
A Realidade como um Modelo Interno para Regulação e Controle
44
1 A realidade como um modelo interno para regulação e controle 1 Ao abrirmos os olhos, desde o início de nossas vidas, deparamo-nos com a realidade. Não há esforço em percebê-la como tal, nem lembranças de percebê-la diferentemente. A realidade se nos apresenta independentemente de nossa vontade: ela se nos impõe, diria o filósofo realista. Nós acreditamos na realidade, buscamos nos relacionar com ela da melhor forma, e, às vezes, entender o sucesso dessa relação. Parece razoável supor que parte do sucesso dessa relação se dá em virtude do uso de dispositivos tais como linguagem, teorias, tecnologia, os quais conseguem entrelaçar de forma tão harmônica diferentes pessoas e a própria realidade. Assim, um dos problemas centrais em filosofia consiste em entender o papel que esses dispositivos cumprem nessa relação. Parece também razoável afirmar que a visão intuitiva acerca dessa relação, a qual espreita diferentes tratamentos filosóficos do problema, seria a de que esses dispositivos contribuem para tal sucesso, pois, representam, ou referem, ou denotam a realidade – onde o grau de acuidade da conexão justifica o maior grau de sucesso. Assim, admitida uma visão de realidade, de um lado, a tarefa seria a de desvendar os liames dessa conexão. No presente texto, argumentaremos que nossa noção comum de realidade consiste num modelo interno elaborado pelo Sistema Nervoso Central para melhorar o processo de regulação e controle de nosso organismo. Esse modelo não consistiria numa representação fiel de nosso entorno, mas apenas num constructo que atende as necessidades de regulação. Entre as consequências que imediatamente obtemos dessa perspectiva, teremos que ao confiarmos em nossa ‘realidade’ frequentemente acreditamos em algo literalmente falso, o que nos conduz a suspeita de que a maneira pela qual esses mecanismos contribuem para nosso sucesso como espécie deve ser distinta de qualquer coisa que chamaríamos de semântica. Nossa argumentação se desenvolverá desde uma perspectiva materialista, onde qualquer evento que adquira significado para nossas vidas deve ser visto, em princípio, como um estado funcional neuronal – ou seja, como um parâmetro de ordem neuronal atendendo a alguma função no organismo. Assim, nosso primeiro esforço será o de compreender o tipo de sistema que caracteriza o Sistema Nervoso Central e seu papel a cumprir no organismo biológico. Para tal fim, e tão comente por razões pragmáticas, a 1 Márlon Henrique dos Santos Teixeira 2 Naturalmente o termo máquina aqui não deve conduzir o leitor a qualquer preconceito cartesiano, visto que esse
Transcript of A Realidade como um Modelo Interno para Regulação e Controle

1
A realidade como um modelo interno para regulação e controle1
Ao abrirmos os olhos, desde o início de nossas vidas, deparamo-nos com a
realidade. Não há esforço em percebê-la como tal, nem lembranças de percebê-la
diferentemente. A realidade se nos apresenta independentemente de nossa vontade: ela
se nos impõe, diria o filósofo realista. Nós acreditamos na realidade, buscamos nos
relacionar com ela da melhor forma, e, às vezes, entender o sucesso dessa relação.
Parece razoável supor que parte do sucesso dessa relação se dá em virtude do uso de
dispositivos tais como linguagem, teorias, tecnologia, os quais conseguem entrelaçar de
forma tão harmônica diferentes pessoas e a própria realidade. Assim, um dos problemas
centrais em filosofia consiste em entender o papel que esses dispositivos cumprem
nessa relação. Parece também razoável afirmar que a visão intuitiva acerca dessa
relação, a qual espreita diferentes tratamentos filosóficos do problema, seria a de que
esses dispositivos contribuem para tal sucesso, pois, representam, ou referem, ou
denotam a realidade – onde o grau de acuidade da conexão justifica o maior grau de
sucesso. Assim, admitida uma visão de realidade, de um lado, a tarefa seria a de
desvendar os liames dessa conexão.
No presente texto, argumentaremos que nossa noção comum de realidade
consiste num modelo interno elaborado pelo Sistema Nervoso Central para melhorar o
processo de regulação e controle de nosso organismo. Esse modelo não consistiria numa
representação fiel de nosso entorno, mas apenas num constructo que atende as
necessidades de regulação. Entre as consequências que imediatamente obtemos dessa
perspectiva, teremos que ao confiarmos em nossa ‘realidade’ frequentemente
acreditamos em algo literalmente falso, o que nos conduz a suspeita de que a maneira
pela qual esses mecanismos contribuem para nosso sucesso como espécie deve ser
distinta de qualquer coisa que chamaríamos de semântica.
Nossa argumentação se desenvolverá desde uma perspectiva materialista, onde
qualquer evento que adquira significado para nossas vidas deve ser visto, em princípio,
como um estado funcional neuronal – ou seja, como um parâmetro de ordem neuronal
atendendo a alguma função no organismo. Assim, nosso primeiro esforço será o de
compreender o tipo de sistema que caracteriza o Sistema Nervoso Central e seu papel a
cumprir no organismo biológico. Para tal fim, e tão comente por razões pragmáticas, a
1 Márlon Henrique dos Santos Teixeira
2 Naturalmente o termo máquina aqui não deve conduzir o leitor a qualquer preconceito cartesiano, visto que esse

2
Cibernética e Teoria da Informação serão ferramentas de uso comum, enquanto que
neurociências, neuropsicologia e psicologia cognitiva servirão de suporte para as
hipóteses e resultados levantados. Porque essa perspectiva privilegiará antes a interação
do sistema com o entorno, do que as propriedades intrínsecas ao sistema, deixe-nos
chamá-la de interacionista.
1. O Sistema Nervoso Central como um Sistema dinâmico
1.1.Cérebro ou Sistema Nervoso Central?
Quando se fala sobre o pensamento, desde uma perspectiva materialista,
frequentemente nossa fala versa sobre o cérebro, o qual é o centro de processamento do
Sistema Nervoso Central (SNC). Porém, na medida em que se fala do cérebro sem
mencionar o SNC, corre-se o risco de pensar que poderíamos entender os eventos
cerebrais fora do contexto do SNC. O cérebro, ou espaço interneural, consiste em um
conjunto de subsistemas do SNC, de tal modo que os eventos ocorridos em seu espaço
devem ser entendidos com relação ao papel desempenhado no sistema como um todo
(MAINZER, 2007). A designação ‘espaço interneural’ deriva do fato de que seus
neurônios têm a função de transmitir informação proveniente dos neurônios aferentes –
órgãos sensoriais – até os neurônios eferentes – que resultam em contrações musculares
(LLINÁS, 2001). O SNC é compreendido pelo cérebro, mais a espinha dorsal, os quais
são bilateralmente simétricos [Figura 1]. A espinha dorsal recebe informação sensorial
da pele através de um conjunto de longos axônios, chamados também de nervos
periféricos, e envia comando motor para os músculos (KANDEL, 2012).
Figura 1 - Sistema Nervoso Central

3
Assim, será esse o sistema sobre o qual falaremos.
1.2. O Sistema Nervoso Central como uma Máquina com Input
Na perspectiva da cibernética, o SNC é concebido como um sistema dinâmico.
Intuitivamente, um sistema dinâmico pode ser entendido como qualquer grupo de
elementos, exibindo algum tipo de relação (LERNER, 1975; ASHBY, 1956). De um
ponto de vista teórico, isso poder ser expresso como qualquer conjunto de variáveis,
onde cada variável descreve um aspecto, por assim dizer, dos elementos. Os valores que
as variáveis do sistema assumem dependem inteiramente dos propósitos inerentes à
investigação. No caso do SNC, podemos descrevê-lo desde uma perspectiva que
considere suas moléculas, ou membrana potencial, ou sinapses, ou neurônios, ou
núcleos, ou circuitos, ou redes, ou mapas, ou sistemas, e, logo, o sistema nervoso central
como um todo. No entanto, para nossos propósitos é preferível introduzirmos uma
definição mais precisa. Então, deixe-nos assumir que as variáveis de nosso sistema
podem ser entendidas como um vetor cujas componentes sejam numéricas, então
podemos definir um sistema da seguinte forma:
[Def. 1.1] Um sistema dinâmico consiste em um espaço de fase e uma transformação
(operador) sobre esse espaço;
o espaço de fase consiste num espaço n-dimensional no qual os operandos e as
transformandas pode ser representados por pontos (ASHBY, 1956).
Dito isso, devemos nos perguntar: que tipo de sistema dinâmico é o SNC?
Segundo Prigogine (1961), de acordo com a termodinâmica os sistemas podem ser
classificados em três categorias, a saber: sistemas isolados como aqueles que não
podem trocar nem energia e nem matéria com entorno; sistemas fechados como aqueles
que trocam energia, mas não trocam matérias; e os sistemas abertos que trocam tanto
energia como matéria com o entorno. O SNC evidentemente será compreendido como
uma estrutura dissipativa – sistema aberto – fora de equilíbrio termodinâmico em um
estado de entropia reduzida (MAINZER, 2007, p. 98; PRIGOGINE & NICOLIS, 1977).
Do ponto de vista da cibernética, isso seria equivalente a dizer que o comportamento do
sistema é dependente de sua relação com o entorno, ou variedade exterior que afeta o
sistema. Para ilustrar esse ponto, lembre-se que perdemos consciência após alguns
segundos sem ar – troca de matéria com o entorno – ou que nosso comportamento não é
totalmente aleatório, mas é, em parte, uma função dos estímulos exteriores que nos
4
afetam – como veremos a seguir é muito importante notar que o SNC é capaz de gerar
estados funcionais totalmente independentes dos estímulos exteriores.
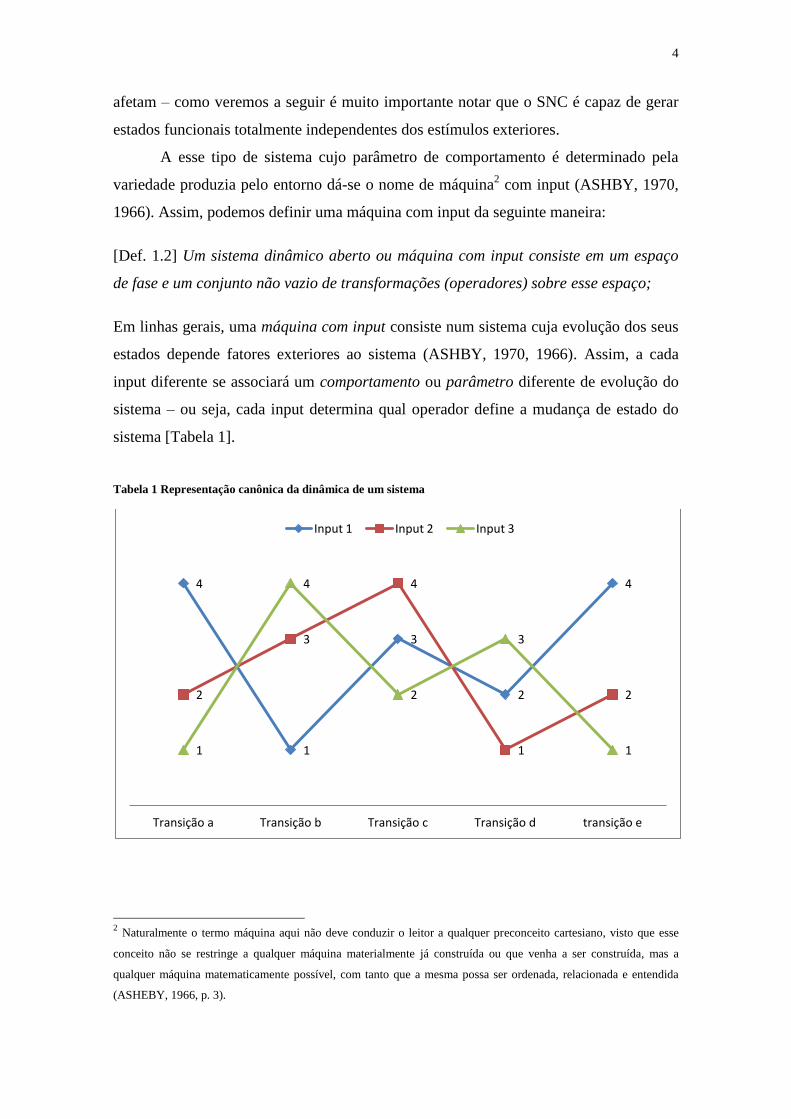
A esse tipo de sistema cujo parâmetro de comportamento é determinado pela
variedade produzia pelo entorno dá-se o nome de máquina2 com input (ASHBY, 1970,
1966). Assim, podemos definir uma máquina com input da seguinte maneira:
[Def. 1.2] Um sistema dinâmico aberto ou máquina com input consiste em um espaço
de fase e um conjunto não vazio de transformações (operadores) sobre esse espaço;
Em linhas gerais, uma máquina com input consiste num sistema cuja evolução dos seus
estados depende fatores exteriores ao sistema (ASHBY, 1970, 1966). Assim, a cada
input diferente se associará um comportamento ou parâmetro diferente de evolução do
sistema – ou seja, cada input determina qual operador define a mudança de estado do
sistema [Tabela 1].
Tabela 1 Representação canônica da dinâmica de um sistema
2 Naturalmente o termo máquina aqui não deve conduzir o leitor a qualquer preconceito cartesiano, visto que esse
conceito não se restringe a qualquer máquina materialmente já construída ou que venha a ser construída, mas a
qualquer máquina matematicamente possível, com tanto que a mesma possa ser ordenada, relacionada e entendida
(ASHEBY, 1966, p. 3).
4
1
3
2
4
2
3
4
1
2
1
4
2
3
1
Transição a Transição b Transição c Transição d transição e
Input 1 Input 2 Input 3

