
A Subset of NKT Cells That Lacks the NK1.1 Marker, Expresses CD1d Molecules, and Autopresents the...
11
of May 11, 2014. This information is current as Antigen -Galactosylceramide α Autopresents the Marker, Expresses CD1d Molecules, and A Subset of NKT Cells That Lacks the NK1.1 Jean-François Bach and André Herbelin Jean-Marc Gombert, Corinne Garcia, Yasuhiko Koezuka, Agathe Hameg, Irina Apostolou, Maria Leite-de-Moraes, http://www.jimmunol.org/content/165/9/4917 doi: 10.4049/jimmunol.165.9.4917 2000; 165:4917-4926; ; J Immunol References http://www.jimmunol.org/content/165/9/4917.full#ref-list-1 , 25 of which you can access for free at: cites 39 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2000 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on May 11, 2014 http://www.jimmunol.org/ Downloaded from by guest on May 11, 2014 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A Subset of NKT Cells That Lacks the NK1.1 Marker, Expresses CD1d Molecules, and Autopresents the...

of May 11, 2014.This information is current as
Antigen-GalactosylceramideαAutopresents the
Marker, Expresses CD1d Molecules, and A Subset of NKT Cells That Lacks the NK1.1
Jean-François Bach and André HerbelinJean-Marc Gombert, Corinne Garcia, Yasuhiko Koezuka, Agathe Hameg, Irina Apostolou, Maria Leite-de-Moraes,
http://www.jimmunol.org/content/165/9/4917doi: 10.4049/jimmunol.165.9.4917
2000; 165:4917-4926; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/9/4917.full#ref-list-1
, 25 of which you can access for free at: cites 39 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

A Subset of NKT Cells That Lacks the NK1.1 Marker,Expresses CD1d Molecules, and Autopresents thea-Galactosylceramide Antigen1
Agathe Hameg,* Irina Apostolou,† Maria Leite-de-Moraes,‡ Jean-Marc Gombert,§
Corinne Garcia,¶ Yasuhiko Koezuka,i Jean-Francois Bach,* and Andre Herbelin2*
In the present report, we characterize a novel T cell subset that shares with the NKT cell lineage both CD1d-restriction and highreactivity in vivo and in vitro to the a-galactosylceramide (a-GalCer) glycolipid. These cells preferentially use the canonicalVa14-Ja281 TCR-a-chain and Vb8 TCR-b segments, and are stimulated bya-GalCer in a CD1d-dependent fashion. However,in contrast to classical NKT cells, they lack the NK1.1 marker and express high surface levels of CD1d molecules. In addition, thisNK1.12 CD1dhigh T subset, further referred to as CD1dhigh NKT cells, can be distinguished by its unique functional features.Although NK1.11 NKT cells require exogenous CD1d-presenting cells to make them responsive toa-GalCer, CD1dhigh NKT cellscan engage their own surface CD1d in an autocrine and/or paracrine manner. Furthermore, in response toa-GalCer, CD1dhigh
NKT cells produce high amounts of IL-4 and moderate amounts of IFN-g, a cytokine profile more consistent with a Th2-likephenotype rather than the Th0-like phenotype typical of NK1.11 NKT cells. Our work reveals a far greater level of complexitywithin the NKT cell population than previously recognized and provides the first evidence for T cells that can be activated uponTCR ligation by CD1d-restricted recognition of their ligand in the absence of conventional APCs. The Journal of Immunology,2000, 165: 4917–4926.
CD1 molecules represent a family of conserved nonpoly-morphic cell surface glycoproteins related to the MHCclass I and class II molecules, but encoded by genes out-
side the MHC (1). In accordance with their MHC Ag-presentingmolecule-like structure, and their expression by APCs, they havebeen described as targets for T cell recognition (1–3). CD1d-re-active T cells have been initially identified in the mouse. They areautoreactive in vitro to CD1d-expressing cells (1, 4) and display alarge specificity pattern toward various CD1d-positive cell typesand transfectants (5, 6). Among them, NKT cells have been arecent focus of interest (2, 7). In the thymus, the NKT cell subset,which includes both double-negative (DN)3 and CD41 T cells, ischaracterized by the expression of NK lineage-specific receptors,namely NK1.1 (CD161), and the preferential usage of the invariantVa14-Ja281 TCR-a-chain paired with Vb8, Vb7, or Vb2 TCR-bsegments (2, 8). These cells can also be distinguished by their
intermediate level of TCR expression and their activated/memory-like phenotype (2, 8). A human counterpart of the mouse NKTcells has been identified. It expresses a restricted repertoire ho-mologous to the one expressed by mouse NKT cells and is reactiveto CD1d (9, 10). The unique capacity of NKT cells to promptlyrelease IFN-g and IL-4 upon TCR engagement is thought to be thebasis of their various regulatory functions during the effector phaseof immune responses, including regression of tumor metastases(11–13), regulation of autoimmune diseases (14–16), and the pro-tection against bacterial or parasitic infections (17–19).
Although the early cytokine response in the spleen of mice hav-ing received an i.v. injection of anti-CD3 mAb can be ascribed toNKT cells (20, 21), little is known about their physiological stim-uli. Recently,a-galactosylceramide (a-GalCer), originally isolatedfrom marine sponge (22), was found to activate most Va14-Ja2811/Vb81 NKT cells in a CD1d-dependent fashion (23–25),suggesting that these cells might recognize unidentified self-gly-colipids (24, 25). The responsiveness toa-GalCer seems to berestricted mainly to Va14-Ja2811 NKT cells because most Va14-Ja2811 hybridomas are activated bya-GalCer, conversely toCD1d-autoreactive hybridomas that do not use the Va14 TCR(24, 25).
We have recently reported that CD1d-restricted T lymphocytesthat share with NK1.11 T cells the capacity to produce largeamounts of IL-4 and IFN-g upon TCR cross-linking but do notexpress the NK1.1 molecule can be identified ex vivo (26). Inkeeping with these findings, other studies have provided evidencefor class-I-restricted and/or CD1d-reactive cells among memory/activated CD41 NK1.12 T lymphocytes (4, 27). Analyses of theCD1d-restricted CD41 compartment in mice lacking MHC class IImolecules (I-Ab2/2 C57BL/6 mice) have shown that it can besubdivided into NK1.11 and NK1.12 cell populations, whoseTCR repertoire was found to be skewed toward Vb8 segments (4,26–28). Moreover, Skold et al. (28) demonstrated that some of
*Institut National de La Sante et de la Recherche Medicale (INSERM) Unite 25 andCentre Claude Bernard, Hopital Necker, Paris, France;†INSERM Unite 277, InstitutPasteur, Paris, France;‡Centre National de la Recherche Scientifique, Unite Mixte deRecherche 8603, Universite Rene Descartes, Hopital Necker, Paris, France;§Labo-ratoire d’Immunologie-Immunopathologie, Centre Hospitalier Universitaire, Poitiers,France;¶INSERM Unite 373, Institut Necker, Paris, France; andiPharmaceuticalResearch Laboratory, Kirin Brewery Company, Gunma, Japan
Received for publication May 1, 2000. Accepted for publication August 3, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by institute funds from Institut National de La Sante´ et dela Recherche Medicale and the Ligue Nationale contre le Cancer (Axe Immunologie,2000). A.H. is supported by the Fondation pour la Recherche Medicale.2 Address correspondence and reprint requests to Dr. Andre Herbelin, Institut Na-tional de La Sante et de la Recherche Medicale Unite 25, Hopital Necker, 161 rue deSevres, 75743 Paris Cedex 15, France. E-mail address: [email protected] Abbreviations used in this paper: DN, double-negative; HSA, heat-stable Ag;a-Gal-Cer, a-galactosylceramide.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
these cells express the Va14-Ja281 rearrangement typical ofNK1.11 T cells.
To date,a-GalCer reactivity during primary cultures has beeninvestigated exclusively in the NK1.11 T cell subset (24). In thepresent study, we addressed the question whether the CD1d-re-stricted NK1.12 T cells were also responsive to this glycolipid Ag.To this end, we isolated NK1.12 CD41 T splenocytes fromI-Ab2/2 C57BL/6 mice in which CD1d-restricted CD41 T cellsare enriched (4) and identified a subset of CD1d-restricted NK1.12
CD41 T cells that, like classical NKT cells, produce high amountsof IL-4 upon stimulation bya-GalCer in a CD1d-dependent fash-ion. We found that, in contrast to the classical NKT population,CD1d-restricted NK1.12 CD41 T cells display high levels ofCD1d molecules on their surface and possess the remarkable ca-pacity of autopresenting thea-GalCer Ag. The implications of thisproperty for the regulatory functions of NKT cells during the im-mune responses will be discussed.
Materials and MethodsMice
Wild-type and mutant I-Ab2/2 C57BL/6 mice were bred and maintainedin our animal facilities under specific pathogen-free conditions. MutantCD1d2/2 C57BL/6 mice were kindly provided by L. Van Kaer (HowardHughes Medical Institute, Vanderbilt University School of Medicine,Nashville, TN) (29).
a-GalCer
a-GalCer [(2S, 3S, 4R)-1-O-(a-D-galactopyranosyl)-2-(N-hexacosanoyl-amino)-1,3,4-octadecanetriol] (22) was synthesized by Pharmaceutical Re-search Laboratories, Kirin Brewery (Gunma, Japan). The preparation hada single dominant peak of the expected m.w. by electrospray mass spec-trometry, ruling out the presence of significant amounts of degradationproducts. The stock solution was dissolved at 10mg/ml in 10% DMSO, anddiluted in culture medium to a final concentration of 100 ng/ml. A 10%DMSO vehicle solution diluted to a 0.1% final concentration was routinelyused as control.
