![Survival of Black-headed Gulls Larus ridibundus wintering in urban areas in The Netherlands [in Dutch, with English summary] (Overleving van overwinterende Kokmeeuwen in Nederlandse](https://static.fdokumen.com/doc/165x107/6316f36bd16b3722ff0d2193/survival-of-black-headed-gulls-larus-ridibundus-wintering-in-urban-areas-in-the.jpg)
A ptilochronological study of carry-over effects of conditions during wintering on breeding...
12
513 A ptilochronological study of carry-over effects of conditions during wintering on breeding performance in the barn swallow Hirundo rustica Nicola Saino, Maria Romano, Manuela Caprioli, Roberto Ambrosini, Diego Rubolini, Chiara Scandolara and Andrea Romano N. Saino ([email protected]), M. Romano, M. Caprioli, D. Rubolini, C. Scandolara and A. Romano, Dipartimento di Biologia, Univ. degli Studi di Milano, via Celoria 26, IT-20133 Milano, Italy. – R. Ambrosini, Dipartimento di Biotecnologie e Bioscienze, Univ. degli Studi di Milano Bicocca, Piazza della Scienza 2, IT-20126 Milano, Italy. e ecological conditions that a bird experiences during any stage of its life cycle may have consequences that become man- ifested at later life stages, and these ‘carry-over effects’ may be major components of variance in individual performance. Condition-dependent feather growth rate, as assessed by growth bars width (GBW), provides a unique, though largely under-exploited tool to investigate carry-over effects of ecological conditions and individual physiological state during molt. In this study of breeding barn swallows Hirundo rustica, which undergo a single complete annual molt of tail and wing feathers during wintering in sub-Saharan Africa, we first show that old ( 2 yr) females have larger GBW than old males and yearlings. GBW was smaller with larger infestations by common ectoparasites of barn swallows, independent of the swallows’ age or sex, and larger GBW was associated with a higher index of body condition during the breeding season in males. Larger GBW predicted higher seasonal reproductive output of older males, but not repro- ductive output of younger males or females of any age class, with this higher reproductive output of older males mediated by higher offspring fledging success. However, no relationship with GBW was observed for seasonal reproductive output of males in the spring preceding the winter when the feathers were grown. Hence, this study suggests that the analysis of the rate of feathers growth (‘ptilochronology’) has a larger potential to serve as a powerful tool in the study of carry-over effects than has been appreciated to date. Specifically, the present results support the idea that conditions experienced during wintering in Africa and proximately reflected by GBW have carry-over effects on body condition and breeding suc- cess. ese effects are sex and age specific, being more pronounced in older males, possibly as a consequence of differences in annual time routines and susceptibility to extrinsic factors among sex and age classes. Journal of Avian Biology 43: 513–524, 2012 doi: 10.1111/j.1600-048X.2012.05622.x © 2012 e Authors. Journal of Avian Biology © 2012 Nordic Society Oikos Subject Editor: Wesley M. Hochachka. Accepted 10 July 2012 Variation in the environmental conditions experienced by animals as they move through space and time can have immediate consequences on their state and viability but also delayed, far-reaching consequences at later life-stages (Sutherland 1996, Lindström 1999, Harrison et al. 2011). At the individual level, life-history components are expected to be reciprocally linked by constraining relationships, because of limitation of the resources that can be allocated to competing functions, resulting in a negative covariation between fitness traits (Roff 1992, Stearns 1992). Ample variation in resource availability and internal state caused by environmental or genetic heterogeneity, however, may differentially relax such constraining relationships eventu- ally resulting in a positive, rather than a negative covaria- tion between fitness components among individuals (van Noordwijk and de Jong 1986, Zera and Harshman 2001). e consequences of benign environmental conditions may thus be expressed as simultaneous increases of phenotypic values at fitness traits and/or as ‘ carry-over effects’ whereby favorable conditions at any given time allow for larger allocation to important life-history traits both at present and future times (Norris 2005, Inger et al. 2010, Legagneux et al. 2011, Stutchbury et al. 2011). While their existence makes considerable intuitive as well as physiological sense, our knowledge of carry-over effects has been hampered by the difficulties of tracking birds and other vagile animals through different stages of their lives, but also by lack of ad hoc research efforts (see Harrison et al. 2011 for a review). In migratory birds, carry-over effects have been inferred, for example, at the population level by comparing mean arrival date or breeding performance in springs following different conditions on the wintering or breeding grounds (Gill et al. 2001, Saino et al. 2004a, b, Gunnarsson et al. 2005, 2006). Analyses at the individual level are still rare because of lack of knowledge of individual state and environment during either annual phase (wintering or breeding) (Norris et al. 2004, Robb et al. 2008, Sedinger et al. 2011). Although carry-over effects are likely to account
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A ptilochronological study of carry-over effects of conditions during wintering on breeding...

513
A ptilochronological study of carry-over effects of conditions during wintering on breeding performance in the barn swallow Hirundo rustica
Nicola Saino , Maria Romano , Manuela Caprioli , Roberto Ambrosini , Diego Rubolini, Chiara Scandolara and Andrea Romano
N. Saino ([email protected]), M. Romano, M. Caprioli, D. Rubolini, C. Scandolara and A. Romano, Dipartimento di Biologia, Univ. degli Studi di Milano, via Celoria 26, IT-20133 Milano, Italy. – R. Ambrosini, Dipartimento di Biotecnologie e Bioscienze, Univ. degli Studi di Milano Bicocca, Piazza della Scienza 2, IT-20126 Milano, Italy.
Th e ecological conditions that a bird experiences during any stage of its life cycle may have consequences that become man-ifested at later life stages, and these ‘ carry-over eff ects ’ may be major components of variance in individual performance. Condition-dependent feather growth rate, as assessed by growth bars width (GBW), provides a unique, though largely under-exploited tool to investigate carry-over eff ects of ecological conditions and individual physiological state during molt. In this study of breeding barn swallows Hirundo rustica , which undergo a single complete annual molt of tail and wing feathers during wintering in sub-Saharan Africa, we fi rst show that old ( � 2 yr) females have larger GBW than old males and yearlings. GBW was smaller with larger infestations by common ectoparasites of barn swallows, independent of the swallows ’ age or sex, and larger GBW was associated with a higher index of body condition during the breeding season in males. Larger GBW predicted higher seasonal reproductive output of older males, but not repro-ductive output of younger males or females of any age class, with this higher reproductive output of older males mediated by higher off spring fl edging success. However, no relationship with GBW was observed for seasonal reproductive output of males in the spring preceding the winter when the feathers were grown. Hence, this study suggests that the analysis of the rate of feathers growth ( ‘ ptilochronology ’ ) has a larger potential to serve as a powerful tool in the study of carry-over eff ects than has been appreciated to date. Specifi cally, the present results support the idea that conditions experienced during wintering in Africa and proximately refl ected by GBW have carry-over eff ects on body condition and breeding suc-cess. Th ese eff ects are sex and age specifi c, being more pronounced in older males, possibly as a consequence of diff erences in annual time routines and susceptibility to extrinsic factors among sex and age classes.
Journal of Avian Biology 43: 513–524, 2012 doi: 10.1111/j.1600-048X.2012.05622.x
© 2012 Th e Authors. Journal of Avian Biology © 2012 Nordic Society Oikos Subject Editor: Wesley M. Hochachka. Accepted 10 July 2012
Variation in the environmental conditions experienced by animals as they move through space and time can have immediate consequences on their state and viability but also delayed, far-reaching consequences at later life-stages (Sutherland 1996, Lindstr ö m 1999, Harrison et al. 2011). At the individual level, life-history components are expected to be reciprocally linked by constraining relationships, because of limitation of the resources that can be allocated to competing functions, resulting in a negative covariation between fi tness traits (Roff 1992, Stearns 1992). Ample variation in resource availability and internal state caused by environmental or genetic heterogeneity, however, may diff erentially relax such constraining relationships eventu-ally resulting in a positive, rather than a negative covaria-tion between fi tness components among individuals (van Noordwijk and de Jong 1986, Zera and Harshman 2001). Th e consequences of benign environmental conditions may thus be expressed as simultaneous increases of phenotypic values at fi tness traits and/or as ‘ carry-over eff ects ’ whereby
favorable conditions at any given time allow for larger allocation to important life-history traits both at present and future times (Norris 2005, Inger et al. 2010, Legagneux et al. 2011, Stutchbury et al. 2011). While their existence makes considerable intuitive as well as physiological sense, our knowledge of carry-over eff ects has been hampered by the diffi culties of tracking birds and other vagile animals through diff erent stages of their lives, but also by lack of ad hoc research eff orts (see Harrison et al. 2011 for a review).
In migratory birds, carry-over eff ects have been inferred, for example, at the population level by comparing mean arrival date or breeding performance in springs following diff erent conditions on the wintering or breeding grounds (Gill et al. 2001, Saino et al. 2004a, b, Gunnarsson et al. 2005, 2006). Analyses at the individual level are still rarebecause of lack of knowledge of individual state and environment during either annual phase (wintering or breeding) (Norris et al. 2004, Robb et al. 2008, Sedinger et al. 2011). Although carry-over eff ects are likely to account

514
for a large proportion of variance in fi tness (Harrison et al. 2011), our ability to capture their impact remains small. On the other hand, traits that are expressed at relatively fi xed times during ontogeny, and particularly those that are periodically renewed like feathers, may provide an inte-grated record of physiological state around the time when the trait is expressed (Bortolotti et al. 2002). For example, diverse forms of perturbation of individual homeostasis during feather molt, and particularly defi cient nutrition, can result in ‘ pathological ’ feather growth, such as forma-tion of fault bars that structurally weaken the feathers and impair aerodynamics (Ritchie et al. 1994, Bortolotti et al. 2002, Jovani and Blas 2004, M ø ller et al. 2009, Romanoet al. 2011). Parasitism and immune response can also aff ect feather growth and molt speed (Ilmonen et al. 2000, Sanz et al. 2004, Moreno-Rueda 2010).