5
O input seleciona, a cada instante t, uma transforma do conjunto de transformadas, a
qual será responsável pela transição de estado do sistema. Na tabela 1, os numerais 1, 2,
3 e 4 representam os estados iniciais possíveis e os numerais abaixo representam os
estados de transição possíveis do sistema relativos aos inputs A, B e C. Cada linha
colorida no Gráfico 1 representa um comportamento distinto do sistema, relativo a cada
input diferente, sendo o conjunto de seus comportamentos possíveis chamado de
representação canônica3. Obviamente, um dos objetivos principais ao examinarmos um
sistema será a determinação de sua representação canônica.
No entanto, os casos em que podemos vislumbrar a representação canônica de
uma máquina prontamente são raros – se de fato existem. O que ocorre, em geral, é que
os sistemas se apresentam como caixas pretas. Em particular, o SNC consiste em um
caso clássico de uma Caixa Preta (ASHBY, 1970; MAINZER, 2007).
1.3. O SNC como uma Caixa Preta: em busca da estrutura da máquina
A teoria da Caixa Preta consiste numa estratégia para lidarmos com sistemas
cuja dinâmica não nos é acessível diretamente; ou seja, com sistemas que se apresentam
propriamente como uma caixa fechada. Essa teoria vem a ser muito mais útil do que a
primeira vista pode parecer, pois a maioria dos sistemas que nos deparamos, sobretudo
na investigação científica, apresenta-se justamente como tal. O SNC é um exemplo
típico de tais sistemas dinâmicos, onde não se pode acompanhar diretamente a transição
dos estados do sistema. Assim, uma abordagem comportamental vem a ser mais
apropriada, uma vez que as únicas coisas que se podem estivar com certa segurança
vêm a ser o conjunto dos inputs e outputs do sistema.
Destarte, a primeira tarefa consiste, basicamente, em precisar os conjuntos de
inputs e de outputs. Por input se quer dizer qualquer evento externo ao sistema que o
modifique de alguma maneira – que cause uma transição em algum de seus estados. E
3 Note que a noção de espaço de fase e representação canônica são sinônimos aqui. Porém, a noção de
espaço de fase é definida em termos das noções de espaço vetorial ou espaço topológico, assim, a noção
de representação canônica consiste numa abstração dessas noções.
↓ 1 2 3 4
InputA 3 4 2 1
InputB 2 3 4 1
InputC 4 3 1 2

6
por output se quer dizer qualquer mudança produzida no entorno pelo sistema
(ROSENBLUETH et al, 1943; ASHBY, 1970). Uma vez fixando esses dois conjuntos,
submete-se o sistema a repetidos distúrbios de tal sorte que se possa estabelecer uma
relação – determinada ou estocástica – entre cada input e cada output. Onde distúrbio
deve ser entendido como a exposição do sistema a diferentes inputs, com vistas a
vislumbrar justamente as diferentes transições de estado.
Uma vez tendo submetido o SNC a um processo de distúrbios, de um ponto de
vista fisiológico e anatômico, o que se verifica como resultado é exclusivamente
motricidade – aparte dos componentes glandulares e neuro-humorais (KANDEL, 2012;
FUSTER, 2006; LLINAS, 2001; ARBIB, 1981; SPERRY, 1952). Em outras palavras, a
motricidade4é única forma por meio da qual o sistema é capaz de afetar seu entorno. Um
dos primeiros registros dessa perspectiva de entendimento sobre o funcionamento do
SNC se encontra em Neurology and the Mind-Brain problem (SPERRY, 1952).
Segundo Sperry, “o inteiro output de nossa máquina de pensamento consiste em nada
mais do que padrões de coordenação motora, [...] a única saída de energia e o único
meio de expressão são pelo canal motor” (grifos do autor) (1952, p. 296-298). Do ponto
de vista psicológico, também se pode verificar um privilégio da ação em detrimento do
pensamento. Estudos acerca de como o SNC reage com relação a distúrbios repentinos –
tal como rebater rapidamente uma bola, frear o carro para evitar um choque, desviar de
um obstáculo, ou fechar os olhos para evitar a entrada de um inseto no olho –
mostraram que o sistema sensorial é estimulado com um atraso de 100ms em detrimento
do sistema motor (EAGLEMANN, 2012; LLINÁS, 2001). Do ponto de vista
filogenético, nota-se que, descendo na escala evolutiva, a atividade puramente mental
vem a ser cada vez mais insignificante, quando comparada com atividade motora –
obviamente o que se toma por ‘mental’ aqui consiste em ‘atividade neural sem uma
corelação motora imediata’. Isso não seria de todo relevante não fosse o fato de que dos
peixes ao homem há somente uma aparente refinamento à elaboração gradual dos
mecanismos cerebrais, sem qualquer alteração radical dos princípios fundamentais de
operação (SPERRY, 1952).
Ainda do ponto de vista filogenético, a existência de uma espécie animal
chamada de ascídias, ou tunicatas (Acideacia) tem sido interpretada pelos
neurocientistas e biólogos evolucionistas como uma prova em favor dessa perspectiva
4 Há também produto glandular, mas esse output por ser desconsiderado uma vez que mantém como
função a regulação fisiológica da homeostase de nosso corpo.

7
sobre a função do SNC. Em sua fase adulta, esses animais são sésseis e ancorados por
um pedículo em um lugar fixo no mar, cujas únicas funções na vida consistem em filtrar
a água em busca de alimento e reproduzir-se por brotamento. Em sua fase larval, as
ascídias produzem uma larva semelhante a um girino, a qual possui células ganglionares
semelhantes a um cérebro – aproximadamente 300 células. Esse sistema nervoso
primitivo recebe informação do entorno por meio de um estatocisto – órgão de
equilíbrio –, um pedaço rudimentar de pele sensitiva à luz e uma corda dorsal – uma
espinha dorsal primitiva – tornando-as, assim, capazes de lidar com o entorno. O fato
surpreendente ocorre assim que a larva acha um local onde possa se estabelecer
definitivamente, pois, uma vez feito isso, a larva enterra sua cabeça e literalmente digere
seu protocérebro, sua corda dorsal, sua calda e a musculatura da calda, voltando assim a
ser uma criatura séssil (LLINAS, 2001; CLONEY, 1982). Ou seja, não havendo mais
locomoção, deixa também de haver a necessidade de se ter um Sistema Nervoso
Central.
Agora, de acordo com Wiener, “[...] a estrutura da máquina ou organismo é um
índice do desempenho que dela se pode esperar” (1989, p. 57). Ou, em outras palavras,
que sendo nosso SNC uma máquina constituída para produzir e conduzir motricidade
inteligente – como oposto a um processo motriz gerado ao acaso – então é de se esperar
que os estágios intermediários nesse processo estejam em função desse propósito. No
entanto, o SNC é apenas um subsistema do organismo biológico como um todo. Assim,
antes de tentarmos entender qualquer estágio intermediário no processo de geração de
motricidade, devemos entender o papel que a motricidade compre nesse contexto mais
amplo, a saber, o organismo vivo.
2. A função do SNC no organismo biológico
2.1. Organismo como um Sistema Complexo
Um dos grandes benefícios das noções de sistema e entorno consiste na
relatividade quanto a seu uso – a qual tem sido pouco explorada em filosofia. Deixe-nos
definir entorno da seguinte maneira:
[Def. 2.1.] entorno consiste no conjunto de variáveis cuja mudança afeta o organismo e
as variáveis que sofrem alteração pelo comportamento do organismo (ASHBY, 1966,
p.86).

8
Assim, as noções de sistema e entorno podem ser relativizadas tendo em vista que é
sempre possível conceber dois sistemas em interação E e O como um sistema complexo
único C, tal que o input do sistema E consiste, nessa nova configuração, no input do
sistema C, enquanto que o output do sistema O consiste no output do sistema C – onde
E pode ser entendido como entorno, O organismo e C o ecossistema, por exemplo.
Desse modo, o organismo biológico é um sistema para o qual o meio ambiente é seu
entorno, o SNC é um sistema para o qual o corpo é parte do entorno, uma rede neural é
entorno para outra rede neural, um neurônio é um sistema para o qual outros neurônios
são entorno e assim por diante (ASHBY, id.; MAINZER, 2010). Podemos considerar,
agora, o organismo biológico como um sistema complexo composto de vários
subsistemas para o qual o SNC faz parte e analisar algumas propriedades importantes
desse sistema complexo, a fim de tentarmos compreender qual o papel que o SNC,
enquanto um de seus subsistemas, cumpre no organismo biológico.
2.1.1 Transmissão de Informação
Antes de introduzirmos a noção de organismo vivo e mostrarmos como tal noção
pode nos ser útil, deixe-nos introduzir algumas noções de Teoria da Informação que
serão necessárias. Em um amplo sentido, Teoria da Informação pode ser entendida
como o estudo quantitativo “[,,,] dos procedimentos por meio dos quais um mecanismo
afeta outro mecanismo” (SHANNON & WEAVER, 1963, p. 3).
Figura 2 - Sistema de Informação
Logo se pode perceber que tal teoria será de grande utilidade em cibernética, tendo em
vista que o tipo de sistemas estudado pela disciplina consiste justamente naqueles em

9
que sua dinâmica depende da troca contínua de matéria ou energia do sistema com seu
entorno – sistemas abertos, como já vimos. De acordo com a teoria da informação, cada
sistema pode ser entendido como uma fonte de informação, onde grau de liberdade com
que um sistema pode afetar outro sistema é definido como a entropia desse sistema.
Entre dois sistemas A e B deve haver ainda um meio por onde A possa afetar B, o qual
se chama de canal transmissão do sistema de informação. A Figura 5 vem a ser uma
típica representação de um sistema de informação. Assim, há uma fonte de informação,
a qual se caracteriza por certa entropia. Uma vez que essa fonte não pode enviar seus
símbolos diretamente para o destino, o transmissor codifica os símbolos da fonte em
termos de um sinal, o qual é enviado por um canal. Do outro lado, há um receptor, o
qual decodifica sinal recebido em termo dos símbolos enviados pela fonte de
informação. Para o envio do sinal é necessário, como já dissemos, um meio de envio
desse sinal, o canal de transmissão do sinal. No processo de transmissão de um sinal por
um canal há comumente um ruído, que consiste em qualquer distúrbio interior ou
exterior ao canal, o qual aumenta a entropia desse sinal, dificultando a decodificação
precisa do sinal enviado.
Para efeito de nossos propósitos não será necessário considerar um sistema de
informação com tal nível de detalhes – em geral, ignoraremos os resultados referentes
aos limites em compressão de dados – mas será suficiente apenas considerar a fonte e a
transmissor como um sistema único A, o canal, o ruído e o receptor e o destino como
outro sistema B. Assim, a Figura 6 ilustra o tipo de sistema que usamos para apresentar
certos resultados.
Figura 3 - Sistema de Informação Simplificado
Shannon (id.) introduz uma medida para o grau de liberdade com que um
sistema pode afetar outro, chamada de entropia do sistema, a qual é dada pela fórmula
, onde X é uma variável aleatória, E é o valor estimado e p é um valor
entre 0 e 1 que a variável aleatória X pode assumir. Uma vez que, em geral, os códigos

10
de compressão usados são binários, usa-se o logaritmo de base 2, pois este coincide
exatamente como o número de símbolos binários necessários para codificar certa fonte
de informação. Assim, se a transmissão incessante através do canal for vista como um
processo estocástico, então a transmissão num lapso de tempo Δt consistirá em uma
sequência φ com uma estrutura estocástica tal que complexidade de φ é dada por H(X).
Notemos que o grau de liberdade com que o entorno pode afetar o sistema – ou seja, seu
conjunto de inputs – pode agora ser tratado em termos da entropia desse sistema, e o
mesmo para o grau de liberdade concernente ao que resulta dessa interação no que se
refere ao sistema afetado – o output do sistema. Em geral, a palavra informação pode
ser usada para designar esse conjunto de inputs que é capaz de afetar o sistema, porém
deve-se estar ciente de que a noção de informação apresenta um sentido um pouco mais
restrito, a saber, aquela variedade resultante do distúrbio – denominada de informação
mútua –, enquanto a variedade possível deve ser denominada de entropia.
2.1.2. O organismo vivo
Passaremos, agora, ao tratamento do organismo vivo como um sistema dinâmico
aberto, ou máquina com input. Da [Def. 1.2.] podemos notar que o organismo pode ser
representado como um sistema cujo comportamento é determinado pela informação que
afeta o sistema. Assim, como é usual, dado um estado, o organismo apresentará, em
geral, um comportamento distinto relativo a cada input distinto; ou seja, haverá uma
transição para um estado diferente. Evidentemente não é necessário que o organismo
reaja diferentemente com relação a cada input, nem tão pouco que sua transição seja
totalmente determinada – ou seja, com probabilidade 1 para um estado e 0 para os
demais – mas é suficiente que a noção de ‘determinado’ aqui tenha o sentido de
respeitar certa distribuição de probabilidade5. O interessante é que o organismo
biológico, assim entendido, coincidirá exatamente com o esquema apresentado na
Figura 6, onde o organismo será visto como um canal de informação. Podemos definir
um canal de informação da seguinte maneira
[Def. 2.2] dado duas variáveis aleatórias E={ } e O={ }, diz-se
haver uma canal transmitindo informação entre E e O se e somente se a entropia
5 Note que, embora se use a noção de determinado como oposto a estocástico, um sistema determinista é
apenas um caso particular de um sistema estocástico no qual as transições ocorrem com probabilidade 1
para um caso e 0 para os demais.