In vivo treatment witha-GalCer
Mice received a single injection of 2–4mg of a-GalCer, (1–2mg i.v. plus1–2mg i.p.), diluted in NaCl. Control mice were injected with an identicalvolume of vehicle solution alone (0.025% polysorbate). Mice were sacri-ficed 2 h after injection.
Abs and flow cytometry analysis
FITC- and PE-anti-CD1d (clone 1B1), PE-anti-CD24 (clone M1/69),FITC-anti-CD44 (clone 1 M7.8), PE-anti-NK1.1 (clone PK136), FITC- orAPC- anti-TCR-ab (clone H57-597), and APC-anti-CD4 (clone RM4.5)mAbs, PE-anti-IL-4 (clone 11B11) and its PE-isotype control (clone R3-34) mAbs were obtained from PharMingen (San Diego, CA). PE-anti-IFN-g (clone XMG1.2) mAb and its PE-isotype IgG1 control mAb wereobtained from Caltag Laboratories (Burlingame, CA). Anti-CD62-L (cloneMEL-14) and anti-CD1d (clone 20H2) mAbs, kindly provided by F. Le-pault (Centre National de la Recherche Scientifique, Unité Mixte de Re-cherche 8603, Institut Necker, Paris, France) and Dr. A. Bendelac (De-partment of Molecular Biology, Princeton University, Princeton, NJ),respectively, were purified and biotinylated in the laboratory. Anti-CD8(clone 53.6.7), anti-TCR-ab (clone H57-597), anti-Vb8 (clone F23.1), andanti-CD24 (anti-heat-stable Ag (HSA), clone J11d) were purified and/orfluoresceinated or biotinylated in our laboratory. For cell surface markeranalysis, four-color staining was performed as described (26). After incu-bation with the appropriate FITC-labeled, PE-labeled, and biotinylatedmAbs, cells were incubated with the appropriate APC-labeled mAb plusstreptavidin-peridinin chlorophyll protein or streptavidin-Cy-Chrome(PharMingen). Control staining with irrelevant Abs was always performedin parallel. For intracellular cytokine staining, 53 105 cells were incubatedwith FITC-anti-CD1d and biotinylated anti-NK1.1 mAbs. After extensivewashing, cells were incubated with APC-anti-TCR-ab mAb and strepta-vidin-Cy-Chrome. They were fixed in 4% paraformaldehyde for 5 min atroom temperature, washed with PBS containing 1% BSA and 0.5% sapo-nin (Sigma, St. Louis, MO), and incubated with either PE-anti-IL-4 orPE-anti-IFN-g (or with the appropriate PE-labeled-isotype control) for 30min at room temperature. The cells were washed again with PBS/BSA/
saponin and then with PBS/BSA without saponin to allow membrane clo-sure. A FACScalibur cytometer (Becton Dickinson, Mountain View, CA)was used and a minimum of 53 104 events gated from viable cells wereacquired with CellQuest software. Results were analyzed using MacCellQuest software. Each analytical gate contained at least 23 103 events.
Cell preparation
Thymi and spleens were carefully removed from exsanguinated mice.Great care was taken to avoid contamination of thymi by cells from bloodor thoracic lymph nodes. DN and CD41 mature thymocytes were enrichedby treating freshly isolated thymocytes with the IgM Abs Y169 (rat anti-mouse CD8) and J11d (rat anti-mouse heat stable Ag, anti-HSA) plus com-plement killing (Low-Tox rabbit Complement, Cedarlane, Ontario, Can-ada) at 37°C for 40 min. In these conditions, mature CD81, CD41CD81,
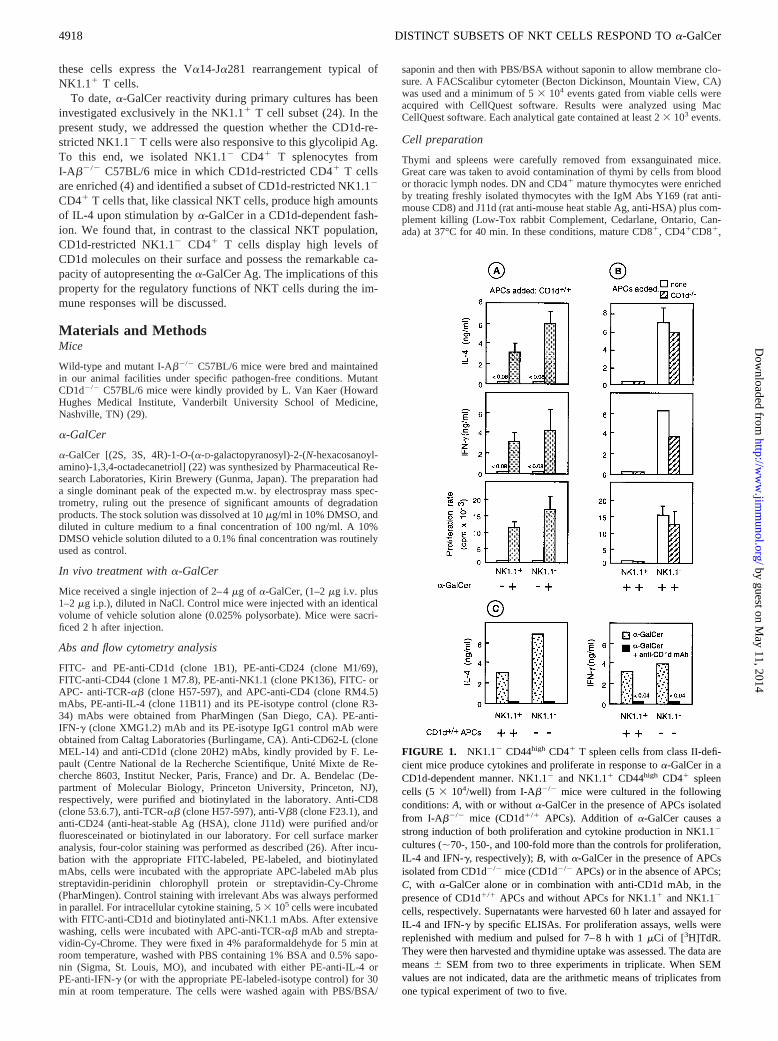
FIGURE 1. NK1.12 CD44high CD41 T spleen cells from class II-defi-cient mice produce cytokines and proliferate in response toa-GalCer in aCD1d-dependent manner. NK1.12 and NK1.11 CD44high CD41 spleencells (5 3 104/well) from I-Ab2/2 mice were cultured in the followingconditions:A, with or withouta-GalCer in the presence of APCs isolatedfrom I-Ab2/2 mice (CD1d1/1 APCs). Addition of a-GalCer causes astrong induction of both proliferation and cytokine production in NK1.12
cultures (;70-, 150-, and 100-fold more than the controls for proliferation,IL-4 and IFN-g, respectively);B, with a-GalCer in the presence of APCsisolated from CD1d2/2 mice (CD1d2/2 APCs) or in the absence of APCs;C, with a-GalCer alone or in combination with anti-CD1d mAb, in thepresence of CD1d1/1 APCs and without APCs for NK1.11 and NK1.12
cells, respectively. Supernatants were harvested 60 h later and assayed forIL-4 and IFN-g by specific ELISAs. For proliferation assays, wells werereplenished with medium and pulsed for 7–8 h with 1mCi of [3H]TdR.They were then harvested and thymidine uptake was assessed. The data aremeans6 SEM from two to three experiments in triplicate. When SEMvalues are not indicated, data are the arithmetic means of triplicates fromone typical experiment of two to five.
4918 DISTINCT SUBSETS OF NKT CELLS RESPOND TOa-GalCer
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
and most immature CD42CD82 thymocytesdied and were further elim-inated by centrifugation on a density gradient (J. Prep.; Techgen, LesUlis, France). Purity of the HSA2CD82 thymocyte preparation waschecked by staining the cells with PE-anti-HSA (clone M.1/69) andFITC-anti-CD8 (clone 53.6.7). In either case, the preparation contained.95% HSA2CD82 cells upon reanalysis. Spleen cell suspensions wereprepared using a homogenizer, and RBCwere lysed in an hemolysisbuffer (NH4Cl, KHCO3, EDTA). Splenocytes were enriched for CD41 Tcells using anti-CD4-coated magnetic beads (Miltenyi Biotech, Bergisch-Gladbach, Germany), as reported (26). Purity of enriched-CD41 cell frac-tions was always greater than 92%, except for I-Ab2/2 mice (75–85%).For cell sorting, MACS enriched-CD41 splenocytes were stained withFITC-anti-CD44, PE-anti-NK1.1, biotinylated-anti-CD62L plus SAv-Red613 (Life Technologies, Grand Island, NY), and APC-anti-CD4.NK1.11 and NK1.12 populations were then sorted (among gated CD44high
CD62L2 CD41 cells) using a FACSvantage cell sorter (Becton Dickin-son). In some experiments, MACS-enriched CD41 splenocytes werestained with FITC-anti-CD1d, PE-anti-NK1.1, and APC-anti-CD4. In theseconditions, CD1dlow NK1.11, CD1dlow NK1.12 and CD1dhigh NK1.12
populations were sorted among gated CD41 cells. Purity of each enrichedcell fraction was$90% upon reanalysis (see inset of Fig. 3A).