Physiological feather growth processes during natural molt result in a typical transversal feather barring pattern( ‘ growth bars ’ ; Riddle 1907, 1908, Michener and Michener 1938, Wood 1950, Grubb 2006). In several species, rectrices and remiges show a regular succession of light and dark bands, a few millimeters in width, that run (sub-)perpendicular to the rachis (Grubb 1995, Takaki et al. 2001, Clarkson 2011). Th is pattern may vary in dis-tinctness among individuals and also plumage regions, being most evident on rectrices in several taxa (Grubb 1995, Takaki et al. 2001, Clarkson 2011, Saino et al. unpubl.). Th e width of these bands has been used in ptilochrono-logical studies as proxy for feather growth rate, as a pair of light/dark bands (a ‘ growth bar ’ ) has been shown to refl ect a 1-d growth interval in the adults of the species where a distinct alternation of light/dark bands is evident (Michener and Michener 1938, Wood 1950, Grubb 1989, 1995, Brodin 1993, Jovani et al. 2011, Jovani and Diaz-Real 2012; but see Kern and Cowie 2002 for a study of nestlings).
Plumage renewal is a highly demanding process, in terms of allocation of potentially limiting nutrients, as feathers may represent up to 40% of a bird ’ s dry mass and methionine- and cysteine-rich proteins account for 95% of feathers ’ mass (Ginn and Melville 1983, Murphy and King 1992, Lindstr ö m et al. 1993, Murphy 1996). Rapid feather growth is positively selected via the fi tness benefi ts it accrues in terms of retention of thermal insu-lation and effi ciency of fl ight and escape from predators (Nilsson and Svensson 1996, Swaddle et al. 1999, Dawson et al. 2000) but also in sexual selection processes and antiparasite defence (Pennycuick 1989, Andersson 1994, Fitzpatrick 1998, Jovani and Blas 2004, Serrano and Jovani 2005, Newton 2008). Parasite-free individu-als in good nutritional conditions are thus predicted to have larger growth bar width (GBW) by allocating more resources to feather biosynthesis per unit time. As long as growth bars refl ect quanta of daily feather growth, they thus constitute a useful proxy to the ecological milieu and physiological state at the time when the feather was grown. In fact, several studies have shown that GBW cova-ries with a host of extrinsic biotic and abiotic factors, such as habitat quality and urbanization (Grubb and Yosef 1994, Carrascal et al. 1998, Perez-Tris et al. 2000, Stratford and Stouff er 2001, Vangestel et al. 2010), social dominance (Carrascal et al. 1998) or nutritional condition (Grubb
1989, 1991, Grubb and Cimprich 1990). Th e proximate mechanistic links between GBW and ecological conditions may occur via a number of pathways, but nutritional condition and stress ultimately produced by habitat and social factors are the most likely candidates (Grubb 1991, White et al. 1991, Ekman et al. 1996, Carrascal et al. 1998, Grubb et al. 1998, Jenkins et al. 2001, Romero et al. 2005, DesRochers et al. 2009; but see Murphy and King 1991, Murphy 1992).
Capitalizing on the evidence that nutritional and other forms of stress can impact on GBW, some studies have tested the straight-forward prediction that individuals with larger GBW have higher phenotypic values at fi tness traits. In the Styan ’ s grasshopper warbler Locustella pleskei , large GBW on tail feathers, which are molted just after breeding, predicts early arrival and seasonal breeding success the fol-lowing year (both sexes) but also annual survival (males only) (Takaki et al. 2001). In Siberian jays Perisoreus infaustus , survival increases with GBW in both sexes (Gienapp and Meril ä 2010). Overall, however, it appears that the poten-tial of ptilochronology in studies of carry-over eff ects has remained largely unexploited.
In order for GBW on any given feather to represent a reliable proxy of individual condition during the entire molt period, GBW of feathers from one body region should covary with GBW of feathers from other body regions that are molted at diff erent times. In fact, consistency of GBW among feathers from an individual relative to other indi-viduals would imply that by measuring GBW on a single feather information can be gathered on condition over a longer period than that required to grow that particu-lar feather. However, this requisite has been seldom tested empirically in ptilochronological studies.
In this study of barn swallows Hirundo rustica , which normally undergo a complete molt of primary and rec-trix feathers during wintering in Africa (Ginn and Melville 1983, Jenni and Winkler 1994, Cramp 1998, Turner 2006), we thus fi rst analyzed the correlation between GBW measured on a rectrix and on a primaryremex. Moreover, we tested whether GBW on a rectrix feather grown in winter of 2010 – 2011 covaried with body condition (measured as residual body mass on body size) and intensity of infestation by ectoparasites, breeding date and seasonal reproductive output in the subsequent breeding season (i.e. in 2011). We predicted that old individuals ( � 2 yr old) had larger GBW than yearlings because experienced birds may be better foragers, and selection should have purged low-quality individuals from the old part of the population. Moreover, earlier spring arrival of old compared to yearling swallows to the breed-ing grounds suggests that molt and thus feather growth is faster in the former class of individuals. Protandrous spring migration and arrival (M ø ller 1994, Saino et al. 2010) should also select for faster molt and thus larger GBW in males, under the assumption that GBW aff ects molt speed and that sexes share the same ecological conditions during molt. Because relative parasite load of diff erent individuals is consistent during the year (Methods), we also expected GBW to be negatively pre-dicted by infestation by an hematophagous louse fl y and by feather chewing lice.

515
Within each sex by age class, the hypothesis of carry-over eff ects from the previous winter led us to expect thatGBW positively predicted reproductive output, and nega-tively predicted breeding date in the subsequent breeding season (i.e. spring 2011), since individuals in better condi-tion are expected to arrive and breed earlier (M ø ller 1994). However, a covariation between breeding performance and GBW could arise because of carry-over eff ects but also because of inter-individual variation in phenotypic/genetic quality, whereby high-quality individuals have both relatively fast feather growth and large breeding output. In order for a positive association between GBW and breeding performance in 2011 to refl ect carry-over eff ects, no or even negative covariation between GBW and breeding success in spring 2010 (i.e. before feathers were grown) should exist. Th is is the case because vari-ation in conditions experienced during wintering, and thus GBW, should aff ect breeding performance the next spring (year 2011) via carry-over eff ects, but should not predict breeding performance of the previous year (2010). If large reproductive eff ort entails severe physiological costs, carry-over eff ects may even be expected to operate between the breeding season and timing of molt, produc-ing a negative association between GBW and breeding out-put in 2010. On the other hand, if the association between GBW and breeding performance is dependent on vari-ation in individual quality, a positive covariation of GBW should also exist with breeding performance in 2010. Hence, for the sex by age groups where a signifi cant relationship between GBW and breeding variables in 2011 emerged, we also tested for a relationship between GBW and breeding variables measured in 2010.
Methods
Field methods
During spring – summer 2010 and 2011 we studied 9 barn swallow colonies ( � farms) in an intensively cultivated study area in the Po Valley, centered ca 50 km WNW of Milano (northern Italy). Th e farms were visited at least weekly to record breeding events and three or more cap-ture sessions in each farm were performed in April – July to individually mark all breeding adults with color rings and patches on the breast and belly feathers. Because breeding individuals normally spend the night inside the buildings, most capture sessions were started before sunrise by plac-ing mist-nets at all entrances of the rural buildings where the colonies were settled, ensuring to effi ciently capture the vast majority of the individuals. Th is was confi rmed by direct observation of the breeding pairs and by the very small proportion ( � 4%) of unmarked individuals that were found at the last capture session in July.
Barn swallows in a neighboring area as well as in southern Switzerland and other European areas (M ø ller 1994, Saino et al. 2011, Scandolara unpubl.) show extremely high breed-ing philopatry, meaning that individuals that have bred in a colony in one year do not move to breed in a diff erent colony in the next year(s). Conversely, off spring have large natal dispersal distances. High breeding philopatry and
capture effi ciency of breeding adults (see above) thus imply that unmarked individuals that were captured in 2011 could be confi dently assumed to be yearlings hatched in 2010 in another or, less frequently, in the same colony. Conversely, individuals that were already ringed upon capture in 2011 were 2 or more years old (i.e. hatched in 2009 or earlier). Adults were thus classifi ed by age (‘yearlings’ or ‘ older ’ indi-viduals) and by sex, as determined by inspection for presence of an incubation patch (indicating female), of a prominent cloacal protuberance (indicating male) upon repeated (up to fi ve) capture sessions, or by observation of singing and incu-bation behavior. Because captures in the study farms started in 2010, no information on age of individuals in 2010 was available.
Upon fi rst capture in 2011 we measured a number of traits including keel length using a digital caliper (expressed in mm � 100; approximation of 0.01 mm), that was used as a proxy for skeletal body size to calculate a body con-dition index (Statistical analyses). At fi rst capture and all recaptures (up to fi ve per individual) we also measured body mass (in g � 10; precision of 0.1 g) of all individu-als. Th e intensity of infestation by louse fl ies Ornithomya biloba was measured by counting the number of fl ies by carefully inspecting all plumage regions in a standard way. Th e abundance of the chewing lice Machaerilaemus malleus was estimated by counting the number of character-istic holes they make on rectrix and remex feathers, which is proportional to the actual number of parasites harbored by an individual (M ø ller 1994, Pap et al. 2005).
