must be
made of targeting sequences and written with some basic rules: the correspondence
between targeting sequence and γ-TuRC receptors must be invariable (constant and
static) while the correspondence between γ-TuRC receptors and cortical
compartments must be univocal (one-to-one) but time-variable: cortical
compartmentalization too is dynamic, variable, nonetheless always under control,
never random. The one-to-one correspondence between targeting signals and γ-TuRC
receptors is absolute, constant and invariable, whereas the one-to-one
correspondence between targeting signals (and related γ-TuRC receptors) and cortical
compartments is strictly univocal but time-variable (it is then a “relative” concept):
by moving (off-centering) the aster, the correspondence between compartments
remains one-to-one (from a given centrosome compartment it is possible to arrive
univocally into only one cortical compartment) but the same centrosome
compartment can correspond, from time to time, to different cortical compartments,
and vice-versa, depending on the controlled position of the centrosome. Off-centering
of centrosomes explains why, although centriole geometry is based on the 40°angle,
the first meridian division planes in sea urchin embryos occur at 90° angle and not at
80° or 120° as expected if the centrosome in the cell maintains a central position.

59
This fits in very well with an evolutionary model of “genetics of morphology”:
purifying selection has firstly approved the choice of a particular targeting sequence
added, for instance, to polarity or adherence factors only if the consequences were not
dangerous, then, through natural selection, the fittest shapes have established.
Evolutionary morphology is a tale of targeting sequences, added, changed, erased, in
order to test and evaluate new spatial disposition of cells; thus genetics of
morphology is the genetics of targeting sequences.
Growth is a highly anisotropic process, in which polarity and direction are always
regulated by precise forecast programs. Identical objects (matches or sticks for
instance) can compose many different geometric plane figures (perimeters) when they
are assembled following each other, each one oriented with a particular tilt angle; if
only the right angle is permitted (and its useful multiples: 0°; 90°; 180°; 270°; four
degrees of freedom) it is easily possible, for example, to compose all the Arabic
numerals (as in digital displays); if only the 45° angle is allowed, (and its multiples:
0°; 45°; 90°; 135°; etc.; eight degrees of freedom) all the capital letters of the English
alphabet can be easily composed. Fibroblasts can control the disposition and
orientation of the collagen fibers they produce, making fasciae of different shapes
that force the homogenous muscular tissue to take the form characteristic and typical
of each skeletal muscle. It is worth recalling the work of the mathematician biologist
D'Arcy Wentworth Thompson: in his famous “On growth and form”, written many
years ago, in 1917, he wrote a chapter -“The comparison of related forms”- to explain
the importance of mathematical transformations of forms in related animals: physical,
mathematical and mechanical laws are the basic determinant of the form; between the
mathematical transformations he examined, “share-mapping” is particularly
intriguing, because it is generated by changing the 2D angle between axes (rectangle
> parallelogram, for example): applied to the shape of organisms, through 2D “share-
mapping” only one of the two coordinates of each landmark (landmark-based
geometric morphometrics) changes and this sustains the difference of forms.
Thompson’s ideas did not have biological and molecular bases; the geometric role of

60
the centrosome can fill this gap: new species can arise because targeting sequences
have been added, changed, erased and new coded values for growth vector directions
have been performed.
Morphogenesis: a tale of angles
Growth velocity vectors are well defined for each point and stage of a developing
organism: vector module (the intensity of growth, or growth “speed”, which can be
considered the “scalar” component of growth “velocity”), can vary around a main µ
value (Gaussian distribution) under the control of many agents, from genetic
programs to growth factors and hormones. On the contrary, growth vector orientation
is always precisely predetermined and fixed so that adult organisms and organs can
reach different size but have always the same characteristic shape (“geometric
similarity”: left and right mammalian limbs, cochleae and eyes are good examples).
Size can be different in two different individuals (in each species the expected or
main value and the standard deviation are defined for each organ) but only very small
variations, and only in the last stage of development, are permitted in the single
(healthy) individual whose pair symmetric organs are highly similar, about identical:
the dimensions (size) of limbs, fingers, ears are always the same in the left and right
half of the same individual whereas the Darwin’s tubercle, on the helix of the
auricular pinna, can be different in the two ears because it arises at the last stage of
pinna formation. A fundamental morphogenetic rule in Metazoa: scalar quantities,
subject to many controllers, can vary continuously around a species-specific
“Gaussian” main µ value (with a typical standard deviation σ value for each process
of organ morphogenesis), whereas vector orientations are precisely predetermined,
chosen in a limited number of values, in each species and for each stage and point of
the developing organism and their very subtle variations are only due to thermal
fluctuations.

61
Illacme plenipes has 750 bilaterally symmetric legs
Illacme plenipes (Myriapoda) has up to 750 legs all arising from a fixed point of the
cylindrical body circumference and all with the same (bilaterally symmetric)
orientation of the proximal-distal axis; if the angle between the proximal-distal and
the anterior-posterior axis were continuously variable about a main value, we would
see 750 legs not parallel, but in Illacme plenipes the legs of the same side are
absolutely parallel, in each individual; two individuals are substantially identical: the
number of legs can vary, like size, around a main value, but the tilt of the proximal-
distal axis in respect to the anterior-posterior axis is always invariable. Different
organs have their own spatio-temporal values for the magnitude (“scalar” component)
of growth in an interval of permitted species-specific values (accordingly with the
species-specific standard deviation): growth magnitude can be positive, null (no
growth) or even negative (apoptosis) and is independent from the intensity of other
organs: as is well known, growth occurs through the addition of many interacting
developmental elementary microprograms or modules (something similar to the
microprogramming in computer architecture), like morphogenetic fields and imaginal
discs: few cells in a restricted space; as a consequence, when the final (macroscopic)
result depends on the interaction of several (microscopic) modules, external forces
can influence the correct angles imposed by growth programs. As we have seen,
microscopic programs are extremely precise, however macroscopic results can show
differences: the more complex (larger and bigger) organisms are, the more variable
the ultimate results become. Large organs and big organisms cal mislead us showing
forms that vary in size and in angle too: if we look at cleavage (from 1 to 16 cells) of
a frog embryo, we can see (control on the Internet) a sequence of precise angles
between two successive division planes; but cells are never identical in size (they are
subject to the action of growth factors whose local concentration is, in turn, subject to
thermal fluctuations and then variable) and therefore can distort the final shape (think

62
to a cube with only one face a little larger than others). In limbs, the bones are the
leading structures and do not compete with others; differently, teeth can be
programmed with a size that does not fit in well with the size of facial bones: so teeth
can be too crowded or too sparse in respect to jaw’s or maxilla’s size and each tooth
can be subject to external forces (by other close teeth) that perturb the original
orientation of growth velocity vectors: variations of directional growth angles are
imposed by the concurrence of external cues.
Which molecules and which organelles are involved in directional architectural
management?
A surprising aspect of development is how cells are capable of forming
morphologically different organs and organisms (millions diverse species, from
worms to humans) by using (often for different purposes) the same (and highly
conserved) few signaling pathways (Wnt, Notch, TGF-β, Hedgehog, etc.),
morphogens (retinoic acid; bicoid and nanos mRNAs etc.) and structural molecules
(tubulin, actin, etc.). Why is it possible to realize so many and so different shapes
through the same molecular gradients? Where is direction-information coded?
The only “driving motor” that moves molecules in solution is thermal fluctuation
(Brownian motion) and the only “helm” that directs molecules in different directions
is the result of enormous numbers of random collisions between molecules that allow
a molecule to find sooner or later the correct orientation and direction: the
randomness is compensated for by the frequency of collisions, the speed of molecules
and the mean free path. Let’s recall some values calculated in gases at ambient
temperature and pressure: the average time between two collisions is of the order of
10-10
s, therefore 10 billion collisions per second; the mean free path between two
consecutive collisions is of the order of 10-8
m, or 10 nm; the average speed has
values of 1000 (103) km/h, higher than the speed of sound in air. Such high numbers
make it possible for each molecule to be quickly directed in the desired direction but
without any predictable time precision: we must consider that the Gaussian
continuous distribution of values in systems based on solutions and concentrations

63
does not agree with the precise 3D order necessary for biological structures that must
be stereotypically reproduced: e.g. the orientation and direction of extracellular
matrix fibers (cornea), or limb and organ shape (inner ear: malleus, incus, stapes,
cochlea, semicircular canals, utricle, saccule); only one out of infinite possible angles
is sought and, to do so, other types of arrangement and order are necessary: enormous
numbers of very small cells always obtain the same architectural result, laying
themselves out correctly to compose very complex organs whose size is
incomparable with cell size. “The spatial resolution of gradients based on diffusible
molecules has fundamental limitations set by the ability of receptors to discriminate
small differences in ligand concentration, such that sensitivity to concentration
changes in one part of the gradient comes at the cost of saturation in the rest of the
gradient. Such considerations lead to the idea that global positional information
might provide at most a low-resolution map of position within the cell that must then
be refined by local determination of structure and organization” (Marshall, 2011). In
addition to these limits we must also wonder how receptors can evaluate directional
information, if any, of ligand concentrations. Two similar geometric figures (a 2D
polygon or a 3D polyhedron) can show different dimensions of sides, areas and
volumes (scaling), but do not change their (plane or dihedral) angles (homothety)
indispensable to maintain their similar shape: by changing the angles, a square is no
longer a square but is transformed into a rhombus, like a rectangle becomes a
parallelogram. From the proposed geometrical model of the centrosome, a cell wired
by the aster emerges, mapped (really and physically compartmentalized under the
direction of the virtual grid line coded and stored in DNA) and capable of precisely
carrying 3D developmental programs out, in which the centrosome plays the role of a
geometric interface between (discrete) address codes, encoded and memorized in
DNA, similar to postcodes, and the relative spatial (also discrete) localizations like
the cortical compartmentalization of proteins into discrete clustered domains. It is
thus possible to obtain the assembly (controlled by DNA) of large and extended
structures and systems (cells, tissues and organs) with very high levels of order and

64
precision, highly stereotypically reproducible in organisms composed of hundreds of
thousands of billions of cells (1014
), bypassing the limits imposed by the
thermodynamics of solutions.
About “precision” and a biological “anti-aliasing” theoretical filter
Centrioles/protractors/goniometers are instruments equipped with discrete, non-
continuous, polarity (with nine degrees of freedom) that limits, but at the same time
makes precise and reproducible, the controlled execution of morphogenetic programs
(oriented cell division, tube morphogenesis, migration, extracellular fibers
disposition, convergence and extension, adhesion, etc.). One-to-one correspondence
between centrosomal and cortical compartments assures the precision of
reproducibility of forms: let’s recall Albrecht-Buehler’s words: “Several principles of
construction of a microscopically small device for locating the directions of signal
sources in microscopic dimensions: it appears that the simplest and smallest device
that is compatible with the scrambling influence of thermal fluctuations as are
demonstrated by Brownian motion is a pair of cylinders oriented at right angles to
each other”: in metazoan organisms we see high precision in reproducing shapes and
executing stereotyped oriented processes but not identity, because thermal
fluctuations cannot be eliminated. To understand how cells control angles and
directions we can observe only those developmental microprograms carried out by a
few cells: cleavage, gastrulation and neurulation furnish clear geometrical
information. Indeed the 9-fold symmetry of the centriole, taken together with the cell
size and the numbers of cells necessary to build metazoan organisms, seems to be the
best choice for building stereotyped organs and organisms (then successful,
controlled and purified by evolutionary selection); if the symmetry of centrioles had
an order of magnitude more than 9, thermal fluctuations (MTs are stiff and bending-
resistant but very long in a viscous cytoplasm crowded with organelles and

65
membranes) would cause overlapping of cortical domains (considered as the area in
which an MT subject to thermal fluctuations can arrive and “land”): during evolution,
“experiments” have been performed (but without success) as showed by Riparbelli
and colleagues (2009) who found in Acerentomon microrhinus centrioles composed
of 14 doublet microtubule blades, used to form cilia and flagella or mitotic and
meiotic spindles; the 9-fold symmetry has been chosen because it is, so to say, the
cellular “anti-aliasing” filter (to borrow something from the well known Nyquist–
Shannon Information Theory sampling theorem) to avoid overlapping (and disorder)
of cortical compartments, and, at the same time, the best choice to obtain the best
topological and morphological reproducible results: maximum output with minimum
effort.
As I have said, the objective and the purpose of development is to realize very large
organs and organisms making use of very small units (cells): the addition of
thousands of little errors, due to continuously variable values of directional
parameters, in each stage of growth, is incompatible with the realization of high-
quality architecture.
We move from a continuous distribution of degrees of freedom, subject to gradient
levels, to a discrete, precisely programmable distribution, limited but precise and
constant for each step, in the execution of a program and therefore resistant to the
“background noise”; thermal fluctuations are therefore restricted and limited around a
fixed (discrete) number of degrees of freedom: subject to thermal fluctuations, the
MT nucleated by a γ-TuRC will not reach precisely the center of the corresponding
cortical domain but it will arrive, like an arrow directed at a target, in a point that is
surely inside the compartment; to better explain this concept it is convenient to recall
the atom model by Bohr (omitting the successive considerations of Quantum
Mechanics): electrons orbit around the proton with canonical periods into different
shells depending on their energy, but they are only permitted to have discrete values
of energy, so they can stay on one delimited shell or on another; similarly, the MT
“landing” point onto the cortex fluctuates around the center of a compartment in 2D

66
annuli or rings of different probability densities (higher near the center) but only one
(discrete) cortical compartment is reachable from one centrosomal compartment
(one-to-one univocal correspondence, although spatio-temporally variable, as we
have seen before); the Gaussian distribution consequent to thermal fluctuations
appears restricted and limited to a small area around the center of the cortical
compartment like a small rifle target: the cell cortex is then composed of several
discrete neighbouring non-overlapping “rifle targets“ (cortical compartments), each
one connected to only one centrosomal compartment (one-to-one discrete
correspondence). In organisms composed of 1014
cells, stochastic processes
algebraically sum variations due to thermal fluctuations so that size varies
continuously but inside an interval; not so forms: a thumb is a thumb and an index
finger is an index finger; a limited “discrete” one-to-one correspondence between
discretely orientated centrosomal compartments and discrete cortical compartments
restricts and limits the effects of thermal fluctuations. Organs are composed of very
large numbers of cells, organized in different tissues: the ultimate result is the sum of
many, many sub-programs or elementary micro-programs so that forms could appear
different; indeed the previous considerations about homothety must be referred only
to elementary programs: the high precision of the control of 3D angles during the
stereotypical pattern of orientation of cleavage division planes furnish a glaring
example of simple basic micro-programs: cleavage is a typical microprogram
whereas gastrulation and neurulation, although simple, are composed of some
elementary programs; organogenesis is the sum of a lot of microprograms. This is a
clearer explanation of the concepts of continuous, controlled distribution of
probability density, high precision and identity. The theoretical definition of “discrete
cortical compartment” is: “the cortical area where MTs arrive, subject to thermal
fluctuations, started from the corresponding centrosomal compartment”. To borrow
something from Quantum Physics, cell cortex polarity is “quantizated”: its
“quantum” could be obtained (only when the centrosome is well-centered in the cell)
by multiplying the squared cell radius by π/45 (0.07 might be the “cell polarity

67
constant”), but the frequent off-centering of the centrosome changes its value. More
than nine triplets imply more than 45 centrosomal and cortical compartments, but
thermal fluctuations overlap the cortical areas where MTs arrive started from the
corresponding centrosomal compartment”, destroying the one-to-one correspondence
(aliasing). I am tempted to add a few words to Albrecht-Buehler’s cited sentence: “It
appears that the simplest and smallest device that is compatible with the scrambling
influence of thermal fluctuations as are demonstrated by Brownian motion is a pair of
9-fold-discrete-symmetry-cylinder oriented at right angles to each other”. To find
solutions for fighting entropy and limiting the disorder caused by thermal fluctuations
is the fundamental extraordinary ability of living beings.
Precision and constant reproducibility, particularly in very small confined spaces like
cells, are reached with difficulty by chemical concentrations (solutions constitute
typical examples of continuous distributions of density rates: where and when they
exceed a threshold value is unpredictable with high precision and out of a 3D
directional control: uncertainty and “noise” are inevitable; “The spatial resolution of
gradients based on diffusible molecules has fundamental limitations” Marshall,
2011); in addition, chemical concentrations, substantially isotropic, are not useful for
transferring or transmitting the angle information of 3D highly anisotropic growth
processes (compare bicoid, nanos, hunchback and caudal mRNAs gradients in
Drosophila embryos to the MT cytoskeleton that can discretely and “precisely” wire
small cells: the firsts are one-dimensionally controlled, whereas a labelled
cytoskeleton is three-dimensional). “Regulated spatial segregation of polarity factor
clusters provides a means to spatio-temporally control cell polarity at the cell cortex”
(Dodgson et al., 2013)
The gradient of a scalar field (density) is a vector pointing in the direction of the
greatest rate of increase: its components are the first partial derivatives (x,y,z) of the
field; supposing that cells could precisely control the rate of increase along a
direction (being able to create a 3D directionally oriented increase of concentrations,
read a 3D directional increase, 3D directionally respond and react), then cells must be

68
able to manage x, y, z coordinates, and a spherical reference system tool would again
be necessary (see the next paragraph: “About the primary cilium”).
A discrete, limited but precise organelle is the essential aspect of the centrosome
geometrical model. By observing the spinal column of mammals (in reptiles and fish
the number of vertebrae is even larger) we can check the preceding statement: we do
not see a variable continuous distribution of the form, but its precise (and discrete)
distribution, so that the more cranial vertebra is always the atlas, followed by the
epistropheus and gradually by the characteristic vertebrae for size and position in the
column, always predictable and discrete (cervical, thoracic, lumbar), that maintain
their typical (cervical, thoracic, lumbar) forms; like polyhedrons, each vertebra (in
different individuals of the same species), can vary in size but not in shape. Through
the geometrical molecular architecture of the centrosome (hardware: γ-TuRC
inclination) and topogenic sequences interacting with γ-TuRC receptors (software),
DNA has acquired the capability to store (and transmit to descendants) geometric
directional developmental instructions (understood and translated by the centrosome)
and has achieved a complete mastery and command to accomplish developmental 3D
programs. The orientation of first cell divisions (cleavage) in Metazoa confirms this
assertion: “When centrioles are experimentally ablated, spindles drift within the cell”
(Azimzadeh and Marshall, 2010); without two orthogonal few-degrees-of-freedom
“geometric” organelles, planarians cannot carry out the orientation of cell division
planes, typical of the spiral cleavage of their ancestors. Gastrulation is also
compromised. As we have observed, growth directions are fixed and invariable in the
elementary programs: when a lot of nested microprograms belonging to different
developmental modules are added up, the final results could show differences, also
considering the random local variability and fluctuation of cell dimensions and their
number (size and division rates are subject to several growth factors and hormones),
but, as we will soon see, the fine control of angles has achieved exceptional results
during evolution.

69
Quercus has about 500 species
Quercus has about 500 species, because of the enormous differences of shape; in each
individual of the same species, branches and roots have locations and orientations
(direction of the proximal-distal axis) whose values are continuously distributed:
branches can arise from any point of the circumference of the trunk and be oriented in
any direction, showing continuously variable tilts (compare to Illacme legs); only in a
few genus - Larix, Abies, Cedrus, Cocos, Phoenix, Agave - branches arise with a
characteristic inclination (but from any point of the circumference of the trunk), and
this order is already lost in twigs: not so for the 750 legs of Illacme pleniples, all
arising, as I have said, from an invariable constant point of the body circumference
and all with the same (bilaterally symmetric) parallel orientation of the proximal-
distal axis. In plants a different reference system, sensitive to different cues, inner
(MT disposal) or outer (light and gravity) but not controlled by “discrete” organelles,
drives their orientation and development, without a fixed (but precise) number of
discrete degrees of freedom (which is the main characteristic of the animal
centrosome): so words like “Ventral, Dorsal, Left, Right”, when referred to plant
anatomy, are meaningless. On the contrary, “tube elaboration in the Drosophila
embryo is highly stereotyped and often genetically determined… Recent studies of the
branching program of the mouse lung reveal that the branching patterns of complex
mammalian tubular organs animals can be highly stereotypical. Metzger and
colleagues (2008) examined hundreds of fixed mammalian lungs to generate a 3D
reconstruction of mouse lung morphogenesis at different developmental stages. They
demonstrated that from animal to animal, the pattern of lung branching is highly
reproducible and involves only three modes of branching used reiteratively in only
three different subroutines to pattern the entire lung. The modes of branching were
described as (1) domain branching, (2) planar bifurcation, and (3) orthogonal
bifurcation“ (Andrew and Ewald, 2010). Unlike plants, metazoan limbs do not arise

70
from any point of the body (and neither do cranial and spinal nerves, etc.), but their
location and their orientation have a fixed, highly reproducible, stereotypical value
(chosen between the permitted degrees of freedom).
A precise order and control and an extraordinary fine tuning of inclination and
direction is strikingly evident in fruit fly imaginal discs (page 14), in feathers
(maintained from the rachis to barbs and barbules) in limbs (leg, paw, claw) and even
in hairs, from flies (bristles) to humans (in our eyebrows and eyelashes, hairs show a
locally controlled - bilaterally symmetric - direction, and also in moustaches where
hair tilt is carefully and locally controlled, as hair inclination is characteristic and
stereotypically reproduced in the beard); Metazoa need a precise architecture for the
locomotion apparatus to perform extraordinarily difficult and highly directional tasks
in order to balance, walk, run, fly, swim. So metazoan organisms are capable of
developing their sophisticated 3D bilateral symmetry because precise, discrete,
stereotyped and programmed instructions for position and direction are “translated”
by an enantiomorphous “discrete” centriole. Illacme plenipes, as I have already said,
has 750 legs arising from a fixed point of the cylindrical body circumference with the
same (bilaterally symmetric) orientation: all individuals are about geometrically
identical. And so is it in Paramecium tetraurelia with its thousands of cilia arranged
in dozens of parallel longitudinal rows in every individual, each hardly
distinguishable from others: the number of basal bodies and rows (similarly to
Illacme legs) can vary, but not their geometrical arrangement. On the contrary, two
trees of the same species are neither similar nor have an identical distribution of
branches, which are free to arise from any point of the trunk and with variable
direction.
Human opposable thumbs: a great success of 3D angle control
Morphogenesis and chemical gradients do not agree: the first requires a discrete and

71
precise disposition and orientation of cells and extracellular fibers, the second do not
possess 3D information.
In nephrons, sweat or sebaceous glands, middle and inner ears we see the result of a
coordinated and precise hystological architecture; in limbs, which in Vertebrates are
composed of a very large number of cells, we can see an even more astonishing
precision in the management of angle geometry.
A very simple experiment: open your hand and point the tip of the thumb towards
your eyes: it is rotated of ~80° in respect to the other fingers (compare to the big toe
(hallux) orientation in your foot which is identical in all the five fingers):
The bilaterally symmetric repositioning of the thumb axes, properly oriented in order
to build an opposable finger, has been a milestone of extraordinary importance for the
evolution of Humans, an extraordinary result achieved by the fine control of angles;
this had been possible because of the high capability of precisely managing 3D
angles, a fundamental characteristic of metazoan cells. Is it conceivable to obtain a
similar result (bilaterally symmetric) through morphogen gradients? In addition: a
simple (and then likely) mutation in only one gene coding for a targeting sequence
recognized by the corresponding γ-TuRC receptor of “right” and “left” symmetric
centrosomes, as proposed by the geometrical model of centrosome functioning (see
later), can sustain the evolutionary variation of an angle (like in right- or left-handed
developing snails): what evolutionary event can represent the molecular base of
morphogen gradients able to realize the appearance of new organisms equipped with
bilateral symmetric opposable thumbs?
The centrosome: an input-output cellular spherical reference system tool
“When centrioles are experimentally ablated, spindles drift within the cell”
(Azimzadeh and Marshall, 2010).
“Our results suggest that selective pressure to maintain the centrosome in animals

72
comes from the need to coordinate specific developmental processes rather than a
fundamental cellular requirement for the organelle” (Azimzadeh, 2012).
No centrosome? No order.
At this point of the theoretical analysis an initial summary is convenient:
compartmentalization of the cell cortex and segregation of mRNAs (localization of
translation) are mediated by MTs; the fundamental role of the centrosome consists in
operating like an interface between DNA coded geometric instructions (the cell
biochemical mapping “software” consisting in targeting or topogenic sequences, or
ligand-“zip-codes” that interact with the corresponding γ-TuRC receptors) and the
corresponding final locations in the cell, reached by oriented MTs, conveniently
nucleated and irradiated by bi-oriented (longitude + latitude) γ-TuRCs to wire the cell
cortex (the cell biochemical mapping “hardware”: orientation and inclination of γ-
TuRC scaffolds).
Now we can lay the theoretical foundation of the model.
A first model of the centrosome
Equivalent or not equivalent, that is the question
“Centriole duplication is part of the mechanism by which the cytoskeleton of the
daughter cell is patterned upon that of the mother. It is probably not correct to say
that one centriole provides a template to make another (and there are instances of
centrioles arising de novo), but some kind of copying appears to be involved...
Examples of self-organization have long been familiar to biochemists under the
heading of self-assembly. Ribosomes, microtubules, microfilaments, virus particles,
and lipid bilayers come to mind” (Harold, 2005).
“Centrioles may act to keep PeriCentriolar Material components in a precise

73
position throughout the cell cycle and so be useful in the control of the position of the
axis of polarity and division.” (Gueth-Hallonet et al., 1993).
“It seems that in Protists and in Metazoa the triplets of basal bodies are not-
equivalent” (Beisson and Jerka-Dziadosz, 1999).
Equivalent or not equivalent, that is the question. Rather, it is the main fundamental
problem we must face. As already observed, the sophisticated 9-fold symmetry
architecture (very difficult to build) would be meaningless, useless and
incomprehensibly costly, if their 9 components were undistinguishable. If Metazoan
centriolar triplets are diverse and not equivalent like in Protists, as supposed by
Janine Beisson and Maria Jerka-Dziadosz, centrioles will operate like “biological”
protractors/goniometers also in Metazoa. Vorobjev and Chentsov (1982) presented in
their cited article, interesting pictures of the appendages of the “mother” centriole in
mammalian cells that may support the idea of the non-equivalence of the mammalian
centriolar triplets: each triplet appears different in shape in comparison with the
others and the nine ribs of the “daughter” centriole are also different. Unfortunately,
studies on centrioles in Metazoa are difficult because centrioles are embedded in the
proteinaceous matrix of the PCM.
”Is it possible to confirm this idea that the circumferential, morphological, structural
and molecular asymmetry of centrioles can be inferred from Mammals ciliated
epithelia? While the circumferential anisotropy of centrioles cannot be ascertained
within the centrosome, its existence can be inferred from the properties they express
during ciliogenesis, be it the formation of a primary cilium or of bona fide 9+2 cilia
in ciliated epithelia, some of which at least derive directly from the centrioles. As in
Ciliates and flagellates, these basal bodies nucleate appendages of various molecular
compositions (basal foot, striated rootlets, alarm sheets, etc, which anchor the basal
body to the membrane and to the cytoskeleton) and these nucleations arise at specific
sites of the basal body cylinder; in particular, the basal foot is located on triplets 5
and 6 corresponding to the side of the effective stroke of the cilium. What is
remarkable is that basal feet develop before the basal bodies reach their membrane

74
site and before they acquire their functional orientation “(Beisson and Jerka-
Dziadosz, 1999).
Some certainty, at last (but does any certainty exist in science?): rather, some clear
experimental evidence. Mammalian centrioles have non-equivalent triplets that
nucleate divers (morphologically and functionally) appendages before reaching the
cell membrane. We have before supposed an informational non-equivalence of
triplets because our theoretical analysis had accompany us to this conviction: now we
have a first demonstration we were right. However this is true only for mammalian
centrioles and basal bodies. What happens inside the centrosome?
It is not hazardous trying to face the previous question, following this path: the
theoretical analysis of a spherical reference system based on two orthogonal
protractors/goniometers gives significance and meaning to the highly sophisticated
structure and rotational polarity of centrioles and indicates how a centrosome could
operate; if centrosome behavior in Metazoa agrees with the functioning of a
theoretical spherical reference system organizer, it is possible to hypothesize that its
geometrical role is likely. Indeed many studies (see references) confirm the
geometrical role of centrosomes in defining cell polarity, orientation of division
planes, differential cell-cell adhesion, migration, apical constriction, convergence and
extension by intercalation, extracellular fiber disposition. Centrosome’s geometry
implies its function: metazoan triplets appear not equivalent.
Centrioles in Ciliates and centrosomes in Metazoa: biophysical and architectural
considerations about non-equivalence
“Among the conserved proteins involved in the biogenesis of the canonical 9-triplet
centriolar structures, SAS-6 and Bld10 have been shown to play central roles in the
early steps of assembly and in establishment/stabilization of the ninefold symmetry.”
(Jerka-Dziadosz et al., 2010).

