
A Phylogenetic Test of Classical Species Groups in Argia (Odonata: Coenagrionidae)
12
A PHYLOGENETIC TEST OF CLASSICAL SPECIES GROUPS IN ARGIA (ODONATA: COENAGRIONIDAE) RYAN M. CAESAR 1,2 AND JOHN W. WENZEL 1 1 Department of Entomology, Ohio State University, Columbus, OH Abstract.—We present the first cladistic analysis of Argia species, focusing on those occurring in North America north of Mexico. Our analysis is based on mitochondrial 16S rDNA and morphological characters of both sexes of adults and immatures. We reexamine classical work on Argia taxonomy and phylogeny. Our results agree considerably with previous hypotheses based morphology in an absence of phylogenetic analysis, and thus our work represents and independent test of these previous hypotheses. Argia is recovered as monophyletic. The clade composed of A. funcki plus A. lugens is basal among the species studied here. The species A. fumipennis, including the three subspecies, appears to be a paraphyletic assemblage, and thus may warrant being considered separate species as originally described. The feasibility of producing a thorough phylogenetic analysis of the entire genus using multiple sources of data is discussed. Key words: Argia, Coenagrionidae, Odonata, damselfly, phylogeny. INTRODUCTION Rambur described the genus Argia (Odonata: Zygoptera: Coenagrionidae: Argiinae) in 1842. Argia is extremely speciose in comparison to other odonate genera, with 118 valid described species (Garrison, 1994; Garrison and von Ellenrieder, 2007). The genus occurs throughout the New World, with its greatest diversity occurring in the neotropical region. There are approximately thirty six species that can be found in North America north of Mexico, with the highest diversity in the southwestern United States; twenty three species are known to occur in southeastern Arizona alone. The remaining species occur in subtropical, tropical and temperate regions of Meso- and South America. Populations of common Argia species can be quite large, and, like all odonates, they are voracious predators in all life stages. As such, they represent vital components of the trophic webs of aquatic ecosystems. Most species of Argia prefer low to mid order streams (Westfall and May, 1996), unlike the remainder of coenagrionids that tend to occur in lentic systems (Dunkle, 1990). Several species are rare, endemic, or threatened, and may be of conservation concern. For example, in Ohio, A. bipunctulata (Hagen) is listed as an endangered species, due to its restriction in the state mainly to the 182 hectare Cedar Bog Preserve and a few other fen habitats (Moody, 2002), although the species is fairly common in adjacent states. The recently described species A. sabino Garrison is known from very few localities in Arizona, most of which are continually threatened by forest fire, and thus it is considered a species of concern by biologists in that state (D. Turner, pers. comm.) Some of the more common species have been thoroughly studied from a behavioral and ecolog- ical perspective (Borror, 1934; Bick and Bick, 1965, 1971, 1982; Robinson et al., 1983; Conrad, 1992; Conrad and Pritchard, 1988, 1990). Little is known about the biology of most of the tropical species, and there has never been a phylogenetic study of the genus. The current taxonomy of Argia species has been established in the absence of an explicit phyloge- netic hypothesis based on modern comparative methods. In his work on Central American Odonata, Calvert treated forty eight species of Argia (Calvert, 1901), eighteen of which occur in the United States (Garrison, 1994). Kennedy included some Argia species in his ‘‘phylogeny’’ of Zygoptera (Kennedy, 1920a), and he described several species (Kennedy, 1918, 1919). Leonora Gloyd did considerable taxonomic work on Argia throughout her life (Gloyd, 1958, 1968a, 1968b), although she died before much of her work was completed (Garrison, 1994). The larvae of some Mexican species were treated by Novelo-Gutierrez (1992). Garrison (1994) provided a thorough 2 E-mail address for correspondence: caesar.6@ osu.edu Entomologica Americana 115(2):97–108, 2009
-
Upload
carnegiemnh -
Category
Documents
-
view
1 -
download
0
Transcript of A Phylogenetic Test of Classical Species Groups in Argia (Odonata: Coenagrionidae)

A PHYLOGENETIC TEST OF CLASSICAL SPECIES GROUPS IN ARGIA(ODONATA: COENAGRIONIDAE)
RYAN M. CAESAR1,2
AND JOHN W. WENZEL1
1Department of Entomology, Ohio State University, Columbus, OH
Abstract.—We present the first cladistic analysis of Argia species, focusing on those occurring in North America
north of Mexico. Our analysis is based on mitochondrial 16S rDNA and morphological characters of both sexes of
adults and immatures. We reexamine classical work on Argia taxonomy and phylogeny. Our results agree
considerably with previous hypotheses based morphology in an absence of phylogenetic analysis, and thus our work
represents and independent test of these previous hypotheses. Argia is recovered as monophyletic. The clade
composed of A. funcki plus A. lugens is basal among the species studied here. The species A. fumipennis, including the
three subspecies, appears to be a paraphyletic assemblage, and thus may warrant being considered separate species as
originally described. The feasibility of producing a thorough phylogenetic analysis of the entire genus using multiple
sources of data is discussed.
Key words: Argia, Coenagrionidae, Odonata, damselfly, phylogeny.
INTRODUCTION
Rambur described the genus Argia (Odonata:
Zygoptera: Coenagrionidae: Argiinae) in 1842.
Argia is extremely speciose in comparison to other
odonate genera, with 118 valid described species
(Garrison, 1994; Garrison and von Ellenrieder,
2007). The genus occurs throughout the New
World, with its greatest diversity occurring in the
neotropical region. There are approximately thirty
six species that can be found in North America
north of Mexico, with the highest diversity in the
southwestern United States; twenty three species
are known to occur in southeastern Arizona
alone. The remaining species occur in subtropical,
tropical and temperate regions of Meso- and
South America. Populations of common Argia
species can be quite large, and, like all odonates,
they are voracious predators in all life stages. As
such, they represent vital components of the
trophic webs of aquatic ecosystems.
