
A Role for the GCC-Box in Jasmonate-Mediated Activation of the PDF1.2 Gene of Arabidopsis1
13
A Role for the GCC-Box in Jasmonate-Mediated Activation of the PDF1.2 Gene of Arabidopsis 1 Rebecca L. Brown, Kemal Kazan*, Ken C. McGrath, Don J. Maclean, and John M. Manners Cooperative Research Centre for Tropical Plant Protection (R.L.B., K.K., K.C.M., D.J.M., J.M.M.), and Department of Biochemistry and Molecular Biology (R.L.B., K.M., D.J.M.), The University of Queensland, Brisbane, Queensland 4072, Australia; and Commonwealth Scientific and Industrial Research Organisation, Plant Industry, Queensland Bioscience Precinct, St. Lucia, Brisbane, Queensland 4067, Australia (K.K., J.M.M.) The PDF1.2 gene of Arabidopsis encoding a plant defensin is commonly used as a marker for characterization of the jasmonate-dependent defense responses. Here, using PDF1.2 promoter-deletion lines linked to the -glucoronidase-reporter gene, we examined putative promoter elements associated with jasmonate-responsive expression of this gene. Using stably transformed plants, we first characterized the extended promoter region that positively regulates basal expression from the PDF1.2 promoter. Second, using promoter deletion constructs including one from which the GCC-box region was deleted, we observed a substantially lower response to jasmonate than lines carrying this motif. In addition, point mutations introduced into the core GCC-box sequence substantially reduced jasmonate responsiveness, whereas addition of a 20-nucleotide-long promoter element carrying the core GCC-box and flanking nucleotides provided jasmonate responsive- ness to a 35S minimal promoter. Taken together, these results indicated that the GCC-box plays a key role in conferring jasmonate responsiveness to the PDF1.2 promoter. However, deletion or specific mutations introduced into the core GCC-box did not completely abolish the jasmonate responsiveness of the promoter, suggesting that the other promoter elements lying downstream from the GCC-box region may also contribute to jasmonate responsiveness. In other experi- ments, we identified a jasmonate- and pathogen-responsive ethylene response factor transcription factor, AtERF2, which when overexpressed in transgenic Arabidopsis plants activated transcription from the PDF1.2, Thi2.1, and PR4 (basic chitinase) genes, all of which contain a GCC-box sequence in their promoters. Our results suggest that in addition to their roles in regulating ethylene-mediated gene expression, ethylene response factors also appear to play important roles in regulating jasmonate-responsive gene expression, possibly via interaction with the GCC-box. Diseases caused by fungal, bacterial, and viral pathogens result in enormous reductions in yield and quality from agricultural crops around the world. However, although plants are continuously chal- lenged by pathogenic microorganisms, the develop- ment of disease is a relatively rare event. It is clear, therefore, that plants possess efficient defense sys- tems. Effective induction of the defense response re- quires pathogen recognition followed by a network of signal transduction processes, resulting in the rapid activation of defense gene expression. A num- ber of secondary signal molecules such as salicylic acid (SA), jasmonic acid (JA) and its methyl ester, methyl jasmonate (MeJA), and ethylene act to am- plify and regulate defense responses after initial ac- tivation (Dong, 1998; Reymond and Farmer, 1998; Schenk et al., 2000). Interestingly, the SA- and jasmonate/ethylene-dependent signaling pathways appear to modulate plant defense responses against different classes of pathogens (for review, see Thomma et al., 2001). Treatment with MeJA provides Arabidopsis plants with protection against subse- quent inoculation with a range of necrotrophic fungi, but not against the biotrophic pathogen Peronospora parasitica (Thomma et al., 1998). Similarly, several studies of Arabidopsis mutants including ein2, coi1, jar1, and fad3/7/8, in which the jasmonate/ethylene- response is disrupted, have indicated that resistance to a variety of necrotrophic pathogens requires a functional jasmonate/ethylene-signaling pathway (Staswick et al., 1998; Vijayan et al., 1998; Thomma et al., 1999). In contrast, resistance to the biotroph P. parasitica but not to the necrotroph Alternaria brassici- cola is enhanced by pretreatment with isonicotinic acid, a functional analog of SA (Thomma et al., 1998). A number of classes of regulatory proteins and transcription factors are known to play important roles in relaying the pathogen-initiated signals to downstream components for the activation of plant defense responses. Among these, ethylene response factor domain-containing transcription factors (ERFs) are implicated as key regulators of plant defense responses. For example, the tomato (Lycopersicon es- culentum) gene Pti4 encodes an ERF protein that is induced by defense-signaling molecules and by the bacterial pathogen Pseudomonas syringae pv. tomato 1 This work was supported by the Grains Research and Devel- opment Corporation (postgraduate fellowship to R.L.B.). * Corresponding author; e-mail [email protected]; fax 61– 7–3214 –2950. Article, publication date, and citation information can be found at www.plantphysiol.org/cgi/doi/10.1104/pp.102.017814. 1020 Plant Physiology, June 2003, Vol. 132, pp. 1020–1032, www.plantphysiol.org © 2003 American Society of Plant Biologists www.plant.org on August 15, 2015 - Published by www.plantphysiol.org Downloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of A Role for the GCC-Box in Jasmonate-Mediated Activation of the PDF1.2 Gene of Arabidopsis1

A Role for the GCC-Box in Jasmonate-MediatedActivation of the PDF1.2 Gene of Arabidopsis1
Rebecca L. Brown, Kemal Kazan*, Ken C. McGrath, Don J. Maclean, and John M. Manners
Cooperative Research Centre for Tropical Plant Protection (R.L.B., K.K., K.C.M., D.J.M., J.M.M.), andDepartment of Biochemistry and Molecular Biology (R.L.B., K.M., D.J.M.), The University of Queensland,Brisbane, Queensland 4072, Australia; and Commonwealth Scientific and Industrial Research Organisation,Plant Industry, Queensland Bioscience Precinct, St. Lucia, Brisbane, Queensland 4067, Australia (K.K.,J.M.M.)
The PDF1.2 gene of Arabidopsis encoding a plant defensin is commonly used as a marker for characterization of thejasmonate-dependent defense responses. Here, using PDF1.2 promoter-deletion lines linked to the �-glucoronidase-reportergene, we examined putative promoter elements associated with jasmonate-responsive expression of this gene. Using stablytransformed plants, we first characterized the extended promoter region that positively regulates basal expression from thePDF1.2 promoter. Second, using promoter deletion constructs including one from which the GCC-box region was deleted,we observed a substantially lower response to jasmonate than lines carrying this motif. In addition, point mutationsintroduced into the core GCC-box sequence substantially reduced jasmonate responsiveness, whereas addition of a20-nucleotide-long promoter element carrying the core GCC-box and flanking nucleotides provided jasmonate responsive-ness to a 35S minimal promoter. Taken together, these results indicated that the GCC-box plays a key role in conferringjasmonate responsiveness to the PDF1.2 promoter. However, deletion or specific mutations introduced into the coreGCC-box did not completely abolish the jasmonate responsiveness of the promoter, suggesting that the other promoterelements lying downstream from the GCC-box region may also contribute to jasmonate responsiveness. In other experi-ments, we identified a jasmonate- and pathogen-responsive ethylene response factor transcription factor, AtERF2, whichwhen overexpressed in transgenic Arabidopsis plants activated transcription from the PDF1.2, Thi2.1, and PR4 (basicchitinase) genes, all of which contain a GCC-box sequence in their promoters. Our results suggest that in addition to theirroles in regulating ethylene-mediated gene expression, ethylene response factors also appear to play important roles inregulating jasmonate-responsive gene expression, possibly via interaction with the GCC-box.
Diseases caused by fungal, bacterial, and viralpathogens result in enormous reductions in yield andquality from agricultural crops around the world.However, although plants are continuously chal-lenged by pathogenic microorganisms, the develop-ment of disease is a relatively rare event. It is clear,therefore, that plants possess efficient defense sys-tems. Effective induction of the defense response re-quires pathogen recognition followed by a networkof signal transduction processes, resulting in therapid activation of defense gene expression. A num-ber of secondary signal molecules such as salicylicacid (SA), jasmonic acid (JA) and its methyl ester,methyl jasmonate (MeJA), and ethylene act to am-plify and regulate defense responses after initial ac-tivation (Dong, 1998; Reymond and Farmer, 1998;Schenk et al., 2000). Interestingly, the SA- andjasmonate/ethylene-dependent signaling pathwaysappear to modulate plant defense responses againstdifferent classes of pathogens (for review, see
Thomma et al., 2001). Treatment with MeJA providesArabidopsis plants with protection against subse-quent inoculation with a range of necrotrophic fungi,but not against the biotrophic pathogen Peronosporaparasitica (Thomma et al., 1998). Similarly, severalstudies of Arabidopsis mutants including ein2, coi1,jar1, and fad3/7/8, in which the jasmonate/ethylene-response is disrupted, have indicated that resistanceto a variety of necrotrophic pathogens requires afunctional jasmonate/ethylene-signaling pathway(Staswick et al., 1998; Vijayan et al., 1998; Thomma etal., 1999). In contrast, resistance to the biotroph P.parasitica but not to the necrotroph Alternaria brassici-cola is enhanced by pretreatment with isonicotinicacid, a functional analog of SA (Thomma et al., 1998).
A number of classes of regulatory proteins andtranscription factors are known to play importantroles in relaying the pathogen-initiated signals todownstream components for the activation of plantdefense responses. Among these, ethylene responsefactor domain-containing transcription factors (ERFs)are implicated as key regulators of plant defenseresponses. For example, the tomato (Lycopersicon es-culentum) gene Pti4 encodes an ERF protein that isinduced by defense-signaling molecules and by thebacterial pathogen Pseudomonas syringae pv. tomato
1 This work was supported by the Grains Research and Devel-opment Corporation (postgraduate fellowship to R.L.B.).
* Corresponding author; e-mail [email protected]; fax 61–7–3214 –2950.
Article, publication date, and citation information can be foundat www.plantphysiol.org/cgi/doi/10.1104/pp.102.017814.
1020 Plant Physiology, June 2003, Vol. 132, pp. 1020–1032, www.plantphysiol.org © 2003 American Society of Plant Biologists www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

(Thara et al., 1999; Gu et al., 2000). Pti4 and othertomato ERFs Pti5 and Pti6 are phosphorylated by Ptokinase, and this provides a direct link betweenR-gene mediated pathogen recognition and subse-quent defense gene activation (Zhou et al., 1997; Guet al., 2000). Transgenic expression of ERFs such asERF1 of Arabidopsis, Pti4 of tomato, and Tsi1 oftobacco (Nicotiana tabacum) activates defense geneexpression and provides enhanced resistance againstfungal and bacterial pathogens (Park et al., 2001;Berrocal-Lobo et al., 2002; Gu et al., 2002). Expressionof ERFs may also provide resistance to abioticstresses (e.g. osmotic stress), indicating that multiplesignaling pathways may converge on ERFs to regu-late gene expression in response to a number of bioticand abiotic stresses (Fujimoto et al., 2000; Park et al.,2001; Memelink et al., 2001).
