
A novel early chorioallantoic membrane assay demonstrates quantitative and qualitative changes...
12
A novel early chorioallantoic membrane assay demonstrates quantitative and qualitative changes caused by antiangiogenic substances SUSAN J. HAZEL ADELAIDE, AUSTRALIA The chicken chorioallantoic membrane (CAM) has been extensively used in the study of angiogenesis. However, the CAM assay can be difficult and time-consum- ing to quantify, provides poor quality images of the results, and is not very repro- ducible. In this study, a novel early CAM assay was developed: It was found to be quantitative through relatively simple methods, enabled high-quality imaging of results, and was reproducible. Additionally, unique qualitative changes in vessel structure were observed, and it was possible to measure veins and arteries sepa- rately. Treatment of the CAM on days 4 and 5 with SU5614, suramin, fumagillin, amiloride, and PI-88 reduced blood-vessel growth. SU5614 (4 g) resulted in signif- icant reductions in artery but not vein length (60% and 111%, respectively, vs control). Suramin tended to increase CAM vasculature at 50 g but caused dra- matic reductions both in vessel length and CAM growth at 100 g. As with SU5614, the effect was greater with regard to arterial compared with venous length (49% and 74%, respectively, vs control). PI-88 (20 g) also decreased artery and vein length (66% and 80%, respectively, vs control). In contrast, fumagillin (5 g) and amiloride (20 g) both reduced arterial growth slightly less than venous growth (67% and 54% and 50% and 44%, respectively, vs control). Each antiangiogenic substance caused a different qualitative pattern of change in vessel branching and structure. The early CAM assay will be useful in the screening of antiangiogenic substances. Further study of the qualitative effects of antiangiogenic treatments may be a valuable tool to increase our understanding of the angiogenic process itself. (J Lab Clin Med 2003;141:217-28) Abbreviations: ANOVA analysis of variance; bFGF basic fibroblast growth factor; CAM chorioallantoic membrane; DMEM Dulbecco’s minimum essential media; DMSO dimeth- ylsulfoxide; PDGF platelet-derived growth factor; SEM standard error of the mean; TGF-1 transforming growth factor-1; TNF- tumor necrosis factor-; uPA urokinase plasminogen activator; uPAR urokinase plasminogen activator receptor; VEGF vascular endothelial growth factor A ngiogenesis, or the formation of new blood vessels from preexisting vasculature, plays a role in pathologic processes such as the growth and metastasis of tumors, diabetic retinopathy, and psoriasis. For this reason, the study of angiogenesis is an active area of research. Blood vessels form through a complex process involving multiple steps and several cell types. In vitro models, although useful in delineating parts of this process, may not be represen- tative of what occurs in vivo. One useful in vivo system that has been used extensively in angiogenesis research is the highly vascularized CAM of the chicken em- bryo. 1 Chicken embryos are less expensive to use than whole animals such as rodents, making the CAM assay From the Division of Veterinary Services, Institute of Medical and Veterinary Science. Submitted for publication June 24, 2002; revision submitted October 31, 2002; accepted November 19, 2002. Reprint requests: Susan J. Hazel, MD, Veterinary Services Division, Institute of Medical and Veterinary Science, Adelaide, Australia, 101 Blacks Road, Gilles Plains, SA 5007; e-mail: [email protected]. Copyright © 2003 by Mosby, Inc. All rights reserved. 0022-2143/2003 $30.00 0 doi:10.1067/mlc.2003.19 217
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A novel early chorioallantoic membrane assay demonstrates quantitative and qualitative changes...

A novel early chorioallantoic membrane assaydemonstrates quantitative and qualitative changescaused by antiangiogenic substances
SUSAN J. HAZEL
ADELAIDE, AUSTRALIA
The chicken chorioallantoic membrane (CAM) has been extensively used in thestudy of angiogenesis. However, the CAM assay can be difficult and time-consum-ing to quantify, provides poor quality images of the results, and is not very repro-ducible. In this study, a novel early CAM assay was developed: It was found to bequantitative through relatively simple methods, enabled high-quality imaging ofresults, and was reproducible. Additionally, unique qualitative changes in vesselstructure were observed, and it was possible to measure veins and arteries sepa-rately. Treatment of the CAM on days 4 and 5 with SU5614, suramin, fumagillin,amiloride, and PI-88 reduced blood-vessel growth. SU5614 (4 �g) resulted in signif-icant reductions in artery but not vein length (60% and 111%, respectively, vscontrol). Suramin tended to increase CAM vasculature at 50 �g but caused dra-matic reductions both in vessel length and CAM growth at 100 �g. As with SU5614,the effect was greater with regard to arterial compared with venous length (49%and 74%, respectively, vs control). PI-88 (20 �g) also decreased artery and veinlength (66% and 80%, respectively, vs control). In contrast, fumagillin (5 �g) andamiloride (20 �g) both reduced arterial growth slightly less than venous growth (67%and 54% and 50% and 44%, respectively, vs control). Each antiangiogenic substancecaused a different qualitative pattern of change in vessel branching and structure. Theearly CAM assay will be useful in the screening of antiangiogenic substances. Further studyof the qualitative effects of antiangiogenic treatments may be a valuable tool to increaseour understanding of the angiogenic process itself. (J Lab Clin Med 2003;141:217-28)
Abbreviations: ANOVA � analysis of variance; bFGF � basic fibroblast growth factor; CAM �chorioallantoic membrane; DMEM � Dulbecco’s minimum essential media; DMSO � dimeth-ylsulfoxide; PDGF � platelet-derived growth factor; SEM � standard error of the mean;TGF-�1 � transforming growth factor-�1; TNF-� � tumor necrosis factor-�; uPA � urokinaseplasminogen activator; uPAR � urokinase plasminogen activator receptor; VEGF � vascularendothelial growth factor
A ngiogenesis, or the formation of new bloodvessels from preexisting vasculature, plays arole in pathologic processes such as the
growth and metastasis of tumors, diabetic retinopathy,and psoriasis. For this reason, the study of angiogenesisis an active area of research. Blood vessels formthrough a complex process involving multiple steps and
several cell types. In vitro models, although useful indelineating parts of this process, may not be represen-tative of what occurs in vivo. One useful in vivo systemthat has been used extensively in angiogenesis researchis the highly vascularized CAM of the chicken em-bryo.1 Chicken embryos are less expensive to use thanwhole animals such as rodents, making the CAM assay
From the Division of Veterinary Services, Institute of Medical andVeterinary Science.
Submitted for publication June 24, 2002; revision submitted October31, 2002; accepted November 19, 2002.