Most species of Argia prefer low to mid order
streams (Westfall and May, 1996), unlike the
remainder of coenagrionids that tend to occur in
lentic systems (Dunkle, 1990). Several species are
rare, endemic, or threatened, and may be of
conservation concern. For example, in Ohio, A.
bipunctulata (Hagen) is listed as an endangered
species, due to its restriction in the state mainly to
the 182 hectare Cedar Bog Preserve and a few
other fen habitats (Moody, 2002), although the
species is fairly common in adjacent states. The
recently described species A. sabino Garrison is
known from very few localities in Arizona, most
of which are continually threatened by forest fire,
and thus it is considered a species of concern by
biologists in that state (D. Turner, pers. comm.)
Some of the more common species have been
thoroughly studied from a behavioral and ecolog-
ical perspective (Borror, 1934; Bick and Bick,
1965, 1971, 1982; Robinson et al., 1983; Conrad,
1992; Conrad and Pritchard, 1988, 1990). Little is
known about the biology of most of the tropical
species, and there has never been a phylogenetic
study of the genus.
The current taxonomy of Argia species has been
established in the absence of an explicit phyloge-
netic hypothesis based on modern comparative
methods. In his work on Central American
Odonata, Calvert treated forty eight species of
Argia (Calvert, 1901), eighteen of which occur in
the United States (Garrison, 1994). Kennedy
included some Argia species in his ‘‘phylogeny’’
of Zygoptera (Kennedy, 1920a), and he described
several species (Kennedy, 1918, 1919). Leonora
Gloyd did considerable taxonomic work on Argia
throughout her life (Gloyd, 1958, 1968a, 1968b),
although she died before much of her work was
completed (Garrison, 1994). The larvae of some
Mexican species were treated by Novelo-Gutierrez
(1992). Garrison (1994) provided a thorough
2 E-mail address for correspondence: caesar.6@
osu.edu
Entomologica Americana 115(2):97–108, 2009

synopsis of the species of Argia occurring north of
Mexico, including taxonomic keys for adults
(these keys are reproduced in Westfall and May
1996), and several informal species groups were
outlined. Forster (2001) provides updated taxo-
nomic keys for some of the more common Central
American species of Argia. Garrison is currently
continuing the work of Leonora Gloyd on
revising the tropical species (pers. comm.). Many
undescribed species are thought to exist, and new
species descriptions continue to be published
(Daigle, 1991, 1995; Garrison, 1994, 1996; Garri-
son and von Ellenrieder, 2007). Here we provide
the first modern phylogenetic hypothesis for Argia
species based on multiple data sources and using
two phylogenetic optimality criteria. These pre-
liminary phylogenetic analyses allows us to test
existing taxonomic hypotheses, contribute an
improved understanding of species relationships,
and provide a foundation for resolving the
phylogeny of the genus.
MATERIALS AND METHODS
Taxon sampling
Our matrix includes thirty eight of the 118
described Argia species, including nearly all of
those found north of Mexico as well as several
from Mesoamerica. A. bipunctulata, A. barretti
Calvert and A. carlcooki Daigle are the only
species currently known to occur north of Mexico
that are missing from our matrix. Collection
records for specimens used in this study are listed
in Table 1, along with GenBank accession num-
bers. Where possible, we include in this study
adult specimens that were recently collected in the
field using a hand held aerial net and deposited
directly into 95% ethyl alcohol for preservation.
Additional specimens were donated by other
collectors utilizing various collection and preser-
vation techniques, or were borrowed from the
International Odonata Research Institute (IORI;
Gainesville, Florida). Additionally, we did not
attempt to collect molecular data from some
species for which few or single specimens were
available.
Species were identified on the basis of published
keys based on morphological characters (Garri-
son, 1994; Westfall and May, 1996; Abbott, 2005.)
A. sp. nov. is an undescribed species collected in
northern Mexico; it has been known for several
years but remains undescribed. The sister group of
Argia is not known; we include six outgroup taxa
representing other damselfly lineages. 16S sequenc-
es for these outgroup species were taken from
GenBank (Table 1.) Specimens used in this study
are deposited as vouchers at either the IORI or the
Charles Triplehorn Insect Collection at the Ohio
State University (OSUC) in Columbus, Ohio.
Character analysis
We coded ten morphological characters from
imagos of both sexes as well as larvae for 35 Argia
species. Characters of the head, thorax and
abdomen, including secondary sexual characters
of both sexes, are represented. Characters were
initially chosen based on information in published
dichotomous keys. These characters are unambig-
uous in their interpretation and do not seem to
vary within species. The morphological matrix is
presented in Table 2.
Character 1. Thorax and head with metallic
copper-red coloration (imagos): absent (0), pre-
sent (1). This condition is also associated with
copper-red to red eyes for specimens in vivo
(imagos.) Species with this coloration are often
referred to as being associated together in the
‘‘metallics’’ group. While pigmentation is gener-
ally not a very useful character for damselflies, as
it is often variable within species, this condition is
largely a result of structural coloration and is
invariant within species.
Character 2. Mesepisternal tubercles (females):
absent (0), reduced (1), prominent (2). See Fig. 1.
Character 3. Posterior lobes of mesostigmal
plates (females): absent (0), broad and flange-like
(1), elongate and finger-like (2). See Fig. 1.
Character 4. Mesothoracic pits (females): absent
(0), shallow (1), deep (2). This character has been
discussed in very limited context in the literature,
but it has been suggested that it might be
informative for phylogenetics (Gloyd, 1958). Here
we code it for the first time and show that it is useful.
Character 5. Hairs lining mesothoracic pits
(females): absent (0), sparse (1), dense (2).
Character 6. Mesepisternal pits costate (fe-
males): absent (0), present (1).