Primary cultures of responder spleen cells
To assess cytokine production and proliferation, cells in RPMI 1640 Glu-tamax culture medium (Life Technologies) supplemented with 10% FCS(Techgen), 2-ME 0.05 mM, penicillin (100 IU/ml), and streptomycin (100mg/ml) were cultured (2–53 104/well; 200 ml final volume) in 96-well
round bottom microplates (Nunc, Roskilde, Denmark) with immobilizedanti-TCR-ab (clone H57-597) or solublea-GalCer (100 ng/ml) for 60 h inthe presence or absence of APCs (autologous or heterologous unseparatedspleen cells at 23 105/well g-irradiated at 2500 rad). The blocking anti-CD1d mAb 20H2 (10mg/ml) was added simultaneously to the responderspleen cells. For control cultures, the responder cells were omitted or ve-hicle alone was used instead ofa-GalCer. No significant proliferation orcytokine production was detected in control wells (Fig. 1A). The superna-tants were harvested 60 h later and stored at270°C until IL-4 and IFN-gassays and wells were replenished with medium and pulsed for 7–8 h with1 mCi of [3H]TdR (5 Ci/mM; Amersham, Buckinghamshire, U.K.). Cellswere then harvested and thymidine uptake was measured. Enriched-CD41
spleen cells (1.253 105/well; 0.5 ml final volume) obtained froma-Gal-Cer- or vehicle-treated mice were cultured in 24-well plates (Falcon, Bec-ton Dickinson) without additional stimulus. Supernatants were harvestedafter 2 h and stored at270°C until cytokine assays were performed.
Analysis of Va14-Ca and Va14-Ja281 mRNA expression
Total RNA was extracted with RNABle reagent (Eurobio, Paris, France),followed by ethanol precipitation and resuspension of pellets in 20ml dis-tilled water. Reverse transcription was performed with random hexamersand oligo(dT) priming using standard methods. Quantitative kinetic ELISAPCR was conducted as described using oligonucleotides specific for Ca,Va14, and Ja281 (30). In this method, for two samples containing the sameamount of Ca, a shift ofn cycles along thex-axis of the two amplificationcurves represents an;1.8n-fold difference in the two samples studied.
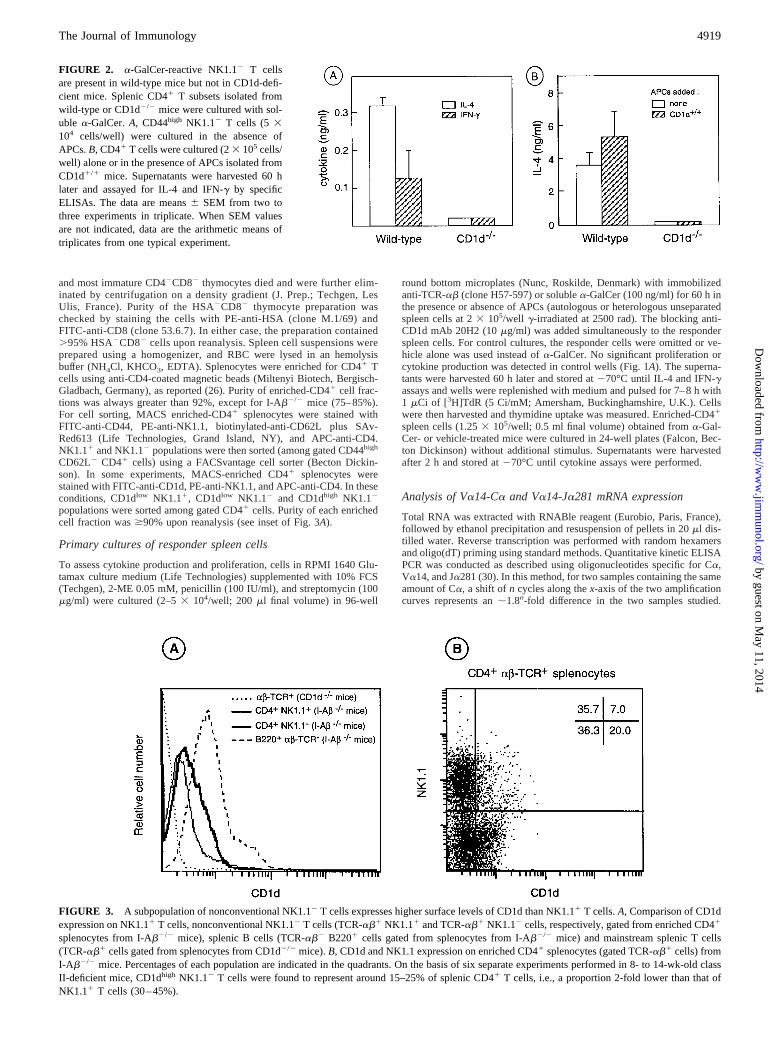
FIGURE 2. a-GalCer-reactive NK1.12 T cellsare present in wild-type mice but not in CD1d-defi-cient mice. Splenic CD41 T subsets isolated fromwild-type or CD1d2/2 mice were cultured with sol-uble a-GalCer.A, CD44high NK1.12 T cells (5 3104 cells/well) were cultured in the absence ofAPCs.B, CD41 T cells were cultured (23 105 cells/well) alone or in the presence of APCs isolated fromCD1d1/1 mice. Supernatants were harvested 60 hlater and assayed for IL-4 and IFN-g by specificELISAs. The data are means6 SEM from two tothree experiments in triplicate. When SEM valuesare not indicated, data are the arithmetic means oftriplicates from one typical experiment.
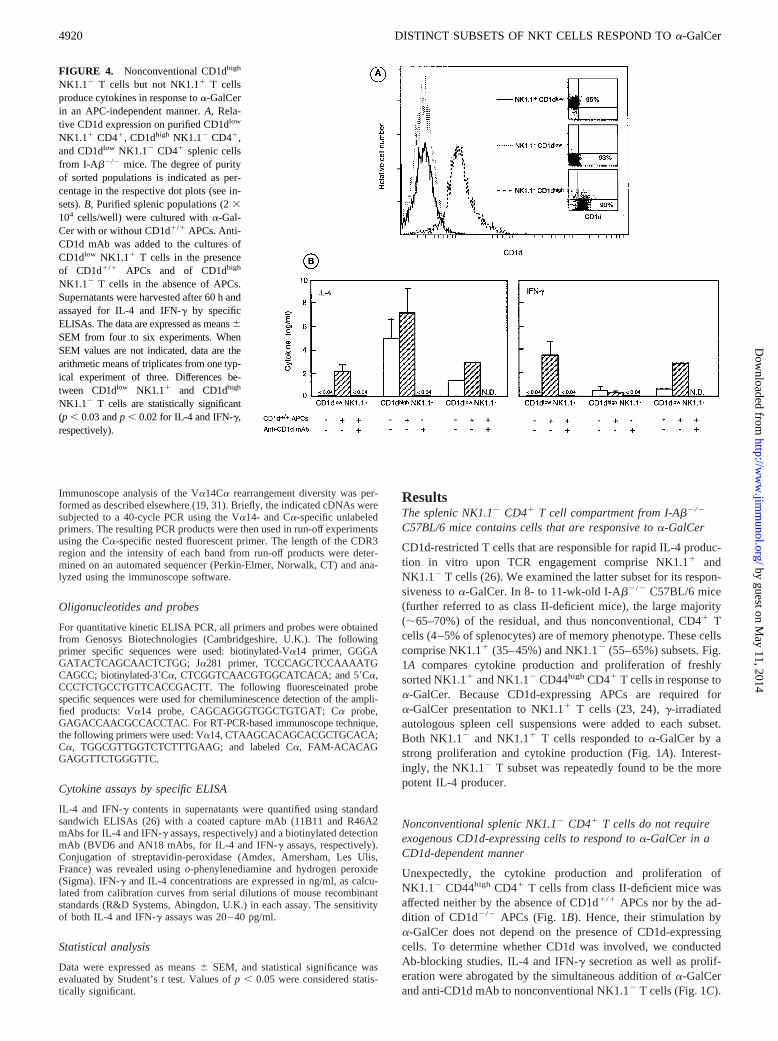
FIGURE 3. A subpopulation of nonconventional NK1.12 T cells expresses higher surface levels of CD1d than NK1.11 T cells.A, Comparison of CD1dexpression on NK1.11 T cells, nonconventional NK1.12 T cells (TCR-ab1 NK1.11 and TCR-ab1 NK1.12 cells, respectively, gated from enriched CD41
splenocytes from I-Ab2/2 mice), splenic B cells (TCR-ab2 B2201 cells gated from splenocytes from I-Ab2/2 mice) and mainstream splenic T cells(TCR-ab1 cells gated from splenocytes from CD1d2/2 mice).B, CD1d and NK1.1 expression on enriched CD41 splenocytes (gated TCR-ab1 cells) fromI-Ab2/2 mice. Percentages of each population are indicated in the quadrants. On the basis of six separate experiments performed in 8- to 14-wk-old classII-deficient mice, CD1dhigh NK1.12 T cells were found to represent around 15–25% of splenic CD41 T cells, i.e., a proportion 2-fold lower than that ofNK1.11 T cells (30–45%).
4919The Journal of Immunology
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
Immunoscope analysis of the Va14Ca rearrangement diversity was per-formed as described elsewhere (19, 31). Briefly, the indicated cDNAs weresubjected to a 40-cycle PCR using the Va14- and Ca-specific unlabeledprimers. The resulting PCR products were then used in run-off experimentsusing the Ca-specific nested fluorescent primer. The length of the CDR3region and the intensity of each band from run-off products were deter-mined on an automated sequencer (Perkin-Elmer, Norwalk, CT) and ana-lyzed using the immunoscope software.