Reprint requests: Susan J. Hazel, MD, Veterinary Services Division, Institute
of Medical and Veterinary Science, Adelaide, Australia, 101 Blacks Road,Gilles Plains, SA 5007; e-mail: [email protected].
Copyright © 2003 by Mosby, Inc. All rights reserved.
0022-2143/2003 $30.00 � 0
doi:10.1067/mlc.2003.19
217
attractive for investigators to screen antiangiogenicsubstances.1
The CAM is an extraembryonic membrane that de-
velops between days 3 and 4 of incubation. Bloodvessels in the CAM expand rapidly until approximatelyday 11, when the endothelial mitotic index declines
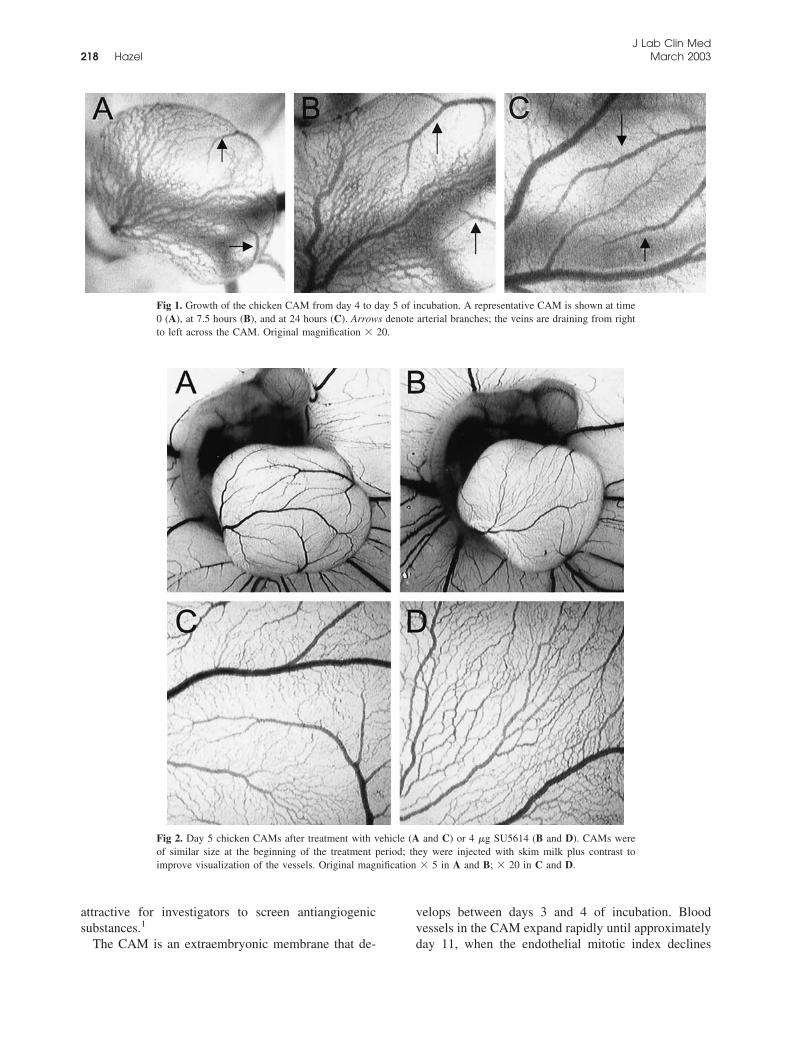
Fig 1. Growth of the chicken CAM from day 4 to day 5 of incubation. A representative CAM is shown at time0 (A), at 7.5 hours (B), and at 24 hours (C). Arrows denote arterial branches; the veins are draining from rightto left across the CAM. Original magnification � 20.
Fig 2. Day 5 chicken CAMs after treatment with vehicle (A and C) or 4 �g SU5614 (B and D). CAMs wereof similar size at the beginning of the treatment period; they were injected with skim milk plus contrast toimprove visualization of the vessels. Original magnification � 5 in A and B; � 20 in C and D.
J Lab Clin Med218 Hazel March 2003
rapidly.2 Many protocols have been used for the CAMassay; treatments have been applied with the use ofgelatin sponges,3 Silastic rings,4 methylcellulose discs,5
silicone rings,6 filter discs with cortisone acetate andbFGF,7 and plastic discs containing cortisone acetate8
or simply by dropping solution onto the CAM surface.9
Quantification of the CAM vasculature has been per-formed with the use of point-counting of photographedimages,6 radiolabeled proline to measure collagenousprotein synthesis,8 fractal analysis of digital images,9
and even subjective observations of reduced vascula-ture.7 The diversity of available protocols illustratesthat no single method offers any major advantage;investigators must compromise between using an assaythat is relatively easy and quick to perform and achiev-ing replicable and quantitative results. In traditionalmethods of performing the CAM assay, the majordrawbacks are that the assay can be (1) difficult andtime-consuming to quantify, (2) not amenable to ac-quiring good quality in situ images of the results, and(3) poorly reproducible. In this study, the aim was todevelop a quantitative and reproducible CAM assay inwhich high-quality in situ images could be obtained.Embryos were used at an early stage, from day 4 ofincubation; hence the method was called the earlyCAM assay.
The early CAM assay was validated with the use ofknown antiangiogenic substances, including SU5614,suramin, fumagillin, amiloride, and PI-88. SU5614 is atyrosine kinase inhibitor with activity against the Flk-1VEGF and PDGF receptors.10 Suramin has multipleactions that may influence angiogenesis, including in-hibition of bFGF and VEGF signaling,11,12; proteinkinase C13; and heparanase.14 Fumagillin causes endo-thelial cell-cycle arrest in the late G1 phase,15 possiblyby covalently binding to methionine aminopeptidasetype 2.16 Amiloride is an inhibitor of angiogenesis thatacts by blocking the Na�-H� antiporter or by inhib-iting the uPA-uPAR complex.17 Finally, PI-88 is asulfate oligosaccharide mimic of heparan sulfate thatinhibits the interaction between FGF and heparan sul-fate and of the enzyme heparanase.18
METHODS
Materials. Suramin, fumagillin, amiloride, and methylcel-lulose were purchased from Sigma Chemical Co (St Louis,Mo), and SU5614 was from Calbiochem (San Diego, Calif).PI-88 was a generous gift from Progen Industries Ltd (Bris-bane, Australia).
