
A novel IRQ ligand-modified nano-carrier targeted to a unique pathway of caveolar endocytic pathway
10
A novel IRQ ligand-modified nano-carrier targeted to a unique pathway of caveolar endocytic pathway Diky Mudhakir, Hidetaka Akita, Erdal Tan, Hideyoshi Harashima ⁎ Laboratory for Molecular Design of Pharmaceutics, Faculty of Pharmaceutical Sciences, Hokkaido University, Kita-12, Nishi-6, Kita-ku, Sapporo 060-0812, Japan Received 26 June 2007; accepted 20 October 2007 Abstract In the present study, the cellular uptake and subsequent intracellular trafficking of liposomes was investigated, in which a novel peptide (IRQ), identified with in vivo phage display, was modified on the surface. Since the novel peptide IRQRRRR is rich in arginine, the cellular uptake mechanism was compared with octaarginine (R8)-modified liposomes, which are known to be taken up by cells via macropinocytosis. The uptake mechanism and intracellular trafficking of peptide-modified liposomes was determined by confocal laser scanning microscopy and flow cytometry analysis. Modification of the liposomal surface with the IRQ peptide (IRQ-Lip), induced internalization via a novel pathway–caveolar endocytosis–in parallel with clathrin-mediated endocytosis. Furthermore, the IRQ peptide stimulated escape from endocytic vesicles, leading to efficient gene silencing. When siRNA was condensed and encapsulated in an IRQ-modified multifunctional envelope-type nano-device (IRQ- MEND), transgene expression was reduced 52% with the fusogenic lipid, DOPE/CHEMS. This result shows that the novel IRQ can be utilized for cytoplasmic delivery of macromolecules. Moreover, the IRQ has the potential to be useful for delivery therapeutic agents to parenchymal cells via caveolar endocytosis, as this uptake pathway also plays an important role in transcytosis. © 2007 Elsevier B.V. All rights reserved. Keywords: IRQ; Cellular uptake mechanism; Caveolar endocytosis; Intracellular fate 1. Introduction Cell-penetrating peptide (CPP) derivatives, such as human immunodeficiency virus (HIV)-1 Tat-(48-60) protein and Dro- sophila Antennapedia (Antp)-(48-60) protein are efficiently translocated across cellular membranes in cell culture systems [1]. These peptides possess protein transduction domains (PTDs), which are arginine-rich basic peptide sequences; peptide internalization is highly dependent on the sequence of the PTDs [2]. PTDs have been shown to successfully mediate efficient cellular uptake of a variety of molecules, such as proteins, peptides and nucleic acids [3]. It also was demon- strated that Tat peptide facilitated internalization into cells of liposomes with a diameter of approximately 200 nm [4]. In addition, complexes between Tat-liposomes and DNA also showed enhanced transfection, both in vitro and in vivo [5]. Recently, efficient translocation of peptides comprised of only 8 arginine residues, such as the Tat peptide, was demonstrated [1]. It is well-known that liposomes modified with R8 at low density are taken up mainly via clathrin- mediated endocytosis. When modified with R8 at high density, liposomes are taken up mainly via macropinocytosis and were less susceptible to lysosomal degradation, resulting in efficient transfection [6]. This result suggests that the mechanism of cellular uptake and the intracellular fate of the carrier are closely related. Furthermore, mechanistic studies of the intracellular events determining the fate of gene vectors provide the basis for designing optimal vector-based delivery systems. R8-Lip also has been demonstrated to efficiently deliver various macro- molecules. Recently, a multifunctional envelope-type nano- device (MEND) was developed by our group for packaging of pharmaceutical agents. This system was originally used as a novel delivery system for plasmid DNA (pDNA) in which core pDNA was condensed using polycations and was covered with a lipid membrane [7]. Modification of the MEND surface with R8 Available online at www.sciencedirect.com Journal of Controlled Release xx (2007) xxx – xxx COREL-04638; No of Pages 10 www.elsevier.com/locate/jconrel ⁎ Corresponding author. Tel.: +81 11 706 3919; fax: +81 11 706 4879. E-mail address: [email protected] (H. Harashima). GENE DELIVERY 0168-3659/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2007.10.020 ARTICLE IN PRESS Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-carrier targeted to a unique pathway of caveolar endocytic pathway, J. Control. Release (2007), doi:10.1016/j.jconrel.2007.10.020
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of A novel IRQ ligand-modified nano-carrier targeted to a unique pathway of caveolar endocytic pathway

Available online at www.sciencedirect.com
e xx (2007) xxx–xxx
COREL-04638; No of Pages 10
www.elsevier.com/locate/jconrel ENEDELIVERYARTICLE IN PRESS
Journal of Controlled Releas
G
A novel IRQ ligand-modified nano-carrier targeted to a unique pathwayof caveolar endocytic pathway
Diky Mudhakir, Hidetaka Akita, Erdal Tan, Hideyoshi Harashima ⁎
Laboratory for Molecular Design of Pharmaceutics, Faculty of Pharmaceutical Sciences, Hokkaido University, Kita-12, Nishi-6, Kita-ku, Sapporo 060-0812, Japan
Received 26 June 2007; accepted 20 October 2007
Abstract
In the present study, the cellular uptake and subsequent intracellular trafficking of liposomes was investigated, in which a novel peptide (IRQ),identified with in vivo phage display, was modified on the surface. Since the novel peptide IRQRRRR is rich in arginine, the cellular uptakemechanism was compared with octaarginine (R8)-modified liposomes, which are known to be taken up by cells via macropinocytosis. The uptakemechanism and intracellular trafficking of peptide-modified liposomes was determined by confocal laser scanning microscopy and flow cytometryanalysis. Modification of the liposomal surface with the IRQ peptide (IRQ-Lip), induced internalization via a novel pathway–caveolarendocytosis–in parallel with clathrin-mediated endocytosis. Furthermore, the IRQ peptide stimulated escape from endocytic vesicles, leading toefficient gene silencing. When siRNA was condensed and encapsulated in an IRQ-modified multifunctional envelope-type nano-device (IRQ-MEND), transgene expression was reduced 52% with the fusogenic lipid, DOPE/CHEMS. This result shows that the novel IRQ can be utilized forcytoplasmic delivery of macromolecules. Moreover, the IRQ has the potential to be useful for delivery therapeutic agents to parenchymal cells viacaveolar endocytosis, as this uptake pathway also plays an important role in transcytosis.© 2007 Elsevier B.V. All rights reserved.
Keywords: IRQ; Cellular uptake mechanism; Caveolar endocytosis; Intracellular fate
1. Introduction
Cell-penetrating peptide (CPP) derivatives, such as humanimmunodeficiency virus (HIV)-1 Tat-(48-60) protein and Dro-sophila Antennapedia (Antp)-(48-60) protein are efficientlytranslocated across cellular membranes in cell culture systems[1]. These peptides possess protein transduction domains(PTDs), which are arginine-rich basic peptide sequences;peptide internalization is highly dependent on the sequence ofthe PTDs [2]. PTDs have been shown to successfully mediateefficient cellular uptake of a variety of molecules, such asproteins, peptides and nucleic acids [3]. It also was demon-strated that Tat peptide facilitated internalization into cells ofliposomes with a diameter of approximately 200 nm [4]. Inaddition, complexes between Tat-liposomes and DNA alsoshowed enhanced transfection, both in vitro and in vivo [5].
⁎ Corresponding author. Tel.: +81 11 706 3919; fax: +81 11 706 4879.E-mail address: [email protected] (H. Harashima).