Oligonucleotides and probes
For quantitative kinetic ELISA PCR, all primers and probes were obtainedfrom Genosys Biotechnologies (Cambridgeshire, U.K.). Thefollowingprimer specific sequences were used: biotinylated-Va14 primer, GGGAGATACTCAGCAACTCTGG; Ja281 primer, TCCCAGCTCCAAAATGCAGCC; biotinylated-39Ca, CTCGGTCAACGTGGCATCACA; and 59Ca,CCCTCTGCCTGTTCACCGACTT. The following fluoresceinated probespecific sequences were used for chemiluminescence detection of the ampli-fied products: Va14 probe, CAGCAGGGTGGCTGTGAT; Ca probe,GAGACCAACGCCACCTAC. For RT-PCR-based immunoscope technique,the following primers were used: Va14, CTAAGCACAGCACGCTGCACA;Ca, TGGCGTTGGTCTCTTTGAAG; and labeled Ca, FAM-ACACAGGAGGTTCTGGGTTC.
Cytokine assays by specific ELISA
IL-4 and IFN-g contents in supernatants were quantified using standardsandwich ELISAs (26) with a coated capture mAb (11B11 and R46A2mAbs for IL-4 and IFN-g assays, respectively) and a biotinylated detectionmAb (BVD6 and AN18 mAbs, for IL-4 and IFN-g assays, respectively).Conjugation of streptavidin-peroxidase (Amdex, Amersham, Les Ulis,France) was revealed usingo-phenylenediamine and hydrogen peroxide(Sigma). IFN-g and IL-4 concentrations are expressed in ng/ml, as calcu-lated from calibration curves from serial dilutions of mouse recombinantstandards (R&D Systems, Abingdon, U.K.) in each assay. The sensitivityof both IL-4 and IFN-g assays was 20–40 pg/ml.
Statistical analysis
Data were expressed as means6 SEM, and statistical significance wasevaluated by Student’st test. Values ofp , 0.05 were considered statis-tically significant.
ResultsThe splenic NK1.12 CD41 T cell compartment from I-Ab2/2
C57BL/6 mice contains cells that are responsive toa-GalCer
CD1d-restricted T cells that are responsible for rapid IL-4 produc-tion in vitro upon TCR engagement comprise NK1.11 andNK1.12 T cells (26). We examined the latter subset for its respon-siveness toa-GalCer. In 8- to 11-wk-old I-Ab2/2 C57BL/6 mice(further referred to as class II-deficient mice), the large majority(;65–70%) of the residual, and thus nonconventional, CD41 Tcells (4–5% of splenocytes) are of memory phenotype. These cellscomprise NK1.11 (35–45%) and NK1.12 (55–65%) subsets. Fig.1A compares cytokine production and proliferation of freshlysorted NK1.11 and NK1.12 CD44high CD41 T cells in response toa-GalCer. Because CD1d-expressing APCs are required fora-GalCer presentation to NK1.11 T cells (23, 24),g-irradiatedautologous spleen cell suspensions were added to each subset.Both NK1.12 and NK1.11 T cells responded toa-GalCer by astrong proliferation and cytokine production (Fig. 1A). Interest-ingly, the NK1.12 T subset was repeatedly found to be the morepotent IL-4 producer.
Nonconventional splenic NK1.12 CD41 T cells do not requireexogenous CD1d-expressing cells to respond toa-GalCer in aCD1d-dependent manner
Unexpectedly, the cytokine production and proliferation ofNK1.12 CD44high CD41 T cells from class II-deficient mice wasaffected neither by the absence of CD1d1/1 APCs nor by the ad-dition of CD1d2/2 APCs (Fig. 1B). Hence, their stimulation bya-GalCer does not depend on the presence of CD1d-expressingcells. To determine whether CD1d was involved, we conductedAb-blocking studies. IL-4 and IFN-g secretion as well as prolif-eration were abrogated by the simultaneous addition ofa-GalCerand anti-CD1d mAb to nonconventional NK1.12 T cells (Fig. 1C).
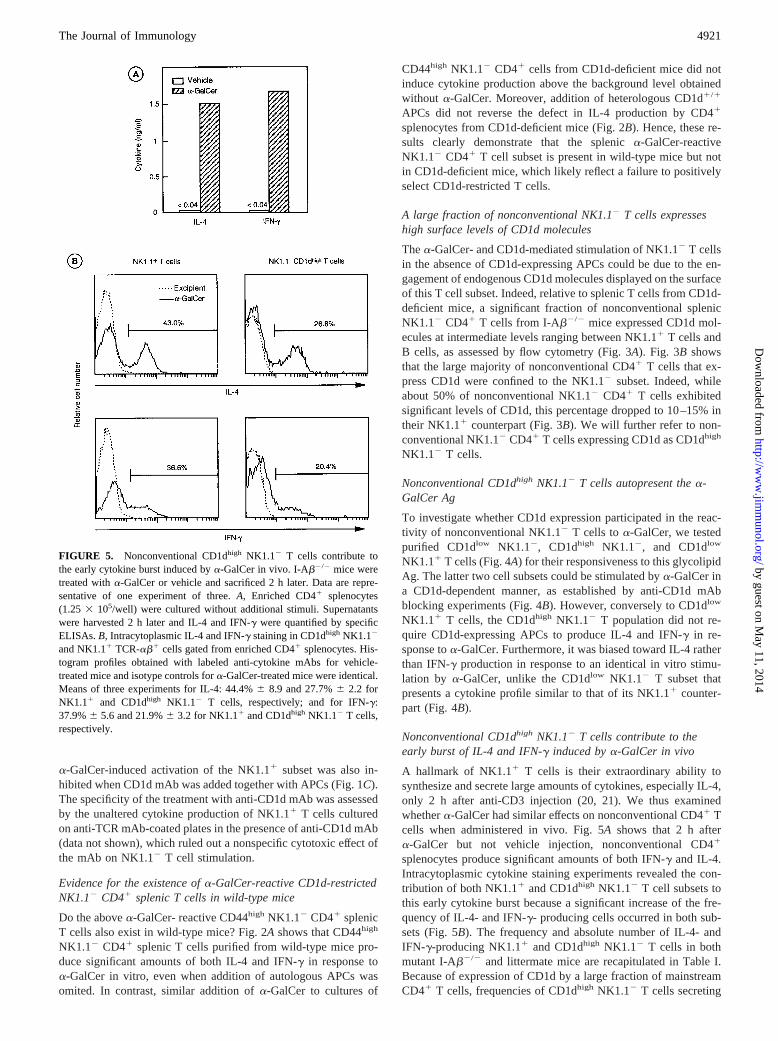
FIGURE 4. Nonconventional CD1dhigh
NK1.12 T cells but not NK1.11 T cellsproduce cytokines in response toa-GalCerin an APC-independent manner.A, Rela-tive CD1d expression on purified CD1dlow
NK1.11 CD41, CD1dhigh NK1.12 CD41,and CD1dlow NK1.12 CD41 splenic cellsfrom I-Ab2/2 mice. The degree of purityof sorted populations is indicated as per-centage in the respective dot plots (see in-sets).B, Purified splenic populations (23104 cells/well) were cultured witha-Gal-Cer with or without CD1d1/1 APCs. Anti-CD1d mAb was added to the cultures ofCD1dlow NK1.11 T cells in the presenceof CD1d1/1 APCs and of CD1dhigh
NK1.12 T cells in the absence of APCs.Supernatants were harvested after 60 h andassayed for IL-4 and IFN-g by specificELISAs. The data are expressed as means6SEM from four to six experiments. WhenSEM values are not indicated, data are thearithmetic means of triplicates from one typ-ical experiment of three. Differences be-tween CD1dlow NK1.11 and CD1dhigh
NK1.12 T cells are statistically significant(p , 0.03 andp , 0.02 for IL-4 and IFN-g,respectively).
4920 DISTINCT SUBSETS OF NKT CELLS RESPOND TOa-GalCer
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
a-GalCer-induced activation of the NK1.11 subset was also in-hibited when CD1d mAb was added together with APCs (Fig. 1C).The specificity of the treatment with anti-CD1d mAb was assessedby the unaltered cytokine production of NK1.11 T cells culturedon anti-TCR mAb-coated plates in the presence of anti-CD1d mAb(data not shown), which ruled out a nonspecific cytotoxic effect ofthe mAb on NK1.12 T cell stimulation.
Evidence for the existence ofa-GalCer-reactive CD1d-restrictedNK1.12 CD41 splenic T cells in wild-type mice
Do the abovea-GalCer- reactive CD44high NK1.12 CD41 splenicT cells also exist in wild-type mice? Fig. 2A shows that CD44high
NK1.12 CD41 splenic T cells purified from wild-type mice pro-duce significant amounts of both IL-4 and IFN-g in response toa-GalCer in vitro, even when addition of autologous APCs wasomited. In contrast, similar addition ofa-GalCer to cultures of
CD44high NK1.12 CD41 cells from CD1d-deficient mice did notinduce cytokine production above the background level obtainedwithout a-GalCer. Moreover, addition of heterologous CD1d1/1
APCs did not reverse the defect in IL-4 production by CD41
splenocytes from CD1d-deficient mice (Fig. 2B). Hence, these re-sults clearly demonstrate that the splenica-GalCer-reactiveNK1.12 CD41 T cell subset is present in wild-type mice but notin CD1d-deficient mice, which likely reflect a failure to positivelyselect CD1d-restricted T cells.
A large fraction of nonconventional NK1.12 T cells expresseshigh surface levels of CD1d molecules
Thea-GalCer- and CD1d-mediated stimulation of NK1.12 T cellsin the absence of CD1d-expressing APCs could be due to the en-gagement of endogenous CD1d molecules displayed on the surfaceof this T cell subset. Indeed, relative to splenic T cells from CD1d-deficient mice, a significant fraction of nonconventional splenicNK1.12 CD41 T cells from I-Ab2/2 mice expressed CD1d mol-ecules at intermediate levels ranging between NK1.11 T cells andB cells, as assessed by flow cytometry (Fig. 3A). Fig. 3Bshowsthat the large majority of nonconventional CD41 T cells that ex-press CD1d were confined to the NK1.12 subset. Indeed, whileabout 50% of nonconventional NK1.12 CD41 T cells exhibitedsignificant levels of CD1d, this percentage dropped to 10–15% intheir NK1.11 counterpart (Fig. 3B). We will further refer to non-conventional NK1.12 CD41 T cells expressing CD1d as CD1dhigh
NK1.12 T cells.