75
As we have already seen, centrioles are present in eukaryotic cells, but not in higher
plants and most fungi; unicellular organisms have only centrioles/basal bodies, each
one independent, whereas in multicellular algae and Metazoa two orthogonal
centrioles are embedded in the PeriCentriolar Material that forms the centrosome.
Protists show rotational, cylindrical (or rather circumferential) symmetry, but also
asymmetry is frequent whereas they never show bilateral symmetry [pay attention to
the difference between “180° rotation” of a geometric 2D figure (the sequence of
marks remains unaltered) that seems, but is not, bilateral symmetry, and “180°
reflection (or overturn)” that is really bilateral symmetry (the sequence is reversed or
opposite): the axes around which the two figures move are diverse, in the first case
being orthogonal to the plane in which the figure lies, whereas in the second case the
axis lies on the plane containing the figure]. On the contrary, Metazoa show bilateral
symmetry and a 3D architecture of their organs and limbs; Ciliates have cylindrical
centrioles, Metazoa have two orthogonal cylindrical centrioles embedded in a
spherical centrosome: the second centriole, orthogonal to the first, adds the capability
of completely controlling the third dimension, indispensable for managing the
arrangement of cells in multicellular organisms. The idea is that Metazoa have
developed new pathways to adapt Ciliates’ sophisticated molecular mechanisms in
order to transmit the 9-fold symmetry and polarity of two orthogonal cylindrical
centrioles to the spherical wedges and sectors of the centrosome (or rather of the
PCM): so Metazoa could assemble once and for all, a tool through which they can
control, direct and organize their 3D anisotropic growth. In fact, to use a spherical
reference instrument based on two orthogonal protractors/goniometers, like a globe, a
two-step process is necessary: firstly the longitude of a point ( coordinate) must be
found, then its latitude (θ coordinate) can be identified, but only after a rotation of the
vertical protractor around its vertical axis that aligns the protractor inner border in
correspondence with the found longitude; this appears particularly complex and
difficult (and frankly unlikely) in a cell; on the contrary, if a little sphere (always
maintained in a controlled position and orientation) like the centrosomal PCM is

76
organized only once and for all in each cell and built with its surface subdivided and
compartmentalized into 45 small areas (as we have seen in the above geometrical
analysis) each one oriented and labelled by molecular receptors that recognise the
geometric (molecular) signals intended for the corresponding longitude and latitude
(and, consequently, for the corresponding cytoplasmic pyramid too), the problem of
spatial orientation in the cell environment has a very easy solution. Like on a globe,
parallels and meridians are marked on its surface once and for all to facilitate the task
of finding a point of given coordinates, so the 9-fold symmetry of two orthogonal
cylindrical centrioles, transmitted and impressed on the centrosome surface once and
for all, permits an easy translation of molecular geometric signals (in which the
intended coordinates are coded) into their corresponding localizations in the
cytoplasm, only by the usual signal-receptor interaction between targeting sequences
and γ-TuRC (or scaffold/docking platform) receptors located on the centrosome
surface. Obviously, this must happen maintaining fixed (or variable but, in any case
under control, that is never random) the orientation of the centrosome in the cell,
which is in accordance with the described mobility of the only DC: after building the
PCM surface, the MC controls its position and orientation, while the DC disengages
and is quite free in the cytoplasm. “Once and for all” are words that sound bad in
biology, because molecular turnover is the fundamental biophysical base for
managing variation of concentrations (thresholds); although many centrosome
components show high turnover rates, centrioles are very stable structures, effectively
built “once and for all”, and so it may be supposed for the PCM skeleton which
organizes orientation and labelling of γ-TuRC scaffolds. Therefore “once and for all”
means that the molecular PCM skeleton responsible for the inclination of γ-TuRC
scaffolds and the localized delivery of γ-TuRC receptors is somehow transferred to γ-
TuRC scaffolds in order to orientate and molecularly label each PCM compartment
only once per cell cycle, just like centrioles are built only once per cell cycle.
A regular polygon of 9 sides (nonagon or enneagon) has 9 internal angles of 140° and
is composed by 9 isosceles triangles (radially disposed) whose vertex (or central)

77
angle measures 40° (Fig. 2 on page 35). Ciliates possess advanced biochemical
mechanisms (with high predisposition for self-assembly) to assemble and build two
angles: the right angle and the angle of 140°; the first is evident in the orthogonal
arrangement of MC and DC during duplication and in the orthogonality between each
cartwheel spoke and the A-microtubule of the corresponding triplet of the centriolar
wall; the second (and its supplemental of 40°) is evident in the 9 fold symmetry of
cartwheel spokes and triplets: in the transition zone of Chlamydomonas basal bodies,
where the axoneme is organized, there is an astonishing “stellate structure” (well
described by Geimer and Melkonian through a beautiful electronic microscope image
in their cited article, free on the Internet) composed of 9 isosceles triangles (nonagon)
with a vertex angle of 40° and two identical angles, each of 70°: the base of each
triangle forms an angle of 140° (70° + 70°) with the base of the consecutive triangle.
In Mammals the right angle is also shown by some Microtubule Associated Proteins
(MAP-2, MAP-4, Tau-MAP). Kitagawa and colleagues (2011), van Breugel and
colleagues (2011) and Gönczy (2012) have shown that the N-terminal domains of
SAS-6 dimers (a conserved protein indispensable for centriole building) naturally
interact between themselves at 140° (internal angle) to form polygonal oligomers,
rings and helices (left- or right-handed); SAS-6 has been found in every model
organism: Chlamydomonas, C.elegans, D. rerio, X. laevis, M. musculus, G. gallus, H.
sapiens. "Intriguingly, when Drosophila SAS-6 is over expressed together with Ana2
(ortholog of SAS-5), the two proteins can assemble into well-ordered tubules that
bear a striking resemblance to the cartwheel" (Cottee et al., 2011). Then, SAS-6 is a
powerful and quasi-autonomous (self-assembling) tool to build angles of about 140°.
The case of the hexapode Acerentomon microrhinus is interesting in which Riparbelli
and colleagues (2009) found ” large centrioles composed of 14 doublet microtubules
that serve as templates for cilia and flagella and organize mitotic and meiotic
spindles”: self-assembly of SAS-6 in cooperation with other molecular complexes
allows cells to build other symmetries (a regular polygon of 14 sides has 14 angles of
154.28°), when necessary: centrosomes and centrioles with non-9-fold symmetry can

78
perform (as well as, or perhaps better) tasks like axoneme or spindle formation,
different from their fundamental morphogenetic role as geometrical interface:
metazoan morphogenetic 3D programs are written in “140° language” (or 40° vertex
angle) and must be interpreted by a “compatible” 9-fold symmetry interface
(remember the considerations about the “biological anti-aliasing filter”, genetics of
targeting sequences and “genetics of morphogenesis”); centrosomes of somatic cells
of Acerentomon microrhinus possess 9-fold symmetry MCs able to organize DCs
equipped with different symmetry to perform particular functions, different from
morphogenetic ones: “Strikingly, daughter centrioles contain a transient cartwheel
that is lost after maturation. The length of radial spokes is like that found in 9-fold
cartwheels, whereas the diameter of the hub varies according to the dimensions of the
centriole cylinder. This suggests that the hub may dictate the master plan for
centriole geometry. Finally, the finding that 14-doublet centrioles arise from 9-
doublet mothers points to an alternative model for centriole assembly” (Riparbelli
and colleagues 2009). Guichard and colleagues (2010) have shown a 110-nm stalk
that connects at 90° the central hub of the DC procentriol to the MC in human
centrosome; Chlamydomonas Bld-10p, a component of the cartwheel-spoke tip,
connects the triplets to the spokes and maintains them orthogonal to the spoke.
Picone and colleagues (2010) have identified dynamic centrosomal MTs that mediate
homeostatic length control in Metazoa cells, like Chlamydomonas manages axoneme
length. Geometrically, the management of 90° and 140° angles, together with the
capability of controlling distances and the predisposition for self-assembly of rings
and curved structures, are sufficient for the cylindrical (or, rather, prismatic)
symmetry of two orthogonal centrioles to be transferred to a spherical (or, rather,
polyhedral) shaped centrosome; some models (schematics in Piel et al., 2000 and in
Bornens, 2002, 2012, free on the Internet) clear and confirm this idea, showing that
centrioles, through the nucleation of a “fan-shaped” structure made up of MTs lying
on planes passing through the axis of the centriole and the A-MT of the triplet-blade
and having different tilts (like some radii of a circle starting with an angle of 40°

79
between themselves: something like the fan-shaped structure schematized in Fig. 11)
can transform the longitudinal rectilinear shape of each blade of their walls into a
curved spherical surface; note that the geometry of 140° (40° supplemental angle) is
respected in this model; the schematics and images proposed by Gopalakrishnan and
colleagues (2011) in “SAS-4 provides a scaffold for cytoplasmic complexes and
tethers them in a centrosome.” (Nature Commun.), Mennella and colleagues (2012)
in: “Subdiffraction-resolution fluorescence microscopy reveals a domain of the
centrosome critical for pericentriolar material organization” (Nature Cell Biol.),
Lawo et al.(2012) in ”Subdiffraction imaging of centrosomes reveals higher-order
organizational features of pericentriolar material” (Nature Cell Biol.), are consistent
with this idea. Centrosome architecture reminds us of fullerenes: both self-manage an
angle (140° in centrosomes, 109.5° in fullerenes) and build cylindrical or spherical
structures taking advantage of their aptitude for self-assembly. Also the self-
assembled “baskets” of clathrin triskelia (vesicles) that show angles of 120° have
something in common with centrosomes: intriguingly, the N-terminal of the human
SAS-6 shows 40% identity (65% positives) with the central domain of the clathrin
light chain; this can support the previous considerations about Acerentomon
microrhinus: 14-fold symmetry (as we have seen a regular polygon of 14 sides has 14
angles of 154.28°) may be realized through the cooperation of molecular complexes
that force the basic interaction of SAS-6, whose most thermodynamically stable
configurations are about 140° ±10%. Rodrigues-Martins and colleagues (2007) have
shown that self-assembly is required in centriole formation: "centriole biogenesis is a
template-free self-assembling process triggered and regulated by molecules that
ordinarily associate with the existing centriole. The MC is not a bona fide template
but a platform for a set of regulatory molecules that catalyzes and regulates DC
assembly". Song and colleagues (2008) arrive to similar considerations, finding a
mutual cooperation between the PCM and the centrioles: “the PCM itself may direct
the formation of the DC”. In mice, first embryonic divisions occur with unpredictable
cleavage plane positions and blastomeres lack centrioles (they will appear only at the

80
blastocyst stage) but possess PCM and γ-TuRCs to form spindles: evidently the
kernel or nucleus of the PCM necessary for mitosis is autonomously self-assembled
in each new cell until the stage of 64-blastomeres, when centrioles begin to be formed
and build a complete centrosome. Kubo and colleagues (1999) and Dammermann and
Merdes (2002) observed that pericentriolar satellites, after recruiting proteins
indispensable for anchoring γ-TuRCs to their centrosomal docking platform (centrin,
pericentrin, ninein, SAS-4 etc.), are transported to the centrosome by MT-linked
dynactin. Centrosomes of Spisula, after chemical disassembling (1.0 M KI) lose γ-
TuRCs, but maintain their PCM skeleton (built once and for all) intact: after
incubation with oocyte extracts containing γ-TuRCs, they recover: "This recovery
process occurs in the absence of microtubules, divalent cations, and nucleotides"
(Schnackenberg et al., 1998). Ou and colleagues (2002) found that the PCM is
organized in an ordered and geometric fashion, showing a characteristic “centrosomal
tube”, closed at one (the inner) end and open at the other end, with a diameter of 1.5
μm and a depth of 2 μm, dimensions that are larger than centrioles height and width
but comparable to the dimensions of the centrosome: "A subset of PCM proteins have
been shown to be arranged in a tubular conformation with an open and a closed end
within the centrosome... Microinjection of antibodies against either CEP110 or ninein
into metaphase HeLa cells disrupted the reformation of the tubular conformation of
proteins within the centrosome following cell division and consequently led to
dispersal of centrosomal material throughout the cytosol (CEP250/c-Nap1,
pericentrin, γ-tubulin and ninein)" (Ou et al., 2002). Alliegro and Alliegro (2008)
have identified in Spisula solidissima several non-coding centrosomal RNAs: the
importance of non-coding RNA in organizing protein complexes is well known
(spliceosomes, ribosomes, and nucleoli). Unlike fullerenes and clathrins vesicles,
which are spherical layers, centrosomes do not possess an accurately spherical
surface: the PCM grows around centrioles (the core components of the centrosome)
and organizes perfectly oriented γ-TuRCs on an irregularly spherical surface. It seems
that only one centriole might be sufficient for organizing the PCM; indeed, a second

81
centriole, orthogonal to the first, is indispensable for transmitting correct “latitudinal”
inclinations and marks (or latitude receptors responsible for angle θ) to the parallel
caps and rings (Fig. 3 and 6). In the geometrical model, as already hypothesized, cells
use mechanisms to manage 90° and 140° angles.
The transition from unicellular Protists to multicellular Metazoa is founded on
the orthogonal disposition of two centrioles.
So, it seems that Metazoa have empowered the sophisticated centriolar molecular
apparatuses of Ciliates (capable of ordering and arranging new cell structures under
the influence of others, already existing: assembly of microtubules, centrioles
duplication, assembly of axonemes and IntraFlagellar Transport) to spherically
organize their centrosomes (the diameter of the centrosome is about only two times
the height of centrioles, then not so different) in order to build 3D complex organs
and organisms: Metazoa arrange spherical disks and wedges around two orthogonal
centrioles, by the only operational control of 90° and 140° angle. In Ciliates
centrioles perform three main functions: 1°) as basal bodies they manage the
assembly of the axoneme, transmitting their 9-fold-symmetry, controlling axoneme
length and organizing the complex IntraFlagellar Transport; 2°) as centrioles they
build and maintain the cytoskeleton; 3°) as Mother centrioles they are used as
platforms by new arising procentrioles. The addition and the successive fusion of
these three functions have prepared and paved the way for the next step: building a
centrosome. The transition from unicellular Protists to multicellular Metazoa is
founded on the orthogonal disposition of two centrioles (see the paragraph “Shared
and coordinated orientation of metazoan centrosomes” on page 46). As in
Chlamydomonas and Paramecium the rotational asymmetry and non-equivalence of
the triplets is used to build axonemes and complex cytoskeletons made up of
different fibers each one linked to a particular triplet, so in Metazoa the two

82
orthogonal centrioles, after assembling a “quasi-spherical” PCM skeleton, recruiting,
attaching, orienting and anchoring γ-TuRCs to impose spherical polarity and
asymmetry, cooperate to “label” γ-TuRCs. So, γ-tubulin rings are anchored, oriented
and labeled by the participation of many proteins –SAS-4, ninein, centrin,
pericentrin– each one present in several isoforms: the high number of these proteins
confirms that the process of γ-TuRCs anchoring is not a simple docking process: this
process establishes the sophisticated (and indispensable) spatial orientation of γ-
TuRCs: this function is the evolution of Ciliates’ 1°)-function of axoneme assembly.
Like in Ciliates’ 2°)-function each centriolar triplet uses its own unique 3D molecular
shape to recognize and bind its particular ligand (one of the several cytoskeletal
components), metazoan triplets have molecular structures to recognize and bind their
proper ligands in order to correctly transmit located receptors to γ-Turcs, already
oriented on their scaffolds/docking platforms. In Ciliates the centrioles (one or more)
manage the cyto-skeleton, in Metazoa the centrioles (two per cell) manage and
organize the PCM-skeleton which, in turn, builds and manages the cyto-skeleton.
Metazoan centrioles organize the geometrical architecture of the PCM (oriented
scaffolds for γ-TuRCs: centrosome “hardware”) and transmit their non-equivalence
by displacing to each scaffold/γ-TuRC the receptors corresponding to its inclination.
The evolutionary process is: from only one centriole > to two orthogonal centrioles
(diplosome); from Protists “centriole>cyto-skeleton” to Metazoa “diplosome>PCM-
skeleton”.

83
The only “true” centriole is the mother centriole: the daughter centriole is an
intermediate product between a procentriole and a “true” centriole, only useful
for cooperating orthogonally with the MC to form the PCM of a 3D reference
system organizer, the centrosome.
Ciliates duplicate their centriole, 3°)-function, using a complete centriole as a
platform near which a new procentriole arises at right angle and, when it is
completely formed disengages: this is already a diplosome, likely used to polarize the
new centriole in the same manner than the “mother” centriole (here we utilize the
term “mother” to underline the similarity of the centriole cycle in Ciliates and in
Metazoa) in such a way that a coordinated orientation (positioning of the “0°” mark)
between mother and daughter centrioles is maintained and the new centriole is
correctly inserted in the cell to avoid that an uncoordinated orientation causes
distortions in their complex cytoskeleton (Paramecium); the task of positioning a
basal body with nine different triplets, each one connected to a specific cytoskeleton
component, has something in common with the insertion and connection of the pins
(several and, above all, functionally different!) of an integrated circuit (microchip)
into an electronic circuit: each pin must be precisely connected to a defined
conductive track on the printed circuit board. It seems that Metazoa have empowered
and developed this very important stage: in Metazoa the centriole cycle is temporarily
stopped at the stage of orthogonality (diplosome) to build and organize the PCM and,
successively, restarts and finishes only in the next S phase. Follow carefully this point
of view: in late S phase, after the process of maturation of the DC, two new MC
generate and correctly orientate two new procentrioles (like in Ciliates) but the
process of maturation of procentrioles is blocked, interrupted, suspended: making use
of this convenient orthogonal disposition, each diplosome forms the PCM and cell
division occurs, each new cell having a MC associated with a “pro-/daughter-“

84
centriole; the process of centriole biogenesis will be re-activated and completed in the
next early S phase (definitive DC maturation into a complete, ultimate MC). The only
“true” centriole is the MC, whereas the DC is an “aid-centriole” an intermediate
product between a procentriole and a “true” centriole, only useful for cooperating
orthogonally with the MC to form the PCM of a 3D reference system organizer, the
centrosome. Before we have used the expression “addition and successive fusion”:
addition of 1°) Ciliates’ function (axoneme assembly) to 2°) function (cytoskeleton
building) occurs during the orthogonal disposition of the arising procentriole and the
“mother” centriole: this stage has been prolonged and, in a sense, is overlapping and
not in phase with the metazoan cell cycle; this can help us understand the strange
centriole/centrosome metazoan cycle.
Whether centrosomes and MTs are involved in AP polarity formation in C. elegans
embryos and in other organisms has been a subject of controversy: Tsai and Ahringer
(2007) have cleared up this problem demonstrating that “polarity only occurs when a
small, late-growing microtubule aster is visible at the centrosome; MTs deliver
positional signals and are required for establishing polarity in many different
organisms and cell types. In C. elegans embryos, posterior polarity is induced by an
unknown centrosome-dependent signal". This is a fundamental milestone: in Metazoa
the spherical polarity of centrosomes and centrioles is transferred through the
cytoskeleton to the whole cell cortex.
As we have seen, to satisfy the basic requirements of a spherical reference system, in
order to recognize coded geometric signals and translate them into their desired final
locations, “cellular protractors/goniometers” must possess: 1) different marks; 2)
constant ordered sequence of marks; 3) one start mark; 4) controlled orientation of
the tool in respect to reference points: “In C. reinhardtii the polypeptide VFL1 coded
by the gene vfl1 (Variable number of FLagella), binds to the triplets 9-1-2 near to the
triplet N° 1… Further evidence of rotational polarity comes from the asymmetric
attachment of various appendages to specific triplets in the basal bodies and from the
site of daughter basal body formation adjacent to defined triplets in the mature

85
organelles” (Silflow et al., 2001): evidently in C. reinhardtii the triplet N° 1 has its
own unique and distinguishable triplet marker. Surprisingly Yoshinori and Yukio
(2010) found a human ortholog of Chlamydomonas Vfl1 protein and other correlated
centrosomal proteins: “the bibliography as well as database searches provided
evidence that the human proteome contains at least seven centrosomal leucine-rich
repeat proteins;… we identified a centrosomal protein called CLERC (Centrosomal
leucine-rich repeat and coiled-coil containing protein) which is a human ortholog of
Chlamydomonas Vfl1 protein. The bibliography as well as database searches
provided evidence that the human proteome contains at least seven centrosomal
leucine-rich repeat proteins including CLERC. CLERC and four other centrosomal
leucine-rich repeat proteins contain the SDS22-like leucine-rich repeat motifs,
whereas the remaining two proteins contain the RI-like and the cysteine-containing
leucine-rich repeat motifs. Individual leucine-rich repeat motifs are highly conserved
and present in evolutionarily diverse organisms”.
There are then surprising correspondences between the geometrical model of the
centrosome (nine meridian wedges, each one subdivided in five parallel areae) and
many reported findings: the compartmentalization of the cortex in Drosophila
embryo during its 14th mitosis (one impair meridian wedge, four right and four left
meridian wedges, each one subdivided in five parallel areae), the
compartmentalization of the cortex in yeast (about 50 cortical domains); the role
played by the centrosome in asymmetric delivery of mRNAs in I. Obsoleta; the role
played by centrosomal microtubules in segregating different mRNAs into different
cytoplasmic regions; the centrosomal proteins found by Yoshinori and Yuko.
Then, now it is the time to set the hypotheses of the centrosome geometrical model.

86
Fig. 6 Centrosome theoretical geometrical model: from 2D rotational polarity to 3D spherical polarity.
A: (top view): the MC (internal circle subdivided in 9 intensely coloured sectors) is responsible for
“longitude” that is transmitted to the whole PCM (external annulus, weakly coloured), whose -TuRCs
(small bars) acquire an inclination parallel to the corresponding centriolar blade; each MC blade faces one
meridian wedge. In each wedge, all the -TuRCs have the same longitudinal inclination. B: after the
intervention of the DC, that imposes a rotational inclination corresponding to that of its blades, each -TuRC
acquires also the latitude inclination which is added to that of longitude. There is a double inclination: first
each -TuRC is parallel to the corresponding blade of MC, then it acquires the inclination parallel to the
corresponding DC blade; the eccentric positioned DC is responsible for “latitude” (two opposed spherical
caps and three parallel spherical disks): this second centriole/protractor is composed of two symmetric hemi-
protractors/goniometers. C: all the -TuRCs contained in the same cap or disc (coloured circles) whatever
their longitudinal orientation, are rotated to acquire the same latitudinal orientation, identical in the same cap
or disk. So, two 2D circumferential-rotational polarities are merged to realize a 3D spherical polarity
First hypotheses:
- Centrioles have a circumferential ordered and discrete asymmetry as a consequence
of macromolecular differences of their triplets.
- Centrioles are platforms for a set of regulatory molecules that assist and facilitate

87
the semi-self-assembly of the PCM.
- The MC transmits its 9-fold symmetry (longitude-dependent inclination:
coordinate of a spherical reference system) to PCM -TuRC scaffolds /docking
platforms.
- The DC transmits its 9-fold symmetry (latitude-dependent inclination: θ coordinate
of a spherical reference system) to PCM -TuRC scaffolds /docking platforms.
- Each -TuRC on a centrosomal scaffold/docking platform is oriented in accord with
the tilt of the local tangent plane: scaffold’s (and -TuRC’s) inclination is the addition
of the latitudinal tilt (DC) to the longitudinal (MC); on the same scaffold there are
several -TuRCs.
- Both centrioles transmit to the PCM their circumferential asymmetry and impress
their molecular non-equivalence.
- Each -TuRC (or its centrosomal scaffold/docking platform) receives from both
centrioles the receptors (longitude and latitude) corresponding to its own position and
orientation.
- Each -TuRC (or its centrosomal docking platform) displays its receptors to
recognise the signal (a molecular ligand with a particular targeting sequence)
corresponding to its own orientation and to the intended location in the cell cortex.
- The direction of centrosomal MTs depends on the orientation (tilt) of the -TuRCs
by which they are nucleated.
- Each signal (ligand) has a 3D shape that recognises only the receptors of that -
TuRC that is oriented in order to nucleate an MT with the desired direction: there is a
precise and invariable one-to-one correspondence between geometric signals and -
TuRC receptors.
- The centrosome is the cytoskeletal organizing center; it is polarized and its
orientation and position in the cell are strictly controlled.
- The correspondence between centrosomal and cortical compartments is univocal
(one-to-one) but time-variable, since it depends on the position and orientation of the

88
centrosome: a same -TuRC can be connected, at different times, to diverse cortical
compartments.
- The one-to-one correspondence between targeting signals and γ-TuRC receptors is
constant and invariable, whereas the one-to-one correspondence between targeting
signals (and related γ-TuRC receptors) and cortical compartments is time-variable.
- The centrosome is a geometric discrete interface that receives geometric coded
signals (input), matches each one with the corresponding -TuRC receptors
(decoding: ligand-receptor interaction) and nucleates oriented MTs (translation) to
reach the required locations (output).
- Aster, cytoskeleton and cell cortex receive from the centrosome the same spherical
asymmetry (mapping or polarization).
- Centrioles are chiral structures as a consequence of their rotational reversible
asymmetry.
About the primary cilium
The primary cilium (only one per cell) is a non-motile cilium whose basal body is the
MC of the centrosome, which, in G0 non-dividing and non-migrating cells, moves
towards the cell membrane (maintaining its orientation) [“non-motile” does not mean
“motionless, static”: motile cilia of the multiciliated cells in the respiratory
epithelium, like propellers, produce an extracellular intense flow; non-motile primary
cilia do not, however they can bend when subject or exposed to flows]; in epithelial
cells it is positioned near the center of the apical surface; if cells re-enter mitosis, the
cilium is reabsorbed and sheds: motile cilia are formed ex novo, primary cilium does
not, it is unique (one-per-cell) as the MC that is its basal body and follows the strange
cycle of the MC. It is involved in cell-cell signaling pathways: in Vertebrates, unlike
other Metazoa, Hedgehog receptors are positioned on the primary cilium. Motile cilia
of the Hensen’s node cells generate a flow, involved in left-right asymmetry, which is

89
“sensed” by primary cilia of neighbouring cells. Avasthi and Marshall (2013) and
Wood et al. (2013) hypothesize that primary cilia, like antennas, are capable of
receiving and transmitting too.
Indeed it seems possible that the direction of provenance (the location of signal
sources) is an understandable “geometrical information” of a signal, like in planar
cell polarity or in cells near the midline: this information does not consist in any
molecular signal but it is simply implicitly furnished by the spatial direction of a
flow: it can be deciphered and decoded only by receptors associated with (or
corresponding with) the oriented blades of the basal body, i.e. receptors located not
randomly but in precise positions of cilium circumference, in correspondence of one
of its 9 doublets. The important task of maintaining the midline in its correct position
or repositioning a new midline (after first blastomeres splitting events or during
regeneration after surgical or accidental injuries) can take advantage of information
about the provenance of signals (deciphering if they arrive from the left or from the
right) in order to drive the process; as we have observed before, Carbajal-Gonzales
(2012) has shown that “in Eukaryotic cilia and flagella, radial spokes are different
from each other, but conserved across species”: the primary cilium radial spokes may
be circumferentially polarized by its polarized MC/basal body. Cao and colleagues
(2012) reported that a sophisticated mechanism organizes the centriole-to-basal body
transition and cilia assembly through miRNA-mediated post-transcriptional
regulation.
Why only one primary cilium per cell? Only one primary cilium is convenient for
“understanding” different directions of provenance of a regular (quasi-laminar) flow
of molecules: many cilia, random or orderly displaced, are not because a space
crowded with cilia cause flow turbulence with unpredictable bounces and collisions
(recall the values of molecular speed seen on page 60) of signalling molecules subject
also to electric (+/- charges) or chemical (hydro-phobic/-philic groups) forces during
impacts with cilia membranes, able to alter and modify the direction of hits between
molecules and cilia (noise). A pinball machine with its cylindrical spring active

90
bumpers illustrates this concept.
Receptors that enrich the primary cilium, exposed to a morphogen flow, are
stimulated by signal kinetics (concentration levels going over threshold). Hedgehog
(Hh) signalling, as I have said, in Vertebrates occurs in primary cilia, involving
kinesin (KIF7) and dynein transport of many molecules (Smoothened, Patched,
SUFU, Gli): downstream Hh signalling, in Vertebrates as in Invertebrates similar
processes occur, but only in Vertebrates (the most complex 3D organisms) does Hh
signalling need receptors positioned on the primary cilium. In order to better
appreciate the following considerations, it is interesting to review some numbers:
concentration of Sonic hedgehog (Shh) in chick embryo neural tube decreases from
15 nM to 0.5 nM; 1 nM concentration corresponds to about 1 molecule/µm3 (0.6 to be
precise; a simple addition of exponents: 1M means 6.022 x 1023
molecules per liter or
dm3; 1 nM is 1 x 10
-9 M or 6.022 x 10
14 molecules; 1 dm
3 contains 10
15 μm
3: that is
6.022 x 10-1
or 0.6 molecules per μm
3); the volume that faces a primary cilium (with
the same size of the cilium: 7 µm length and 0.3 µm diameter) measures about 2 µm3;
molecules of Shh in a volume of 2 µm3
vary from 18 to 0.6; Shh gradient is capable
of signaling at a distance of 30 cell diameters; protein diameter average size is about
3-6 nm; primary cilium circumference measures 1,900 nm (1.9 µm) , and its height is
(as just seen) 7,000 nm (7 µm); compare to Acetylcholine concentration [5*10 -4
M] in
the synaptic space [1 µm3] -> 300,000 molecules, and to hormones concentration
[10-8
M] -> 6 molecules/µm3 (average density of receptor: 300/µm
2, 9*10
4/cell); recall
also the values of molecular speed and collisions (page 60), considering that 1 µm3 of
gas at standard conditions (0°C or 273,15° K) contains a number of molecules of an
order of 107
(like Acetylcholine) but the temperature in Vertebrate cells is higher. The
gradient of Hedgehog is attentively controlled through N-terminus palmitoylation
which increases its hydrophobicity and facilitates its binding to cell membranes,
limiting its diffusion. Could receptors, randomly positioned on the cilium surface
(even on the opposite face), be able to interact with very few (and so small in respect
to primary cilium dimensions) molecules? Often in cell biology only small quantities