Character 7. Pronotal pits (females): absent (0),
shallow (1), deep (2). This character is not utilized
in keys or discussed much in the literature, but
seems to be variable enough within Argia to be
useful. Indeed, this region corresponds to the
placement of the dorso-posterior potion of the
98 ENTOMOLOGICA AMERICANA Vol. 115(2)

male paraprocts during tandem linkage and
copulation (see Fig. 1.)
Character 8. Shape of cerci (males): entire (0),
bifid (1), trifid (2). Cerci are a secondary sexual
character, utilized as part of the clasping process
during copulation. The cerci contact the female
mesostigmal plates and mesepisternal tubercules
when linked in copula and may be part of the
Table 1. Taxonomic information, collection records and GenBank accession numbers for Argia and outgroup
species used in phylogenetic analyses.
Taxon Authority Sampling locality Collector
GenBank
accession
number
Calopteryx aequibilis Say, 1839 unknown GenBank AF170961
C. maculata (Beauvois, 1805) unknown GenBank AF170960
Hetaerina americana (Fabricius, 1798) unknown GenBank AF170951
Neoneura esthera Williamson, 1917 unknown GenBank AF170948
Ceriagrion nipponicum Asahina, 1967 unknown GenBank AB127067
Argia agrioides Calvert, 1895 USA: Oregon R. Caesar FJ592218
A. alberta Kennedy, 1918 USA: New Mexico J. Abbott FJ592211
A. anceps Garrison, 1996 Mexico: Hidalgo K. Tennessen FJ592233
A. apicalis (Say, 1839) USA: Ohio R. Caesar FJ592212
A. cuprea (Hagen, 1861) USA: Texas T. Gallucci FJ592227
A. emma Kennedy, 1915 USA: California C. Barrett FJ592228
A. extranea (Hagen, 1861) Mexico: Sonora R. Behrstock FJ592231
A. fumipennis atra (Burmeister, 1839) USA: Florida K. Holt FJ592230
A. fumipennis violacea (Burmeister, 1839) USA: Ohio R. Caesar FJ592232
A. funcki (Selys, 1854) Mexico: Sonora D. Paulson FJ592197
A. garrisoni Daigle, 1991 Mexico: Tamaulipas R. Behrstock FJ592213
A. harknessi Calvert, 1899 Mexico: Sonora D. Paulson FJ592199
A. hinei Kennedy, 1918 USA: Texas J. Abbott FJ592207
A. immunda (Hagen, 1861) USA: Texas A. Cognato FJ592214
A. lacrimans (Hagen, 1861) USA: Arizona R. Behrstock FJ592216
A. leonorae Garrison, 1994 USA: Texas R. Behrstock FJ592226
A. lugens (Hagen, 1861) USA: Arizona R. Caesar FJ592215
A. moesta (Hagen, 1861) USA: Texas A. Cognato FJ592229
A. munda Calvert, 1902 USA: Arizona R. Caesar FJ592223
A. nahuana Calvert, 1902 USA: Texas T. Gallucci FJ592225
A. sp. nov. Not applicable Mexico: Sonora D. Paulson FJ592198
A. oculata Hagen in Selys, 1865 Mexico: Tamaulipas R. Behrstock FJ592221
A. oenea Hagen in Selys, 1865 Mexico: San Luis Potosi R. Behrstock FJ592217
A. pallens Calvert, 1902 USA: Arizona W. Mauffray FJ592224
A. pima Garrison, 1994 USA: Arizona J. Daigle FJ592208
A. plana Calvert, 1902 USA: Texas J. Abbott FJ592196
A. pulla Hagen in Selys, 1865 Nicaragua: Jinotega J. Abbott FJ592222
A. rhoadsi Calvert, 1902 Mexico: San Luis Potosi R. Behrstock FJ592206
A. sabino Garrison, 1994 USA: Arizona J. Daigle FJ592202
A. sedula (Hagen, 1861) USA: Oklahoma H. Song FJ592209
A. tarascana Calvert, 1902 USA: Arizona R. Caesar FJ592200
A. tezpi Calvert, 1902 Honduras: Francisco
Morazan
S. Dunkle FJ592220
A. tibialis (Rambur, 1842) USA: Ohio R. Caesar FJ592203
A. tonto Calvert, 1902 USA: Arizona R. Caesar FJ592204
A. translata Hagen in Selys, 1865 USA: Texas A. Cognato FJ592210
A. ulmeca Calvert, 1907 Mexico: Tamaulipas R. Behrstock FJ592205
A. vivida Hagen in Selys, 1865 USA: California J. Abbott FJ592201
A. westfalli Garrison, 1996 Mexico: Tamaulipas R. Behrstock FJ592219
2009 ARGIA PHYLOGENY 99

mechanism by which females recognize and
evaluate males.
Character 9. Shape of paraprocts (males): entire
(0), bifid (1), trifid (2).
Character 10. Lateral gills with marginal fringe
of stout setae (larvae): present (0), absent (1).
Genomic DNA was extracted from specimens
that were either freshly collected and preserved in
95% ethyl alcohol, or older museum specimens
that were acetone-dried (up to 20 years old). A
modification of the animal tissue protocol for
Quiagen DNeasy extraction kits (Quiagen, Valen-
cia, CA, USA; Caesar et al., 2005) was used to
extract DNA from leg and thoracic muscle tissue
that was isolated from specimens using sterile
techniques. Dried specimens from which DNA
was extracted were labeled as DNA vouchers.
DNA template vouchers are stored at 280uC in
the Wenzel Laboratory at the Museum of
Biological Diversity, Ohio State University
(MBD.)
16S ribosomal DNA (0.6 kb) from the mito-
chondria was amplified by the polymerase chain
reaction (PCR) run at 35 cycles with annealing
temperature of 50uC. We used the primers LR-J-
12887 (59 CCGGTCTGAACTCAGATCACGT
39) and LR-N-13398 (59 CGCCTGTTTAA-
CAAAAACAT 39) (Simon et al., 1994), known
to be useful in several odonate studies (e.g., Misof
et al., 2000). Reactions were carried out in 25 ml
volumes, with 12.5 ml of a TAQ master mix
(Quiagen, Inc. Valencia, CA, USA), 3.5 ml of
H2O, 2.0 ml of each primer (0.5 pmol/ml), and
5.0 ml genomic DNA template. A layer of mineral
oil was applied to each sample, and reactions
Table 2. Morphological character matrix used for
parsimony analysis of Argia. See Methods for coding.