Growth bar width of a rectrix feather (Ptilochronological measurements) was modeled in relation to ectoparasite load assuming that parasites can aff ect feather development, as suggested by previous studies (Introduction). Because para-site load was measured during the breeding season, this analysis rests on the assumption that the intensity of an individual ’ s parasite infestation relative to other individu-als is consistent in winter and spring. Albeit this assump-tion cannot be tested directly, its validity is supported by a signifi cant positive, though weak correlation of parasite loads of the same individuals measured in consecutive breeding seasons (M ø ller 1994, Saino et al. unpubl.), which suggests consistency also at intermediate, wintering times.
Th e adults attending individual clutches/broods were identifi ed by direct observation of behavior at the nest (see also below), and timing of breeding events and breed-ing performance (laying date, clutch size and number of nestlings) were determined by visiting the nests every week or more frequently. When date of laying of the fi rst egg could not be observed directly, it was inferred from clutch size, assuming that consecutive eggs are laid at one day intervals (M ø ller 1994, Saino et al. 2002, Turner 2006). Th roughout the paper we refer to the number of fl edglings as ‘ breeding success ’ and to the ratio between the number of fl edglings and the number of eggs as ‘ fl edging success ’ . Dates are expressed as Julian days (1 January � 1). Because nests were not always accessible due to e.g. presence of cattle in the stable or could be diffi cult to observe due to their position, and because of time limitation in assigning indi-viduals to nests, the number of individuals for which infor-mation on breeding performance was available was smaller than that on which feather growth bars were measured

516
Hence, body mass measurement was centered on average on the start of laying. Th e analyses on body condition were restricted to individuals for which information on breed-ing date was available because in the analyses we aimed at controlling for BCI variation according to breeding stage.
All dependent variables were modeled assuming a normal error distribution, except for number of broods ( � number of clutches initiated) per season (Poisson distri-bution) and fl edging success ( � proportion of eggs that gen-erated a fl edgling; binomial distribution). Data for breeding performance of swallows from the same colony may be considered as non-independent and may therefore have to be modeled including farm as random factor. Likelihood ratio tests comparing Gaussian linear mixed models including farm as random factor with the corresponding linear models excluding the random eff ect, however, showed that the mixed model did not provide a better fi t compared to the linear model ( χ 2 � 3.60, DF � 1, always p � 0.057). Moreover, in all analyses the inclusion or, respectively, exclusion of the random term ‘ colony ’ led to qualitatively very similar results (i.e. the eff ects that were signifi cant in the generalized linear model were also signifi cant in the corresponding mixed model, and vice versa). Hence, for simplicity in the Results we present the outcome of (generalized) linear models, i.e. the models excluding the random eff ect of farm.
Ideally, breeding performance variables of pairs could be analyzed by entering GBWR4 and age of both members of the pair in the same model. However, this was imprac-tical for diverse reasons. First, no information on GBWR4 was available for the (fi rst clutch) mate of 38 individuals (see also Ptilochronological measurements). Second, 8% of the adults changed mate between consecutive clutches, so that pairs ’ breeding performance variables (e.g. a pair ’ s sea-sonal number of fl edglings) could not always be unequivo-cally defi ned. Th ird, in barn swallows there is strong positive assortative mating for age (M ø ller 1994, Saino et al. unpubl.). Hence, models including age of both partners would incorporate collinear independent variables, poten-tially biasing error estimates. We therefore analyzed breed-ing performance and BCI in relation to GBWR4 and age of each sex separately. In the models, we initially included the interaction term between age and GBWR4 , and subsequently excluded it when its eff ect was non-signifi cant. Post-hoc comparisons were done by the least signifi cant diff erence method (LSD). We note that laying date was not included as a covariate in the models of breeding performance because laying date is highly associated with age (Results).
Statistics are reported with their standard error where appropriate.
Results
Sex and age effects on GBW
We analyzed the pattern of covariation between GBWR4 and GBW P7 using a subsample of 42 males and 41 females where both characters were measured. In a linear model with sex and age as factors, GBW P7 was strongly positively related to GBWR4 (F1,79 � 29.39, p � 0.001, coeffi cient � 0.710
(see Results for a detailed account of the size of the sample of individuals involved in the analyses).
Ptilochronological measurements
On all individuals included in the study, in 2011 we mea-sured GBW on the 4th rectrix feather (counting outwards). Th is feather was chosen because at a preliminary screen-ing of several individuals it was the feather where bands could be identifi ed most clearly than on the other tail feath-ers or on primary wing feathers. GBW has been measured on rectrix feathers also in most studies using ptilochrono-logical traits. However, on approximately 15% of the birds, too few or no distinct growth bars could be distin-guished. Th ese individuals were excluded from the study. At fi rst capture, we fi rst identifi ed the proximal and dis-tal limits of a feather segment including 8 – 11 bars, using the proximal limit of dark bands as reference. While it can be speculated that the greater the number of bars included in the measures, the more complete the infor-mation on an individual condition is expected to be, we decided to limit to 8 – 11 the number of bars included in the segment because at a preliminary screening these were the numbers of bars that could be clearly seen in most indi-viduals. Th e segment started from the fi rst/second clearly visible band at the distal end of the feather and extended to cover the next 8 – 11 bars towards the proximal end of the feather. Th e limits of the segment where marked on the rachis with a fi ne white fi bre tip pen. We then measured the length of the segment with a digital caliper (precision of 0.01 mm, measures expressed in mm). GBW on the 4th rectrix feather (GBWR4 ) was expressed as the length of the segment divided by the number of growth bars it included, as commonly done in ptilochronological studies (Grubb 1989, Kern and Cowie 2002). Hence, large GBWR4 indicates rapid feather growth. At a following recapture, in a subsample of 41 individuals we re-measured the same bands to estimate repeatability, which proved to be high (intraclass correlation coeffi cient � 0.86). Moreover, in a subsample of 42 males and 41 females we did the same measurements on 8 – 11 growth bars on the 7th (counting outwards) primary wing feather and computed GBW P7 . Th ese data were used to test for a correlation between GBW recorded on diff erent plumage regions (Results).
Statistical analyses
All analyses were run with SPSS 13.0 or SAS 9.2. We mainly relied on linear and generalized linear models to test the eff ect of sex and age (factors) or parasite counts (covariate) on GBWR4 and of GBWR4 and age on breeding date, breed-ing performance variables, and on body condition index (BCI). Th e latter was estimated as residual from a reduced major axis regression of body mass on keel length (see Green 2001 for a discussion of the merits of this approach relative to other methods). In all cases, body mass recorded at fi rst capture of individuals in 2011 was used. Th e mean stage in the breeding cycle (defi ned as the diff erence between date of measurement and date of laying of the fi rst egg in the fi rst clutch) when body mass data used in the analyses were collected was day 0.63 (12.77 SD, n � 238).
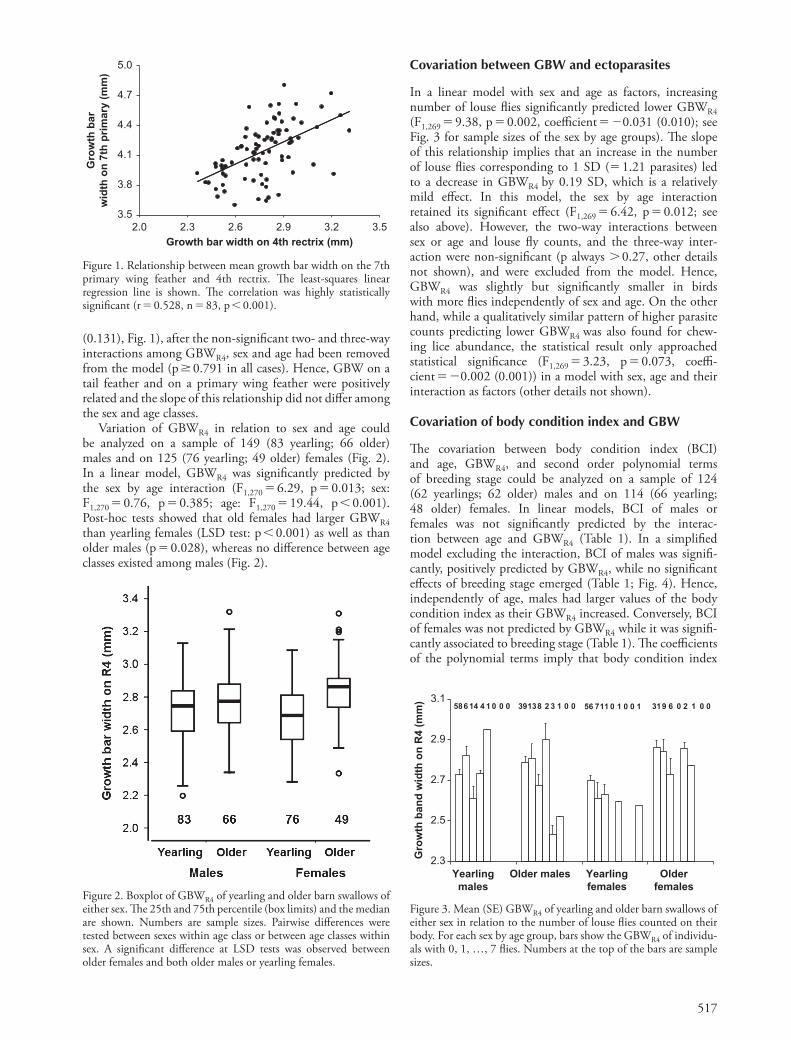
517
Covariation between GBW and ectoparasites
In a linear model with sex and age as factors, increasing number of louse fl ies signifi cantly predicted lower GBWR4 (F1,269 � 9.38, p � 0.002, coeffi cient � �0.031 (0.010); see Fig. 3 for sample sizes of the sex by age groups). Th e slope of this relationship implies that an increase in the number of louse fl ies corresponding to 1 SD ( � 1.21 parasites) led to a decrease in GBWR4 by 0.19 SD, which is a relatively mild eff ect. In this model, the sex by age interaction retained its signifi cant eff ect (F1,269 � 6.42, p � 0.012; see also above). However, the two-way interactions between sex or age and louse fl y counts, and the three-way inter-action were non-signifi cant (p always � 0.27, other details not shown), and were excluded from the model. Hence, GBWR4 was slightly but signifi cantly smaller in birds with more fl ies independently of sex and age. On the other hand, while a qualitatively similar pattern of higher parasite counts predicting lower GBWR4 was also found for chew-ing lice abundance, the statistical result only approached statistical signifi cance (F1,269 � 3.23, p � 0.073, coeffi -cient � �0.002 (0.001)) in a model with sex, age and their interaction as factors (other details not shown).