Early CAM assay. Fertilized chicken eggs (HiChick Breed-ing Co, Kapunda, Australia) were incubated at 39.5°C for 3days in an automatically turning egg incubator (Incubators
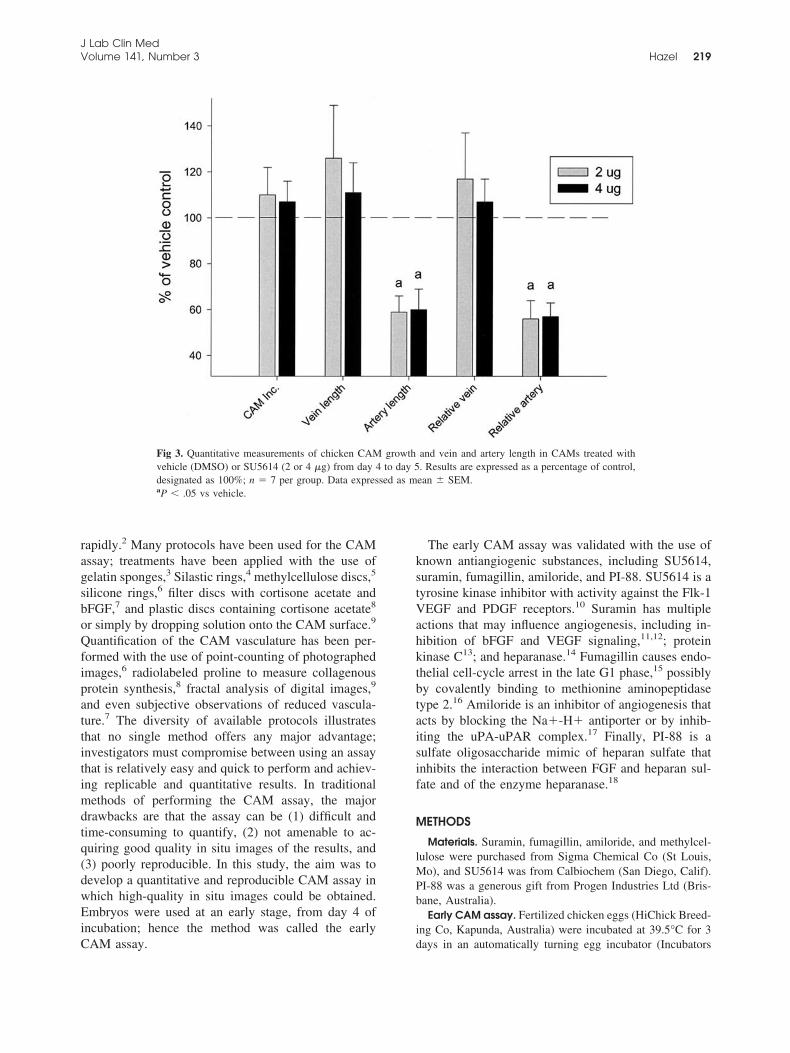
Fig 3. Quantitative measurements of chicken CAM growth and vein and artery length in CAMs treated withvehicle (DMSO) or SU5614 (2 or 4 �g) from day 4 to day 5. Results are expressed as a percentage of control,designated as 100%; n � 7 per group. Data expressed as mean � SEM.aP � .05 vs vehicle.
J Lab Clin MedVolume 141, Number 3 Hazel 219
and More, Adelaide, Australia). Embryos were then crackedfrom the eggshell into an external apparatus. A modificationof a previously published egg-cracking method was used.19 Inbrief, the shell above the air sac was broken and the surround-ing shell peeled away with forceps; care was taken not tobreak the inner shell membrane. The egg was tilted and theinner shell membrane broken, which allowed the contents todrain. On average, the number of eggs successfully crackedfrom the egg was 84%, with approximately equal lossesstemming from infertility, developmental abnormalities, andbreakage. The external apparatus, a modification of previousstudies, consisted of a 77 � 60-mm PVC pipe with plasticwrap (O-So Easy Cling Wrap, Clorox Australia Pty Limited,Padstow, Australia) spread on the top and secured with a77 � 10-mm PVC pipe ring.20,21 Two milliliters of DMEMcontaining penicillin/gentamicin (12 and 16 �g/mL, respec-tively) was added to the plastic wrap beforehand, and a sterilePetri-dish lid was placed on top to prevent contamination. Theex ovo eggs were then placed in a humidified 37°C incubatorfor approximately 24 hours. Of the eggs successfully crackedout, only approximately 81% were available for experimen-tation; most exclusions were made because of developmentalabnormalities or leakage of the yolk. Therefore approximately68% of the original eggs were used for experimentation.
On day 4, the embryos were photographed at 5� magni-fication with the use of a dissecting microscope (SZ-CTVOlympus Optical Co, Ltd, Tokyo, Japan) with an attacheddigital camera (Panasonic GP_KR222, Panasonic, Osaka, Ja-pan). Images were captured with image-analysis software(Video Pro; Leading Edge Australia Pty Ltd, Adelaide, Aus-
tralia). The area of the CAM was measured for each embryo,after which the embryos were stratified into treatment groupsof similar size, with a vehicle control in each group. It wasnecessary to compare CAMs that started at similar sizesbecause relatively small differences in size on day 4 becamelarger differences on day 5.
Treatments were applied to 5- to 8-mm diameter methyl-cellulose discs that had been previously dried on Petri dishes.The substances were applied to the discs in a volume of 2 to10 �L, after which the discs were dried overnight undervacuum. Vehicle discs consisted of the diluent used for eachsubstance (eg, DMSO, ethanol, and water). Because eachgroup contained a vehicle-treated CAM, treated CAMs werecompared with a control CAM started at a similar size. Discswere placed on top of the CAMs; these were approximatelythree times larger than the CAM area at the time of applica-tion, meaning that the substance was diffused over the entireCAM surface. Incubation was continued for another 18 to 24hours. Treatment groups included replicates of at least twoseparate experiments.
Quantification of CAM growth and vasculature. On day5, images of the CAM vasculature were captured. To improveimage contrast, a 30-gauge needle was used to inject skimmilk (10% wt/vol in water) with 0.2% blue food coloring(Queen Fine Foods Pty Ltd, Enoggera, Australia) into theCAM. Quantification was performed on 5� images with theuse of image analysis. Vein and artery lengths were tracedseparately, and the CAM area also measured. At this level ofmagnification, only larger vessel branches were included.