Taxon Characters (1–10)
Het.ameri ??????????A.plana1 0111202?11A.extrane 0101?01111A.vivida1 0211100011A.emma1 0222111111A.munda1 00???1?010A.anceps2 0110000000A.westfal 011120000?A.sabino1 011110211?A.lacrima 0?10001110A.tonto1 0010001110A.pima1 001??0?11?A.f.atra1 0121201211A.f.viola 0121201211A.pallens 0010001211A.funcki1 00212?110?A.lugens1 0122102100A.moesta1 0222201100A.hinei1 0020002211A.agrioid 0120001011A.alberta 0000000010A.leonora 000??0?01?A.nahuana 0110001011A.pulla1 0000001121A.rhoadsi 0022101001A.sedula1 0010102011A.tarasca 0221101110A.apicali 0100001210A.tezpi 0221101010A.tibiali 0000000110A.immunda 0001001111A.transla 0222102010A.ulmeca1 0101002110A.n.sp.1 0??????11?A.garriso 002000111?A.harknes 0?????????A.oculata 001100201?A.cuprea1 1221102100A.oenea1 1021102110Cal.aequa ??????????Cal.macul ??????????Ceri.nipp ??????????Neo.esthe ??????????
Fig. 1. Photo of the pro- and mesothorax of a
female of Argia emma, lateral oblique angle. Arrows
indicate some of the characters coded in the
morphological matrix.
100 ENTOMOLOGICA AMERICANA Vol. 115(2)

occurred on a PTC-100 Peltier thermal cycler (MJ
Research, Ramsey, Minnesota, USA). Amplified
DNA was separated in an agarose gel via
electrophoresis and visualized under UV light
for verification of presence of amplicon and to
check for contamination. PCR products were
purified using QIAquick PCR purification kit
(Quiagen, Valencia, CA, USA), and purified
DNA was sequenced at the Plant Microbe
Genomics Facility (Ohio State University, Co-
lumbus, OH) or at Cogenics (Houston, TX).
Editing and assembly of raw sequences was
performed using Sequencher 4.1 (Gene Codes
Corp., Ann Arbor, Michigan, USA). We se-
quenced PCR products in both directions, and
and differences or ambiguities between strands
were resolved by eye after examining the elecro-
pherograms of each sequence. Edited sequences
were initially aligned by eye in Sequencher 4.1,
then submitted to CLUSTALW (Thompson et al.,
1994) for further alignment using a 1:1 gap
opening: extension cost. After editing and align-
ment, we utilized 551 base pairs (bp) as our
molecular data matrix. All new sequences gener-
ated in this study were deposited in GenBank
(Table 1).
Phylogenetic analysis
Calopteryx was designated as outgroup for all
phylogenetic analyses. Parsimony analysis was
performed on the molecular data only, and in
combination with the morphological data. We
used the parsimony ratchet implemented in
NONA/WinClada (Nixon, 2000). By default, this
implements TBR branch swapping. Gaps in the
molecular matrix were treated as missing data.
Bremer support values were generated in NONA
using the command ‘‘.hold 15,000; bsupport 5;’’
to estimate relative clade support. This command
generates as many as 15,000 trees that are up to
five steps longer than the most parsimonious
trees.
Concordance among trees produced by differ-
ent methods of phylogenetic reconstructions is
considered by some to be a good test of
phylogenetic accuracy (Huelsenbeck, 1997). Thus,
we also performed Maximum Likelihood (ML)
analysis on the molecular data. We did this using
the program GARLI, version 0.96 (Zwickl, 2006),
which uses a genetic algorithm to rapidly search
for the nucleotide substitution model parameters,
branch lengths, and topology that maximize the
log likelihood score. Default parameters were
used for 100 search replicates.
RESULTS
In some cases, DNA extraction and/or PCR
amplification of the target sequence failed for
older museum specimens that were otherwise
available for study. Thus, our analyzed matrices
do not include all available species. The ClustalW-
aligned molecular matrix contains 126 informative
sites and yields 38 most parsimonious trees (489
steps, CI 5 0.54, RI 5 0.61) in the MP analysis.
The strict consensus (499 steps) of these is shown
in Fig. 2. A monophyletic Argia is recovered with
good support, but a basal polytomy with several
weakly supported clades follows. The ML analysis
of the molecular data produced two trees that
differ only in the resolution of the clade contain-
ing the three species A. apicalis (Say), A. tarascana
Calvert, and A. tezpi Calvert. The ML tree with
the best likelihood score is shown in Fig. 3.
Analysis of the morphological matrix (Table 2)
alone yields trees (not shown) that are poorly
resolved and not informative. Combining the
morphological and molecular characters, we
obtained six most parsimonious trees (573 steps,
CI 5 0.49, RI 5 0.58). The consensus (578 steps)
is shown in Fig. 4. These morphological data in
the combined analysis improved resolution in
comparison with the molecular data alone, with
few changes in topology. Again Argia is recovered
as monophyletic, although the Bremer support
value improves from three to five. The basal
polytomy resulting from the molecular only data
is resolved, and we recover the clade of A. funcki
(Selys) + A. lugens (Hagen) as basal to the rest of
the genus. In the combined tree, A. moesta
(Hagen) is more basal, closer to A. translata
Hagen; A. munda Calvert falls out of the clade to
which it belonged, sister to the component
including A. pima Garrison and A. rhoadsi
Calvert. Some taxa are derived more apically in
the combined tree compared to the molecular tree,
such as A. oculata Hagen, which is no longer sister
to A. oenea Hagen in the basal paraphyly at the
ingroup node of Fig. 2. The polytomy in the
middle of the molecular tree (the component
including A. alberta Kennedy, A. nahuana Calvert,
A. leonorae Garrison, to A. anceps Garrison) has
better resolution, and the A. pima-A. westfalli
Garrison component is now sister to the remain-
2009 ARGIA PHYLOGENY 101

Fig. 2. Strict consensus of 38 equally parsimonious trees resulting from analysis of 16S data alone. Clades with
dots are recovered in both ML and parsimony analyses. Numbers below branches are Bremer support values.