Several members of the ERF family bind specifi-cally to the sequence AGCCGCC through the con-served ERF domain (Hao et al., 1998). This cis-actingsequence, known as the GCC-box, is found in thepromoters of many pathogen-responsive genes suchas PDF1.2, Thi2.1, and PR4 (basic chitinase; Zhou etal., 1997; Manners et al., 1998). The GCC-box has beenshown to function as an ethylene-responsive elementthat is necessary, and in some cases sufficient, for theregulation of transcription by ethylene (Ohme-Takagi and Shinshi, 1995; Shinshi et al., 1995). Inaddition, the GCC-box is implicated in ozone-responsive expression of tobacco basic-typepathogenesis-related PR-1 protein gene via ethylene-dependent signaling (Grimmig et al., 2003).
More recently, an ERF subfamily of transcriptionfactors called octadecanoid-responsive CatharanthusAP2s (ORCAs) has been implicated in jasmonate-regulated expression of secondary metabolic path-ways in Madagascar periwinkle (Catharanthus roseus;Menke et al., 1999a; van der Fits and Memelink,2000). MeJA treatment and fungal elicitors rapidlyinduce transcript levels of Orca2 and Orca3. TheORCA2 and ORCA3 proteins regulate overlappingbut distinct sets of genes associated with secondarymetabolism via specific binding to a promoter ele-ment called the jasmonate- and elicitor-responsiveelement, which contains a core GCC-box (Menke etal., 1999a; van der Fits and Memelink, 2000).
The PDF1.2 gene of Arabidopsis encodes a plantdefensin, and its expression is induced by pathogenchallenge both locally at the site of inoculation byincompatible fungal pathogen and systemically inremote noninoculated regions of the plant (Pen-ninckx et al., 1996). This activation occurs via ajasmonate/ethylene-mediated signaling pathway,rather than via the SA-dependent pathway of defensegene activation (Penninckx et al., 1996, 1998). Al-though the promoter of the PDF1.2 gene containsseveral putative cis-acting sequence elements, suchas a GCC-box (Manners et al., 1998), the roles of theseelements and their respective trans-acting factors in
regulation of the PDF1.2 gene by jasmonates have notbeen investigated directly. We previously reportedon the jasmonate and pathogen-mediated activationof the PDF1.2 promoter as promoter-reporter trans-gene that mimicked the expression patterns of thenative PDF1.2 gene (Manners et al., 1998; Mitter et al.,1998). Examination of the 1,183-bp region upstreamof the transcription start site of the PDF1.2 generevealed several putative motifs with homology toknown cis-elements involved in the transcriptionalregulation of other genes, including those with par-ticular relevance for jasmonate-mediated expression.Two of these motifs, located between �277 and �495of the PDF1.2 promoter, included the jasmonate-responsive element, TGACG (�392 to �396) presentin the Lox1 gene of barley (Hordeum vulgare; Rousteret al., 1997), and a 9-bp sequence, AAATGTTGT(�410 to �419), similar to a sequence within thejasmonate-responsive region of the promoter of thesoybean (Glycine max) gene VspB (Mason et al., 1993).
In the present study, we set out to delineate theregions of the PDF1.2 promoter that control jas-monate responsiveness. We report here on the iden-tification of regions of the PDF1.2 promoter includinga GCC-box that actively modulate the expression ofthis gene in response to jasmonates. In addition, weidentified a jasmonate-, ethylene-, and pathogen-responsive Arabidopsis ERF transcription factorwhose transgenic overexpression in Arabidopsis re-sulted in transcriptional activation of the PDF1.2gene.
RESULTS
Production of Transgenic Lines Carrying PDF1.2Promoter Deletion Constructs and Identification of aPositive Regulatory Element in the PDF1.2 Promoter
We generated a series of 5� deletions of the PDF1.2promoter fused in frame with the �-glucoronidase(GUS, UidA) reporter gene in a binary vector for usein plant transformation (Fig. 1C). The locations of theputative cis sequences that may be associated withjasmonate responses were taken into account whendesigning the 5�-deletion series. The deletion seriesincluded, in addition to the complete P1 construct(�1,183), three constructs P2 (�789), P3 (�495), andP4 (�278), in which relatively large deletions of 394,688, and 905 nucleotides from the P1 construct weremade. The constructs P5 (�262) and P6 (�255) re-moved only 16 and 23 nucleotides from the previoussmallest construct P4 and were designed to examinethe role of the GCC-box (GCCGCC between �255 to�261). We generated between seven and 12 indepen-dent transgenic lines stably transformed for each ofthe PDF1.2-promoter deletion constructs. We ran-domly selected five independently transformed linesfor each PDF1.2 promoter deletion construct and an-alyzed GUS activity in homogenates from wholesterile-grown seedlings as described in “Materials
GCC-Box and Jasmonate-Mediated Activation of the PDF1.2 Gene
Plant Physiol. Vol. 132, 2003 1021 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.
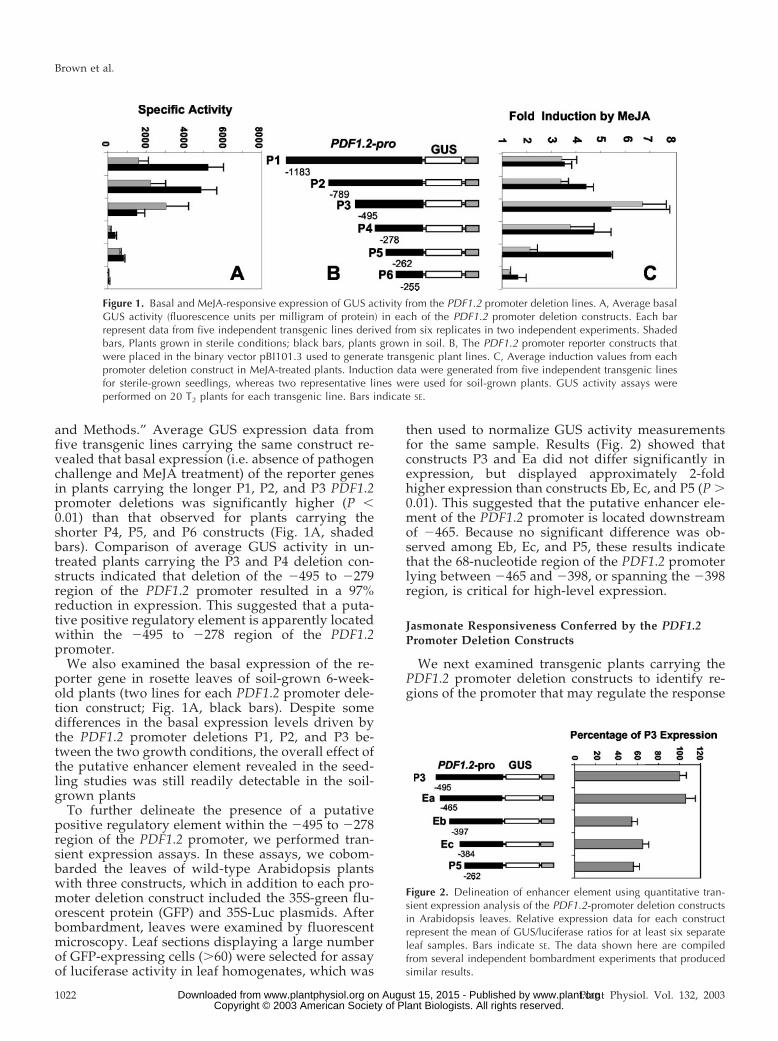
and Methods.” Average GUS expression data fromfive transgenic lines carrying the same construct re-vealed that basal expression (i.e. absence of pathogenchallenge and MeJA treatment) of the reporter genesin plants carrying the longer P1, P2, and P3 PDF1.2promoter deletions was significantly higher (P �0.01) than that observed for plants carrying theshorter P4, P5, and P6 constructs (Fig. 1A, shadedbars). Comparison of average GUS activity in un-treated plants carrying the P3 and P4 deletion con-structs indicated that deletion of the �495 to �279region of the PDF1.2 promoter resulted in a 97%reduction in expression. This suggested that a puta-tive positive regulatory element is apparently locatedwithin the �495 to �278 region of the PDF1.2promoter.
We also examined the basal expression of the re-porter gene in rosette leaves of soil-grown 6-week-old plants (two lines for each PDF1.2 promoter dele-tion construct; Fig. 1A, black bars). Despite somedifferences in the basal expression levels driven bythe PDF1.2 promoter deletions P1, P2, and P3 be-tween the two growth conditions, the overall effect ofthe putative enhancer element revealed in the seed-ling studies was still readily detectable in the soil-grown plants
To further delineate the presence of a putativepositive regulatory element within the �495 to �278region of the PDF1.2 promoter, we performed tran-sient expression assays. In these assays, we cobom-barded the leaves of wild-type Arabidopsis plantswith three constructs, which in addition to each pro-moter deletion construct included the 35S-green flu-orescent protein (GFP) and 35S-Luc plasmids. Afterbombardment, leaves were examined by fluorescentmicroscopy. Leaf sections displaying a large numberof GFP-expressing cells (�60) were selected for assayof luciferase activity in leaf homogenates, which was
then used to normalize GUS activity measurementsfor the same sample. Results (Fig. 2) showed thatconstructs P3 and Ea did not differ significantly inexpression, but displayed approximately 2-foldhigher expression than constructs Eb, Ec, and P5 (P �0.01). This suggested that the putative enhancer ele-ment of the PDF1.2 promoter is located downstreamof �465. Because no significant difference was ob-served among Eb, Ec, and P5, these results indicatethat the 68-nucleotide region of the PDF1.2 promoterlying between �465 and �398, or spanning the �398region, is critical for high-level expression.
Jasmonate Responsiveness Conferred by the PDF1.2Promoter Deletion Constructs
We next examined transgenic plants carrying thePDF1.2 promoter deletion constructs to identify re-gions of the promoter that may regulate the response
Figure 1. Basal and MeJA-responsive expression of GUS activity from the PDF1.2 promoter deletion lines. A, Average basalGUS activity (fluorescence units per milligram of protein) in each of the PDF1.2 promoter deletion constructs. Each barrepresent data from five independent transgenic lines derived from six replicates in two independent experiments. Shadedbars, Plants grown in sterile conditions; black bars, plants grown in soil. B, The PDF1.2 promoter reporter constructs thatwere placed in the binary vector pBI101.3 used to generate transgenic plant lines. C, Average induction values from eachpromoter deletion construct in MeJA-treated plants. Induction data were generated from five independent transgenic linesfor sterile-grown seedlings, whereas two representative lines were used for soil-grown plants. GUS activity assays wereperformed on 20 T2 plants for each transgenic line. Bars indicate SE.
Figure 2. Delineation of enhancer element using quantitative tran-sient expression analysis of the PDF1.2-promoter deletion constructsin Arabidopsis leaves. Relative expression data for each constructrepresent the mean of GUS/luciferase ratios for at least six separateleaf samples. Bars indicate SE. The data shown here are compiledfrom several independent bombardment experiments that producedsimilar results.
Brown et al.
1022 Plant Physiol. Vol. 132, 2003 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

to jasmonate. In both sterile- and soil-grown plants,although GUS activity levels were higher after MeJAtreatment, differences between constructs appearedto mirror the results from untreated plants in thatspecific GUS activities were considerably higher inlines carrying constructs P1, P2, and P3 than linescarrying constructs P4, P5, and P6 (data not shown).However, a different pattern emerged when -foldinduction ratios were compared (Fig. 1C). In sterile-grown plants, constructs P3 and P4 did not differsignificantly from one another in -fold induction ra-tios and showed significantly higher -fold inductionby MeJA than P1 and P2 (P � 0.01). Similarly, insoil-grown plants, lines carrying the P3 constructshowed a higher level of induction than lines carry-ing P1 and P2 constructs.