91
of solutes are necessary: 250 protons are enough for lysosome acidification. In order
to sense, “measure” and graduate the responses to very low concentrations of
important signaling molecules which move at high speed (addition of Brownian
motion and flow speed), cells use sophisticated strategies: the HER family of
receptors for epidermal growth factors can dimerize in such a manner that only one
molecule can activate some dimers while two ligand molecules are necessary to
trigger other types of dimers: then different dimers are activated by different
concentrations. Flows of very low concentrations of signalling molecules can better
and more quickly interact with receptors if these are correctly positioned in
correspondence of no more than two or three consecutive doublets, oriented towards
the flow and directly exposed to it: this is possible through the geometric role of the
rotationally polarized MC/basal body. “A structure at the ciliary base appears to have
all the features required for compartmentalization and which we thus call the ciliary
partitioning system (CPS). This complex consists of the terminal plate, which serves
as a cytosolic ciliary pore complex (CPC)… a plate-shaped structure containing nine
pores through which the microtubule doublets of the basal body pass. Each pore
expands from the doublet B-tubule into an opening well suited for the passage of
intraflagellar transport particle” (Ounjai et al., 2013). Receptors can be placed only
in correspondence to one or two doublets in order to coordinate the cell fast response
to a forecast source direction, and reduce noise: when a very weak signal is expected
from a forecast provenance, it is convenient to orient receptors towards the foreseen
direction (in migrating cells, for example). “In vertebrates, centriole position appears
to respond to planar cell polarity cues, consistent with the localization of some
planar cell polarity proteins at centrioles” (Montcouquiol and Kelley, 2003). When
receptors are placed everywhere on the cilium, through intra-flagellar transport,
activated signal molecules or signal transduction molecules can be carried, along the
corresponding doublets (primary cilium compartments), to the circumferential
polarized basal body, labelled with a targeting sequence coding the “number” of the
doublet and sent to the nucleus to be elaborated: qualitative analysis (spatial

92
orientation and type of morphogen gradients) can be added to quantitative analysis
(number of receptors stimulated in a temporal interval). Such a mechanism is
probably indispensable to allow epithelial cells, which have only one side free to
receive signalling molecules, to feel and estimate the direction of origin of very low
concentrated morphogens or to “sense” very few morphogen molecules from a
particular (forecast) direction. Primary cilium basal body is supposed to have been
previously oriented in the “mapped” cortex in such a way that each axonemal doublet
is located in a fixed and controlled position in respect to the cytoskeleton and
coordinated with the other cells of the epithelium: the basal body of the primary
cilium is the MC of the centrosome, that, already oriented during the last cell
division, has migrated to the cortex, maintaining its orientation, so that it can easily
arrange and orient its transition fibers, basal feet, and ciliary rootlets to anchor itself
to the membrane and maintain the correct orientation. So through a polarized primary
cilium, morphogen gradients (mathematically defined as vectors oriented toward the
source) can be deciphered, interpreted and translated by basal bodies: a controlled
rotational disposition of receptors can cooperate to increase the differential response
to variation of concentration rates (using thus three factors: concentration gradient,
receptor density and receptor circumferential position). Signaling pathways can
interfere with the number and circumferential position of primary cilium receptors in
order to better fine-tune the cell response to gradients of morphogens or to flows (see
left-right and the role of Hensen’s node cells): a primary cilium, circumferentially
polarized by its polarized basal body, can organize its receptors in such a way that it
can operate as a directional antenna or, rather, a directionally programmable antenna.
Left-Right
A role of centrosomes and centrioles in left-right patterning has been proposed by
many researchers:
“What sub cellular component is responsible for the crucial orientation event that

93
defines “leftward”? One likely possibility is that the coordination of the 3 axes is
performed by a cytoskeletal organizing center such as the centriole or basal
body...The sharp midline separation suggests that the first cell cleavage in X. laevis
may produce L and R halves that inherit differential chiral information.“ (Vanderberg
and Levin, 2009).
“An intrinsically chiral structure, perhaps the centrosome, serves as a template for
directing polarity in the absence of spatial cues. Such a template could help to
determine left–right asymmetry and planar polarity in development” (Xu et al.,
2007).
Cilia are considered the initiators of asymmetry: the flow generated by the motile
cilia of Hensen's node cells induces asymmetry in many internal organs (heart, liver).
What role can centrioles play in left-right patterning? Brown and Wolpert (1990)
hypothesized a molecule or a cellular chiral structure, named “F”, capable of adding a
Left-Right axis perpendicularly to the AP and DV axes.
An extraordinary answer to this question arrives from the geometrical model of the
centrosome, considering its role of interface that translates targeting sequences into
their location in cells, through labelled γ-TuRCs: MCs, if assembled with reverse
circumferential sequence (clockwise/counterclockwise), can generate a real true
enantiomorphous organelle (likely the only one in the cell) right-/left-handed, able to
symmetrically translate morphogenetic instructions (topogenic sequences). Let’s
come back to the set (a blind or a visually impaired person and a Braille clock dial
maintained horizontal) which symbolizes biological protractor/goniometer
functioning; let’s suppose that two blind or visually impaired individuals start from
the same point of the plane: one has a normal Braille dial whereas the second has a
dial built with a counterclockwise disposition of the Braille characters (“12” is in
front of him like in the other instrument, but the other eleven numerals are disposed
counterclockwise: “1” is in the sector normally reserved for “11”, “2” in the segment
normally occupied by “10”, and so on); vocal signals indicate the direction of their
steps: the two individuals will execute the received (identical) instructions in a

94
symmetric fashion: their paths will be bilaterally symmetric. Therefore the answer to
the left-right patterning question has been defined “extraordinary”: a little change in a
cell organelle instead of thousands and thousands of changes in morphogenetic
programs is capable of generating bilateral symmetry. Same signals, two symmetric
pathways: same morphogenetic programs, two symmetric organs.
A new hypothesis
- Reverse or opposite rotational polarity of the MC is the base of Left-Right
patterning and bilateral symmetry.
Both the problems about Left-Right patterning and bilateral symmetry are easily
resolved by the centrosome geometrical model: a centriole with a reversible
circumferential polarity (default or opposite, standard or overturned, clockwise or
counter-clockwise) is a clear chiral, enantiomorphous structure; its role as an
interface whose output is the spatial delivering of targeted molecular complexes
assures the realization of bilaterally symmetric architecture (output) as a response to
the same input that drives the default architecture (realized by non-reverse or default
MC).
Mirror symmetry of two 3D objects is very much easier than one might suppose: it
consists of the opposite sign of only one coordinate: any point P (with coordinates “x,
y, z”) belonging to one object, is symmetrical to the point P’ belonging to the
symmetric object (in this case the plane of symmetry is the “y z” plane) whose
coordinates are “ –x, y, z ” (note the sign “–“ before the “x” only); in a spherical
reference system, only the sign of the coordinate changes. The MC of the
centrosome, if built with reverse polarity (mirror symmetric in respect to a “default”
MC) matches each targeting sequence with the same corresponding mark which is
symmetrically positioned in respect to the default MC, and assembles aster and

95
cytoskeleton with the same reverse polarity of its triplets, and this is so for the
disposition of cell landmarks, polarity complexes, receptors and signalling molecules
and, above all for the direction of extracellular fibers: the whole polarization of cells
and tissues is symmetric and the morphogenetic processes of paired and unpaired
organs are carried out mirror-symmetrically: the reverse circumferential polarity of
the MC is transmitted to tissues and organs that grow bilaterally symmetric. In snails,
only one maternal gene is responsible for the left- or right-handed spiral cleavage: it
is surprising that only one gene is able to completely reverse (with perfect mirror
symmetry) the arrangement and orientation of cleavage planes in many many cells,
however mirror symmetry consists only in the sign “-“ before the coordinate “”; a
simple molecular mechanism must necessarily exist capable of reversing MC
circumferential polarity: this matter will be examined more deeply later.
Plants do not show bilateral symmetry or left-right polarity. Unlike animals, plants
and trees do not have evident and true bilateral symmetry, a left and a right half or
side; only the shape of leaves and flowers could seem nearly symmetric (although 2D
only), but the curvature of the two leaf edges (particularly near the petiole and apex),
the edge indentation, the position of petiole, the apex tilt, the arrangement of the
veins that start alternately from the central vein and, above all, meristem histology
and developmental biology exclude the existence of a true bilateral symmetry. Also
the flowers of zygomorphous plants (orchids for example), described as roughly 2D
bilaterally symmetric, after a thorough morphological and developmental analysis,
lead to the same conclusions: bilateral symmetry of paired sepals and petals, as
symmetry of the two halves of unpaired structures (especially for the arrangement of
veins and pigments) is only apparent, not supported by anatomical, histological and
developmental foundation; the two cotyledons (in dicots) have different shapes too;
other symmetries frequently appear, with several reflection planes. A similar apparent
but false symmetry, like in the petals of zygomorphous flowers, appears also in each
of our fingers: a finger seems itself a symmetric object composed of two symmetric
halves and shows an “apparent” midline and an “apparent” (sagittal, proximal-distal)

96
plane of symmetry, but anatomy, histology and embryology do not support this idea:
each finger is mirror symmetric to its opposite finger of the other limb. It is known
that the apparent bilateral symmetry of zygomorphous flowers has an advantage in
attracting pollinator insects (which curiously, take note, are Bilateria): thus, during
evolution, zygomorphous plants have developed quasi-symmetric flowers, modifying
their plans for building laminae and layers, as we will see later. Indeed, plants
dispose, under their cell wall, parallel MT rings that compose a real grid line; so they
can manage somehow both the third dimension and anything like mirror symmetry:
fixing some points of reference along their cortical MTs after cell divisions, and
controlling the distance from the equator, through MAPs, they have the possibility of
using the third dimension (orthogonal to the array of cortical MTs) in order to locate
the spindle poles along the circumference of cortical MTs. It is possible to model how
2D bilateral symmetry could be generated in laminar tissues without centrosomes by
using MTs and MAPs, like plants, devoid of centrioles, or like acentrosomal
planarians: after the first (or another “particular” division in plant meristem or in
regeneration blastemas) the two new cells can mark their respective side of contact:
this is already a midline and both cells, in respect to this side, are bilaterally
symmetric between themselves; this side-mark and the opposite spindle pole position
(symmetric, in the two cells, in respect to the midline), reproduced and transmitted by
the offspring, represent the cues that orient the planes of division; descendant cells
orient the spindle poles following autonomous independent programs (automatically
alternating longitudinal and transversal divisions), to generate division planes
bilaterally symmetric between two close or distant cells having the same lineage but
located one in the “right” and the other in the” left” half. In effect, in plants, this
supposed apparent mirror symmetry is local-depending: unlike animals, in which
bilateral symmetry is globally oriented in respect to the sagittal plane (shared
orientation of centrosomes), the orientation of “quasi-mirror-symmetric” organs
(flowers) is random and local (not global and shared), so they can look up or down,
or toward East or West. Without centrosomes, 3D developmental processes cannot be

97
realized: Planarians do not perform gastrulation. Compare plant development to that
of Drosophila wings (although the fruit fly possesses only an “ancient” and
incomplete type of centrosome with doublets instead of triplets and is a simple
organism with a little connective tissue, without bones and cartilages): when a cell is
genetically engineered to grow faster than others, its clone of descendant cells forms
a larger part of the wing, but the final result are two bilaterally symmetric wings
showing the canonical shape: a “geometrical” interactive cross talking between
genetic programs and extracellular environment has happened to form two
stereotypical normal wings (in addition mirror symmetric). So in zygomorphous
plants, a quasi-bilateral 2D symmetry of flowers can be achieved through non-
regulative development. “Bilateral (3D) symmetry” means that two (possibly distant)
structures (the hands or the ears, for instance) show anatomical, histological, cellular
and sub-cellular mirror symmetry; in the two mammalian cochleae of the same
individual, the general shape is clearly symmetric: furthermore also symmetric is the
disposition and orientation of membranes (basilar, tectorial, Reissner’s), of cells
(Corti’s, Deiters’, Koelliker’s) and of their cytoskeletal MTs too (MT bundles in pillar
cell). In C.elegans, morphogenesis of the intestine shows that the two first cells (Ea
and Ep) divide transversally (the plane of cytodieresis contains the AP axis) and
generate their bilateral counterparts: from now on, right and left cells will have mirror
symmetric shape, polarity, dimensions, movements, orientation of cleavage planes,
and cell-cell adhesions; on the contrary, during C.elegans vulva formation, the first
divisions occur longitudinally (the plane of cytokinesis is orthogonal to the AP axis)
and an “apparent” bilateral symmetry (anterior-posteriorly oriented) appears:
however developmental differences are detectable between the anterior and posterior
halves, the anterior one growing faster than the posterior, and cells have different
dimensions, different cell-cell contacts and adhesions.
As we have already said, centrioles are present in eukaryotic cells, but not in higher
plants and most fungi; unicellular organisms have only centrioles/basal bodies, each
one independent, while in algae and Metazoa two orthogonal centrioles are embedded

98
in the PeriCentriolar Material that forms the centrosome: only the centrosome can be
equipped with left/right MC.
An accurate bilateral symmetry and a definite left-right polarity have been established
only in Metazoa.
Drosophila right and left wings reproduce, with mirror-like feature, the same shape,
edge curvature, compartments and arrangement of tracheae and their anastomoses
(symmetry breaking and superimposition of asymmetry is reserved at the latest stages
of wing formation: see Levin M., Palmer A.R. 2007: “Left-right patterning from the
inside out: widespread evidence for intracellular control”, BioEssays); Ultrabithorax
mutants, instead of halters, develop a pair of additional wings that are built with the
same bilateral symmetry. Inserting the human ortholog gene LHX2 into apterous
mutant embryos (the mutant phenotype lacks wings) the normal wild-type phenotype
is rescued, with two bilaterally symmetric wings (Strachan and Read, 2010); it is
clear that, upstream, the same genes and the same signals act, but downstream there
are two different ways (or, rather, two symmetric tools) left- and right-handed that
symmetrically carry the same instructions out; development confirms that wings
originate from symmetrical primordia, the imaginal discs: after introducing the idea
that MC reverse symmetry is the most likely organizer and builder of metazoan
bilateral symmetry it is not surprising that imaginal discs topology not only
corresponds to that of developed appendages, but is bilaterally symmetric too.
Bilateral symmetry of Metazoa has clear anatomical, histological and developmental
foundations: if Metazoa (bilaterally symmetric) possess centrosomes and plants (non
mirror-symmetric) do not, this cannot be a fortuitous and meaningless coincidence.
Obviously, the inevitable variability of biological processes, due to several cues,
thermal fluctuations above all, especially when enormous numbers of cells are
involved, (very high precision, as we have seen, does not mean identity) can originate
very small differences between left and right: our faces, frankly bilaterally
symmetric, can show a slight (controlled) asymmetry (but our legs have the same
precise -bilaterally symmetric- length!).

99
Bilateral symmetry in Metazoa is more marked than it may appear: it is evident in the
unpaired bones, cranium, vertebral column, pelvis and thoracic cage, but also the
unpaired internal organs (e.g. digestive or respiratory apparatuses), originated from
unpaired (e.g. archenteron) or symmetrical paired primordia (e.g. somites, somatic
and splanchnic mesoderm), keep this symmetry until, during development,
asymmetric movements, rotations and twists, oriented in only one of the two possible
directions (clockwise or counterclockwise), intervene, or developmental adaptations
are superimposed in only one half of the body. Indeed Metazoa are often described as
superficially symmetric but interiorly asymmetric: my personal opinion is that in
Metazoa bilateral symmetry is a fundamental basic property (and an essential
requirement) of their locomotive and sensorineural (movement driving) apparatuses:
bilateral symmetry is the simplest and the most efficient way to assure balance, drive
and control the direction of movements and localize perceived signals (differential
stimulation of two equal bilateral effectors or receptors); without bilateral symmetry,
the control of balance in fast-running/-flying animals (prey and predator change
directions at a great rate) would be a quite difficult problem; this seems to have been
the reason for the extraordinary evolutive success of bilateral symmetry in mobile
organisms; on the contrary, the asymmetry of high brain functions, like hemispheric
lateralization of language or face perception, is superimposed on the basic sensorial
bilateral symmetry; in effect the initial inevitable bilateral symmetry of internal
organs is only a consequence of the establishment of a general symmetry plan
(gastrulation, neurulation, gut formation) but, unlike in locomotive and sensorineural
apparatuses, does not have functional significance: Vertebrates have developed and
adopted asymmetry in many internal organs to solve, for example, the non-
elementary problems of the anatomy and physiology (fluid dynamics) of the great
vessels (systemic and pulmonary circulation). Also in other organs and in other Phyla
limited asymmetries have been described, for instance in Drosophila wings (Levin
and Palmer, 2007), confirming that a downstream process of symmetry breaking is
superimposed to an upstream bilateral symmetry plan.

100
Summarizing, the extraordinary property of two reversely polarized MCs is: “same
input, two symmetrical outputs”.
Bryant (1981: for review see also: Gilbert, 2010, Baker, 2011) proposed the “Polar
Coordinate Model of Positional Information in the Developing and Regenerating
Limb”: in this model, cells are supposed to have a circumferential organization and
adjacent cells “sense” neighbouring signals (recall also the considerations about the
primary cilium and its role as a rotationally programmable –receiving and
transmitting– antenna). When developing tissues or cells, normally non-adjacent, are
juxtaposed, duplications arise: Bryant's polar coordinate model forecasts the
orientation of duplicated limbs; the graft of a left limb bud (or a left regeneration
blastema) on the contralateral right stump, causes three areas of re-growth that
produce three limbs; a central left limb, composed of the transplanted left cells that
maintain their circumferential value (reversed regarding right stump cells) and their
characteristic left-handed tilt and (shared common) orientation of their three axes;
other two abnormal external right limbs growth, composed of stump right cells,
conserving the AP and DV axes typical of the right limb, but with the PD axis
described as rotated respectively of about ± 90°; cells try to normalize circumferential
cell-cell contacts inducing the arising of correctly polarized cells interpolated
between graft and stump cells); cells that are differently patterned (left/right) respond
differently to the same morphogenetic stimulus: they build the forecast structures
(cells maintain the state of differentiation that is permitted in the species: re-growth is
species specific), but through their own polarity; the orientation of the proximal-distal
axis in limb buds depends on the spatial topological position (orientation of
circumferential values) of the neighbouring cells from which the blastema is induced.
Bryant’s “shortest intercalation rule” shows that the juxtaposition of surface receptors
(circumferentially ordered; remember again the considerations made about the
primary cilium) “senses” incorrect cell-cell contacts and stimulates the correct
differentiation and intercalation of cells with correctly oriented rotational polarity.
Bryant’s model completely agrees with the statements of the geometrical model of

101
centrosome functioning: i) cells possess a circumferential global polarity, imposed by
the centrosome which organizes the cytoskeleton and compartmentalizes cell cortex
and membrane (polarity and adhesion factors, signalling receptors); ii) reverse
polarity of the MC is the molecular foundation of left-right patterning and bilateral
symmetry. Bryant's model indicates that the “chiral structure” that determines the
left-right pattern (Xu et al., 2007) and the “cellular component” that specifies the
difference between left and right sides (Vandenberg and Levin, 2009) must possess
not only a simple generic enantiomorphism but a complete “rotational” chirality: this
supports the idea that only the MC (which is circumferentially polarized) has such
properties.
Germ cells and, consequently, every zygote, have a MC built with “default” rotational
polarity, clockwise- or counterclockwise-oriented, depending on the species: in
Xenopus zygotes, Danilchik and colleagues (2006) filmed in vivo their
circumferential asymmetry: rotational equatorial cortical movements
(pharmacologically induced) are oriented in a single fixed (“default”) direction in the
zygote and in parthenogenetically activated oocytes, whereas, after the first mitosis,
the two blastomeres show similar equatorial cortical movements but with opposite
direction between them; a left-right symmetric polarity had been established in left-
and right-handed symmetric blastomeres. Every zygote has the same “default”
pattern: so situs viscerum solitus is the same in every adult, not 50% as expected in
case of random left/right patterned zygotes.
Breaking of bilateral symmetry
During the early stages of development, cilia of cells in the Hensen node (or its
functional equivalent in other phyla) rotate unidirectionally, producing a (default)
flow that activate genes encoding transcription factors only in the cells of the left part

102
of the organism that are reached by this flow. Thus many internal organs in
Vertebrates are asymmetrical: heart, great vessels, lungs, spleen, cerebral cortex,
digestive apparatus and its accessories (liver, pancreas). Situs viscerum solitus is
established: the spleen and the heart on the left, the liver on the right. Studies carried
out on the Zebra fish (Danio Rerio) have filmed the rotation and migration events
that occur during heart early development and that are superimposed on the original
bilateral symmetry. In these organs, originated from primordia with bilateral
symmetry, during a certain phase of development different processes are performed
that involve only one half of the organism: heterochelia, the difference in shape
between the two (right and left) chelae in Crustaceans, is a good example; one of the
two chelae, used in courtship, is larger and has a more suitable shape for cutting,
while the other specializes in grasping. It appears that there is a common “default”
program for both bilateral primordia: later, during development, in the left half this
“default” program is silenced and replaced (or overlapped) by a modified program,
reserved uniquely for the left half (left-reserved program), that conserves the left-
handed modality of execution (carried out by left patterned cells). Errors in this phase
cause “heterotaxia”: in the rare cases of cardiac right isomerism, two both right atria
develop (the “default” program is not silenced in the left half and the left-reserved
program is not activated), while in left cardiac isomerism two both left atria develop
(the “default” program is silenced also in the right half and the left-reserved program
is activated in the right half too): surprisingly, in both cases, the two atria (whatever
they are both right or both left) are mirror-images of each other (bilateral symmetry):
thus in right isomerism there are two symmetric right atria (both with the
characteristic anatomical features of a right atrium) one built by left-handed cells, the
other built by right-handed cells and therefore bilaterally symmetric; and this is so in
left isomerism (Hildreth et al., 2009); this can confirm that cells, determined to be
right (circumferentially polarized by the MC), transmit their polarity to aster and
cytoskeleton, and execute any morphogenetic program in right-handed modality,
whereas left cells translate the same instructions in left-handed modality.

103
It is remarkable the importance of gap junctions between left- and right-patterned
adjacent cells: the pioneering experiments on frogs performed by the first
embryologists (Wilhelm Roux in 1888, Oskar Hertwig in 1893) more than a century
ago showed that if one of the two first blastomeres is burnt off while the other one
continues to sense its membrane receptors (gap junctions), only a half embryo
develops; on the other hand, if the two first cells are separated, each cell can no
longer “sense” the other one through the gap-junctions and two complete organisms
arise. “When separated at the 2-cell stage, Newt embryos exhibit 89% incidence of
organ laterality reversal in one of the twins”. (Takano et al 2007): one blastomere is
“default” patterned, the other one is already oppositely patterned (situs viscerum
inversus). “The sharp midline separation suggests that the first cell cleavage in X.
laevis may produce L and R halves that inherit differential chiral information”
(Vanderberg and Levin, 2009): left and right centrosome have real differential chiral
information contained and “coded” in the reverse sequence of MC non-equivalent
triplets. However, left and right cells with chiral information (during the very first
stages of development) are able to “reset” and restart from an “original” condition: in
fact, 16-cells-blastulae of sea urchin, artificially divided into two halves (one Ventral
and one Dorsal) produce two complete larvae: the Ventral one conserves Animal-
Vegetal and Dorsal-Ventral axes, whereas the Dorsal one maintains only the original
A-V axis, but acquires a new repositioned D-V axis and reverse L-R polarity
(resetting of the cells which will differentiate into blastopore primordium); the graft
of the Spemann organizer shows that the blastopore possesses L-R inductive
capability, being able to induce receptor cells to form duplicated ectopic structures,
bilaterally symmetric along a new midline, from neural tubes to complete heads. In
these experiments the blastopore appears to control the positioning of the midline and
to possess a unique role in managing L-R polarity (symmetry conservation or
symmetry breaking induction); so, normally, it can maintain the natural L-R
patterning with a fixed midline or can intervene to “reset and restart” imperfect
blastulae in order to save very early splitting-events (twins): this is a powerful

104
mechanism that greatly improves the survival of defective blastulae and the success
of the reproduction. The blastopore, unpaired positioned on the midline, its bilateral
symmetric inducing activity and its developmental connection with the Hensen node
(or its equivalent) shows its important role in L-R symmetry and in symmetry
breaking mechanisms; in Mammals, during gastrulation, left-right patterned cells
from the epiblast primitive node (on the midline) invaginate and, directed from the
midline to the Left or Right embryo half, replace the previous hypoblast cells (likely
not left-right determined) to form the embryo endoderm; a second wave of entering
of left-right patterned cells forms the mesoderm, many cells moving laterally and
some directed forwards to form the notochord on the midline, whose inducing
activity is well known (neural plate, somites, intermediate and lateral plate
mesoderm, prechordal plate: all these structures are formed through bilaterally
symmetric morphogenetic processes); also well known is the role of nodal cells in
Left-Right symmetry breaking (unidirectional flow of perinodal fluid due to
unidirectional/default rotation of nodal cells primary cilium). In Bryant’s “Polar
Coordinate Model of Positional Information in the Developing and Regenerating
Limb” it appears that cells try to normalize circumferential cell-cell contacts, by
inducing the arising of correctly polarized cells that interpolate between graft and
stump cells: this indicates a left-right pattering-inducing activity also in limb buds
and in regenerating blastemas (probably triggered by signals from gap-junctions as
we have seen in the experiments of Roux and Hertwig).
Left-right determination is an epigenetic process that has something in common with
sex chromosomes and sex determination: in the two sexes the majority of organs have
the same shape but the morphogenetic programs characteristic of genital organs of
one sex are strictly reserved to and expressed in this one only and silenced in the
other one. Similarly, in the two halves of an organism, many organs are similar
(symmetric) but others are characteristic and strictly reserved to one half only
(chelae, spleen, bicuspid and tricuspid valve, left and right heart ventricle). In both
cases this means that only in one sex or in one half of the body, is access allowed to

105
that part of the genome which is reserved only to that sex or that half. The link
between alterations in the normal functioning of sex chromosomes and left-right
patterning is interesting: it is present in CHILD syndrome (images presented by
Vanderberg and Levin, 2009, free on the Internet) and in gynandromorphism
(astonishing images are presented by Palmer, 2010, also free on the Internet). It is
useful to remember that teratomas, when derived from germ cells, occur in testes or
ovaries, whereas, when derived from embryonic cells, occur on the midline.
Symmetry breaking and asymmetry establishment have singular features in
Echinoderms like sea stars: larvae are clearly bilaterally symmetric (plutei) whereas
adults show an apparent and misleading 5-fold symmetry: indeed the adult is almost
entirely composed of left-half derived cells; rather than a radial symmetry, this
phylum seems to have adopted an unusual circular metamerism (different from the
longitudinal metamerism, like segments in D.melanogaster, or somites in Vertebrates)
and to have enormously amplified the left-right asymmetry. Also in fruit flies left-
right patterning shows singular and surprising features: as we have seen in Foe’s
study, the domains described occur as pairs and, paired or not, each domain is
bilaterally symmetric: so a clear midline is evident dorsally and ventrally; the
preblastodermic syncytial development in Drosophila is not so unusual: it is a
bilateral symmetric blastula quite similar to that of other Metazoa (Xenopus, sea
urchin, etc.) but with blastomeres without a membrane: its polarity and global
compartmentalization is dictated by its centrosomes.