102 ENTOMOLOGICA AMERICANA Vol. 115(2)

der, with the A. emma Kennedy-A. extranea
(Hagen) group coming out next.
DISCUSSION
All but three clades from the MP consensus
from the combined matrix are found in the ML
trees, as indicated by the dots on the shared nodes
of the MP trees (Figs. 2, 4), and the ML trees
(Fig. 3), demonstrating that the solution to our
16S data is generally stable, despite optimality
criterion. The main difference is that the MP tree
places A. funcki and A. lugens rather basally, and
the A. rhoadsi and A. sedula (Hagen) group rather
apically, whereas the ML tree places these lineages
in the middle of the tree. This rather modest
difference has the undesirable effect of compro-
mising the monophyly of most groups along the
spine of the tree even if the great majority of
fundamental relationships remain the same.
Monophyly of Argia is strongly supported by
both data partitions and in all phylogenetic
analyses conducted for this study (Figs. 2, 3, and
4). This is not surprising, and there are additional
morphological features that are autapomorphic
for Argia relative to other Coenagrionidae that
support monophyly of Argia. These include the
long length of tibial spines (Westfall and May,
1996) and absence of an angulate frons (Carle et
al., 2008). O’Grady and May also recovered Argia
as monophyletic in their morphological analysis
of Coenagrionidae (2003), although only three
species were included. We think that this result
will be stable to the addition of data in future
analyses.
The solutions to the molecular-only (Fig. 2) or
combined (Fig. 4) matrices appear very different,
and a strict consensus of these trees provides little
resolution. Strict consensus trees are usually used
to illustrate groups that are monophyletic for all
solutions, but it is not very useful when there is a
great degree of agreement for certain networks of
taxa among otherwise competing solutions. Prob-
lematic taxa and rooting issues that interrupt
networks are often not disputed among very
different solutions. While we are always interested
in recovering monophyletic of lineages, especially
for taxonomic work, analysis of ‘‘species groups’’
should also focus on stable networks of terminals,
even if their placement is ambiguous. We offer
Fig. 5 to demonstrate that the solutions of Figs. 2
and 4 are easily interpretable to be closely related,
despite a poorly resolved consensus. Our discus-
sion is based on three agreement subtrees, each of
which is common to all solutions.
We concentrate on the clade that is sister to A.
oculata, where the appearance of discord is most
evident. Excluded from discussion are A. munda
and the pair A. agrioides Calvert and A. hinei
Kennedy, because they are not informative for
demonstrating agreement among trees. The first
comparison is the sedula-pulla-rhoadsi component
Fig. 3. Best maximum likelihood tree generated by analysis of the 16S data using GARLI.
2009 ARGIA PHYLOGENY 103

(Fig. 5A). This component can be placed either
basally near A. oculata (Fig. 2), or apically in the
clade of interest (Fig. 4). In both cases the root of
this component is between A. sedula and the pair
A. pulla Hagen in Selys plus A. rhoadsi. Next we
discuss the component that includes the clades
emma-extranea, and pima-westfalli (Fig. 5B). This
combination of ten species is topologically iden-
tical in both Figs. 2 and 4, and also rooted in the
same place. The difference is that in Fig. 2 the
component is apical in the tree and monophyletic,
whereas in Fig. 4 it is deeper in the tree and
Fig. 4. Strict consensus of six equally parsimonious trees generated from combined 16S and morphology data.
Clades with dots are recovered in ML analyses of 16S alone. Numbers below branches are Bremer support values.
104 ENTOMOLOGICA AMERICANA Vol. 115(2)
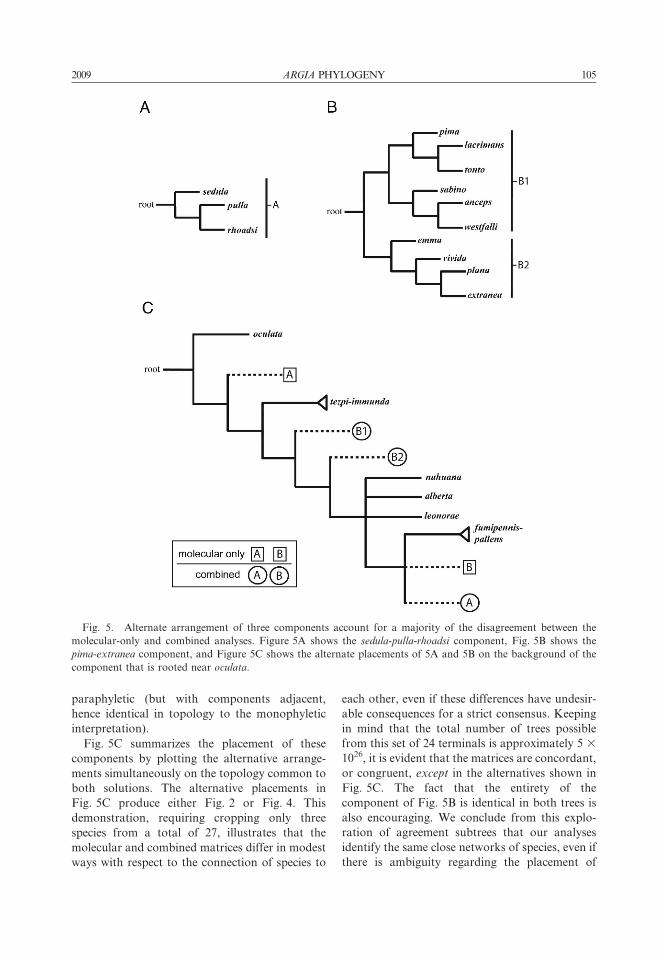
paraphyletic (but with components adjacent,
hence identical in topology to the monophyletic
interpretation).
