
Integrated Pest Management for Brassica Production in East ...
Upload
independentCategory
view
0download
0
ORIGINAL PAPER
A passage through in vitro culture leads to efficientproduction of marker-free transgenic plants in Brassicajuncea using the Cre–loxP system
N. Arumugam Æ Vibha Gupta ÆArun Jagannath Æ Arundhati Mukhopadhyay ÆAkshay K. Pradhan Æ Pradeep Kumar Burma ÆDeepak Pental
Received: 27 April 2006 / Accepted: 20 November 2006 / Published online: 12 January 2007� Springer Science+Business Media B.V. 2006
Abstract The Cre–loxP site-specific recombina-
tion system was deployed for removal of marker
genes from Brassica juncea (Indian mustard).
Excision frequencies, monitored by removal of
nptII or gfp genes in F1 plants of crosses
between LOX and CRE lines, were high in
quiescent, differentiated somatic tissues but
extremely poor in the meristematic regions
(and consequently the germinal cells) thus pre-
venting identification and selection of marker-
free transgenic events which are devoid of both
the marker gene as well as the cre gene, in F2
progeny. We show that a passage through
in vitro culture of F1 leaf explants allows effi-
cient development of marker-free transgenics in
the F2 generation addressing current limitations
associated with efficient use of the Cre/loxP
technology for marker gene removal.
Keywords Marker-free plants � Brassica juncea �Cre–loxP � Transgenic crop
Introduction
The use of marker genes conferring resistance to
antibiotics or herbicides is essential for the
selection of transformed cells and tissues in
genetic transformation of plants. However, their
continued presence in transgenics, particularly
crop plants that are used for human and/or animal
consumption, has been a matter of concern for
nutritionists and environmentalists alike, with
member countries of the European Union re-
cently advocating a complete moratorium on the
use of transgenics containing superfluous marker
genes (Konig 2003). Retention of a marker gene
in a transgenic crop plant also prevents its
subsequent use for introducing additional genes
of agronomic value in a transgenic stock. Stacking
agronomically important genes by sexual crosses
using different transgenic stocks raised indepen-
dently using the same marker gene/promoters is
also not a viable proposition as it may lead to
homology-based silencing of the marker gene
N. Arumugam and Vibha Gupta have contributed equallyto this work.
N. Arumugam � V. Gupta � A. Jagannath �A. Mukhopadhyay � A. K. Pradhan � D. Pental (&)Centre for Genetic Manipulation of Crop Plants,University of Delhi South Campus, Benito JuarezRoad, New Delhi 110 021, Indiae-mail: [email protected]
Present Address:N. ArumugamDepartment of Biotechnology, PondicherryUniversity, Pondicherry 605 014, India
A. K. Pradhan � P. K. Burma � D. PentalDepartment of Genetics, University of Delhi SouthCampus, Benito Juarez Road,New Delhi 110 021, India
123
Transgenic Res (2007) 16:703–712
DOI 10.1007/s11248-006-9058-7

and/or the gene of interest cloned in cis (Vauc-
heret 1993; Kumpatla et al. 1998).
With the recent progress in plant genomics, the
development of strategies for marker gene
removal has become very relevant to plant
scientists (Hare and Chua 2002; Halpin 2005).
Several systems such as co-transformation (Math-
ews et al. 2001; Park et al. 2004; Chen et al. 2005),
transposons (Goldsbrough et al. 1993), homolo-
gous recombination (Iamtham and Day 2000),
site-specific recombination (Ow 2002; Lyznik
et al. 2003) and use of P-DNA (Rommens et al.
2004) have been developed for the removal of
marker genes from transgenic plants. Among the
various strategies, site-specific recombination sys-
tems have been tested extensively with the
bacteriophage P1 Cre–loxP system being one of
the most studied systems till date. The Cre–loxP
system consists of two components: (a) two loxP
sites each consisting of 34 bp-inverted repeats
cloned in direct orientation flanking a DNA
sequence and (b) the cre gene encoding a 38-
kDa recombinase protein that specifically binds
to the loxP sites and excises the intervening
sequence along with one of the loxP sites.
Although the Cre–loxP system has been tested
in several plants including Arabidopsis (Zuo et al.
2001), Nicotiana (Odell et al. 1990; Dale and Ow
1991; Gleave et al. 1999), Zea mays (Zhang et al.
2003) and Oryza sativa (Hoa et al. 2002; Sreekala
et al. 2005), none of these studies have reported
successful development of marker-free transgen-
ics, which are devoid of the marker as well as the
cre genes but contain the gene of interest (GI), in
the F2 progeny.