11
condicional de E dado O for menor que entropia de E , o que é expresso pela fórmula
H(E|O)<H(E)6.
Ou seja, suponhamos que E seja o conjunto de inputs por meio dos quais o entorno é
capaz de afetar o organismo e O seja os possíveis estados do organismo resultantes da
relação, então a entropia condicional H(E|O) será máxima – igual a H(E) – apenas no
caso em que E e O sejam completamente independentes um do outro. Em outras
palavras, se dois pontos mantiverem qualquer relação comportamental, então existe um
canal entre esses dois pontos (ASHBY, 1956, p. 180). Como se pode perceber, o
organismo biológico satisfaz essa condição.
Agora, devemos nos perguntar que tipo de informação o organismo está
continuamente transmitindo? E que tipo de informação, em especial, podemos dizer que
o organismo deseja transmitir? Como foi dito, um sistema pode ser entendido como um
conjunto de variáveis – onde o conjunto de pontos no espaço de fase consiste no
conjunto de vetores estado do sistema – cada uma descrevendo certo aspecto do sistema.
Assim, podemos ter variáveis estando para o comportamento geométrico do organismo
– sua movimentação –, variáveis para o volume ocupado pelo organismo no espaço –
altura, comprimento do cabelo –, variáveis para a pressão sanguínea do organismo,
variáveis para a quantidade de sangue do organismo – e quantidade de glóbulos
vermelhos, etc. –, variáveis para quantidade de oxigênio por litro de sangue etc.. O
conjunto de variáveis pode ser entendido como um vetor, e a dinâmica do sistema
durante um lapso de tempo Δt pode ser representado por uma variável aleatória
. Assim, podemos denotar o vetor estado do organismo como o vetor
. O mesmo pode ser dito das várias formas pelas quais o entorno pode afetar o valor
das componentes do vetor e podemos denotar essa variedade também por um vetor
, onde possa ocorrer. Agora, dentro dos possíveis valores que as componentes
do vetor podem assumir, é razoável definirmos um subconjunto desses possíveis
6 Intuitivamente podemos ilustrar a definição 2.2. da seguinte maneira. Imagine que a posição da mão
direita de um sujeito A em um espaço de fase tridimensional, relativo a um eixo de coordenadas definido
pela linha dos ombros e outra linha perpendicular a ela. Assim, poderíamos dizer que a posição de mão do
sujeito A define um ponto em cada instante t, e que esse espaço de fase pode definir uma fonte de
informação. Agora imaginemos um sujeito B, no Japão, na mesma situação. Poderíamos, então,
perguntar: qual a probabilidade de que, dado que o sujeito A, em um tempo t’, está com a mão levantada
(ou seja, que a posição de sua mão defina um ponto pertencendo a um subconjunto δ de pontos nesse
espaço), o sujeito B esteja com a mãe abaixada (ou seja, que a posição de sua mão defina um ponto
pertencendo a um subconjunto γ de pontos nesse espaço)? Uma vez que não há qualquer relação entre
ambos A e B, seria razoável supor que a probabilidade de Pr(B|A) seria igual para todos pontos definidos
por B – ou seja, H(A|B)=H(A).

12
valores como aqueles nos quais o organismo está vivo, o que define também um
subconjunto dos vetores , digamos os vetores . Assim, para qualquer
transição na qual o valor em que i for menor do que certo índice k e maior do que o
índice m, diremos que o organismo morreu. Esse subconjunto de vetores
compreenderá aqueles estados cujas variáveis essenciais para a sobrevivência do
organismo – tais como pressão sanguínea, níveis de oxigênio no sangue, integridade
anatômica, etc. – se mantiveram dentro de certos valores desejados.
A informação que o organismo deseja transmitir, é razoável assumir, consiste no
organismo vivo. Como o subconjunto de vetores em que o organismo está vivo,
, define um subconjunto dos estados possíveis de O, então se pode dizer que o
que se deseja é manter o organismo num estado de equilíbrio estável. Onde a
estabilidade de um estado de equilíbrio pode ser definida, segundo Lyapunov (1959), da
seguinte maneira:
[Def. 2.3.] Um estado de equilíbrio será estável se para qualquer domínio dado ε de
desvios permitidos do estado de equilíbrio v é possível indicar um domínio δ, que inclui
o estado de equilíbrio, tal que a trajetória de qualquer transição começando na região
definida por δ nunca alcançará os limites do domínio ε.
Um estado de equilíbrio é um no qual nenhuma das coordenadas do sistema sofre
mudança (LERNER, 75, p.40). Agora, deixemos que o conjunto de vetores
esteja para o domínio δ da definição [Def. 2.3.] e assim, podemos dizer que o que se
quer é que o organismo esteja em um estado de equilíbrio estável com relação a um
estado v.
Figura 4 - Organismo como um canal de informação

13
Uma vez que estamos falando de vida e morte, não podemos tomar como
exemplo apenas um único organismo, mas será necessário falarmos de um conjunto de
organismos. Assim, deixe-nos introduzir uma segunda variável aleatória V(v), de vida,
tal que (V=0), caso um organismo se mantenha vivo, e (V=1) caso um organismo
morra. Assim, pode-se dizer que o entorno está transmitindo uma mensagem, por meio
do organismo, de entropia H(V), tal que resultará numa entropia nula ou próxima de
zero caso todos os organismo permaneçam vivos – ou casos todos morram (Figura 4).
Tal comportamento dá-se, pois,
quando p=1 é igual à zero.
Porém, eventualmente eventos indesejados ameaçam o organismo. Por exemplo,
um carro em alta velocidade em trajetória coincidente com o organismo, uma infecção
bacteriana, uma redução drástica na temperatura do ambiente, etc.. Todas essas
informações, entre outras, podendo conduzir nosso sistema para um estado fora do
domínio definido como estável, conduzindo a organismo a uma instabilidade
irreversível – morte. Uma vez que nós definimos o organismo como um canal por meio
do qual se deseja transmitir uma única mensagem, qualquer variedade adicional que
venha a aumentar sua entropia H(O)˃0 será considerado como ruído indesejado na
transmissão. Nesse caso, então, é necessário algum mecanismo por meio do qual
possamos barrar essa informação adicional de modo que nosso sistema receba apenas a
mensagem desejada, a saber, v. Tais mecanismos, em Cibernética, são mecanismos de
regulação e controle e, como veremos, nosso SNC pode plausivelmente ser entendido
como um eficiente mecanismo desse tipo.
2.2. O SNC como um Sistema de Regulação e Controle
Um sistema de regulação e controle consiste em um sistema acoplado a outro
sistema, o sistema controlado, cuja função é evitar qualquer desvio do sistema regulado
de seu estado de equilíbrio desejado. O sistema regulador, por meio da emissão de um
input no sistema acoplado, reconduz o sistema controlado a seu estado de equilíbrio, ou
evita que o mesmo se afaste desse estado. Sendo assim, o input emitido pelo sistema de
regulação não pode ser aleatório, mas deve ser escolhido com vistas ao objetivo de
regulação. O sistema regulador tipicamente usa duas fontes de informação para escolher
seu input. A primeira fonte consiste na variedade resultante do desvio do sistema
controlado. A segunda fonte de informação pode ser qualquer variedade por meio da
qual se possa estimar um possível desvio futuro do estado de equilíbrio. Assim, o
segundo caso pode ser chamado de extrapolativo, uma vez que extrapola a situação

14
presente, e, o primeiro, não-extrapolativo como oposto ao primeiro7 (ASHBY, 1956;
ROSENBLUETH et al., 1943; LERNER, 1975; ARBIB, 1981). Como veremos SNC
desempenha ambos os papéis, porém, para levar a cabo o processo de regulação com
sucesso, se lhe impõe ainda o requerimento de um sistema de antecipação – forward
(KAWATO, 1999; GRUSH, 2004).
2.2.1. Sistemas de Resposta Não-Extrapolativos e Extrapolativos – feedback e
feedforward
Em um sistema de resposta não-extrapolativo, a informação que o sistema
escolhe para regulação resulta do desvio ocasionado pela interação do organismo com o
entorno. Por exemplo, no caso do tremor de frio, tão logo nosso corpo começa a baixar
sua temperatura da faixa dos 36,5ºC, o SNC emite um tremor como resposta na tentativa
de reconduzir sua temperatura para o estado desejado – sistema de resposta não-
extrapolativo [Figura 5]. Uma vez que o sistema não-extrapolativo age em função do
desvio provocado pelo entorno no sistema controlado, ele torna-se pouco efetivo na
preservação da vida, pois, quando o desvio for tal que provoque a morte do organismo,
o sistema obviamente já não poderá realizar o controle.
Figura 5 Sistema de Regulação de Resposta
Assim, para que o organismo consiga uma regulação efetiva da informação
proveniente do entorno, torna-se necessária outra estratégia de controle, a saber, uma
extrapolativa. No caso sistema de controle extrapolativo [Figura 6], ao invés de colher
informação do desvio resultante a interação do organismo com o entorno, ele colhe
informação sobre os possíveis desvios do sistema de seu estado de equilíbrio. Assim, ao
invés de emitir um input para reconduzir o organismo controlado ao estado desejado, o
7 Os termos extrapolativo e não-extrapolativos são empregados por Rosenblueth et al. (1943), mas são
sinônimos de feedback e feedforward.

15
sistema extrapolativo previamente emite um input tal que, sob o parâmetro da variedade
prestes a afetar o organismo, irá manter o sistema controlado no estado desejado. Na
Tabela 1 nós vimos que o mesmo estado, sujeito a diferentes inputs, em geral, conduz o
sistema a diferentes outputs. Assim, podemos perceber que, em geral, diferentes estados
conduzem o sistema ao estado 4, por exemplo, relativo a diferentes inputs. Destarte, se
o input 1, por exemplo, estivesse na iminência de afetar o sistema e o estado desejado
fosse o 4, o sistema de controle emitiria um input no sistema controlado conduzindo-o
ao estado 2, o qual, reativo ao input 1, conduz o sistema ao estado 4.
Figura 6 Sistema de Regulação Antecipatório
Um sistema extrapolativo pode explorar diferentes estratégias de regulação de
acordo com as características do sistema controlado. Por exemplo, pode haver um
estado privilegiado em um sistema controlado tal que a transição, sob uma considerável
variedade de parâmetros, ocorra para estado de equilíbrio desejado. Assim, sistemas
extrapolativos podem ainda ser divididos em estáticos e ativos (ASHBY, 1956;
LERNER, 1975). Um sistema extrapolativo estático é aquele que mantém o sistema
controlado sempre no estado privilegiado. Exemplos desse tipo de sistema são o casco
da tartaruga, a carapaça dos mariscos e caramujos, armaduras e coletes a prova de bala,
e a lataria dos automóveis. Porém, uma vez que a o entorno costuma caracterizar-se por
uma entropia superior aquela expressa pelo conjunto de parâmetros definido pelo estado
privilegiado, sistemas extrapolativos estáticos não são a melhor estratégia.
Outra alternativa seria a de um sistema extrapolativo ativo. Nesse caso, o
sistema é capaz de escolher, relativo a cada input prestes a afetar o sistema, o estado
mais apropriado para a sobrevivência ou manutenção da estabilidade do organismo.
Pode-se perceber que a efetividade em regulação, nesse caso, está diretamente
relacionada à variedade de opções disponível para escolha, assim como à eficiência na