Nonconventional CD1dhigh NK1.12 T cells autopresent thea-GalCer Ag
To investigate whether CD1d expression participated in the reac-tivity of nonconventional NK1.12 T cells toa-GalCer, we testedpurified CD1dlow NK1.12, CD1dhigh NK1.12, and CD1dlow
NK1.11 T cells (Fig. 4A) for their responsiveness to this glycolipidAg. The latter two cell subsets could be stimulated bya-GalCer ina CD1d-dependent manner, as established by anti-CD1d mAbblocking experiments (Fig. 4B). However, conversely to CD1dlow
NK1.11 T cells, the CD1dhigh NK1.12 T population did not re-quire CD1d-expressing APCs to produce IL-4 and IFN-g in re-sponse toa-GalCer. Furthermore, it was biased toward IL-4 ratherthan IFN-g production in response to an identical in vitro stimu-lation by a-GalCer, unlike the CD1dlow NK1.12 T subset thatpresents a cytokine profile similar to that of its NK1.11 counter-part (Fig. 4B).
Nonconventional CD1dhigh NK1.12 T cells contribute to theearly burst of IL-4 and IFN-g induced bya-GalCer in vivo
A hallmark of NK1.11 T cells is their extraordinary ability tosynthesize and secrete large amounts of cytokines, especially IL-4,only 2 h after anti-CD3 injection (20, 21). We thus examinedwhethera-GalCer had similar effects on nonconventional CD41 Tcells when administered in vivo. Fig. 5Ashows that 2 h aftera-GalCer but not vehicle injection, nonconventional CD41
splenocytes produce significant amounts of both IFN-g and IL-4.Intracytoplasmic cytokine staining experiments revealed the con-tribution of both NK1.11 and CD1dhigh NK1.12 T cell subsets tothis early cytokine burst because a significant increase of the fre-quency of IL-4- and IFN-g- producing cells occurred in both sub-sets (Fig. 5B). The frequency and absolute number of IL-4- andIFN-g-producing NK1.11 and CD1dhigh NK1.12 T cells in bothmutant I-Ab2/2 and littermate mice are recapitulated in Table I.Because of expression of CD1d by a large fraction of mainstreamCD41 T cells, frequencies of CD1dhigh NK1.12 T cells secreting
FIGURE 5. Nonconventional CD1dhigh NK1.12 T cells contribute tothe early cytokine burst induced bya-GalCer in vivo. I-Ab2/2 mice weretreated witha-GalCer or vehicle and sacrificed 2 h later. Data are repre-sentative of one experiment of three.A, Enriched CD41 splenocytes(1.25 3 105/well) were cultured without additional stimuli. Supernatantswere harvested 2 h later and IL-4 and IFN-g were quantified by specificELISAs.B, Intracytoplasmic IL-4 and IFN-g staining in CD1dhigh NK1.12
and NK1.11 TCR-ab1 cells gated from enriched CD41 splenocytes. His-togram profiles obtained with labeled anti-cytokine mAbs for vehicle-treated mice and isotype controls fora-GalCer-treated mice were identical.Means of three experiments for IL-4: 44.4%6 8.9 and 27.7%6 2.2 forNK1.11 and CD1dhigh NK1.12 T cells, respectively; and for IFN-g:37.9%6 5.6 and 21.9%6 3.2 for NK1.11 and CD1dhigh NK1.12 T cells,respectively.
4921The Journal of Immunology
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
IL-4 and IFN-g were consistently reduced in wild-type mice ascompared with the ones found in I-Ab2/2 mice. Despite this, inwild-type as in I-Ab2/2 mice, the frequency of IL-4- and IFN-g-producing CD1dhigh NK1.12 T cells is of about one-half and one-third that of CD1dlow NK1.11 T cells, respectively, in response toa-GalCer. Because splenic NK1.11 T cells are present at the samefrequency in wild-type and I-Ab2/2 mice, it can be concluded thatin the two types of mice thea-GalCer-reactive subset within theCD1dhigh NK1.12 population is represented in equivalent numberof cells and contributes to the early burst of cytokines (Table I).
The phenotype of nonconventional CD1dhigh NK1.12 T cellsresembles that of NK1.11 T cells
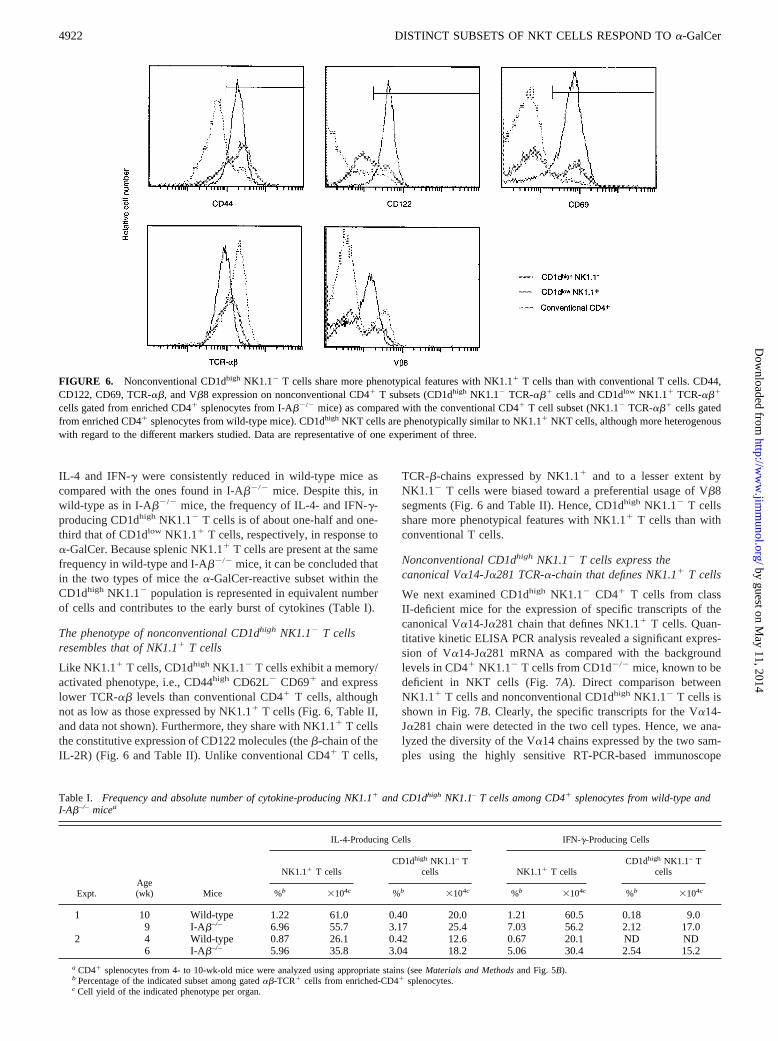
Like NK1.11 T cells, CD1dhigh NK1.12 T cells exhibit a memory/activated phenotype, i.e., CD44high CD62L2 CD691 and expresslower TCR-ab levels than conventional CD41 T cells, althoughnot as low as those expressed by NK1.11 T cells (Fig. 6, Table II,and data not shown). Furthermore, they share with NK1.11 T cellsthe constitutive expression of CD122 molecules (theb-chain of theIL-2R) (Fig. 6 and Table II). Unlike conventional CD41 T cells,
TCR-b-chains expressed by NK1.11 and to a lesser extent byNK1.12 T cells were biased toward a preferential usage of Vb8segments (Fig. 6 and Table II). Hence, CD1dhigh NK1.12 T cellsshare more phenotypical features with NK1.11 T cells than withconventional T cells.
Nonconventional CD1dhigh NK1.12 T cells express thecanonical Va14-Ja281 TCR-a-chain that defines NK1.11 T cells
We next examined CD1dhigh NK1.12 CD41 T cells from classII-deficient mice for the expression of specific transcripts of thecanonical Va14-Ja281 chain that defines NK1.11 T cells. Quan-titative kinetic ELISA PCR analysis revealed a significant expres-sion of Va14-Ja281 mRNA as compared with the backgroundlevels in CD41 NK1.12 T cells from CD1d2/2 mice, known to bedeficient in NKT cells (Fig. 7A). Direct comparison betweenNK1.11 T cells and nonconventional CD1dhigh NK1.12 T cells isshown in Fig. 7B. Clearly, the specific transcripts for the Va14-Ja281 chain were detected in the two cell types. Hence, we ana-lyzed the diversity of the Va14 chains expressed by the two sam-ples using the highly sensitive RT-PCR-based immunoscope
FIGURE 6. Nonconventional CD1dhigh NK1.12 T cells share more phenotypical features with NK1.11 T cells than with conventional T cells. CD44,CD122, CD69, TCR-ab, and Vb8 expression on nonconventional CD41 T subsets (CD1dhigh NK1.12 TCR-ab1 cells and CD1dlow NK1.11 TCR-ab1
cells gated from enriched CD41 splenocytes from I-Ab2/2 mice) as compared with the conventional CD41 T cell subset (NK1.12 TCR-ab1 cells gatedfrom enriched CD41 splenocytes from wild-type mice). CD1dhigh NKT cells are phenotypically similar to NK1.11 NKT cells, although more heterogenouswith regard to the different markers studied. Data are representative of one experiment of three.