In this study, we tested the efficiency of the
Cre–loxP system for marker gene removal in
Brassica juncea (Indian mustard), a major oilseed
crop of the Indian subcontinent, which requires
introduction of many desirable agronomic traits
using transgenic technology to enhance its yield
potential (Grover and Pental 2003). Independent
transgenic lines homozygous for the marker gene
(NPT-LOX plants with the kanamycin resistance-
conferring nptII gene between the loxP sites) and
the recombinase source (CRE plants with the cre
gene driven by the CaMV35S promoter) were
developed in B. juncea. PCR analysis of the
marker gene status in NPT-LOX · CRE F1
hybrids revealed efficient excision of the marker
gene in leaf tissues. However, no marker-free
transgenics could be obtained in the F2 progeny of
such marker-free F1 plants. Studies on the
dynamics of Cre-mediated excision using the gfp
gene (GFP-LOX plants containing the gfp gene
within the loxP sites) showed efficient excision in
the quiescent somatic cells of cotyledons, leaves
and roots of GFP-LOX · CRE F1 hybrids. Exci-
sion frequencies were, however, very poor in
actively dividing meristematic tissues including
the shoot apical meristems, which are the pro-
genitors of germinal cells. We tested the possibil-
ity of obtaining marker-free individuals by
in vitro regeneration of plants from marker-free,
somatic (leaf) tissues of F1 events. We report that
these regenerants (designated ‘‘F1LR’’ events)
showed unambiguous segregation of marker-free
individuals in the F2 progeny thereby enabling
successful deployment of the Cre–loxP system in
the crop.
Materials and methods
Development of transformation vectors
pCGMCP-npt(lox)
A new binary vector (pCGMCP22; Fig. 1a) was
constructed in this study by modification of the
binary vector pPZP200 (Hajdukiewicz et al.
1994). The BclI-ScaI fragment of pPZP200 was
replaced with a 1.4-kb synthetic T-DNA se-
quence comprising the 25 bp consensus left and
right border repeats of T-DNA, a number of
unique restriction sites, loxP recombination sites
in direct orientation and a consensus overdrive
sequence from an octopine Ti plasmid outside
the right border. The synthetic DNA sequence
was designed to maintain an overall GC-content
of 42% in consonance with the GC-content of a
large number of dicot genomes (Salinas et al.
1988). Methylation-prone sequences (CGs and
CNGs) were also removed to the possible extent.
Keeping in view the polar transfer of T-DNA
from right to left border, the loxP sites for the
removal of marker genes were placed towards
the left border. The 1.4-kb sequence was
704 Transgenic Res (2007) 16:703–712
123

assembled using five sub-fragments (300–350 bp
in length) with unique restriction sites at their
ends for directional assembly. These sub-frag-
ments were constructed by recursive PCR (Dil-
lon and Rosen 1990) using six overlapping
oligonucleotides each of which was 60–80 nucle-
otides in length. The 1.4-kb synthetic sequence
was cloned as a BclI-ScaI fragment at the
corresponding sites of pPZP200 to generate the
plasmid pCGMCP22 (Fig. 1a).
The Pnos-nptII-ocspA fragment (used as the
selectable marker) was cloned at the EcoRV site
between the loxP sites of pCGMCP22. The
CaMV35S-gus-35SpA cassette was cloned in the
Ecl136II site (outside the loxP site) and used as a
reporter as well as the passenger gene of interest.
This construct, termed pCGMCP-npt(lox), is
schematically represented in Fig. 1b.
pPZP200-cre (Fig. 1c)
The cre gene was PCR amplified from bacterio-
phage P1 and mobilized between the CaMV35S
promoter and the 35S polyA signal. The
CaMV35S-cre-35SpA cassette was cloned at the
HindIII site of pPZP200. The nos-nptII-ocspA
BamHI(6182) SphI(7645)
T-DNA Region
LB RBlox lox PacI(7058)PflMI(6803)
PmeI(7310)
ScaI(4)
SpeI(7041)SspI(7029)
XbaI(7291)
AvrII(6721)
BclI(6161)
Ecl136II(7279)
EcoRV(6708)HindIII(6689)
SfiI(6443)
SfiI(7462)
AflII(6562) AflII(7400)BglII(6574)
BstXI(6466) BstXI(7482)
MscI(6552)
MscI(7390)
MspI(6540)
MspI(7412)
T-DNA Region
SwaI(7363)XmnI(6590) BglII(7376)
a
d
LB
bar probegfp probe
HindIIISpeI
HindIII
EcoRVEcoRV
RB
EcoRV
Amv loxP loxP
P35S gfp 35SpA P35S bar 35SpA
c
cre probe
RB
HindIII
LB
EcoRI
P4
P5
HindIII
EcoRV EcoRV
BamHI
EcoRI
XbaINcoI NcoI
Pnos nptII OcspA P35S cre 35SpA
XbaI
RB
gus probenptII probe P3
b
LB
EcoRVHindIII
P2
NcoINcoI
P1
EcoRVXbaI
XhoII
EcoRV
loxPloxP
P35S gus 35SpAPnos nptII OcspA
Fig. 1 Constructs usedfor testing the Cre–loxPsystem for marker generemoval in B. juncea. (a)pCGMCP22, (b)pCGMCP-npt(lox), (c)pPZP200-cre and (d)pCGMCP-gfp(lox).Unique restriction sitesare shown in bold inFig. 1(a). LB and RB:Left and Right borders,respectively of the T-DNA, P1 to P5 are PCRprimers specific foramplification of pre-excision (P2–P3), post-excision (P1–P3) and cre(P4–P5) gene products
Transgenic Res (2007) 16:703–712 705
123

cassette was cloned at the EcoRI site of pPZP200
and used as the in vitro selection marker.