Using sterile-grown transgenic plants, the deletionof the 15-bp region between constructs P4 and P5resulted in a significant decrease in induction byMeJA (P � 0.01; Fig. 1C). A further decrease in in-duction was observed in P6 with the deletion of anadditional 7 bp comprising the core GCC motif ofAGCCGCC. In contrast to sterile-grown plants, thestably transformed soil-grown plants carrying the P5construct gave a similar -fold induction of the re-porter gene to plants carrying the P3 or P4. However,similar to sterile-grown plants, deletion of the coreGCC-box (P6 construct) significantly reduced thelevel of MeJA-mediated induction of GUS activity.Although these results indicate the importance of thisGCC-box motif for induction of the PDF1.2 gene byjasmonate, deletion of this box did not completelyabolish jasmonate responsiveness, indicating thatother elements downstream of �255 may also beinvolved in the regulation of this response. The bio-
logical significance of the difference between sterile-and soil-grown plants carrying construct P5 is notknown but if confirmed, could be related to addi-tional positive signaling pathways initiated by bioticor abiotic factors.
In an effort to confirm the role of the GCC-box inmediating jasmonate responsiveness, we generatedtwo additional PDF1.2 promoter constructs, mGCCand pIB (Fig. 3B). The mGCC consisted of the �495PDF1.2 promoter fragment with substitution of thecore GCC-box sequence (GCCGCC) with TCCTCA.The pIB construct was aimed at determining whetherthe GCC region alone was sufficient to conferjasmonate-responsive gene expression. This con-struct was a fusion of the extended GCC-box region(�250 to �270) upstream of a truncated 35S (�46)promoter derived from CaMV.
Analysis of the mean GUS expression in MeJA-treated sterile-grown whole seedlings stably trans-formed with the mGCC construct indicated that therewas no significant induction by MeJA compared withmock-treated controls (Fig. 3C). This experiment in-cluded plants stably transformed with construct P3as control, which showed the same highly significantincrease in MeJA-responsive induction (P � 0.01) asthe previous experiment described in Figure 1. Thisresult directly indicates that the GCC-box sequenceconfers jasmonate responsiveness to the PDF1.2promoter.
Next, we examined the jasmonate responsivenessconferred by the pIB construct linking the GCC-boxto a minimal 35S promoter in stably transformedsterile-grown plants. After MeJA treatment, theseplants showed some jasmonate response, but thelevel of induction was significantly less than that
Figure 3. A through C, Further confirmation that the GCC-box confers jasmonate responsiveness to the PDF1.2 promoterusing quantitative analysis of stably transformed plants. C, Results are presented as average -fold induction values from twoto five transgenic lines harboring each PDF1.2 promoter deletion construct. A, The basal expression of GUS from eachconstruct is also given. The mGCC construct is the same as construct P3 but otherwise contains three substitutions in the first(G–T), fourth (G–T), and sixth (C–A) nucleotides (shown in bold) to convert the core GCCGCC box sequence to TCCTCA.B, Construct pIB contains a 20-nucleotide-long sequence including the GCC-box and the surrounding nucleotides from the�270 to �250 of the PDF1.2 promoter linked to a truncated 35S cauliflower mosaic virus (CaMV) promoter. SE bars are alsoshown for each construct.
GCC-Box and Jasmonate-Mediated Activation of the PDF1.2 Gene
Plant Physiol. Vol. 132, 2003 1023 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

measured for construct P3 (P � 0.05). Plants trans-formed with a 35S minimal promoter construct alonedid not show any significant induction of GUS activ-ity after MeJA treatment (Fig. 3C).
Identification of MeJA-Responsive Arabidopsis ERFs
Jasmonate-responsive expression of GUS activityin plants stably transformed with PDF1.2 promoterconstructs including the GCC-box suggested that thisconserved sequence element might act as binding sitefor jasmonate-responsive ERF transcription factors.To identify gene family members encoding putativeArabidopsis ERFs, we used the consensus amino acidsequence of the DNA-binding domain of all knownERF members that bind to the GCC-box domain as aquery sequence for BLASTP searches of Arabidopsissequences available on GenBank. The searches re-turned 145 sequences representing unique proteins(see also Sakuma et al., 2002), of which 131 containeda single AP2/ERF-type DNA-binding domain. Be-cause of the relatively large size of this transcriptionfactor family, we selected representatives from se-quence clusters for expression profiling. With theexception of APETALA2, all of the genes were se-lected from the subfamily that contains only a singleERF domain. The selected sequences included ERF1(Solano et al., 1998), AtERF2, AtERF3, AtERF4,AtERF5 (Fujimoto et al., 2000), and 12 other ERFs (see“Materials and Methods” for locus numbers) forwhich no expression data were available in the liter-ature. Of the latter, AP6 was selected because it hadthe highest homology of any Arabidopsis ERF toORCA2. DNA sequences for each of these 18 ERFswere generated by PCR amplification from divergentregions outside the conserved DNA-binding domain,to avoid cross-hybridization among the differentmembers of the gene family.
To identify jasmonate-responsive ArabidopsisERFs, we first examined the expression patterns ofthe 18 selected Arabidopsis ERFs by reverse northernhybridization analysis of a macroarray of the variablesequence from each gene. Expression patterns ofERFs showing induction after MeJA treatment werethen confirmed by northern-blot analysis. Only five(ERF1, AtERF2, AtERF3, AtERF4, and RAP2.10) of the18 AP2/ERF gene family members present on themacroarray, hybridized with cDNA prepared fromMeJA-treated plants and their associated controls.All of these five genes showed induction by MeJA at0.5 h, with a peak in induction after 1 h of MeJAexposure (data not shown). With the exception ofAtERF4, which remained induced until 4 h, the tran-scription levels of all genes had returned to thoseobserved in control plants by 2 h (data not shown).Examination of expression patterns by northern blot-ting confirmed the results obtained from macroarrayanalysis (Fig. 4A, left panel). Induction of PDF1.2 wasobserved after 4 h of exposure to MeJA in theseexperiments (Fig. 4A, left panel).
We further quantified the induction ratios of ERF1,AtERF2, and PDF1.2 after MeJA treatment by quan-titative real-time RT-PCR, which is particularly suit-able for specific detection of transcripts from themembers of multigene families (Charrier et al., 2002).In these time-course experiments, we used RNA iso-lated from plants grown and treated at separate oc-casions. Results from these experiments which in-cluded early as well as late time-points (e.g. 24 and48 h) were in agreement with the results fromnorthern-blot and macroarray analyses as both ERF1and AtERF2 displayed distinct yet overlapping ex-pression patterns (Fig. 4B). AtERF2 was substantiallymore responsive to MeJA treatment than ERF1, espe-cially at earlier time points (e.g. 3 and 6 h), whereasa sustained level of expression was observed for both
Figure 4. A, Expression profiling of selected ERF genes by northernblot in a time-course analysis. Left panel, Expression in response toMeJA; right panel, expression in response to ethylene for each gene.Top and bottom panels represent RNA from control (C) and treated(T) plants, respectively. B, Induction ratios of the Arabidopsis ERF1,AtERF2, and PDF1.2 genes obtained by real-time quantitative reversetranscriptase (RT)-PCR after MeJA treatment.
Brown et al.
1024 Plant Physiol. Vol. 132, 2003 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.
genes at late time points. Consistent with knownexpression pattern of PDF1.2, we detected a stronginduction of the PDF1.2 transcripts starting at 6 to12 h after treatment.
We also tested the ethylene response profiles ofthose ERFs that were induced by MeJA by northern-blot analysis. These analyses showed that AtERF2,AtERF3, AtERF4, and RAP2.10 responded in a similarmanner to ethylene as observed for MeJA (Fig. 4A,right panel). They displayed rapid induction on eth-ylene exposure, with expression returning to controllevels after one to 2 h. ERF1 expression after ethylenetreatment was more sustained and continued to in-crease over the time points tested.
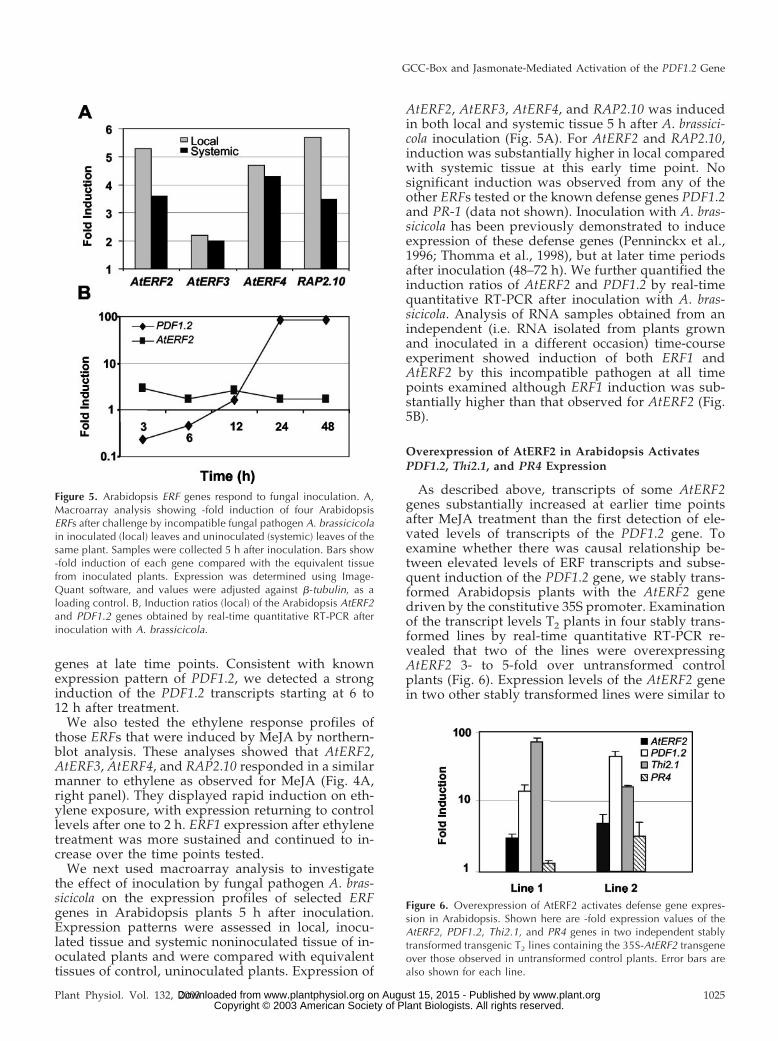
We next used macroarray analysis to investigatethe effect of inoculation by fungal pathogen A. bras-sicicola on the expression profiles of selected ERFgenes in Arabidopsis plants 5 h after inoculation.Expression patterns were assessed in local, inocu-lated tissue and systemic noninoculated tissue of in-oculated plants and were compared with equivalenttissues of control, uninoculated plants. Expression of
AtERF2, AtERF3, AtERF4, and RAP2.10 was inducedin both local and systemic tissue 5 h after A. brassici-cola inoculation (Fig. 5A). For AtERF2 and RAP2.10,induction was substantially higher in local comparedwith systemic tissue at this early time point. Nosignificant induction was observed from any of theother ERFs tested or the known defense genes PDF1.2and PR-1 (data not shown). Inoculation with A. bras-sicicola has been previously demonstrated to induceexpression of these defense genes (Penninckx et al.,1996; Thomma et al., 1998), but at later time periodsafter inoculation (48–72 h). We further quantified theinduction ratios of AtERF2 and PDF1.2 by real-timequantitative RT-PCR after inoculation with A. bras-sicicola. Analysis of RNA samples obtained from anindependent (i.e. RNA isolated from plants grownand inoculated in a different occasion) time-courseexperiment showed induction of both ERF1 andAtERF2 by this incompatible pathogen at all timepoints examined although ERF1 induction was sub-stantially higher than that observed for AtERF2 (Fig.5B).