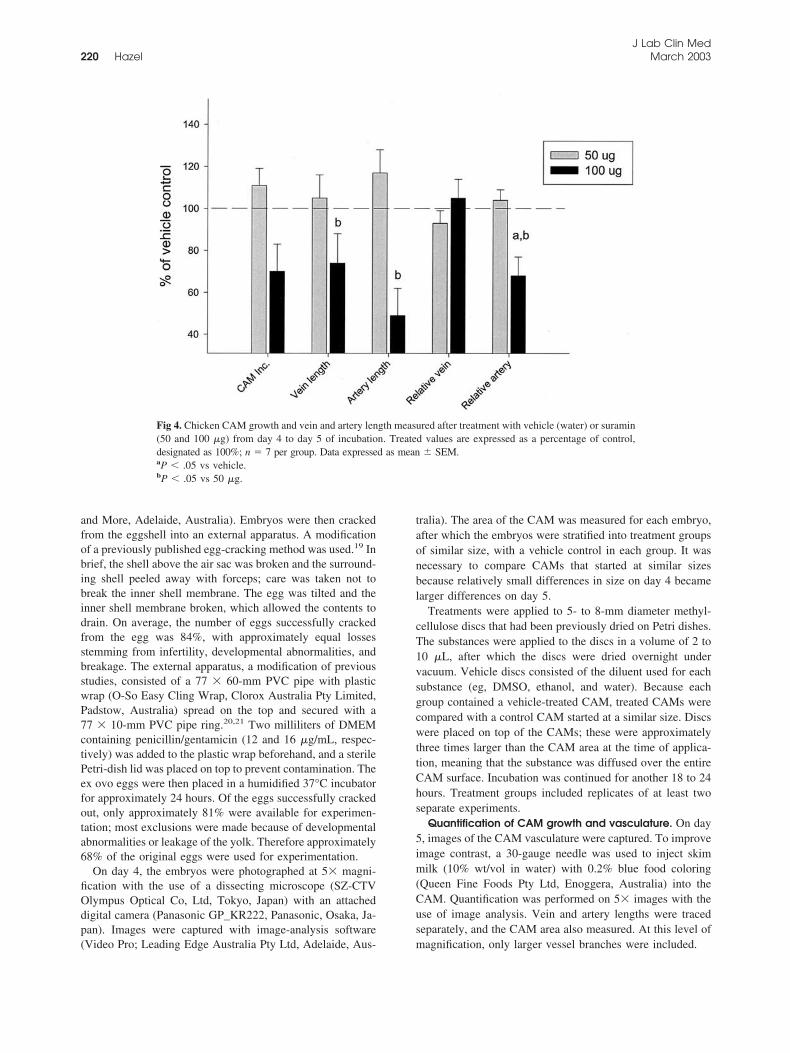
Fig 4. Chicken CAM growth and vein and artery length measured after treatment with vehicle (water) or suramin(50 and 100 �g) from day 4 to day 5 of incubation. Treated values are expressed as a percentage of control,designated as 100%; n � 7 per group. Data expressed as mean � SEM.aP � .05 vs vehicle.bP � .05 vs 50 �g.
J Lab Clin Med220 Hazel March 2003
Statistical analysis. Statistical analysis was performed withthe use of SigmaStat for Windows version 2.03 (SPSS Inc,Chicago, Ill). One-way ANOVA was used, as were pairwisemultiple comparisons employing the Student-Newman-Keulsmethod. When a group was nonnormally distributed,ANOVA on ranks was performed, followed by Tukey’s test.P values of less than .05 were considered significant.
RESULTS
Normal development of CAM. The CAM first becamevisible late on day 3 or early on day 4 of incubation. Onday 4, the CAM area was between 2.5 and 9.5 mm2; 18to 21 hours later, this area had grown to 27 to 75mm2,an increase of approximately seven- to 11-fold. Notonly were increases in the number and length of CAMvessels noted, but dramatic remodeling and reorganiza-tion of these vessels were also observed. In the newlyvisible CAM, vessels formed a primitive plexus ofsimilarly sized tubes, with veins and arteries indistin-guishable from each other. Within hours, the CAM
vessels could be clearly identified as venous or arterial.In Fig 1, a representative CAM is shown during a24-hour period stretching from day 4 to day 5. At theearliest time point, the vein is draining from right to leftand consists of multiple branches of similar size (Fig 1,A). Two straighter arteries are visible above and belowthe vein on the right side of the CAM. Seven-and-a-halfhours later, the CAM edges are outside the image, andtwo main vein branches of a significantly larger diam-eter than the surrounding venules are visible. Note thedramatic increase in length of the upper artery duringthis period. Twenty-four hours later, the mature inter-digitating pattern of vessels is present, with veins andarteries in parallel. Arterioles branch into smaller cap-illaries, which are not obvious in the captured image butare clearly visible, with single blood cells passing throughbeneath the dissecting microscope. On day 5, capillarieshad migrated up into a superficial level in the CAM, withthe main arteries and veins deeper in the membrane.
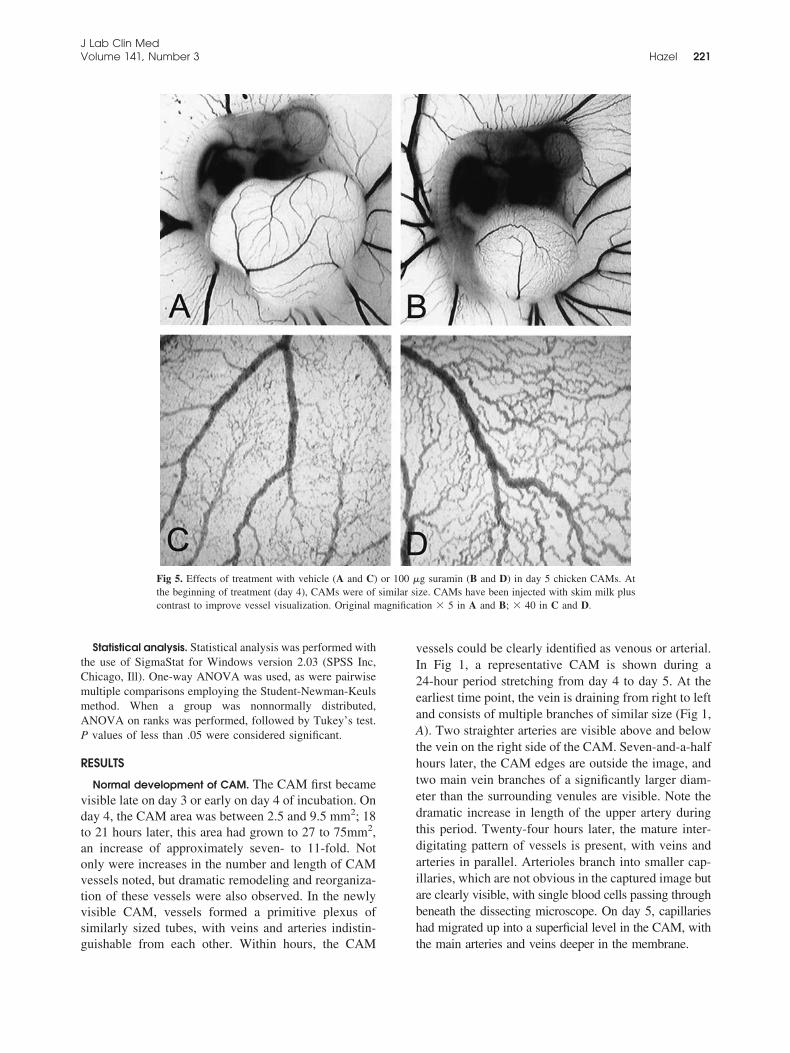
Fig 5. Effects of treatment with vehicle (A and C) or 100 �g suramin (B and D) in day 5 chicken CAMs. Atthe beginning of treatment (day 4), CAMs were of similar size. CAMs have been injected with skim milk pluscontrast to improve vessel visualization. Original magnification � 5 in A and B; � 40 in C and D.