pCGMCP-gfp(lox) (Fig. 1d)
The CaMV35S-gfp-35SpA cassette was cloned at
the HindIII site between the two loxP sites of
pCGMCP22 and used for monitoring Cre-medi-
ated excision in plant tissues. The CaMV35S-bar-
35SpA cassette (conferring phosphinothricin
resistance) was cloned as a blunt-ended fragment
at the Ecl136II site (outside the loxP site) towards
the right border and used as an in vitro selection
marker.
Transformation of Brassica juncea
The pCGMCP-npt(lox), pCGMCP-gfp(lox) and
pPZP200-cre constructs were mobilized indepen-
dently into the Agrobacterium strain GV3101 by
electroporation (Mattanovich et al. 1989). B.
juncea was transformed using hypocotyl explants
following Mehra et al (2000). Regenerated shoots
were transferred for rooting on a selection
medium containing 50 mg/l kanamycin (for
pCGMCP-npt(lox) and pPZP200-cre constructs)
or 20 mg/l phosphinothricin (for pCGMCP-
gfp(lox) construct). Only one T0 transformant
per explant was selected. Transgenics were trans-
ferred to soil and grown under containment in
accordance with guidelines of the Department of
Biotechnology, Government of India.
Development of F1LR events: shoot
regeneration from leaf explants of NPT-
LOX · CRE F1 progeny
Five mm2 segments of healthy leaves from 15-
day-old F1 shoot cultures were placed on a
regeneration medium consisting of MS salts
supplemented with 1 mg/l NAA, 1 mg/l BAP
and 20 lM AgNO3. Culture conditions for
obtaining regenerants were as described by Me-
hra et al (2000). The regenerated shoots (F1LRs)
were maintained on MS medium with 2 mg/l IBA,
transferred to soil under containment and self-
pollinated to raise F2 progeny.
Molecular analysis: Southern hybridization
and tissue PCR
Single copy transgenics were identified by South-
ern hybridization. Total DNA was isolated from
leaf tissues of putative T0 transgenics by the
CTAB method (Rogers and Bendich 1994). For
NPT-LOX plants, 5 lg of total DNA was inde-
pendently restricted with HindIII and XbaI,
electrophoresed and blotted on to nylon mem-
branes following standard procedures (Sambrook
et al. 1989). The HindIII and XbaI blots were
hybridized with gus and npt gene probes (Fig. 1b),
respectively. For CRE plants, blots were prepared
after digestion of genomic DNA with EcoRI and
BamHI and hybridized with the cre and npt gene
probes (Fig. 1c), respectively. The copy number
of T-DNA in GFP-LOX plants was determined
by digesting the total DNA with SpeI and
hybridization with bar and gfp gene probes
(Fig. 1d). Plants showing single copy integrations
on both flanks of the T-DNA were taken further
for subsequent studies.
For confirming removal of the marker gene,
genomic DNA isolated from the leaf tissues of
relevant F1, F2 and homozygous parent transgenic
lines was digested with HindIII and the blots were
hybridized with nptII or gus gene probes (Fig. 1b)
for confirming the presence or absence of the
respective genes.
PCR primers were designed for specific ampli-
fication of pre-excision (presence of the marker
gene), post-excision (presence of empty donor
site after removal of the marker gene from NPT-
LOX plants) and cre gene products (Fig. 1b, c).
The primer sequences and the expected sizes of
PCR products are given in Table 1. Template
DNA for tissue PCR was prepared from leaf
tissues and/or shoot tips of in vitro grown trans-
genic plants and relevant controls following Kli-
myuk et al (1993). PCR conditions for
amplification of pre-excision products were: ini-
tial denaturation at 94oC for 5 min followed by 40
cycles each of 94oC, 1 min, 55oC, 1 min and 72oC,
1 min. A final extension at 72oC for 5 min was
given. PCR conditions for amplification of post-
excision and cre gene products were: initial
denaturation at 94oC for 5 min followed by 40
706 Transgenic Res (2007) 16:703–712
123

cycles each of 94oC, 1 min, 54oC, 1 min and 72oC,
1 min. A final extension at 72oC for 5 min was
given.