Overexpression of AtERF2 in Arabidopsis ActivatesPDF1.2, Thi2.1, and PR4 Expression
As described above, transcripts of some AtERF2genes substantially increased at earlier time pointsafter MeJA treatment than the first detection of ele-vated levels of transcripts of the PDF1.2 gene. Toexamine whether there was causal relationship be-tween elevated levels of ERF transcripts and subse-quent induction of the PDF1.2 gene, we stably trans-formed Arabidopsis plants with the AtERF2 genedriven by the constitutive 35S promoter. Examinationof the transcript levels T2 plants in four stably trans-formed lines by real-time quantitative RT-PCR re-vealed that two of the lines were overexpressingAtERF2 3- to 5-fold over untransformed controlplants (Fig. 6). Expression levels of the AtERF2 genein two other stably transformed lines were similar to
Figure 5. Arabidopsis ERF genes respond to fungal inoculation. A,Macroarray analysis showing -fold induction of four ArabidopsisERFs after challenge by incompatible fungal pathogen A. brassicicolain inoculated (local) leaves and uninoculated (systemic) leaves of thesame plant. Samples were collected 5 h after inoculation. Bars show-fold induction of each gene compared with the equivalent tissuefrom inoculated plants. Expression was determined using Image-Quant software, and values were adjusted against �-tubulin, as aloading control. B, Induction ratios (local) of the Arabidopsis AtERF2and PDF1.2 genes obtained by real-time quantitative RT-PCR afterinoculation with A. brassicicola.
Figure 6. Overexpression of AtERF2 activates defense gene expres-sion in Arabidopsis. Shown here are -fold expression values of theAtERF2, PDF1.2, Thi2.1, and PR4 genes in two independent stablytransformed transgenic T2 lines containing the 35S-AtERF2 transgeneover those observed in untransformed control plants. Error bars arealso shown for each line.
GCC-Box and Jasmonate-Mediated Activation of the PDF1.2 Gene
Plant Physiol. Vol. 132, 2003 1025 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

that observed in untransformed control plants (datanot shown). We also detected a 16- and 51-fold in-crease in PDF1.2, an 80- and 50-fold increase inThi2.1, and 2- and 3-fold increase in PR4 (basic chiti-nase) transcript levels, respectively, in these twoAtERF2 overexpressing lines (Fig. 6), whereas nosuch increase in the expression of these genes wasevident in untransformed control plants and in twoother transgenic lines with wild-type levels ofAtERF2 transcripts. Overall, these results suggestthat the AtERF2 protein is capable of activating thePDF1.2, Thi2.1, and PR4 genes when overexpressedin transgenic plants.
AtERF2 Transcript Level Remains Unchanged inArabidopsis Mutants Constitutively Expressing thePDF1.2 Gene
To determine whether enhanced levels of theAtERF2 transcript can be found in Arabidopsisdefense-signaling mutants that constitutively expressthe PDF1.2 and Thi2.1 genes, we examined the tran-script levels of AtERF2 by real-time quantitative RT-PCR analysis in Arabidopsis defense-signaling mu-tants cev-1 and cpr5 (Bowling et al., 1997; Ellis andTurner, 2001). These analyses showed no significantchange in the transcript level of AtERF2 in thesemutants (data not shown). However, consistent withprevious reports, we detected enhanced transcriptlevels of PDF1.2 in both cev-1 and cpr5 mutants (datanot shown).
DISCUSSION
The GCC-Box Is a Major Element Conferring theJasmonate Responsiveness of the PDF1.2 Promoter
In this report, we have identified the �255 to �277promoter region as being critical for regulation of thePDF1.2 gene by jasmonate. Plant lines carrying con-structs with deletions in this region (P5 and particu-larly P6) showed significantly lower induction byMeJA in comparison with all other deletion con-structs. Penninckx et al. (1998) and Manners et al.(1998) have reported that the level induction ofPDF1.2 by both MeJA and ethylene in tissue culture-grown plants was increased by growth of the plantsunder non-sterile conditions. Therefore, we usedboth types of growth conditions in assessing the ex-pression from the promoter-deletion constructs. Ingeneral, results reported in this manuscript sup-ported this observation for the induction of PDF1.2promoter-driven reporter gene expression by MeJAin soil-grown plants. Augmented induction in soil-grown plants may be a consequence of physiologicaldifferences between plants grown in soil and in vitroor as a result of potentiation by soil-associated bio-logical factors.
The �255 to �277 region contains the GCC-box,which is a common motif in the promoters of various
genes encoding defense-related proteins, and hasbeen previously identified as the specific binding sitefor members of the ERF subfamily of AP2/ERF tran-scription factors (Ohme-Takagi and Shinshi, 1995;Buttner and Singh, 1997; Zhou et al., 1997). Analysisof Arabidopsis lines stably transformed with aPDF1.2 promoter-GUS reporter transgene revealed astepwise reduction in induction by jasmonate as thesequences immediately preceding the GCC-box (P4and P5) were deleted, with a further reduction ofinduction after deletion of the core GCC motif itself(P6). In other studies, a 21-nucleotide sequence con-taining the GCC-box in the promoter of the PDF1.2promoter provided jasmonate responsiveness whenlinked to a minimal 35S CaMV promoter, and muta-tions introduced into the GCC-box significantly re-duced the jasmonate responsiveness of the plantscarrying P3 construct. Taken together, these resultsindicate that the GCC-box in the promoter of thePDF1.2 gene modulates the jasmonate responsive-ness of the PDF1.2 gene. Similar to our findingsreported here, Menke et al. (1999a) have recentlyshown that jasmonate- and elicitor-responsive ele-ments found in the promoter of the secondary me-tabolite biosynthetic gene, Str, of Madagascar peri-winkle, contains a GCC-like box that is bothnecessary and sufficient for jasmonate- and elicitor-responsive expression.
It has been well established that members of ERFtranscription factor family interact with the GCC-boxsequence. Detailed analysis of the in vitro bindingrequirements of three ERF transcription factors fromtobacco indicated that whereas the contact of specificamino acid residues of the ERFs are confined to the6-bp core GCCGCC region of the GCC-box, sur-rounding nucleotides are necessary for high-affinitybinding (Hao et al., 1998). These sequences may alsomodulate differential binding by different ERFs. Thismay explain the observation that plants carrying theP5 construct, with a deletion of the 5�-flanking se-quence to the core GCC motif, showed decreasedresponsiveness to jasmonate but significantly higherbasal expression levels compared with the P4 plantscarrying an extended GCC motif. The removal of thenucleotides specifying recruitment of the specificERFs that up-regulate PDF1.2 transcription in re-sponse to jasmonate could allow competitive bindingof other ERF subfamily members, which althoughunresponsive to jasmonate, may increase basal geneexpression.
Although the results presented here identify the�255 to �277 region, containing the core GCC-box,and flanking sequences as critical for gene regulation,they do not exclude the additional significance ofcis-elements located further downstream in thePDF1.2 promoter. In fact, other cis-elements also ap-pear to contribute to jasmonate responsiveness, asindicated by residual induction displayed by linescarrying the P6 construct. Interestingly, in P6, a par-
Brown et al.
1026 Plant Physiol. Vol. 132, 2003 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

tial GCC-box sequence (AGCCGC, �219 to �214)occurs in the PDF1.2 promoter downstream from theconsensus GCC-box at �255 to �277. It is possiblethat some transcription factors can bind to this GCC-box like region and activate gene expression throughinteractions with ERFs. Coregulation of gene expres-sion by the GCC-box and other promoter elementshas also been observed previously. For example, theGCC-box is required but not sufficient for high-levelinduction of the tobacco osmotin gene by ethylene(Raghothama et al., 1997). Also, a G-box is associatedwith jasmonate responsiveness in defense-associatedgenes in various plant species (Kim et al., 1992;Mason et al., 1993). In the PDF1.2 promoter, theGCC-box also directly follows a putative G-box mo-tif (�255 to �250) with the sequence CATGTG(Manners et al., 1998). Close proximity of theseG- and GCC-box elements is a common feature oftobacco PR genes and is postulated to allow cross-linking of ERF and bZIP transcription factors forregulation of gene expression during plant defenseresponses (Sessa et al., 1995; Buttner and Singh,1997). Although the G-box has been shown to beimportant in regulating the expression of certainjasmonate-responsive genes (Kim et al., 1992; Masonet al., 1993), the putative G-box in the PDF1.2 pro-moter lacks the exact ACGT core sequence charac-teristic of bZIP transcription factor binding sites(Izawa et al., 1993) and thus may not represent afunctional G box. Further work would be needed todetermine the role of these additional elements inthe regulation of the PDF1.2 gene.
AtERF2, a Jasmonate/Ethylene- and Pathogen-InducibleERF, Constitutively Activates Transcription from theGCC-Box Containing Defense Genes
The deletion analysis of the PDF1.2 promoter sug-gested that the GCC-box is critical for jasmonate-mediated regulation of the PDF1.2 gene expression. Itthus seemed a reasonable hypothesis that one ormore members of the Arabidopsis ERF transcriptionfactor family homologous to the periwinkle ORCA2and ORCA3 transcription factors (Menke et al.,1999a; van der Fits and Memelink, 2000) might act toregulate the expression of PDF1.2 in response to jas-monates. Therefore, we examined the mRNA levelsof selected ERFs in Arabidopsis plants after treat-ment with inducers of PDF1.2 expression and iden-tified ERFs that show distinct induction profiles.ERF1, AtERF2, AtERF3, AtERF4, and RAP2.10 all dis-played immediate early induction, with mRNA accu-mulating after only 0.5 h of exposure to MeJA. MeJA-induced expression of each of these genes occurredearlier than that observed for PDF1.2, suggesting thatone or more of these ERFs may regulate PDF1.2expression. Rapid and transient induction of AtERF2by MeJA mimicked the expression patterns of theOrca2 and Orca3 transcripts, which also showed a
strong induction 0.5 h after exposure to MeJA(Menke et al., 1999a, 1999b; van der Fits andMemelink, 2000, 2001). In addition, the deducedamino acid sequence of AtERF2 is highly similar tothat of the ORCA3, particularly around the Ser-richregion, a putative phosphorylation site located at theC terminus of ORCA3 protein (van der Fits andMemelink, 2000). These similarities between AtERF2and ORCA2/3 suggested that AtERF2 might play arole in MeJA-mediated induction of the PDF1.2 gene.We specifically quantified expression from thePDF1.2 gene by quantitative real-time RT-PCR todistinguish the expression of PDF1.2 from three otherPDF1.2-like expressed sequences that exist in Arabi-dopsis. The predicted mature peptides encoded bygenes PDF1.2a (same as PDF1.2), PDF1.2b, andPDF1.2c are identical (R.L. Brown and K. Karzan,unpublished data; Thomma et al., 2002). Moreover,the upstream regulatory sequences of the latter twogenes contain regions with substantial homology tothe GCC-box and neighboring sequences found inthe promoter of the PDF1.2 gene (R.L. Brown and K.Karzan, unpublished data). The identification of mul-tiple family members homologous to PDF1.2 meansthat studies of the expression of this gene basedsolely on hybridization signals may be misleading.This was especially critical to rule out the possibilitythat in AtERF2-overexpressing plants, expression ofPDF1.2 but not any other PDF1.2-like genes is de-tected because it is possible that other PDF1.2-likegenes might also be activated due to having GCC-boxlike sequences in their promoters. Our analysisshowed that overexpression of AtERF2 activatedtranscription of defense genes such as PDF1.2, Thi2.1,and PR4. All of these genes contain GCC-boxes intheir promoters (Epple et al., 1995; Zhou et al., 1997;Manners et al., 1998), and their activation by AtERF2probably occurs via the GCC-box. Fujimoto et al.(2000) have demonstrated the ability of AtERF2 tointeract with the GCC-box for the activation of re-porter gene expression in transient expression assays.