Table I. Frequency and absolute number of cytokine-producing NK1.11 and CD1dhigh NK1.1– T cells among CD41 splenocytes from wild-type andI-Ab–/– micea
Expt.Age(wk) Mice
IL-4-Producing Cells IFN-g-Producing Cells
NK1.11 T cellsCD1dhigh NK1.1– T
cells NK1.11 T cellsCD1dhigh NK1.1– T
cells
%b 3104c %b 3104c %b 3104c %b 3104c
1 10 Wild-type 1.22 61.0 0.40 20.0 1.21 60.5 0.18 9.09 I-Ab–/– 6.96 55.7 3.17 25.4 7.03 56.2 2.12 17.0
2 4 Wild-type 0.87 26.1 0.42 12.6 0.67 20.1 ND ND6 I-Ab–/– 5.96 35.8 3.04 18.2 5.06 30.4 2.54 15.2
a CD41 splenocytes from 4- to 10-wk-old mice were analyzed using appropriate stains (seeMaterials and Methodsand Fig. 5B).b Percentage of the indicated subset among gatedab-TCR1 cells from enriched-CD41 splenocytes.c Cell yield of the indicated phenotype per organ.
4922 DISTINCT SUBSETS OF NKT CELLS RESPOND TOa-GalCer
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
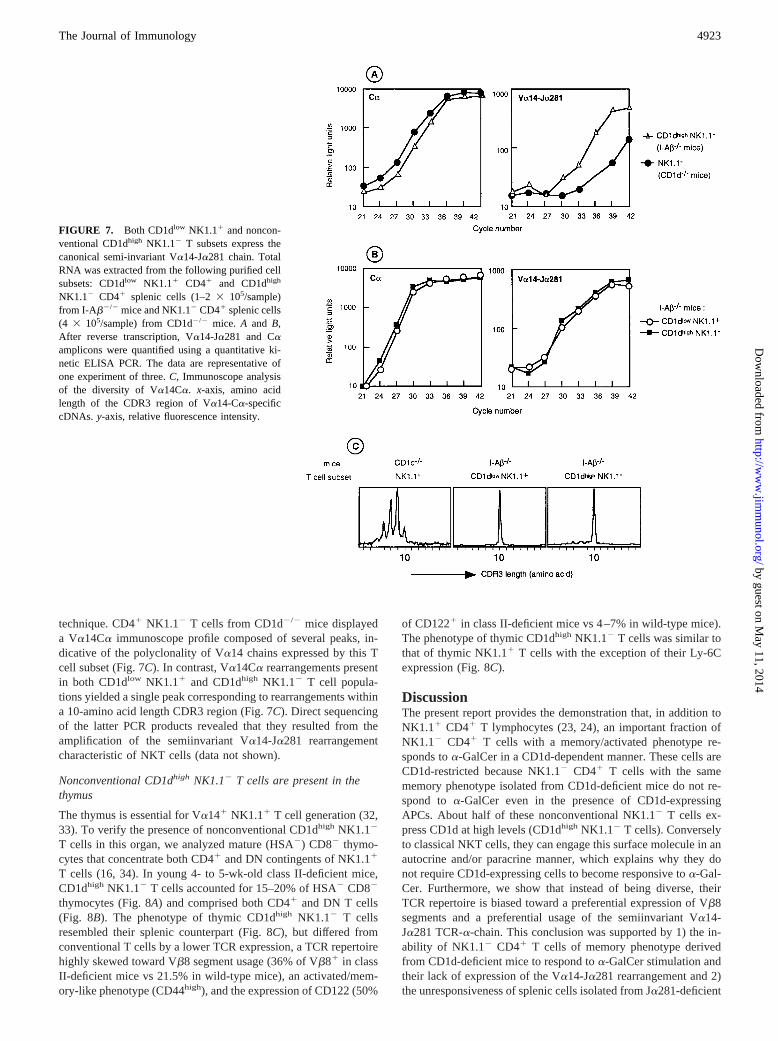
technique. CD41 NK1.12 T cells from CD1d2/2 mice displayeda Va14Ca immunoscope profile composed of several peaks, in-dicative of the polyclonality of Va14 chains expressed by this Tcell subset (Fig. 7C). In contrast, Va14Ca rearrangements presentin both CD1dlow NK1.11 and CD1dhigh NK1.12 T cell popula-tions yielded a single peak corresponding to rearrangements withina 10-amino acid length CDR3 region (Fig. 7C). Direct sequencingof the latter PCR products revealed that they resulted from theamplification of the semiinvariant Va14-Ja281 rearrangementcharacteristic of NKT cells (data not shown).
Nonconventional CD1dhigh NK1.12 T cells are present in thethymus
The thymus is essential for Va141 NK1.11 T cell generation (32,33). To verify the presence of nonconventional CD1dhigh NK1.12
T cells in this organ, we analyzed mature (HSA2) CD82 thymo-cytes that concentrate both CD41 and DN contingents of NK1.11
T cells (16, 34). In young 4- to 5-wk-old class II-deficient mice,CD1dhigh NK1.12 T cells accounted for 15–20% of HSA2 CD82
thymocytes (Fig. 8A) and comprised both CD41 and DN T cells(Fig. 8B). The phenotype of thymic CD1dhigh NK1.12 T cellsresembled their splenic counterpart (Fig. 8C), but differed fromconventional T cells by a lower TCR expression, a TCR repertoirehighly skewed toward Vb8 segment usage (36% of Vb81 in classII-deficient mice vs 21.5% in wild-type mice), an activated/mem-ory-like phenotype (CD44high), and the expression of CD122 (50%
of CD1221 in class II-deficient mice vs 4–7% in wild-type mice).The phenotype of thymic CD1dhigh NK1.12 T cells was similar tothat of thymic NK1.11 T cells with the exception of their Ly-6Cexpression (Fig. 8C).
DiscussionThe present report provides the demonstration that, in addition toNK1.11 CD41 T lymphocytes (23, 24), an important fraction ofNK1.12 CD41 T cells with a memory/activated phenotype re-sponds toa-GalCer in a CD1d-dependent manner. These cells areCD1d-restricted because NK1.12 CD41 T cells with the samememory phenotype isolated from CD1d-deficient mice do not re-spond to a-GalCer even in the presence of CD1d-expressingAPCs. About half of these nonconventional NK1.12 T cells ex-press CD1d at high levels (CD1dhigh NK1.12 T cells). Converselyto classical NKT cells, they can engage this surface molecule in anautocrine and/or paracrine manner, which explains why they donot require CD1d-expressing cells to become responsive toa-Gal-Cer. Furthermore, we show that instead of being diverse, theirTCR repertoire is biased toward a preferential expression of Vb8segments and a preferential usage of the semiinvariant Va14-Ja281 TCR-a-chain. This conclusion was supported by 1) the in-ability of NK1.12 CD41 T cells of memory phenotype derivedfrom CD1d-deficient mice to respond toa-GalCer stimulation andtheir lack of expression of the Va14-Ja281 rearrangement and 2)the unresponsiveness of splenic cells isolated from Ja281-deficient
FIGURE 7. Both CD1dlow NK1.11 and noncon-ventional CD1dhigh NK1.12 T subsets express thecanonical semi-invariant Va14-Ja281 chain. TotalRNA was extracted from the following purified cellsubsets: CD1dlow NK1.11 CD41 and CD1dhigh
NK1.12 CD41 splenic cells (1–23 105/sample)from I-Ab2/2 mice and NK1.12 CD41 splenic cells(4 3 105/sample) from CD1d2/2 mice. A and B,After reverse transcription, Va14-Ja281 and Caamplicons were quantified using a quantitative ki-netic ELISA PCR. The data are representative ofone experiment of three.C, Immunoscope analysisof the diversity of Va14Ca. x-axis, amino acidlength of the CDR3 region of Va14-Ca-specificcDNAs. y-axis, relative fluorescence intensity.
4923The Journal of Immunology
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

mice toa-GalCer (23). Thus, NKT cells, as defined by their CD1drestriction and Va14-Ja281 TCR-a-chain expression, can be di-vided into NK1.11 and CD1dhigh NK1.12 subsets. For the sake ofclarity, we will further refer to these subsets as NK1.11 NKT andCD1dhigh NKT cells, respectively.
The present study was performed with CD1dhigh NKT cells pu-rified from class II-deficient mice. Characterization of this NKTsubset in wild-type mice was complicated by the expression ofCD1d molecules by some conventional CD41 T cells (data notshown) and by their low frequency among CD41 NK1.12 T cells.Nevertheless,a-GalCer-reactive CD1dhigh NKT cells do also existin wild-type mice because their splenic CD44high NK1.12 CD41
T cells produce significant amounts of both IL-4 and IFN-g inresponse toa-GalCer, even in the absence of CD1d-expressingcells. In accordance with these data, CD41 T splenocytes fromwild-type mice respond toa-GalCer in the absence of exogenousAPCs. Moreover, Burdin et al. (24) have recently identified aVa14-expressing CD1d-autoreactive NKT cell hybridoma derivedfrom wild-type mice that expresses CD1d at high levels and is
capable of autopresentinga-GalCer. Finally, intracytoplasmic cy-tokine staining performed following a short-term treatment of miceby a-GalCer allowed us to determine that the ratio ofa-GalCer-responding CD1dhigh NKT cells to a-GalCer-responding NK1.11
NKT cells is identical in wild-type mice and I-Ab2/2 mice.The unique functional properties of CD1dhigh NKT cells dem-
onstrated herein raise the question of the physiological relevanceof either NKT subset in various immune responses, including thosepreviously ascribed to NK1.11 NKT cells. Because the activationof CD1dhigh NKT cells bya-GalCer in vitro occurs in the absenceof APCs it is likely that they are also autonomous in vivo. If so,their independence from costimulatory signals delivered by APCswould endow them with the advantage of reacting to glycolipidAgs more rapidly and more efficiently than their NK1.11 counter-part. Surprisingly and in accordance with this hypothesis,CD1dhigh NKT cells, but not NK1.11 NKT cells, were found toexpress CD40 and CD86, two costimulatory molecules usuallyexpressed by APCs (A. Herbelin and A. Hameg, unpublishedobservations).