Covariation of body condition index and GBW
Th e covariation between body condition index (BCI) and age, GBWR4 , and second order polynomial terms of breeding stage could be analyzed on a sample of 124 (62 yearlings; 62 older) males and on 114 (66 yearling; 48 older) females. In linear models, BCI of males or females was not signifi cantly predicted by the interac-tion between age and GBWR4 (Table 1). In a simplifi ed model excluding the interaction, BCI of males was signifi -cantly, positively predicted by GBWR4 , while no signifi cant eff ects of breeding stage emerged (Table 1; Fig. 4). Hence, independently of age, males had larger values of the body condition index as their GBWR4 increased. Conversely, BCI of females was not predicted by GBWR4 while it was signifi -cantly associated to breeding stage (Table 1). Th e coeffi cients of the polynomial terms imply that body condition index
(0.131), Fig. 1), after the non-signifi cant two- and three-way interactions among GBWR4 , sex and age had been removed from the model (p � 0.791 in all cases). Hence, GBW on a tail feather and on a primary wing feather were positively related and the slope of this relationship did not diff er among the sex and age classes.
Variation of GBWR4 in relation to sex and age could be analyzed on a sample of 149 (83 yearling; 66 older) males and on 125 (76 yearling; 49 older) females (Fig. 2). In a linear model, GBWR4 was signifi cantly predicted by the sex by age interaction (F1,270 � 6.29, p � 0.013; sex: F1,270 � 0.76, p � 0.385; age: F1,270 � 19.44, p � 0.001). Post-hoc tests showed that old females had larger GBWR4 than yearling females (LSD test: p � 0.001) as well as than older males (p � 0.028), whereas no diff erence between age classes existed among males (Fig. 2).
3.5
3.8
4.1
4.4
4.7
5.0
2.0 2.3 2.6 2.9 3.2 3.5
Gro
wth
b
ar
wid
th
o
n 7th
p
rim
ary (m
m)
Growth bar width on 4th rectrix (mm)
Figure 1. Relationship between mean growth bar width on the 7th primary wing feather and 4th rectrix. Th e least-squares linear regression line is shown. Th e correlation was highly statistically signifi cant (r � 0.528, n � 83, p � 0.001).
Figure 2. Boxplot of GBWR4 of yearling and older barn swallows of either sex. Th e 25th and 75th percentile (box limits) and the median are shown. Numbers are sample sizes. Pairwise diff erences were tested between sexes within age class or between age classes within sex. A signifi cant diff erence at LSD tests was observed between older females and both older males or yearling females.
2.3
2.5
2.7
2.9
3.1
Yearling
males
Older males Yearling
females
Older
females
Gro
wth
b
an
d w
id
th
o
n R
4 (m
m)
58 39 56 316 14 4 1 0 0 0 138 2 3 1 0 0 711 0 1 0 0 1 9 6 0 2 1 0 0
Figure 3. Mean (SE) GBWR4 of yearling and older barn swallows of either sex in relation to the number of louse fl ies counted on their body. For each sex by age group, bars show the GBWR4 of individu-als with 0, 1, … , 7 fl ies. Numbers at the top of the bars are sample sizes.
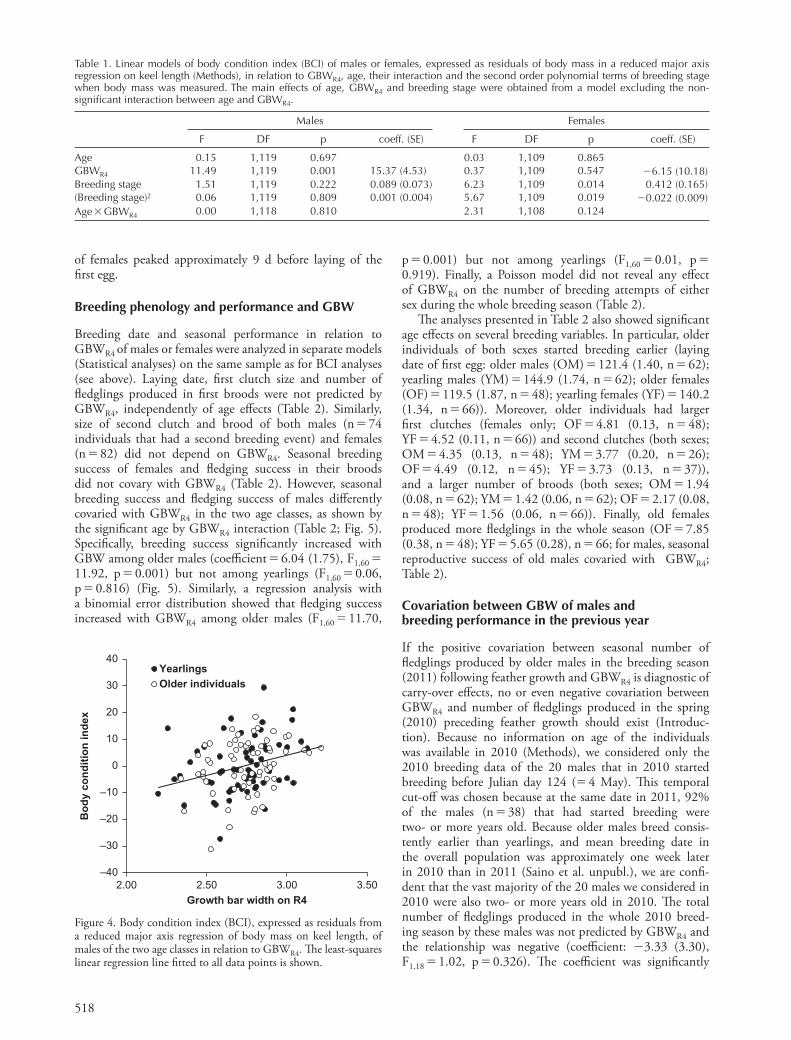
518
p � 0.001) but not among yearlings (F1,60 � 0.01, p � 0.919). Finally, a Poisson model did not reveal any eff ect of GBWR4 on the number of breeding attempts of either sex during the whole breeding season (Table 2).
Th e analyses presented in Table 2 also showed signifi cant age eff ects on several breeding variables. In particular, older individuals of both sexes started breeding earlier (laying date of fi rst egg: older males (OM) � 121.4 (1.40, n � 62); yearling males (YM) � 144.9 (1.74, n � 62); older females (OF) � 119.5 (1.87, n � 48); yearling females (YF) � 140.2 (1.34, n � 66)). Moreover, older individuals had larger fi rst clutches (females only; OF � 4.81 (0.13, n � 48); YF � 4.52 (0.11, n � 66)) and second clutches (both sexes; OM � 4.35 (0.13, n � 48); YM � 3.77 (0.20, n � 26); OF � 4.49 (0.12, n � 45); YF � 3.73 (0.13, n � 37)), and a larger number of broods (both sexes; OM � 1.94 (0.08, n � 62); YM � 1.42 (0.06, n � 62); OF � 2.17 (0.08, n � 48); YF � 1.56 (0.06, n � 66)). Finally, old females produced more fl edglings in the whole season (OF � 7.85 (0.38, n � 48); YF � 5.65 (0.28), n � 66; for males, seasonal reproductive success of old males covaried with GBWR4; Table 2).
Covariation between GBW of males and breeding performance in the previous year
If the positive covariation between seasonal number of fl edglings produced by older males in the breeding season (2011) following feather growth and GBWR4 is diagnostic of carry-over eff ects, no or even negative covariation between GBWR4 and number of fl edglings produced in the spring (2010) preceding feather growth should exist (Introduc-tion). Because no information on age of the individuals was available in 2010 (Methods), we considered only the 2010 breeding data of the 20 males that in 2010 started breeding before Julian day 124 ( � 4 May). Th is temporal cut-off was chosen because at the same date in 2011, 92% of the males (n � 38) that had started breeding were two- or more years old. Because older males breed consis-tently earlier than yearlings, and mean breeding date in the overall population was approximately one week later in 2010 than in 2011 (Saino et al. unpubl.), we are confi -dent that the vast majority of the 20 males we considered in 2010 were also two- or more years old in 2010. Th e total number of fl edglings produced in the whole 2010 breed-ing season by these males was not predicted by GBWR4 and the relationship was negative (coeffi cient: �3.33 (3.30), F1,18 � 1.02, p � 0.326). Th e coeffi cient was signifi cantly
of females peaked approximately 9 d before laying of the fi rst egg.