16
escolha mais adequada – chamada Lei da Variedade Requerida [4.1.]. A qualificação
‘ativo’ deriva do fato de que, em geral, a motricidade é o meio pelo qual o sistema de
regulação afeta o sistema controlado. Ou seja, o sistema de regulação detecta a
informação do entorno e, através do movimento, determinada o estado mais apropriado
para a manutenção da estabilidade do sistema. Notemos que aqui existe uma clara
relação entre a caracterização dada ao SNC no início de nosso texto – como um sistema
cuja única forma de afetar seu entorno ocorre através da motricidade – e o papel que
esse sistema cumpre no organismo com fins da preservação da vida8.
Uma vez sugerido que o SNC seja visto como um sistema de regulação e
controle da vida do organismo, somos naturalmente conduzidos a hipótese de que nossa
noção de realidade seja um componente no processo de regulação. Em geral, uma
combinação entre sistemas extrapolativos e não-extrapolativos são admitidos como
elementos componentes do processo de regulação pelo SNC (KAWATO, 1999;
GRUSH, 2004). Porém, seriam esses conceitos suficientes para o processo de regulação
com sucesso? Haveria a necessidade de uma noção de realidade no processamento da
informação proveniente do entorno? Veremos que os sistemas de resposta não-
extrapolativo e extrapolativo não seriam suficientes para um processo de regulação com
sucesso, exigindo a elaboração de um emulador ou sistema antecipatório - forward.
Nossa noção de realidade pode ser entendida como um emulador ou sistema
antecipatório.
3. Regulando a informação: requerimentos
Veremos nessa sessão quais requerimentos se impõe para que Sistema Nervoso
Central execute sua função regulatória com sucesso com vistas a identificar alguma
evidência que obrigasse o sistema regulatório à elaboração de concepção de entorno –
nossa realidade.
8 Parece sugestivo, ainda que especulativo, que a vida transite, na medida em que aumenta sua
complexidade, de sistemas extrapolativos estáticos para sistemas extrapolativos ativos. Nossos
automóveis são exemplos clássicos de sistemas extrapolativos estáticos, os quais paulatinamente rumam
para tornarem-se sistemas extrapolativos ativos, onde mecanismos são projetados para lidar com situações
que fogem a capacidade de nosso SNC, tal como detecção de condições de derrapagem, estabilidade e
tráfego. Pode-se perfeitamente especular que a natureza de nosso sistema de regulação – nosso SNC –
poderia ser diferente, seja porque nossa realidade física permitisse outros meios de regulação ativa, ou
porque a informação relevante para regulação fosse de natureza distinta. Assim, podemos imaginar que,
fosse o teletransporte, algo efetivamente realizável, ou pelo menos facilmente realizável, nosso sistema
poderia, em vez de motricidade, resultar justamente na capacidade de se teletransportar para os lugares
desejados. E caso fosse, em vez da massa, a velocidade e a trajetória as informações mais relevantes para
regulação, a atração gravitacional, a qual é desconsiderável no que tangencia o trato com as entidades
mundanas, então talvez nossa intuição básica fosse a do campo gravitacional dos objetos em vez de suas
relações espaço temporais – modificando nossas sacrossantas intuições básicas do entendimento.

17
3.1. Memória
O SNC, foi sugerido, deve ser entendido como um sistema extrapolativo ativo.
Assim, o sistema continuamente colhe informação do entorno com vistas à
determinação de possíveis desvios indesejados futuramente. Porém, note que a distinção
entre aqueles estados cuja transição levaria o sistema a desvios indesejados e aqueles
estados cuja transição manteria o sistema em seu estado de equilíbrio requer a existência
de memória. Mais especificamente, assuma que o entorno caracteriza-se pelo vetor
estado num dado tempo t, a inexistência de qualquer informação prévia é consistente
com a possibilidade de que, sob o efeito da variedade , um dos estados
transite, num tempo t+1, com a mesma probabilidade para qualquer um de seus n
estados possíveis, o que tornaria a escolha um completo acaso. Assim, podemos dizer
que, para lidar com o presente, o sistema precisa de uma noção de passado (memória) e
futuro (possíveis desvios). A memória consiste no modo pelo qual o sistema é capaz de
escapar da contingência radical inerente ao fato isolado para uma realidade possível e
previsível [Imagem 10]. Essa interpretação do papel da memória é consistente com os
seguintes fatos: que nossas memórias da infância são esquecidas, pois, devido às
exageradas mudanças nas dimensões corpóreas, elas antes nos atrapalhariam no
processo de regulação (WOLPERT, vídeo); e que nossa vida subjetiva transcorre no
plano do futuro possível, e não do agora isolado. O agora já não é interessante para o
processo de regulação (LLINÁS, 2001; FRITH, 2006; EAGLEMANN, 2012).
O que ocorre, assim, é que o sistema interioriza certas características da
dinâmica do entorno de modo que essa informação possa restringir a variedade dos
eventos futuros – interiorização a qual chamamos de memória. Nosso organismo dispõe
de diferentes tipos de memória – memória a curto prazo, memória longo prazo e suas
subclassificações (TULVING & SCHACTER, 1990) . Uma distinção mais abrangente
consiste em entendê-las em termos de memórias filogenéticas e ontogenéticas.
Compreende-se como memória ontogenética qualquer conhecimento adquirido durante
a vida do indivíduo particular, independente da latência temporal – episódios,
aprendizagens, o número de telefone memorizado até a discagem. Compreendem-se por
memória filogenética todas as capacidades cognitivas que herdamos no decorrer da
evolução de nossa espécie – tais como as capacidades de distinguir os objetos quanto ao
padrão de reflexão, movimentos, reconhecimento de faces, etc. (WOLF, 2009; LLINÁS,
2001).

18
Imagem 7. Qual a trajetória possível dos objetos nessa foto? (Fotografia Dali Atomicus (1948), por Philippe
Halsman)
Contudo, a exigência de memória pelo sistema de regulação não está
condicionada a elaboração de uma concepção de realidade, pois, seria suficiente que os
estados fossem escolhidos com base no padrão da informação prestes a afetar o sistema.
De fato, essa perspectiva, na qual nossas representações internas não comprem papel na
interação com o entorno, já foi defendida em psicologia (GIBSON, 1966, 1967). De
outro modo, os sistemas de regulação artificiais construídos pelo homem em robótica
são quase que inteiramente sistemas de regulação de resposta (KAWATO, 1999). Mas
seria possível que o sistema pudesse escolher entre os estados desejados e indesejados
com base no padrão da informação proveniente do entorno? A comparação com os
sistemas de controle em robótica será útil para que possamos perceber os limites que se
impõem no processamento de informação pelo SNC.
3.3. Ruído
Frequentemente, fatores podem intervir negativamente na decodificação da
informação proveniente do entorno prejudicando no processo de regulação, o que exige
do regulador estratégias para contornar essa interferência – a essa interferência negativa
se dá o nome de ruído. O ruído numa mensagem ou sinal deve ser entendido como uma
variedade adicional indesejada que é incluída no sinal no processo de transmissão. Essa

19
variedade adicional se define com relação à variedade característica da mensagem
transmitida (ASHBY, 1956; SHANNON, 1964). Assim, qualquer variedade adicional
que altere em alguma medida o sinal recebido deve ser considera como ruído e, logo,
indesejada; pois, havendo ruído no sinal recebido, o SNC pode ser levado a escolher um
estado inapropriado, causando a morte do organismo.
3.2.1. Emulando a realidade para corrigir o atraso temporal
Em sistemas de regulação em robótica, a informação colhida pelos mecanismos
de transdução é transmitida até o centro de controle através de circuitos eletrônicos a
uma frequência de 500-1000 Hz, com um baixíssimo nível de ruído. A transmissão de
informação nesses sistemas de controle é da ordem de 1-2ms, o que torna o sistema
competente para operar diretamente com base na informação colhida do entorno pelos
canais sensoriais. Diferentemente, no caso do organismo biológico, tal como o
conhecemos, a informação é transmitida através de um processo eletrofisiológico
denominado de sinapse, cuja taxa de transmissão é da ordem dos 150-250ms – nos
casos mais rápidos, na espinha dorsal, ficando ainda na ordem dos 30-50ms
(KAWATO, id.; FELDMAN, 2006). Porém, tendo em vista que a velocidade de nossos
movimentos mais rápidos é da ordem dos 150ms e dos movimentos intermediários é da
ordem dos 500ms, esse atraso na transmissão do sinal no SNC deve ser considerado
como ruído, uma vez que o mesmo interfere na decodificação da mensagem pelo SNC,
a saber, a escolha do estado desejado (KAWATO, id.; NIJHAWAN, 2008). Caso o
SNC atuasse diretamente com base na informação proveniente do entorno, não seríamos
capazes de rebater uma bola de tênis em um jogo, pois estaríamos sempre tentando
rebater a bola no passado.
Disso segue-se fatalmente que o SNC não pode efetuar a regulação com base na
informação recebida diretamente do entorno, pois, tendo em vista que as estimativas do
sistema estão baseadas no estado presente do entorno, elas inevitavelmente seriam
prejudicadas pelo atraso temporal na transmissão da informação. Assim, o sistema
precisa de uma estratégia para corrigir esse ruído imposto pelo atraso em transmissão.
Para tal o SNC elabora um emulador dos eventos exteriores, um modelo interno do real,
o qual atua antecipadamente com relação à informação proveniente do entorno, usando-
a antes como sinal de correção (GRUSH, 2004; DESMURGET & GRAFTON, 2000;
KAWATO, 1999; WOLPERT, 1995). Desse modo, o SNC não efetua sua regulação
baseado diretamente sobre a informação fornecida pelo entorno, mas sim, sobre a

20
informação fornecida pelo emulador da realidade. A informação provida pelo entorno
serve para a atualização antecipatória do emulador, o qual fornece a informação
relevante para o SNC efetuar seu papel regulatório. A realidade que temos consciência,
e sobre a qual atuamos, deliberamos, pensamos, etc. consiste no modelo criado pelo
SNC, com base nas memórias filogenéticas e ontogenéticas, para suprimir o ruído
imposto pelo atraso em transmissão.
Assim, nossa noção de realidade pode ser vista como uma estratégia que o SNC
elabora para correção de um tipo de ruído na transmissão – o atraso temporal. Porém,
não haveria alguma forma de ruído que afetasse nosso próprio emulador da realidade?
Pois, sendo nosso emulador da realidade, um sistema de antecipação – forward –
baseado sobre a informação proveniente do entorno, pode haver ainda diferentes formas
de ruído que afetem o próprio emulador.
3.2.3. O caráter indeterminado do SNC
Todo sistema neural exibe um caráter profundamente aleatório, desde a
percepção dos sinais sensoriais até a geração de atividade motora, o que tem conduzido
os neurocientistas à hipótese de que haveria ruído presente em todos os níveis do
processamento de informação do SNC (DESTEXHE & RUDOLPH-LILITH, 2012;
FAISAL et al., 2008; STEIN et al. 2005; WOLPERT et al. 2003). Esse consiste num
dos pontos de maior atenção nos presentes dias em neurociências, pois, uma vez que
ainda não se conhece a linguagem neuronal, pode-se estar tratando como ruído uma
variedade que consita, na verdade, em informação – por exemplo, descobriu-se que a
variedade temporal inerente ao sinal, a qual antes era considerada como ruído, é parte
importante da informação veiculada (FAISAL et al., id). Essa aleatoriedade
característica do processamento de informação do SNC esta presente tanto na
transmissão de informação neuronal, quanto no sinal que é transmitido do entorno para
o sistema nervoso.
Referente aos sinais transmitidos do entorno para o sistema nervoso – ou seja, o
feixe de fótons que afeta nossas células fotossensíveis, o choque das moléculas de ar nas
células ciliadas da cóclea, ou os processos químicos envolvidos nos sistemas gustativo e
olfativo –, todos estão repletos de ruído indesejado. Por exemplo, os fótons que afetam
nossa retina chegam até as células fotossensíveis de forma totalmente aleatória – o que
pode ser pensado como um Processo de Poisson –, e não exibindo um padrão perfeito
característico de um código (PIRENNE, 1959). As ondas sonoras que afetam nossas

21
células ciliadas do sistema auditivo estão sujeitas ao Movimento Browniano criando
flutuações consistentes com uma variedade adicional (HARRIS, 1968). Da mesma
forma, nossos órgãos sensoriais químicos estão sujeitos ao ruído termodinâmico,
também causando flutuações adicionais (BERG & PURCELL, 1977; BIALEK &
SETAYESHGAR, 2005).
Referente ao ruído na transmissão intrínseca ao SNC se verifica uma variedade
compatível com ruído em todos os níveis do processamento de informação – ao nível
molecular, celular e das redes neuronais. Por exemplo, as células neuronais não
respondem uniformemente a uma sucessão de estímulos uniforme, disparidade a qual é
percebida pelos neurônios corticais. E, desde que se percebeu que o intertalo temporal
entre um potencial de ação e outro veicula informação, é plausível supor que essa
variedade adicional seja considerada como ruidosa (SHADLEN & NEWSOME, 1998;
SOFTKY & KOCH, 1993). Além disso, o potencial de membrana das células neuronais
está sujeito a contínuas flutuações, alterando continuamente os limites do potencial de
ação. Esse fato faz com o potencial de ação ocorra na ausência de atividade pré-
sináptica, ou com que uma célula não responda à atividade de outras em certos casos
(DIBA et al. 2004). Essas flutuações, que se devem em parte ao ruído do canal de íon,
aumentam inversamente proporcional ao tamanho das células, uma vez que a resistência
do input da membrana aumenta rapidamente com a diminuição do diâmetro (RALL,
1969; FAISAL et al., 2005). Ainda, quando um impulso elétrico é enviado por longos
axônios, a taxa de falha no envio do sinal é da ordem dos 5-80% (DEBANNE, 2004).
Os exemplos acima dão apenas uma pequena amostra da complexidade dos processos
envolvidos na transmissão neuronal, os quais afetam os processos de percepção,
cognição e motricidade.
Flutuações e estabilidade são conceitos que andam lado a lado, pois, quanto mais
instável for um sistema relativo à informação proveniente pelo entorno, mais esse
sistema estará sujeito a flutuações. Assim, se levarmos em consideração o alto grau de
estabilidade dos sistemas eletrônicos, em contraste com os sistemas biológicos, e a
inevitável presença de uma taxa de ruído em transmissão nesses sistemas, torna-se
sensato aceitarmos que as variedades características do processamento de informação
neuronal devem ser em boa medida tomadas como ruído.
Como o cérebro lida com tal sinal ruidoso de modo que nosso modelo nos
apresenta uma representação detalhada do entorno? Duas estratégias parecem jazer
nesse processo, a saber, a utilização de canais em paralelo e o uso de conhecimento