106
Discussion of the model
Critical points of the proposed geometrical model
The previous theoretical structural hypotheses about the centrosome have highlighted
how accurately they fit in with several geometric, morphogenetic and thermodynamic
requirements of complex organisms composed of very large numbers of very small
units (cells): for instance, bilateral symmetry and left-right patterning can be easily
resolved by the centrosome geometrical theoretical model in agreement and accord
with the statements of cell and evolutionary biology. However, from the proposed
(theoretical) model some important criticalness (biochemical feasibility) emerges and
questions arise: then answers (and new questions) must be searched for and, if it

107
possible, found.
Here our aim is to ascertain the biological, biochemical and biophysical foundation of
our hypotheses.
Q&A
Above all there is the problem of the non-equivalence of the nine centriole triplets:
what is the biochemical basis of such an unusual biological process, the assembly of
an ordered sequence of nine similar but non-equivalent structures?
In addition, other queries:
i) The symbols for the hours on a clock are orderly sequenced (circumferential
polarity) and the symbol for “12 o’clock” is always at the top to guarantee the global
orientation of the whole structure: which known biochemical mechanism can create
the circumferential polarity of the nine centriole triplets, ordering, arranging and
aligning them in accordance to a predefined sequence and a pre-established global
orientation?
ii) In the proposed model, the reverse orientation of centriole polarity is the basis of
3D bilateral symmetry: which known biochemical mechanism can carry this function
out? In other words, how is it possible to assemble in an opposite sequence
(clockwise > counterclockwise) nine radial spokes and nine centriolar blades?
iii) The centrosome has been supposed to be spherical while the PCM is organized in
cylinders, coaxial to the centrioles: how is the correct quasi-spherical -TuRC
inclination achieved?
iv) How can the centrosome map the whole cortex if the nucleus, to which it is often
close and which is not crossed by MTs, hides more than half of the cortex?

108
v) How do the macromolecular complexes (3-6 nm in diameter), destined for a
precise position in the cortex or the cell membrane, quickly find their respective -
TuRC receptors, interspersed among many others over the large (2x106 nm
2)
centrosome surface?
i) Molecular basis of triplet non-equivalence (centrosome software)
From Geimer and Melkonian (2004) ”The ultrastructure of the Chlamydomonas
reinhardtii basal apparatus: identification of an early marker of radial asymmetry
inherent in the basal body”:
“At the distal end of the basal body (bb), exactly at the transition between the
microtubular triplets of the bb and the doublets of the transitional region, a filament
of about 10 nm diameter is attached to the microtubular doublets (rarely triplets) in a
rotationally asymmetric pattern linking A-tubules of doublets 7, 8, 9, 1 and 2. From
doublet 2, the filament arches back to doublet 7, passing through the lumen of the bb.
The overall outline of this closed structure resembles that of an (capless) acorn with
its broad base located between doublets 1 and 2, and its pointed tip at doublet 7. At
doublets 2 and 7, the acorn has a prominent knob through which linkage with the A-
tubule is established. The side of the acorn that is not attached to bb doublets/triplets
will, in the following, be called the `lumenal' side of the acorn. A second, V-shaped,
system of filaments of about 8 nm diameter is present in this region of the bb. Two
filaments extend from triplets (rarely doublets) 4 and 5 into the bb lumen, bisect the
lumenal side of the acorn and, in the center of the bb lumen, connect to another
filament that runs perpendicular to the two filaments, extending down from the
proximal end of the stellate structure in the transitional region. The positional
relationship of the acorn to these filaments is best seen in longitudinal sections
through bbs. In such sections, the acorn is seen as two asymmetrically positioned

109
electrondense dots. The filaments extend from triplets 4 and 5 at the level of the acorn
and bend downwards with an angle of about 35° after reaching the lumenal side of
the acorn. They continue to the other side of the bb, making contact with triplets at
the level of the distal connecting fiber attachment with the bb. In cross sections
through bbs, depending on the sectional plane, these filaments can be seen to contact
triplets 8, 9 and 1.
Most interestingly, both systems of filaments were already present at the distal end of
the probasal bodies.”
The last words completely agree with the geometrical model of centrosome
functioning: the process of centriole duplication through the characteristic orthogonal
disposition and the MC role as a platform are considered the mechanisms responsible
for transmitting the same coordinated orientation of rotational asymmetry
(correspondence of MC and DC “0° marks); as we have seen in “Centriole/Basal
bodies in Ciliates” (on page 53, about Paramecium) the distal end of the basal body
(immediately under the axoneme) is the same end which, during centriole formation,
is close to the pre-existing centriole, used as a platform for self-assembly; a new
centriole, once correctly and coordinately oriented by its mother/platform, can be
easily inserted into the fragile structure of the cytoskeleton maintaining a coordinated
mother/daughter orientation without causing any distortion: 4000 basal bodies, made
up of nine different triplets each one having high propensity for binding different
elements of the cytoskeleton, must be guided and oriented to avoid a tangled jam. To
underline its importance, we have compared before this apparently strange but
fundamental and unique process of centriole duplication to the introduction of a new
trolley-bus into the aerial-suspended electric town-circuit or to the insertion of an
integrated circuit onto a printed circuit board.
We will come back to this theme.
The idea underlying the completion of the geometrical model of the centrosome has
something in common with the process of coordination of the rotational polarity of
the arising procentriole and the MC; the idea is that the biochemistry of non-

110
equivalence does not consist in a structural difference of triplets (or radial spokes),
but in a mechanism able to differentiate and “decorate” nine identical sectors,
apposing nine different and orderly sequenced molecular complexes, which, in turn,
distinguish and render different both triplets and radial spokes. Thus, non-equivalence
is not imposed by the tertiary structure of the centriolar blades and spokes, on the
contrary their quaternary structure is responsible for their non-equivalence. A ring is
certainly the most likely structure and the “acorn” described by Geimer and
Melkonian is particularly appealing; a ring of an informational macromolecule able to
contain information (a linearly ordered sequence of equi-spaced markers) appears to
be the most suitable structure: a ring of RNA? Indeed no polypeptidic ring has been
found in centrioles and centrosomes, whereas “within the core of the basal body a
twisted or looped 90 Å diameter fiber, or more probably pair of fibers, RNase-
sensitive, in association with dense granules is observable (Dippel, 1976):
1 RNA loop + 9 protein dense granules = 9 RiboNucleoProteins (RNPs) markers.
The work “Structure and non-structure of centrosomal proteins” (Dos Santos et al.,
2013) presents the finding that “centrosomal proteins tend to be larger than generic
human proteins since their genes contain in average more exons (20.3 versus 14.6).
They are rich in predicted disordered regions, which cover 57% of their length,
compared to 39% in the general human proteome… These residues predicted to be
both disordered and coiled-coil may represent disordered regions that lack stable
structure unless they interact with a binding partner, and take coiled-coil structure
only upon binding”. Disordered protein domains frequently bind nucleic acids
(Vuzman and Levy, 2012): then the propensity of centrosomal proteins to bind RNA
is exciting (like CLERC, centrosomal leucine rich and coiled coil containing proteins,
studied, as we will see soon, by Yoshinori and Yukio: leucine rich domains and coiled
coils have high propensity to bind RNA): RNAs appear to be the fittest candidate to
be the biochemical basis of the centrosome “software”. Nine triplets (and also nine
cartwheel radial spokes), if structurally identical, can be easily self-assembled around
the SAS-6 ring without a fixed order of appearance, and, in a later moment, each of

111
them acquires its own individuality: so identical triplets and spokes become
distinguished (and therefore non-equivalent) because of the presence of nine different
marks (like RNA hairpins, stems, loops, etc.) aligned in an orderly sequence, easily
coded, memorized and stored in a linear macromolecule. A ring of RNA is therefore a
very good candidate to realize the molecular basis of triplet non-equivalence: nine
diverse marks can be linearly coded in DNA (realizing an orderly sequence) and
distanced by “spacers”; the mark defined as the “0° mark” (the reference mark used
to orient the ring like the numeral “12” on a clock) can be formed by the
juxtaposition of the two ends of the molecule, then in a fixed position in respect to the
other eight marks and arranged in the centriole under the control of the MC used as a
platform: this fits in with the idea that the reason of the strange centrosome cycle
consists in the process for coordinating shared orientation). Many biochemical
pathways constitute the complex cross-talking between DNA and centrosome to
control and regulate the cell cycle: an RNA ring, transcribed but not translated, is
faster and fitter than a protein. A DNA sequence transcribed into an RNA ring allows
nine RNA-based markers (receptors), different from one another, to be arranged and
equally spaced around the SAS-6 ring (cartwheel): other PCM macromolecular
complexes, like miRNAs or RiboNucleoProteins (RNPs) recognize these triplet
markers in order to be arranged into that sector of the centrosome which is radially
contiguous to (or in front of) each centriolar blade (Fig 6). Indeed self-assembly is a
property characteristic of identical macromolecules, monomers, or of a few different
units (heterodimers and heterotrimers) that form the quaternary structure of proteins:
nine MT blades, which, in addition, are different and must be orderly sequenced, are
too many to be self-assembled together, especially because the precise constant
sequential order is a very difficult and frankly unlikely self-assembly process. “To get
deeper insight into centriolar wall formation, we examined the order of appearance
of the nine A-microtubules in the nascent procentriole. The formation of the first A-
microtubules was not directly related to a specific position. Moreover, the position of
each A-microtubule at the centriole wall did not correlate with its size, arguing

112
against a sequential growth process. These findings suggest that each A-microtubule,
and thus each blade in a procentriole, assembles in an independent manner”
(Guichard et al., 2010). According to this finding, the nine triplets are, originally,
structurally identical: after their (random-arising) self-assembling, they become
different and orderly distinguishable because of the decoration by an RNA ring. As
already cited, “In C. reinhardtii the polypeptide VFL1 coded by the gene vfl1
(Variable number of FLagella), binds only to the triplet N° 1” (Silflow et al., 2001):
evidently in C. reinhardtii the triplet N° 1 has its own unique and distinguishable
triplet marker. Yoshinori and Yukio (2010) “identified a centrosomal protein called
CLERC (Centrosomal leucine-rich repeat and coiled-coil containing protein) which
is a human ortholog of Chlamydomonas Vfl1 protein. The bibliography as well as
database searches provided evidence that the human proteome contains at least seven
centrosomal leucine-rich repeat proteins including CLERC. CLERC and four other
centrosomal leucine-rich repeat proteins contain the SDS22-like leucine-rich repeat
motifs, whereas the remaining two proteins contain the RI-like and the cysteine-
containing leucine-rich repeat motifs. Individual leucine-rich repeat motifs are highly
conserved and present in evolutionarily diverse organisms”. Coiled-coil containing
proteins and leucine-/cysteine-rich regions have highly propensity to bind RNA.
Centrosomal RNA
“The centrosome and spindle as a ribonucleoprotein complex” (Alliegro, 2011).
“An intriguing morphological and developmental parallel between certain
procentriolar elements and an RNA-containing component of the Paramecium
kinetosome occurs during centriolar production and development in embryonic chick
tracheal epithelial cells” (Dippel, 1976).
Hiedemann and colleagues (1977) reported their observations on the formation of an
incomplete and not functional aster, following the implant of basal bodies of
Chlamydomonas Rheinardtii in Xenopus eggs: after the treatment of the basal bodies
with weak concentrations of RNase, the formation of a partial aster was found to be

113
completely suppressed and only the capacity for lengthening of the centriole MTs
remained; the -TuRC function (nucleation of astral MTs) appeared to be sensitive to
the RNase, whereas only the MTs of the triplets of the centriole wall, after removal of
the CP110 cap, could still grow in length (in vitro) because of the addition of α and β
tubulin dimers at the “+” end. What is the functional association between RNA and
centrosomal -TuRCs ? Laane and Haugli (1974) had already studied the association
of RNA with centrosomes in Physarum, and Hartman and colleagues (1974) in
Tetrahymena pyriformis; Snyder (1980) showed that RNase inhibited microtubule
nucleation in PtK1 cells. Finally centrosomal RNAs (cnRNAs), not coding and absent
from the remaining cytoplasm was identified in the centrosome of Spisula
Solidissima (organism clearly bilaterally symmetric) (Alliegro et al., 2006).
Chichinadze and colleagues (2012) have continued the studies on Spisula cnRNAs
concentrating on characteristics that are particularly worthy of in-depth examination,
following the observation by Alliegro and colleagues (2006) that “a substantial ORF
[Open Reading Frame] was readily mapped in the antisense strand. The predicted
polypeptide encoded in this ORF includes a highly conserved 200 amino acid-long
reverse transcriptase domain. This domain exhibits striking homology (P=e-75
) to Zea
mays putative polyprotein, a member of the RNA-dependent nucleotide polymerase
family of proteins, Pfam00078. In addition, a ribonucleoprotein consensus RNA-
binding site (RNP-1) was found downstream from the reverse transcriptase domain.”
The importance of RNA for centrosome to work correctly emerges from another
study (Ishigaki et al., 2013): “RBM8A (Y14) is carrying RNA-binding motif and forms
the tight heterodimer with MAGOH. The heterodimer is known to be a member of
exon junction complex on exporting mRNA and is required for mRNA metabolisms
such as splicing, mRNA export and nonsense-mediated mRNA decay. Almost all
RBM8A-MAGOH complexes localize in nucleoplasm and shuttle between nuclei and
cytoplasm for RNA metabolism. Recently, the abnormality of G2/M transition and
aberrant centrosome regulation in RBM8A- or MAGOH-deficient cells has been
reported. These results prompt us to the reevaluation of the localization of RBM8A-

114
MAGOH in human cells. Interestingly, our immunostaining experiments showed the
localization of these proteins in centrosome in addition to nuclei. Furthermore, the
transiently expressed eYFP-tagged RBM8A and Flag-tagged MAGOH also co-
localized with centrosome signals. In addition, the proximity ligation in situ assay
was performed to detect the complex formation in centrosome. Our experiments
clearly showed that Myc-tagged RBM8A and Flag-tagged MAGOH formed a
complex in centrosome. GFP-tagged PLK1 also co-localized with Myc-RBM8A. Our
results show that RBM8A-MAGOH complex is required for M-phase progression via
direct localization to centrosome rather than indirect effect”.
Hypothesis: circumferential polarity is generated by an RNA ring
“Structural and genetic studies suggest that asymmetry of the centriole (basal body)
plays a critical determining role in organizing the internal organization of algal cells,
through the attachment of microtubule rootlets and other large fiber systems to
specific sets of microtubule triplets on the centriole…To understand cell organization,
it will be critical to understand how the different triplets of the centriole come to have
distinct molecular identities” (Marshall, 2012).
“The overall outline of this closed structure resembles that of an (capless) acorn with
its broad base located between doublets 1 and 2, and its pointed tip at doublet 7. At
doublets 2 and 7, the acorn has a prominent knob through which linkage with the A-
tubule is established” (Geimer and Melkonian, 2004).
“Within the core of the basal body a twisted or looped 90 Å diameter fiber, or more
probably pair of fibers, RNase-sensitive, in association with dense granules is
observable…The apparent extraction, from the above structures, of RNA-containing
material by pronase and of protein-containing material by RNase is particularly
puzzling and intriguing… The apparent removal of both RNA and protein from this
basal body structure by either of the two corresponding enzymes suggests an unusual

115
organization of the two components.… authors suggest that the filament might
contain "informational macromolecules" for assembly of the cores and centrioles… If
the RNA plays a structural rather than a translational role, then does it provide
scaffolding or dictate the order of assembly of the subunits in protofilament
formation, or serve as a recognition center for protein aggregations giving rise to
nucleation sites? Or does the RNA have some unique morphogenetic function which
our present preconceptions about assembly mechanisms have not permitted us to
envisage?”(Dippel, 1976). The “acorn” of Geimer and Melkonian and the “looped
fiber” of Dippel show similar diameters: 10 nm (acorn) and 90 Å (remember the
equivalence: 1 nm = 10 Å).
“Perhaps less tangible but still a likely cause for lingering controversy is that the
presence of nucleic acids in the spindle or centrosomes will require us to look
differently at these structures from a functional, and more to the point, evolutionary
standpoint ” (Alliegro, 2011).
Warning: the non-equivalence of the triplets concerns exclusively the “software” of
centrosome functioning, i.e. γ-TuRCs labelling; the orientation and inclination of γ-
TuRCs (centrosome “hardware”) is not controlled by the mechanisms examined in
this section.
To better develop the geometrical model of centrosome functioning, it is necessary to
deeply analyze the idea that the biochemistry of non-equivalence does not consist in a
(biochemically unlikely) structural difference of triplets (or cartwheel radial spokes),
but in a single linear macromolecule - RNA - capable of assuming different 3D
conformation: such molecule is able to bind different proteins (markers or receptors)
in order to decorate the base of the centriole and differently label the nine triplets and
the nine spokes, juxtaposing nine different 3D structures (ribonucleoproteins, RNPs:
Figs. 7 and 8) to spokes and triplets: as we have seen, the idea that the non-
equivalence of triplets and spokes does not consist in their different structure or
molecular composition (primary, secondary and tertiary structures are identical and
equivalent: this is a more likely and canonical property of the centriolar blades), but

116
in the collocation of nine different polyribonucleotidic-marks at their base that can, in
turn, bind different proteins (non-equivalence of the quaternary structure) seems
plausible and convincing.
A single-stranded filament of RNA, because of its chemical-physical properties and
the frequent modification of the bases, exhibits, like proteins, a variety of
thermodynamically favourable secondary structures (hairpins, stems, loops, knots,
pseudoknots: Fig. 7): these, in turn, create particular 3D tertiary configurations.
Transfer-RNA is a well-known example: its secondary (two-dimensional) cloverleaf
structure (three different stems, each ending in a loop, and a fourth variable loop), is
encoded linearly (i.e. one-dimensionally sequenced) in DNA and in the pre-tRNA
transcript; a 3D tertiary “L” shaped conformation similar to a chaise longue is the
final result after splicing: these properties enable it to interact with extreme precision
with its cognate amino-acyl-tRNA-synthase and with the appropriate site of the
ribosome. This is an evident discrete system, highly resistant to noise, based on codes
recognized by their 3D structure, with an almost negligible margin of error. Similar
secondary and tertiary structures have been highlighted also for introns and in many
small nuclear and nucleolar RNAs. “The acridine-orange fluorescence studies
showed reduction of the yellow-green basal body fluorescence by RNase but not by
DNase, and a shift from yellow-green (indicative of double-stranded RNA or DNA) to
orange-red (indicative of single stranded RNA) when staining followed
mercaptoethanol or pronase treatment” (Dippel, 1976): a ring of single-stranded
RNA with nine different double-stranded hairpins or knots (Figs. 6, 7) linked to
proteins (RNPs) is strongly consistent with these observations. Centriolar triplets
non-equivalence, supported by RNPs, is transmitted to the whole neighbouring
sector of the corresponding PCM and especially to the elaborate scaffold of γ-TuRCs:
γ-TuRCs are similar to cones, with the circumference of the base composed of a γ-
tubulin ring; γ-tubulin is incorporated into a heterotetramer (γ-tubulin small complex
or γ-TuSC) which, in turn, is composed of two molecules of γ-tubulin and two other
conserved proteins, DGrip84 -91 in Drosophila; [Grips means γ-Ring Proteins also

117
called GCPs, acronym of γ-Complex Proteins; other different Grips, Dgrip-75, -128
and -163 in Drosophila, link γ-TuSCs to organize -TuRCs]; pericentrin fibrils
(Mennella, 2012) and SAS-4 (Gopalakrishnan, 2011) connect radially centriolar
blades and γ-TuRC scaffolds.
As before stated, to satisfy the basic requirements of a spherical reference system
organizer, in order to recognize coded geometric signals and translate them into their
desired final locations, “cellular protractors/goniometers” must possess: 1) different
marks; 2) constant ordered sequence of marks; 3) one start mark; 4) controlled spatial
orientation of the entire structure. A single-stranded RNA ring has all the
characteristics to be the molecular platform of the non-equivalence of the nine
centriolar blades: (a) it is memorized linearly in DNA, which can thus encode nine
different molecular labels; (b) these different molecular labels (hairpins or knots) are
orderly (linearly) sequenced and equally spaced (DNA spacers), suitably distanced so
that they fit in structurally with each of the nine spokes/triplets; this is comparable to
a strip of conveniently distanced numerals that follow each other in an orderly
manner and that can decorate the face of a clock, comprising the characteristic
numeric ring; (c) an RNA ring is equipped with structures that allow it an
unambiguous orientation: like the acceptor stem of tRNA, 5’ and 3’ cnRNA ends can
be used as anything like a handle-stem that identifies the so-called “0°” reference or
start-mark (it can be compared to the figure “12” which is always at the top on a
clock). Indeed a mechanism to form a ring of single-stranded RNA, connecting its
ends (3’ poly(A)-tail and 5’ m7G cap) is used during the first stages of translation in
eukaryotes: several poly(A)-tail-binding proteins, joined to the 3’mRNA poly(A)-tail,
are bound by the eukaryotic translation Initiation Factor 4G (eIF4G), while the
5’mRNA m7G-cap is bound by eIF4E; both eIF4G and eIF4E are joined by eIF4B,
producing a circular molecule (often very large) of mRNA. It is well known that
many regions of human non protein-coding DNA are transcribed into pre-mRNA:
cnRNA11 (see later) possess 3’UTR- poly(A)-tails and 5’UTR- m7G caps and can be
assembled into a ring conformation, following usual mRNA processing.

118
-------------------------------------------------------------------------------------------------------
Fig. 7 RNA 3D structures.
Because of its chemical-physical characteristics and the frequent modification of its bases, an RNA
molecule, from its earlier linear configuration (A), acquires diverse local secondary, thermodynamically
favourable, structures (stems, loops, hairpins, knots as showed in: B > I) with canonical and non-canonical
base-pairings, and corresponding different tertiary 3D structures
-------------------------------------------------------------------------------------------------------
ii) Chirality and bilateral symmetry: RNA, through an event of
retrotransposition, might have generated the reverse polarity of the MC
Let’s summarize again the hypothesis about left-right patterning, which will be faced
here. In the geometrical model of centrosome functioning, the MC is considered the
chiral structure responsible for realizing bilateral symmetry: indeed the MC,
polarized with reversed, opposite circumferential sequence (clockwise/
counterclockwise), seems to be the only rotationally enantiomorphous organelle, the
unique biological structure able to play a geometrical role in left-right patterning and
bilateral symmetry establishing, which are indisputably geometrical processes.

119
Centrosomes, with opposite polarity of the MC, are geometrical interfaces which
receive the same input (signals with the information for the value of the coordinate)
and translate them symmetrically (output); during organogenesis, they transmit their
intrinsic chirality to each mirror symmetric organ. As we have already said, only the
MC has the property of being assembled with opposite polarity, on the contrary, the
DC has always the same rotational polarity in every cell: mirror symmetry changes
only the longitude ( coordinate) of a point, never its latitude (θ coordinate). An MC
with reversed sequence of the triplets and therefore, with circumferential polarity,
organizes a “counterclockwise polarized PCM”, transmitting its reverse polarity to
the γ-TuRC receptors. In each meridian wedge of the “reverse” centrosome, the γ-
TuRC receptors remain structurally the same but are bilaterally symmetric in respect
to the “default” centrosome, like in a ”reverse” clock dial the numeral “1” takes the
place of the numeral “11”, “2” takes the place of “10” and so on; longitudinal -
TuRC receptors are always the same but reversely (symmetrically) positioned: their
3D configuration, their shape and form maintain their characteristic and remain
unaltered and unchanged. -TuRC inclination too remains unaltered and so the
associated latitudinal receptors: in a clock dial decorated by Roman numerals, “II”
identifies a sector positioned at “60° -clockwise rotation- from XII”, whereas in a
reverse/symmetric dial (overturned) the Roman numeral “II” does not change its
shape, only identifies a mirror symmetric position “60° -counterclockwise rotation-
from “XII”) [Overturning Roman numerals creates some problem: IV,VI,VII,VIII,
and IX, when overturned, are different; not so if we make use of capital letters: A, H,
I, M, O, T, U, V, W, X,Y]. The reversely polarized centrosome, compared to a
“default” centrosome, has the extraordinary property to translate the same DNA
morphogenetic geometric instructions – 3D signals or targeting sequences - into 3D
bilaterally symmetrical locations: one input > two reversely rotationally polarized
MCs > two symmetric outputs. Note that left and right centrosomes are built through
a variation in a part of their “software” (opposite or reverse polarity of longitudinal -

120
TuRC receptors) whereas their “hardware” (-TuRC scaffold inclination and
orientation) remains completely unaltered. This hypothesis fits in very well with
evolutionary biology statements: a simple, small, easy change is much more likely
than conspicuous and large genomic variations: to change only the sequence of nine
marks of an RNA ring is very much more likely than to collect, store and conserve “a
lot” of variations in “a lot” of morphogenetic programs, until two complete sets of
instructions (one Left and one Right, coding for the correct bilateral symmetry of “a
lot” of symmetrical organs) make it possible to build an entire bilaterally symmetric
organism: and this for millions species (million is much more than “a lot”). Ease
passes the examination of natural selection: like in computer programming simple
languages and concise algorithms are preferred, so in evolutive competition easy
solutions are more stable and sure than complicated programs, and, to say, “win the
award”.
A question: to be effectively evolutionarily likely, a change must be simple, small and
easy: how “simple”? At this moment of our analysis, to reverse a sequence of
nucleotides does not seem so “easy”.
Each geometric signal for the longitude or angle “” (encoded in the related targeting
sequence), recognized by a -TuRC receptor on a counter clockwise labeled
centrosome, is decoded and translated as the symmetrical negative angle “-”,
whatever is its angle θ (its latitude). By reversing the only angle , perfect bilateral
symmetry is realized: “Basal bodies/centrioles are an excellent candidate for an
intracellular “F-molecule” [the chiral molecule hypothesized by Brown and Wolpert
(1990)], because they function as a microtubule-organizing center to direct
asymmetric localization of maternal components” (Levin and Palmer, 2007).
Left/right centrosomes are capable of building bilaterally symmetric cytoskeletons
that wire and map cell cortex compartments in a corresponding mirror symmetric
fashion.
As just seen, the biochemistry of non-equivalence does not consist in structural
differences of triplets (or radial spokes), but, more likely, in a single ring of RNA: to

121
change only the sequence of nine marks of an RNA ring is easier than to change the
orderly sequence of nine bulky radial spokes or nine different awkward triplets, each
made of three long, hulking, unwieldy MTs: how “easier”? What known biochemical
mechanism can easily carry out the overturn of this ring or the reversal of its
nucleotide sequence, indispensable to realize an opposite, reversed polarity in the
MC? In other words, how is it possible to assemble in an opposite sequence
(clockwise > counterclockwise) nine RNA/RNP structures which decorate nine radial
spokes and nine centriole blades? In this respect it is convenient to imagine the RNA
ring like a very large molecule of tRNA having 9 loops in lieu of 3 and with a short
handle-stem similar to the tRNA acceptor stem, containing different 3’ and 5’ ends
(like a plug equipped with two differently shaped prongs or blades or pins).
What known biochemical mechanism can carry out the reversion of MC polarity?
“Gedankenexperiment” (thought experiments)
This term, introduced by the Danish physicist Hans Christian Ørsted, became famous
through Einstein’s and Schrödinger’s mental (or conceptual) experiments.
We too will perform three virtual experiments in which an observer is in front to
different couples of similar (near identical) objects: two similar thin plugs (having flat
large faces and two short pins), two similar house keys (having a large bow like a thin
disk and a very short blade), two similar images of a tRNA secondary structure,
designed on a sheet.
1) Both plugs have the name of the manufacturer impressed on one face only, and
nothing impressed on the opposite face: both pugs lay on the plane of a table, have
the pins pointing toward the observer, and show the side with the manufacturer’s
name; watching them the observer sees the name of the manufacturer on both the
plugs and notes a little difference in the pins: one plug (that we name “A”) has on the

122
left (of the observer) a short and thick pin, whereas the other pin is longer and
thinner; the second plug (“B”) has similar but reversed pins: the short and thick one is
on the right (of the observer), the longer and thinner on the left; to enter into their
socket which has two vertical holes (at the top the large one, at the bottom the small
one) the pins must fit in well with the holes; after entering, the observer sees that the
name of the manufacturer in the “A” and “B” plugs appears on the mirror (or
opposite) side, on the left side in one, on the right side in the other: the plugs appear
as symmetric.
2) Both keys are a little strange: they have a very large bow (disk shaped) and a very
short flat blade; on one side of the bow there is a “head”, on the other a “cross”;
watching the keys, the observer sees that both show the same side with the head but
in the blades (which point both toward him) the observer notes a little difference: one
key (that we name “A”) has a blade showing at the left (of the observer) a smooth
straight border or edge, whereas the other border has a knob or a little tooth; the
second key (“B”) has its blade edges reversed: the knob or tooth is at the left of the
observer; to slide into the keyhole or keyway (supposed horizontal) and enter into the
lock, the keys must have the knob on the same side, on the right of the observer (Fig.
8); after entering, the “heads” in the “A” and “B” keys are on the mirror/opposite
sides (one up, one down) and the observer sees “head” on one key bow and “cross”
on the other; the bows (disks) have been overturned.
3) Both tRna secondary structure images are designed with the anticodon loop at the
bottom, the “D” loop at the left and the “T” loop at the right of the observer; only a
little difference in the stems(top) is perceived by the observer: in one image (named
“A”) the terminal sequence “CCA” is at the right of the observer in the 3’ tRNA end,
while in the figure “B” the terminal sequence “CCA” is at the right of the observer, in
the 5’ tRNA end; the acceptor stems can enter and interact with the active site of the