Confocal microscopy
F1 seeds of the GFP-LOX · CRE cross were
germinated on MS medium and seedlings were
transferred to a medium consisting of MS salts
supplemented with 2 mg/l IBA, 20 mg/l phosph-
inothricin and 50 mg/l kanamycin. Seedlings
which rooted on the above double selection
medium were grown further for 15 days in a liquid
medium of the same composition but without any
selection pressure. The parental lines were also
grown on respective selection media and subse-
quently on the liquid medium. Shoot tips were
dissected under an Olympus Stereo Zoom micro-
scope and mounted on slides in PBS buffer (80 g/l
NaCl, 2 g/l KCl, 6.09 g/l Na2HPO4, 2 g/l KH2PO4
and pH 7.4) as whole mounts. Five to ten
millimeter long root segments along with the root
tips were also whole mounted in PBS buffer. GFP
imaging was done using a LSM5 Pascal Confocal
Laser Scanning Microscope set at 488 nm excita-
tion and 510 nm emission. Photographs were
taken using Image pro-plus 4.0 software.
Results
Transformation of B. juncea and development
of homozygous LOX and CRE lines
Transformants were generated in B. juncea using
the pCGMCP-npt(lox) and pPZP200-cre con-
structs. Ten NPT-LOX and eight CRE transgen-
ics with single copy integrations of the T-DNA
were identified by Southern analysis of 64 NPT-
LOX and 40 CRE plants, respectively using the
restriction enzymes and probes described in
Materials and methods (data not shown). Single
copy transgenics were self-pollinated to obtain T1
seeds. T1 seedlings of most of the selected T0
plants segregated in the expected 3:1 ratio for
kanamycin resistance:sensitivity. T1 kanamycin-
resistant plants were transferred to soil, self-
pollinated and the T2 progeny of those plants that
did not segregate for kanamycin resistance and
susceptibility were identified as homozygous
NPT-LOX and CRE lines (data not shown).
Analysis of marker gene (nptII) excision in F1
and F2 progeny of NPT-LOX · CRE crosses
Seven homozygous CRE lines were crossed with
seven homozygous NPT-LOX lines in 49 different
combinations. Screening a representative popula-
tion of the F1 progeny of each of the above
crosses by Tissue-PCR of leaf tissue using primers
specific for pre-excision, post-excision and cre
gene products (Table 1), indicated excision of the
marker gene in ~53% of F1 plants which showed
no pre-excision product but tested positive for
post-excision and cre gene products. To obtain
marker-free transgenics (which are also devoid of
the cre gene), F2 plants were generated from two
selected F1 lines (NPT-LOX1 · CRE2 showing
maximum and NPT-LOX1 · CRE4 showing least
expression levels of the cre protein). These two F1
lines lacked the marker gene within the loxP sites
as revealed by PCR analysis of leaf tissues.
However, PCR analysis did not show any
marker-free plants among the analyzed F2 prog-
eny (Table 2). Amplification of post-excision
PCR products (indicating excision of the marker
gene) was always observed to be associated with
the presence of the cre gene. We also observed
Table 1 Primers used for PCR analyses and expected size of PCR products
Product Primer Primer code Sequence Expected sizeof PCR product
Pre-excision Forward P2 5¢-GCTAGCTGATAGTGACCTTAGGCGACTT-3¢ 745Reverse P3 5¢-GCTACTTGGAGGTTCAACATTGTAGGACA-3¢
Post-excision Forward P1 5¢-ATGCATTCATGATGGTCACTCTCTCA-3¢ 292Reverse P3 5¢-GCTACTTGGAGGTTCAACATTGTAGGACA-3¢
cre gene Forward P4 5¢-CAGCAACATTTGGGCCAGCTAAAC-3¢ 368Reverse P5 5¢-TCTCTACACCTGCGGTGCTAACCAG-3¢
Transgenic Res (2007) 16:703–712 707
123

two new classes of plants: chimeric plants (con-
taining pre-excision, post-excision and cre PCR
products) and plants containing only the pre-
excision product without the cre gene (Table 2).
PCR analysis of excision profiles in apical meri-
stematic tissues (which are the progenitors of
germinal cells) of the F1 plants showed the
presence of both pre- and post-excision products
(Fig. 2) indicating poor efficiency of marker gene
excision in the same. We also analyzed by tissue
PCR, shoot tips of 48 randomly selected F2 plants
showing post-excision and cre products in the leaf
tissues. All these events were also found to
contain pre-excision product in their shoot tips
(data not shown).
Analysis of cre-mediated excision pattern in
plants using a reporter gene (gfp)
To monitor the excision pattern in plant tissues
more accurately, we generated transgenics in B.
juncea using a construct containing the green
fluorescent protein-encoding gfp gene between
the loxP sites (pCGMCP-gfp(lox); Fig. 1d). Sin-
gle copy GFP-LOX transgenics were identified
and crossed with the CRE lines used earlier.