22
prévio (memória). A utilização de canais em paralelo consiste num meio de introduzir
redundância no sinal de modo a restringir a ação do ruído. A redundância deve ser
entendida como qualquer redução do grau de liberdade de uma mensagem (ASHBY,
1956; SHANNON, 1964). A estratégia consiste em enviar o mesmo sinal por meio de
diferentes canais para um mesmo receptor. Assim, assumindo que o ruído interfira de
forma independente em cada canal, o receptor – um neurônio ou rede neuronal – pode
usar a redundância inerente aos diferentes sinais – uma vez que se esperaria veicularem
todos a mesma informação – para decodificar o sinal (FAISAL et al., 2005). Verificam-
se indicações de que esses procedimentos sejam adotados pelo SNC na correção do
sinal interior nos sistemas auditivo e visual, por exemplo (KOZLOV et al. 2007;
KANDEL et al. 2000). De outro modo, testes cognitivos mostram que a interpretação
do sinal proveniente do entorno é realizada também por via de diferentes canais, tal que,
por exemplo, em ambientes acusticamente ruidosos, o CNS processa a compreensão da
fala com o uso das características gestuais da boca no dialogo (MA et al. 2009). Essa
estratégia de usar canais adicionais para diminuir o ruído e aumentar a capacidade de
transmissão parece ser muito mais ubíqua no processamento de informação do SNC e
pode ser útil para explicar processos cognitivos mais complexos e menos intuitivos, tais
como o uso de maquinários conceituais e de computação.
Contudo, a introdução de redundância no sinal por meio do uso de canais em
paralelo pode não ser suficiente se os canais forem muito ruidosos ou afetarem não
uniformemente os diferentes sinais (JACOBS, 1999; KNILL, 2003). Assim, parece que
o SNC usa também outra estratégia, a saber, conhecimento prévio. Deve-se perceber
que somente é possível que se realize previsões uma vez que o entorno mostre certa
redundância (LAUGHLIN, 1981; FIELD, 1987). A Imagem 10 não se mostra como um
caos total, pois nós sabemos que certos eventos são mais prováveis, enquanto outras são
altamente improváveis. Esse conhecimento acerca da estrutura probabilística de um
sinal, o qual é denominado de conhecimento prévio, serve para que o cérebro possa
decodificar o sinal mesmo que a introdução de redundância pelo sistema não seja
suficiente para eliminar todo o ruído do sinal (FAISAL et al., 2005; WOLPERT &
GHAHRAMANI, 2009).
O que essas análises sugerem é que o conteúdo de nossas representações é, em
larga medida, baseado no conhecimento prévio que temos de situações similares, tanto
quanto da informação proveniente do entorno. Essa é a razão pela qual a adoção de
modelos estatísticos bayesianos tem se mostrada adequada para modelagem de nossos

23
raciocínios – em contraste com modelos deterministas, tais como lógica clássica
(WOLPERT & GHAHRAMANI, 2009; DOYA et al. 2007; SHADMEHR & MUSSA-
IVALDI, 2012). Segundo Faisal et al., os modelos bayesianos de inferência, podem ser
entendidos da seguinte maneira: “[...] os modelos bayesianos de inferência atribuem
probabilidades às proposições acerca na realidade exterior (crenças). Essas crenças são
calculadas combinando conhecimento prévio (por exemplo, que um animal seja
predador) e as ruidosas observações (a aproximação do animal) para inferir a
probabilidade da proposição (por exemplo, o ataque do animal)” (FAISAL et al. .p 299,
2008). Deixe-nos parafrasear a descrição acima pelos termos aqui utilizados, a qual
figuraria da seguinte maneira: <<os modelos bayesianos de inferência atribuem
probabilidades aos estados do modelo interno (o regulador) acerca da informação
proveniente do entorno (distúrbio). Esses estados são combinados com o conhecimento
prévio e suas ruidosas estimativas para inferir a probabilidade do estado do modelo em
relação ao distúrbio>>. O raciocínio poder ser expressar através da seguinte fórmula:
Essa estrutura matemática tem se mostrada empiricamente adequada para modelar
nossas experiências sensoriais e tomada de decisões (KORDING & WOLPERT, 2004;
STOCKER & SIMONCELLI, 2006).
Nessa perspectiva, a informação proveniente pelos canais sensoriais não confere
conteúdo ao modelo interno, mas antes modula sua dinâmica antecipatória, a qual tenta
estar o máximo possível de acordo com o sinal proveniente do entorno – a modulação
do modelo interno é o que se entende por percepção (LLINÁS, 1992, 2001, 2009;
GRUSH, 2004; CHURCHLAND et al. 1994).
3.4. Quão dependente da realidade nosso modelo interno do real é?
Nós viemos evidenciando os requerimentos que se impõem para que nosso SNC
possa regular com sucesso a estabilidade do organismo que somos: ou seja, mantê-lo
vivo. Assim, vimos que além de memória, uma série de estratégias é requerida para
suprimir o ruído na transmissão de informação, o qual pode afetar fatalmente a
regulação. A elaboração de um modelo interno da realidade foi vista como uma dessas
estratégias. Agora devemos nos perguntar: qual é o grau de dependência que nosso
modelo interno mantém do estímulo proveniente do entorno? A visão intuitiva,

24
introduzida por William James (1890), nos diz que nossa visão da realidade é produto
da afecção exterior, onde conteúdo representacional consiste no reflexo causado pelo
estímulo sensorial. Assim, que tipo de evidências pode haver para estimamos essa
relação?
Desde a perspectiva que estamos assumindo, perguntar pela dependência
existente entre aquilo que experimentamos como realidade e a informação proveniente
do entorno é sinônimo de perguntar pela dependência existente entre a geração de certos
estados funcionais no SNC concomitantes com nossa experiência subjetiva enquanto
acordado e os estímulos transmitidos pelos canais sensoriais. Uma estratégia para lançar
luz a esse ponto consiste em perceber as similaridades fisiológicas e neuropsicológicas
existentes entre os estados funcionais característicos do estágio de sono REM9 e aqueles
característicos do indivíduo acordado. Essas similaridades sugerem fortemente que
nossa vida subjetiva caracteriza-se adequadamente como um sonho lúcido gerado pelas
propriedades intrínsecas do SNC e apenas modulada pelos sentidos.
Distinguiram-se, com base comportamental e fisiológica, quatro estágios de
sono, entre os quais as variáveis do sistema flutuam de maneira bem definida –
eletrorritmicidade, caráter de resposta autônoma, tolerância sensorial para acordar. De
um ponto de vista eletrofisiológico, os estados funcionais característicos do estágio de
sono REM e do indivíduo acordado demonstram profundas similaridades, distintos dos
demais estágios de sono. Em ambos os casos, os estados funcionais apresentam uma
eletrorritmicidade dessincronizada, caracterizada por ondas de alta amplitude e baixa
frequência e alto caráter de resposta a estímulos sensoriais, embora no caso do sono
REM não se verifique reatividade no período final de latência, na faixa dos 150-300ms,
característico das áreas mais exteriores do córtex (LLINÁS & PARÉ, 1991; STERIADE
& MACCARLEY, 1990).
De um ponto de vista neuropsicológico, os indivíduos acordados nesse estágio
de sono relatam estarem sonhando. Ainda, o declínio de atividades cognitivas mais
elaboradas seguindo lesões circunscritas do córtex também é refletido no conteúdo dos
sonhos. Por exemplo, pacientes afligidos com negligência unilateral resultante de dano
no lóbulo parietal direito, no qual o lado oposto de campo visual não é percebido,
reportam uma falta de percepção similar em seus sonhos (MESULAM, 1981).
9 Sono REM consiste no período de sono no qual indivíduos acordados nesse estágio relatam estar
sonhando. REM refere-se aos rápidos movimentos nos olhos (rapid eye moviment) característicos desse
estágio.

25
Similarmente, pacientes com prosopagnosia, desordem na qual o sujeito se torna
incapaz de reconhecer faces, relatam que as pessoas que habitam seus sonhos são
completamente sem face (LLINÁS & PARÉ, 1991). De outro modo, sujeitos que
sofrem de cegueira total devido ao descolamento da retina ocorrido na fase adulta,
relatam um imaginário visual totalmente detalhado durante os sonhos, e voltam à
cegueira ao despertar (HULL, 1990). Essas evidências apontam para a conclusão de que
os estados funcionais responsáveis pelos nossos sonhos e aqueles responsáveis pela
nossa concepção de realidade são estruturalmente equivalentes, e funcionalmente
distintos exceto pelo fato de que, nos sonhos, o SNC volta sua atenção para a memória,
enquanto que, no estado acordado, ele volta sua atenção para os estímulos sensoriais
(LLINÁS & PARÉ, 1991). Ainda, essas evidências mostram que o SNC é capaz de
gerar uma representação equivalente a que temos da realidade de forma completamente
independente dos estímulos sensoriais. Assim, e dado essa capacidade de autonomia do
SNC na geração de estados funcionais e as limitações já tratadas no parágrafo 2.3.,
podemos imaginar que o conteúdo de nossas representações, talvez, não seja tão
dependente da informação proveniente pelos sentidos como intuitivamente se imagina.
De fato existem evidências morfológicas que suportam a ideia de que a geração
dos estados funcionais do cérebro se deve mais às propriedades oscilatórias intrínsecas
do SNC, do que em virtude dos estímulos que afetam nossos órgãos sensoriais
(LLINÁS, 2001, 2009; LLINÁS & PARÉ, 1991, 2006; LLINÁS et al. 1994). Os
estados funcionais que caracterizam nossas capacidades cognitivas exibem alta
sincronia temporal e espacial, a qual suscita, por hipótese, um sistema de orquestração
desses estados10
. O tálamo tem sido visto como o candidato mais proeminente para essa
função, pelas seguintes razões: i) todas as mensagens sensoriais, exceto a olfativa,
passam pelos núcleos talâmicos; ii) o tálamo, juntamente com a amídala, consiste no
lugar de onde o maior número de axônios em direção ao córtex se projetam de forma
bidirecional; e iii) lesões nos núcleos específicos do tálamos, responsáveis pela
transmissão da informação proveniente dos órgãos sensoriais, ocasionam a perda parcial
de determinada modalidade sensorial, enquanto que lesões nos núcleos inespefícicos
ocasionam coma – ausência de atividade subjetiva (LLINÁS, 2001, 2009; LLINÁS &
PARÉ, 1991, 2006; LLINÁS et al. 1994). Porém, verifica-se que apenas uma minoria
das sinapses que se originam no sistema tálamo-cortical é proveniente dos núcleos
10
Isso não deve ser visto como um entendimento no qual o cérebro teria um sistema central de comando.
Mas apenas de distribuição da informação.

26
específicos, o que indicaria que nossa experiência subjetiva dependeria de forma
minoritária da informação proveniente dos canais sensoriais (LLINÁS, 2001, 2009;
LLINÁS & PARÉ, 1991, 2006; LLINÁS et al. 1994).
Cabe mencionar que essa perspectiva, na qual o cérebro tem certa autonomia na
geração do modelo interno, tem oferecido meios de explicação de casos como
alucinação, esquizofrenia e depressão – o que corrobora a perspectiva.
3.2.5. O modelo interno como um parâmetro pré-motor
Nas precedentes análises nos detemos exclusivamente com as evidências no que
se referem a nossa percepção. Isso, pois, essa é a dimensão acessível à introspecção e,
logo, familiar ao filósofo. Porém, tão desafiante de ser explicado quanto a origem da
subjetividade é o meio pelo qual o cérebro coordena a atividade motora. A dimensão do
problema é excluída do imaginário filosófico pelo mesmo motivo que sempre esteve
ausente da reflexão do homem: ela faz parte da esfera inconsciente da atividade do
SNC. Levemos em consideração que, ainda que de forma diferente, fomos capazes de
criar máquinas que pudessem resolver problemas abstratos de forma muito mais
eficiente que o homem – jogos de xadrez, e a computação de problemas matemáticas.
Porém, não se está nem sequer próximo de criarmos robôs cuja excelência motora
supere a de uma criança de cinco anos (WOLPERT, Vídeo).
O problema da motricidade, como é de se esperar, é bastante detalhado. Mas
será suficiente apenas ilustrarmos sua magnitude. Levante sua mão para pegar um copo
sobre a mesa. Ao fazê-lo, 50 ou mais músculos serão simultaneamente contraídos. Cada
músculo compreende centenas de unidades motoras, as quais devem ser compreendidas
como um conjunto de fibras musculares sob o comando de um específico neurônio
motor. Assim, nosso cérebro teria possíveis modos de levar a cabo a ação. Porém,
uma vez que nossos movimentos não são bruscos, mas suaves e passíveis de mudança
de trajetória ao longo de uma ação, isso indica que nosso cérebro escolhe a cada instante
a melhor trajetória. Desse modo, se o cérebro escolher uma alternativa a cada
milissegundo, então o cérebro faria decisões por segundo. Mas o cérebro contém
apenas aproximadamente neurônios! Ou seja, mesmo se cada neurônio estivesse
envolvido no processo de decisão de pegar um copo, o que não é o caso, isso ainda não
seria suficiente (LLINÁS & ROY, 2012). Agora, essa é uma tarefa bastante simples, e
nosso cérebro escolhe a todo instante, pois, postura, fala, e qualquer forma de ser
envolve uma decisão de caráter motriz.