Fig. 5C summarizes the placement of these
components by plotting the alternative arrange-
ments simultaneously on the topology common to
both solutions. The alternative placements in
Fig. 5C produce either Fig. 2 or Fig. 4. This
demonstration, requiring cropping only three
species from a total of 27, illustrates that the
molecular and combined matrices differ in modest
ways with respect to the connection of species to
each other, even if these differences have undesir-
able consequences for a strict consensus. Keeping
in mind that the total number of trees possible
from this set of 24 terminals is approximately 5 3
1026, it is evident that the matrices are concordant,
or congruent, except in the alternatives shown in
Fig. 5C. The fact that the entirety of the
component of Fig. 5B is identical in both trees is
also encouraging. We conclude from this explo-
ration of agreement subtrees that our analyses
identify the same close networks of species, even if
there is ambiguity regarding the placement of
Fig. 5. Alternate arrangement of three components account for a majority of the disagreement between the
molecular-only and combined analyses. Figure 5A shows the sedula-pulla-rhoadsi component, Fig. 5B shows the
pima-extranea component, and Figure 5C shows the alternate placements of 5A and 5B on the background of the
component that is rooted near oculata.
2009 ARGIA PHYLOGENY 105

those networks. The main lesson is that morpho-
logical data provide adequate phylogenetic signal
to move the sedula-pulla-rhoadsi group apically,
away from A. oculata Hagen in Selys, and place
the component of Fig. 5b to a more basal portion
of the tree, even in combination with molecular
data. These findings demonstrate the importance
of morphological data to complement molecular
data in Argia, and warrant further investigation of
potentially informative morphological characters.
The combined parsimony tree (Fig. 4) in many
ways accords with traditional groupings of species
based on morphology alone. Having a phylogeny
with which to compare to these previous non-
phylogenetic hypotheses of relationship allows us
to provide a test, and a revised hypothesis.
Because our phylogeny is based on different
characters and analytical methods, it represents
an independent test of the classical morphology-
based species groups of Calvert, Kennedy, Gloyd,
Garrison and others.
The close relationship between A. funcki and A.
lugens, and their sister relationship to the remain-
ing Argia, reflects the original placement of these
species in the genus Hyponeura by Selys (1865) on
the basis of veinational characters. Hyponeura,
along with Diargia Calvert (which included the
species bicellulata Calvert), was synonomized with
Argia after it was determined that venational
characters are unreliable for species distinction
(Gloyd, 1968a). Kennedy (1920b) divided about
55 species of Argia into five subgenera on the
basis of the morphology of the penes; indeed,
omitting those taxa that do not fall into the
geographic range of our study, we recover many
of Kennedy’s groups in our analyses, both
molecular and combined: Cyanargia includes A.
lacrimans (Hagen) and A. tonto Calvert; Heliargia
is composed of A. vivida Hagen, A. plana Calvert,
(plus A. immunda, which we pull out of this clade);
Chalcargia, including A. translata, A. harknessi, A.
cuprea, A. oenea, A. ulmeca, A. occulata, (A.
garrisoni and A. sp. nov. were not available to
Kennedy). He also placed here A. tezpi and A.
sedula although our analyses do not support this.
Kennedy defined his species groups solely on
morphology of the intromittent penes, so it is
satisfying that we find similar patterns based on
molecular and morphological characters not used
by Kennedy.
Additionally, taxonomists have proposed close
relationships among (A. vivida, A. plana, A.
extranea), (A. fumipennis and A. pallens), (A.
westfalli and A. anceps), and (A. tonto and A.
lacrimans) (Gloyd, 1958; Garrison, 1994, 1996;
Westfall and May, 1996), which our analyses
support. A. plana was originally described as a
variety of A. vivida by Calvert (1901), and Gloyd
later elevated it to species status on the basis of
clasper morphology (1958). Our results validate
Gloyd, as we recover A. vivida as sister to A. plana
plus A. extranea. The close relationship between
the latter was suggested on the basis of coloration
and morphology by Garrison (1994).
In a thorough study of some Argia larvae,
Novelo-Gutierrez separated species into groups
on the basis of a single character: the degree of
convexity of the ligula (1992). He placed species
into three groups: those with very prominent
ligulae (including A. emma, harknessi, insipida,
moesta, oenea, tezpi, translata and ulmeca),
moderately prominent ligulae (A. munda, taras-
cana, and tonto), and slightly prominent ligulae
(A. fumipennis, lacrimans, nahuana, plana, pulla,
rhoadsi, and sedula). Our analysis suggests that
this character is not homologous (Fig. 4), or at
least is insufficient to accurately infer phylogenetic
relationship. Larvae of Argia species remain
poorly studied. Our limited inclusion of larval
characters indicates their potential value as
sources of phylogenetic characters, but further
study of larval morphology is needed. Of the 118
species of Argia currently recognized, larvae are
known from relatively few species.
Argia fumipennis is composed of the three
subspecies atra, fumipennis, and violacea. These
were described as separate species largely on the
basis of wing color; A. f. violacea has the typical
clear wing color, A. f. fumipennis can have smoky-
hyaline wing color in parts of its range, while A. f.
atra has dark brown wing pigmentation. These
species were unified on the basis of several
morphological characters (Gloyd, 1968b). Closely
related to A. fumipennis, on the basis of general
size, appearance, and clasper morphology, is A.
pallens (Westfall and May, 1996), a species
restricted in the U.S. to southeastern Arizona.