Breeding phenology and performance and GBW
Breeding date and seasonal performance in relation to GBWR4 of males or females were analyzed in separate models (Statistical analyses) on the same sample as for BCI analyses (see above). Laying date, fi rst clutch size and number of fl edglings produced in fi rst broods were not predicted by GBWR4 , independently of age eff ects (Table 2). Similarly, size of second clutch and brood of both males (n � 74 individuals that had a second breeding event) and females (n � 82) did not depend on GBWR4 . Seasonal breeding success of females and fl edging success in their broods did not covary with GBWR4 (Table 2). However, seasonal breeding success and fl edging success of males diff erently covaried with GBWR4 in the two age classes, as shown by the signifi cant age by GBWR4 interaction (Table 2; Fig. 5). Specifi cally, breeding success signifi cantly increased with GBW among older males (coeffi cient � 6.04 (1.75), F1,60 � 11.92, p � 0.001) but not among yearlings (F1,60 � 0.06, p � 0.816) (Fig. 5). Similarly, a regression analysis with a binomial error distribution showed that fl edging success increased with GBWR4 among older males (F1,60 � 11.70,
Table 1. Linear models of body condition index (BCI) of males or females, expressed as residuals of body mass in a reduced major axis regression on keel length (Methods), in relation to GBWR4 , age, their interaction and the second order polynomial terms of breeding stage when body mass was measured. The main effects of age, GBWR4 and breeding stage were obtained from a model excluding the non-signifi cant interaction between age and GBWR4 .
Males Females
F DF p coeff. (SE) F DF p coeff. (SE)
Age 0.15 1,119 0.697 0.03 1,109 0.865GBWR4 11.49 1,119 0.001 15.37 (4.53) 0.37 1,109 0.547 �6.15 (10.18)Breeding stage 1.51 1,119 0.222 0.089 (0.073) 6.23 1,109 0.014 0.412 (0.165)(Breeding stage)2 0.06 1,119 0.809 0.001 (0.004) 5.67 1,109 0.019 �0.022 (0.009)Age � GBWR4 0.00 1,118 0.810 2.31 1,108 0.124
–40
–30
–20
–10
0
10
20
30
40
2.00 2.50 3.00 3.50
Bo
dy co
nd
itio
n in
dex
Growth bar width on R4
Yearlings
Older individuals
Figure 4. Body condition index (BCI), expressed as residuals from a reduced major axis regression of body mass on keel length, of males of the two age classes in relation to GBWR4 . Th e least-squares linear regression line fi tted to all data points is shown.
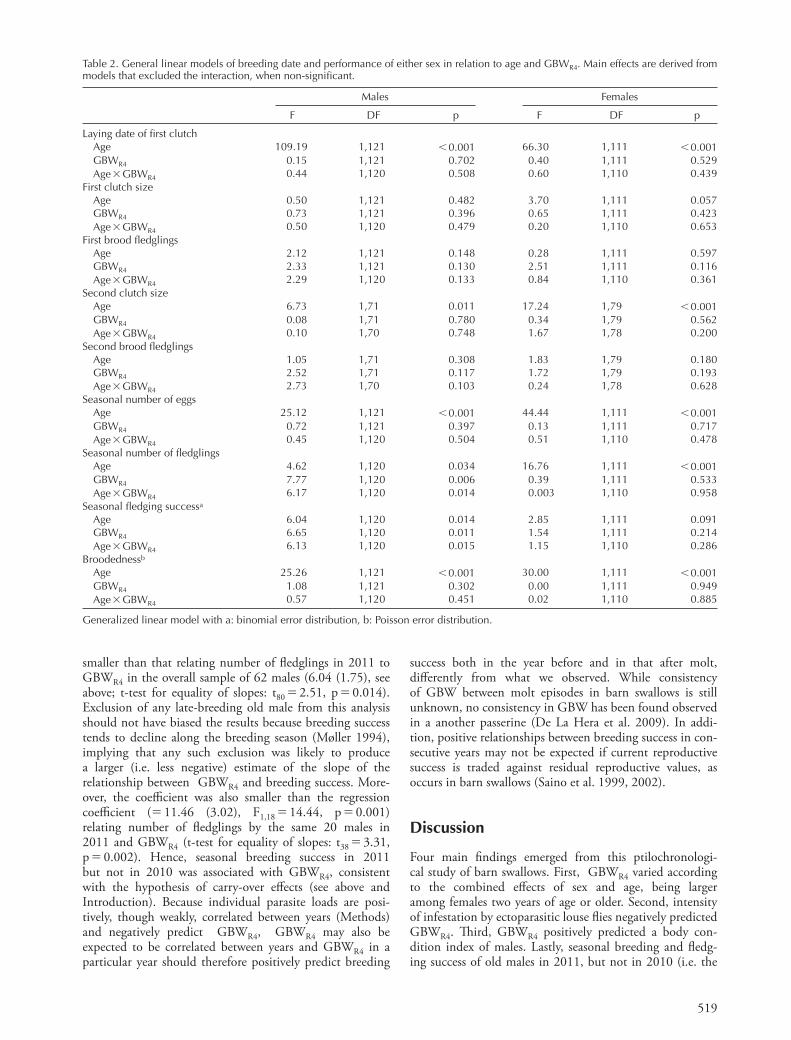
519
Table 2. General linear models of breeding date and performance of either sex in relation to age and GBWR4 . Main effects are derived from models that excluded the interaction, when non-signifi cant.
Males Females
F DF p F DF p
Laying date of fi rst clutchAge 109.19 1,121 � 0.001 66.30 1,111 � 0.001GBWR4 0.15 1,121 0.702 0.40 1,111 0.529Age � GBWR4 0.44 1,120 0.508 0.60 1,110 0.439
First clutch sizeAge 0.50 1,121 0.482 3.70 1,111 0.057GBWR4 0.73 1,121 0.396 0.65 1,111 0.423Age � GBWR4 0.50 1,120 0.479 0.20 1,110 0.653
First brood fl edglingsAge 2.12 1,121 0.148 0.28 1,111 0.597GBWR4 2.33 1,121 0.130 2.51 1,111 0.116Age � GBWR4 2.29 1,120 0.133 0.84 1,110 0.361
Second clutch sizeAge 6.73 1,71 0.011 17.24 1,79 � 0.001GBWR4 0.08 1,71 0.780 0.34 1,79 0.562Age � GBWR4 0.10 1,70 0.748 1.67 1,78 0.200
Second brood fl edglingsAge 1.05 1,71 0.308 1.83 1,79 0.180GBWR4 2.52 1,71 0.117 1.72 1,79 0.193Age � GBWR4 2.73 1,70 0.103 0.24 1,78 0.628
Seasonal number of eggsAge 25.12 1,121 � 0.001 44.44 1,111 � 0.001GBWR4 0.72 1,121 0.397 0.13 1,111 0.717Age � GBWR4 0.45 1,120 0.504 0.51 1,110 0.478
Seasonal number of fl edglingsAge 4.62 1,120 0.034 16.76 1,111 � 0.001GBWR4 7.77 1,120 0.006 0.39 1,111 0.533Age � GBWR4 6.17 1,120 0.014 0.003 1,110 0.958
Seasonal fl edging successa
Age 6.04 1,120 0.014 2.85 1,111 0.091GBWR4 6.65 1,120 0.011 1.54 1,111 0.214Age � GBWR4 6.13 1,120 0.015 1.15 1,110 0.286
Broodednessb
Age 25.26 1,121 � 0.001 30.00 1,111 � 0.001GBWR4 1.08 1,121 0.302 0.00 1,111 0.949Age � GBWR4 0.57 1,120 0.451 0.02 1,110 0.885
Generalized linear model with a: binomial error distribution, b: Poisson error distribution.
smaller than that relating number of fl edglings in 2011 to GBWR4 in the overall sample of 62 males (6.04 (1.75), see above; t-test for equality of slopes: t80 � 2.51, p � 0.014). Exclusion of any late-breeding old male from this analysis should not have biased the results because breeding successtends to decline along the breeding season (M ø ller 1994), implying that any such exclusion was likely to produce a larger (i.e. less negative) estimate of the slope of the relationship between GBWR4 and breeding success. More-over, the coeffi cient was also smaller than the regression coeffi cient ( � 11.46 (3.02), F1,18 � 14.44, p � 0.001) relating number of fl edglings by the same 20 males in 2011 and GBWR4 (t-test for equality of slopes: t38 � 3.31, p � 0.002). Hence, seasonal breeding success in 2011 but not in 2010 was associated with GBWR4 , consistent with the hypothesis of carry-over eff ects (see above and Introduction). Because individual parasite loads are posi-tively, though weakly, correlated between years (Methods) and negatively predict GBWR4 , GBWR4 may also be expected to be correlated between years and GBWR4 in a particular year should therefore positively predict breeding
success both in the year before and in that after molt, diff erently from what we observed. While consistency of GBW between molt episodes in barn swallows is still unknown, no consistency in GBW has been found observed in a another passerine (De La Hera et al. 2009). In addi-tion, positive relationships between breeding success in con-secutive years may not be expected if current reproductive success is traded against residual reproductive values, as occurs in barn swallows (Saino et al. 1999, 2002).