27
Já deve estar claro o papel que nosso modelo interno do real cumpre no SNC –
um sistema de regulação cujo único meio de afetar o entorno, aparte das secreções
glandulares, consiste em motricidade. Nessa perspectiva nosso modelo do real consiste
num parâmetro pré-motor de ação possível, o qual pode ser levado a cabo ou não. Essa
perspectiva, além de ampliar a noção de representação elabora pelo SNC, também nos
ajuda a estimar o tipo de informação que ganha significado no contexto de nosso
modelo interno. Entender nosso modelo interno como um parâmetro pré-motor amplia
nosso conceito de modelo interno do real, pois, inclui não somente aquele tipo de
representação pictórica, relacionada especificamente com a visão, ou audição, mas
também aquelas representações proprioceptivas, tais como a localização da perna
embaixo da mesa, ou o exato lugar nas costas onde se deseja coçar.
Quanto à informação relevante, deve-se perceber que a função do SNC é a de
assegurar a vida do organismo – enquanto indivíduo e espécie – e que, assim, é
plausível que ganhe significado no seu espaço interno aquela informação relevante para
esse propósito. Não é de se surpreender que não tenhamos mecanismos de transdução de
raio-x, por exemplo, uma vez que esse tipo de energia eletromagnética não oferece
perigo, no modo como a encontramos naturalmente disposta. Outra forma de representar
o que é a mesma ideia, tal como sugere Arbib (1981), consiste em ver nosso sistema
pré-motor como um programa de solução geral de problemas (GPS), tal que as únicas
categorias relevantes para o programa são os estados indesejados, os operadores a
disposição para a solução do problema, e uma tabela com a relação entre os operadores
e os estados a serem evitados. Assim, o que nós representamos são apenas aqueles
operadores relevantes para evitar os estados indesejados e, logo, solucionar o problema.
Há uma miríade de evidências neuropsicológicas suportando a existências de
modelos internos no cérebro para regulação e controle, as quais abarcam tanto a
dimensão sensorial como motora do SNC (JORDAN & RUMELHAR, 1992; MIALL et
al. 1993; WOLPERT, 1995; FLANAGAN & WING, 1997; SNYDER, 1999;
MERFELD et al. 1999; KAWATO, 1999; DESMURGET & GRAFTON, 2000;
GRUSH, 2004) . Porém, uma vez que nossos propósitos filosóficos estiveram
endereçados exclusivamente na dimensão sensorial do modelo interno, a qual, por
hipótese, consistia em nosso sentido mais íntimo de realidade, conceitos e evidências
relacionados a aspectos motor foram deliberadamente omitidos.
4. Os limites da Regulação

28
Agora é chegada a hora de buscarmos por razões e evidências por meio das
quais possamos estimar o grau de acuidade com que nosso modelo interno emula a
realidade exterior. Primeiro, veremos uma razão de caráter teórico, por meio qual
podemos ter uma ideia quantitativa dessa acuidade. Posteriormente veremos evidências
que atestam a limitação nessa fidelidade, as quais parecem estar relacionadas com as
limitações impostas na transmissão e processamento.
4.1. Teorema da Variedade Requerida e o limite em regulação
Deixe-nos representar a relação de regulação por três variáveis aleatórias não
independentes D={ }, R={ } e V={0,1}, correspondentes ao
entorno, o SNC e o organismo.
O que buscamos é um meio de medir a eficiência do SNC enquanto um
decodificador da informação proveniente do entorno; em outras palavras, buscamos
uma função indicadora de erro. Anteriormente falamos de uma variável aleatória V(v),
tal que V era definida da seguinte maneira:
[Def. 3.2.]
Lembremo-nos de que a tarefa do SNC, enquanto um sistema regulador extrapolativo, é
a de escolher corretamente um estado no sistema tal que, somado ao parâmetro dado
pelo entorno, mantenha o sistema vivo. Evidentemente que essa escolha envolve a
decodificação da informação proveniente do entorno de tal modo que um erro em
decodificação pode ocasionar na escolha de um estado inapropriado e, logo, na morte do
organismo. Assim, podemos interpretar a variável aleatória V(v) como a função

29
indicadora de erro que necessitávamos. Desse modo, a entropia da variável aleatória V
pode ser vista como a entropia do equívoco11
ou acidente12
.
3.1.
Assim, percebe-se uma relação entre a entropia da variável aleatória e a incerteza
no processo regulatório .
Entre as possíveis relações algébricas que podem se seguir do processo
regulatório, percebe-se que H(D|R,V=0)=0. Ou seja, dado que o organismo teve certa
experiência (R) e continuou vivo (V=0), não há qualquer incerteza quanto à mensagem
recebida. Assim, podemos assumir que o SNC decodificou corretamente a informação
proveniente do entorno. De outro modo,
3.2.
onde 1, do lado direito da inequação, consiste num valor que define a mensagem
erroneamente decodificada pelo regulador13
, L consiste em todos as mensagens
possíveis do alfabeto da fonte D, exceto aquela apontado erroneamente por R e no
probabilidade de erro. Ou seja, dada certa experiência (R) e a morte do organismo
(V=1), podemos dizer que o organismo se equivocou acerca da informação proveniente
do entorno – onde o equívoco pode ser ‘não perceber’, ou ‘perceber inadequadamente’,
ou ‘perceber adequadamente e não escolher o meio de regulação apropriado’, etc..
A questão que nos ocupa pode ser expressa pela seguinte fórmula H(D,V|R). Ou
seja: dado certa representação ou percepção, qual a probabilidade de que não se esteja
incorrendo em erro – ou qual a probabilidade de que minha representação interna seja
um modelo adequado, onde ‘adequado’ deve ser entendido no sentido estrito de ‘que me
mantenha vivo’? Algebricamente podemos inferir o seguinte dessa probabilidade
condicional:
3.3.
11
O termo ‘equívoco’ aqui não ocorre com o mesmo sentido dado por Shannon, o qual entende
‘equívoco’ como a entropia condicional na expressão da informação mútua; ou seja: I(X;Y)=H(X)-
H(X|Y). 12
A notação se mantém semelhante àquela usada por Massey (1998). 13
O logaritmo de base 2 aqui se requer pela definição de entropia.

30
Notemos que H(V|D,R)=0, pois, uma vez que se tenha conhecimento do conteúdo da
percepção, e da informação do entorno, V assumirá um valor determinado14. Resultando
assim em:
[Lema 3.1.]
o que vem a ser bastante intuitivo.
Porém, e também por necessidade algébrica, da pergunta pela relação entre
nosso modelo interno e o real, obtemos a seguinte relação:
3.4.
3.5.
3.6.
A expressão 3.5. é especialmente útil, pois, pelo resultado 3.1. e 3.2. obtemos o seguinte
teorema:
[Lema de Fano
3.3.]
Podemos interpretar a inequação 3.3. como a afirmação de que o limite mínimo de erro
ou engano que pode haver entre o regulador e a informação proveniente do entorno é
dado por H(D|R). Porém, notemos que H(D|R), quando não for nulo, apenas firma que
há uma incerteza quanto a D; de outro modo, nada é afinado sobre a natureza dessa
incerteza ou como reduzi-la.
No entanto, notemos que I(D;R)=I(R;D), por necessidade algébrica, e logo que
3.7.
3.8.
Substituindo 3.8. na expressão dada pelo Lema de Fano obtemos o famoso Teorema da
Variedade Requerida, devido a Ross Ashby (1956),
[Teorema da Variedade
Requerida 3.] 14
Note que nesse ponto, não é comprometedor se certas relações parecerem demasiado especulativos, tais
como ‘a relação entre o estado mental (R) e a informação exterior ao sistema (D)’, pois essa relação surge
por necessidade algébrica, donde seus valores quantitativos podem ser obtidos a partir de outras variáveis
– tais como V e D.

31
Agora, o Teorema da Variedade Requerida mostra que a única forma de fazermos com
que o erro H(V)= diminua consiste em aumentar a informação dentro do sistema
H(R); ou seja, fazer com que a relação entre o entorno, D, e o modelo interno, R, seja o
mais próximo de uma bijeção. De outro modo, o teorema deixa claro que, a existência
de acidentes pode ser tomada como um indicador de que o léxico do regulador H(R) é
menor do que o léxico do entorno, tal que H(D|R) tenha valor positivo. Dizemos ‘um
indicador’, pois, mesmo que houvesse uma bijeção entre o modelo interno da realidade
e a ‘realidade’, poderiam haver incertezas com relação a precisão com que R distingue
cada ‘símbolo15
’ emitido pela fonte D.
Por fim, pode-se ter a impressão de o argumento só serviria para representar
situações imediatas de regulação, excluindo, assim, a dimensão de planejamento e
expectativa em longo prazo. Porém, o argumento não restringe a latência no tempo de
transmissão, onde a única coisa que se requer para que um sinal seja transmitido através
do tempo, é memória (LERNER, 1975).
Contudo, devemos buscar evidências que deem suporte para nosso argumento
teórico, pois, um argumento matemático, se em desacordo com a realidade, consiste na
forma mais exuberante de retórica – elegante, difícil e convincente. Assim, para que os
resultados oferecidos pelo Teorema da Variedade Requerida tenham o direito de
vangloriar-se de seus louros é preciso haver evidências que os confirmem.
4.2. Ilusões: quando o modelo falha
Ilusões são casos clássicos de erro no processamento de informação do SNC.
Nos últimos tempos, e com a ascensão das ciências cognitivas, descobriu-se, e vem
descobrindo-se, que nossa percepção da realidade é repleta de ilusões (GREGORY,
1968, 2009; FRITH, 2006; EAGLEMAN, 2011). Alguns exemplos de ilusões serão
mostrados para ilustrar como esses casos de erro se conectam com as dificuldades
existentes no processamento de informação pelo SNC apontadas previamente. No
entanto, o ponto central sobre ilusões será o de que aquilo que entendemos por ilusões
são erros em processamento que se tornam perceptíveis por exibirem incoerência com
um contexto mais amplo de conhecimento, o que sugere fortemente que: i) nossa
percepção da realidade tem uma carga muito maior de ilusões do que podemos verificar
15
A palavra ‘símbolo’ deve ser entendida no sei mais amplo sentido como qualquer coisa que possa afetar
o sistema; seja um comprimento de onda, ou uma quantidade de força, etc..

32
de forma consciente e ii) que essas ilusões, em muitos casos, podem ser vistas como
falsidades convenientes.
De acordo com Gregory (1968), nossas ilusões podem ser divididas entre três
tipos de ilusões, a saber, ilusões óticas, sensoriais e perceptuais. Ilusões óticas seriam
aquelas decorrentes da quantidade excessiva de ruído no sinal proveniente do entorno,
tais como miragens e ilusões de reflexão e refração16
. Ilusões sensoriais seriam aquelas
decorrentes de ruído e limitações na transmissão e processamento no sinal interno ao
sistema. Por exemplo, o Efeito Cascata [Figura 11] decorrente da atividade
desequilibrada dos neurônios detectores de movimento no sistema visual, em virtude da
exposição prolongada ao estímulo, faz com que vejamos algo que é impossível:
movimento sem mudança de posição (GREOGRY, 1961; EAGLEMAN, 2012; FRITH,
2007). As ilusões perceptuais seriam aquelas decorrentes de inferências mal sucedidas
baseadas em conhecimento prévio. Ilusões perceptuais consistem nos casos mais
interessantes, por serem as mais ubíquas no processo de percepção, e difíceis de
detectar.
Ilusões perceptuais abrangem todas nossas categorias representacionais
(GREGORY, 2009). Por exemplo, nós experimentamos objetos menores até sessenta
por cento mais pesados do que objetos maiores na mesma escala de peso (GREGORY,
1968). Ou seja, de acordo com uma densidade inesperadamente maior ou menor se sofre
uma ilusão de peso. Nossa percepção visual é determinantemente afetada por
informações prévias tais como, simetria e profundidade. Na ilusão do Quarto de Ames
16
É interessante notar que o grau de equívoco concernente a sinais transduzidos por canais térmicos ou
acústicos, por exemplo, é tão grande que se torna redundante estender o termo para essas modalidades dos
sentidos.
Figura 8 - Efeito Cascata. Ao girar a imagem se perceberá a espiral diminuindo ou aumentando, de acordo
com a direção do giro, após algum tempo, ao contemplar-se a imagem parada, ter-se-á a ilusão de que ela está
aumentando ou diminuindo.