Analysis of the fluorescence pattern of GFP in the
F1 progeny of the above crosses showed strong
fluorescence in the shoot as well as in the root
apical meristems (indicating non-excision of the
gfp gene) while no fluorescence could be detected
in any of the other somatic tissues adjacent to the
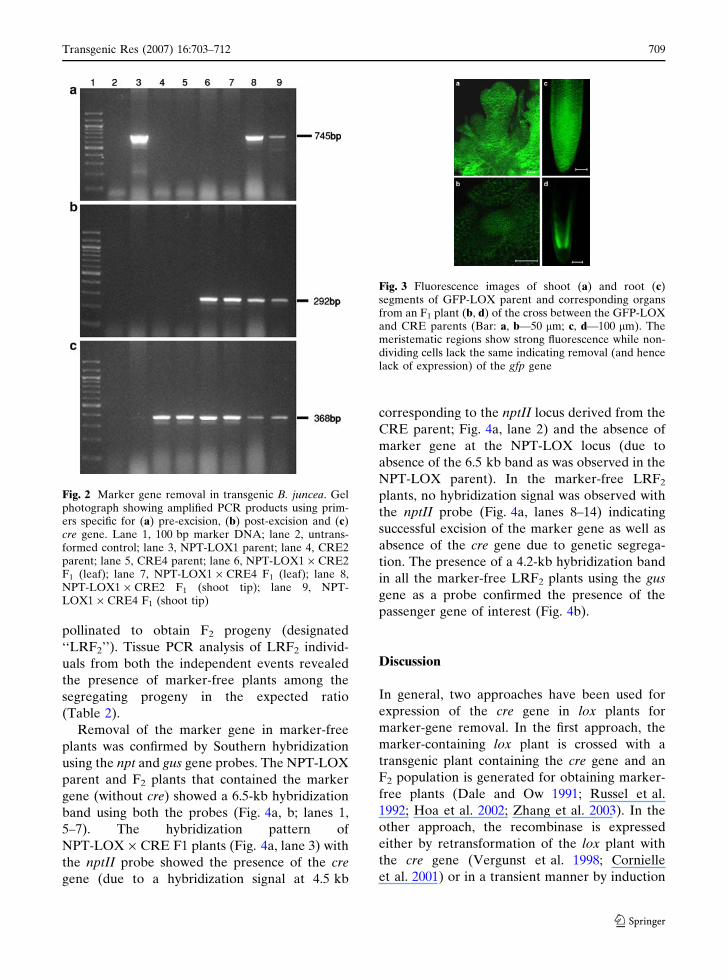
meristematic regions (Fig. 3).
In vitro shoot regeneration from leaf explants
of NPT-LOX · CRE F1 plants and generation
of marker-free transgenics
To obtain marker-free individuals, we regener-
ated plants from the leaf (somatic) tissues of the
same NPT-LOX1 · CRE2 and NPT-
LOX1 · CRE4 F1 plants that were used earlier
for development of F2 progeny (see above).
Tissue PCR analysis of the F1 regenerants thus
obtained (designated ‘‘F1LR’’) showed the pres-
ence of only post-excision and cre products
indicating that these shoots have regenerated
from marker-free cells of the F1 plant. Such
regenerants were transferred to the field and self-Ta
ble
2S
eg
reg
ati
on
of
po
st-e
xci
sio
n,
pre
-ex
cisi
on
,cr
ea
nd
nu
llP
CR
ev
en
tsin
F2
pro
ge
ny
de
riv
ed
by
selfi
ng
F1
pla
nts
an
db
yse
lfin
gle
af
reg
en
era
nts
(F1L
R)
of
F1
pla
nts
Ev
en
tN
o.
of
pla
nts
an
aly
zed
Nu
mb
er
of
pla
nts
sho
win
gP
CR
am
pli
fica
tio
nfo
r
Po
st-e
xci
sio
na
nd
crea
Po
st-e
xci
sio
nw
ith
ou
tcr
eacr
eaN
ull
aP
re-e
xci
sio
n,
po
st-e
xci
sio
na
nd
cre
Pre
-ex
cisi
on
wit
ho
ut
cre
F1
(NP
T-L
OX
1·
CR
E2
)fi
F2
20
47
4(1
15
)0
(38
)3
7(3
8)
13
(13
)3
2(0
)4
8(0
)F
1(N
PT
-LO
X1
·C
RE
4)
fiF
22
24
86
(12
6)
0(4
2)
52
(42
)1
9(1
4)
28
(0)
39
(0)
F1
(NP
T-L
OX
1·
CR
E2
)fi
LR
fiF
2b5
43
3(3
0)
8(1
0)
9(1
0)
4(3
)0
(0)
0(0
)F
1(N
PT
-LO
X1
·C
RE
4)
fiL
Rfi
F2b
13
18
0(7
4)
19
(25
)2
2(2
5)
10
(8)
0(0
)0
(0)
Da
taw
as
ob
tain
ed
by
tiss
ue
PC
Ru
sin
ge
xp
an
de
dle
av
es
of
inv
itro
gro
wn
F2
pla
nts
aT
he
sea
reth
ee
xp
ect
ed
cla
sse
sa
cco
rdin
gto
the
ind
ep
en
de
nt
ass
ort
me
nt
of
two
loci
;n
um
be
rsin
pa
ren
the
ses
ind
ica
teth
ee
xp
ect
ed
fre
qu
en
cie
sb
Fit
tora
tio
9:3
:3:1
708 Transgenic Res (2007) 16:703–712
123
pollinated to obtain F2 progeny (designated
‘‘LRF2’’). Tissue PCR analysis of LRF2 individ-
uals from both the independent events revealed
the presence of marker-free plants among the
segregating progeny in the expected ratio
(Table 2).