123
amino-acyl-tRNA synthase only if the “CCA” sequence is (we suppose, here) on the
right (of the observer); after taking the correct position into the enzyme housing, in
both secondary RNA structures, the sequence “CCA” will be at the right (of the
observer) and the two molecules will show “T” and “D” loops symmetrically located
in respect to the observer and an usual anticodon in one image while in the other it is
opposite (reversed).
Fig. 8 Centrosome theoretical geometrical model: chirality.
A: a key with nine clockwise sequenced numbers on the edge of its bow, and with a short blade that has a
bulge or a knob pointing toward the number “3” can correctly fit in its lock; B: a second similar key built
with a reverse, symmetric, blade (the bulge points towards the number “9”) has the same clockwise sequence
of numbers on the edge of the bow, but cannot enter (large X) in its lock; C: after overturning the second
key, it becomes compatible with its lock but the sequence of the 9 numbers on the edge of the bow is now
reversed and symmetric to the ring in “A” (the nine numbers are counter clockwise sequenced).
D, E: an RNA molecule has all the characteristics to be the molecular basis of chirality: it is memorized
linearly in the genome, which can thus encode the ordered sequencing of nine different molecular “labels”
(see also Fig.6); this molecule can assume a 3D shape made of a ring of 9 orderly sequenced “labels” and a
“stem” (used to orientate the ring) which fit in the housing (active site) of its scaffold ; two cnRNA rings

124
with the same sequence, but having their short “handles/stems” symmetric (D,E), after fitting into the
housing structure are reversely oriented, showing a bilaterally symmetric sequence of their nine loops,
hairpins etc.
These experiments demonstrate that very little changes only in a small part of a large
object are responsible for the overturn of the whole object: by changing only four or
five nucleotides in a tRNA molecule it is possible to realize the overturn of the entire
molecule. As already observed, a little evolutive change is more likely than a big one.
A ring of RNA composed of a little stem and a large circle (like a tRNA with nine
loops), can be easily front > rear overturned like a coin (head > tail): this overturned
ring, with a short stem and a large circle containing 9 equi-spaced different 3D loops
is symmetric in respect to the not overturned ring (Fig. 8); what molecular process
can overturn an RNA ring? Changing few nucleotides at the ends of a RNA sequence
is three time easier than for an polypeptide sequence (one amino-acid is coded by
three nucleotides): from this point of view an RNA ring is better than a protein ring.
The Brown and Wolpert’s “F” chiral structure

125
Fig. 9 theoretical geometrical model: chirality.
A: model of the cnRNA default ring (here arbitrarily named “right”); its handle/stem (3’ + 5’
juxtaposed ends) is the “F” Brown and Wolpert chiral structure.
B: “left” version of the cnRNA ring; the sequence of the nine markers is unaltered, identical to A;
the only difference consists in the reverse (or opposite) direction of the two horizontal hyphens of
the capital letter “F”.
C: the same “left” ring of B after the overturn necessary to position correctly the F shaped
handle/stem to fit in well and interact with the housing of the corresponding molecular structure: the
sequence of the nine markers is opposite, clockwise>counter-clockwise reversed, mirror (or
bilaterally) symmetric.
Let’s imagine, as just seen, the RNA ring and its handle-stem like a common house
key having only the border (or the circumference) of the bow (a large “ring shaped”
bow instead of a “disk shaped” bow) with nine different (radially outward or inward
directed) jutting notches, and a short asymmetric blade (something similar to a big
“F”); to better explain this concept, let’s suppose that the nine different jutting
notches have shapes which do not change when they are head>tail reversed or
overturned, like the following capital letters “A; H; I; O; M; T; V; X; Y” (more
convenient than Roman numerals, we before used in a clock dial); the large key bow
(with its nine different notches/letters) symbolizes the large cnRNA ring with nine 3D
structures (loops or hairpins shaped in such a way that their form remains the same
when they are overturned) while the short blade is the short cnRNA stem (Fig. 9).
This handle stem is exactly the Brown and Wolpert’s “F” chiral structure composed

126
of a vertical segment and two horizontal hyphens: in the “default” configuration
(which, here, we name “right”) these hyphens are in the usual position showed by the
letter “F”, whereas in the “left” configuration they are reversely (or oppositely)
oriented. The Brown and Wolpert’s “F” chiral structure is made up of few nucleotides
belonging to the 3’ and 5’ ends of the cnRNA transcribed sequences.
This is a first answer to the precedent question: “how simple is this small, easy
change?” The change is small because it concerns only a minimum part of the whole
object. Probably the easiest solution of the problem of two opposite or reversed –
clockwise > counterclockwise – sequences of RNA consists in a very little difference
of the stem/handle: two identical large RNA rings with nine marks, identically
orderly sequenced, but with little differences in their short 3’-5’ termini, so that their
two short stems have symmetric 3D conformations; these two rings can be managed
as two symmetric RNA rings (Fig. 8); when their symmetric stems are arranged in
such a way that their 3D configurations appear identical to the observer and are both
compatible with the housing-structure in which the stems must be positioned (the
active site of a molecular complex), the two RNA rings are overturned, head-tailed,
symmetric, with opposite and reversed -clockwise > counterclockwise- sequence of
their 9 double stranded structures. These nine 3D loops, like the jutting notches/letters
of the key bow, after overturning can interact with the same molecular complexes that
interact with the not overturned loops. Then, little differences in the terminal ends of
the cnRNA ring (about 1% of nucleotides are different) realize the overturn of the
entire RNA ring that can easily decorate symmetrically the MC with “opposite”,
reversely oriented, clockwise > counter clockwise disposition of its nine marks.
Something similar can be observed in cytoplasmic dynein, where two monomers
containing a ring-shaped structure and a stem, bind together, juxtaposing their stems
in such a fashion that the rings are symmetrically disposed (one “default”, one
overturned).
Summarizing: the mirror symmetry of the short handle-stems of two large identical

127
rings (like two protractor/goniometers) is responsible for completely overturn the
entire structure (protractor/goniometers oppositely graduated).
What mechanism can perform a little change only in the ends of a sequence,
maintaining conserved the whole long sequence? The most likely (known) process is
a transposition event, followed by independent evolution of 5’ and 3’ ends: two loci,
having the same sequence and free to evolve separately, maintain the same general
sequence through purifying selection while a little change, permitted only in their 3’-
5’ ends, can create two short stems having a symmetric 3D shape: it is not necessary
that the two terminal sequences are precisely identical and reversed because different
short sequences of RNA, after covalent modifications of their nucleotides and non-
canonical pairing, can assume the same 3D shape in order to realize two 3D
symmetrical configurations compatible with the housing-structure in which the stem
must be positioned (Fig.8). This is a second answer to the precedent question: “how
simple is this little, easy change?” Such an evolutionary event able to introduce
bilateral symmetry in Metazoa is extremely easy, little and simple, then very likely:
mutation of only a few nucleotides at the ends of the DNA sequence coding for
cnRNA. Paraphrasing Descartes‘famous sentence “Cogito, ergo sum”, we may say:
“Facile, ergo verisimile” [easy, therefore likely]. Indeed it is quite more conceivable
and plausible that a single and simple, casual, random, fortuitous event (mutation of
four or five nucleotides) is the base of bilateral symmetry of “all” the organs (paired
or unpaired) of “all” metazoan species, instead of an infinite number of complicated
casual mutations (in millions of species and for each organ) able to “write” bilaterally
symmetric building programs, choosing, step by step, the proper symmetrical cell
location.
Left ring and Right ring, left MC and Right MC, Left centrosome and Right
centrosome, Left half and Right half of organisms. The overturned RNA ring
transmits its opposite, reversely oriented sequence of marks to the PCM surrounding
the MC and to the already orientated -TuRCs (as we have said, orientation and

128
inclination of -TuRCs in left and right centrosome are the same; only the longitude
labels of -TuRCs are symmetrically positioned): from the reverse MC, -TuRCs
receive reversely ordered longitude receptors: two identical cnRNA rings with
opposite sequence of their 3D structures create two bilaterally symmetrical
centrosome; bilateral symmetry is not a property of -TuRC orientation but a property
of -TuRC longitude labelling. “The data are consistent with models in which the
early cytoskeleton is nucleated by a chiral structure that orients itself with respect to
the other two axes, and thus biases the intracellular transport of key determinants
along the LR axis” (Lobikina et al., 2012). “In the intracellular model, an early
component of the cell-polarity system (likely involving the cytoskeleton) orients the
transport of key molecules within cells that ultimately creates an LR difference in the
embryo… This intracellular model shows how physiological mechanisms integrate to
produce large-scale LR gradients from initial subcellular polarities. Although the
orienting cytoskeletal element is not yet known, this model provides a comprehensive,
quantitative synthesis of all the molecular and biophysical steps leading from LR
orientation within single cells to asymmetric gene expression in the early embryo,
and it does not depend on ciliary motion” (Levin and Palmer, 2007).
My personal opinion is that in Bilateria, during very early cleavage stages, probably
during the first zygote division itself (but this depends on the Clade), Left-Right
pattern is defined, and a midline imposed, through a genetic/epigenetic process
memorized in each cell, so that many genes are, at the same time, switched “ON” in
left cells and “OFF” in right, while other are switched “OFF” in left cells and “ON”
in right: the difference between “Left” and “Right” cells consists only in the state
“ON/OFF” of probably only one upstream gene (or an enhancer/silencer sequence)
which controls a cascade of many other genes and DNA sequences (some of them are
“ON” in Left cells and “OFF” in right and others are “OFF” in Left and “ON” in right
cells). So, some signal molecules, produced only in left cells and deciphered by
receptors directionally positioned on a polarized primary cilium of right cells, or
transmitted through gap junctions (Levin and Mercola, 1999) interact with nuclear

129
receptors (expressed only in the right half), producing a “left-right interactively cross-
talking” in order to confirm and maintain a correct midline and to face events of
embryo splitting by restarting, resetting or recovering left/right patterning.
Downstream there are different cascades of left/right gene expression: lefty and
nodal, for example, are expressed normally, only in the left half. One component of
this differential cascade is the transcription, from only one of the two different loci, of
the Left or Right cnRNA molecule, which drives the assembling of Left or Right
MCs and allows Left cells to carry morphogenetic instructions out, mirror
symmetrically compared to Right cells. This transcription event not necessarily must
occur already at the stage of two blastomeres, it is simply one of the events that
belong to the Left or to the Right cascade of downstream left/right genes: in mice,
centrosomes appear at the stage of 64 blastomeres, but cells maintain epigenetic
memory of their Left or Right identity, previously acquired.
“Yet one key question remains: how is the chiral cytoskeletal architecture interpreted
to localize ion transporters to the left or right side?... we found that endogenous
Rab11 mRNA and protein are expressed symmetrically in the early embryo. We
conclude that Rab11-mediated transport is responsible for the movement of cargo
within early blastomeres, and that Rab11 expression is required throughout the early
embryo for proper LR patterning” (Vandenberg et al., 2013).
Left-right determination, as we have already seen, is an epigenetic process that has
something in common with sex chromosomes and sex determination: Left and Right
cells run the same morphogenetic programs in a mirror symmetrically fashion
(bilateral symmetry) or perform different programs which are reserved only to one
half of the organism and, consequently, asymmetrically expressed (asymmetry).
Then, during later stages, in the development of some organs, Left-Right bilateral
symmetry is broken (in a species-specific fashion, not always conserved) and
asymmetry is defined (situs viscerum solitus/inversus) in such a way that cells in the
left half follow different developmental instructions than “right” cells and run left-
reserved programs (differential cascades of DNA transcription and signalling

130
pathways, “ON/OFF” switched genes). Perturbing the cytoskeleton, the primary
cilium, and, above all, MT structure causes problems to the mechanism (the
centrosome “hardware”) that firstly imposes and, later, breaks Left-Right symmetry:
“Recent findings in Arabidopsis thaliana have shown that mutations in α-tubulin and
in a γ-tubulin-associated protein (Tubgcp2 [a -TuRC component]) play an important
role in the symmetry properties of the plant axial organs. Wild-type A. thaliana axial
organs do not twist during normal elongation, and its flowers are radially
symmetrical. This symmetry can be broken by mutations in tubulin and tubulin-
associated protein complexes. The tubulin mutations spiral1, spiral2, and spiral3
produce right-handed helical growth mutants. Lefty (lefty1 and lefty2) mutants were
found to be suppressor mutants of spiral1 and when outcrossed displayed a
prominent left-handed helical growth. Both α-tubulin and γ-tubulin complexes are
ubiquitous in eukaryotes and are involved in the formation and nucleation of
microtubules. Here, we characterize the laterality phenotypes induced by the same
mutations in a vertebrate (the frog Xenopus laevis), the nematode Caenorhabditis
elegans, and mammalian cells, supporting a fundamental role for tubulin in the cilia-
independent generation of LR asymmetry” (Lobikina et al., 2012). Left-Right
“asymmetry” (that consists in “bilateral symmetry” breaking), imposed by a
genetic/epigenetic mechanism, is performed through any chiral structure (primary
cilium or other centriolar or MT derived structures). As we have already said chirality
is not the same as “bilateral symmetry”, it is the basis to build bilaterally symmetrical
organs; α-tubulin and γ-tubulin mutations interfere with MT rings disposition in
plants and with centrosome building in Metazoa.
Assuming that cnRNA is the organizer for the creation of circumferential polarity, the
concepts of non-equivalence of the centriolar triplets and chirality (left-right
patterning) overcome the barrier of their biochemical feasibility.

131
cnRNA11 bioinformatics
Alliegro and colleagues have described various centrosomal RNAs (cnRNAs),
including two cnRNA11: an mRNA11 of 3863 nucleotides (nt), GenBank:
DQ359732, coded in the Plus strand and another (“Minus” strand coded) RNA11,
GenBank: DQ363378, of 638 nt. Data banks conventionally supply the sequence of
the Plus/+ filament, numbering nucleotides from 5’ to 3’. Here “long dcnRNA11” is
the short form for “DNA locus coding for the long cnRNA11”: in this case the locus
is on the Plus/+ strand; “coding locus” here is the sense, non-template and non-
transcribed 5’>3’ sequence, whereas its antiparallel complementary strand is the
antisense, template and transcribed 3’>5’ sequence; “short dcnRNA11” is the short
form for “DNA locus coding for the short cnRNA11”: in this case it is on the Minus/-
strand. The words “sense/non-sense and template/non-template” (referred to the
entire filament) are not used because transcription may happen on both filaments,
depending on each locus: the long dcnRNA11sense sequence is on the Plus filament,
whereas the short dcnRNA11 sense sequence is on the Minus; Plus and Minus strands
are antiparallel (and so are their respective numbering by data banks): if imagined
like written lines (as presented by data banks) in the Plus strand the order of the
letters is (and numbering increases) from the left end to the right end (5’>3’) whereas
in the Minus strand the letters ( and numbering) start from the right end. Aligning
(with blastn algorithm) these two dcnRNA11 coding sequences, the longest one (from
the 2255th nt to the 2880th nt of its Plus filament) corresponds to the sequence from
the 630th nt to the 3rd of the shortest one (Minus filament), with clearly reverse
alignment (Minus/Plus), a non-perfect correspondence (93%), 6 indels (in-sertion del-
etion) and, above all, the substitution of some original terminal nucleotides (2 from
the 3’ end, 8 from the 5’ end of the short dcnRNA11): these are therefore two very
similar but not identical sequences, then coded and stored in two different loci, one,

132
long, in the Plus strand and the other, short, in the Minus strand. Pay attention to the
alignment direction: both sequences, although not identical but very highly similar,
have their corresponding nucleotides aligned in the same 5’>3’ direction, respectively
on the Plus and Minus filament: as we have said these sequences are antiparallel,
left>right on the Plus strand, right>left on the “Minus” one, but both 5’>3’. They are
not perfectly identical to each other (especially because of the two deletions and four
insertions in the short dcnRNA11), and are reversely oriented in a sense as we might
expect from a Plus > Minus retro-transposition event (transposition does not change
the nucleotide sequence: original and transposed loci, if read in the same 5’ > 3’
direction (that in Plus and Minus strands are opposite or antiparallel) have the same
identical sequence; therefore the reverse alignment of two sequences, one on the Plus
filament and the other on the Minus is only apparent). Taken together these findings
support the retro-transposition hypothesis, which maintains 5’ > 3’ order, requires the
intervention of nucleases (reverse transcriptase is an RNase family member) and,
strikingly and surprisingly, completely agree with the previous considerations about
the little changes in 3’-5’ ends necessary and sufficient for an RNA ring to create two
short 3D symmetric stems. Therefore, we must deduce that the 638 nt cnRNA11 is
not a part of the 3863 nt cnRNA11: these two dcnRNA11 loci, after transposition,
have independently evolved with significant changes (6 indels in little more than 600
nt are not negligible), however maintaining their substantial sequence identity
through purifying selection (except for the ends). The different ends of the transposed
short cnRNA11 can assemble a stem with a 3D shape that is bilaterally symmetric in
respect to the stem of the original cnRNA1: it is not necessary that the two terminal
sequences are precisely identical and reverse because not identical short sequences of
RNA, as we have already said, after covalent modifications of their nucleotides and
non-canonical pairing, can assume similar 3D shapes; after entering into the
housing/active site of the MC, the transposed cnRNA11 ring is necessarily overturned
(Fig. 8) and the sequence of its nucleotides is opposite and reversed. The long
cnRNA11 is transcribed from its antisense non-template (in this case the 3’ > 5’

133
Minus filament) strand, and shows a sequence corresponding to its 5’ > 3’ sense
(“Plus”) coding strand (as known RNA polymerases “walk” on the antisense template
filament in direction 3’ > 5’ and synthesize a new RNA chain from 5’ to 3’), whereas
the short cnRNA11 is transcribed from its antisense non-template (in this case the 5’
> 3’ Plus filament) strand, read by RNA polymerase from the 3’ end, and shows a
sequence corresponding to its 3’ > 5’ sense (“Minus”) coding strand (Fig. 8). [Do not
confuse the expression “ 5’>3’ “ when it is used to show a direction and when it is
used to identify a DNA strand: the 5’>3’ filament is the leading “Plus” filament
during DNA replication, whereas the 3’>5’ filament is the lagging or “Minus”
strand]. Exploring the nucleotides which flank the matching sequence in both
dcnRNA11, very surprising findings emerge: the two stems show a really symmetric
3D sequence because of the symmetric position of a consensus sequence. It is better
to establish some point of reference: let us consider as “START” (the beginning of
the region of similarity) the sequence (5’>3’: left>right) “TGACG” which in the long
dcnRNA11 Plus (coding) strand goes from the 2255th to the 2259th nt, and as
“STOP” (the end of the region of similarity) the sequence (5’>3’: left>right)
“GCTAG” which in the long dcnRNA11 Plus strand goes from the 2876h to the
2880th nt (Fig. 9). On the Plus strand of the short dcnRNA11 (the only one reported in
data banks; in this case it is the non-coding antisense template filament) we find the
sequence (5’>3’: left>right) “CTAGC” (3rd >7th nt) that is complementary but
opposite (antiparallel) to the previous “STOP”, and the sequence (5’>3’: left>right)
“CGTCA” (626th >630th nt) that is complementary and reverse (also antiparallel) to
the previous “START”. Reading the Minus (coding) sense strand of the short
dcnRNA11 (Fig. 9) and following the direction 5’>3’( now right>left), we can read
identical sequences as in the long dcnRNA11: the “START” “TGACG” sequence
(630th >626th nt; Fig. 8 reports the sequence 3’ > 5’; numbering is always the same
as the Plus strand) and the “STOP” sequence “GCTAG” (7th >3th nt). If now we
read (always 5’>3’) the nucleotides which are after the “STOP” sequence on the long
dcnRNA11 Plus strand, we find “TTGAGTTA”. A closely corresponding (5’>3’:

134
right>left) sequence precedes the “START” in the short dcnRNA11 Minus strand
“ATGGAAG”: because their opposite antiparallel alignment (5’>3’ vs 3’>5’), the
sequences (both read in 5’ > 3’ direction) “AGTTA” and “AGGTA” are surprising
similar (consensus sequence) but positioned on the opposite ends (3’ end of the long
cnRNA, 5’ end of the short cnRNA: Fig. 9). If we represent the two rings of
cnRNA11 with their stems corresponding (not symmetric, both showing the
consensus sequence at the right of the observer, like in the third virtual experiment),
their sequences are clearly reversed (Fig. 10): the overturn of the RNA ring is
obtained through only little nucleotide differences in the stem. in effect only very few
nucleotides are normally enough to precisely fit in with the active site of a
polypeptide.
Still more extraordinary is the fact that the antisense filament that codes the long
3863 nt cnRNA11, along its length, contains a sequence similar to a reverse
transcriptase, therefore indicating the concrete possibility of a retro-transposition
event. But there are still more surprises because, as we have already seen, “a
ribonucleoprotein consensus RNA-binding site (RNP-1) was found downstream from
the reverse transcriptase domain” (Alliegro et al., 2006): this long mRNA appears to
be similar to a LINE (Long Interspersed Nuclear Element, a not Long Terminal
Repeat retro transposon) even if shorter. Thus, the previous analysis confirms the
possibility that a transposition has originated the short cnRNA ring which can be used
in an overturned fashion (front > rear or head > tail in respect to the corresponding
similar sequence of the long cnRNA11), creating a symmetric decoration of the MC:
this might represent the (easy, simple and evolutionary likely) biochemical basis of
bilateral symmetry: transposition has generated two DNA loci (cnRNA-coding) that
have evolved differently, generating two similar cnRNAs, displaceable, through their
diverse ends, in two different symmetrical fashions (head/tail, front/rear).
What length should we expect for an RNA ring that ensures a receptor at the base of
each triplet? The ring composed of nine SAS-6 N-terminal dimers (Kitagawa, 2011)
has a diameter of 23 nm and a circumference of 72 nm; the ring formed by 9

135
complete SAS-6 dimers (with their radially arranged coiled-coil C-terminal domains)
has a diameter of 43 nm and a circumference of 135 nm; (the “acorn” of Geimer and
Melkonian has a diameter of 10 nm: it is possible that the RNA ring is inscribed into
the SAS-6 ring); estimating a linear length of 0.34 nm occupied by each nucleotide in
a linear polynucleotide chain, and about 16 bases for each of the 9 stem-loops (as in a
tRNA arm), 400 nt should be sufficient to cover the entire circumference of 9 SAS-6
N-terminal dimers and to generate the 9 signals (600 nt for the complete ring, C-
terminals included). These dimensions correspond to those of the various cnRNAs
described by Alliegro and colleagues (Alliegro et al., 2006, 2010; Alliegro and
Alliegro, 2008; Alliegro, 2011), except for the long 3863 nt RNA, whose meaning
seems much more general; in fact: 1) it can no longer be seen after the first zygote
division; 2) it is present during the formation of the first two centrosomes which, in
the author’s model, in agreement with the observations of Danilchik and colleagues
(2006), Levin and Mercola (2007) and Venderberg and colleagues (2013), distinguish
the first two blastomeres in a Left-Right sense (in species whose first blastomeres
contain centrosomes); 3) it includes the reverse transcriptase; 4) it contains a
sequence that is quite similar but not identical to the 638 nt cnRNA 11.
“In Spisula oocytes, most of these [cnRNAs] are not detectable before oocyte
activation, but several were found to be present in unactivated oocytes as a distinct
hybridization patch associated with the nucleolinus. As the nuclear envelope breaks
down, the newly formed centrosomes are found to be embedded within, or closely
apposed to, the nucleolinar patch…The nucleolinus is an RNA-rich compartment,
closely apposed to or embedded within the nucleolus…. The nucleolinus is a unique
cellular compartment containing centrosome and spindle-associated RNAs” (Alliegro
et al., 2010). Because of this long cnRNA11 (3863 nt) location, the nucleolinus (in
which firstly centrosomes are embedded) appears to be the (temporary: only once per
cell cycle) nuclear location for pre-cnRNAs maturation (like the nucleolus is the
location for pre-rRNAs maturation and spliceosomes are the locations for pre-
mRNAs maturation): using several gene-finding programs - GenScan (MIT pattern

136
scanning gene finder), GeneMark (Georgia Tech gene finder), NCBI ORF finder and
FGENESH (HHM-based gene finder from Soft Berry - to predict exons + introns,
this long cnRNA11 shows to possess introns and then it may reasonably be
considered as a very long and immature pre-cnRNA.
How many different types of cnRNA should there be in the centrosome? We
obviously expect three distinct types of cnRNAs, one for the MC (invertible to
support the Left-Right difference: therefore 2, slightly different, MC cnRNAs) and
one invariable, for the DC. These three cnRNAs should have the duty of marking
each radial spoke and each triplet in order to be able to transmit the non-equivalence
to other structures, like the PCM wedges. Other cnRNAs may comprise the receptors
associated to the -TuRCs (hypothesizing, as seems logical, that these receptors are
also polyribonucleic or ribonucleopolypeptidic): 9 different cnRNAs would comprise
the receptors for the longitude (nine centrosomal meridian wedges) and 5 more for
the latitude (two centrosomal caps and three parallel rings); a further receptor that
“captures” all the macromolecules that must be distributed in the cell by the
centrosome and display a centrosomal localization targeting sequence (see below:
centrosome trafficking) can also be envisaged. These are 18 different cnRNAs; it
cannot be excluded, and is in effect very probable, that the wiring system generated
by centrioles, centrosome and aster MTs, if used also in a centripetal sense (with
dynein molecular motors), as already mentioned, might require a similar number of
small RNAs (located near the -TuRCs) that would accompany molecular messengers
coming from particular compartments of the cell membrane (and taking the
fundamental information of “cortical compartment of provenience”) to the nucleus;
this makes it possible to code the geometrical information (cortical compartment of
origin) into a molecular structure (a particular miRNA able to bind with suitable DNA
sequences or with other proteins involved in the regulation of the genes). In their
work, Alliegro and colleagues (2008) found a similar number of different cnRNAs:
“This subset of 39 sequences that exhibited database identities could be divided into
three categories. The largest group (19) matched uncharacterized BAC [bacterial

137
artificial chromosome] clones, hypothetical proteins, or other uncharacterized
chromosomal sequences from a wide variety of species. Sixteen of the 39 identities
were related to nucleic acid or genome structure and metabolism. Examples include
nucleotide polymerases (cnRNAs 65, 118), an Entamoeba snRNP (cnRNA48), clam
(Venus) and pig microsatellite sequences (cnRNAs 123, 154, 226), and retroelements
from a variety of species (cnRNAs 15, 93, 142, 171). The third category containing
only four sequences had no clear relationship to centrosomes, the cytoskeleton, or
nucleic acid metabolism...A subsequent screen for centrosome-associated RNAs in
Ilyanassa embryos resulted in the identification of approximately 50 additional
molecules” (Alliegro 2011).
A) Short dcnRNA11 “Plus” 5’ ag|ctagc…………cgtca|cttccatc 3’
B) Short dcnRNA11 “Minus” 3’ tc|gatcg…………gcagt|gaaggtag 5’
C) Long dcnRNA11 “Plus” 5’ ttcatctaa|tgacg... ..gctag|ttgagttac 3’
D) Long RNA11 3’ AUU|ACUGC……CGAUC|AACUCAAU 5’
Short RNA11 3’ CUACCUUC|ACUCGC……CGAUC|GA 5’

138
E) Long RNA11 3’ AUU|ACUGC……CGAUC|AACUCAAU 5’
Short RNA11 5’ AG|CUACG……CGUCA|CUUCCAUC 3’
Fig. 10.1 dcnRNA11.
“START” sequence is underlined; vertical bars separate the matching sequence.
A) Plus / + / antisense / template strand locus for the short cnRNA11 (638 nt): 5’ (left) > 3’(right).
B) Minus / - / sense / coding strand locus for the short cnRNA11 (638 nt): 3’(left) > 5’( right).
C) Plus /+ / sense / coding strand locus for the long cnRNA11 (3863 nt): 5’ (left) > 3’(right).
D) 5’ (left) > 3’(right) long and short cnRNA11 alignment: “START” and “STOP” coincide. Consensus
sequence (bold) at opposite ends.
E) Opposite (reversal) alignment of both cnRNA11: “START” and “STOP” are at opposite ends (reversed).
Consensus sequence (bold) at the same side.
long cnRNA11 short cnRNA11
5’ 3’
U U
A A
A C
C C
3’ U 5’ U
A:::::::C A:::::::U
U:::::::A G:::::::C
U:::::::A c A
A c u C
C u a U
U a c G
G c g C
C g .. ..
.. .. .. ..
.. .. ……………
…………… Fig. 10.2 cnRNA11 stems.
When the stems of both short and long cnRNA11 correspond (consensus sequence in bold) the two rings are
overturned: “START” and “STOP” sequences are mirror symmetric.