FIGURE 8. Nonconventional CD41 and DNCD1dhigh NK1.12 T cells are found in the thymusand share similar phenotypical features withNK1.11 thymocytes. Freshly isolated HSA- andCD8-depleted thymocytes were from mutantI-Ab2/2 and wild-type mice.A, Distribution ofCD1d and NK1.1 surface markers on gated TCR-ab1 thymocytes from I-Ab2/2 mice. B, Distri-bution of CD41 and DN cells among gated TCR-ab1 CD1dhigh NK1.12 and TCR-ab1 CD1dlow
NK1.11 thymocytes from I-Ab2/2 mice. C,CD44, CD122, Ly-6C, TCR-ab, and Vb8 expres-sion on nonconventional cell subsets (gatedCD1dhighNK1.12 TCR-ab1 and CD1dlow NK1.11
TCR-ab1 thymocytes from I-Ab2/2 mice) and onconventional CD41 T cells (gated NK1.12 TCR-ab1 CD41 thymocytes from wild-type mice). Datarepresent a typical experiment of two.
Table II. Frequency (%) of cells expressing Vb8, CD122, and memory activation markers among CD1dhigh NK1.12 T cellsa
CD41 T Cells Vb8 CD69 CD122 CD44
CD1dlow NK1.11 60.46 0.9 65.26 12.9 94.26 1.4 90.66 3.5CD1dhigh NK1.1– 36.36 0.5 29.66 1.5 36.86 2.8 78.96 3.7Conventional 19.56 0.4 6.46 1.6 9.06 3.0 16.36 2.6
a CD41 TCR-ab1 cells gated from CD41-enriched splenocytes from C57BL/6 wild-type and I-Ab–/– mice were stained as described inMaterials and Methodsand analyzedfor the expression of CD44, CD122, CD69, and Vb8 according to CD1d and NK1.1 expression. Percentages were calculated as the frequency of positive cells in the gaterepresented in Fig. 5. Conventional CD41 T cells correspond to CD41 NK1.1– TCR-ab1 splenocytes of wild-type mice. The data are arithmetic means6 SEM of threeexperiments.
4924 DISTINCT SUBSETS OF NKT CELLS RESPOND TOa-GalCer
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Althougha-GalCer is a synthetic glycosylceramide, it might behypothesized that it mimics endogenous Ag(s) expressed on nor-mal/activated, stressed, apoptotic, or tumorized cells (25).CD1dhigh NKT cells might thus be envisioned as an early sentinelto survey at any moment the homeostasis or integrity of cells ex-pressing such endogenous Ags. Through the expression of CD1d,they could also exert other putative functions, such as Ag presen-tation to other CD1d-restricted T cells, whether they belong to theNKT subset or not, thus rendering them capable of participating intheir thymic and/or peripheral selection/education process. Be-cause NK1.11 NKT cells can recognize exogenous glycolipids,such as phosphatidylinositolmannosides derived from mycobacte-rial cell walls (19) and glycosylphosphatidylinositols isolated fromthe membrane of parasites (35), CD1dhigh NKT cells might be ableto do the same, thus counteracting systemic infections by respond-ing to soluble parasite or mycobacterial derivatives in an APC-independent manner. Do NK1.11 and CD1dhigh NKT cells possessdifferent Ag specificities, thus enabling them to exert a large spec-trum of recognition? As far as putative endogenous Ags are con-cerned, it would be informative to test whether CD1dhigh NKTcells, like NK1.11 NKT cells (6, 24, 25) display tissue-specificity.Future identification of natural ligand(s) for these two NKT celltypes in mammals will provide further insights into these issues.
Recent studies have suggested thata-GalCer might emerge as auseful tool to manipulate the balance of adaptive immune re-sponses by directing conventional T cells to a Th2 differentiationpathway (36, 37). It would be of interest to determine the respec-tive contribution of NK1.11 and CD1dhigh NKT cell subsets in thepolarization of the immune response. In vivo, we show that bothsubsets contribute to the burst of IL-4, whereas a clear dichotomybetween the two populations in terms of cytokine production oc-curs in vitro. Indeed, NK1.11 NKT cells produce IL-4 and IFN-gat equivalent amounts whereas CD1dhigh NKT cells generate muchmore IL-4 than IFN-g. After a-GalCer administration, the ratio ofIL-4-producing cells to IFN-g-producing cells was for CD1dhigh
NKT cells and NK1.11 NKT cells of 1.536 0.12 (mean6 SEM,n 5 4) and 1.166 0.07 (mean6 SEM, n 5 4) in I-Ab2/2 mice( p , 0.006), respectively, and of 1.716 0.01 (mean6 SEM,n 52) and 1.166 0.14 (mean6 SEM, n 5 2) in wild-type mice,respectively. Taken together, it suggests that NK1.11 NKT cellsbehave like Th0 cells, whereas CD1dhigh NKT cells behave likeTh2-cells. The identification of glycolipid Ags stimulatingCD1dhigh NKT cells but not NK1.11 NKT cells would be usefulfor studies of autoimmune diseases for which a shift toward a Th1profile and a deficit in NKT cells have been evidenced (15, 16).
Another important issue of this work concerns the possible lin-eage relationship betweena-GalCer-reactive NK1.11 and NK1.12
T cells. Knowing that NKT cells can upon in vitro activationdown-regulate the NK1.1 molecule (38) and up-regulate CD1d ina transitory manner (A. Herbelin and A. Hameg, unpublished ob-servations), it could be argued that CD1dhigh NKT cells correspondto activated splenic NK1.11 NKT cells. In naive mice, a low butsignificant fraction (10–15%) of the NK1.11 pool of cells expresshigh levels of CD1d. Because of the increased size of the lattercells, one can assume that they are blastic at a high frequency(51.3 6 4.7%, n 5 4), which contrasts with the 2-fold reducedfrequency of blastic cells among the CD1dhigh NKT cells (24.265.6%;n 5 4). If the CD1dhigh NKT cells were exclusively arisingfrom the NK1.11 NKT cells, one would expect to find about 50%of blastic cells within the CD1dhigh NKT cell compartment. Thus,although we cannot exclude that a fraction of CD1dhigh NKT cellsderives from NK1.11 NKT cells having up-regulated CD1d anddown-regulated NK1.1 molecules, our data support the idea that alarge majority of CD1dhigh NKT cells express constitutively
CD1d. The presence of CD1dhigh NKT cells in thymus and theiridentical activated/memory phenotype in thymus and spleen areconsistent with the notion that their phenotype is not exclusivelydue to activation and/or education processes in secondary lym-phoid organs. These latter observations along with the CD1d re-striction of both populations are in accordance with a shared se-lection pathway whose subtle differences might explain the distinctphenotype. It might indeed be hypothesized that double positive(DP) thymocytes, which express CD1d, lose their expression ac-cording to their TCR specificity after interaction with differentendogenous Ags. An alternative possibility is that CD1dlow andCD1dhigh populations diverge at early stage of differentiation andcould exist at DP stages before positive selection occurs.
In conclusion, this work provides evidence for a novel subset ofCD1d-restricted anda-GalCer-reactive NKT cells and reveals anadditional level of complexity within the NKT cell population.Interestingly, two very recent reports (which were published dur-ing the preparation of this manuscript) demonstrated the existenceof a-GalCer-reactive NK1.12 NKT cells (39, 40). The respectiveroles of each NKT subset in the regulation of homeostasis andpathophysiological responses need to be further investigated.
AcknowledgmentsWe thank C. Gouarin for her excellent technical assistance. We thankDr. L. Van Kaer (Howard Hughes Medical Institut, Vanderbilt UniversitySchool of Medicine, Nashville, TN) for providing mutant CD1d2/2 mice.We thank Dr. A. Bendelac (Department of Molecular Biology, PrincetonUniversity, Princeton, NJ) for generously providing anti-CD1d mAb. Wealso thank Drs. Michel Dy and Elke Schneider for critical review of themanuscript.
References1. Porcelli, S. A., and R. L. Modlin. 1999. The CD1 system: antigen-presenting
molecules for T cell recognition of lipids and glycolipids.Annu. Rev. Immunol.17:297.
2. Bendelac, A., M. N. Rivera, S. H. Park, and J. H. Roark. 1997. Mouse CD1-specific NK11 T cells: development, specificity, and function.Annu. Rev. Im-munol. 15:535.
3. Maher, J. K., and M. Kronenberg. 1997. The role of CD1 molecules in immuneresponses to infection.Curr. Opin. Immunol. 9:456.
4. Cardell, S., S. Tangri, S. Chan, M. Kronenberg, C. Benoist, and D. Mathis. 1995.CD1-restricted CD41 T cells in major histocompatibility complex class II-defi-cient mice.J. Exp. Med. 182:993.
5. Brossay, L., S. Tangri, M. Bix, S. Cardell, R. Locksley, and M. Kronenberg.1998. Mouse CD1-autoreactive T cells have diverse patterns of reactivity toCD11 targets.J. Immunol. 160:3681.
6. Chiu, Y. H., J. Jayawardena, A. Weiss, D. Lee, S. H. Park, A. Dautry-Varsat, andA. Bendelac. 1999. Distinct subsets of CD1d-restricted T cells recognize self-antigens loaded in different cellular compartments.J. Exp. Med. 189:103.
7. Bendelac, A., O. Lantz, M. E. Quimby, J. W. Yewdell, J. R. Bennink, andR. R. Brutkiewicz. 1995. CD1 recognition by mouse NK11 T lymphocytes.Sci-ence 268:863.