Removal of the marker gene in marker-free
plants was confirmed by Southern hybridization
using the npt and gus gene probes. The NPT-LOX
parent and F2 plants that contained the marker
gene (without cre) showed a 6.5-kb hybridization
band using both the probes (Fig. 4a, b; lanes 1,
5–7). The hybridization pattern of
NPT-LOX · CRE F1 plants (Fig. 4a, lane 3) with
the nptII probe showed the presence of the cre
gene (due to a hybridization signal at 4.5 kb
corresponding to the nptII locus derived from the
CRE parent; Fig. 4a, lane 2) and the absence of
marker gene at the NPT-LOX locus (due to
absence of the 6.5 kb band as was observed in the
NPT-LOX parent). In the marker-free LRF2
plants, no hybridization signal was observed with
the nptII probe (Fig. 4a, lanes 8–14) indicating
successful excision of the marker gene as well as
absence of the cre gene due to genetic segrega-
tion. The presence of a 4.2-kb hybridization band
in all the marker-free LRF2 plants using the gus
gene as a probe confirmed the presence of the
passenger gene of interest (Fig. 4b).
Discussion
In general, two approaches have been used for
expression of the cre gene in lox plants for
marker-gene removal. In the first approach, the
marker-containing lox plant is crossed with a
transgenic plant containing the cre gene and an
F2 population is generated for obtaining marker-
free plants (Dale and Ow 1991; Russel et al.
1992; Hoa et al. 2002; Zhang et al. 2003). In the
other approach, the recombinase is expressed
either by retransformation of the lox plant with
the cre gene (Vergunst et al. 1998; Cornielle
et al. 2001) or in a transient manner by induction
Fig. 2 Marker gene removal in transgenic B. juncea. Gelphotograph showing amplified PCR products using prim-ers specific for (a) pre-excision, (b) post-excision and (c)cre gene. Lane 1, 100 bp marker DNA; lane 2, untrans-formed control; lane 3, NPT-LOX1 parent; lane 4, CRE2parent; lane 5, CRE4 parent; lane 6, NPT-LOX1 · CRE2F1 (leaf); lane 7, NPT-LOX1 · CRE4 F1 (leaf); lane 8,NPT-LOX1 · CRE2 F1 (shoot tip); lane 9, NPT-LOX1 · CRE4 F1 (shoot tip)
Fig. 3 Fluorescence images of shoot (a) and root (c)segments of GFP-LOX parent and corresponding organsfrom an F1 plant (b, d) of the cross between the GFP-LOXand CRE parents (Bar: a, b—50 lm; c, d—100 lm). Themeristematic regions show strong fluorescence while non-dividing cells lack the same indicating removal (and hencelack of expression) of the gfp gene
Transgenic Res (2007) 16:703–712 709
123

using physical and chemical agents (Zuo et al.
2001; Zhang et al. 2003; Yuan et al. 2004; Sree-
kala et al. 2005). In cases wherein the recom-
binase was expressed by external induction, the
cre gene was cloned as a part of the construct
containing the marker gene. Although previous
studies have shown efficient excision of the
marker gene in F1 plants (Hoa et al. 2002;
Zhang et al. 2003), none of them provide data
on the production of marker-free transgenics
(which lack the marker as well as the cre genes
but contain the passenger gene of interest) in the
F2 generation.
In our studies on deployment of the Cre–loxP
system in B. juncea, we found that although
excisions occurred with very high efficiency in
quiescent, differentiated somatic tissues of F1
plants, no marker-free transgenics could be
obtained in the F2 progeny. We hypothesized
that this could be due to lack of excisions in the
apical meristems, which are the progenitors of
germinal cells. Experiments using gfp as a
reporter gene to track Cre-mediated excision
and PCR analysis of meristematic tissues of F1
plants conclusively established that excision fre-
quencies are poor in meristematic tissues. Similar
observation in F2 plants (containing pre-excision
product in meristematic tissues and post-excision
and cre products in leaf tissues) also indicates that
it may not be possible to obtain marker-free
transgenics in the F3 generation also. To address
the above limitation, we regenerated plants from
marker-free, quiescent, differentiated somatic
cells of F1 progeny (F1LR) and successfully
generated marker-free segregants among the F2
progeny of such regenerants.