The AtERF2 protein also shares high amino acidsequence identity with the tomato ERF, Pti4, whoseoverexpression in Arabidopsis also results in ele-vated expression of PDF1.2, Thi2.1, and PR4 andenhanced resistance against the fungal pathogen Ery-siphe orontii and the bacterial pathogen P. syringae pv.tomato (Gu et al., 2002). Defense gene activation andan enhanced disease resistance phenotype were alsoreported for Arabidopsis plants overexpressing ERF1(Berrocal-Lobo et al., 2002; Lorenzo et al., 2003). Thismay indicate that ERF1 and AtERF2 may have over-lapping or redundant functions. Although we havenot tested the level of disease resistance in plantsoverexpressing AtERF2, we predict that these plantsmay also show elevated levels of disease resistancedue to constitutive overexpression of a number ofdefense genes which may be further induced duringinfection due to availability of additional positive
GCC-Box and Jasmonate-Mediated Activation of the PDF1.2 Gene
Plant Physiol. Vol. 132, 2003 1027 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

regulators of defense genes, which may require in-duction by a pathogen.
Convergence of Multiple Signaling Pathways on ERFs
Recent studies (Fujimoto et al., 2000; Ohta et al.,2000; Park et al., 2001; Cheong et al., 2002; Gu et al.,2002; Mazarei et al., 2002; Nishiuchi et al., 2002;Onate-Sanchez and Singh, 2002; Wu et al., 2002) sug-gest that ERFs are regulated differentially at both thetranscriptional and posttranscriptional levels andthat different ERF proteins may regulate various sub-sets of GCC-box-containing genes. For instance, theERF genes studied here including AtERF2 were re-sponsive to the known inducers of the PDF1.2 gene,namely MeJA and ethylene (Fujimoto et al., 2000;Lorenzo et al., 2003; this study), and inoculation withfungal pathogen (this study). Identification ofjasmonate-, ethylene-, pathogen-, wounding-,nematode-, and stress-inducible ERF transcriptionfactors suggest that ERFs can spatially and tempo-rally regulate gene expression in response to bioticand abiotic cues. As recently suggested by Fujimotoet al. (2000), both ethylene and stress-related signal-ing pathways seem to converge on a subset of differ-ent ERFs, which subsequently regulate a subset ofGCC-box containing genes. Results reported here,together with studies on ORCA2/3 proteins, andERF1 (Lorenzo et al., 2003) suggest that componentsof jasmonate-signaling pathways converge withthose of other pathways (e.g. abiotic stress, wound-ing, ethylene, and pathogen/elicitor) on ERF tran-scription factors in regulating gene expression.
Interestingly, so far a number of ERF proteins havebeen shown to be able to trans-activate the PDF1.2expression when expressed stably or transiently inArabidopsis (Fujimoto et al., 2000; Ohta et al., 2001;Berrocal-Lobo et al., 2002; Gu et al., 2002; Wu et al.,2002; this study). However, it is not clear which ERFsmight be responsible for activating the PDF1.2 ex-pression in planta in response to jasmonate or patho-gen signals. More recently, Lorenzo et al. (2003) havesuggested that ERF1 and AtERF2 may have redun-dant functions. It is also possible that the activationof the PDF1.2 gene in transgenic plants constitutivelyexpressing a GCC-box binding protein occurs some-what nonspecifically. In this study, AtERF2, ajasmonate-, pathogen-, and ethylene-inducible ERF,was able to activate the expression from the PDF1.2gene when overexpressed in transgenic plants. How-ever, examination of the constitutive PDF1.2-expressing Arabidopsis mutants, cev1 and cpr5, didnot reveal any increase in the level of AtERF2 tran-script. These results are consistent with those ofOnate-Sanchez and Singh (2002), who have recentlyreported that the expression of a number ofjasmonate-, pathogen-, and SA-inducible ERFs, in-cluding AtERF2 have not changed in cep1 and cpr5-2mutants. Taken together, these data suggest that
PDF1.2 overexpression observed in these mutantsmay be mediated by other ERFs. Alternatively, assuggested by Gu et al. (2002), relatively low levels ofAtERF2 may be activated posttranslationally by aconstitutively expressed putative kinase without anydetectable increase at the transcript level. Such aphosphorylation event could influence the localiza-tion of AtERF2 to nucleus and modulate its DNA-binding ability or its interaction with other transcrip-tion factors. Phosphorylation by Pto kinase enhancesbinding of the tomato ERF Pti4 to the GCC-box (Gu etal., 2000). Several other AP2/ERFs, such as CBF1,CBF2, CBF3 ORCA2, ORCA3, and AtERF5, containpotential recognition sites for protein kinases, whichmay play important roles in modulating the functionof these proteins (Medina et al., 1999; Yamamoto etal., 1999; Fujimoto et al., 2000).
Alternatively to ERF activation, inactivation of aputative repressor protein, perhaps also an ERF,might be responsible for constitutive expression ofthe PDF1.2 gene in the cev1 and cpr5 mutants notedabove. Recent studies have indicated that some ERFsmay act as repressors rather than activators of tran-scription (Fujimoto et al., 2000; Ohta et al., 2001). Aconserved sequence motif called ethylene-associatedamphiphilic repression within the C-terminal regionof these transcription factors seems to be essential forrepression by such repressor ERFs (Ohta et al., 2001).For example, the AtERF3, AtERF4, and RAP2.10 pro-teins all contain the ethylene-associated amphiphilicrepression motif, and AtERF3 and AtERF4 have beenshown to function as repressors in in vitro assays(Fujimoto et al., 2000). Thus, even though the MeJA-,ethylene-, and A. brassicicola-induced expression ofAtERF3, AtERF4, and RAP2.10 precedes induction ofPDF1.2 in the research reported herein, it cannot beconcluded that these genes are involved in the spe-cific activation of PDF1.2.
Enhancer Region in the PDF1.2 Promoter
In this paper, we have also reported on promoterelements other than the GCC region, that appear toplay roles in regulating the overall levels of expres-sion of the PDF1.2 gene of Arabidopsis. Analysis ofreporter gene expression in multiple transgenic linesfor each promoter-reporter deletion line showed thata strong enhancer element is located within the �277to �495 region of the PDF1.2 promoter. Further de-letion analysis of the PDF1.2 promoter using thetransient expression system allowed delineation ofthis element to a 68-nucleotide region located be-tween �398 and �465. This region of the PDF1.2promoter contains several motifs with similarity toregulatory elements of other genes, and/or knownrecognition sites for transcription factors. These in-cluded the sequences TTCGAC (�398 to �404), in-volved in the SA responsiveness to the PR-2d gene oftobacco (Shah and Klessig, 1996), and CGACG (�399
Brown et al.
1028 Plant Physiol. Vol. 132, 2003 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

to �403), necessary for high-level expression of therice (Oryza sativa) Amy3D gene under conditions ofsugar starvation (Hwang et al., 1998). In addition,this 68-nucleotide region contains the sequenceCAACA (�454 to �458), which was revealed by invitro binding experiments as the DNA recognitionsequence for the RAV1 transcription factor of Arabi-dopsis (Kagaya et al., 1999). The RAV1 protein bindsto this motif through a protein domain that stronglyresembles the AP2 domain found in AP2/ERF pro-teins (Kagaya et al., 1999). Also located within the68-nucleotide putative enhancer region of the PDF1.2promoter are sequences with homology to the GT-and GATA (I box) elements, which mediate light-responsive expression of many genes (Zhou, 1999).GT-elements also exist in many light-unresponsivegenes, indicating a diverse range of functions and,depending on promoter structure, may have a posi-tive or negative effect on transcription (Zhou, 1999).Further experimentation is necessary to determinewhether one or more of the elements described abovemodulate high-level expression on the PDF1.2 pro-moter, or whether the putative enhancer containedwithin this region of the PDF1.2 promoter representsa novel element.
In conclusion, our results have demonstrated a rolefor the GCC-box in conferring jasmonate responsive-ness to the PDF1.2 promoter. This motif and its sur-rounding bases most probably interact with severalmembers of the ERF family of transcription factors,some of which may be also induced by jasmonates.Analysis of the PDF1.2 promoter has also indicatedthe existence of other, probably interactive, cis ele-ments necessary for high-level jasmonate-mediatedregulation of PDF1.2 expression during plant defenseresponses.
MATERIALS AND METHODS
Production of the PDF1.2 Promoter Constructs
The cloned promoter of the PDF1.2 gene described in Manners et al.(1998) was used as template for the production of all PDF1.2-promoterdeletions. Promoter fragments of varying lengths were amplified by PCRusing the following forward primers for constructs P2, P3, P4, P5, and P6,respectively (Fig. 1B): 5�-ATCAAAGCT TTGAGCCTCTCACTTGCGGTC-3�,5�-ATCAAA GCTTTGCTGCTCTTGAGATCAACC-3�, 5�-GCCGAAGCTT-CCATTCAGATTAACCAGCC-3�, 5�-ATCAAAGCTTAGCCGCCCATGTG-AACGATG-3�, 5�-ATCAAAGCTTCATGTGAACGATGTAGCATTAGC-3�,and a common reverse primer 5�-GAGAGAGGATCCTGATGGAAGCAAA-CTTAGCCATG-3� for all constructs. The PDF1.2-promoter deletions werecloned into the HindIII-BamHI site of the binary vector pBI101.3 (CLON-TECH, Hampshire, UK), in frame with the UidA gene, using standardmolecular biology techniques (Sambrook et al., 1989). The pA35S constructwas made by amplifying the �46 to �10 region of the CaMV 35S promoterusing the primer pair 5�-CACTATGTCGACCAAGACCTTTCCTCTATA-TAAG-3� and 5�-CCCGGGGATCCGTCCTCTCAAAATGAATGGAAC-3�.Amplification product was digested with SalI and BamHI and inserted infront of the UidA gene in pBI101.3 also digested with SalI and BamHI. Thisconstruct is termed pB35S. The transgene cassette in pB35S was then re-moved by digesting HindIII and EcoRI and inserted into HindIII/EcoRIdigested binary vector pAOV, which carries a gene for Basta resistance(Mylne and Botella, 1998). The pIB construct was made by amplifying the�270 to �250 region of the PDF1.2 promoter using primers 5�-
TTAATCAAGCTTGATTAACCAGCCGCC-3� and 5�-TGCTACGTCGACC-ACATGGGCGGCTGG-3�. Amplification product was then digested withSalI and HindIII and ligated onto SalI/HindIII-digested pB35S, which wascalled pB6. The transgene cassette in pB6 was then removed by digestingwith HindIII/EcoRI and cloned into HindIII/EcoRI-digested pAOV. ThemGCC construct was generated via a two-step PCR process using primerpairs: (a) 5�-ATCAAAGCTTTGCTGCTCTTGAGATCAACC-3� and 5�-CGTTCACATGTGAGGATGGTTAATC-3� and (b) 5�-GATTAACCATCC-TCACATGTGAACG-3� and 5�-GAGAGAGGATCCTGATGGAAGCAAA-CTTAGCCATG-3�. The amplification product was digested with BamHI andHindIII and was cloned into BamHI/HindIII-digested binary vectorPBI101.3. The transgene cassette was then removed from this plasmid byHindIII/EcoRI digest and cloned into HindIII/EcoRI digested pAOV. Plas-mids were introduced into Escherichia coli DH5-� cells via heat shock trans-formation (Sambrook et al., 1989). The fusion constructs were verified bysequencing using the antisense primer (5�-CAGTTTTCGCGATCCAG-ACTG-3�) for the UidA gene. Binary vectors were transferred to Agrobacte-rium tumefaciens via triparental mating. The procedure outlined by Valvek-ens et al. (1988) was used for the production of transgenic Arabidopsisplants of C24 ecotype, and transformants were selected based on kanamycinresistance. The mGCC, pIB, pA35, and an additional P3 construct wereintroduced into Col-0 plants by using the flower-dip transformation proce-dure (Clough and Bent, 1998).