J Lab Clin MedVolume 141, Number 3 Hazel 221

Effects of antiangiogenic substances in the early CAMassay. The VEGF receptor inhibitor SU5614 had adramatic effect on CAM vasculature (Fig 2). The pat-tern of major vessels in normal day 5 CAMs wasuniform, with interdigitating arteries and veins. Theveins drain from right to left across the CAM surfaceand disappear over the left edge. Artery branches ap-pear over the other edges. In the vehicle control(DMSO; Fig 2, A), the arteries are on the right and atthe bottom edge of the CAM. Unlike the control, in theSU5614-treated CAM the major vein branches weresurrounded by a network of venules of similar diame-ters, running in parallel (Fig 2, B and D). Arterybranches were poorly developed on the CAM edges.Blood from the arteries drained directly into the veinsbecause no capillary network had formed. Quantifica-tion confirmed that the artery lengths were reduced to60% of control (P � .05), whereas vein lengths wereconversely increased to 111% of control in the grouptreated with 4 �g SU5614 (Fig 3). Results were similarfor CAMs treated with 2 and 4 �g of SU5614 (7.3 and14.7 nmol, respectively). Although the vasculature wassignificantly affected, increases in CAM area werecomparable between the SU5614 and control groups.
At a dose of 50 �g (35 nmol), suramin tended toincrease CAM size and vessel length, but 100 �g (70nmol) had an antiangiogenic effect, evidenced by re-duced vessel length and CAM size (Fig 4). Greater
reduction in artery versus vein length (49% and 74%,respectively) was noted, and artery lengths were alsoreduced to 68% of control relative to the size of theCAM (P � .05). CAM growth decreased to 70% ofvehicle control (water). A CAM treated with 100 �g ofsuramin is shown in Fig 5, with the blood vesselsgreatly reduced compared with the matched control(Fig 5, A and B). The major arteries and veins wereconnected by highly tortuous vessels of similar size(Fig 5, D), unlike the ordered branching of the controlCAM (Fig 5, C). As seen with SU5614, no superficialcapillary network had formed.
Fumagillin was also antiangiogenic in the earlyCAM, with a different qualitative pattern of changes.Unlike the results seen with SU5614 and suramin, therewas a slightly greater effect of 5 �g of fumagillin onvein length versus artery length (54% and 67%, respec-tively, Fig 6). Relative vessel lengths were also reducedcompared with those in the vehicle control group (P �.05; 70% ethanol). Effects were similar between 2.5and 5 �g of fumagillin (5.5 and 11 nmol, respectively),with the higher dose somewhat more active. Vascula-ture in the fumagillin-treated CAMs was generally re-duced, without dramatical alteration of vessel organi-zation (Fig 7). At higher magnification, the major veinand artery branches of the fumagillin-treated CAMappeared to have shrunk compared with those in thecontrol CAM (Fig 7, C and D). Large artery and vein
Fig 6. Effects of vehicle (70% ethanol) and fumagillin (2.5 and 5 �g) on chicken CAM growth and vein andartery length. Treated values are expressed as a percentage of the control CAM, designated as 100%. CAMs weretreated from day 4 to day 5 of incubation; n � 5 per group. Data expressed as mean �SEM.aP � .05 vs vehicle.
J Lab Clin Med222 Hazel March 2003
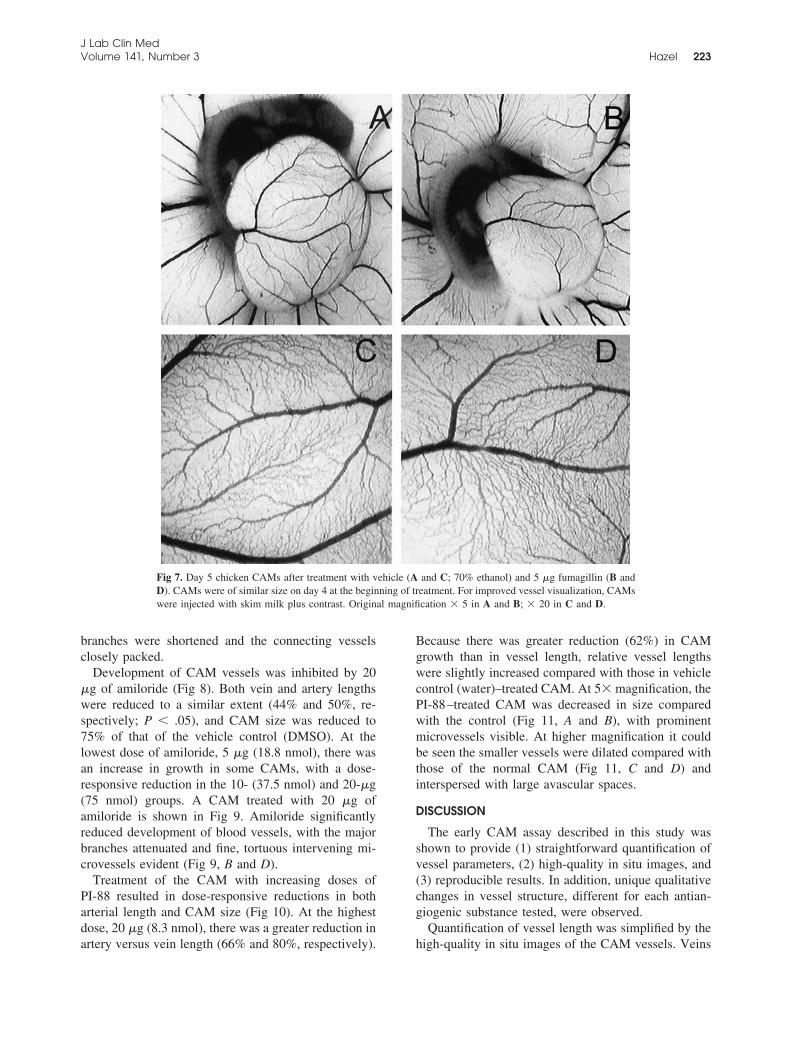
branches were shortened and the connecting vesselsclosely packed.