0168-3659/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jconrel.2007.10.020
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
Recently, efficient translocation of peptides comprised ofonly 8 arginine residues, such as the Tat peptide, wasdemonstrated [1]. It is well-known that liposomes modifiedwith R8 at low density are taken up mainly via clathrin-mediated endocytosis. When modified with R8 at high density,liposomes are taken up mainly via macropinocytosis and wereless susceptible to lysosomal degradation, resulting in efficienttransfection [6]. This result suggests that the mechanism ofcellular uptake and the intracellular fate of the carrier are closelyrelated. Furthermore, mechanistic studies of the intracellularevents determining the fate of gene vectors provide the basis fordesigning optimal vector-based delivery systems. R8-Lip alsohas been demonstrated to efficiently deliver various macro-molecules. Recently, a multifunctional envelope-type nano-device (MEND) was developed by our group for packaging ofpharmaceutical agents. This system was originally used as anovel delivery system for plasmid DNA (pDNA) in which corepDNAwas condensed using polycations and was covered with alipid membrane [7]. Modification of the MEND surface with R8
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

GENEDELIVERY
2 D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
(R8-MEND) proved to efficiently deliver pDNA into cells,resulting in a high transfection rate in vitro [6,7]. Subsequently,other therapeutic agents, such as oligodeoxynucleotide (ODN)and siRNA [8,9], were successfully packaged. R8-MEND candeliver siRNA into cells, efficiently and persistently, silencingtarget genes with minimum toxicity.
For the application of vectors in in vivo delivery, it isimportant to find a novel ligand that targets specific organs.Therefore, we attempted to find a peptide ligand that targetsskeletal muscle by in vivo phage display. As a result, we found anovel peptide, which we referred to as the IRQ peptide. Thispeptide consists of 7 amino acids that are arginine-rich andpossesses 1 isoleucine and 1 glutamine (IRQRRRR). Since thecellular uptake pathway is closely related to the subsequentintracellular trafficking and application of the carrier, asexplained above, in the present study we characterized in detailthe intracellular events of IRQ-modified liposomes (IRQ-Lip)in relation to the uptake mechanism based on a comparison withan R8-modified one. Furthermore, to evaluate IRQ function indelivery of therapeutic agents, such as siRNA, IRQ wasmodified on the surface of MEND (IRQ-MEND). Intracellulartrafficking of siRNA-encapsulated in the IRQ-MEND wasmodified by changing the lipid membranes to provide asophisticated gene-silencing effect.
In the present study, a novel IRQ peptide ligand, which wastaken up using a new uptake mechanism–caveolar endocyto-sis–in parallel with clathrin-mediated endocytosis, was devel-oped. To the best of our knowledge, this is the first report todescribe a peptide as a targeting ligand for the caveolar pathway.
2. Materials and methods
2.1. Materials
A Ph.D.-7 phage display peptide library kit was purchasedfrom New England Biolabs Inc., Beverly, Maryland. Ethylen-diamine-N,N,Nl,Nl-tetraacetic acid-disodium salt-dehydrate(EDTA), 2-amino-2-hydroxymethyl-1,3-propanediol (Tris(hydroxymethyl) amino-methane), 2-[4-(2-hydroxyethyl)-1-piperazinyl]ethanesulfonic acid (HEPES), bacto trypton, bactoyeast extract, agar, agarose S, polyethylene glycol 8000, sucrose,dimethyl sulfoxide dehydrated (DMSO), tetracycline, magne-sium chloride hexahydrate (MgCl2×6H2O), N,N-dimethylfor-mamide (DMF), heparin, and Hoechst 33342 were purchasedfrom Wako Pure Chemical Industries, (Osaka, Japan). Isopropylb-D-thiogalactoside (IPTG), 5-bromo-4-chloro-3-indolyl-b-D-galactoside (Xgal), lithium chloride (LiCl), cesium chloride(CsCl) and sodium chloride (NaCl) were purchased fromNacalaiTesque, Inc. (Kyoto, Japan). Spectra/por biotech membranesFED 3.1: MWCO 3500 was purchased from Funakoshi (Tokyo,Japan). Bigdye terminator was purchased from AppliedBiosystems (Tokyo, Japan). Phenylmethylsulfonyl fluoride(PMSF), aprotinin, leupeptin, amiloride, filipin III from Strep-tomyces filipinensis, PKH67 Green Fluorescent Cell Linker and5-cholesten-3β-ol 3-hemisuccinate (CHEMS) were purchasedfrom Sigma-Aldrich (St. Louis, MO). Egg phosphatidylcholine(EPC), cholesterol (Cho), dioleoylphosphatidyl ethanolamine
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
(DOPE), 1,2-dioleoyl-sn-glycero-3-phosphoethanol amine-N-(7-nitro-2-1,3-benzoxadiazol4-yl) (NBD-DOPE), and N-(lissa-mine rhodamine B sulfonyl)-1,2-dioleyl-sn-glycero-3-phos-phoethanolamine (Rh-DOPE) were purchased from AvantiPolar Lipids (Alabaster, AL). Syto-24 was purchased fromMolecular Probes (Eugene, Oregon, USA). Stearylated-octaar-ginine (STR-R8) was synthesized as described previously [4].Synthesis of STR-IRQ was performed following the procedureused for the synthesis of STR-R8, in which the stearyl moietywas attached to the N-terminal of the IRQ peptide. Anti-luciferase siRNA (21-mer, 5′-GCGCUGCUGGUGCCAACCC-TT-3′, 5′-GGGUUGGCACCAGCAGCAGCGCTT-3′) andanti-green fluorescent protein (GFP) siRNA (5′-GCUGACCC-UGAAGUUCAUCTT-3′, 5′-GAUGAACUUCAGGGUCAGCTT-3′) were obtained from Thermo Electron GmbH (Ulm,Germany). Luciferase assay reagents and reporter lysis bufferwere obtained from Promega Co (Madison, WI). OPTI-MEM Iwas purchased from Invitrogen Corp. (Carlsbad, CA). siRNAanti luciferase 3′-Alexa Fluor 546 was purchased from QiagenInc. (Valencia, CA). NIH3T3 cells were obtained from theAmerican Type Culture Collection (Manassas, VA).
2.2. Animals
Male Balb/c mice, aged 8 weeks old, were purchased fromSankyo Labo Service (Sapporo, Japan). The mice were housedat 23±3 °C and 50±10% relative humidity and weremaintained on a 12-h light/dark cycle. During the acclimatiza-tion, the mice were allowed free access to food and water. Allanimals were used without fasting before all experiments. Theexperimental protocols were reviewed and approved by theHokkaido University Animal Care Committee in accordancewith the “Guide for the Care and Use of Laboratory Animals”.
2.3. Methods
2.3.1. Screening of novel peptide ligands by in vivo phagedisplay method
A random 7-mer linear with 2×1011 pfu of phages peptidelibrary was injected into mice via the tail vein. After allowingphages to bind to the organ for 10 min, each mouse waseuthanized and its heart was perfused with EDTA/PBS (100 ml)to remove non-bonding phages. Skeletal muscle was collectedand homogenized. Recovered phages were determined in analiquot of homogenized tissue. For the second and thirdpanning, after recovered phages were amplified and purified,phages were injected individually to enhance their binding tothe target tissue. In the study of targeting ability of peptide toskeletal muscle, the protocol experiment was the same asexplained above, with the exception of dosage. The dosage ofpurified phage clones and non-selective phage was 1×1010 pfu.In addition, an amplification step was unnecessary after thedetermination of recovered phages.