Plant Treatments for Promoter Activity Assays
For sterile-grown plants, Arabidopsis seeds were surface sterilized andevenly positioned across petri dishes containing germination medium(Valvekens et al., 1988) at a density of 70 seeds per plate. Seeds werestratified at 4°C for 3 d, and the petri dishes were transferred to growthcabinets with a 12-h photoperiod, at 24°C. Induction experiments werecarried out on 16-d-old seedlings. For soil-grown plants, seeds were germi-nated on germination medium in petri dishes, and 10-d-old seedlings weretransferred to soil, with five individuals planted in each 4- � 4-cm pot forinduction experiments using exogenous chemicals. For ease of inoculation,only four individuals were transplanted per pot for experiments whenplants were challenged by the fungal pathogen Alternaria brassicicola. Plantswere grown under short days (8-h photoperiod) to delay bolting. Daytimetemperature was 21°C with 70% humidity, and nighttime temperature was24°C, with 60% humidity. Induction experiments were performed on 5- to6-week-old plants.
For plant treatments, the plants pots or tissue culture plates were placedinto 1-L phytocon vessels (Sigma-Aldrich, St. Louis), which were sealedaround the rim with plasticine. Ethylene was applied at a concentration of200 �L L�1 by injection of gaseous ethylene through a rubber septum. MeJAwas applied at a concentration of 0.1 �mol L�1 air, by inoculation of 5 �L ofa 21.8 mm solution of MeJA (in ethanol) to a ball of cotton wool taped to theinside of the container. Control plants were placed in identical chambers,but without the addition of inducing agents. Samples were collected after24 h of incubation in continuous light. For the seedling-based assay, 20individual whole transgenic plants (T2 generation) were bulked for eachreplicate sample. For each treatment, three replicate sets of samples weretaken from each of two independent experiments. Five plants were bulkedfor each replicate in the mature plant assays. MeJA treatment of plants usedin additional real-time quantitative RT-PCR time-course experiments wasdone according to Schenk et al. (2000).
Fungal Inoculation
Leaves of soil-grown plants were inoculated with 5-�L drops of a freshlyharvested conidial spore suspension (5 � 105 spores mL�1 in water) of A.brassicicola (isolate UQ4273). The plants were placed in a propagator flatunder a clear polystyrene lid to maintain high humidity. The treated leaveswere collected separately from the untreated leaves of the same plant, 5 hafter inoculation.
Quantitative GUS Assays for Promoter Activity
Transgenic plants at T2 generation (20 individuals) stably transformedwith each promoter reporter construct were used. Promoter activity wasassessed by quantitative GUS assays of the UidA gene, using 4-methyl-
GCC-Box and Jasmonate-Mediated Activation of the PDF1.2 Gene
Plant Physiol. Vol. 132, 2003 1029 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

umbelliferyl-�-d-glucuronide (Sigma-Aldrich) as a substrate. The assay wascarried out according to the procedure of Jefferson (1987), adapted for usewith microtitre plates and the Fluoroskan Ascent fluorometer (Labsystems,Helsinki). The total protein content of samples was assessed using thedye-binding protein assay adapted for microtitre plates (Bio-Rad Laborato-ries, Hercules, CA).
Transient Expression Assays
The pBINm-GFP5-ER plasmid (Haseloff et al., 1997) containing a syn-thetic GFP gene under the control of the 35S promoter (CaMV) was used asa transformation control in all experiments. The pSP35Slucnos plasmidcontaining the firefly luciferase gene under the control of the 35S promoter(CaMV) was a gift from Dr. Robert Birch (Department of Botany, TheUniversity of Queensland, Australia). PCR amplified PDF1.2-promoterdeletion fragments using the forward primers 5�-AACAAGCTTTTTAGT-CCACAACG-3�, 5�-TGTAAGCTTATGACGAAGGTC-3�, and 5�-AAG-CTTGACTATGAACTGC-3� for constructs Ea, Eb, and Ec, respectively, anda common reverse primer 5�-GAGAGAGGATCCTGATGGAAGCAAA-TTAGCCATG-3� for all constructs was cloned, in frame with the UidA gene,into the 4.87-kb vector pGEM4.GUS3 using standard molecular biologytechniques (Sambrook et al., 1989). Bombardments were carried out using acustom-made helium pressure-driven particle inflow gun as previouslydescribed (Schenk et al., 1998). GFP-expressing cells in the bombardedleaves were viewed using an MZ6 stereomicroscope with fluorescence GFPPlus filter module (Leica Microscopy and Scientific Instruments, Bannock-burn, IL), containing a 480/40-nm excitation filter, a 505-nm long passdichromatic beam-splitting mirror, and a 510-nm long pass barrier filter.Luciferase activity was detected using Luciferase Assay Reagent (Promega,Madison, WI) in a Fluoroskan Ascent FL luminometer (Labsystems).
Macroarray Analysis
The consensus amino acid sequence for the DNA-binding domain of theAP2/ERF transcription factor family (YRGV RRPW GKWA AEIR DPKRVWLG TFTA EEAA RAYD AARG KALN FP; Riechmann and Meyerowitz,1998) was used as a query sequence in a BLASTP search of the nonredundantGenBank protein database, limited to Arabidopsis. Subfamily allocation wasdetermined by searching for the conserved linker sequence (E-V—LRR-S—F-RG; Klucher et al., 1996) occurring between the two conserved AP2 domainsof AP2 subfamily members. ClustalW was used to analyze sequence similar-ity within the conserved domain and to produce a guide tree of relatedness.For macroarray construction, genomic DNA was extracted from Arabidopsisecotype Col-0 plants using the CTAB method. The following forward andreverse primers sequences were used for amplification of divergent regions ofAP2/ERF sequences from the Arabidopsis genome: for ERF1 (At3g23240),5�-CGGCGGAGAGAGTTCAAGAGTC-3� and 5�-TCCCACTATTTTCAGAA-GACCCC-3�; for AtERF2 (At5g47220), 5�-TTGGGGAGGTTTGCCATTG-3�and 5�-TCCATCGCCGTAAAGTTCTCAG-3�; for AtERF3 (At1g50640),5�-CCGACCATCAACAACAGTTCCC-3� and 5�-CGACGAATCG-CAATCGCTG-3�; for EREBP4/RAP2.5/AtERF4 (At3g15210), 5�-TC-TACTTTTTGGACCTGATGGGG-3� and 5�-TCGCTTTGGGCACCACAAG-3�; for AtERF5 (At5g47230), 5�-ACGAGGCTTCTCCTGTGGCTAC-3� and5�-GCGAAATCTTCAATGGCGG-3�; for putative EREBP (At2g44970), 5�-CGATAACTGGAGCGACTTGCC-3� and 5�-GGAGAGGTAACGGGAG-GAACG-3�; for TINY isolog (At1g22810), 5�-TCCTTCACTCGTTTCCA-GAACTTC-3� and 5�-GCCGCCCAGATAATCATACACTG-3�; for genesimilar to tobacco EREBP3 (At1g12980), 5�-AGAAATCTTCGCCGTCT-GCTC-3� and 5�-TCTTGATACCCCCACTCGTTTG-3�; for AP9 (At1g80580),5�-AACCCGAAAACACGCATC-3� and 5�-GCTTCACACCACCTTCTT-TGG-3�; for RAP2.10 (At4g36900), 5�-AGGAGGAGTGAACGGTGGTG-3�and 5�-GATGATGATGATGACGATTCCC-3�; for putative AP2 (At1g63030),5�-ATTCTGCTTGGAGGTTGCCG-3� and 5�-CTCCAAAGTGACA-AATCTTC-3�; for putative AP2 (At2g35700), 5�-CATCTCCCACAGTTACG-GAAAC-3� and 5�-AAAAGAGTCCCAGAAGCCATC-3�; for putative AP2(At1g63040), 5�-TCTTACCCCATTCCCCTTTCC-3� and 5�-AGCCACCAT-CATCCCTTTGG-3�; for putative AP2 (At2g46310), 5�-CGAGGGAATCG-CAAATCAGTC-3� and 5�-CGAGAAACAAAGGGTCAGGGG-3�; for AP2transcription factor (At2g40340), 5�-CAAGTTCAGGTTTTGGTCAGGTG-3�and 5�-GCAATCTCCATAGGGTTGAGGC-3�; for putative AP2 (At2g22200),5�-TGAATCCTCTCCCTTCCTCTGTTG-3� and 5�-CATCGCTTCTCGGT-
GACTCATTAG-3�; for putative protein (At4g31060), 5�-AGGCAAC-CAAAAAGTGGCG-3� and 5�-CAACCATTCCGTGTTCTCCATC-3�; and forAPETALA2 (At4g36920), 5�-AACGACGCACCACACCAAAC-3� and 5�-AAAGCCAGAAGCAACACCACC-3�. For normalization purposes, themacroarray filters also contained DNA sequences from the following genesamplified using the following primer pairs: PR-1, 5�-CGGCGACAAGAC-TACCTTGATG-3� and 5�-TGGCACATCCAACCCACTCTG-3�; PDF1.2,5�-CGCACCGGCAATGGTGGAAG-3� and 5�-CACACGATTTAGC-ACCAAAG-3�; BGL-2, 5�-GTAAAACGACGGCCAGT-3� and 5�-GGAAA-CAGCTATGACCATG-3�; UidA, 5�-ACCCTTACGCTGAAGAGATGC-3�,and 5�-AACGTATCCACGCCGTATTCG-3�; Actin, 5�-GGATGCTTATGTTG-GCGATG-3� and 5�-GCTGGTTTTGGCTGTCTCG-3�; and Tubulin �-8 chain,5�-GTAAAACGACGGCCAGT-3� and 5�-GGAAACAGCTATGACCATG-3�.Macroarray filters were prepared as follows. Probe DNA (40 ng spot�1) wasdenatured and transferred in duplicate to a Hybond N� membrane (Am-ersham Biosciences, Piscataway, NJ) using the Bio-Dot microfiltration ap-paratus (Bio-Rad Laboratories), according to the manufacturer’s instruc-tions. Total cDNA synthesis was performed using 750 ng of purifiedpoly(A�) RNA using Superscript II RT (Invitrogen, Carlsbad, CA) as rec-ommended by the manufacturer. The total cDNA prepared was labeledeither using the Megaprime DNA labeling system (Amersham Biosciences)or the Strip-EZ system (Ambion, Austin TX) as instructed by the manufac-turer. Blots were hybridized as previously described (Mitter et al., 1998) andwere visualized by exposure to a PhosporImaging system, and data wereprocessed with the ImageQuant analysis software (Molecular Dynamics,Sunnyvale, CA). For each hybridization blot, a reference figure was calcu-lated by averaging the hybridization signal from the duplicate �-tubulinspots minus the background. Data for probes from all other genes weredetermined similarly and then divided by the �-tubulin reference for thatblot to allow comparison of relative expression levels of the different genes.