Development of CAM vessels was inhibited by 20�g of amiloride (Fig 8). Both vein and artery lengthswere reduced to a similar extent (44% and 50%, re-spectively; P � .05), and CAM size was reduced to75% of that of the vehicle control (DMSO). At thelowest dose of amiloride, 5 �g (18.8 nmol), there wasan increase in growth in some CAMs, with a dose-responsive reduction in the 10- (37.5 nmol) and 20-�g(75 nmol) groups. A CAM treated with 20 �g ofamiloride is shown in Fig 9. Amiloride significantlyreduced development of blood vessels, with the majorbranches attenuated and fine, tortuous intervening mi-crovessels evident (Fig 9, B and D).
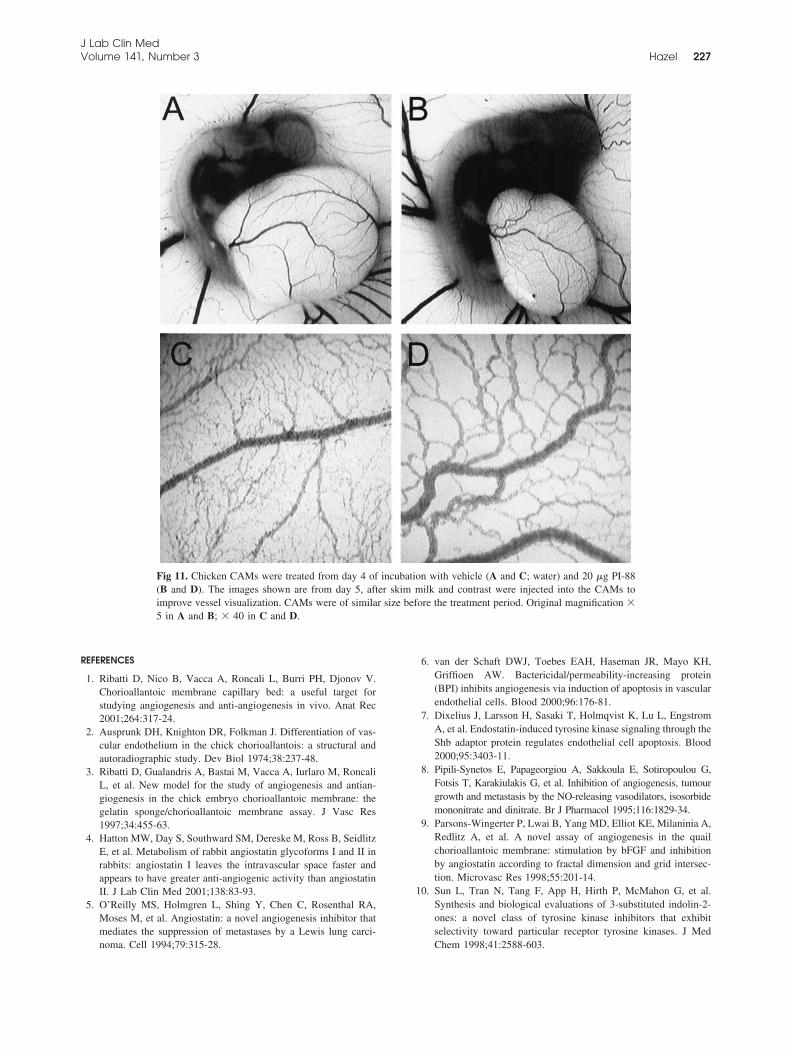
Treatment of the CAM with increasing doses ofPI-88 resulted in dose-responsive reductions in botharterial length and CAM size (Fig 10). At the highestdose, 20 �g (8.3 nmol), there was a greater reduction inartery versus vein length (66% and 80%, respectively).
Because there was greater reduction (62%) in CAMgrowth than in vessel length, relative vessel lengthswere slightly increased compared with those in vehiclecontrol (water)–treated CAM. At 5� magnification, thePI-88–treated CAM was decreased in size comparedwith the control (Fig 11, A and B), with prominentmicrovessels visible. At higher magnification it couldbe seen the smaller vessels were dilated compared withthose of the normal CAM (Fig 11, C and D) andinterspersed with large avascular spaces.
DISCUSSION
The early CAM assay described in this study wasshown to provide (1) straightforward quantification ofvessel parameters, (2) high-quality in situ images, and(3) reproducible results. In addition, unique qualitativechanges in vessel structure, different for each antian-giogenic substance tested, were observed.
Quantification of vessel length was simplified by thehigh-quality in situ images of the CAM vessels. Veins
Fig 7. Day 5 chicken CAMs after treatment with vehicle (A and C; 70% ethanol) and 5 �g fumagillin (B andD). CAMs were of similar size on day 4 at the beginning of treatment. For improved vessel visualization, CAMswere injected with skim milk plus contrast. Original magnification � 5 in A and B; � 20 in C and D.
J Lab Clin MedVolume 141, Number 3 Hazel 223
and arteries were measured separately, making it pos-sible to observe differences in the effects of antiangio-genic substances on vein growth versus artery growth.Artery growth was preferentially reduced by SU5614;conversely, fumagillin reduced vein length to a greaterextent. The early CAM assay was also reproducible,with replicate experiments included for each treatmentgroup and statistically significant results obtained ingroups of five to seven CAMs.
The qualitative changes in vessel structure after an-tiangiogenic treatment were the most unique observa-tions of the early CAM assay. These changes may havebeen permitted by the earlier stage of CAM develop-ment in which the substances were applied (ie, day 4versus day 5 or later) in most published CAM assays.Between days 4.5 and 5 there is an abrupt increase inendothelial permselectivity22 that would reduce macro-molecular access to the endothelium. Hence earliertreatment of the CAM may allow higher local concen-trations of substances in the endothelium versus latertreatment periods. In addition, the high-quality in situimages allowed the qualitative changes to be demon-strated; once the CAM is removed, the smallest vesselsare no longer visible.