139
Further researches have been carried out on the 3863 nt cnRNA11: the short sequence
between the 2054th and the 2089th nt
“CTGGTGCTGTTGCTGCGGTTGCTGCTGTTGCTGCTG”
has an enormous diffusion in all the genomes and is present in dozens loci in all
organisms. If we examine this sequence, it does not appear to be similar to any
known localization targeting sequence (mitochondrial, lysosomal, etc. actually
present in “Signal Peptid Resourches Data Base” [proline.bic.nus.edu.sg/spdb] and in
“Prediction of signal peptides” [www.predisi.de]), it is not similar to a “CG isle”, and
cannot be compared with sequences often found in introns, enhancers and silencers; it
does not contain tandem repeated trinucleotides (trinucleotide tandem sequences in
Homo sapiens, although not frequent nor very long are much more ordered, with very
few, if any, variations in the repeated trinucleotide sequence).
It seems not to be itself, in toto, a repeated sequence (no repeats found using
“Tandem repeat finder Program”): it is 98% identical, from nt 1 to 36, to Jockey-
6_DK, (from nt 511 to 546) which is a non-Long Terminal Repeat /Jockey in
Drosophila kikkawai, defined as a "apurinic-like endonuclease and reverse
transcriptase"; this strengthens the supposition that the long cnRNA11 is a non-LTR
retrotransposon. Inserting this cnRNA as a query and using “blastn” to look for
generic correspondences both in the “Nucleotide collection” and in the “Genome”
databases (specifying each time the restriction of the research to one single model

140
organism) a great number of correspondences appears every time; it is astonishing
that this sequence is the only section of the long cnRNA11 with such an abundant
correspondence: being a brief sequence, there is always the suspicion that it is a case
of random correspondence, but in 3863 nt there should be other 36 nt sequences that
are randomly aligned with various other sequences of several genomes. For example:
by using” blastn” to seek alignments corresponding to this 36 nt sequence (from the
2054th to the 2089th nt of the long cnRNA11) in the chromosomes of a model
organism like Mus musculus (taxid:10090), we obtain similarities in dozens of
different loci for each chromosome, belonging to many proteins (some of which very
interesting because they concern, for example, histone-methylases, transcription
factors, nuclear receptors, extracellular sulphatases) or situated at different distances
from genes; there are correspondences also in short interspersed repeated sequences
(SINE), this last circumstance being very interesting from the point of view of
development and morphology, if we think that the morphological differences between
canine races are almost exclusively dictated by genomic differences relating to
SINEs, as demonstrated by Wang and Kirkness (2005).
Alliegro consider this cnRNA 11 an mRNA: therefore, trying to translate this
sequence into a polypeptide and, considering that the lack of a start codon does not
allow an Open Reading Frame (ORF) to be identified, we obtain the following 6
amino-acid sequences:
1) 5'>3' Frame 1st: L V L L L R L L L L L L;
2) 3’>5' Frame 2nd: S S N S S N R S N S T;
3) 5'>3' Frame 3rd: G A V A A V A A V A A;
4) 3'>5' Frame 4th: Q Q Q Q Q Q P Q Q Q H Q;
5) 5'>3' Frame 5th: W C C C C G C C C C C;
6) 3'>5' Frame 6th: A A T A A T A A T A P.
The meaning of these short polypeptides is very problematic: it may be a question of
a consensus sequence of centrosome localization, similar to those present on the
fibers surrounding nuclear pores in which phenylalanine and glycine, both non-polar,

141
alternate; five sequences are either completely hydrophobic (the first, third and fifth:
Leucine, Alanine or Cysteine) just like the filaments connected to nuclear pores or
decidedly hydrophilic (the second and the fourth: Serine and asparagiNe or
glutamine, Q); the sixth sequence alternates two amino acids, but one is hydrophobic
(Alanine) and the other is hydrophilic (Threonine) producing an amphiphilic
sequence. Since this 36 nt sequence recurs in positions far away from the genes, it
seems probable that it is part of regulatory domains and, then, non-coding and
untranslated.
In conclusion, the idea that non-equivalence is a property of the quaternary structure
of nine triplets with identical tertiary structure seems to be supported by common and
known biological mechanisms; same observation can be made for the second idea,
the reversion of the orderly sequence of non-equivalent markers, whose opposite
disposition may be the structural base of bilateral symmetry. Then, if both ideas are
sustained by simple and common biochemical processes, they cannot be speculative
suppositions: they appear plausible. The findings and evidences discussed, taken
together, turn the proposed ideas into something more than a hypothesis. I hope.
iii) PCM molecular architecture (centrosome hardware)
“In top-views of centrioles, of the 18 proteins analysed in this study, 4 (Centrin, SAS-
6, STIL and Plk4) revealed compact dots. All other proteins were disposed in ring-
like patterns around the centriole and, as shown for γ-Tubulin and NEDD1, CPAP
showed both ring and central dot localization” (Sonnen et al., 2012).

142
Now it’s the turn of the centrosome “hardware”. In the centrosome, the two centrioles
maintain their orthogonal configuration during S and G2 phase and in the first stages
of mitosis: at this time the centrosomes and their PCM are formed and acquire all
their geometrical properties.
The main task of both metazoan centrioles appears to be the assembly of the PCM;
the MC is also responsible for forming cilia and maintaining centrosome orientation
during G1: in non-dividing cells the MC becomes the primary cilium basal body.
The centrosome, which has long been described as a spherically-shaped organelle, on
the contrary, appears to be not spherical but structured in cylindrical, coaxial, layers
around the centrioles; then two considerations must be made:
- the aster is a spherical structure and the direction of the astral MTs is clearly
spherical-radial, centrifugal: this implies that the orientation of -TuRCs must be
parallel, at the point of nucleation of each MT, to the centrosome “spherical /
polyhedral” surface (or rather, parallel to the local surface-tangent plane); the surface
would then be spherical and the PCM too would therefore have a spherical /
polyhedral shape; [warning: here and later, the word “γ-TuRC” stays for “the ring-
shaped base of γ-TuRCs”: only this ring, lying on a plane, can be “parallel” to a
tangent plane; the word “parallel” would be meaningless if referred to the whole
structure of a γ-TuRC, similar to a cone].
- in contrast, the organization of the PCM around the MC is indisputably cylindrical:
Sonnen and colleagues (2012), using 3D-structured illumination microscopy to
analyse the molecular architecture of the PCM, have demonstrated that its structure
consists of coaxial layers around the MC, confirming a similar description already
expressed by Ou and colleagues (2002). The quantity of PCM around the DC is much
less than around the MC and the position of the DC is decidedly eccentric (Fu and
Glover, 2012). The DC too organizes its modest PCM circumferentially and
orthogonally to the MC.

143
Both centrioles possess a 9-fold symmetry: the PCM and the whole centrosome,
organized by them, must necessarily conserve the same 9-fold symmetry. The MC
organizes circumferentially the PCM and subdivides it into 9 sectors (meridian
wedges) corresponding to the nine MT blades of the MC (Fig. 6; this schema is
confirmed by the images presented by Mennella and colleagues 2012, free on the
Internet); the DC also organizes circumferentially the PCM and subdivided it into two
caps and three parallel rings or discs (Figs. 6B, 6C,).
Then there are two main geometrical problems to be addressed and faced:
(1) MTs show an astral, centrifugal, radiating 3D (spherical) and, above all, non-
intersecting direction: then, a biochemical mechanism must exist capable of orienting
-TuRCs in such a way that each one is parallel to the centrosome’s surface tangent-
plane in order to nucleate a radially oriented microtubule towards the cell cortex: in
other words, -TuRCs do not show the same cylindrical symmetry as centrioles, on
the contrary each -TuRC shows an inclination that is the addition of two
inclinations, one (longitude) imposed by the MC, one (latitude) by the DC. (2) The
longitude inclination and the relative labeling information can be transferred radially
(cylindrically) by the MC whose position in the PCM is central: not so for the
latitude-information that from each DC blade must reach all the -TuRCs included on
the same cap- or disc-sector (comprised between two “parallels”) and positioned on
different meridian wedges: a biochemical mechanism must exist capable of
transmitting the same “latitude-information” to -TuRCs which have the same
latitude but different “longitudinal location”.
A logical explanation might be that the spherical / polyhedral shape is not a property
of the whole centrosome but that it is restricted only to -TuRCs, whereas the
centrosomal PCM structure consists of roughly cylindrical, coaxial layers around the
MC. It seems that -TuRCs are orientated in two step modality: whatever their
latitude, because of the organization imposed by the MC and its central pivotal

144
position inside the centrosome, like a hub, -TuRCs are initially parallel to the MC
blades (centrioles should be imagined as prismatic rather than cylindrical), acquiring
the orientation corresponding only to the longitude (Fig. 6 A): the contribution of the
DC would then consist in revolving -TuRCs, so as to transmit and superimpose the
orientation relative to their latitude. This hypothesis is based on the observation that
the aster is composed of MTs that do not intersect (O’Toole et al., 2012) but have an
ordered radial, 3D (spherical, centrifugal) direction, necessarily resulting from the
orientation of the -TuRC from which each MT originates: like orientation (longitude
+ latitude), like direction (or, rather: one -TuRC “discrete” orientation, one MT
“discrete” direction; one-to-one correspondence between -TuRC “discrete”
orientation and MT “discrete” direction). Molecular complexes capable of orienting
each -TuRC must therefore exist and the cylindrical organization of the PCM around
both centrioles agrees with the hypothesis of centriole-driven orientation of γ-TuRCs;
γ-TuRC structure fits in well with this hypothesis: its centripetal portion is a long
cone that is inserted into the PCM and therefore comes in contact with various other
molecules. “Several centrosomal proteins have γ-TuRC anchoring function at the
centrosome, including pericentrin B, CG-Nap (also known as AKAP350 and AKAP
450)….Additional proteins including PCM-1, CEP135 or BBS4 are required for the
formation and maintenance of a radial microtubule array anchored at the centrosome
in interphase” (Delgehyr et al., 2005); “Complex containing the centrosome proteins
CAP350 and FOP, and EB1, has also been proposed to play a role in anchoring MTs
at the centrosome” (Azimzadeh and Bornens, 2007).
It seems, as before observed, that Metazoa have improved the molecular control of
three quantities (already well managed by Protists): the right angle, the angle of 140°
(and its supplemental of 40°) and MT length: combining these three geometric
quantities by assembling their biochemical supports, they can manage the architecture
to realize and regulate the double inclination (longitude and latitude) of centrosomal
-TuRCs. Control of the angle of inclination corresponding to the longitude and

145
latitude can only (or, rather, must necessarily) derive from the transmission of the
orientation corresponding to the geometry of the centriolar blades (9-fold symmetry):
each MC blade is in effect already oriented (parallel) towards the corresponding
wedge/meridian sector, and each DC blade is already oriented towards the
corresponding cap or a parallel disk (Figs. 6, 2); transmission of this information
must take place through simple known mechanisms: “The centrosome from G1 cells
is composed of a MC and a DC linked by a matrix. Matrix assembly is assumed to be
triggered by centrioles through two subsets of microtubule-binding proteins, one able
to bind centriole proximal minus ends, and one able to bind centriole walls in a
polyglutamylation-dependent manner….A large pericentriolar cloud of additional
components can be accumulated in a microtubule-dependent manner, leading to an
extended boundary” (Bornens, 2002). The presence of -TuRCs with different
functional value has been suspected for a long time (Fouquet et al., 1998): some of
them act as MTOCs (nucleating cytoplasmic MTs) while others only have functions
in organizing the centrosome. “The dual distribution of the γ-tubulin could indicate
two different abilities of MT nucleation. PCM γ-tubulin is considered to be involved
in the nucleation of cytoplasmic MTs. Centriolar γ-tubulin must be involved in the
unknown functions of the centrioles … Moreover this γ-tubulin γ-TuRCs labelling [by
immunogold electron microscopy] was initially present only in centrioles and then it
becomes predominant in the PCM” (Fouquet et al., 1998). The role of this “unknown
functions of centriolar γ-tubulin “ must be faced. From two different populations of γ-
TuRCs, one of centriolar identity (nucleation of centrosomal MTs that remain
confined in the centrosome), and one of PCM identity (nucleation of cytoplasmic
MTs), arise two diverse populations of MTs, PCM-MTs, confined and restricted in
the PCM, useful to impose the correct orientation to each γ-TuRC scaffold and
transfer to each γ-TuRC, through kinesin-mediated transport, the receptors of
longitude and latitude, accordingly with γ-TuRC position in respect to MC and DC
triplets, and cytoplasmic-MTs that reach the cell cortex. It is important remembering
that several MAPs (e.g.Tau and MAP-2, both neuronal, or MAP-4 that is expressed in

146
many cell types) have a typical “right angle” shape, with a side parallel and
longitudinally bound to the MT and the other orthogonal to the MT axis: one or more
“right-angle MAPs” at the tip of a radial “centrosomal, PCM-restricted” MT can be
the molecular structure able to orientate γ-TuRCs orthogonally to the “centrosomal
virtual radius”. The cylindrical organization around the MC may occur by
maintaining and transmitting the geometrical subdivision of the centriole wall and its
9-fold symmetry. The poor use of the DC as a cytoplasmic-MTOC in G1, and the
presence of γ-tubulin in close proximity to its wall (Sonnen et al., 2012), suggest that
the few MTs (nucleated by -TuRCs of centriolar identity only) originating from this
centriole are useful to transmit latitude inclination (and corresponding receptors) to
the -TuRCs of the PCM, reached with centrifugal and straight-line direction during
last stages of mitosis (centrosome maturation): ”There are no foci of convergence of
microtubules around the second (daughter) centriole in interphase until the middle of
the G2 period ….the centriole as such is not the center of microtubule formation;
instead, microtubule-organizing centers are formed on it or in contact with it”
(Vorobjev and Chentsov, 1982). The problem still needs to be addressed of how this
information about latitude is transmitted from the DC to all the γ-TuRCs included in
the same parallel disk, but located in different meridian wedges (of different
longitude: Fig. 6 C). The MC can easily transmit its 9-fold symmetry to the entire
PCM, independently from latitude, because of its central/axial position in a
cylindrically organized PCM (Fig. 6 A); the DC, because of its eccentric position
(Fig. 6 B), must transfer the latitude-information of its blades to γ-TuRCs having
different longitude: a simple “radial” transmission from an eccentric DC is not
sufficient: it would only reach the γ-TuRCs of two (opposed) meridian wedges. As
we have already said, this is the main geometrical problem: plant cells, larger than
animal cells, make use of parallel cortical MTs (very similar to the parallels on a
globe) and control the MT distance from each other (e.g. to form the pre-prophase-
band) in order to manage, in some way, the third dimension; metazoan small cells
must transfer the circumferential symmetry of their orthogonal centrioles to -TuRCs,

147
being successful in imposing on them the 3D spherical symmetry indispensable to
organize their 3D cytoskeleton; then a molecular 3D structure must exist capable of
forming something like a little “aster” inside the PCM, with its centre at the proximal
end of the DC, that reaches all the -TuRCs, whatever their longitude.
The cylindrical layers around the MC reach a diameter of about 560 nm, accordingly
with Sonnen and colleagues (2012) and Fu and Glover (2012) and form almost the
whole PCM. On the other hand, the DC is surrounded by layers with maximum
thickness of 200-250 nm containing also γ-tubulin (Fu and Glover 2012); both the
centrioles have an external diameter of 170-180 nm: the DC is surrounded by very
little PCM, which is nevertheless able to nucleate few MTs originating from -TuRCs
placed almost on the centriole wall itself. This is consistent with the notion that the
DC is not a real MTOC; even the distance between the γ-tubulin and the DC wall
(Sonnen et al., 2012) fits in with the preceding observations. So what is the use of an
orthogonal centriole (DC) if it produces only very few MTs (Bornens, 2002; Vorobjev
and Chentsov, 1982)?
The DC eccentricity leads me to think that on the proximal end of each MT blades
there exists an MT structure (the “large pericentriolar cloud of additional
components can be accumulated in a microtubule-dependent manner, leading to an
extended boundary” hypothesized by Bornens) able to transmit, by means of PCM-
restricted-MTs, the latitudinal inclination and the relative receptors (characteristic of
each DC triplet) to all the γ-TuRCs of each corresponding disk (or cap) whatever
their longitude. Since the 40° angle-based geometry must be respected (as in
centriolar 9 fold symmetry: 360°/9 = 40°; the central dihedral angle of each wedge
measures 40°) it seems logical to take into consideration a structure composed of five
SAS-6 dimers, whose molecular structure is responsible for the 9-fold symmetry
constructed on an internal angle of 40° and on its supplemental external angle of
140° (polymerization angle of SAS6 dimers N-termini: Kitagawa et al., 2011; Van
Breugel et al., 2011; Gönczy, 2012); these authors have demonstrated the capacity of
SAS-6 dimers to self-assemble in polymers from 3 to 10 units whose N-terminal ends

148
bind together to form a 140° angle. As already anticipated, unlike fullerenes and
clathrins vesicles, which are quasi-spherical objects, centrosomes do not build an
accurately spherical surface: the PCM grows around the centrioles (the core
components of the centrosome) and organize perfectly oriented -TuRCs on an
irregularly spherical surface. Surprisingly SAS-6 is not only inside the DC lumen
(during DC formation) where it appears in a larger quantity to organize the cartwheel
or the central hub, which have a certain length, but it is also outside the same
procentriole, in close proximity to the proximal end of the centriole wall.
Gopalakrishnan, Guichard and colleagues in “Self-assembling SAS-6 multimer is a
core centriole building block” (2010) show very interesting electronic images and
histograms (free on the Internet) about the clear presence of SAS-6 immediately
outside the centriolar wall: the differences between cross and longitudinal sections
agree with the hypothesis that SAS-6 localizes also in the PCM near the proximal end
of the procentriole; the authors furnish interesting diagrams to show the percentage of
SAS-6 found inside and outside the centriole wall, and these values agree with the
expected ones. Also the localization of CEP 135/Bld10 (which acts as a bridge
between SAS-6 and radial spokes) is more external than was previously believed, γ-
TuRCs are very close to the surface of the DC and γ-tubulin is present inside the
centriole (Sonnen et al., 2012) and immediately outside (Fouquet et al., 1998).
A model to explain how the “about-spherical” shape of the centrosome might be
organized is conceivable: a half ring of 25 nm diameter (Fig.11) of five SAS-6 dimers
(through 140° interaction between their N-termini ), lying on the plane containing the
axis of the daughter centriole, and the A-MT of each triplet-blade, positioned
(outside, but very close to the centriole wall) near the proximal centriole pole, along
the MT blade, can radiate five MTs (from the C-terminals of the SAS-dimers) starting
with an angle of 40° between themselves, like the cartwheel spokes. These MTs, like
radii, are lye on a plane orthogonal to the centrosomal surface, and a molecular
complex similar to Chlamydomonas Bld-10 or MAP-2/ MAP-4 can assure the
orientation of the corresponding -TuRCs (or their docking platforms), orthogonal to

149
MTs and parallel to the local tangent plane. This can be the skeleton of a semicircular
(half-round) structure lying on this plane: a similar process from each blade (near the
proximal poles, then close to an ideal centre of the centrosome) can constitute the
structural basis of a “spherical” centrosome, or rather the platform on which
centrosomal proteins are “spherically” assembled. This is not far from the
“hypothetical model of how centrioles could organize the centrosome matrix”
proposed by Bornens (2002). Several other proteins cooperate with SAS-6 to build
the 9-fold centriole structure and the 140° angle: together they organize, as we have
said, along each DC blade, a half-ring fan-shaped structure that reach, radially, the γ-
TuRCs and superimpose their DC inclination (latitudinal) on that already imposed by
the MC cylindrical structure (longitudinal); so the DC, through this 140°-fan-shaped
SAS-6 half ring and centrosomal MTs, can transmit its latitudinal symmetry and
relative receptors to γ-TuRCs having different longitude.
“In late G2 or M phase cells, Cep135 staining could be seen to extend from MCs to
the area occupied by SAS-6 and STIL. This indicates that Cep135 progressively
associates with the proximal ends of DC, concomitant with their growth during the
cell cycle…taken at face value our data suggest that the number of Cep135 proteins
at cartwheel structures is likely to be low in comparison to the amount of Cep135 that
can be detected by 3D-SIM at MCs. It is tempting to speculate that the latter Cep135
population is important for centriole stabilization” (Sonnen et al., 2012).
The DC 140°-fan-shaped structure is likely composed of few layers of half ring of
SAS-6 and related proteins for a total thickness of 36 nm, compatible with the blade
width. The procentriole central hub has a periodicity of 12 nm because of its
composition of about ten layers of SAS-6 rings: “the central tubule is a polymer of
repeating subunits” (Gopalakrishnan, 2010, Guichard et al., 2010). Nine (temporary)
such 140°-fan-shaped structures along each DC blade constitute a small aster inside
the PCM able to correctly orient all the γ-TuRCs (Fig. 11). It is also known (Jakobsen
et al., 2001) that the PCM has a high turnover of its components: the repeated
movements of splitting and joining of the rather more mobile DC (Piel et al., 2000),

150
in respect to the MC, which is always well anchored in its fixed position in the cell,
ensure the participation of the DC in the correct turnover of the PCM components
under its control.
Considering again Albrecht-Buehler’s cited (and already modified) sentence: “It
appears that the simplest and smallest device that is compatible with the scrambling
influence of thermal fluctuations as are demonstrated by Brownian motion is a pair of
9-fold-symmetry-cylinder oriented at right angles to each other”, I am tempted to
add some other words: in effect it is very difficult to think to a tool like the
centrosome equipped with two orthogonal centrioles, one of which is not eccentric;
necessarily, one centriole, the MC, is a central hub while the second cannot form with
it a cross (literally crossing and intersecting the MC wall: two orthogonal hubs are
very difficult to build and seem useless because a clutched contiguity or an extreme
proximity cause barrier/hiding problems –Fig. 11–) but must assume any eccentric
configuration: the most simple arrangement appears to be the known juxtaposition of
the ends because, as it is possible to appreciate in Fig. 11, the MC is not a barrier and
does not impede that the DC reaches every PCM wedge and transmits to each wedge
its inclination information; furthermore this type of eccentricity allows the MC to
control the orientation (shared and coordinate tissue polarity) of the DC at the time of
its maturation into a new MC. Therefore the DC requires and needs the particular
“140°-fan shaped structure” to realize “a large pericentriolar cloud of additional
components accumulated in a microtubule-dependent manner, leading to an extended
boundary” to borrow Bornens’ words for my own use. So the Albrecht-Buehler’s
sentence (I hope in his indulgent permission) can sound: ”It appears that the simplest
and smallest device that is compatible with the scrambling influence of thermal
fluctuations as are demonstrated by Brownian motion is a pair of 9-fold-symmetry-
cylinder oriented at right angles to each other, one axial and one eccentric whose
base faces the distal wall of the first”. The particular property of the DC to be an
intermediate product between a procentriole and a “true” centriole, extremely useful
because of its orthogonal position in respect to the MC, is strengthened.

151
In conclusion: in the theoretical geometrical model of the centrosome, the MC
arranges cylindrically γ-TuRCs, corresponding to its 9 “prismatic” faces; the
eccentric DC “compels” all the γ-TuRCs contained in the same cap or disk (between
two “parallels”) to assume also the inclination corresponding to their latitude,
creating the spherical polarity (double inclination) of γ-TuRC scaffolds.
Evolutionary “work in progress”
In my opinion cells without centrioles and centrosomes can perform only simple
laminar morphogenetic processes (including the creation of elementary tubes) making
use of the cytoskeleton and through their ability to control their simple and rough
polarity and MT disposition by different cues: light, gravity, “automatic
repositioning” of cell-cell contacts organized through MAPs, etc. (Wasteneys and
Ambrose 2009: ” Spatial organization of plant cortical microtubules: close encounters
of the 2D kind”); as we have seen “the overall structural organization of plants is
generally simpler than that of animals. For instance, plants have only four broad
types of cells, which in mature plants form four basic classes of tissues…organized
into just four main organ systems” (Lodish et al., 2012, Ch. 20). Plants possess and
use many cytoskeleton factors common with Metazoa: “Remarkably, formins appear
to be not only important regulators of the actin cytoskeleton, but also prime
candidates for mediating the co-ordination between microfilaments and microtubules
also in plants, despite using mechanisms of microtubule interaction different from
those of their metazoan counterparts” (Cvrčková, 2012). Microtubule Associated
Proteins (MAPs) can arrange MTs parallel and constantly distanced from each other:
“Cellulose synthases in etiolated Arabidopsis thaliana hypocotyl cells move
bidirectionally in the plasma membrane along tracks that are typically defined by

152
cortical microtubules. However, examples are known in which, vice versa, cellulose
microfibrils impose orientation on newly formed cortical microtubules….At near
weightlessness, tubulin polymerizes into microtubules in vitro, but these microtubules
do not self-organize in the ordered patterns observed at 1g. Likewise, at near
weightlessness cortical microtubules in protoplasts have difficulty organizing into
parallel arrays, which are required for proper plant cell elongation. However, intact
plants do grow in space and therefore should have a normally functioning
microtubule cytoskeleton. Since the main difference between protoplasts and plant
cells in a tissue is the presence of a cell wall, we studied single, but walled, tobacco
Fig. 11 Centrosome theoretical geometrical model: transmitting the same latitudinal receptors to
γ-TuRCs with different longitude.
A) Along each DC blade a 140°-fan-shaped MT-structure, SAS6 and -tubulin based, is positioned near the
proximal centriole end and lies on the plane containing the axes of the centriole and the A-MT of each
triplet-blade: it is supported by a half ring of five SAS-6 dimers (reverse “T”) and other proteins cooperating
to build the 140° angle; through SAS-6 dimers C-terminal coiled coils (“T” stems) and -tubulin/MT , this
140°-fan-shaped MT-structure reaches and organizes -TuRCs (ellipsoidal disks) localized in different
meridian wedges (with different longitude) and superimposes DC radiality (latitudinal inclination) on the
longitudinal inclination already imposed by the MC cylindrical structure.
B) Schematic centrosome cross section at MC proximal end with a nascent procentriole (DC): view from the
MC distal end. The 140°-fan-shaped structures (dotted lines) starting from the DC PCM (close to one triplet
blade) can reach five (1-5) wedges (separated by lines) of the MC PCM (-TuRCs: ellipsoidal disks); a
similar structure (not represented) on the opposite (symmetric) DC PCM (close to the corresponding triplet
blade) can reach five corresponding wedges of MC PCM: two symmetric DC blades transmit the same
latitude; 140° geometry is the same for MC PCM and DC 140°-fan-shaped structure. As depicted, the
presence of the cylindrical MC is not a barrier.
C) View from the DC distal end of a schematic orthogonal (in respect to B) representation of geometrical
disposition of the nine 140°-fan-shaped structures (dotted lines): from the DC PCM each one reaches the
corresponding cap or parallel ring of the MC linked PCM (-TuRCs: disks); symmetric DC blades transmit
the same latitude; 140° geometry is the same as in A, and the presence of the cylindrical MC is not a barrier.

153

154
BY-2 suspension-cultured cells during an 8-day space-flight experiment on board of
the Soyuz capsule and the International Space Station during the 12S mission
(March-April 2006)…. Microtubules could be aligned by cues from an anisodiametric
cell shape, anisotropical mechanical stress, moving cellulose synthase complexes, a
pre-existing cell wall pattern, or maybe gravity” (Sieberer et al., 2009). “One
emerging concept is that the ability to organize decentralized microtubule arrays
depends to a large extent on microtubule self-organization. This may be coordinated
largely by the activities of motor proteins, which in plants include a remarkable
variety of minus- and plus-end directed kinesins…Without centrosomes, plant
microtubule arrays are largely self-organized by the relative activities of microtubule-
associated proteins. The recent discoveries of katanin p60 and MOR1 provide
important clues about how microtubule assembly and stabilization are achieved,
particularly in the cortical arrays that regulate growth polarity and cell wall
deposition in interphase and terminally differentiating cells” (Wasteneys, 2002).
One centriole alone can organize organisms with circumferential (rotational)
symmetry; the intervention of a second orthogonal centriole enables the careful
management of the 3D of tissues and organs, because it governs the orientation of γ-
TuRCs in accordance to the latitude angle, which is added and superimposed to that
of longitude. This is true also for some particular Metazoa: during Drosophila
segmentation, centrosomes are indispensable (segmentation is a 3D process) but in
later stages of fly development they are dispensable: laminar (2D) morphogenetic
processes and creation of elementary tubes do not require 3D precise controls.
There is a wide taxonomic range of “work in progress” in which centrioles and
centrosome have different degrees of development with different structures (the
unicellular green alga Chlamydomonas has centrioles with triplets, like other
unicellular Protists, but Caenorhabditis elegans centriole has singlets and Drosophila
has doublets). As observed by Azimzadeh and colleagues (2012), cellular and
developmental processes are conserved in flatworms, but planarians have lost the
centrosome and develop by making use of different pathways which allow them to

155
build only laminar tissues (like the leaves in Plantae) or elementary tubes and to
maintain an extraordinary ability for regeneration. Acerentomon microrhinus builds
axonemes with 14-fold symmetry; Cnidaria and Ctenophora appear radially
symmetrical, but they are quite different: they are Radiata, not Bilateria and have
only two germ layers, ectoderm and endoderm (Diploblasts); they have centrioles and
cilia, but the behaviour of their centrosomes is quite different than in Bilateria
(Bornens, 2012): Ctenophora fertilization is naturally polyspermic and many
centrioles are utilized contemporaneously in the same cell; Cnidaria zygotes lack
cytasters (Salinas-Saavedra and Vargas. 2011). In this taxonomic area - and in the
corresponding evolutionary phase - some genetic changes need to complete the final
centrosome structures take place. It is clear that although the number of genes is quite
similar in different metazoan Clades, on the contrary, their 3D complex organization
goes in parallel, hand in hand, with non-coding DNA amount and centrosome
structure development.
“Πάντα ῥ εῖ ” [everything flows] (Heraclitus)
The continuous flow of developing and not completely mature mechanisms and
structures (and their molecular base) is characteristic of evolution, and allows what
has been generated previously to be conserved and reused by superimposing small
but important modifications (transposons, duplications, exon exchange between
different genes –gene shuffling–, unequal cross-over, etc.) in order to achieve
surprising results. So, depending on the success in surviving (purifying selection)
different species show differently developed centrosomes and different pathway to
control centrosome’s “hardware and software”: if the origin of bilateral symmetry
may be a simple evolutionary event, quite more complicate are the control
mechanisms; in every species coding DNA is much shorter than the set of sequences
that control gene expression. Everything flows…
But, sometimes, something comes back: “ Nihil sub sole novum” [nothing new under
the sun] (Ecclesiastes).