Recent studies on marker gene removal using
the Cre–loxP system in Arabidopsis (Zuo et al.
2001), maize (Zhang et al. 2003) and tobacco
(Wang et al. 2005) describe the use of constructs
containing both the cre and the marker genes
within the same loxP sites. In these studies,
simultaneous removal of cre and the marker
genes was effected by transient, chemical or
heat-inducible expression of the recombinase
under in vitro conditions. Based on our observa-
tions, it is evident that the success of this
approach is not due to the use of an inducible
system per se but because of regeneration of
plants through in vitro culture following transient
induction of the recombinase in transgenic ex-
plants. However, such inducible systems lack
stringent regulatory controls, which lead to de-
regulated auto-excision of the marker gene and
consequently, a drastic reduction in transforma-
tion frequencies (Wang et al. 2005). Using such
Fig. 4 Hybridization profile of HindIII-digested totalDNA using (a) nptII and (b) gus gene probes. Lane 1,NPT-LOX1 parent; lane 2, CRE4 parent; lane 3, (NPT-LOX1 · CRE4) F1 plant without the marker gene; lane 4,untransformed control; lanes 5–7, marker-containing F2
plants obtained by direct self-pollination of F1 plants; andlanes 8–14, marker-free plants obtained by self-pollinationof F1LR plants (obtained by in vitro regeneration from
leaf explants of F1 plants). The absence of hybridizationwith nptII probe in lanes 8–14 indicates that these plantslack both the cre gene and the nptII marker. The presenceof a low molecular weight (4.2 kb) hybridization signal inthese plants as compared to that in the NPT-LOX1 parent(6.5 kb) on hybridization with the gus gene furtherconfirmed the marker-free status of these transgenic plants
710 Transgenic Res (2007) 16:703–712
123

an approach, generation of transformed chimeras
also cannot be ruled out.
The introduction of the recombinase through
pollen transfer (sexual crosses) is preferable over
inducible expression in tissue culture as it avoids
repeated use of the cre gene in subsequent
transformation experiments. Development of a
single cre stock for a crop will suffice. Our results
clearly suggest that unambiguous segregation of
the marker-free trait and cre gene in F2 plants can
be obtained only by in vitro regeneration of
plants from the quiescent, somatic (leaf) tissues
of F1 (NPT-LOX · CRE) plants followed by self-
pollination of the regenerants. If our observations
on B. juncea hold true in general, removal of
marker genes from transgenic stocks could
become a routine affair addressing all the current
bottlenecks in the development of marker-free
transgenics.
Acknowledgements Financial support for this researchwas provided by DOFCO, a subsidiary of the NationalDairy Development Board (NDDB). B. S. Yadavprovided technical support. The gfp gene was a kind giftfrom Prof Jen Sheen. We are grateful to the Departmentof Botany, University of Delhi for extending use of theconfocal microscope facility.
References
Chen S, Li X, Liu X, Xu H, Meng K, Xiao G, Wei X, WangF, Zhu Z (2005) Green fluorescent protein as a vitalelimination marker to easily screen marker-freetransgenic progeny derived from plants co-trans-formed with double T-DNA binary vector system.Plant Cell Rep 23:625–631
Cornielle S, Lutz K, Svab Z, Maliga P (2001) Efficientelimination of selectable marker genes from theplastid genome by CRE-lox site-specific recombina-tion system. Plant J 27:171–178
Dale EC, Ow DW (1991) Gene transfer with subsequentremoval of the selection gene from the host genome.Proc Natl Acad Sci USA 88:10558–10562
Dillon PJ, Rosen CA (1990) A rapid method for theconstruction of synthetic genes using the polymerasechain reaction. Biotechniques 9:298–299
Gleave AP, Mitra DS, Mudge SR, Morris BAM (1999)Selectable marker-free transgenic plants without sex-ual crossing: transient expression of cre recombinaseand use of a conditional lethal dominant gene. PlantMol Biol 40:223–235
Goldsbrough AP, Lastrella CN, Yoder JI (1993) Transpo-sition mediated re-positioning and subsequent elimi-
nation of marker genes from transgenic tomato. Bio/technology 11:1286–1292
Grover A, Pental D (2003) Breeding objectives andrequirements for producing transgenics for majorfield crops of India. Curr Sci 84:310–320
Hajdukiewicz P, Svab Z, Maliga P (1994) The small,versatile pPZP family of Agrobacterium binary vec-tors for plant transformation. Plant Mol Biol 25:939–994