Discussion
Four main fi ndings emerged from this ptilochronologi-cal study of barn swallows. First, GBWR4 varied according to the combined eff ects of sex and age, being larger among females two years of age or older. Second, intensity of infestation by ectoparasitic louse fl ies negatively predicted GBWR4 . Th ird, GBWR4 positively predicted a body con-dition index of males. Lastly, seasonal breeding and fl edg-ing success of old males in 2011, but not in 2010 (i.e. the

520
traits. In order to accomplish early migration and arrival, which accrues sexual selection advantages in this protan-drous species (M ø ller 1994, Spottiswoode and Saino 2010), males may be forced to advance molt, possibly molt-ing under worse ecological conditions. Admittedly, this interpretation is a posteriori, as there is no clue to how seasonal changes in ecological conditions during wintering may aff ect feather growth rate. In theory, an alternative interpretation is that diff erent feather growth rate results from spatial, rather than temporal segregation at the time of molt. In some migratory birds, the sexes spatially segre-gate during wintering, and males almost invariably winter closer to the breeding areas (Newton 2008, Spottiswoode and Saino 2010). Barn swallows breeding in northern Italy winter in Sahel and sub-Sahel regions, mostly north of the Equator (Saino et al. 2004a, b). Wintering in drier, less hospitable areas north in Sahel may enforce slower feather growth on males (Norris et al. 2004). Another alternative interpretation is that females may molt later and thus be forced to grow feather faster than males.
Juvenile barn swallows molt later than adults (Ginn and Melville 1983, Rubolini et al. 2002). Lack of sex dif-ference in GBW among juveniles may refl ect looser pres-sure for early start of molt among juveniles, which arrive to the breeding grounds on average weeks after older birds (M ø ller 1994, Saino et al. unpubl.). Finally, larger GBW of older compared to juvenile females may depend on age-related diff erences in body condition owing to e.g. diff er-ences in foraging effi ciency, as suggested by the positive covariation of molt scores with body condition (M ø ller et al. 1995), which may imply that birds in good body con-dition molt faster. We have no explanation for the fact that this age-related variation was found only among females.
year before the focal feathers were grown) were positively predicted by GBWR4 .
An open issue in ptilochronological studies is the extent of time over which GBW measured on a single feather provides information. While the rate at which any feather is produced may also be sensitive to earlier, besides con-tingent, physiological state, it seems plausible that GBW refl ects state over a period not much longer than the feather ’ s growth time span. We showed that GBW measured on a rectrix predicts GBW on a primary remex. Because tail and wing feathers are molted at partly diff erent times (Ginn and Melville 1983), an important methodological impli-cation is that GBW on one plumage region may also pro-vide information on physiological state over molt periods diff erent from that when the focal feather was grown, likely because an individual ’ s condition relative to other individu-als is consistent during molt.
Sex- and age-related variation in GBW
Individuals of either sex are expected to diff er in feath-ers growth rate. Proximately, this can be predicted because males and females may diff er in migration and molt phenology (Ginn and Melville 1983, M ø ller 1994, Rubolini et al. 2002, Newton 2008, Saino et al. 2010), and may also diff er in susceptibility to ecological condi-tions during wintering (Weatherhead 2005). In the pres-ent study, older females had larger GBW than males of the same age class, contrary to our expectation. Male barn swallows are believed to molt their body, tail and wing feathers earlier than females (M ø ller et al. 1995, 2011, Rubolini et al. 2002), although these results may partly be infl uenced by sexing procedures based on morphological
0
2
4
6
8
10
12
2.0 2.3 2.6 2.9 3.2 3.5Seaso
nal n
um
ber o
f fled
glin
gs
Yearling males
0
2
4
6
8
10
12
2.0 2.3 2.6 2.9 3.2 3.5
Older males
0
2
4
6
8
10
12
2.0 2.3 2.6 2.9 3.2 3.5Seaso
nal n
um
ber o
f fled
glin
gs
Growth bar width on R4
Yearling females
0
2
4
6
8
10
12
2.0 2.3 2.6 2.9 3.2 3.5Growth bar width on R4
Older females
Figure 5. Total number of off spring produced during the breeding season by males or females in the two age classes in relation to GBWR4 . Th e lines are fi tted by least-squares linear regression. Th e continuous line indicates that the relationship was signifi cant. Dashed lines indicate non-signifi cant relationships.

521
good body conditions and into better parental performance. Th is interpretation is corroborated by the observation that GBWR4 predicted breeding output recorded in the breeding season following but not in that preceding the winter when feathers were grown. Th is observation is consistent with the hypothesis of carry-over eff ects whereas it contradicts the alternative hypothesis that the positive association between GBWR4 and breeding performance was the spu-rious by-product of phenotypic/genetic variation among individuals because this should have translated into a con-sistently positive association between GBWR4 and breed-ing performance in both years (Introduction). However, better body conditions did not directly determine breeding success as there was no correlation between these variables (r � �0.081, n � 62, p � 0.529). Further, there was no hint that fast feather growth translated into early arrival, because GBWR4 did not predict breeding date, which is strongly positively correlated with arrival date. Th is may occur because feather growth rate is unrelated to molt speed or because migration schedule is not constrained by molt.
Our study suggests that the patterns of covariation between feather growth rate, body condition and breeding performance are sex- and age-specifi c. Although the analy-ses did not always provide statistically signifi cant evidence for such diff erential associations, it appears that these were steeper among males, particularly older ones. Older males have perhaps the tightest annual routine: they arrive from spring migration earlier than females and yearling males, have more breeding attempts and may molt fi rst. Tight annual routines may make older males more susceptible to ecological conditions and parasitism in winter and thus the expression of carry-over eff ects more likely, particularly in a period when climate change may exacerbate ecological constraints and favor earlier arrival (M ø ller et al. 2010).
Still unknown is the identity of the specifi c mechanisms mediating the eff ect of environmental conditions on feather growth. In birds, response to stress, including nutritional defi ciency, involves corticosterone-mediated activation of the hypothalamo-pituitary-adrenal axis (Sapolsky et al. 2000). Elevation of corticosterone levels can interfere with molt, and the decrease in circulating corticosterone levels during pre-basic molt in passerines has been interpreted as an adaptation to prevent proteolytic eff ects of corticos-terone on feather growth (Romero 2002, Romero et al. 2005). In fact, corticosterone elevation has major eff ects on energetic metabolism by inhibiting protein biosynthesis and promoting gluconeogenesis via protein catabolism (Eigler et al. 1979, Sapolsky et al. 2000). Increased corti-costerone levels may therefore be the proximate cause of reduced feather growth rate under adverse conditions (Romero et al. 2005, DesRochers et al. 2009). Because parasitism can also aff ect the energetic balance directly or elicit a stress response, corticosterone may also aff ord the mechanistic link between large parasite infestation and low feather growth rate (Apanius 1998). However, causation between parasitism and feather growth rate could also be reversed, because stress from extrinsic factors may elevate corticosterone levels, which in turn have detrimental eff ects on immune system functioning.
Variation in GBW may thus be largely environmental in origin, and arise as a consequence of diff erences among
However, no study has investigated the relationship between GBW and molt duration, and faster growth rate may or may not imply shorter time to complete molt depending on the extent of overlap in molt of diff erent feathers.
Sex and age related variation in feather growth rate is not a novel fi nding. In Siberian jays, diff erential sex-related variation according to age has also been observed (Gienapp and Meril ä 2010). While older Siberian jays were found to have larger GBW than yearlings, consistently with the pattern observed here, relative GBW of male and female older barn swallows was reversed compared to Siberian jays. Th at variation in sex diff erences may exist between these species is not surprising, given their extensive diff erences in life-history, although we have no explanation for the specifi c pattern of diff erences we observed.
Covariation between parasitism, body condition and GBW
Large GBWR4 was associated with low levels of parasitism by louse fl ies and non-signifi cantly negatively correlated to infestation by chewing lice in both sexes. Th ough ectopara-sitism was measured in spring, we interpret these fi ndings as evidence that louse fl ies reduced GBWR4, because ecto-parasite loads are consistent within individuals in consecu-tive years and ectoparasite counts in any given spring may therefore be a reliable proxy for ectoparasitism during the previous year. Hence, low numbers of louse fl ies during winter or in the previous spring may have translated into rapid feather growth, as suggested by studies showing a negative eff ect of parasitism and immune response on molt speed (Ilmonen et al. 2000, Sanz et al. 2004, More-no-Rueda 2010). In addition, large GBWR4 predicted good body condition of males but this pattern did not emerge among females. Females may be less susceptible to con-ditions during molt. Alternatively, any clear relationship between GBWR4 and body condition among females could have been blurred by variation in body mass during thebreeding cycle, though we accounted for the eff ect of breed-ing stage in the analyses.
Breeding performance and GBWR4
Seasonal breeding output of older males positively covar-ied with GBWR4 . Th e large total number of off spring pro-duced by males with large GBWR4 was achieved by means of larger fl edging success, rather than by an increase in the number of broods or fecundity of their mates. Nor did males with large GBWR4 achieve large fl edging success by breed-ing early, when ecological conditions are better (Ambrosini et al. 2006, Gr ü ebler and Naef-Daenzer 2008, Verhulst and Nilsson 2008).
Ecological conditions in the winter quarters predict variation in arrival phenology and inter-annual variation in mean breeding success: after years with high primary production in the winter quarters, older barn swallows advance arrival and mean breeding success at the population level is larger (Saino et al. 2004a, b). Th us, one interpre-tation of the results of the present study is that favorable conditions that some males experienced during wintering, and were refl ected by larger GBWR4, carried-over both into

522
Bortolotti, G. R., Dawson, R. D. and Murza, G. L. 2002. Stress during feather development predicts fi tness potential. – J. Anim. Ecol. 71: 333 – 342.
Brodin, A. 1993. Radio-ptilochronology: tracing radioactively labelled food in feathers. – Ornis Scand. 24: 167 – 173.
Carrascal, L. M., Senar, J. C., Mozetich, I., Uribe, F. and Domenech, J. 1998. Interactions among environmental stress, body condi-tion, nutritional status, and dominance in great tits. – Auk 115: 727 – 738.
Clarkson, C. E. 2011. Applicability of ptilochronology as a conservation tool in waterbirds studies. – Ecol. Indic. 11: 1707 – 1709.