156
iv) mapping the whole cortex
Two different cycles must be followed: the centriole and the centrosome cycle (pay
attention to distinguish “centriole cycle” and “centrosome cycle” when you read the
following lines).
Centriole cycle:
Vorobjev and Chentsov (1982) have clearly described the centriole cycle as follows
(Fig. 1):
“- The structures of the two centrioles differ throughout interphase; the MC has
appendages, the DC does not.
- In the distal part of the DC, there are peculiar outgrowths (ribs). Unlike
appendages, these ribs remain unaltered in two to four serial sections. The above
structure of the DC remains constant to the end of telophase.
- Throughout the S period, differences are observed in the structure of the two MCs,
i.e., one of them contains satellites and appendages, while the other has note. The
structure of the second centriole changes somewhat in the S period: it loses ribs
situated in the distal part.
- Thus, in the next cycle, the new (ex-daughter) centriole differs from the former
(mother) only by the absence of appendages. This difference, however, is functionally
significant…Appendages are probably needed for bridging the centriole to the
membrane, which is also evidenced by the fact that when they become attached to the
membrane their position and outward appearance are somewhat changed.
- Two MCs situated in the two poles of a cell differ somewhat in structure: one
contains distinctly visible appendages, whereas the other lacks them…In anaphase,
the general structure of spindle poles hardly changes. But by this time both MCs have
appendages which are less distinct than in metaphase.
- Despite the fact that in G2 two MCs become equivalent with respect to

157
microtubules, the difference between them, i.e., the presence of appendages on one
and their absence from the other, persists throughout the G2 period and during the
first half of mitosis”.
Centrosome cycle:
Ou and colleagues (2002) have described the centrosome cycle as follows (Fig.1):
“- Thus, in late S phase cells that had two separated centrosomes, the MC centrosome
had two regions of CEP110 (centriolin) staining, whereas the Daughter centrosome
had only one site.
- In random cultures, centrosomes initiating duplication can be identified by the
appearance of a structure at the margin of the centrosome. When cells displaying this
stage of duplication were stained for CEP110, this protein was found at the site of
duplication as well as at the open and closed ends of the MC centrosome tube.
- Further, we showed that, following centriole separation at G1-S, the DC moves
along the MC and come to reside at the site of centrosome duplication. Importantly,
the addition of these proteins to the open end of the Daughter centrosome tube
coincides with the transition of the Daughter centrosome to a mature centrosome
which is capable of duplication and acts as a microtubule organizing center.
- Thus, our study, for the first time, identifies a specific region of the centrosome that
is modified in its protein composition during the process of centrosome maturation, a
process that is completed following cell division”.
During the process of maturation (Fig. 1), the DC moves along the MC towards the
distal end of the MC (where MC appendages are): after losing its ribs, it is formed by
9 equivalent and identical triplets: then, the old DC, now new MC, matures near the
old MC to maintain a coordinated orientation of centrosomes (based on non-
equivalence of triplets), indispensable to coordinate the development of complex
organs. Let’s recall Harold’s words: “Centriole duplication is part of the mechanism
by which the cytoskeleton of the daughter cell is patterned upon that of the mother”.

158
A question: is it possible that the proximal end of the ex-DC becomes the distal end
of the newly formed (or, rather, maturated) MC? In this case some receptor might
distinguish the old and the new centrosome and perform asymmetrical delivery of
transcription factors and mRNAs during mitosis, as we have seen in Ilyanassa.
“Further, we showed that CEP110 appeared at only one site (the site of CEP250/c-
Nap1) within the Daughter centrosome from the time of its initial formation until
prophase” (Ou et al., 2002). “In anaphase, the general structure of spindle poles
hardly changes. But by this time both MCs have appendages which are less distinct
than in metaphase” (Vorobjev and Chentsov, 1982). So, in late anaphase/beginning
telophase, when the new centrosome is mature but the nucleus has not yet formed and
DNA is still packed into chromosomes, there is the physical space for the two
centrosomes, through their asters, to be able to map and wire cortical compartments
in the two daughter cells without the impediment of the nucleus which will, only
subsequently, distort the perinuclear MTs: these, unlike other MTs, will no longer be
substituted in G1, because their intrinsic rigidity does not allow new ones, incapable
of bending in order to move around the nucleus, to be nucleated: “Our study suggests
that perinuclear microtubules are a separate and relatively stable subpopulation of
the total population of cytoplasmic microtubules and may serve a function different
from that of the more variable not perinuclear microtubules” (Park et al., 1984).
Stable MTs show acetylation of a lysine and C-terminal detyrosylation of α-tubulin.
As we have already seen, aster MTs constitute a particular population of stable MTs
whose function consists in mapping and wiring the whole cell.
The maturation timing of the DC and new centrosome (that forms around it, using it
as a “new” MC), gives a response to query iv): the nucleus does not obstruct the
“mapping” process of the entire daughter cell because the process takes place in
anaphase-telophase, when the chromosomes are still compact, endoplasmic reticulum
and Golgi membranes are restricted in small vesicles, both centrosomes are mature
and both asters can easily reach the whole daughter cell cortex.

159
v) Centrosomal trafficking
“Functional centrioles and intact subdistal appendages are required for the
recruitment of PCM proteins in animals” (Esparza et al., 2013).
“In eukaryotic cells, targeted vesicle fusion requires, in addition to the SNAREs, both
a delivery system and a secretory apparatus” (Harold, 2005).
As we have seen, Kubo and colleagues (1999) and Dammermann and Merdes (2002)
observed that pericentriolar satellites, after recruiting proteins indispensable for
anchoring γ-TuRCs to their centrosomal docking platform (centrin, pericentrin,
ninein, SAS-4 etc.), are transported to the centrosome by MT-linked dynactin.
The fifth query to be dealt with concerns the “crowded” presence of many
differently-oriented astral MTs, originating from centrosomal γ-TuRCs: how are the
macromolecular complexes, destined for a precise position in the cell membrane,
quickly guided toward the respective γ-TuRCs, interspersed among many other on a
large polyhedral surface? Up to now, we have discussed the subject of wiring
precision: to assure the needed speed, each γ-TuRC and its MT must be reached
easily and quickly. The “astral” MTs are nucleated by γ-TuRCs that are suitably
oriented and equipped, in accordance to the model, with receptors corresponding to
their latitude and longitude. As we have seen, the only “motor” that moves molecules
in solution is thermal fluctuation and the only “helm” that directs the molecules in
different directions is the result of random collisions with other molecules. The
process by which a macromolecular complex finds the γ-TuRC it is destined for,
localized on a spherical structure that in many cases makes a complete
“circumnavigation” of the centrosome necessary, frankly seems to be too complicated
and slow, if not impossible: protein diameter average size is about 3-6 nm,
centrosome diameter is about 800 nm, its equator then measures approximately 2,500
nm and its surface about 2,000,000 nm2. It appears necessary a multi-step process:
firstly a centrosomal receptor seems to be necessary, situated at one pole of the
centrosome, able to recognize (and to be recognized by) a targeting sequence placed

160
on those molecular complexes that must use centrosome and aster to be correctly
located in the cell.
“The identification and localization of two populations of both CEP110 and ninein
within the centrosome has some functional implications. First, the function of
CEP110 and ninein is, at least in part, centriole-based. Second, previous studies have
suggested that ninein plays a role in microtubule anchoring at the centriole and acts
as a cap for microtubule ends. If so, the presence of ninein at the open end of the
centrosomal tube suggests that microtubule anchoring or capping functions may
occur within this region as well, at least at some point during the cell cycle. The
absence of ninein from the daughter centrosomes would therefore limit their ability to
anchor or cap microtubules at the open end of the tube. Such a relationship may
explain in part why previous studies failed to identify microtubules in association
with immature centrosomes. The observations described here raise the possibility that
ninein plays a larger, and perhaps more diverse, role within the centrosome than
previously thought” (Ou et al., 2002). These authors seems to adapt the hypothesis
supported, amongst others, by Bornens (2002) about PCM restricted MTs (MTs
would start only from MC subdistal appendages) and have developed the
observations by Vorobjev and Chentsov (1982) about the role of the whole MC-
linked-PCM (from which cytoplasmic MTs would start) and MC appendages, where
ninein could play the role of catching the molecular complexes directed to the
centrosome (ninein plays a larger, and perhaps more diverse, role within the
centrosome).
Studies on centrosomal targeting sequences of different ninein isoforms and on the
ability of ninein to bind different kinases and γ-TuRCs themselves, carrying out a
larger, and perhaps more diverse, role within the centrosome than previously thought,
are attractive. “Our study of GFP-tagged full-length hNinein reveals that four
domains are required and sufficient for centrosomal targeting” (Lin et al., 2006). In
this sense, the Cep 164 complexes of distal appendages and of ninein-CEP110
(centriolin), present on the 9 subdistal appendages of the MC, may have the role of

161
sites for recognizing centrosome localization signals, presented by macromolecular
complexes, with the consequent capacity to route these complexes, after binding them
to the centrosome, on the sector having the required longitude. We have already seen
that Kubo and colleagues (1999) and Dammermann and Merdes (2002) observed that
pericentriolar satellites, after recruiting proteins indispensable for anchoring γ-TuRCs
to their centrosomal docking platform (centrin, pericentrin, ninein, SAS-4 etc.), are
transported to the centrosome by MT-linked dynactin.
“We propose that Cep164 is targeted to the apical domain of the MC to provide the
molecular link between the MC and the membrane biogenesis machinery that
initiates cilia formation” (Schmidt et al., 2012). This same molecular link could have
been adapted to drive molecular complexes (centrosome targeted) onto the desired
centrosome wedge.
An in-depth study on the role of ninein “in MT nucleation and MT anchoring at the
centrosome” was carried out by Delgehyr and colleagues (2005): “Ninein is targeted
to both centrioles by its C-terminus and stabilized at the MC by its N-
terminus...These data suggest that a subset of microtubules could be nucleated and
later anchored at the subdistal appendages. Based on our results, we propose that
microtubules nucleated in a γ-TuRC-dependent manner at the centrosome would have
different behaviours. Microtubules nucleated far from the subdistal appendages
would be transiently anchored by the γ-TuRC but then released from the centrosome
in a manner possibly dependent on specific factors. Indeed, many microtubule seeds
are produced and released from the centrosome. By contrast, microtubules nucleated
at the vicinity or on the subdistal appendages would become anchored more readily
at these structures on the MC. Therefore, ninein, a component of these appendages,
might favour microtubule nucleation (by docking the γ-TuRC), followed by
anchoring”.
In accordance to this hypothesis, subdistal appendages might have the role of
capturing the complexes directed to the centrosome through their own “centrosomal”
MTs (arising from γ-TuRCs of centriolar identity: Fouquet et al., 1998) like

162
mechanical molecular antennas and distributing them, through “centrosomal” actin to
the final corresponding γ-TuRC from which the “cytoplasmic” MT, directed to the
cortex, will originate. “The recycling endosome associates with the appendages of the
MC” (Hehnly et al., 2012). As we have seen, during its first stages of cleavage, in
Ilyanassa obsoleta long RNAs are linked to only one of the two centrosome before
mitosis to be delivered exclusively and asymmetrically to micromeres: differential
targeting leads the new arising centrosome in that sector of the cell where the position
of the spindle pole is programmed and the same targeting sequence guide RNAs to
the same sector (together with the centrosome).
It is an attractive hypothesis that several centrosome targeting sequences exist, placed
on those macromolecular complexes (RNA 3’ UTR or N-terminus of polypeptides)
that must be routed towards prefixed cell locations through the appropriate MTs.
Such mechanism is quite similar to that used for translocation of proteins in
mitochondria, where different targeting sequences, present on the proteins that must
enter the mitochondrion, are sequentially decoded in order to reach the expected
location (the external mitochondrial membrane, the internal one, the inter-membrane
space or the matrix); it is well known that in fly oocyte bicoid and nanos mRNA are
localized through their 3’UTR sequence and delivered by MT transport. When a
polypeptide or polyribonucleotide complex (spindle poles binding proteins, cell
adhesion complexes, integrins and other extra-cellular fiber receptors, maternal
mRNAs, etc.) must be sent by means of the centrosomal MTs into a certain cortical
compartment, should first of all have a centrosomal localization signal that is easily
recognizable in a particular (easy accessible) area of the centrosome: the cap where
the distal end of the MC is present with its subdistal appendages, where the open end
of the centrosomal tube (Ou et al., 2002) is and where the ninein-linked MTs are, is
the most likely one. Here a second receptor placed on the appropriate “wedge”
appendage can recognize a second targeting sequence that identifies the meridian and
can send the macromolecular complex along the expected sector (wedge), where
molecular motors present in the centrosome (centrosome localization of myosin-V

163
and centractin - an ARP i.e. Actin Related Protein - is well known) can lead it towards
the latitudinal receptor corresponding to its third targeting sequence. At each
recognition of a targeting sequence by the appropriate receptor, different kinases or
proteases (Urbé et al., 2012) may deactivate or remove the targeting sequence that
has been read and recognized, leaving room for the next recognition, exactly as
happens in mitochondria, in which the choice of one among many destinations poses
quite similar problems.
From its ancestral function of anchoring to the membrane and generating the
axoneme, the MC distal end has evolved to acquire new structures in order to link to
adjacent centrioles (Chlamydomonas, Paramecium, Tetrahymena) and interface with
them and with other cytoplasmic fibers, in accordance to a precise geometric design;
Vorobjev and Chentsov (1982) observed that “appendages are probably needed for
bridging the centriole to the membrane, which is also evidenced by the fact that when
they become attached to the membrane their position and outward appearance are
somewhat changed”. In non-ciliated cells the appendages (distal and subdistal) seem
to perform other and new functions, such as orienting the centrosome in the cell
(coordinately, as we have already seen, with the polarity of neighboring cells),
anchoring it to the nucleus, correctly orientating the daughter centrosome, capturing
and routing of macromolecular complexes equipped with “address-signal” (targeting
sequences) towards the correct γ-TuRC; membrane linked centrosome maintain their
orientation and can perform their geometrical role of biological interfaces.

164
Conclusions
Let’s then summarize the results:
i) The centrosome is an ordered tool whose surface is made up of 45 oriented
scaffolds for γ-TuRCs; each scaffold is labelled by its own receptor able to recognize
(and interact with) the targeting sequence intended for the cortical compartment
reachable through the MT nucleated by its oriented γ-TuRCs. One-to-one invariable
correspondence is established between targeting sequences and γ-TuRC molecular
labels. One-to-one univocal but time-variable correspondence is established between
γ-TuRs and cortical compartments, depending on the (known and controlled)
centrosome position in the cell.
ii) Two centrioles, arranged with eccentric orthogonality during S G2 and M phase,
transmit their 9-fold symmetry and non-equivalence of the triplets to the PCM (γ-
TuRC scaffolds).
iii) A ring of RNA, composed of nine different and equi-spaced loops (markers) is
responsible for the non-equivalence of the centriolar triplets.
iv) A transposition event of the gene coding for the RNA sequence responsible for the
non-equivalence of the centriolar triplets has generated a similar sequence with a
little difference in its ends so that its transcript’s 3’ and 5’ terminals, once juxtaposed,
compose a stem whose 3D structure is mirror symmetric to that of the original
transcript. The interaction between the RNA ring stem and the corresponding
molecular centriolar complex must occur through a typical disposition of the stem:
this causes the overturn of the ring transcribed from the transposed locus with
consequent reversal of the sequence of the triplet markers.

165
Future perspectives: From cell geometry to organ geometry. Toward a global theory
of morphogenesis in Metazoa
In fibroblasts, procollagens and proteoglycans are delivered through trans-Golgi
derived secretion vesicles, carried to the cell membrane by MT linked kinesins: these
vesicles converge and coalesce to form long and deep grooves on the outer face of the
cell membrane, where procollagens auto-assemble into long fibrils. The orientation of
grooves and surface collagen binding molecules determine the spatial orientation of
future collagen fibers. Taken together, the geometrical model of the centrosome, its
role in mapping and wiring the cell cortex (one-to-one correspondence between
centrosomal and cortical compartments), the coordination of the shared common
centrosome orientation in every cell of a tissue make it possible to hypothesize that
the shape of organs is modeled by connective tissues through genetically
programmed orientation of extracellular matrix fibers, whose geometrical orientation
and direction are achieved by means of controlled exocytosis of procollagen and
proteoglycan: this may be possible because of the centrosome and its role of
geometric, noise resistant, 3D interface that translates morphogenetic signals into
precise locations in the cell. Intriguingly, fibroblast secretory granules and grooves,
membrane areas forecast by the centrosome model and cortical domains found in
eukaryotic cells have compatible size. This may be the transition from cell geometry
to organ geometry.

166
References
Albrecht-Buehler G. (1981) Does the geometric design of centrioles imply their
function? Cell Motil. 1: 237-245.
Alliegro M.C. (2011) The centrosome and spindle as a ribonucleoprotein complex.
Chrom. Res. 19: 367-376.
Alliegro M.A., Henry J.J., Alliegro M.C. (2010) Rediscovery of the nucleolinus, a
dynamic RNA-rich organelle associated with the nucleolus, spindle, and centrosomes.
Proc. Natl. Acad. Sci. USA. 107: 13718-13723.
Alliegro M.C., Alliegro M.A. (2008) Centrosomal RNA correlates with intron-poor
nuclear genes in Spisula oocytes. Proc. Natl. Acad. Sci. USA 105: 6993-6997.
Alliegro M.C., Alliegro M.A., Palazzo R.E. (2006) Centrosome-associated RNA in
surf clam oocytes. Proc. Natl. Acad. Sci. USA 103: 9034-9038.
Almeida J., Galego L. (2005) Flower symmetry and shape in Antirrhinum. Int. J.
Dev. Biol. 49: 527-537.
Andreuccetti L., Carnevali O., Dini L., Falugi C., Filona S., Kaltoff K., Viscoso R.
(2010) Biologia dello Sviluppo. McGraw-Hill, Milano. Pp. 10-11, 132, 227.
Andrew D.J., Ewald A.J. (2010) Morphogenesis of epithelial tubes: insights into tube
formation, elongation and elaboration. Dev. Biol. 341: 34-55.
Avasthi P., Marshall W. (2013) Ciliary secretion: switching the cellular antenna to
transmit. Curr. Biol. 23: 906-911.
Azimzadeh J., Bornens M. (2007) Structure and duplication of the centrosome. J.
Cell Sci.120, 2139-2142.

167
Azimzadeh J., Marshall W.F. (2010) Building the centriole. Curr. Biol. 20: R816-
R825.
Azimzadeh J., Wong M.L., Downhour D.M., Alvarado A.S., Marshall W.F. (2012)
Centrosome loss in the evolution of planarians. Science 335: 461-463.
Baena-López L.A., Baonza A., García-Bellido A. (2005) The orientation of cell
divisions determines the shape of Drosophila organs. Curr. Biol. 15: 1640-1644.
Baker J., Theurkauf W.E., Sehubiger G. (1993) Dynamic changes in microtubule
configuration correlate with nuclear migration in the preblastoderm Drosophila. J.
Cell Biol. 122: 113-121.
Baker N.E. (2011) Proximodistal patterning in the Drosophila leg: models and
mutation. Genetics 187: 1003-1010.
Basto R., Lau J., Vinogradova T., Gardiol A., Woods C.G., Khodjakov A., Raff J.W.
(2006) Flies without centrioles. Cell 125: 1375-1386.
Beisson J. (2008) Preformed cell structure and cell heredity. Prion 2: 1-8.
Beisson J., Jerka-Dziadosz M. (1999) Polarities of the centriolar structure:
morphogenetic consequences. Biol. Cell 91: 367-378.
Bellett G., Carter J.M., Keynton J., Goldspink D., James C., Moss D.K., Mogensen
M.M. (2009) Microtubule plus-end and minus-end capture at adherens junctions is
involved in the assembly of apico-basal arrays in polarised epithelial cells. Cell
Motil. Cytoskeleton 66: 893-908.
Benson G. (1999) Tandem repeats finder: a program to analyze DNA sequences.
Nucleic Acids Res. 27: 573-580.
Bettencourt-Dias M. (2013) Q&A: Who needs a centrosome? BMC Biol. 11: 28.
Bienkowska D., Cowan C.R.(2012) Centrosomes can initiate a polarity axis from any
position within one-cell C. elegans embryos. Curr. Biol. 22: 583-589.
Blower M.D. (2013) Molecular insights into intracellular RNA localization. In.t Rev.
Cell. Mol. Biol. 302:1-39.
Bornens M. (2012) The centrosome in cells and organisms. Science 335: 422-426.
Bornens M. (2002) Centrosome composition and microtubule anchoring

168
mechanisms. Curr. Op. Cell Biol. 14: 25-34.
Brodu V., Baffet A.D., Le Droguen P.M., Casanova J., Guichet A. (2010) A
developmentally regulated two-step process generates a noncentrosomal microtubule
network in Drosophila tracheal cells. Dev. Cell 18: 790-801.
Brown N.A., Wolpert L. (1990) The development of handedness in left/right
asymmetry. Development 109: 1-9.
Brugmann S.A., Cordero D.R., Helms J.A. (2010) Craniofacial ciliopathies: a new
classification for craniofacial disorders. Am. J. Med. Genet. A 152A: 2995-3006.
Bryant P.J. (1981) Distal regeneration and symmetry. Science 212: 993-1002.
Bui K.H., Sakakibara H., Movassagh T., Oiwa K., Ishikawa T. (2009) Asymmetry of
inner dynein arms and inter-doublet links in Chlamydomonas flagella. J. Cell Biol.
186: 437-446.
Cagan R. (2009) Principles of Drosophila eye differentiation. Curr. Top. Dev. Biol.
89: 115-135.
Cao J., Shen Y., Zhu L., Xu Y., Zhou Y., Wu Z., Li Y., Yan X., Zhu X. (2012) miR-
129-3p controls cilia assembly by regulating CP110 and actin dynamics. Nat. Cell
Biol. 14: 697-706.
Carbajal-González B. “Similar differences”: radial spokes are different from each
other, but conserved across species. (2012) On line at:
http://blogs.brandeis.edu/science/2012/02/08/similar-differences, last accessed
September 2, 2013.
Cernuda-Morollón E., Millán J., Shipman M., Marelli-Berg F.M., Ridley A.J. (2010)
Rac activation by the T-cell receptor inhibits T cell migration. PLoS One 5: e12393.
Chichinadze K., Lazarashvili A., Tkemaladze J. (2013) RNA in centrosomes:
structure and possible functions. Protoplasma 250: 397-405
Cottee M.A., Raff J.W., Lea S.M., Roque H. (2011) SAS-6 oligomerization: the key
to the centriole? Nature Chem. Biol. 7: 650-653.
Cowan C.R., Hyman A.A. (2004) Centrosomes direct cell polarity independently of
microtubule assembly in C. elegans embryos. Nature 431: 92-96.

169
Cvrčková F. (2012) Formins: emerging players in the dynamic plant cell cortex.
Scientifica Article ID 712605; doi: 10.6064/2012/712605.
Dammermann A., Merdes A. (2002) Assembly of centrosomal proteins and
microtubule organization depends on PCM-1. J. Cell Biol. 159: 255-266.
Danilchik M.V., Brown E.E., Riegert K. (2006) Intrinsic chiral properties of the
Xenopus egg cortex: an early indicator of left-right asymmetry? Development 15:
4517-4526.
Delgehyr N., Sillibourne J., Bornens M. (2005) Microtubule nucleation and
anchoring at the centrosome are independent processes linked by ninein function. J.
Cell Sci. 118: 1565-1575.
Dippel R.V. (1976) Effects of nuclease and protease digestion on the ultrastructure of
Paramecium basal bodies. J. Cell Biol. 69: 622-637.
Dodgson J., Chessel A., Yamamoto M., Vaggi F., Cox S., Rosten E., Albrecht D.,
Geymonat M., Csikasz-Nagy A., Sato M., Carazo-Salas R.E. (2013) Spatial
segregation of polarity factors into distinct cortical clusters is required for cell
polarity control. Nat. Commun. 4: 1834.
Dos Santos H.G., Abia D., Janowski R., Mortuza G., Bertero M.G., Boutin M.,
Guarín N., Méndez-Giraldez R., Nuñez A., Pedrero J.G., Redondo P., Sanz M.,
Speroni S., Teichert F., Bruix M., Carazo J.M., Gonzalez C., Reina J., Valpuesta J.M.,
Vernos I., Zabala J.C., Montoya G., Coll M., Bastolla U., Serrano L. (2013) Structure
and non-structure of centrosomal proteins. PLoS ONE 8: e62633.
Drevensek S., Goussot M., Duroc Y., Christodoulidou A., Steyaert S., Schaefer E.,
Duvernois E., Grandjean O., Vantard M., Bouchez D., Pastuglia M. (2012) The
Arabidopsis TRM1-TON1 interaction reveals a recruitment network common to plant
cortical microtubule arrays and eukaryotic centrosomes. Plant Cell 24: 178-191.
Dupin I., Camand E., Etienne-Manneville S. (2009) Classical cadherins control
nucleus and centrosome position and cell polarity. J. Cell Biol. 185: 779–786.

170
Eddy R.J., Pierini L.M., Maxfield F.R. (2002) Microtubule asymmetry during
neutrophil polarization and migration. Mol. Biol. Cell 13: 4470-4483.
Efroni I., Esheda Y., Lifschitz E. (2010) Morphogenesis of simple and compound
leaves: a critical review. The Plant Cell 22: 1019-1032.
Emlen D.J., Allen C.E. (2003) Genotype to Phenotype: Physiological Control of Trait
Size and Scaling in Insects. Integr. Comp. Biol. (2003) 43: 617-634.
Engel B.D., Ishikawa H., Wemmer K.A., Geimer S., Wakabayashi K., Hirono M.,
Craige B., Pazour g.J., Witman G.B., Kamiya R., Marshall W.F. (2012) The role of
retrograde intraflagellar transport in flagellar assembly, maintenance, and function. J.
Cell Biol. 199: 151-167.
Erkang A., Poole D.S., Skop A.R. ( 2011) Long astral microtubules and RACK-1
stabilize polarity domains during maintenance phase in Caenorhabditis elegans
embryos. PLoS One 6: e19020.
Esparza J.M., O’Toole E., Li L., Giddings T.H. Jr, Kozak B., Albee A.J., Dutcher
S.K. (2013) Katanin localization requires triplet microtubules in Chlamydomonas
reinhardti. PLoS One 8:e53940.
Feldman J.L., Geimer S., Marshall W.F. (2007) The MC plays an instructive role in
defining cell geometry. PLoS Biol. 5: e149.
Feldman J.L., Priess J.R. (2012) A role for the centrosome and PAR-3 in the hand-off
of microtubule organizing center function during epithelial polarization. Curr. Biol.
22: 575-582.
Foe V.E. (1989) Mitotic domains reveal early commitment of cells in Drosophila
embryos. Development 107: 1-22.
Foethke D., Makushok T., Brunner D., Nédélec F. (2009) Force- and length-
dependent catastrophe activities explain interphase microtubule organization in
fission yeast. Mol. Syst. Biol. 5: 241.
Fouquet J.P., Kann M.L., Combeau C., Meilki R. (1998) -Tubulin during the
differentiation of spermatozoa in various mammals and man. Mol. Hum. Reprod. 4:
1122-1129.