2.3.2. DNA sequenceFrom the third round plate, 96 individual phage clones were
randomly isolated. Each plaque was grown for 4.5 h in 1 ml LB.
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

GENEDELIVERY
3D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
The phages were isolated and purified by precipitation usingPEG 8000. Phage pellets were resuspended in 100 ml iodidebuffer and 250 ml ethanol was added to precipitate the single-stranded phage DNA. Phage DNA was recovered by centrifu-gation for 10 min at 15,000 rpm. Finally DNAwas resuspendedin 30 ml TE buffer (10 mM Tris–HCl pH 8.0, 1 mM EDTA).Phage DNA was amplified by PCR (using primer phage-F,sequence (5′–3′), GCAATTCCTTTAGTGGTACC and primerphage-R, sequence (5′–3′), CCCTCATAGTTAGCGTAACG).PCR products were then reacted by florescent markers (bigdye)for DNA sequencing. The data were then analyzed using aGenetic Max software program.
2.3.3. Preparation of IRQ-Liposomes and R8-LiposomesTwo types of liposomes, IRQ-Lip and R8-Lip, were prepared
by the hydration method. Liposomes were composed of Cho andEPC in a molar ratio of 3:7. In this study, liposomes weremodified by direct conjugation of IRQ or R8 to the surface ofliposomes. To prepare IRQ-Lip and R8-Lip, STR-IRQ or STR-R8 was incorporated at 5 mol% of total lipid. To label the lipidphase, Rh-DOPE was incorporated at 1 mol% of total lipid.Briefly, lipids, totaling 0.6 mmol, were first dissolved in ethanol.After 5 mol% of the STR-IRQ or STR-R8 and 1mol% of the Rh-DOPE in ethanol were added into the bottom of a glass tube, thesolvent was removed with nitrogen gas under reduced pressure.The lipid film was then hydrated with 0.6 ml of phosphate buffersaline (PBS), followed by incubation for 10 min to hydrate thelipids. The glass tube was then sonicated for approximately1 min in a bath-type sonicator (125 W, Branson Ultrasonics,Danbury, CT). Liposomes were stored at 4 °C for a maximum ofone week. The size distribution and zeta-potential of eachsample were determined using a Zetasizer Nano ZS ZEN3600(MALVERN Instrument, Worchestershire, UK).
2.3.4. Encapsulation of siRNA into IRQ-modified liposomesFor the intracellular trafficking and transgene expression
studies, siRNA was encapsulated in IRQ-Lip, which wasprepared by the lipid hydration method previously reported forthe preparation of siRNA-encapsulated R8-MEND [9]. siRNAand STR-R8, as a polycation agent, were dissolved in RNase-free water. To condense the siRNA, the siRNA solution wasadded to the polycation solution while vortexing at roomtemperature. The concentrations of both solutions were fixed at0.1 mg/ml. The siRNA/STR-R8 complexes were prepared at anitrogen/phosphate (N/P) ratio of 2.9. A 0.25 ml aliquot ofcondensed siRNA suspension was added to the lipid film, whichwas formed by evaporation of a chloroform solution of 137.5 nmlipids, on the bottom of a glass tube followed by a 10-minincubation at room temperature to hydrate the lipids. In thisstudy, two different lipid compositions were used: DOPE/CHEMS and EPC/CHEMS, both in molar ratios of 9:2. The finallipid concentration was 0.55 mM. To coat condensed siRNAwith lipids, the glass tube was sonicated for approximately 1 minin a bath-type sonicator (125 W, Branson Ultrasonics, Danbury,CT). Finally, the surface of the liposomes was modified withIRQ by incubation with a STR-IRQ (5 mol% lipids) solution for30 min at room temperature. This siRNA-encapsulating IRQ-
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
Lip was termed an IRQ-modified multifunctional envelope-typenano-device (IRQ-MEND). The size distribution and zeta-potential of each sample were determined using a Zetasizer NanoZS ZEN3600 (MALVERN Instrument, Worchestershire, UK).
2.3.5. The uptake mechanism study
2.3.5.1. Qualitative study of uptake of liposomes by confocallaser scanning microscopy (CLSM). To investigate themechanism of internalization of IRQ-Lip and R8-Lip, 2×105
NIH3T3 cells were seeded on a 35-mm glass-base dish (Iwaki,Chiba, Japan) in 2 ml of Dulbecco's modified Eagle's medium(DMEM) containing 10% fetal calf serum (FCS) for 24 h. Beforetransfection, the cells were washed once with 1 ml of PBS andwere pre-incubated with serum-free medium in the absence orpresence of the following three compound for various times:sucrose (0.4 M) at 37 °C for 30 min; amiloride (5 mM) for10 min; or filipin (1 mg/ml) for 1 h. IRQ-Lip and R8-Lip labeledwith Rh-DOPE corresponding to 0.050 mmol lipids then wereadded and the cells were incubated for 1 h at 37 °C in thepresence or absence of inhibitors. The cells were washed 3 timeswith 1 ml of ice-cold phosphate buffer saline (PBS) supple-mented with heparin (20 units/ml) to completely removesurface-bound R8-Lip, as reported previously [10]. BecauseIRQ also is rich in arginine residues, cell surface-bound IRQ-Lipwas removed according to the protocol used for the removal ofR8-Lip. The cells then were washed once with 1 ml of Krebsbuffer and were kept on ice in 2ml of Krebs buffer until observedby confocal microscopy. Nuclei were stained with Syto-24 (finalconcentration 0.5 mM) for 20 min before microscopic analysis.
2.3.5.2. Quantification of cellular association of liposomes byflow cytometric analysis. To investigate the cellular uptake ofIRQ-Lip, 5×105 NIH3T3 cells were seeded on a 6-well plate in2 ml of DMEM containing 10% FCS for 24 h. Beforetransfection, the culture medium was removed and the cellswere washed with 1 ml of serum-free medium. To examine thecellular uptake mechanism, cells were pre-incubated at 37 °C inserum-free medium in the absence or presence of amiloride(5 mM) for 10 min, or with sucrose (0.4 M) for 30 min, or withfilipin III (1 μg/ml) for 1 h. R8-Lip and IRQ-Lip labeled withRh-DOPE corresponding to 0.1 mmol of lipids/well then wereadded and incubated for 1 h at 37 °C in the presence or absenceof inhibitors. At the end of the incubation, the culture mediumwas removed, and the cells were washed once with ice-coldPBS supplemented with heparin (20 units/ml). The cells thenwere trypsinized and collected into an Eppendorf tube. The cellsthen were washed twice after pelleting by centrifugation(1500 rpm, 4 °C, 5 min) and were suspended in 1 ml ofheparin–PBS. Finally, cells were suspended in 0.5 ml of PBSand filtered through a nylon mesh. The cells then were analyzedby a flow cytometer (FACScan, Becton Dickinson).
2.3.6. Co-localization of IRQ-MENDs with plasmacompartment
1×105 NIH3T3 cells were seeded on a 35-mm glass-basedish (Iwaki, Chiba Japan) in 2 ml of DMEM containing 10%
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

Fig. 1. Accumulation of selected phage clones in the skeletal muscle.Accumulation was expressed as the number of recovered phages per gram oftissue (pfu/g). Open bars, light gray bars, and closed bars indicate non-selectivephages, the clone number with the highest accumulation of phages, and group ofclone number of phages screened by matching frequency respectively, after anindividual administration of a purified phage via i.v. injection at a dosage of1×1010 pfu.