Cramp, S. 1998. Th e complete birds of the Western Palearctic on CD-ROM. – Oxford Univ. Press.
Dawson, A., Hinsley, S. A., Ferns, P. N., Bonser, R. H. C. and Eccleston, L. 2000. Rate of moult aff ects feather quality: a mechanism linking current reproductive eff ort to future survival. – Proc. R. Soc. B 267: 2093 – 2098.
De La Hera, I., Perez-Tris, J. and Telleria, J. L. 2009. Repeatable length and mass but not growth rate of individual feathers between moults in a passerine bird. – Acta Ornithol. 44: 95 – 99.
DesRochers, D. W., Reed, J. M., Awerman, J., Kluge, J. A., Wilkinson, J., van Griethuijsen, L. I., Aman, J. and Romero, L. M. 2009. Exogenous and endogenous corticosterone alter feather quality. – Comp. Biochem. Physiol. A 152: 46 – 52.
Eigler, N., Sacca, L. and Sherwin, R. S. 1979. Synergistic interactions of physiologic increments of glucagons epine-phrine and cortisol in the dog: a model for stress-induced hyperglycemia. – J. Clin. Invest. 63: 114 – 123.
Ekman, J. A., Brodin, A., Bylin, A. and Sklepkovych, B. 1996. Selfi sh long-term benefi ts of hoarding in the Siberian jay. – Behav. Ecol. 7: 140 – 144.
Fitzpatrick, S. 1998. Birds ’ tails as signaling devices: markings, shape, length, and feather quality. – Am. Nat. 151: 157 – 173.
Gienapp, P. and Meril ä , J. 2010. Genetic and environmental eff ects on a condition-dependent trait: feather growth in Siberian jays. – J. Evol. Biol. 23: 715 – 723.
Gienapp, P. and Meril ä , J. 2011. Sex-specifi c fi tness consequences of dispersal in Siberian jays. – Behav. Ecol. Sociobiol. 65: 131 – 140.
Gill, J. A., Norris, K., Potts, P. M., Gunnarsson, T. G., Atkinson, P. W. and Sutherland, W. J. 2001. Th e buff er eff ect and large-scale population regulation in migratory birds. – Nature 412: 436 – 438.
Ginn, H. B. and Melville, D. S. 1983. Moult in birds. – British Trust for Ornithology.
Green, A. J. 2001. Mass/length residuals: measures of body condition or generators of spurious results? – Ecology 82: 1473 – 1483.
Grubb, T. C. Jr 1989. Ptilochronology: feather growth bars as indicators of nutritional status. – Auk 106: 314 – 320.
Grubb, T. C. Jr 1991. A defi cient diet narrows growth bars on induced feathers. – Auk 108: 725 – 727.
Grubb, T. C. Jr 1995. Ptilochronology. A review and prospectus. – Curr. Ornithol. 12: 89 – 114.
Grubb, T. C. Jr 2006. Ptilochronology: feather time and the biology of birds. – Oxford Univ. Press.
Grubb, T. C. Jr and Cimprich, D. A. 1990. Supplementary food improves the nutritional condition of wintering woodland birds: evidence from ptilochronology. – Ornis Scand. 21: 277 – 281.
Grubb, T. C. Jr and Yosef, R. 1994. Habitat-specifi c nutritional condition in loggerhead shrikes (Lanius ludovicianus ): evi-dence from ptilochronology. – Auk 111: 756 – 759.
Grubb, T. C. Jr, Woolfenden, G. E. and Fitzpatrick, J. W. 1998. Factors aff ecting nutritional condition of fl edgling Florid scrub-jays: a ptilochronology approach. – Condor 100: 753 – 756.
individuals in wintering strategies. Th e relative contribution of environmental and additive genetic sources of variation in this trait has seen little investigation. Signifi cant, but low narrow-sense heritability estimates have been obtained in animal models of GBW in Siberian jays (Gienapp and Meril ä 2011). While low estimates do not necessarily imply that a trait harbors small additive genetic variation, these estimates may be biased upwards by environmental covariation among related individuals and maternal eff ects, and indicate that environmental (and measurement error) components are large (Gienapp and Meril ä 2011). Hence, variation in GBW in barn swallows may also have a large environmental component. Future parent – off spring analyses may provide heritability estimates, though they will hardly control for environmental covariance between relatives.
In conclusion, this study showed that the growth rate of feathers molted in winter predicts breeding performance and body condition in the next but not in the previous spring. Th ese results are consistent with the hypothesis of carry-over eff ects whereby individuals in good physical con-ditions during winter perform better at the breeding sites. Th ese carry-over eff ects were stronger among older males, possibly because their tight annual routines make them more susceptible to variation in ecological conditions and thus in the expression of condition-dependent feather traits. Importantly, we also provided evidence that GBW is posi-tively correlated between plumage regions, implying that this trait may integrate information on body condition over a relatively large period during molt.
As a fi nal remark, we feel that ptilochronological stud-ies have the potential to provide a larger contribution to bird behavioral and ecological studies than has occurred to date. While GBW may appear as a crude descriptor of an individual ’ s condition, it is among the few condition-dependent traits that we currently have at our disposal to capture individual variation in general state during molt in studies of e.g. carry-over eff ects or life-history trade-off s. Although other methods, such as incorporation of stress markers in the growing feathers, may aff ord a valuable alternative, GBW may remain one among the few proxies of condition during molt for a long time. As a non-negligible advantage, feather growth rate integrates infor-mation on condition over several days or weeks, depending on synchrony of molt among plumage regions. How-ever, more eff ort needs to be devoted to identifying the mechanisms behind variation in GBW, to improve standard-ization of the measurement protocols and to better charac-terize the components of physiological condition that are refl ected by ptilochronological variables.
References
Ambrosini, R., Ferrari, R. P., Martinelli, R., Romano, M. and Saino, N. 2006. Seasonal, meteorological, and micro-habitat eff ects on breeding success and off spring phenotype in the barn swallow, Hirundo rustica . – Ecoscience 13: 298 – 307.
Andersson, S. 1994. Costs of sexual advertising in the lekking Jackson’s widowbird. – Condor 96: 1 – 10.
Apanius, V. 1998. Stress and immune response. – Adv. Study Behav. 27: 133 – 154 .

523
Murphy, M. E. 1996. Energetics and nutrition in molt. – In: Carey, C. (ed.), Avian energetics and nutritional ecology. Chapman and Hall, pp. 158 – 198.
Murphy, M. E. and King, J. R. 1991. Ptilochronology: a critical evaluation of assumptions and utility. – Auk 108: 695 – 704.
Murphy, M. E. and King, J. R. 1992. Energy and nutrient use during moult by the white-crowned sparrow Zonotrichia leucophrys gambelii . – Ornis Scand. 23: 304 – 313.
Newton, I. 2008. Th e migration ecology of birds. – Academic Press.
Nilsson, J.- Å . and Svensson, M. 1996. Sibling competition aff ects nestling growth strategy in marsh tits. – J. Anim. Ecol. 65: 825 – 836.
Norris, D. R. 2005. Carry-over eff ects and habitat quality in migra-tory populations. – Oikos 109: 178 – 186.
Norris, D. R., Marra, P. P., Kyser, T. K., Sherry, T. W. and Ratcliff e, L. M. 2004. Tropical winter habitat limits reproduc-tive success on the temporal breeding grounds in a migratory bird. – Proc. R. Soc. B 271: 59 – 64.
Pap, P. L., Tokolyi, J. and Sz é p, T. 2005. Frequency and conse-quences of feather holes in barn swallows Hirundo rustica . – Ibis 147: 169 – 175.
Pennycuick, C. J. 1989. Bird fl ight performance: a practical calcu-lation manual. – Oxford Univ. Press.
Perez-Tris, J., Carbonell, R. and Telleria, J. L. 2000. Abundance distribution, morphological variation and juvenile condition of robins, Erithacus rubecula (L.), in their Mediterranean range boundary. – J. Biogeogr. 27: 879 – 888.
Riddle, O. 1907. A study of fundamental bars in feathers. – Biol. Bull. 12: 165 – 174.
Riddle, O. 1908. Th e genesis of fault bars in feathers and the cause of alternation of light and dark fundamental bars. – Biol. Bull. 14: 328 – 371.
Ritchie, B. W., Harrison, G. J. and Harrison, L. R. 1994. Avian medicine: principles and application. – Wingers.
Robb, G. N., McDonald, R. A., Chamberlain, D. E., Reynolds, S. J., Harrison, T. J. E. and Bearhop, S. 2008. Winter feeding increases productivity in the subsequent breeding season. – Biol. Lett. 4: 220 – 223.
Roff , D. A. 1992. Life history evolution. – Sinauer. Romano, A., Rubolini, D., Caprioli, M., Boncoraglio, G.,
Ambrosini, R. and Saino, N. 2011. Sex-related eff ects of an immune challenge on growth and begging behavior of barn swallow nestlings. – PLoS One 6: e22805.
Romero, L. M. 2002. Seasonal changes in plasma glucocorticoid concentrations in free-living vertebrates. – Gen. Comp. Endocrinol. 128: 1 – 24.
Romero, L. M., Strochlic, D. E. and Wingfi eld, J. C. 2005. Corticosterone inhibits feather growth: potential mechanism explaining seasonal down regulation of corticosterone during molt. – Comp. Biochem. Physiol. A 142: 65 – 73.
Rubolini, D., Massi, A. and Spina, F. 2002. Replacement of body feathers is associated with low premigratory energy stores in a long-distance migratory bird, the barn swallow (Hirundo rustica ). – J. Zool. 258: 441 – 447.