Previously published studies involving suramin andamiloride with alternative CAM assay protocols can beused for comparison with the early CAM assay. In arecent study,23 suramin was added to CAMs exposed
by a window in the shell between days 5 and 6. Areasof major vessels were determined with the use of im-ages of glutaraldehyde-fixed CAMs and were reducedto approximately 50% of control with 500 �g ofsuramin, with capillaries also decreased on vesselcounting of histologic sections. In comparison, totalvessel lengths were reduced to 61% of control by 100�g of suramin in this study. Blood-vessel branchingwas reported to be abnormal, but no images of theCAMs were shown, and veins and arteries were com-bined. In another study, agarose discs with suraminwere applied to day 6 CAMs, and the experimentcontinued to day 8.24 Fat emulsion was used to increasecontrast, images of the CAM were captured and mea-surements of gray scale were used to compare vesseldensity. A 35% inhibition in vessel density was re-ported with 50 �g of suramin, with no change in vesselstructure, and veins and arteries were combined. As afurther example, amiloride was added in solution to day6 CAMs exposed by a window in the shell, and imagesof the windowed area were captured on day 7 or 8.17
With the use of image analysis, measurements weremade of vessel density (ie, the number of grid blocksintersected times the linear size of the grid blocks) andfractal dimension, an indication of the degree of com-plexity of the vascular tree. Significant reductions invascular density of up to 67% of control were found;however, there were no images of treated CAMs and no
Fig 8. Effects of vehicle (DMSO) and amiloride (5, 10, and 20 �g) on chicken CAM growth and vein and arterylength. CAMs were treated from day 4 and to day 5 of incubation. Treated values are expressed as a percentageof control, designated as 100%; n � 5 per group. Data expressed as mean � SEM.aP � .05 vs vehicle.
J Lab Clin Med224 Hazel March 2003

qualitative effects on vessel structure, and veins and ar-teries were combined. In addition, 150 �mol of amiloridewas used, compared with 75 nmol in this study, in whichtotal vessel lengths were reduced to 47% of control.
The period between days 4 and 5 is a time of activegrowth and reorganization of the CAM vasculature.Not only do the CAM vessels increase in length, theydifferentiate from irregularly spaced tubules to a morecomplex system of capillaries, arteries, and veins.25 Inaddition, between days 4 and 5 the capillaries migrateup to a position beneath the ectodermal layer of theCAM. Remodeling of blood vessels during angiogene-sis is a poorly understood event that involves thegrowth of new vessels and regression of others. Devel-opment of the mature vascular network requires pre-cisely coordinated growth and migration of endothelialand surrounding cells; depending on their mechanismsof action, antiangiogenic substances could be predictedto disturb different parts of this process. Hence it wasnot surprising in this study that the qualitative changes
caused by the different antiangiogenic substances weredissimilar, because each substance tested had a differ-ent mechanism of action. Thus qualitative changes inthe early CAM assay may predict the mechanism of actionof an unknown substance. However, to determine whetherthis is the case, further studies will be needed to comparequalitative patterns of change in substances with knownsimilarities in their mechanism of action.
Several growth factors are known to regulate angio-genesis in the CAM. VEGF is a relatively specificendothelial growth factor.26 The avian homologue ofVEGFR-2, Quek1, is expressed in the CAM as amarker for endothelial cells,27 and VEGF isoforms arepotent inducers of vascularization and permeability inthe chicken embryo.28 The angiogenic factor bFGF isalso present in the developing CAM,29,30 and bFGFstimulates, whereas bFGF antibodies inhibit, CAM an-giogenesis.30,31 Unlike VEGF, whose activity is rela-tively specific to the endothelial cell, bFGF has effectson several cell types.32 Other stimulators of angiogen-
Fig 9. Day 5 chicken CAMs after treatment with vehicle (A and C) and 20 �g amiloride (B and D). CAMs wereof similar size before treatment on day 4. CAMs were injected with skim milk plus contrast to improvevisualization of the vessels. Original magnification � 5 in A and B; � 20 in C and D.
J Lab Clin MedVolume 141, Number 3 Hazel 225

esis in the CAM include TNF-�,33 lipoprotein a,34 thechemokine I-309,35 and adenosine.36 The presence ofseveral angiogenic pathways on the CAM makes it acomplex model for the study of angiogenesis.
The differential effect of antiangiogenic substanceson vein or arterial growth likely reflects differences insignaling pathways operating in these vessels. An in-creasing number of molecules have been shown to beexpressed at different levels in veins and arteries, in-cluding the Eph/ephrin family,37,38 neuropilin-1 and�2,39 and CX40.40 The greater effect of VEGF inhibi-tion (SU5614) on arterial compared with venousgrowth may be the result of blocking the arterial com-mitment of endothelial cells, as VEGF, produced bySchwann or neuronal cells, increases expression of theartery-specific marker ephrinB2.40
Quantification was based on measurement of vessellength. These measurements probably underestimatedthe effects of the antiangiogenic substances; along withthe reduction in total length, the diameter of the majorvessels was also decreased. Because blood flow isproportional to the fourth power of the vessel’s diam-eter,41 the differences in blood flow between the treatedand control CAMs would be greater than differences inthe vessel length. In the CAMs in which the vesselswere highly tortuous (eg, those treated with suramin),blood flow was noticeably slowed.
The VEGF receptor inhibitor SU5614 had effects
only on CAM vasculature, whereas the other sub-stances tested also resulted in reductions in overallCAM size. It is possible that these other substancesdecreased CAM membrane growth and only indirectlyreduced growth of CAM vessels. However, each of thesubstances tested has been previously shown to haveantiangiogenic effects.10,16,18,42,43 Furthermore, angio-genesis shares signaling pathways with processes suchas hematopoiesis and inflammation, and angiogenicstimulators such as bFGF and adenosine also have effectson nonendothelial cells.32 The early CAM assay may noteasily determine whether a substance is inhibiting angio-genesis directly through endothelial cells or indirectlythrough other cell types. However, this is the case withany in vivo assay in which many cell types are present.
The early CAM assay will be a useful addition to theassays available for screening antiangiogenic agents.Information was obtained that had not been availablethrough traditional CAM assays, including differentialartery- and vein-length measurements and qualitativechanges in vessel structure. Future studies to define thequalitative changes arising from various antiangiogenicagents in the early CAM assay may be useful in increasingour understanding of the angiogenic process itself.