GENEDELIVERY
4 D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
FCS for 24 h. Before transfection, the culture medium wasremoved and the cells were washed with 1 ml of serum-freemedium. Plasma membranes then were stained with PKH67Green Fluorescent Cell Linker (Sigma-Aldrich; St. Louis, MOUSA) following the manufacturer's protocol with minormodifications. After the cells were washed with serum-freemedium, 100 ml of PKH67 Linker solution (10 mM) wasadded, and the cells were incubated for an additional 5 min atroom temperature. Staining was terminated by dilution of themedium with 100 ml of serum. The cells then were washedtwice with 1 ml PBS and twice with 1 ml serum-free medium.After washing, IRQ-MEND composed of EPC/CHEMS orDOPE/CHEMS corresponding to 0.4 mg siRNA were imme-diately added, followed by incubation for 1 h at 37 °C. The cellsthen were washed 3 times with 1 ml PBS supplemented withheparin (20 units/ml), and the cell culture medium wasexchanged with 1 ml of Krebs-Henseleit Buffer. Confocalimages were captured by a Zeiss Axiovert 2000 objective (CarlZeiss Co. Ltd., Jena, Germany).
2.3.7. Imaging of de-coating process of IRQ-MENDsNIH3T3 cells were seeded on a 35-mm glass-base dish at
1×105 cells in 2 ml of DMEM containing 10% FCS for 24 h.Before transfection, the culture medium was removed and thecells were washed once with 1 ml serum-free medium. Toexamine the de-coating phenomena, cells were incubated for 1 hat 37 °C with 1 ml serum-free medium containing IRQ-MENDs, corresponding to 1.6 mg siRNA. MENDs werelabeled with the lipid-phase marker, NBD-DOPE (1% of totallipid), and Alexa 546 was used to label the siRNA. One hourafter transfection, the cells were washed with 1 ml PBSsupplemented heparin (20 units/ml) and then were incubatedwith 2.5 ml of Hoechst 33342 (1 mg/ml) in 1 ml serum-freemedium for 10 min at room temperature. At the end ofincubation, cells were washed two times with 1 ml PBSsupplemented with heparin. The cells then were washed with1 ml of Krebs Ringer solution and were observed by CLSM in2 ml of Krebs Ringer.
2.3.8. Transgene expression of IRQ-MENDsFor the transgene expression study, 5×104 HeLa cells stably
expressing luciferase (∼108 RLU/mg protein) were seeded ontoa 24-well plate in 0.6 ml of DMEM containing 10% FCS andwere incubated for 24 h. Before transfection, cells were washedwith 0.5 ml PBS. IRQ-MENDs corresponding to 0.4 mg siRNAwere suspended in 0.25 ml OPTI-MEM without serum and thenwere incubated with cells for 3 h at 37 °C. Next, 1 ml mediumsupplemented with serum was added to the cells, followed byincubation for an additional 21 h. After 24 h, cells were washedwith 0.5 ml PBS and then were solubilized with reporter lysisbuffer (Promega, Madison, WI). Luciferase activity wasinitiated by addition of 50 ml of luciferase assay reagent(Promega, Madison, WI) to 20 ml of cell lysate and measuredby means of a luminometer (Luminescencer-PSN, ATTO,Japan). The amount of protein in the cell lysate was measuredusing a BCA protein assay kit (PIERCE, Rockford, IL). Resultswere expressed in relative light units (RLU)/mg protein.
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
3. Results
3.1. Screening of novel peptide ligands by in vivo phagedisplay method
By repeating the in vivo biopanning, numbers of recoveredphages were gradually enhanced, suggesting that phages potentin targeting of the tissues were enriched. After the third panning,it was confirmed that the number of recovered phages in skeletalmuscle was 10-fold higher than that of non-selective phages.Therefore, 96 phage clones were isolated and the coded aminoacid sequence was determined. The candidate peptides werethen picked up based on the homology information. As criteria,we introduced a parameter of matching frequency. It is definedas the number of peptides having three or more matching aminoacid sequences when compared with the other 95 peptides. Inaddition, the positions of matching amino acids need not beconsecutive. As a result, 20 peptides with the highest matchingfrequencies were selected as candidate clones. To confirm thetargeting ability of the peptides, accumulation of each phageclone in skeletal muscle was compared with non-selectivephages. As control, non-selective phages lacking selective-presenting peptides were used. As a result, a majority of thephages exceeded the non-selective phages. In particular, clone52 whose match frequency was 32 showed the highestenhancement in the skeletal muscle up to 10-fold compared tothat of the non-selective phages (Fig. 1). Clone 52 has an aminoacid sequence of IRQRRRR (IRQ). Therefore, we focused onthe IRQ peptide for the following studies.
3.2. Investigation of cellular uptake mechanism of liposomesby CLSM
The cellular uptake of IRQ-Lip was investigated todetermine the intracellular fate of the particles for use insiRNA delivery. The size and zeta-potential of the prepared
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.
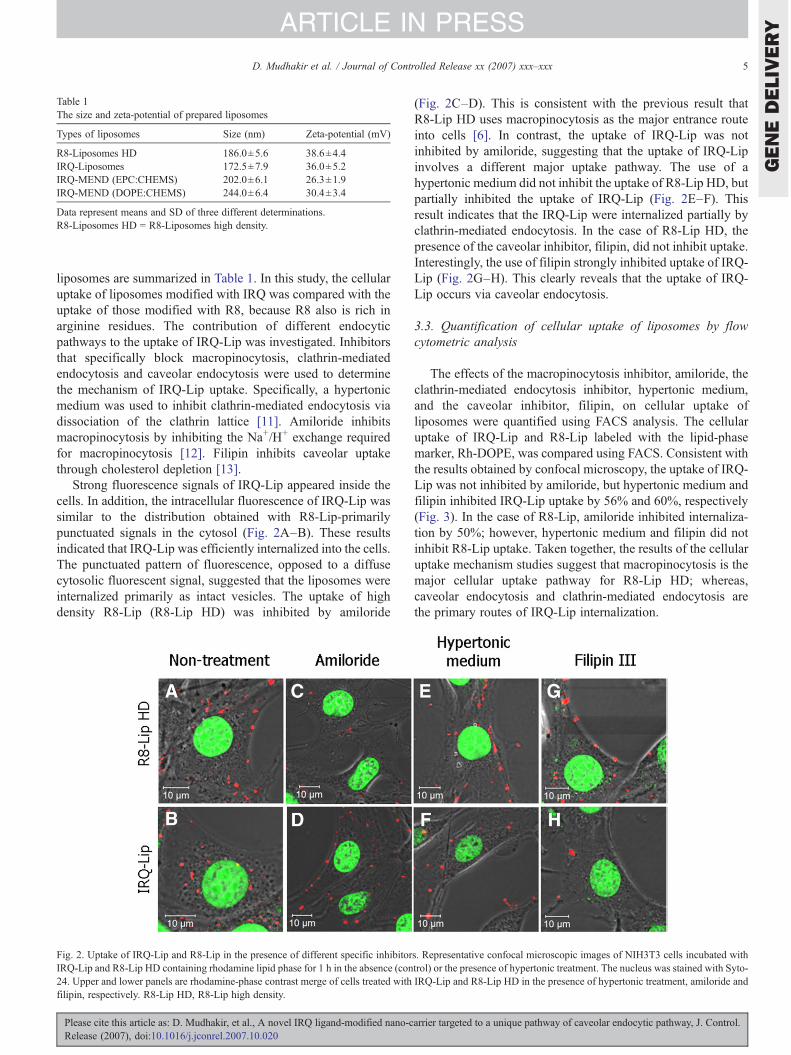
Table 1The size and zeta-potential of prepared liposomes
Types of liposomes Size (nm) Zeta-potential (mV)
R8-Liposomes HD 186.0±5.6 38.6±4.4IRQ-Liposomes 172.5±7.9 36.0±5.2IRQ-MEND (EPC:CHEMS) 202.0±6.1 26.3±1.9IRQ-MEND (DOPE:CHEMS) 244.0±6.4 30.4±3.4
Data represent means and SD of three different determinations.R8-Liposomes HD = R8-Liposomes high density.