33
[Figura 12] a perspectiva simétrica cria uma ilusão de que as pessoas dentro do quarto
de Ames têm tamanhos distintos. Informações interpretadas pelo cérebro como
Figura 9 Figura 9 Quarto de Ames – todos os homens no quarto são do mesmo tamanho, porém em
proximidades distintas do observador.
indicando profundidade também conduzem a inferências errôneas acerca do tamanho
dos objetos. Assim, a ilusão de Ponzo [Figura 13], na qual a impressão de convergência
em linhas paralelas resulta na percepção de que os objetos de tamanho igual sejam
vistos com tamanhos diferentes. A informação prévia também influencia no modo como
percebemos a intensidade no brilho das cores [Figura 14] (ANDERSON & WINAWER,
2005).
Os casos aqui mencionados são apenas alguns entre uma infinidade de casos
conhecidos hoje (GREGORY, 2009; EAGLEMAN, 2011; FRITH, 2007). A existência
de ilusões, em si, já consiste numa evidência de erro no emulador, assim como numa
prova de que o sistema de regulação, de fato, criaria um modelo do real, uma vez que se
tem a experiência, persistente em muitos casos, claramente incoerente com o restante do
contexto de informação. A incoerência flagrante nos casos de ilusão pode nos conduzir

34
a perceber um importante aspecto no que se refere ás ilusões.
Pois, como se podem distinguir aquelas experiências ditas ilusórias daquelas entendidas
como uma interpretação adequada dos eventos exteriores? Perceber-se-á que o que
entendemos por ilusão tipicamente se define pelas experiências que demonstram
incoerência com um episódio instantâneo de percepção, ou que se torna incoerente na
medida em que a experiência se desenvolve no tempo. A experiência do Efeito Cascata,
talvez uma das primeiras que se tem registro – tendo sido mencionada por Aristóteles –,
demonstra flagrante incoerência com nosso conhecimento filogenético acerca do mundo
– que não há movimento sem mudança de posição. O Quarto de Ames se revela como
uma ilusão tão logo o espectador mudar de perspectiva, tornando-se capaz de perceber
as assimetrias do quarto. As ilusões referentes ao peso dos objetos e a ilusão de Ponzo
requerem mecanismos de medição, por meio dos quais possamos comparativamente
identificá-las como ilusão. E, no caso mais difícil, a ilusão do brilho das peças de xadrez
exige que as peças sejam recortadas e comparadas, alheias ao ambiente, para que se
possa convencer de que, de fato, trata-se de peças com o mesmo brilho. Nesse mesmo
sentido, uma série ilusões cuja identificação depende de engenhosos experimentos são
paulatinamente descobertas (NIJHAWAN, 2008; GREGORY, 2009).

35
Figura 11 – Os dois conjuntos de peças de Xadrez em ambos os ambientes são idênticos. Em ambos os casos, as
figuras sobre ambiente escuro aparecem como objetos claros visíveis através da névoa escura, enquanto as
figuras sobre o ambiente claro aparecem como objetos escuros através da névoa clara.
O que se deve perguntar é: não poderia nossa percepção da realidade ser em
larga medida ilusória não havendo comprometimento, em muitos casos, do processo de
regulação do SNC? Baseado sobre o que vimos até então, nossa hipótese é a de que
experimentamos ilusões a todo instante. Se aceitarmos que a falha em representar
semelhantes experiências – mas não idênticas! – como distintas deve ser entendida
como uma ilusão, então nossa hipótese parece seguramente verdadeira. Pois, o que se
tem verificado é que o conhecimento prévio acerca de experiências passadas (memória)
é antes uma característica ubíqua de nossa percepção, tal que as experiências prévias
preparam as futuras experiências – ‘priming perception’ (BERNSTEIN et al. 1989;
TULVING & SCHACTER 1990; RATEY, 1994; KAWAKAMI et al. 2003). Esse
preparo em nossa percepção consiste em experimentar novos estímulos como estímulos
passados, de tal modo que as dessemelhanças são eliminadas de nosso modelo interno.
No entanto, provavelmente seria um exagero entendermos essa característica, de
preparar nossas futuras experiências, como um defeito no processamento de informação.
Pode-se conjeturar que o SNC tem algo a ganhar nessa simplificação, tal que essa ilusão
ubíqua seja pragmaticamente desejável. De fato, se levarmos em consideração que,
enquanto organismos biológicos, somos um sistema aberto em constante troca de
energia e matéria com o entorno, o que ocasiona uma constante dinâmica em nossa
constituição física, e que assim também o são todos os objetos interessantes para a vida
humana, tal capacidade de discriminação pelo SNC ocasionaria a exigência de uma

36
desnecessária sobrecarga no processamento de informação17
. Isso vem a ser bastante
plausível uma vez que essas dessemelhanças, além de dificultarem o processamento de
informação, são irrelevantes para o processo de regulação e, logo, não devem ser
consideradas.
5. Implicações Filosóficas
Antes de sugerirmos algumas implicações filosóficas que parecem resultar dessa
perspectiva, lembremos o caminho percorrido para chegarmos até aqui. Na primeira
parte do presente texto, introduzimos a noção de sistema aberto cujo comportamento se
define, de forma determinada ou não, com relação à informação proveniente do entorno.
Essa noção serviu para que pudéssemos vislumbrar o tipo de sistema característico do
SNC, cujo único meio de afetar seu entorno consiste em motricidade.
Subsequentemente, e depois de introduzirmos alguns conceitos de Teoria da
Informação, vimos que nós, enquanto organismo biológico, também poderíamos ser
vistos como um sistema em estado de equilíbrio estável, e chamamos esse estado de ‘o
organismo vivo’. Vimos que a manutenção desse estado requeria um subsistema de
regulação e controle, identificando, assim, a função do SNC no organismo vivo que
somos. No que se seguiu, passamos a investigar quais eram os requerimentos que se
colocavam para o SNC pudesse levar a cabo o processo de regulação com sucesso, e
vimos que esse sistema requeria memória e algumas estratégias para contornar o ruído
no processamento de informação, entre as quais figurava a elaboração de um modelo
interno do real. O modelo interno do real foi entendido como aquilo que nós
compreendemos intuitivamente como realidade. Passamos, então, a investigar o grau de
acuidade desse modelo interno. Primeiro expusemos um argumento teórico, com uso de
teoria da informação, o qual afirmava que a existência de erro, ou enganos, deflagrava o
grau de desacordo entre o modelo e o entorno. Em seguida, e na busca por evidências,
passamos a analisar as ilusões características do processo de percepção, as quais
sugeriram fortemente que nossa noção de realidade consiste num modelo com certo
grau de erro, o que nem sempre deve ser vistos de forma indesejada.
Essa perspectiva parece-nos ser demasiada revolucionária em filosofia, pois abre
caminho para entendermos diferentemente uma série de temas filosóficos. Entre esses
figuram, por exemplo, temas como a natureza da racionalidade humana, o papel que
17
Imaginemos os problemas que haveriam para identificar um organismo, em dois instantes diferentes,
como sendo o mesmo.

37
teorias científicas comprem em nossa interação com o entorno, um critério para
distinção entre epistemologia e ontologia. Porém, em particular, essa perspectiva, se
aceita, parece ser fatal para aquelas formas de reflexão que se aglutinam sob o nome de
Realismo Semântico. Uma concepção realista semântica, em linhas gerais, apoia-se
sobre a noção de verdade como correspondência com a realidade para justificar o
significado de termos linguísticos ou as inferências realizadas com sucesso pelas teorias
científicas (SHALBOWSKI, 1995; BROWN, 1982; GAMBLE, 2002; PUTNAN 1975).
Entende-se que a unidade no significado das expressões linguísticas ou a justificação no
sucesso da ciência está ancorada na relação que se estabelece entre as entidades
linguísticas, de um lado, e a realidade dotada das propriedades e entidades, de outros. A
pergunta que se deve fazer é: que realidade é essa? De nosso ponto de vista, enquanto
seres humanos o único sentido de realidade que nós conhecemos é aquele elaborado
pelo SNC para controle motor. Mas nós evidenciamos que nosso cérebro constrói um
modelo do real, o qual é, em larga medida, ilusório e que essa característica, em muitos
casos, não seria de todo condenável. Baseado nisso, nem a unicidade do significado das
expressões linguísticas, nem o sucesso da ciência podem ser justificados com relação a
nossa realidade, uma vez que a relação é, em muitos casos, ilusória. Antes, seria um
milagre que a ciência tivesse o sucesso que, de fato, tem, estivesse ela baseada nessa
relação.
De nosso ponto de vista, linguagem, significado, realidade – no sentido robusto
empregado pelo realista – e a relação que se estabelece entre ambos figuram dentro o
espaço neural. Mas isso não quer dizer que estejamos endossando uma concepção
idealista – lembre-se, somos um sistema aberto cuja estabilidade depende da troca
contínua de informação com o entorno – mas que insistir nesse modelo representacional
consiste em perseguir o próprio rabo. Mais especificamente, significado e referência,
por exemplo, consistem, em última instância, na relação que se estabelece entre um
padrão de ação neural concomitante entre redes neurais nas áreas de Wernicke, Broca e
no sistema visual no lóbulo occipital, realizada por longos axônios conectando essas
áreas. Mas se isso for o caso, a ocorrência de danos neurais nessas áreas devem
comprometer essas capacidades. E é exatamente o que acontece quando indivíduos
sofrem danos neurais nas áreas responsáveis pela linguagem – área de Wernicke e
Broca – havendo severo comprometimento nas capacidades linguísticas
(GESCHWIND, 1970, 1974; FADIGA et al. 2009; RATEY, 1994).

38
A menção ao realismo semântico, aqui, não tem o objetivo de figurar como uma
argumentação contra essa concepção – ainda que o exposto pareça oferecer suporte para
tal – o que nos levaria longe demais no presente texto. Mas serve para mostrar o
contraste que há entre uma concepção representacionalista do mente e uma concepção
interacionista, e como elas comprometem nossas estratégias de conceitualização de
diferentes domínios. Em suma, não é que nossa visão da realidade não seja adequada –
não há com o que comparar. A tese é ainda mais fundamental. É que pensar a realidade
em termos representacionais é um modo antropomórfico de pensar essa relação. Isso
não quer dizer que devamos parar de buscar uma, ou diferentes, representações da
realidade para fins pragmáticos – na verdade, esse é o melhor mecanismo que temos
para interagir com a ‘realidade’ com sucesso. É que, filosoficamente, seguir esse
caminho consiste em confundir epistemologia e ontologia e tomar a primeira como
sendo a segunda. Contudo, o filósofo da ciência nos diria que a realidade com a qual ele
lida nunca foi essa realidade particular de nossas percepções, mas aquela realidade
abstrata captada pelas teorias científicas. E isso nos levaria a tarefa de examinar como o
cérebro computa informação abstrata e se essas computações têm alguma relação com
essa concepção comum de realidade que tratamos aqui. Mas isso é assunto para outra
ocasião.
Bibliografia:
ARBIB, A. M. ‘Perceptual Structures and Distributed Motor Control’. Comprehensive
Physiology, Supplement 2: Handbook of Physiology, The Nervous System, Motor
Control, 1981.
ASHBY, W. R. An Introduction to Cybernetics. London: Chapman & Hall, 1956.
Internet (1999): http://pcp.vub.ac.be/books/IntroCyb.pdf
_____. Design for a Brain. New York: John Wiley & Sons. Inc, 1966.
AZZOUNI, J. ‘A new characterization of scientific theories’, 2010
BERG, H. C. & PURCELL, E. M. ‘Physics of chemoreception’. Journal of Biophysics,
Vol. 20, pp. 193–219, 1977.
BERNSTEIN, I. H., BISSONNETTE, V., VYAS, A., BARCLAY, P. ‘Semantic
priming: subliminal perception or context?’. Perception & Psychophysics, Vol. 45, pp.
153-151, 1989.