8. Leite-de-Moraes, M. C., and M. Dy. 1997. Natural killer T cells: a potent cyto-kine-producing cell population.Eur. Cytokine Network 8:229.
9. Dellabona, P., E. Padovan, G. Casorati, M. Brockhaus, and A. Lanzavecchia.1994. An invariant V 24-J Q/V 11 T cell receptor is expressed in all individualsby clonally expanded CD4–8- T cells.J. Exp. Med. 180:1171.
10. Exley, M., J. Garcia, S. P. Balk, and S. Porcelli. 1997. Requirements for CD1drecognition by human invariant Va1 CD42CD82 T cells.J. Exp. Med. 186:109.
11. Takeda, K., S. Seki, K. Ogasawara, R. Anzai, W. Hashimoto, K. Sugiura,M. Takahashi, M. Satoh, and K. Kumagai. 1996. Liver NK1.11 CD41 T cellsactivated by IL-12 as a major effector in inhibition of experimental tumor me-tastasis.J. Immunol. 156:3366.
12. Cui, J., T. Shin, T. Kawano, H. Sato, E. Kondo, I. Toura, Y. Kaneko, H. Koseki,M. Kanno, and M. Taniguchi. 1997. Requirement for Va2 14 NKT cells inIL-12-mediated rejection of tumors.Science 278:1623.
13. Smyth, M. J., K. Y. Thia, S. E. Street, E. Cretney, J. A. Trapani, M. Taniguchi,T. Kawano, S. B. Pelikan, N. Y. Crowe, and D. I. Godfrey. 2000. Differentialtumor surveillance by natural killer (NK) and NKT cells.J. Exp. Med. 191:661.
14. Mieza, M. A., T. Itoh, J. Q. Cui, Y. Makino, T. Kawano, K. Tsuchida, T. Koike,T. Shirai, H. Yagita, A. Matsuzawa, H. Koseki, and M. Taniguchi. 1996. Selec-tive reduction of V 141 NK T cells associated with disease development inautoimmune-prone mice.J. Immunol. 156:4035.
15. Wilson, S. B., S. C. Kent, K. T. Patton, T. Orban, R. A. Jackson, M. Exley,S. Porcelli, D. A. Schatz, M. A. Atkinson, S. P. Balk, J. L. Strominger, andD. A. Hafler. 1998. Extreme Th1 bias of invariant V 24J Q T cells in type 1diabetes.Nature 391:177.
4925The Journal of Immunology
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

16. Gombert, J. M., A. Herbelin, E. Tancrede-Bohin, M. Dy, C. Carnaud, andJ. F. Bach. 1996. Early quantitative and functional deficiency of NK11-like thy-mocytes in the NOD mouse.Eur. J. Immunol. 26:2989.
17. Denkers, E. Y., T. Scharton-Kersten, S. Barbieri, P. Caspar, and A. Sher. 1996.A role for CD41 NK1.11 T lymphocytes as major histocompatibility complexclass II independent helper cells in the generation of CD81 effector functionagainst intracellular infection.J. Exp. Med. 184:131.
18. Flesch, I. E., A. Wandersee, and S. H. Kaufmann. 1997. IL-4 secretion by CD41
NK11 T cells induces monocyte chemoattractant protein-1 in early listeriosis.J. Immunol. 159:7.
19. Apostolou, I., Y. Takahama, C. Belmant, T. Kawano, M. Huerre, G. Marchal,J. Cui, M. Taniguchi, H. Nakauchi, J. J. Fournie, P. Kourilsky, and G. Gachelin.1999. Murine natural killer T(NKT) cells [correction of natural killer cells] con-tribute to the granulomatous reaction caused by mycobacterial cell walls [pub-lished erratum appears in1999 Proc. Natl. Acad. Sci. USA 96:7610.]Proc. Natl.Acad. Sci. USA 96:5141.
20. Yoshimoto, T., and W. E. Paul. 1994. CD4pos, NK1.1pos T cells promptly pro-duce interleukin 4 in response to in vivo challenge with anti-CD3.J. Exp. Med.179:1285.
21. Leite-de-Moraes, M., A. Herbelin, F. Machavoine, A. Vicari, J. M. Gombert,M. Papiernik, and M. Dy. 1995. MHC class I-selected CD42CD82TCR-ab1 Tcells are a potential source of IL-4 during primary immune response.J. Immunol.155:4544.
22. Morita, M., K. Motoki, K. Akimoto, T. Natori, T. Sakai, E. Sawa, K. Yamaji,Y. Koezuka, E. Kobayashi, and H. Fukushima. 1995. Structure-activity relation-ship ofa-galactosylceramides against B16-bearing mice.J. Med. Chem. 38:2176.
23. Kawano, T., J. Cui, Y. Koezuka, I. Toura, Y. Kaneko, K. Motoki, H. Ueno,R. Nakagawa, H. Sato, E. Kondo, H. Koseki, and M. Taniguchi. 1997. CD1d-restricted and TCR-mediated activation ofa NKT cells by glycosylceramides.Science 278:1626.
24. Burdin, N., L. Brossay, Y. Koezuka, S. T. Smiley, M. J. Grusby, M. Gui,M. Taniguchi, K. Hayakawa, and M. Kronenberg. 1998. Selective ability ofmouse CD1 to present glycolipids:a-galactosylceramide specifically stimulatesVa141 NK T lymphocytes.J. Immunol. 161:3271.
25. Gumperz, J. E., C. Roy, A. Makowska, D. Lum, M. Sugita, T. Podrebarac,Y. Koezuka, S. A. Porcelli, S. Cardell, M. B. Brenner, and S. M. Behar. 2000.Murine CD1d-restricted T cell recognition of cellular lipids.Immunity 12:211.
26. Hameg, A., C. Gouarin, J. M. Gombert, S. Hong, L. Van Kaer, J. F. Bach, andA. Herbelin. 1999. IL-7 up-regulates IL-4 production by splenic NK1.11 andNK1.1- MHC class I-like/CD1-dependent CD41 T cells.J. Immunol. 162:7067.
27. Chen, H., and W. E. Paul. 1998. A population of CD62Llow Nk1.12 CD41 Tcells that resembles NK1.11 CD41 T cells.Eur. J. Immunol. 28:3172.
28. Skold, M., A. Rytter, F. Ivars, and S. Cardell. 1999. Characterization of sub-populations of T-cell receptor intermediate (TCRint) T cells.Scand. J. Immunol.49:611.
29. Mendiratta, S. K., W. D. Martin, S. Hong, A. Boesteanu, S. Joyce, andL. Van Kaer. 1997. CD1d1 mutant mice are deficient in natural T cells thatpromptly produce IL-4.Immunity 6:469.
30. Taoufik, Y., D. Froger, S. Benoliel, C. Wallon, E. Dussaix, J. F. Delfraissy, andO. Lantz. 1998. Quantitative ELISA-polymerase chain reaction at saturation us-ing homologous internal DNA standards and chemiluminescence revelation.Eur.Cytokine Network 9:197.
31. Pannetier, C., J.-P.Levraud, A.Lim, J.Even, and P. Kourilsky. 1997. InThe T-CellReceptor: Selected Protocols and Applications. J. R. Oksenznberg, L. Wang, andJ.-Y.Y. Jeffery, eds. Chapman and Hall, New York, p. 287.
32. Tilloy, F., J. P. Di Santo, A. Bendelac, and O. Lantz. 1999. Thymic dependenceof invariant Va 141 natural killer-T cell development.Eur. J. Immunol. 29:3313.
33. Hammond, K., W. Cain, I. van Driel, and D. Godfrey. 1998. Three day neonatalthymectomy selectively depletes NK1.11 T cells. Int. Immunol. 10:1491.
34. Bendelac, A., N. Killeen, D. R. Littman, and R. H. Schwartz. 1994. A subset ofCD41 thymocytes selected by MHC class I molecules.Science 263:1774.
35. Schofield, L., M. J. McConville, D. Hansen, A. S. Campbell, B. Fraser-Reid,M. J. Grusby, and S. D. Tachado. 1999. CD1d-restricted immunoglobulin Gformation to GPI-anchored antigens mediated by NKT cells.Science 283:225.
36. Burdin, N., L. Brossay, and M. Kronenberg. 1999. Immunization witha-galac-tosylceramide polarizes CD1-reactive NK T cells towards Th2 cytokine synthe-sis.Eur. J. Immunol. 29:2014.
37. Singh, N., S. Hong, D. C. Scherer, I. Serizawa, N. Burdin, M. Kronenberg,Y. Koezuka, and L. Van Kaer. 1999. Cutting edge: activation of NK T cells byCD1d and -galactosylceramide directs conventional T cells to the acquisition ofa Th2 phenotype.J. Immunol. 163:2373.
38. Chen, H., H. Huang, and W. E. Paul. 1997. NK1.11 CD41 T cells lose NK1.1expression upon in vitro activation.J. Immunol. 158:5112.
39. Benlagha, K., A. Weiss, A. Beavis, L. Teyton and A. Bendelac. 2000. In vivoidentification of glycolipid antigen-specific T cells using fluorescent CD1d tet-ramers.J. Exp. Med. 191:1895.
40. Matsuda, J. L., O. V. Naidenko, L. Gapin, T. Nakayama, M. Taniguchi, C.-R.Wang, Y. Koezuka, and M. Kronenberg. 2000. Tracking the response of naturalkiller T cells to a glycolipid antigen using cold tetramers.J. Exp. Med 192:741.
4926 DISTINCT SUBSETS OF NKT CELLS RESPOND TOa-GalCer
by guest on May 11, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from