GENEDELIVERY
5D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
liposomes are summarized in Table 1. In this study, the cellularuptake of liposomes modified with IRQ was compared with theuptake of those modified with R8, because R8 also is rich inarginine residues. The contribution of different endocyticpathways to the uptake of IRQ-Lip was investigated. Inhibitorsthat specifically block macropinocytosis, clathrin-mediatedendocytosis and caveolar endocytosis were used to determinethe mechanism of IRQ-Lip uptake. Specifically, a hypertonicmedium was used to inhibit clathrin-mediated endocytosis viadissociation of the clathrin lattice [11]. Amiloride inhibitsmacropinocytosis by inhibiting the Na+/H+ exchange requiredfor macropinocytosis [12]. Filipin inhibits caveolar uptakethrough cholesterol depletion [13].
Strong fluorescence signals of IRQ-Lip appeared inside thecells. In addition, the intracellular fluorescence of IRQ-Lip wassimilar to the distribution obtained with R8-Lip-primarilypunctuated signals in the cytosol (Fig. 2A–B). These resultsindicated that IRQ-Lip was efficiently internalized into the cells.The punctuated pattern of fluorescence, opposed to a diffusecytosolic fluorescent signal, suggested that the liposomes wereinternalized primarily as intact vesicles. The uptake of highdensity R8-Lip (R8-Lip HD) was inhibited by amiloride
Fig. 2. Uptake of IRQ-Lip and R8-Lip in the presence of different specific inhibitorIRQ-Lip and R8-Lip HD containing rhodamine lipid phase for 1 h in the absence (con24. Upper and lower panels are rhodamine-phase contrast merge of cells treated withfilipin, respectively. R8-Lip HD, R8-Lip high density.
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
(Fig. 2C–D). This is consistent with the previous result thatR8-Lip HD uses macropinocytosis as the major entrance routeinto cells [6]. In contrast, the uptake of IRQ-Lip was notinhibited by amiloride, suggesting that the uptake of IRQ-Lipinvolves a different major uptake pathway. The use of ahypertonic medium did not inhibit the uptake of R8-Lip HD, butpartially inhibited the uptake of IRQ-Lip (Fig. 2E–F). Thisresult indicates that the IRQ-Lip were internalized partially byclathrin-mediated endocytosis. In the case of R8-Lip HD, thepresence of the caveolar inhibitor, filipin, did not inhibit uptake.Interestingly, the use of filipin strongly inhibited uptake of IRQ-Lip (Fig. 2G–H). This clearly reveals that the uptake of IRQ-Lip occurs via caveolar endocytosis.
3.3. Quantification of cellular uptake of liposomes by flowcytometric analysis
The effects of the macropinocytosis inhibitor, amiloride, theclathrin-mediated endocytosis inhibitor, hypertonic medium,and the caveolar inhibitor, filipin, on cellular uptake ofliposomes were quantified using FACS analysis. The cellularuptake of IRQ-Lip and R8-Lip labeled with the lipid-phasemarker, Rh-DOPE, was compared using FACS. Consistent withthe results obtained by confocal microscopy, the uptake of IRQ-Lip was not inhibited by amiloride, but hypertonic medium andfilipin inhibited IRQ-Lip uptake by 56% and 60%, respectively(Fig. 3). In the case of R8-Lip, amiloride inhibited internaliza-tion by 50%; however, hypertonic medium and filipin did notinhibit R8-Lip uptake. Taken together, the results of the cellularuptake mechanism studies suggest that macropinocytosis is themajor cellular uptake pathway for R8-Lip HD; whereas,caveolar endocytosis and clathrin-mediated endocytosis arethe primary routes of IRQ-Lip internalization.
s. Representative confocal microscopic images of NIH3T3 cells incubated withtrol) or the presence of hypertonic treatment. The nucleus was stained with Syto-IRQ-Lip and R8-Lip HD in the presence of hypertonic treatment, amiloride and
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

Fig. 3. Cellular association of IRQ-Lip and R8-Lip in the presence of different specific inhibitors. NIH3T3 cells were incubated with IRQ-Lip or R8-Lip containingrhodamine lipid phase for 1 h in the absence (control) or presence of amiloride, hypertonic medium and filipin, respectively. Flow cytometric analysis was performed1-h post transfection.
GENEDELIVERY
6 D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
3.4. Evaluation of intracellular trafficking of IRQ-MEND
After clarifying the contribution of the endocytic uptakepathways for gene-delivery via IRQ-modified carriers, themechanism of endosomal release of siRNA encapsulated inIRQ-MEND was investigated. To assess the ability of IRQ-MEND to escape from endocytic compartments (endosomes andcaveosomes), the intracellular fate of IRQ-MEND prepared withthe non-fusogenic lipid, EPC, or with the fusogenic lipid, DOPE,was investigated. The effect ofDOPE,which enhances endosomalescape of particles trapped in endosomal vesicles through fusionwith the endosomal membrane [14], was examined. As a result,red fluorescence was observed in a punctuated pattern in thecytosol of cells transfected with IRQ-MEND composed of bothDOPE/CHEMS and EPC/CHEMS (Fig. 4). This result indicatesthat the IRQ peptide also promotes endosomal escape.
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
3.5. De-coating of IRQ-MENDs
In this study, the lipid envelope was labeled with the lipid-phase marker, NBD-DOPE, and siRNAwas labeled with Alexa546. The de-coating study was performed after incubatingviable cells with IRQ-MENDs for 1 h. If fusion plays animportant role in endosomal escape, encapsulated siRNA mustbe released from the lipid layer. However, if fusion is notrequired for escape, siRNA must co-localize with the lipidmembrane. The intracellular fluorescence of siRNAwas highlyco-localized with the non-fusogenic lipid envelope, EPC/CHEMS (Fig. 5A). This result revealed that de-coating didnot occur due to low fusion activity of lipid. When a non-fusogenic lipid was replaced with a fusogenic lipid, DOPE/CHEMS, in the envelope, most of the intracellular siRNAfluorescence was strongly detected in the cytosol as punctuated
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.