171
Fu J., Glover D.M. (2012) Structured illumination of the interface between centriole
and peri-centriolar material. Open Biol. 2: 120104.
Galli M., van den Heuvel S. (2008) Determination of the cleavage plane in early C.
elegans embryos. Annu. Rev. Genet. 42: 389-411.
Geimer S., Melkonian M. (2004) The ultrastructure of the Chlamydomonas
reinhardtii basal apparatus: identification of an early marker of radial asymmetry
inherent in the basal body. J. Cell Sci. 117: 2663-2674.
Gibson M.C., Schubiger G. (2000) Peripodial cells regulate proliferation and
patterning of Drosophila imaginal discs. Cell 103: 343-350.
Gibson W.T., Veldhuis J.H., Rubinstein B., Cartwright H.N., Perrimon N., Brodland
G.W., Nagpal R., Gibson M.C. (2011) Control of the mitotic cleavage plane by local
epithelial topology. Cell 144: 427-438.
Gilbert F.S. (2010) The polar coordinated model of positional information in the
developing and regenerating limbs. 9th edn. Sinauer Associates, Sunderland. Ch.
15.6. Pp 165-9.
Gittes F., Mickey B., Nettleton J., Howard J. (1993) Flexural rigidity of microtubules
and actin filaments measured from thermal fluctuations in shape. J. Cell Biol. 120:
923-934.
Gönczy P.(2012) Towards a molecular architecture of centriole assembly. Nature Rev.
Mol. Cell Biol. 13, 425-435.
Gönczy P., Rose L.S. (2005) Asymmetric cell division and axis formation in the
embryo. On line at: Wormbook:
http://www.wormbook.org/chapters/www_asymcelldiv/asymcelldiv.html
Gopalakrishnan J., Guichard P., Smith A.H., Schwarz H., Agard D.A., Marco S.,
Avidor-Reiss T. (2010) Self-assembling SAS-6 multimer is a core centriole building
block. J. Biol. Chem. 285: 8759-8770.
Gopalakrishnan J., Mennella V., Blachon S., Zhai B., Smith A.H., Megraw T.L.,

172
Nicastro D., Gygi S.P., Agard D.A., Avidor-Reiss T. (2011) Sas-4 provides a scaffold
for cytoplasmic complexes and tethers them in a centrosome. Nat Commun. 21;2:359
Griffiths G., Wathne G. (2012) Hitting the dock. On line at:
http://bpod.mrc.ac.uk/archive/2012/3/14, last accessed September 2, 2013.
Gueth-Hallonet C., Antony C., Aghion J., Santa-Maria A., Lajoie-Mazenc I., Wright
M., Maro B. (1993) -Tubulin is present in acentriolar MTOCs during early mouse
development. J. Cell Sci. 105: 157-166.
Guichard P., Chrétien D., Sergio M., Tassin A. (2010) Procentriole assembly
revealed by cryo-electron tomography. EMBO J. 29: 1565-1572.
Guichard P., Hachet V., Majubu N., Neves A., Demurtas D., Olieric N., Fluckiger I.,
Yamada A., Kihara K., Nishida Y., Moriya S., Steinmetz M.O., Hongoh Y., Gönczy P.
(2013) Native architecture of the centriole proximal region reveals features
underlying its 9-fold radial symmetry. Curr. Biol. 23: 1620-1628.
Harris T.J., Peifer M. (2007) aPKC controls microtubule organization to balance
adherens junction symmetry and planar polarity during development. Dev. Cell 12:
727-738.
Harold F.M. (2005) Molecules into cells: specifying spatial architecture. Microbiol.
Mol. Biol. Rev. 69: 544-564.
Hartman H., Puma J.P., Gurney T. (1974) Evidence for the association of RNA with
the ciliary basal bodies of Tetrahymena. J. Cell Sci. 16: 241-259.
Harumoto T., Ito M., Shimada Y., Kobayashi T.J., Ueda H.R., Lu B., Uemura T.
(2010) Atypical cadherins Dachsous and Fat control dynamics of noncentrosomal
microtubules in planar cell polarity. Dev. Cell 19: 389-401.
Hehnly H., Chen C-T., Powers C.M., Liu H-L., Doxsey S. (2012) The centrosome
regulates the Rab11- dependent recycling endosome pathway at appendages of the
MC. Curr. Biol.
22: 1944-1950.

173
Hiedemann S.R., Sander G., Kirschner M.W. (1977) Evidence for a functional role
of RNA in centrioles. Cell 10: 337-350.
Hildreth V., Webb S., Chaudhry B. (2009) Left cardiac isomerism in the Sonic
hedgehog null mouse. J. Anat. 214: 894-904.
Hiraki M., Nakazawa Y., Kamiya R., Hirono M. (2007) Bld10p constitutes the
cartwheel-spoke tip and stabilizes the 9-fold symmetry of the centriole. Curr. Biol.
17: 1778-1783.
Hoops H., Witman G. (1983) Outer doublet heterogeneity reveals structural polarity
related to beat direction in Chlamydomonas flagella. J. Cell Biol. 97: 902-908.
Huang J., Huang L., Chen Y.J., Austin E., Devor C.E., Roegiers F., Hong Y. (2011)
Differential regulation of adherens junction dynamics during apical-basal
polarization. J. Cell Sci. 124: 4001-
4013.
Ibañes M., Izpisúa-Belmonte J.C. (2008) Theoretical and experimental approaches to
understand morphogen gradients. Mol. Syst. Biol. 4: 176.
Ishigaki Y., Nakamura Y., Tatsuno T., Hashimoto M., Iwabuchi K.,Tomosugi N.
(2013) RNA-binding protein RBM8A (Y14) and MAGOH localize to centrosome in
human A549 cells. Histochem. Cell Biol. (E-pub ahead of print).
Ivanov A., McCall I., Babbin B., Samarin S., Nusrat A., Parkos C. (2006)
Microtubules regulate disassembly of epithelial apical junctions. BMC Cell Biol. 7:
12.
Jakobsen L., Vanselow K., Skogs M., Toyoda Y., Lundberg E., Poser., Falkenby
L.G., Bennetzen M., Westendorf J., Nigg E.A., Uhlen M., Hyman A.A., Andersen J.S.
(2011) Novel asymmetrically localizing components of human centrosomes identified
by complementary proteomics methods. EMBO J. 30: 1520-1535.
Jerka-Dziadosz M., Gogendeau D., Klotz C., Cohen J., Beisson J., Koll F. (2010)
Basal body duplication in Paramecium: the key role of Bld10 in assembly and

174
stability of the cartwheel. Cytoskeleton 67: 161-171.
Jonsdottir A.B., Dirks R.W., Vrolijk J., Ögmundsdottir H.M., Tanke H.J., Eyfjörd
J.E., Szuhai K. (2010) Centriole movements in mammalian epithelial cells during
cytokinesis. BMC Cell Biol. 11: 34.
Kadir S., Astin J.W., Tahtamoun L., Martin P., Nobes C.D. ( 2011) Microtubule
remodelling is required for the front–rear polarity switch during contact inhibition of
locomotion. J. Cell Sci. 124: 2642-2653.
Karr T.L., Alberts B.M. (1986) Organization of the Cytoskeleton in Early Drosophila
Embryos. J. Cell Biol. 102: 1494-1509.
Katsaros C., Karyophyllis D., Galatis B. (2006) Cytoskeleton and morphogenesis in
brown algae. Oxford J. Life Sci. Ann. Botany 97: 679-693.
Keller R. (2006) Mechanisms of elongation in embryogenesis. Development. 133:
2291-2302.
Kirschner M., Mitchison T. (1986) Beyond self-assembly: from microtubules to
morphogenesis. Cell 45: 329-342.
Kitagawa D., Vakonakis I., Olieric N., Hilber M., Keller D., Olieric V., Bortfeld M.,
Erat M.C., Flückiger I., Gönczy P., Steinmetz M.O. (2011) Structural basis of the 9-
fold symmetry of centrioles. Cell 144: 364-375.
Kondo S. (1992) A mechanistic model for morphogenesis and regeneration of limbs
and imaginal discs. Mech. Dev. 39: 161-170.
König A., Happle R., Bornholdt D., Engel H., Grzeschik K.H. (2000) Mutations in
the NSDHL gene, encoding a 3beta-hydroxysteroid dehydrogenase, cause CHILD
syndrome. Am. J. Med. Genet. 14; 90: 339-346.
Kotadia S., Crest J., Tram U., Riggs B., Sullivan W. (2010) Blastoderm formation
and cellularisation in Drosophila melanogaster. In: Encyclopedia of Life Sciences
(ELS). John Wiley & Sons, Ltd: Chichester.
Kubo A., Sasaki H., Yuba-Kubo A., Tsukita S., Shiinaa N. (1999) Centriolar

175
satellites. J. Cell Biol. 147: 969-980.
Laane M.M., Haugli F.B. (1974) Division centers in mitotic nuclei of Physarum
polycephalum plasmodia. Norw. J. Bot. 21: 309-318.
Lacomble S., Vaughan S., Gadelha C., Morphew M.K., Shaw M.K., McIntosh J.R.,
Gull K. (2010) Basal body movements orchestrate membrane organelle division and
cell morphogenesis in Trypanosoma brucei. J. Cell Sci. 123: 2884-2891.
Lander A.D., Lo W., Nie Q., Wan F. (2009) The measure of success: constraints,
objectives, and tradeoffs in morphogen-mediated patterning. Cold Spring Harb.
Perspect. Biol. I-.a002022.
Lawo S., Hasegan M., Gupta G.D., Pelletier L. (2012) Subdiffraction imaging of
centrosomes reveals higher-order organizational features of pericentriolar material.
Nat. Cell Biol. 14: 1148-1158.
Lee J., Harland R. (2007) Actomyosin contractility and microtubules drive apical
constriction in Xenopus bottle cells. Dev. Biol. 311: 40-52.
Levin M., Mercola M. (1999) Gap junction-mediated transfer of left - right patterning
signals in the early chick blastoderm is upstream of Shh asymmetry in the node.
Development 126: 4703-4714.
Levin M., Palmer A.R. (2007) Left-right patterning from the inside out: widespread
evidence for intracellular control. BioEssays 29: 271-287.
Lin C.C., Cheng T.S.,Hsu C,H., Wu C.H.,Chang L.S.,Shen Z.S., Yeh H.M., Chang L-
K., Howng S.H., Hong Y.R. (2006) Characterization and functional aspects of human
ninein isoforms that regulated by centrosomal targeting signals and evidence for
docking sites to -tubulin. Cell Cycle 5: 2517-2527.
Linnaeus C. (1751) Philosophia botanica. Stockholm, Godofr. Kiesewetter.
Lobikina M., Wang G., Xu J., Hsieh Y-W., Chuang C-F., Lemirea J.M., Levin M.
(2012) Early, notciliary role for microtubule proteins in left-right patterning is
conserved across kingdoms. Proc. Natl. Acad. Sci. U.S.A. 109: 12586-12591.
Lodish A.B. (2012) Molecular Cell Biology. W.H. Freeman & Co., New York.
Loncarek J., Khodjakov A. (2009) Ab ovo or de novo? Mechanisms of centriole

176
duplication. Mol. Cells 27: 135-142.
López de Heredia M., Jansen R.P. (2004) mRNA localization and the cytoskeleton.
Curr. Opin. Cell Biol. 16: 80-85.
Marshall W.F. (2011) Origins of cellular geometry. BMC Biol. 9: 57.
Marshall W.F. (2012) Centriole asymmetry determines algal cell geometry. Curr.
Opin. Plant Biol. 15: 632-637.
McGhee J.D. (2007) The C. elegans intestine. On line at:
http://www.wormbook.org/chapters/www_intestine/intestine.html, accessed June
29,2012.
Mennella V., Keszthelyi B., McDonald K.L., Chhun B., Kan F., Rogers G.C., Huang
B., Agard D.A. (2012) Subdiffraction-resolution fluorescence microscopy reveals a
domain of the centrosome critical for pericentriolar material organization. Nat. Cell
Biol.14: 1159-1168.
Metzger R.J., Klein O.D., Martin G.R., Krasnow M.A. (2008) The branching
program of mouse lung development. Nature 453: 745-750.
Miller J., Fraser S.E., McClay D. (1995) Dynamics of thin filopodia during sea
urchin gastrulation.
Development 121: 2501-2511.
Mirth C., Akam M. (2002) Joint development in the Drosophila leg: cell movements
and cell populations.
Dev. Biol. 246: 391-406.
Mogensen M.M., Malik A., Piel M., Bouckson-Castaing V., Bornens M. (2000)
Microtubule minus-end anchorage at centrosomal and non-centrosomal sites: the role
of ninein. J. Cell Sci. 113: 3013-3023.
Montcouquiol M., Kelley M.W. (2003) Planar and vertical signals control cellular
differentiation and patterning in the mammalian cochlea. J. Neurosci. 23: 9469-9478.
Morris V.B. (2007) Origins of radial symmetry identified in an echinoderm during
adult development and the inferred axes of ancestral bilateral symmetry. Proc. Biol.
Sci. 27: 1511-1516.

177
Morris V.B., Selvakumaraswamy P., Whan R., Byrne M. (2009) Development of the
five primary podia from the coeloms of a sea star larva: homology with the echinoid
echinoderms and other deuterostomes. Proc. Biol. Sci. 276: 1277-1284.
Moss D., Bellett G., Carter J., Liovic M., Keynton J., Prescott P., Lane E., Mogensen
M. (2007) Ninein is released from the centrosome and moves bi-directionally along
microtubules. J. Cell Sci. 120: 3064-3074.
Motegi F., Seydoux G. (2007) Revisiting the role of microtubules in C. elegans
polarity. J. Cell Biol. 179: 36736-36739.
Müller-Reichert T., Greenan G., O’Toole E., Srayko M. (2010) The elegans of
spindle assembly. Cell Mol. Life Sci. 67: 2195-2213.
Nishimura T., Honda H., Takeichi M. (2012) Planar cell polarity links axes of spatial
dynamics in neural-tube closure Cell 149: 1084-1097.
Okabe N., Xu B., Burdine R.D. (2008) Fluid dynamics in zebrafish Kupffer's vesicle.
Dev. Dyn. 237: 3602-3612.
Oma Y., Harata M. (2011) Actin-related proteins localized in the nucleus. Actin-
related proteins localized in the nucleus. Nucleus. 2: 38-46.
O'Toole E., Greenan G., Lange K.I., Srayko M., Müller-Reichert T. (2012) The role
of γ-tubulin in centrosomal microtubule organization. PLoS One 7: e29795.
O'Toole E., Giddings T.H., McIntosh R.J., Dutcher S.K. (2003) Three-dimensional
organization of basal bodies from wild-type and δ-tubulin deletion strains of
Chlamydomonas reinhardtii. Mol. Biol. Cell 14: 2999 -3012.
Ou Y.Y., Mack G.J., Zhang M., Rattner J.B. (2002). CEP110 and ninein are located in
a specific domain of the centrosome associated with centrosome maturation. J. Cell
Sci. 115: 1825-1835.
Ounjai P., Kim K.D., Liu H., Dong M., Tauscher A.N., Witkowska H.E., Downing
K.H. (2013) Architectural insights into a ciliary partition. Curr. Biol. 23: 339-344, 31
Palmer A.R. (2010) Anatomical midplane: fact or fiction? On line at:
http://www.biology.ualberta.ca/Palmer/asym/axes/midplane.htm, accessed June
29,2012.

178
Paradez A., Wright A., Ehrhardt D.W. (2006) Microtubule cortical array organization
and plant cell morphogenesis. Curr. Opin. Plant Biol. 9: 571-578.
Park R.S., Légier M.F., Cartwright J. Jr, Goldstein M.A. (1984) Perinuclear
microtubules in postnatal rat heart. J. Morphol. 179: 13-19.
Pearson C.G., Winey M. (2009) Basal body assembly in ciliates: the power of
numbers. Traffic 10:
461-471.
Peyre E., Jaouen F., Saadaoui M., Haren L., Merdes A., Durbec P., Morin X. (2011) A
lateral belt of cortical LGN and NuMA guides mitotic spindle movements and planar
division in neuroepithelial cells. J. Cell Biol. 193: 141-154.
Picone R., Ren X., Ivanovitch K., Clarke J., McKendry A., Baum B. (2010) A
polarised population of dynamic microtubules mediates homeostatic length control in
animal cells. PLoS Biol. 8: e1000542.
Piel M., Meyer P., Khodjakov A., Rieder C.L., Bornens M. (2000) The respective
contributions of the MC and DC to centrosome activity and behavior in vertebrate
cells. J. Cell
Pope K.L., Harris T.J. (2008) Control of cell flattening and junctional remodeling
during squamous epithelial morphogenesis in Drosophila. Development 135: 2227-
2238.
Pulina M.V., Hou S., Mittal A., Julich D., Whittaker C.A., Holley S.A., Hynes R.O.,
Astrof S. (2011) Essential roles of fibronectin in the development of the left-right
embryonic body plan. Dev. Biol. 354: 208-220.
Qiao R., Cabral G., Lettman M.M., Dammermann A., DongG. (2012) SAS-6 coiled-
coil structure and interaction with SAS-5 suggest a regulatory mechanism in C.
elegans centriole assembly. The EMBO J. 31: 4334-4347.
Rabinowitz J.S., Lambert J.D. (2010) Spiralian quartet developmental potential is
regulated by specific localization elements that mediate asymmetric RNA

179
segregation. Development 137:4039-49.
Rauzi M., Lenne P.F., Lecuit T. (2010) Planar polarized actomyosin contractile flows
control epithelial junction remodelling. Nature 468: 1110-1114.
Regolini M. (2013) Centrosome: is it a geometric, noise resistant, 3D interface that
translates morphogenetic signals into precise locations in the cell? Ital. J. Anat. Embr.
118: 19-66.
Riparbelli M.G., Dallai R., Mercati D., Bu Y., Callaini G. (2009) Centriole
symmetry: a big tale from small organisms. Cell Motil. Cytoskeleton 66:1100-5.
Riparbelli M.G., Callaini G. (2011) Male gametogenesis without centrioles. Dev.
Biol. 349: 427-439.
Ritter A., Griffiths G. (2012) http://www.cell.com/cell_picture_show-cancer.
Rodrigues-Martins A., Riparbelli M., Callaini G., Glover D.M., Bettencourt-Dias
M. (2007) Revisiting the role of the MC in centriole biogenesis. Science 316: 1046-
1050.
Rodrigues-Martins A., Bettencourt-Dias M., Riparbelli MM., Ferreira C., Ferreira I.,
Callaini G., Glover D.M. (2007) DSAS-6 organizes a tube-like centriole precursor,
and its absence suggests modularity in centriole assembly. Curr. Biol. 17: 1465-1472.
Roque H., Wainman A., Richens J., Kozyrska K., Franz A., Ra J.W. (2012)
Drosophila Cep135/Bld10 maintains proper centriole structure but is dispensable for
cartwheel formation. J. Cell Sc. 125: 5881-6.
Rosenbaum J.L., Moulder J.E., Ringo D.L. (1969) Flagellar elongation and
shortening in Chlamydomonas. The use of cycloheximide and colchicine to study the
synthesis and assembly of flagellar proteins. J. Cell Biol. 41: 600-619.
Roy S., Hsiung F., Kornberg T.B. (2011) Specificity of Drosophila cytonemes for
distinct signalling pathways. Science 332: 354-358.
Salinas-Saavedra M., Vargas A.O. (2011) Cortical cytasters: a highly conserved
developmental trait of Bilateria with similarities to Ctenophora. Evodevo 1: 23.
Salman H., Abu-Arish A., Oliel S., Loyter A., Klafter J., Granek R., Elbaum M.
(2005) Nuclear localization signal peptides induce molecular delivery along

180
microtubules. Biophys. J. 89: 2134-2145.
Sànchez Alvarado A.
http://planaria.neuro.utah.edu/publications/Current%20BIology.pdf
http://www.sciencedaily.com/releases /2012/01/120105141138.htm, related to:
Azimzadeh et al., 2012.
Sardet C., Paix A., Prodon F., Dru P., Chenevert J. (2007) From oocyte to 16-cell
stage: cytoplasmic and cortical reorganizations that pattern the ascidian embryo. Dev.
Dyn. 236: 1716-1731.
Savriama Y., Klingenberg C.P. (2011) Beyond bilateral symmetry: geometric
morphometric methods for any type of symmetry. BMC Evol. Biol. 11: 280.
Sawyer J.M., Harrell J.R., Shemer G., Sullivan-Brown J., Roh-Johnson M.,
Goldstein B.( 2010) Apical constriction: a cell shape change that can drive
morphogenesis. Dev. Biol. 341: 5-19.
Schatten H. The mammalian centrosome and its functional significance. (2008)
Histochem. Cell Biol. 129: 667-686.
Schenk C., Bringmann H., Hyman A., Cowan C. (2010) Cortical domain correction
repositions the polarity boundary to match the cytokinesis furrow in C. elegans
embryos. Development 13 7: 1743-1753.
Schliwa M., Pryzwansky K.B., Euteneuer U. (1982) Centrosome splitting in
neutrophils: an unusual phenomenon related to cell activation and motility. Cell 31:
705-717.
Schmidt K.N., Stefanie K., Neuner A., Hub B., Zentgraf H., Pereira G. (2012)
Cep164 mediates vesicular docking to the MC during early steps of ciliogenesis. J.
Cell Biol. 199: 1083-1101.
Schnackenberg B.J., Khodjakov A., Rieder C.L., Palazzo R.E. (1998) The
disassembly and reassembly of functional centrosomes in vitro. Proc. Natl. Acad. Sci.

181
USA 95: 9295-9300.
Scott F.G. (2014) Developmental biology 10th Edition, Sinauer Associates,
Sunderland USA.Chapt. 5 p. 165
Sieberer B.J., Kieft H., Franssen-Verheijen T., Emons A.M., Vos J.W. (2009) Cell
proliferation, cell shape, and microtubule and cellulose microfibril organization of
tobacco BY-2 cells are not altered by exposure to near weightlessness in space. Planta
230: 1129-1140.
Silflow C.D., LaVoie M., Tam L., Tousey S., Sanders M., Wu C.W., Borodovsky M.,
Lefebvrea P.A. (2001) The Vfl1 protein in Chlamydomonas localizes in a rotationally
asymmetric pattern at the distal ends of the basal bodies. J. Cell Biol. 153: 63-74.
Simões S., Denholm B., Azevedo D., Sotillos S., Martin P., Hombría J., Jacinto A.
(2006) Compartmentalisation of Rho regulators directs cell invagination during tissue
morphogenesis. Development 133: 4257-4267.
Smith K.A., Chocron S., von der Hardt S., de Pater E., Soufan A., Bussmann J.,
Schulte-Merker S., Hammerschmidt M., Bakkers J. (2008) Rotation and asymmetric
development of the zebrafish heart requires directed migration of cardiac progenitor
cells. Dev. Cell 14: 287-297.
Snyder J.A. (1980) Evidence for a ribonucleoprotein complex as a template for
microtubule initiation in vivo. Cell Biol. Int. Rep. 4: 859-868.
Song M.H., Miliaras N.B., Peel N., O'Connell K.F. (2008) Centrioles: some self-
assembly required. Curr. Opin. Cell Biol. 20: 688-693.
Sonnen K.F., Schermelleh L. Leonhardt H., NiggE.A. (2012) 3D-structured
illumination microscopy provides novel insight into architecture of human
centrosomes. Biol. Open 1: 965-976.
Strutt D. (2009) Gradients and the specification of planar polarity in the insect
cuticle. Cold Spring Harb Perspect. Biol. 1: a000489.
Strachan T., Read A. (2010) Human Molecular Genetics. Garland Science, New
York. Chapters 9.3, 10 and p. 334.
Sugiyama Y., Lovicu F.J., McAvoy J.W. (2011) Planar cell polarity in the mammalian

182
eye lens. Organogenesis 7: 191-201.
Takano K., Ito Y., Obata S., Oinuma T., Komazaki S., Nakamura H., Asashima M.
(2007) Heart formation and left-right asymmetry in separated right and left embryos
of a newt. Int. J. Dev. Biol. 51: 265-272.
Takesono A., Heasman S., Wojciak-Stothard B., Garg R., Ridley A. (2010)
Microtubules regulate migratory polarity through Rho/ROCK signalling in T cells.
PLoS One 5: e8774.
Taylor J., Adler P.N. (2008) Cell rearrangement and cell division during the tissue
level morphogenesis of evaginating Drosophila imaginal discs. Dev. Biol; 313: 739–
751.
Thompson D'A.W. (1917) On Growth and Form. Cambridge, Cambridge University
Press.
Tsai M.C., Ahringer J. (2007) Microtubules are involved in anterior-posterior axis
formation in C. elegans embryos. J. Cell Biol. 179: 397-402.
Tsukaya H. (2005) Leaf shape: genetic controls and environmental factors. Int. J.
Dev. Biol. 49: 547-555.
Urbéa S., Liua H., Hayesa S.D., Heridea C., Rigdenb D.J., Claguea M.J. (2012)
Systematic survey of deubiquitinase localization identifies USP21 as a regulator of
centrosome- and microtubule-associated functions. Mol. Biol. Cell 23: 1095-1103.
van Breugel M., Hirono M., Andreeva A., Yanagisawa H.A., Yamaguchi S.,
Nakazawa Y., Morgner N., Petrovich M., Ebong I.O., Robinson C.V., Johnson C.M.,
Veprintsev D., Zuber B. (2011) Structures of SAS-6 suggest its organization in
centrioles. Science 331: 1196-1199.
Vandenberg L.N., Levin M. (2009) Perspectives and open problems in the early
phases of left-right patterning. Semin. Cell Dev. Biol. 20: 456-463.
Vandenberg L.N., Morrie R.D., Seebohm G., Lemire J.M., Levin M. (2013) Rab
GTPases are required for early orientation of the left-right axis in Xenopus. Mech.
Dev. 130: 254-271.

183
Vellutini B.C., Migotto A.E. (2010) Embryonic, larval, and juvenile development of
the sea biscuit Clypeaster subdepressus (Echinodermata: Clypeasteroida). PLoS One
5: e9654.
Vinogradova T., Paul R., Grimaldi A.D., Loncarek J., Miller P.M., Yampolsky D.,
Magidson V., Khodjakov A., Mogilner A., Kaverina I. (2012) Concerted effort of
centrosomal and Golgi-derived microtubules is required for proper Golgi complex
assembly but not maintenance. Mol. Biol. Cell 23: 820-833.
Vladar E.K., Antic D., Axelrod J.D. (2009) Planar cell polarity signalling: the
developing cell’s compass. Cold Spring Harb. Perspect. Biol. 1: a002964.
Vorobjev A., Chentsov Y.S. (1982) Centrioles in the cell cycle. I. Epithelial cells. J.
Cell Biol. 98: 938-949.
Vuzman D., Levy Y. (2012) Intrinsically disordered regions as affinity tuners in
protein-DNA interactions. Mol Biosyst. 8:47-57.
Wakida N., Botvinick E., Lin J., Berns M. (2010) An intact centrosome is required
for the maintenance of polarization during directional cell migration. PLoS 5:
e15462.
Wang W., Kirkness E.F. (2005) Short interspersed elements (SINEs) are a major
source of canine genomic diversity. Genome Res. 15: 1798-1808.
Wargo M.J., Smith E.F. (2003) Asymmetry of the central apparatus defines the
location of active microtubule sliding in Chlamydomonas flagella. Proc. Natl. Acad.
Sci. USA 100: 137-142.
Wasteneys G.O. (2002) Microtubule organization in the green kingdom: chaos or
self-order? J. Cell Sci. 115: 1345-1354.
Wasteneys G.O., Ambrose J.C. (2009) Spatial organization of plant cortical
microtubules: close encounters of the 2D kind. Trends Cell Biol. 19: 62-71.
Waterman-Storer C., Dueya D., Weberb K., Keechb J., Cheneyd R., Salmone E.,
Bement W. (2000) Microtubules remodel actomyosin networks in Xenopus egg
extracts via two mechanisms of F-actin transport. J. Cell Biol. 150: 361-376.
Wolpert L. quoted by Gilbert F.S. (2010) The polar coordinated model of positional

184
information in the developing and regenerating limbs. 9th edn. Sinauer Associates,
Sunderland. Ch. 15 Page 153.
Wood C.R., Huang K., Diener D.R., Rosenbaum J.L. (2013) The cilium secretes
bioactive ectosomes. Curr. Biol. 23: 906-911.
Woodard G.E., Huang N.N., Cho H., Miki T., Tall G.G., Kehrl J.H. (2010) Ric-8A
and Gi alpha recruit LGN, NuMA, and dynein to the cell cortex to help orient the
mitotic spindle. Mol. Cell Biol. 30: 3519-3530.
Wühr M., Tan E., Parker S., Detrich H., Mitchison T. (2010) A model for cleavage
plane determination in early amphibian and fish embryos. Curr. Biol. 20: 2040-2045.
Xu J., Van Keymeulen A., Wakida N.M., Carlto P., Berns M.W., Bourne H.R. (2007)
Polarity reveals intrinsic cell chirality. Proc. Natl. Acad. Sci. USA 104: 9296-9300.
Yoshinori M., Yukio O. (2010) CLERC and centrosomal leucine-rich repeat proteins.
Cent. Eur. J. Biol. 5: 1-10.
Zallen J.A. (2007) Planar polarity and tissue morphogenesis. Cell. 129: 1051-1063.
Ziegle A., Fabe C., Bartolomaeus T. (2009) Comparative morphology of the axial
complex and interdependence of internal organ systems in sea urchins
(Echinodermata: Echinoidea). Front Zool. 6: 10.




