Our analyses do not include A. f. fumipennis, but
based on the molecular data we place A. pallens
within the A. fumipennis group, sister to A. f.
violacea (Figs. 2, 3). The morphological data pull
A. pallens out as sister to A. fumipennis (Fig. 4) in
a monophyletic clade. Both results are only
weakly supported, so further investigation of
106 ENTOMOLOGICA AMERICANA Vol. 115(2)

these relationships is warranted, as taxonomic
changes may be justified.
A somewhat surprising result is that we fail to
recover in our analyses a monophyletic assem-
blage of the ‘‘metallic’’ species- those that have the
cupreous coloration in the head and thorax (A.
cuprea, A. oenea, A. orichalcea). We only code one
morphological character related to the metallic
condition in this study. Further analysis of this
group is needed, and more detailed character
analysis may provide additional synapomorphy
for these species that have traditionally been
considered close relatives.
A plethora of additional characters, including
several morphological features unique to Argia,
are available for further study. For example,
males have a unique pair of pad-like structures
called ‘‘tori,’’ located posterodorsally on the tenth
abdominal segment between the cerci; the mor-
phology of these, and of the cerci and paraprocts,
are critical for species identification. These char-
acters are used in published taxonomic keys
(Garrison, 1994; Westfall and May, 1996; Forster,
2001), yet have not been thoroughly explored as
phylogenetic characters. Species of Argia also
differ dramatically in morphology of the penes
(Kennedy, 1920a), and while Kennedy used some
of these structures in his ‘‘phylogeny’’ of Zygop-
tera, his work predated by many years the formal
development of phylogenetic methodology. A
careful reexamination of these structures in Argia
is warranted. Additional mitochondrial genes, as
well as both ribosomal and protein-coding genes
such as 12S, 16S, cytochrome oxidases I and II,
are already known to be informative for odonate
phylogenetics (Chippendale et al., 1999; Misof et
al., 2000; Turgeon et al., 2005; Bybee et al., 2008).
In addition, it has been demonstrated that nuclear
genes such as Histone 3, Elongation Factor 1-
alpha, 18S and 28S rDNA show adequate
variation for species-level analyses in odonates
(Ware et al., 2007; Bybee et al., 2008; Carle et al.,
2008). We continue to code, refine, and expand
upon our data set as we continue our work on the
systematics and evolution of Argia.
ACKNOWLEDGMENTS
We thank several collectors who kindly donatedspecimens that were used in this study, listed inTable 1. Bill Mauffray of the IORI made thecollection available on several occasions and provid-ing and generously loaned specimens for this study.For discussion of various aspects of this study, we
thank Joe Gillespie, Dennis Paulson, Mark McPeek,Ola Fincke, and members of the Wenzel Lab at OSU.The suggestions of two anonymous reviewers im-proved an earlier version of our manuscript. We owespecial gratitude to Rosser Garrison for sharing hisimmense expertise on Neotropical Odonata with us.
LITERATURE CITED
Abbott, J. C. 2005. Dragonflies and Damselflies of Texas
and the South-Central United States. Princeton
University Press, Princeton, New Jersey, USA.
Borror, D. J. 1934. Ecological studies of Argia moesta
Hagen (Odonata: Coenagrionidae) by means of
marking. Ohio Journal of Science 34: 97–108.
Bick, G. H. and J. C. Bick. 1965. Color variation and
significance of color in reproduction in the
damselfly, Argia apicalis (Say) (Zygoptera: Coena-
griidae) (sic). Canadian Entomologist 97(1): 32–41.
Bick, G. H. and J. C. Bick. 1971. Localization, behavior,
and spacing of unpaired males of the damselfly
Argia plana Calvert (Odonata: Coenagrionidae).
Proceedings of the Entomological Society of
Washington 73: 146–152.
Bick, J. C. and G. H. Bick. 1982. Behavior of adults of
dark-winged and clear-winged subspecies of
Argia fumipennis (Burmeister) (Zygoptera: Coa-
nagrionidae). Odonatologica 11(2): 99–107.
Bybee, S. M., T. H. Ogden, M. A. Branham and M. F.
Whiting. 2008. Molecules, morphology and
fossils: a comprehensive approach to odonate
phylogeny and the evolution of the odonate wing.
Cladistics 24(4): 477–514.
Caesar, R. M., N. Gillette and A. I. Cognato. 2005.
Population Genetic Structure of an Edaphic
Beetle (Ptiliidae) Among Late Successional Re-
serves within the Klamath-Siskiyou Ecoregion,
California. Annals of the Entomological Society
of America 98(6): 931–940.
Calvert, P. P. 1901. Odonata, in Biologia Centrali
Americana: Insecta Neuroptera. R. H. Porter
and Dulau and Co., London, UK.
Carle, F. L., K. M. Kjer and M. L. May. 2008.
Evolution of Odonata, with special reference to
Coenagrionoidea (Zygoptera). Arthropod Sys-
tematics & Phylogeny 66(1): 37–44.
Chippendale, P. T., D. Varshal, D. H. Whitmore and J. V.
Robinson. 1999. Phylogenetic relationships of
North American damselflies of the genus Ischnura
(Odonata: Zygoptera: Coenagrionidae) based on
sequences of three mitochondrial genes. Molecular
Phylogenetics and Evolution 11: 110–121.
Conrad, K. F. 1992. Relationships of larval phenology
and imaginal size to male pairing success in Argia
vivida Hagen (Zygoptera: Coenagrionidae). Odo-
natologica 21(3): 335–342.
Conrad, K. F. and G. Pritchard. 1988. The mating
behavior of Argia vivida Hagen: as an example of
2009 ARGIA PHYLOGENY 107

female-control mating system (Zygoptera: Coe-
nagrionidae). Odonatologica 17: 179–185.
Conrad, K. F. and G. Pritchard. 1990. Pre-oviposition
mate guarding and mating behavior of Argia
vivda (Odonata: Coenagrionidae). Ecological
Entomology 15: 363–370.