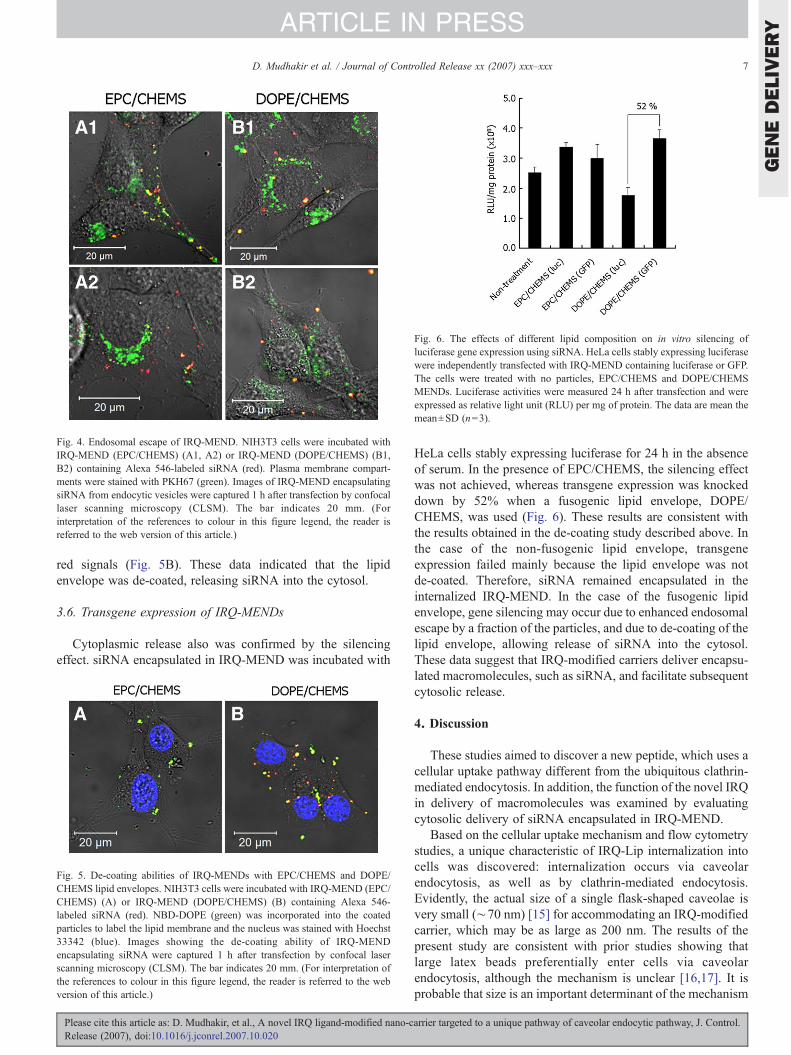
Fig. 4. Endosomal escape of IRQ-MEND. NIH3T3 cells were incubated withIRQ-MEND (EPC/CHEMS) (A1, A2) or IRQ-MEND (DOPE/CHEMS) (B1,B2) containing Alexa 546-labeled siRNA (red). Plasma membrane compart-ments were stained with PKH67 (green). Images of IRQ-MEND encapsulatingsiRNA from endocytic vesicles were captured 1 h after transfection by confocallaser scanning microscopy (CLSM). The bar indicates 20 mm. (Forinterpretation of the references to colour in this figure legend, the reader isreferred to the web version of this article.)
Fig. 6. The effects of different lipid composition on in vitro silencing ofluciferase gene expression using siRNA. HeLa cells stably expressing luciferasewere independently transfected with IRQ-MEND containing luciferase or GFP.The cells were treated with no particles, EPC/CHEMS and DOPE/CHEMSMENDs. Luciferase activities were measured 24 h after transfection and wereexpressed as relative light unit (RLU) per mg of protein. The data are mean themean±SD (n=3).
GENEDELIVERY
7D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
red signals (Fig. 5B). These data indicated that the lipidenvelope was de-coated, releasing siRNA into the cytosol.
3.6. Transgene expression of IRQ-MENDs
Cytoplasmic release also was confirmed by the silencingeffect. siRNA encapsulated in IRQ-MEND was incubated with
Fig. 5. De-coating abilities of IRQ-MENDs with EPC/CHEMS and DOPE/CHEMS lipid envelopes. NIH3T3 cells were incubated with IRQ-MEND (EPC/CHEMS) (A) or IRQ-MEND (DOPE/CHEMS) (B) containing Alexa 546-labeled siRNA (red). NBD-DOPE (green) was incorporated into the coatedparticles to label the lipid membrane and the nucleus was stained with Hoechst33342 (blue). Images showing the de-coating ability of IRQ-MENDencapsulating siRNA were captured 1 h after transfection by confocal laserscanning microscopy (CLSM). The bar indicates 20 mm. (For interpretation ofthe references to colour in this figure legend, the reader is referred to the webversion of this article.)
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
HeLa cells stably expressing luciferase for 24 h in the absenceof serum. In the presence of EPC/CHEMS, the silencing effectwas not achieved, whereas transgene expression was knockeddown by 52% when a fusogenic lipid envelope, DOPE/CHEMS, was used (Fig. 6). These results are consistent withthe results obtained in the de-coating study described above. Inthe case of the non-fusogenic lipid envelope, transgeneexpression failed mainly because the lipid envelope was notde-coated. Therefore, siRNA remained encapsulated in theinternalized IRQ-MEND. In the case of the fusogenic lipidenvelope, gene silencing may occur due to enhanced endosomalescape by a fraction of the particles, and due to de-coating of thelipid envelope, allowing release of siRNA into the cytosol.These data suggest that IRQ-modified carriers deliver encapsu-lated macromolecules, such as siRNA, and facilitate subsequentcytosolic release.
4. Discussion
These studies aimed to discover a new peptide, which uses acellular uptake pathway different from the ubiquitous clathrin-mediated endocytosis. In addition, the function of the novel IRQin delivery of macromolecules was examined by evaluatingcytosolic delivery of siRNA encapsulated in IRQ-MEND.
Based on the cellular uptake mechanism and flow cytometrystudies, a unique characteristic of IRQ-Lip internalization intocells was discovered: internalization occurs via caveolarendocytosis, as well as by clathrin-mediated endocytosis.Evidently, the actual size of a single flask-shaped caveolae isvery small (∼70 nm) [15] for accommodating an IRQ-modifiedcarrier, which may be as large as 200 nm. The results of thepresent study are consistent with prior studies showing thatlarge latex beads preferentially enter cells via caveolarendocytosis, although the mechanism is unclear [16,17]. It isprobable that size is an important determinant of the mechanism
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

GENEDELIVERY
8 D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
by which particles enter cells. However, other studies showedthat particles with sizes exceeding 200 nm were internalized viamacropinocytosis, rather than via caveolar endocytosis [18].Taken together, these results suggest that IRQ-Lip particle sizeitself is not the only determinant of the preferred cellular uptakepathway.
It has been demonstrated that caveolae may serve as a portalof entry for certain viruses, such as SV40 (Simian virus), and formacromolecules, such as albumin [19,20]. To our knowledge,IRQ is the first peptide which uses caveolar endocytosis.However, at this stage, the cell surface receptor molecule thatmediates caveolar endocytosis of IRQ-Lip is yet to beidentified. It is well-known that caveolae are invaginated,flask-shaped plasma membrane domains, which are highlyenriched in cholesterol and sphingolipids [15]. In addition, it iswell-documented that several proteins are expressed incaveolae, such as the folic acid receptor, GPI anchor, gp60,autocrine motility factor (AMF) receptor, IL-2 receptor, GM1gangliosides, sialic acid, α2β1-integrin, platelet-derived growthfactor (PDGF), epidermal growth factor (EGF), bradykinin, andthe CCK receptor [21,22]. Presumably, IRQ-Lip target onethese proteins in the caveolae or even target an unidentifiedmolecule. Therefore, additional studies to elucidate themolecular target of IRQ are needed. It is important to emphasizethat substituting only two amino acids, i.e., replacing argininewith isoleucine or glutamine, dramatically changed the cellularuptake mechanism of IRQ-Lip. Presumably, I and/or Q residuesare very important for internalization via the caveolar pathway.It was reported that the HIV-1 Tat peptide and proteinsconjugated with Tat may be taken up via caveolar endocytosis[23,24]. However, the mechanism by which endocytosis-mediated uptake of peptides, or of its modified carrier occurs,remains controversial. Different studies have demonstrated theinvolvement of various endocytic pathways in the uptake ofthese peptides or their cargo [23–26]. In the present study, theIRQ peptide was consistently found to result in caveolarendocytosis of IRQ-Lip in both qualitative and quantitativestudies. The RGD motif, known to be important for integrin-binding, has an adhesive interaction specific for tripeptiderecognition motifs [27]. Therefore, it is thought that there is aspecific sequence, comprised of at least three amino acids on theIRQ peptide that direct the peptide to the caveolar endocyticpathway.