39
BIALEK, S. W. & SETAYESHGAR, S. ‘Physical limits to biochemical signaling’.
Proceeding of National Academy of Science of the United States of America. May, pp.
10040–10045, 2005.
BROWN, J. R. ‘The Miracle of Science’. The Philosophical Quarterly, Vol. 32, pp.
232-244, 1982.
CLONEY, R. ‘Ascidean Larvae and the Events of Metamorphosis’. American
Zoologist, Vol. 22, No. 4, pp. 817-826, 1982.
CHURCHLAND, P. S., RAMACHANDRAN, V. S., & SEJNOWSKI, T. J. ‘A Critics
of Pure Vision’. Em: Large-Scale Neuronal Theories of the Brain. Cambridge,
Massachusetts: The MIT Press, 1994.
COVER, T. M. & THOMAS, J. A. Elements of Information Theory. Hoboken: New
Jersey, John Wiley & Sons, inc., 2006.
DEBANNE, D. ‘Information processing in the axon’. Nature Review Neuroscience.
Vol. 5, pp. 304–316, 2004.
DESTEXHE, A. & RUDOLPH-LILITH, M., Neuronal Noise. New York: Springer,
2012.
DESMURGET, M. & GRAFTON, S. ‘Forward modeling allows feedback control for
fast reaching movements’. Trends in Cognitive Science, Vol. 11, pp. 423-31, 2000.
DIBA, K., LESTER, H. A. & KOCH, C. ‘Intrinsic noise in cultured hippocampal
neurons: experiment and modeling’. Journal of Neuroscience. Vol.24, pp. 9723–9733,
2004.
DOYA, K., ISHII, S., POUGET, A., RAO, R. P. N. Bayesian Brain: probabilistic
approach to neural coding. Cambridge, Massachusetts: The MIT Press, 2007.
EAGLEMAN, D. Incognito: the secret lives of the brain. London: Canongate, 2011.
FADIGA, L., GRAIGHERO, L.,D’AUSILIO, A. ‘Broca’s Area in Language, Action,
and Music’. The Neuroscience and Music III – Disorder and Plasticity: Ann. N.Y. Acad.
Sci., Vol. 1169, pp. 449-458, 2009.
FAISAL, A. A., SELEN, L. P. J., WOLPERT, D., ‘Noise in the nervous system’.
Nature Review Neuroscience, Vol. 9, pp. 292-303, 2008.
FAISAL, A. A., WHITE, J. A. & LAUGHLIN, S. B. ‘Ion-channel noise places limits
on the miniaturization of the brain’s wiring’. Current Biology. Vol. 15, pp. 1143–1149,
2005.
FELDMAN, G. A. From Molecule to Metaphor: a neural theory of language.
Cambridge, Massachusetts: MIT Press, 2006.

40
FIELD, D. ‘Relations between the statistics of natural images and the response
properties of cortical cells’. Journal of Optic Society of America. A 4, pp. 2379–2394,
1987.
FLANAGAN, J. R. & WING, A. M. ‘The role of internal models in motion planning
and control: evidence from grip force adjustments during movements of hand-held
loads’. Journal of Neuroscience, Vol. 17, pp. 1519-1528, 1997.
FRITH, Christopher D. Making up the Mind: how the brain creates our mental world.
Malden: Blackwell Publishing, 2007.
FUSTER, J. ‘The Cognit: A network model of cortical representation’. International
Journal of Psychophysiology, nº 60, 2006, p. 125-132.
GAMBLE, D. ‘Defending Semantic Realism’. Language and Communication, Vol. 22,
pp. 243-258, 2002.
GESCHWIND, N. Selected Papers on Language and the Brain. Boston: D. Reidel
Publishing Company, 1974.
______. ‘The Organization of Language and the Brain’. Science, Vol. 170, pp. 940-944,
1970.
GIBSON, J. J. The senses considered as perceptual system. Boston: Houghton Mifflin,
1966.
_____. The ecological approach to visual perception. New York: Psychology Press,
1986.
GREGORY, R. L. ‘Perceptual illusions and brain models’. Proceedings of the Royal
Society B, Vol. 171, pp. 279-296, 1968.
______. Seeing Through Illusions. London: Oxford, Oxford University Press, 2009.
GRUSH, R. ‘The emulation theory of representation: Motor control, imagery, and
perception’. Behavioral and Brain Science, pp. 377-442, 2004.
HAKEN, H. Information and Self-Organization. 2th ed.. Springer, 1999.
HARRIS, G. G. ‘Brownian Motion in the Cochlear Partition’. Journal of the Acoustical
Society of America. Volume 44, Issue 1, pp. 176-186, 1968.
HELD, R. ‘Plasticity in sensory-motor systems’. Scientific America, 212: pp. 84-94.
HULL, J. M. Touching the Rock: an Experience of being Blind. London: S.P.C.K.,
1990.
JACOBS, R. A. ‘Optimal integration of texture and motion cues to depth’. Vision Res.
Vol. 39, pp. 3621–3629, 1999.
JAMES, W. The Principals of Psychology. London: Henry Holt, 1890.

41
JOLIOT M., RIBARY, U., & LLINAS, R. ‘Human oscillatory brain activity near 40 Hz
coexists with cognitive’. Em: Neurobiology, Proc. Nati. Acad. Sci. USA Vol. 91, pp.
11748-11751, November 1994
JORDAN, M. I. & RUMELHART, D. E. ‘Forward models: supervised learning
with a distal teacher’. Cogn Sci, Vol. 16, pp. 307-354, 1992.
KANDEL, E. The Agen of Insite: the quest to understand the unconscious in art, mind
and brain, from Vienna 1900 to the present. New York: Handle House, 2012.
KANDEL, E. R., SCHWARTZ, J. H. & JESSELL, T. M. Principles of Neural Science.
McGraw–Hill/Appleton & Lange, New York, 2000.
KAWAKAMI, K., DOVIDIO, J. F., DIJKSTERHUIS, A. ‘Effect Social Category
Priming on Personal Attitudes’. Psychological Science, Vol. 14, pp. 315-319, 2003.
KAWATO, M. ‘Internal model for motor control and trajectory planning’. Current
Opinion in Neurobiology, nº 9, pp. 718-727, 1999.
KNILL, D. C. ‘Mixture models and the probabilistic structure of depth cues’. Vision
Res. Vol.43, pp. 831–854, 2003.
KOHLER, I. ‘Experiments with Goglles’, Scientific America, 205: pp. 62-72.
KORDING, K. P. & WOLPERT, D. M. ‘Bayesian integration in sensorimotor
learning’. Nature, 427, pp. 244–247, 2004.
KOZLOV, A. S., RISLER, T. & HUDSPETH, A. J. ‘Coherent motion of stereocilia
assures the concerted gating of hair-cell transduction channels’. Nature Neuroscience.
Vol. 10, pp. 87–92, 2007.
LAUGHLIN, S. ‘A simple coding procedure enhances a neuron’s information capacity’.
Z. Naturforsch. [C] 36, pp. 910–912, 1981.
LERNER, A. Ya. Fundamentals of Cybernetics. London: Plenum Press, 1975.
LLINAS, R. The I of the Vortex: from neuron to self. Cambridge, Massachusetts: The
MIT Press, 2001.
_____. ‘Umwelt: A Psychomotor Functional Event’. Em: Neurobiology of “Umwelt”:
How Living Beings Perceive the World. Berlin Heidelberg: Springer, pp 29-37, 2009.
LLINÁS, R. & PARÉ, D. ‘El Cerebro como un Sistema Cerrado Modulado por los
Sentidos’. Em: LLINÁS, R. & CHURCHLAND, P. El Continuum Mente-cerebro:
procesos sensoriales. Bogotá: Universidad Nacional de Colombia, 2006.
______. ‘Of Dreaming and Wakefulness’. Neuroscience, Vol. 44, No. 3, pp. 521-535,
1991.

42
LLINÁS, R., RIBARY, U., JOLIOT, M., WANG, X.-J. ‘Content and context in
temporal thalamocortical binding’. Temporal coding in the brain. Berlin : Springer-
Verlag, page: 251, stat: Chapter, 1994.
LLINÁS, R. & ROY, S. ‘The ‘prediction imperative’ as the basis for self-awareness.
Philosophical Transactions of Royal Society, Biological Science, 364, pp. 1301-1307,
2009.
MA, W. J., ZHOU, X., ROSS, L. A., FOXE, J. J., PARRA, L. C. ‘Lip-Reading Aids
Word Recognition Most in Moderate Noise: A Bayesian Explanation Using High-
Dimensional Feature Space’. PLoS ONE 4(3): e4638.
doi:10.1371/journal.pone.0004638, 2009.
MAINZER, K. Thinking in Complexity. 6th
ed.. Berlin: Springer-Verlag, 2007.
MESULAM, M.-M. ‘A cortical network for directed attention and unilateral neglect’.
Annals of Neurology, Vol. 10, pp. 309-325, 1981.
NIJHAWAN, R. ‘Visual Prediction: psychophysics and neuropsychology of
compensation for time delays. Em: Behavioral and Brain Sciences, nº31, pp. 179-239,
2008.
MERFELD, D. M., ZUPAN, L., PETERKA, R. J. ‘Humans use internal models to
estimate gravity and linear acceleration’. Nature, Vol. 398, pp. 615-618, 1999.
MIALL, R. C., WEIR, D. J., WOLPERT, D. M., STEIN, J. F. ‘Is the cerebellum a
Smith Predictor?’. Journal of Motor Behavior, Vol. 25, pp. 203-216, 1993.
PIRENNE. M. H. ‘Some aspects of the sensibility of the eye’. Annals of the New York
Academy of Science. Vol. 74, pp. 377-384, 1959.
PITTS, W. C. & MCCULLOCH, W. S. ‘How we know universals, the perception of
auditory and visual forms’. Bull. Math. Biophys. 9: 127-147, 1947.
PRIGOGINE, I. Introduction to Thermodynamics of Irreversible Processes (2th Ed.).
New York: John Wiley & Sons, 1961.
PRIGOGINE, I. & NICOLIS, G. Self-Organization in Non-Equilibrium Systems. New
York: John Wiley & Sons, 1977.
PUTNAM, H. Philosophical Papers: Vol. I. Cambridge: Cambridge University Press,
1975.
RALL, W. ‘Time constants and electrotonic length of membrane cylinders and
neurons’. Journal of Biophysics. Vol. 9,
pp. 1483–1508, 1969.

43
ROSENBLUETH, A.; WIENER, N.; BIGELOW, J. ‘Behavior, Purpose and Teleology’.
Phylosophy of Science, Volume 10, Issue, p. 18-24, 1943.
SINGER, W. ‘The Brain’s View of the World Depends on What it has to Know’. Em:
Berthoz, A. & Christen, Y. (eds.), Neurobiology of “Umwelt”: How Living Beings
Perceive the World. Research and Perspectives in Neurosciences, Springer-Verlag
Berlin Heidelberg, 2009.
SHADLEN, M. N. & NEWSOME, W. T. ‘The variable discharge of cortical neurons:
implications for connectivity, computation, and information coding’. Journal
Neuroscience. Vol. 18, pp. 3870–3896, 1998.
SHADMEHR, R. & MUSSA-IVALDI, S. Biological Learning and Control: how the
brain builds representations, predicts events, and makes decisions. Cambridge,
Massachusetts: The MIT Press, 2012.
SHALBOWSKI, S. A. ‘Semantic Realism’. The Review of Metaphysics, Vol. 48, pp.
511-538, 1995.
SHANNON, C. & WEAVER, W. The Mathematical Theory of Communication. The
University of Illinois Press – Urbana, 1964.
RATEY, J. The User’s Guide to the Brain: perception, attention and the four theaters of
the brain. New York: Little, Brown and Company, 1994.
SNYDER, L. ‘This way up: illusions and internal models in the vestibular system’.
Nature Neuroscience, Vol. 2, pp. 396-398, 1999.
SOFTKY, W. R. & KOCH, C. ‘The highly irregular firing of cortical cells is
inconsistent with temporal integration of random EPSPs’. Journal Neuroscience.
Vol.13, pp. 334–350, 1993.
SPERRY, R. W. ‘Neurology and The Mind-Brain Problem’. American Scientist, Vol.
40, nº2, April, 1952.
STEIN, R.; GOSSEN, E. R.; JONES, K. E. ‘Neuronal Variability: noise or part of the
signal?’. Nature Review: neuroscience. Nature Publishing Group, Vol.6, May, 2005.
STERIADE, .M & MACCARLEY, R. W. Brain Control of Wakefulness and Sleep.
New York: Plenum Press, 1990.
STERIADE, M., CURRÓDOSSI, R., PARÉ, D., OAKSON, G. ‘Fast oscillations (20–
40 Hz) in thalamocortical systems and their potentiation by mesopontine cholinergic
nuclei in the cat’. Proc Natl AcadSci USA 88: pp. 4396–4400, 1991.
STEWART, I. & COHEN, J. Figments of Reality: the evolution of curious mind.
Cambridge: Cambridge University Press, 1997.

44
STOCKER, A. A. & SIMONCELLI, E. P. ‘Noise characteristics and prior expectations
in human visual speed perception’. Nature Neuroscience. Vol. 9, pp. 578–585, 2006.
TULVING, E. & SCHACTER, L. ‘Priming and Human Memory System’. Science, Vol.
247, pp. 301-306, 1990.
VAN FRAASSEN, B. The Scientific Image. 1980.
WIENER, N. The Human Use of Human Beings: cybernetics and society. London: Free
Association Books, 1989.
_____. Cybernetics or Control and Communication in the Animal and the Machine.
Cambridge, Massachusetts: The M.I.T. Press, 1985.
WOLPERT,D.http://www.ted.com/talks/daniel_wolpert_the_real_reason_for_brains.ht
ml
WOLPERT, D. & GHAHRAMANI, Z. ‘Bayes rule in perception, action and cognition’.
http://learning.eng.cam.ac.uk/Public/Wolpert/Publications, 2009.
WOLPERT, D., GHAHRAMANI, Z. & JORDAN, M. I. ‘An internal modelo for
sensoriomotor integration’. Science, Vol. 269, pp. 1880-82, 1995.
WOLPERT, D., DOYA, K., & KAWATO, M., ‘A unifying computational framework
for motor control and social interaction’. Em: The Neuroscience of Social Interaction:
decoding, imitating, and influencing the actions of the others. FRITH, C. &
WOLPERT, D. (ed.). New York: Oxford University Press, 2003.




