Halpin C (2005) Gene stacking in transgenic plants – thechallenge for 21st century plant biotechnology. PlantBiotech J 3:141–155
Hare PD, Chua N-H (2002) Excision of selectable markergenes from transgenic plants. Nat Biotechnol 20:575–580
Hoa TT, Bong BB, Huq E, Hodges TK (2002) Cre-lox site-specific recombination controls the excision of atransgene from the rice genome. Theor Appl Genet104:518–525
Iamtham S, Day A (2000) Removal of antibiotic resistancegenes from transgenic tobacco plastids. Nat Biotech-nol 18:1172–1176
Klimyuk VI, Carroll BJ, Thomas CM, Jones JDG (1993)Alkali treatment for rapid preparation of plant tissuefor reliable PCR analysis. Plant J 3:493–494
Konig A (2003) A framework for designing transgeniccrops—science, safety and citizen’s concerns. NatBiotechnol 21:1274–1279
Kumpatla SP, Chandrasekharan MB, Iyer LM, Li G, HallTC (1998) Genome intruder scanning and modulationsystems and transgene silencing. Trends Plant Sci3:97–104
Lyznik LA, Gordon-Kamm WJ, Tao Y (2003) Site-specificrecombination for genetic engineering in plants. PlantCell Rep 21:925–932
Mattanovich D, Riiker F, Machado A, Laimer M, RegnerF, Steinkellner H, Himmler G, Katinger H (1989)Efficient transformation of Agrobacterium spp byelectroporation. Nucleic Acids Res 17:6747
Matthews PR, Wang MB, Waterhouse PM, Thornton S,Fieg SJ, Gubler F, Jacobson JV (2001) Marker geneelimination from transgenic barley, using co-transfor-mation with adjacent ‘twin T-DNA’ on a standardAgrobacterium transformation vector. Mol Breed7:195–202
Mehra S, Pareek A, Bandyopadhyay P, Sharma P, BurmaPK, Pental D (2000) Development of transgenics inIndian oilseed mustard (Brassica juncea) resistant toherbicide phosphinotricin. Curr Sci 78:1358–1364
Odell J, Caimi P, Sauer B, Russell S (1990) Site-directedrecombination in the genome of transgenic tobacco.Mol Gen Genet 223:369–378
Ow DW (2002) Recombinase-directed plant transforma-tion for the post-genomic era. Plant Mol Biol 48:183–200
Park J, Lee YK, Kang BK, Chung WH (2004) Co-transformation using a negative selectable markergene for the production of selectable marker gene-free transgenic plants. Theor Appl Genet 109:1562–1567
Transgenic Res (2007) 16:703–712 711
123

Rogers SO, Bendich AJ (1994) Extraction of total cellularDNA from plants, algae and fungi. In: Gelvin SB,Schilperoort RA (eds) Plant molecular biologymanual, 2nd edn, vol D1. Kluwer Academic Publish-ers, Dordrecht, The Netherlands, 1–8
Rommens CM, Humara JM, Ye J, Yan H, Richael C,Zhang L, Perry R, Swords K (2004) Crop improve-ment through modification of plant’s own genome.Plant Physiol 135:421–431
Russel SH, Hoopes JL, Odell JT (1992) Directed excisionof a transgene from the plant genome. Mol GenGenet 243:49–59
Salinas J, Matassi G, Montero LM, Bernardi G (1988)Compositional compartmentalization and composi-tional patterns in nuclear genomes of plant. NucleicAcids Res 16:4269–4285
Sambrook J, Fritsch EF, Maniatis T (1989) Molecularcloning: a laboratory manual. Cold Spring HarborLaboratory Press, New York
Sreekala C, Wu L, Gu K, Wang D, Tian D, Yin Z (2005)Excision of selectable marker in transgenic rice(Oryza sativa L.) using a chemically regulated Cre/loxP system. Plant Cell Rep 24:86–94
Vaucheret H (1993) Identification of a general silencer for19S and 35S promoters in a transgenic tobacco plants:90 bp of homology in the promoter sequence aresufficient for trans-inactivation. CR Acad Sci316:1471–1483
Vergunst AC, Jansen LET, Hooykaas PJJ (1998) Site-specific integration of Agrobacterium T-DNA inArabidopsis thaliana mediated by Cre recombinase.Nucleic Acids Res 26:2729–2734
Wang Y, Chen B, Hu Y, Li J, Lin Z (2005) Inducibleexcision of selectable marker gene from transgenicplants by Cre/lox site-specific recombination system.Transgenic Res 14:605–614
Yuan Y, Yun-Jun L, Tao W (2004) A new Cre/lox systemfor deletion of selectable marker gene. Acta Bot Sin46:862–866
Zhang W, Subbarao S, Addae P, Shen A, Armstrong C,Peschke V, Gilbertson L (2003) Cre/lox-mediatedmarker gene excision in transgenic maize (Zea maysL.) plants. Theor Appl Genet 107:1157–1168
Zuo J, Niu Q-W, Moller SG, Chua N-H (2001) Chemical-regulated, site specific DNA excision in transgenicplants. Nat Biotechnol 19:157–161
712 Transgenic Res (2007) 16:703–712
123
Copyright © 2022 FDOKUMEN




