An interesting question arises based on this phenomena: whydoes IRQ lose its ability to be taken up by macropinocytosis?IRQ peptides are rich in arginine residues, albeit fewer thanpossessed by the R8 peptide. Five arginine residues in the IRQpeptide remain to interact with heparin sulfate proteoglycans,but it is hypothesized that they are not sufficient to inducecytoskeleton rearrangement. Therefore, IRQ-Lip are not inter-nalized via macropinocytosis. Considering that IRQ areinternalized both via caveolar endocytosis and clathrin-mediatedendocytosis, it is possible that there is a specific sequence in theIRQ that is recognized by receptor molecules in caveolae. Thepart of the sequence that is not recognized by the caveolarreceptor, then, is responsible for uptake of IRQ-Lip via clathrin-mediated endocytosis. We hypothesize that IRQ peptide would
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
be useful for in vivo application since it is well accepted that thecaveolar endocytosis pathway can mediate transcytosis[15,20,28]. Therefore, IRQ-modified carriers have the potentialto traverse endothelial cells, in the luminal to basal direction,thereby delivering therapeutic genes to parenchymal cells.
The method used to investigate the potential use of IRQ-Lipin siRNA delivery was based on forming a core–shell structure,which resembled an envelope-type virus. The siRNA wascondensed and coated with a lipid envelope modified with IRQpeptide on the surface of liposomes. For efficient cytoplasmicdelivery of siRNA encapsulated in IRQ-MEND, several rate-limiting steps were considered, in particular endosomal escapeand de-coating of lipid membranes. The rate-limiting steps wereclarified bymodifying the vectors via changes in the type of lipidmembrane used. Two kinds of lipid layer were used in this study,a non-fusogenic lipid, EPC/CHEMS, and a fusogenic-lipid,DOPE/CHEMS. It is generally accepted that inclusion of DOPEallows endosomal membrane fusion [14]. In acidic conditions,the lipid structure changes from a lamellar to a non-lamellarphase, such as hexagonal HII, HI, or to a cubic phase [29].Therefore, IRQ escaped from endosomes when DOPE/CHEMSwas used. Interestingly, strong red punctuated intracellularfluorescence signals of Alexa 546-labeled siRNA also wereobserved in the cytosol when IRQ-MEND composed of non-fusogenic EPC/CHEMSwere used (Fig. 4A and B). This clearlyindicated that the IRQ peptide promoted endosomal escape ofthe MENDs. It is hypothesized that if the red signals of Alexa546-siRNA are more abundant in the use of DOPE/CHEMS thanthat of EPC/CHEMS, then the escape mechanism of the IRQ-modified liposome is via a fusion-dependent manner. However,if the detected red fluorescence of Alexa 546-siRNA in thecytosol is comparable between DOPE/CHEMS and EPC/CHEMS, then the mechanism of the IRQ-modified liposomeis related to the fusion-independent manner. Our results showedthat the comparable fraction of siRNA was detected as a redcluster between fusogenic and non-fusogenic lipids. Further-more, we quantified the efficiencies of endosomal escapemechanisms of liposomes by the confocal image-assisted three-dimensionally integrated quantification (CIDIQ) method[30,31]. In this method, the pixel areas of pDNA were used asthe index of the amount of pDNA. The efficiencies of endosomalescape of IRQ-MENDs were calculated as the fraction of escapefrom endosome/lysosome (nuclear fraction plus cytoplasmicfraction) divided by the total cellular uptake. As a result, thefraction of endosomal escape for IRQ-MEND composed ofEPC/CHEMS and DOPE/CHEMS was 64.20±3.89% and62.48±2.61%, respectively. In addition, statistical calculationby t-test showed that the efficiency of endosomal escape of bothtypes of lipid was not significantly different (p value=0.72).Consistent with the results obtained by qualitative study, thequantification of endosomal escape efficiencies of EPC/CHEMSis considerably comparable with DOPE/CHEMS. The IRQpeptide promoted an escape process of the carrier via a fusion-independent mechanism, presumably via disruption. Therefore,the data presented here suggest that the novel IRQ has uniquecharacteristics: not only is it internalized via caveolar endocy-tosis, but it also facilitates endosomal escape.
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

GENEDELIVERY
9D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
Another important issue than endosomal escape for efficientcytosolic delivery of siRNA is de-coating of lipid membranethat we considered. Although the IRQ peptide mediated escapefrom the endocytic vesicles of MEND composed of EPC/CHEMS, all intracellular siRNAwas co-localized with the lipidenvelope of the MEND. This indicates that siRNAwas trappedinside the particles because the lipid film was not de-coated.De-coating ability is enhanced by the use of pH-sensitivefusogenic lipids, such as DOPE. When DOPE was used, manystrong, red fluorescence signals were detected in the cytosol,suggesting an effective de-coating (Fig. 5B). This resultindicates that, after the IRQ-MEND composed of DOPE/CHEMS trafficked to endosomes, the lipid membrane was de-coated during endosomal escape. Therefore, the novel IRQ hasa function to facilitate cytoplasmic delivery of macromoleculesfor therapeutic agents.
5. Conclusion
The results of the present study indicate that a specificsequence on the ligand determines the internalization mechan-ism. Substitution of only two amino acids in the arginine-richpeptide sequence altered the route of internalization. The IRQpeptide promoted internalization cells via a unique pathway—caveolar endocytosis in the NIH3T3 cells tested in the presentstudy. In addition, the novel IRQ peptide mediates escape fromthe endocytic compartment. The remarkable differences in genesilencing by siRNA using different lipid vectors can beexplained principally by differences in the intracellular traffick-ing of the carriers.
Acknowledgements
This work was supported by the Core Research for Evolutionof Science and Technology (CREST), Japan Science andTechnology Corporation (JST). The authors also wish to thankDr. Pam Hinton and Dr. James L. McDonald for their helpfuladvice in writing the English manuscript.
References
[1] S. Futaki, Arginine-rich peptides: potential for intracellular delivery ofmacromolecules and the mystery of the translocation mechanism, Int. J.Pharm. 245 (2002) 1–7.
[2] S. Futaki, W. Ohashi, T. Suzuki, M. Niwa, S. Tanaka, K. Ueda, H.Harashima, Y. Sugiura, Stearylated arginine-rich peptides: a new class oftransfection systems, Bioconjug. Chem. 12 (2001) 1005–1011.
[3] M. Lindgren, M. Hallbrink, A. Prochiantz, U. Langel, Cell-penetratingpeptides, Trends Pharmacol. Sci. 21 (2000) 99–103.