Northern-Blot Analysis
Arabidopsis ecotype Col-0 plants were grown under short-day conditions(8 of h light, 16 h of dark) for 3 weeks and treated with MeJA, ethylene, ornothing (controls), as described above for GUS activity assays. Total cellularRNA was extracted from the aerial portion of the plants using the guanidinethiocyanate method and a 20-�g sample of total RNA was run on the gel andblotted onto a Hybond N� membrane (Amersham Biosciences). Probes wereamplified by PCR and purified using the Qiaquick PCR purification kit(Qiagen USA, Valencia, CA). The ERF probes were amplified using the primersequences given above. The PDF1.2 probe was amplified from Clone 10a(Manners et al., 1998) using the primers 5�-CGCACCGGCAATGG-TGGAAG-3� and 5�-CACACGATTTAGCACCAAAG-3�. The tubulin �-8 chainprobe was amplified from EST clone number 108K21T7 (GenBank accessionno. T41808; Arabidopsis Biological Resource Center, Columbus, OH) usingthe M13 forward and M13 reverse primers. The hybridizations and analysis ofhybridization signals were done as described for macroarray hybridizations.
Real-Time Quantitative RT-PCR Analysis
Total cellular RNA was extracted from the aerial portion of the plantsusing Plant RNA extraction kit (Bio-Rad Laboratories). First-strand cDNAwas synthesized using 2 �g of total RNA, Superscript II RT (Invitrogen), andrandom hexamer primers to a total volume of 10 �L. The reaction mix wasincubated at 42°C for 50 min, before heat inactivation of the enzyme at 70°Cfor 15 min. The cDNA template was then diluted by adding 240 �L of milli-Qwater, to give a final concentration of 8 ng �L�1 for use in real-time PCRassays. A second 2-�g aliquot of each RNA sample was subjected to the sameconditions, without the addition of RT (the minus RT controls). Real-time PCRamplification and detection was carried out in an ABI model 7700 SequenceDetection System (Applied Biosystems, Foster City, CA). Reactions were infinal volumes of 25 �L and contained 40 ng of cDNA template and 5 �L ofprimer mix containing a 1 �m solution of each of the AtERF2 primers,5�-TGAGGTTAATTCCGGTGAACC-3� and 5�-TCAACTTCCCGTTTTCA-GACGA-3�; ERF1 primers, 5�-CGAGAAGCTCGGGTGGTAGT-3� and5�-GCCGTGCATCCTTTTCC-3�; PDF1.2 (At5g44420) primers, 5�-TTGCT-GCTTTCGACGCA-3� and 5�-TGTCCCACTTGGCTTCTCG-3�; Thi2.1(L41244.1) primers (Epple et al., 1995), 5�-CTCAGCTGATGCTACCAA-TGAGC-3� and 5�-GCTCCATTCACAATTTCACTTGC-3� ; or PR4(At3g12500) primers, 5�-ATCAGCGCTGCAAAGTCCTTC-3� and 5�-
Brown et al.
1030 Plant Physiol. Vol. 132, 2003 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

GTGCTGTAGCCCATCCACCTG-3�; and 12.5 �L of SYBR Green PCR mas-ter mix (Applied Biosystems). The thermal cycling conditions were: 10 minat 95°C, 45 cycles of 15 s at 95°C and 1 min at 60°C. Duplicate SYBR Greenassays for each gene target were performed on both plus and minus RTsamples. No-template controls for each primer pair were included in eachrun. Expression detected from three �-actin genes of Arabidopsis, �-actin-2(At3g18780), �-actin-7 (At5g09810), and �-actin-8 (At1g49240) used thefollowing mixture of primers (R. Simpson, personal communication; auniversal actin forward primer 5�-AGTGGTCGTACAACCGGTATTGT-3�and specific reverse primers 5�-GATGGCATGGAGGAAGAGAGAAAC-3�, 5�-GAGGAAGAGCATTCCCCTCGTA-3�, and 5�-GAGGATAGCATGT-GGAACTGAGAA-3�, respectively) were used as combined internalstandards to normalize small differences in template amounts. Real-timePCR results were captured and analyzed using the Sequence DetectorSoftware (SDS v1.7, Applied Biosystems) and data analysis proceduresdescribed in the User Bulletin 2 for the ABI model 7700 Sequence DetectionSystem.
Construction of the 35S-AtERF2 Binary Vector andPlant Transformation
The AtERF2 (AT5g47220) gene of Arabidopsis was amplified from theCol-0 ecotype using the primers 5�-CTGAAAATGTACGGACAGTGC-3�and 5�-AACTTATGAACCAATAACTC-3�. The amplification product wastreated with 1 unit of polynucleotide kinase (Roche Diagnostics, Mannheim,Germany), ligated into the binary vector pKEN cut by HindIII, filled in byKlenow DNA polymerase (Roche Diagnostics), and dephosphorylated.pKEN was constructed by cloning a SacI/XhoI fragment containing thedouble 35S promoter/terminator cassette from the plasmid pJIT163 (Hellenset al., 2000) into SacI/XhoI-digested pGreen229 (Hellens et al., 2000). Therecombinant plasmid was electroporated into competent A. tumefaciens(strain AGL-1) cells previously transformed with pSoup plasmid (Hellens etal., 2000). Arabidopsis Col-0 plants were then transformed using the floraldip transformation procedure (Clough and Bent, 1998), and the transfor-mants were selected based on their resistance to Basta.
Statistical Analysis of Data
The JMP IN Statistics package v3.2.6 (SAS Institute Inc., Cary, NC) wasused for all statistical analyses. The significance of differences betweentransgenic plants carrying the various promoter deletion constructs andindependent transgenic lines within these constructs was determined bynested ANOVA. The non-parametrical Wilcoxen-Mann-Whitney test wasused to determine statistical significance of differences between constructsin bombardment experiments. Biological significance was assigned at P �0.05.
ACKNOWLEDGMENTS
We thank Dr. John Turner and the Arabidopsis Stock Center at Ohio StateUniversity for cev1 and cpr5 seeds, respectively; Jonathan Anderson for cpr5cDNA samples; Dr. Peer Schenk for advice in plasmid construction andsome of the cDNA samples; Anca Rusu for GUS activity analyses of some ofthe promoter-reporter lines; Dr. Philip Mullineaux for the pGreen0229,pSoup, and pJIT163 plasmids; Dr. Robert Birch for the pSP35Slucnos plas-mid; Dr. Jimmy Botella for the pAOV plasmid; Dr. Jim Haseloff for thepBINm-GFP5-ER plasmid; and Dr. Willem Broekaert for his advice on planttreatments and collaboration.
Received November 14, 2002; returned for revision January 4, 2003; acceptedMarch 5, 2003.
LITERATURE CITED
Berrocal-Lobo M, Molina A, Solano R (2002) Constitutive expression ofETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance toseveral necrotrophic fungi. Plant J 29: 23–32
Bowling SA, Clarke JD, Liu YD, Klessig DF, Dong XN (1997) The cpr5mutant of Arabidopsis expresses both NPR1-dependent and NPR1-independent resistance. Plant Cell 9: 1573–1584
Buttner M, Singh KB (1997) Arabidopsis thaliana ethylene-responsive ele-ment binding protein (AtEBP), an ethylene-inducible, GCC-box DNA-binding protein interacts with an ocs element binding protein. Proc NatlAcad Sci USA 94: 5961–5966
Charrier B, Champion A, Henry Y, Kreis M (2002) Expression profiling ofthe whole Arabidopsis shaggy-like kinase multigene family by real-timereverse transcriptase-polymerase chain reaction. Plant Physiol 130:577–590
Cheong YH, Chang H-S, Gupta R, Wang X, Tong Z, Luan S (2002) Tran-scriptional profiling reveals novel interactions between wounding,pathogen, abiotic stress, and hormonal responses in Arabidopsis. PlantPhysiol 129: 661–667
Clough SJ, Bent AF (1998) Floral dip: a simplified method forAgrobacterium-mediated transformation. Plant J 16: 735–743
Dong X (1998) SA, JA, ethylene, and disease resistance in plants. Curr OpinPlant Biol 1: 316–323
Ellis C, Turner JG (2001) The Arabidopsis mutant cev1 has constitutivelyactive jasmonate and ethylene signal pathways and enhanced resistanceto pathogens. Plant Cell 13: 1025–1033
Epple P, Apel K, Bohlmann H (1995) An Arabidopsis thaliana thionin gene isinducible via a signal transduction pathway different from that forpathogenesis-related proteins. Plant Physiol 109: 813–820
Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M (2000) Arabi-dopsis ethylene-responsive element binding factors act as transcriptionalactivators or repressors of GCC-box-mediated gene expression. Plant Cell12: 393–404
Grimmig B, Gonzalez-Perez MN, Leubner-Metzger G, Vogeli-Lange R,Meins F, Hain R, Penuelas J, Heidenreich B, Langebartels C, Ernst D etal. (2003) Ozone-induced gene expression occurs via ethylene-dependentand -independent signalling. Plant Mol Biol 51: 599–607
Gu YQ, Wildermuth MC, Chakravarthy S, Loh YT, Yang C, He X, Han Y,Martin GB (2002) Tomato transcription factors Pti4, Pti5, and Pti6 acti-vate defense responses when expressed in Arabidopsis. Plant Cell 14:817–831
Gu YQ, Yang CM, Thara VK, Zhou JM, Martin GB (2000) Pti4 is inducedby ethylene and salicylic acid, and its product is phosphorylated by thePto kinase. Plant Cell 12: 771–785
Hao D, Ohme-Takagi M, Sarai A (1998) Unique mode of GCC-box recog-nition by the DNA-binding domain of ethylene-responsive element-binding factor (ERF domain) in plant. J Biol Chem 273: 26857–26861
Haseloff J, Siemering KR, Prasher DC, Hodge S (1997) Removal of acryptic intron and subcellular localization of green fluorescent proteinare required to mark transgenic Arabidopsis plants brightly. Proc NatlAcad Sci USA 94: 2122–2127
Hellens RP, Edwards EA, Leyland NR, Bean S, Mullineaux PM (2000)pGreen: a versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol Biol 42: 819–832
Hwang YS, Karrer EE, Thomas BR, Chen L, Rodriguez RL (1998) Threecis-elements required for rice alpha-amylase Amy3D expression duringsugar starvation. Plant Mol Biol 36: 331–341
Izawa T, Foster R, Chua NH (1993) Plant bZIP protein DNA bindingspecificity. J Mol Biol 230: 1131–1144
Jefferson RA (1987) Assaying chimeric genes in plants: the GUS gene-fusionsystem. Plant Mol Biol Rep 5: 387–405
Kagaya Y, Ohmiya K, Hattori T (1999) RAV1, a novel DNA-binding protein,binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res 27:470–478
Kim SR, Choi JL, Costa MA, An GH (1992) Identification of G-box sequenceas an essential element for methyl jasmonate response of potato protein-ase inhibitor II promoter. Plant Physiol 99: 627–631
Klucher KM, Chow H, Reiser L, Fischer RL (1996) The AINTEGUMENTAgene of Arabidopsis required for ovule and female gametophyte devel-opment is related to the floral homeotic gene APETALA2. Plant Cell 8:137–153
Lorenzo O, Piqueras R, Sanchez-Serrano JJ, Solano R (2003) ETHYLENERESPONSE FACTOR1 integrates signals from ethylene and jasmonatepathways in plant defense. Plant Cell 15: 165–178
Manners JM, Penninckx I, Vermaere K, Kazan K, Brown RL, Morgan A,Maclean DJ, Curtis MD, Cammue BPA, Broekaert WF (1998) The pro-moter of the plant defensin gene PDF1.2 from Arabidopsis is systemicallyactivated by fungal pathogens and responds to methyl jasmonate but notto salicylic acid. Plant Mol Biol 38: 1071–1080
GCC-Box and Jasmonate-Mediated Activation of the PDF1.2 Gene
Plant Physiol. Vol. 132, 2003 1031 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.