Daigle, J. J. 1991. Argia garrisoni spec. nov. from
Mexico (Zygoptera: Coenagrionidae). Odonato-
logica 20(3): 337–342.
Daigle, J. J. 1995. Argia carlcooki spec. nov. from
Mexico (Zygoptera: Coenagrionidae). Odonato-
logica 24(4): 467–471.
Dunkle, S. W. 1990. Damselflies of Florida, Bermuda
and the Bahamas. Scientific Publishers,
Gainesville, Florida, USA.
Forster, S. 2001. The dragonflies of Central America
exclusive of Mexico and the West Indies. Gunnar
Rehfeldt, Wolfenbuttel, Germany.
Garrison, R. W. 1994. A synopsis of the genus Argia of the
United States with keys and descriptions of new
species, Argia sabino, A. leonorae, and A. pima
(Odonata: Coenagrionidae). Transactions of the
American Entomological Society 120(4): 287–368.
Garrison, R. W. 1996. A synopsis of the Argia fissa
group, with descriptions of two new species, A.
anceps sp. n. and A. westfalli sp. n. (Zygoptera:
Coenagrionidae). Odonatologica 25(1): 31–47.
Garrison, R. W. and N. von Ellenrieder. 2007. The true
Argia difficilis Selys, 1865, with the description of
Argia yungensis sp. nov. (Odonata: Coenagrioni-
dae). Transactions of the American Entomolog-
ical Society 133(1): 189–204.
Gloyd, L. K. 1958. The dragonfly fauna of the Big Bend
region of Trans-Pecos Texas. Occasional Papers
of the University of Michigan Museum of
Zoology 658.
Gloyd, L. K. 1968a. The union of Argia fumipennis
(Burmeister, 1839) with Argia violacea (Hagen,
1861), and the recognition of three subspecies
(Odonata). Occasional Papers of the University
of Michigan Museum of Zoology.
Gloyd, L. K. 1968b. The synonomy of Diargia and
Hyponeura with the genus Argia (Odonata:
Coenagrionidae: Argiinae). Michigan Entomolo-
gist 1(8): 271–274.
Huelsenbeck, J. P. 1997. Is the Felsenstein zone a fly
trap? Systematic Biology 46: 69–74.
Kennedy, C. H. 1918. New species of Odonata from the
southwestern United States. Part I. Three new
Argias. Canadian Entomologist 50(7): 256–261.
Kennedy, C. H. 1919. A new species of Argia (Odonata).
Canadian Entomologist 51: 17–18.
Kennedy, C. H. 1920a. The phylogeny of the zygopter-
ous dragonflies as based on the evidence of
the penes. Ohio Journal of Science 21: 19–32.
Kennedy, C. H. 1920b. Forty-two hitherto unrecognized
genera and subgenera of Zygoptera. The Ohio
Journal of Science 21: 83–88.
Misof, B., C. L. Anderson and H. Hadrys. 2000. A
Phylogeny of the Damselfly Genus Calopteryx
(Odonata) Using Mitochondrial 16S rDNA
Markers. Molecular Phylogenetics and Evolution
15(1): 5–14.
Moody, D. 2002. Coenagrionidae: Narrow-winged dam-
selflies. In Glotzhober, R. C. and D. McShaffrey
(eds.), The Dragonflies and Damselflies of Ohio.
Ohio Biological Survey, Columbus, Ohio, USA.
Nixon, K. C. 2000. Winclada version 0.99 (BETA).
Published by the author, Ithaca, New York
(available at www.cladistics.com).
Novelo-Gutierrez, R. 1992. Biosystematics of the larvae
of the genus Argia in Mexico (Zygoptera:
Coenagrionidae). Odonatologica 21: 39–71.
O’Grady, E. W. and M. L. May. December 2003. A
phylogenetic reassessment of the subfamilies of
Coenagrionidae (Odonata, Zygoptera). Journal
of Natural History 37: 2807–2834.
Rambur, M. P. 1842. Histoire Naturelle des Insectes
Neuropteres. Librarie Encyclopedique de Roirot,
Paris, France.
Robinson, J. V., J. E. Dickerson and D. R. Bible. 1983.
The demographics and habitat utilization of
adult Argia sedula (Hagen) as determined by
mark-recapture analysis (Zygoptera: Coenagrio-
nidae). Odonatologica 12(2): 167–172.
Simon, C., F. Frati, A. Beckenbach, B. Crespi, H. Liu and
P. Flook. 1994. Evolution, weighting, and phylo-
genetic utility of mitochondrial gene sequences and
a compilation of conserved polymerase chain
reaction primers. Annals of the Entomological
Society of America 87(6): 651–701.
Thompson, J. D., D. G. Higgins and T. J. Gibson. 1994.
CLUSTAL W: improving the sensitivity of
progressive multiple sequence alignment through
sequence weighting, positions-specific gap penal-
ties and weight matrix choice. Nucleic Acids
Research 22: 4673–4680.
Turgeon, J., R. Stoks, R. A. Thum, J. M. Brown and M.
A. McPeek. 2005. Simultaneous Quaternary
Radiations of Three Damselfly Clades across the
Holarctic. American Naturalist 165: E78–E107.
Ware, J. M., M. L. May and K. Kjer. 2007. Phylogeny
of the higher Libelluloidea (Anisoptera: Odo-
nata): An exploration of the most speciose
superfamily of dragonflies. Molecular Phyloge-
netics and Evolution 45: 289–310.
Westfall, M. J. and M. L. May. 1996. Damselflies of North
America. Scientific Publishers, Gainsville, FL.
Zwickl, D. J. 2006. Genetic algorithm approaches for the
phylogenetic analysis of large biological sequence
datasets under the maximum likelihood criterion.
Ph.D. dissertation, The University of Texas at
Austin.
Received 10 February 2009; accepted 22 November 2009
108 ENTOMOLOGICA AMERICANA Vol. 115(2)




