[4] V.P. Torchilin, R. Rammohan, V. Weissig, T.S. Levchenko, Tat peptide onthe surface of liposomes affords their efficient intracellular delivery even atlow temperature and in the presence of metabolic inhibitors, Proc. Natl.Acad. Sci. U. S. A. 98 (2001) 8786–8791.
[5] V.P. Torchilin, T.S. Levchenko, R. Rammohan, N. Volodina, B.Papahadjopoulos-Sternberg, G.G. D'Souza, Cell transfection in vitro andin vivo with nontoxic TAT peptide–liposome–DNA complexes, Proc.Natl. Acad. Sci. U. S. A. 100 (2003) 1972–1977.
[6] I.A. Khalil, K. Kogure, S. Futaki, H. Harashima, High density ofoctaarginine stimulates macropinocytosis leading to efficient intracellulartrafficking for gene expression, J. Biol. Chem. 282 (6) (2006) 3544–3551.
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
[7] K. Kogure, R. Moriguchi, K. Sasaki, M. Ueno, S. Futaki, H. Harashima,Development of a non-viral multifunctional envelope-type nano device bya novel lipid film hydration method, J. Control. Release 98 (2) (2004)317–323.
[8] Y. Nakamura, K. Kogure, Y. Yamada, S. Futaki, H. Harashima, Significantand prolonged antisense effect of a multifunctional envelope-type nanodevice encapsulating antisense oligodeoxynucleotide, J. Pharm. Pharma-col. 58 (4) (2006) 431–437.
[9] Y. Nakamura, K. Kogure, S. Futaki, H. Harashima, Octaarginine-modifiedmultifunctional envelope-type nano device for siRNA, J. Control. Release119 (2007) 360–367.
[10] I.A. Khalil, S. Futaki, M. Niwa, Y. Baba, N. Kaji, H. Kamiya, H.Harashima, Mechanism of improved gene transfer by the N-terminalstearylation of octaarginine: enhanced cellular association by hydrophobiccore formation, Gene Ther. 11 (2004) 636–644.
[11] J.E. Heuser, R.G. Anderson, Hypertonic media inhibit receptor-mediatedendocytosis by blocking clathrin-coated pit formation, J. Cell Biol. 108(1989) 389–400.
[12] L.J. Hewlett, A.R. Prescott, C. Watts, The coated pit and macropinocyticpathways serve distinct endosome populations, J. Cell Biol. 124 (1994)689–703.
[13] C. Lamaze, S.L. Schmid, The emergence of clathrin-independent pinocyticpathways, Curr. Opin. Cell Biol. 7 (1995) 573–580.
[14] L. Wasungu, D. Hoekstra, Cationic lipids, lipoplexes and intracellulardelivery of genes, J. Control. Release 116 (2006) 255–264.
[15] J.E. Schnitzer, Caveolae: from basic trafficking mechanism to targetingtranscytosis for tissue-specific drug and gene delivery in vivo, Adv. DrugDeliv. Rev. 49 (2001) 265–280.
[16] J. Rejman, V. Oberle, I.S. Zuhorn, D. Hoekstra, Size-dependentinternalization of particles via pathways of clathrin- and caveolae-mediated endocytosis, Biochem. 377 (2004) 159–169.
[17] J. Rejman, M. Conese, D. Hoekstra, Gene transfer by means of lipo- andpolyplexes: role of clathrin and caveolae-mediated endocytosis,J. Liposome Res. 16 (3) (2006) 237–247.
[18] S. Grosse, Y. Aron, G. Thevenot, D. Francois, M. Monsigny, I. Fajac,Potocytosis and cellular exit of complexes as cellular pathways for genedelivery by polycations, J. Gene Med. 7 (2005) 1275–1286.
[19] L. Pelkmans, J. Kartenback, A. Helenius, Caveolar endocytosis of Simianvirus 40 reveals a novel two-step vesicular transport pathway to the ER,Nat. Cell Biol. 3 (2001) 473–483.
[20] J.E. Schnitzer, P. Oh, Albondin-mediated capillary permeability toalbumin. Differential role of receptors in endothelial transcytosis andendocytosis of native and modified albumins, J. Biol. Chem. 269 (1994)6072–6082.
[21] L. Pelkmans, A. Helenius, Endocytosis via caveolae, Traffic 3 (2002)311–320.
[22] P.W. Shaul, R.G.W. Anderson, Role of plasmalemmal caveolae in signaltransduction, Am. J. Physiol. 275 (1998) L843–L851.
[23] A. Ferrari, V. Pellegrini, C. Arcangeli, A. Fittipaldi, M. Giacca, F. Beltram,Caveolae-mediated internalization of extracellular HIV-1 Tat fusionprotein visualized in real time, Molec. Ther. 8 (2) (2003) 284–294.
[24] A. Fittipaldi, A. Ferrari, M. Zoppe, C. Arcangeli, V. Pellegrini, F. Beltram,M. Giacca, Cell membrane lipid rafts mediate caveolar endocytosis ofHIV-1 Tat fusion proteins, J. Biol. Chem. 278 (2003) 34141–34149.
[25] I. Nakase, M. Niwa, T. Takeuchi, K. Sonomura, N. Kawabata, Y.Koike, M. Takehashi, S. Tanaka, K. Ueda, J.C. Simpson, A.T. Jones, Y.Sugiura, S. Futaki, Cellular uptake of arginine-rich peptides: roles formacropinocytosis and actin rearrangement, Molec. Ther. 10 (2004)1011–1022.
[26] M. Lundberg, S. Wikstrom, M. Johansson, Cell surface adherence andendocytosis of protein transduction domains, Molec. Ther. 8 (2003)143–150.
[27] K. Temming, R.M. Schiffelers, G. Molema, R.J. Kok, RGD-basedstrategies for selective delivery of therapeutics and imaging agents to thetumour vasculature, Drug Resist. Updat. 8 (2005) 381–402.
[28] S. Matveev, X. Li, W. Everson, E.J. Smart, The role of caveolae andcaveolin in vesicle-dependent and vesicle-independent trafficking, Adv.Drug Deliv. Rev. 49 (2001) 237–250.
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.

GENEDELIVERY
10 D. Mudhakir et al. / Journal of Controlled Release xx (2007) xxx–xxx
ARTICLE IN PRESS
[29] D. Hoekstra, J. Rejman, L. Wasungu, F. Shi, I. Zuhorn, Gene delivery bycationic lipids: in and out of an endosome, Biochem. Soc. Trans. 35 (2007)68–71.
[30] H. Akita, R. Ito, I.A. Khalil, S. Futaki, H. Harashima, Quantitative three-dimensional analysis of the intracellular trafficking of plasmid DNAtransfected by a nonviral gene delivery system using confocal laserscanning microscopy, Molec. Ther. 9 (3) (2004) 443–451.
Please cite this article as: D. Mudhakir, et al., A novel IRQ ligand-modified nano-cRelease (2007), doi:10.1016/j.jconrel.2007.10.020
[31] S. Hama, H. Akita, R. Ito, H. Mizuguchi, T. Hayakawa, H. Harashima,Quantitative comparison of intracellular trafficking and nuclear transcrip-tion between adenoviral and lipoplex systems, Molec. Ther. 13 (4) (2006)786–794.
arrier targeted to a unique pathway of caveolar endocytic pathway, J. Control.




