Saino, N., Calza, S., Ninni, P. and M ø ller, A. P. 1999. Barn swallows trade survival against off spring condition and immunocompetence. – J. Anim. Ecol. 68: 999 – 1009.
Saino, N., Ambrosini, R., Martinelli, R. and M ø ller, A. P. 2002. Mate fi delity, senescence in breeding performance and repro-ductive trade-off s in the barn swallow. – J. Anim. Ecol. 71: 309 – 319.
Saino, N., Sz é p, T., Ambrosini, R., Romano, M. and M ø ller, A. P. 2004a. Ecological conditions during winter aff ect sexual selec-tion and breeding in a migratory bird. – Proc. R. Soc. B 271: 681 – 686.
Saino, N., Sz é p, T., Romano, M., Rubolini, D., Spina, F. and M ø ller, A. P. 2004b. Ecological conditions during winter
Gr ü ebler, M. U. and Naef-Daenzer, B. 2008. Fitness consequences of pre- and post-fl edging timing decisions in a double-brooded passerine. – Ecology 89: 2736 – 2745.
Gunnarsson, T. G., Gill, J. A., Newton, J., Potts, P. M. and Sutherland, W. J. 2005. Seasonal matching of habitat quality and fi tness in a migratory bird. – Proc. R. Soc. B 272: 2319 – 2323.
Gunnarsson, T. G., Gill, J. A., Atkinson, P. W., Gelinaud, G., Potts, P. M., Croger, R. E., Gudmundsson, G. A., Appleton, G. F. and Sutherland, W. J. 2006. Population-scale drivers of individual arrival times in migratory birds. – J. Anim. Ecol. 75: 1119 – 1127.
Harrison, X. A., Blount, J. D., Inger, R., Norris, D. R. and Bearhop, S. 2011. Carry-over eff ects as drivers of fi tness diff erences in animals. – J. Anim. Ecol. 80: 4 – 18.
Ilmonen, P., Taarna, T. and Hasselquist, D. 2000. Experimental activated immune defence in females pied fl ycatchers results in reduced breeding success. – Proc. R. Soc. B 267: 665 – 670.
Inger, R., Harrison, X. A., Ruxton, G. D., Newton, J., Colhoun, K., Gudmundsson, G. A., Mcelwaine, G., Pickford, M., Hogson, D. and Bearhop, S. 2010. Carry-over eff ects reveal cryptic reproductive costs in a long distance migrant. – J. Anim. Ecol. 79: 974 – 982.
Jenkins, K. D., Hawley, D. M., Farabaugh, C. S. and Cristol, D. A. 2001. Ptilochronology reveals diff erences in condition of captive white-throated sparrows. – Condor 103: 579 – 586.
Jenni, L. and Winkler, R. 1994. Moult and ageing of european passerines. – Academic Press.
Jovani, R. and Blas, J. 2004. Adaptive allocation of stress-induced deformities on bird feathers. – J. Evol. Biol. 17: 294 – 301.
Jovani, R. and Diaz-Real, J. 2012. Fault bars timing and duration: the power of studying feather fault bars and growth bands together. – J. Avian Biol. 43: 97 – 101.
Jovani, R., Blas, J., Navarro, C. and Mougeot, F. 2011. Feathers growth bands and photoperiod. – J. Avian Biol. 42: 1 – 4.
Kern, M. D. and Cowie, R. J. 2002. Ptilochronology proves unreliable in studies of nestling pied fl ycatchers. – Ibis 144: 23 – 29.
Legagneux, P., Gauthier, G., Chastel, O., Picard, G. and Bety, J. 2011. Do glucocorticoids in droppings refl ect baseline level in birds captured in the wild? A case study in snow geese. – Gen. Comp. Endocrinol. 172: 440 – 445.
Lindstr ö m, A., Visser, G. H. and Daan, S. 1993. Th e energetic cost of feather synthesis is proportional to basal metabolic rate. – Physiol. Zool. 66: 490 – 510.
Lindstr ö m, J. 1999. Early development and fi tness in birds and mammals. – Trends Ecol. Evol. 14: 343 – 348.
Michener, H. and Michener, J. R. 1938. Bars in fl ight feathers. – Condor 40: 149 – 160.
M ø ller, A. P. 1994. Sexual selection and the barn swallow. – Oxford Univ. Press.
M ø ller, A. P., Magnhagen, C., Ulfstrand, A. and Ulfstrand, S. 1995. Phenotypic quality and molt in the barn swallow, Hirundo rustica . – Behav. Ecol. 6: 242 – 249.
M ø ller, A. P., Erritz ø e, J. and Nielse, J. T. 2009. Frequency of fault bars in feathers of birds and susceptibility to predation. – Biol. J. Linn. Soc. 97: 334 – 345.
M ø ller, A. P., Fiedler, W. and Berthold, P. 2010. Eff ects of climate change on birds. – Oxford Univ. Press.
M ø ller, A. P., Nuttall, R., Piper, S. E., Sz é p, T. and Vickers, E. J. 2011. Migration, moult and climate change in barn swallows Hirundo rustica in South Africa. – Clim. Res. 47: 201 – 205.
Moreno-Rueda, G. 2010. Experimental test of a trade-off between moult and immune response in house sparrows Passer domesticus. – J. Evol. Biol. 23: 2229 – 2237.
Murphy, M. E. 1992. Ptilochronology: accuracy and reliability of the technique. – Auk 109: 676 – 680.

524
Stutchbury, B. J. M., Gow, E. A., Done, T., MacPherson, M., Fox, J. W. and Afanasyev, V. 2011. Eff ects of post-breeding moult and energetic condition on timing of songbird migration into the tropics. – Proc. R. Soc. B 278: 131 – 137.
Sutherland, W. J. 1996. From individual behaviour to population ecology. – Oxford Univ. Press.
Swaddle, J. P., Williams, E. V. and Rayner, J. M. V. 1999. Th e eff ect of simulated fl ight feather moult on escape take-off performance in starlings. – J. Avian Biol. 30: 351 – 358.
Takaki, Y., Eguchi, K. and Nagata, H. 2001. Th e growth bars on tail feathers in the male Styan ’ s grasshopper warbler may indicate quality. – J. Avian Biol. 32: 319 – 325.
Turner, A. 2006. Th e barn swallow. – T and AD Poyser. van Noordwijk, A. J. and de Jong, G. 1986. Acquisition and
allocation of resources: their infl uence on variation in life his-tory tactics. – Am. Nat. 128: 137 – 142.
Vangestel, C., Braeckman, B. P., Matheve, H. and Lens, L. 2010. Constraints on home range behaviour aff ect nutritional condition in urban house sparrows ( Passer domesticus ). – Biol. J. Linn. Soc. 101: 41 – 50.
Verhulst, S. and Nilsson, J. A. 2008. Th e timing of birds ’ breeding seasons: a review of experiments that manipulated timing of breeding. – Proc. R. Soc. B 363: 399 – 410.
Weatherhead, P. J. 2005. Long-term decline in a red-winged blackbird population: ecological causes and sexual selection consequences. – Proc. R. Soc. B 272: 2313 – 2317.
White, D. W., Kennedy, E. D. and Stouff er, P. C. 1991. Feather regrowth in female European starlings rearing broods of diff erent sizes. – Auk 108: 889 – 895.
Wood, H. B. 1950. Growth bars in feathers. – Auk 67: 486 – 491.
Zera, A. J. and Harshman, L. G. 2001. Th e physiology of life history trade-off s in animals. – Annu. Rev. Ecol. Syst. 32: 95 – 126.
predict arrival date at the breeding quarters in a trans-Saharan migratory bird. – Ecol. Lett. 7: 21 – 25.
Saino, N., Rubolini, D., Serra, L., Caprioli, M., Morganti, M., Ambrosini, R. and Spina, F. 2010. Sex-related variation in migration phenology in relation to sexual dimorphism: a test of competing hypotheses for the evolution of protandry. – J. Evol. Biol. 23: 2054 – 2065.
Saino, N., Caprioli, M., Romano, M., Boncoraglio, G., Rubolini, D., Ambrosini, R., Bonisoli-Alquati, A. and Romano, A. 2011. Antioxidant defenses predict long-term survival in a passerine bird. – PLoS One 6: e19593.
Sanz, J. J., Moreno, J., Merino, S. and Tom á s, G. 2004. A trade-off between two resource-demanding functions: post-nuptial moult and immunity during reproduction in male pied fl ycatcher. – J. Anim. Ecol. 73: 441 – 447.
Sapolsky, R. M., Romero, L. M. and Munck, A. U. 2000. How do glucocorticoids fl uence stress-responses? Integrating permis-sive, suppressive, stimulatory, and adaptive actions. – Endocr. Rev. 21: 55 – 89.
Sedinger, J. S., Schamber, J. L., Ward, D. H., Nicolai, C. A. and Conant, B. 2011. Carryover eff ects associated with winter location aff ect fi tness, social status, and population dynamics in a long-distance migrant. – Am. Nat. 178: E110–E123.
Serrano, D. and Jovani, R. 2005. Adaptive fault bar distribution in a long-distance migratory, aerial forager passerine? – Biol. J. Linn. Soc. 85: 455 – 461.
Spottiswoode, C. N. and Saino, N. 2010. Sexual selection and climate change. – In: M ø ller, A. P., Fiedler, W. and Berthold, P. (eds), Sexual selection and climate change. Oxford Univ. Press, pp. 169 – 189.
Stearns, S. C. 1992. Th e evolution of life histories. – Oxford Univ. Press.
Stratford, J. A. and Stouff er, P. C. 2001. Reduced feather growth rates of two common birds inhabiting central Amazonian forest fragments. – Conserv. Biol. 15: 721 – 728.




