Mason HS, Dewald DB, Mullet JE (1993) Identification of a methyljasmonate-responsive domain in the soybean vspB promoter. Plant Cell 5:241–251
Mazarei M, Puthoff DP, Hart JK, Rodermel SR, Baum TJ (2002) Identifi-cation and characterization of a soybean ethylene-responsive element-binding protein gene whose mRNA expression changes during soybeancyst nematode infection. Mol Plant-Microbe Interact 15: 577–586
Medina J, Bargues M, Terol J, Perez-Alonso M, Salinas J (1999) TheArabidopsis CBF gene family is composed of three genes encoding AP2domain-containing proteins whose expression is regulated by low tem-perature but not by abscisic acid or dehydration. Plant Physiol 119:463–469
Memelink J, Verpoorte R, Kijne JW (2001) ORCAnization of jasmonate-responsive gene expression in alkaloid metabolism. Trends Plant Sci 6:212–219
Menke FLH, Champion A, Kijne JW, Memelink J (1999a) A noveljasmonate- and elicitor-responsive element in the periwinkle secondarymetabolite biosynthetic gene Str interacts with a jasmonate- and elicitor-inducible AP2-domain transcription factor, ORCA2. EMBO J 18:4455–4463
Menke FLH, Parchmann S, Mueller MJ, Kijne JW, Memelink J (1999b)Involvement of the octadecanoid pathway and protein phosphorylationin fungal elicitor-induced expression of terpenoid indole alkaloid biosyn-thetic genes in Catharanthus roseus. Plant Physiol 119: 1289–1296
Mitter N, Kazan K, Way HM, Broekaert WF, Manners JM (1998) Systemicinduction of an Arabidopsis plant defensin gene promoter by tobaccomosaic virus and jasmonic acid in transgenic tobacco. Plant Sci 136:169–180
Mylne J, Botella J (1998) Binary vectors for sense and antisense expressionof Arabidopsis ESTs. Plant Mol Biol Rep 16: 257–262
Nishiuchi T, Suzuki K, Kitajima S, Sato F, Shinshi H (2002) Woundingactivates immediate early transcription of genes for ERFs in tobaccoplants. Plant Mol Biol 49: 473–482
Ohme-Takagi M, Shinshi H (1995) Ethylene-inducible DNA binding pro-teins that interact with an ethylene-responsive element. Plant Cell 7:173–182
Ohta M, Matsui K, Hiratsu K, Shinshi H, Ohme-Takagi M (2001) Repres-sion domains of Class II ERF transcriptional repressors share an essentialmotif for active repression. Plant Cell 13: 1959–1968
Ohta M, Ohme-Takagi M, Shinshi H (2000) Three ethylene-responsivetranscription factors in tobacco with distinct transactivation functions.Plant J 22: 29–38
Onate-Sanchez L, Singh KB (2002) Identification of Arabidopsis ethylene-responsive element binding factors with distinct induction kinetics afterpathogen infection. Plant Physiol 128: 1313–1322
Park JM, Park CJ, Lee SB, Ham BK, Shin R, Paek KH (2001) Over-expression of the Tobacco Tsi1 gene encoding an EREBP/AP2-type tran-scription factor enhances resistance against pathogen attack and osmoticstress in tobacco. Plant Cell 13: 1035–1046
Penninckx IAMA, Eggermont K, Terras FRG, Thomma BPHJ, De Sam-blanx GW, Buchala A, Metraux JP, Manners JM, Broekaert WF (1996)Pathogen-induced systemic activation of a plant defensin gene in Arabi-dopsis follows a salicylic acid-independent pathway. Plant Cell 8:2309–2323
Penninckx IAMA, Thomma BPHJ, Buchala A, Metraux JP, Broekaert WF(1998) Concomitant activation of jasmonate and ethylene response path-ways is required for induction of a plant defensin gene in Arabidopsis.Plant Cell 10: 2103–2113
Raghothama KG, Maggio A, Narasimhan ML, Kononowicz AK, Wang G,D’Urzo MP, Hasegawa PM, Bressan RA (1997) Tissue-specific activationof the osmotin gene by ABA, C2H4 and NaCl involves the same promoterregion. Plant Mol Biol 34: 393–402
Reymond P, Farmer EE (1998) Jasmonate and salicylate as global signals fordefense gene expression. Curr Opin Plant Biol 1: 404–411
Riechmann JL, Meyerowitz EM (1998) The AP2/EREBP family of planttranscription factors. Biol Chem 379: 633–646
Rouster J, Leah R, Mundy J, Mills CV (1997) Identification of a methyljasmonate-responsive region in the promoter of a lipoxygenase 1 geneexpressed in barley grain. Plant J 11: 513–523
Sakuma Y, Liu Q, Dubouzet JG, Abe H, Shinozaki K, Yamaguchi-Shinozaki K (2002) DNA-binding specificity of the ERF/AP2 domain ofArabidopsis DREBs, transcription factors involved in dehydration- andcold-inducible gene expression. Biochem Biophys Res Commun 290:998–1009
Sambrook J, Fritsch EF, Maniatis TA (1989) Molecular Cloning: A Labora-tory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NY
Schenk PM, Elliott AR, Manners JM (1998) Assessment of transient geneexpression in plant tissues using the green fluorescent protein as areference. Plant Mol Biol Rep 16: 313–322
Schenk PM, Kazan K, Wilson I, Anderson JP, Richmond T, Somerville SC,Manners JM (2000) Coordinated plant defense responses in Arabidopsisrevealed by microarray analysis. Proc Natl Acad Sci USA 97: 11655–11660
Sessa G, Meller Y, Fluhr R (1995) A GCC element and a G-box motifparticipate in ethylene-induced expression of the PRB-1b gene. Plant MolBiol 28: 145–153
Shah J, Klessig DF (1996) Identification of a salicyclic acid-responsiveelement in the promoter of the tobacco pathogenesis-related beta-1,3-glucanase gene, PR-2d. Plant J 10: 1089–1101
Shinshi H, Usami S, Ohme-Takagi M (1995) Identification of an ethylene-responsive region in the promoter of a tobacco class I chitinase gene.Plant Mol Biol 27: 923–932
Solano R, Stepanova A, Chao Q, Ecker JR (1998) Nuclear events in ethylenesignaling: A transcriptional cascade mediated by ethylene-insensitive3and ethylene-response-factor1. Genes Dev 12: 3703–3714
Staswick PE, Yuen GY, Lehman CC (1998) Jasmonate signaling mutants ofArabidopsis are susceptible to the soil fungus Pythium irregulare. Plant J15: 747–754
Thara VK, Tang X, Gu YQ, Martin GB, Zhou JM (1999) Pseudomonassyringae pv tomato induces the expression of tomato EREBP-like genesPti4 and Pti5 independent of ethylene, salicylate and jasmonate. Plant J20: 475–483
Thomma BP, Eggermont K, Tierens KF, Broekaert WF (1999) Requirementof functional ethylene-insensitive 2 gene for efficient resistance of Ara-bidopsis to infection by Botrytis cinerea. Plant Physiol 121: 1093–1102
Thomma BP, Penninckx IA, Broekaert WF, Cammue BP (2001) The com-plexity of disease signaling in Arabidopsis. Curr Opin Immunol 13: 63–68
Thomma BPHJ, Cammue BPA, Thevissen K (2002) Plant defensins. Planta216: 193–202
Thomma BPHJ, Eggermont K, Penninckx IAMA, Mauch-Mani B, Vogel-sang R, Cammue BPA, Broekaert WF (1998) Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Ara-bidopsis are essential for resistance to distinct microbial pathogens. ProcNatl Acad Sci USA 95: 15107–15111
Valvekens D, Van Montagu M, Van Lijsebettens M (1988) Agrobacteriumtumefaciens-mediated transformation of Arabidopsis thaliana root explantsby using kanamycin selection. Proc Natl Acad Sci USA 85: 5536–5540
van der Fits L, Memelink J (2000) ORCA3, a jasmonate-responsive tran-scriptional regulator of plant primary and secondary metabolism. Science289: 295–297
van der Fits L, Memelink J (2001) The jasmonate-inducible AP2/ERF-domain transcription factor ORCA3 activates gene expression via inter-action with a jasmonate-responsive promoter element. Plant J 25: 43–53
Vijayan P, Shockey J, Levesque CA, Cook RJ, Browse J (1998) A role forjasmonate in pathogen defense of Arabidopsis. Proc Natl Acad Sci USA95: 7209–7214
Wu K, Tian L, Hollingworth J, Brown DCW, Miki B (2002) Functionalanalysis of tomato Pti4 in Arabidopsis. Plant Physiol 128: 30–37
Yamamoto S, Suzuki K, Shinshi H (1999) Elicitor-responsive, ethylene-independent activation of GCC-box-mediated transcription that is regu-lated by both protein phosphorylation and dephosphorylation in cul-tured tobacco cells. Plant J 20: 571–579
Zhou D (1999) Regulatory mechanism of plant gene transcription by GT-elements and GT-factors. Trends Plant Sci 4: 210–214
Zhou JM, Tang XY, Martin GB (1997) The Pto kinase conferring resistanceto tomato bacterial speck disease interacts with proteins that bind acis-element of pathogenesis-related genes. EMBO J 16: 3207–3218
Brown et al.
1032 Plant Physiol. Vol. 132, 2003 www.plant.org on August 15, 2015 - Published by www.plantphysiol.orgDownloaded from Copyright © 2003 American Society of Plant Biologists. All rights reserved.




