I thank Julianna Balogh and Andrea Talbot for their excellenttechnical assistance and Denis Podger for his helpful criticisms of themanuscript.
Fig 10. Effects of PI-88 (5, 10, and 20 �g) on chicken CAM growth and vein and artery length. CAMs weretreated from day 4 to day 5 of incubation. Results are expressed as a percentage of the vehicle control (water), designatedas 100%; n � 5 per group. Data expressed as mean � SEM.aP � .05 vs vehicle.bP � .05 vs 5 �g.cP � .05 vs 10 �g.
J Lab Clin Med226 Hazel March 2003
REFERENCES
1. Ribatti D, Nico B, Vacca A, Roncali L, Burri PH, Djonov V.Chorioallantoic membrane capillary bed: a useful target forstudying angiogenesis and anti-angiogenesis in vivo. Anat Rec2001;264:317-24.
2. Ausprunk DH, Knighton DR, Folkman J. Differentiation of vas-cular endothelium in the chick chorioallantois: a structural andautoradiographic study. Dev Biol 1974;38:237-48.
3. Ribatti D, Gualandris A, Bastai M, Vacca A, Iurlaro M, RoncaliL, et al. New model for the study of angiogenesis and antian-giogenesis in the chick embryo chorioallantoic membrane: thegelatin sponge/chorioallantoic membrane assay. J Vasc Res1997;34:455-63.
4. Hatton MW, Day S, Southward SM, Dereske M, Ross B, SeidlitzE, et al. Metabolism of rabbit angiostatin glycoforms I and II inrabbits: angiostatin I leaves the intravascular space faster andappears to have greater anti-angiogenic activity than angiostatinII. J Lab Clin Med 2001;138:83-93.
5. O’Reilly MS, Holmgren L, Shing Y, Chen C, Rosenthal RA,Moses M, et al. Angiostatin: a novel angiogenesis inhibitor thatmediates the suppression of metastases by a Lewis lung carci-noma. Cell 1994;79:315-28.
6. van der Schaft DWJ, Toebes EAH, Haseman JR, Mayo KH,Griffioen AW. Bactericidal/permeability-increasing protein(BPI) inhibits angiogenesis via induction of apoptosis in vascularendothelial cells. Blood 2000;96:176-81.
7. Dixelius J, Larsson H, Sasaki T, Holmqvist K, Lu L, EngstromA, et al. Endostatin-induced tyrosine kinase signaling through theShb adaptor protein regulates endothelial cell apoptosis. Blood2000;95:3403-11.
8. Pipili-Synetos E, Papageorgiou A, Sakkoula E, Sotiropoulou G,Fotsis T, Karakiulakis G, et al. Inhibition of angiogenesis, tumourgrowth and metastasis by the NO-releasing vasodilators, isosorbidemononitrate and dinitrate. Br J Pharmacol 1995;116:1829-34.
9. Parsons-Wingerter P, Lwai B, Yang MD, Elliot KE, Milaninia A,Redlitz A, et al. A novel assay of angiogenesis in the quailchorioallantoic membrane: stimulation by bFGF and inhibitionby angiostatin according to fractal dimension and grid intersec-tion. Microvasc Res 1998;55:201-14.
10. Sun L, Tran N, Tang F, App H, Hirth P, McMahon G, et al.Synthesis and biological evaluations of 3-substituted indolin-2-ones: a novel class of tyrosine kinase inhibitors that exhibitselectivity toward particular receptor tyrosine kinases. J MedChem 1998;41:2588-603.
Fig 11. Chicken CAMs were treated from day 4 of incubation with vehicle (A and C; water) and 20 �g PI-88(B and D). The images shown are from day 5, after skim milk and contrast were injected into the CAMs toimprove vessel visualization. CAMs were of similar size before the treatment period. Original magnification �5 in A and B; � 40 in C and D.
J Lab Clin MedVolume 141, Number 3 Hazel 227

11. Waltenberger J, Mayr U, Frank H, Hombach V. Suramin is apotent inhibitor of vascular endothelial growth factor: a contri-bution to the molecular basis of its antiangiogenic action. J MolCell Cardiol 1996;28:1523-9.
12. Zugmaier G, Favoni R, Jaeger R, Rosen N, Knabbe C. Polysul-fated heparinoids selectively inactivate heparin-binding angio-genesis factors. Ann N Y Acad Sci 1999;886:243-8.
13. Mahoney CW, Azzi A, Huang KP. Effects of suramin, an anti-human immunodeficiency virus reverse transcriptase agent, onprotein kinase C: differential activation and inhibition of proteinkinase C isozymes. J Biol Chem 1990;265:5424-8.
14. Nakajima M, DeChavigny A, Johnson CE, Hamada J, Stein CA,Nicolson GL. Suramin: a potent inhibitor of melanoma hepara-nase and invasion. J Biol Chem 1991;266:9661-6.
15. Abe J, Zhou W, Takuwa N, Taguchi J, Kurokawa K, Kumada M,et al. A fumagillin derivative angiogenesis inhibitor, AGM-1470,inhibits activation of cyclin-dependent kinases and phosphoryla-tion of retinoblastoma gene product but not protein tyrosyl phos-phorylation or protooncogene expression in vascular endothelialcells. Cancer Res 1994;54:3407-12.
16. Sin N, Meng L, Wang MQ, Wen JJ, Bornmann WG, Crews CM.The anti-angiogenic agent fumagillin covalently binds and inhib-its the methionine aminopeptidase, MetAP-2. Proc Natl Acad SciU S A 1997;94:6099-103.
17. Knoll A, Schmidt S, Chapman M, Wiley D, Bulgrin J, Blank J,et al. A comparison of two controlled-release delivery systemsfor the delivery of amiloride to control angiogenesis. MicrovascRes 1999;58:1-9.
18. Parish CR, Freeman C, Brown KJ, Francis DJ, Cowden WB.Identification of sulfated oligosaccharide-based inhibitors of tu-mor growth and metastasis using novel in vitro assays for an-giogenesis and heparanase activity. Cancer Res 1999;59:3433-41.
19. Dugan JD, Lawton MT, Glaser B, Brem H. A new technique forexplanation and in vitro cultivation of chicken embryos. AnatRec 1991;229:125-8.
20. Dunn BE, Fitzharris TP, Barnett BD. Effects of varying chamberconstruction and embryo pre-incubation age on survival andgrowth of chick embryos in shell-less culture. Anat Rec 1981;199:33-43.
21. Sells PG, Richards AM, Laing GD, Theakston RD. The use ofhens’ eggs as an alternative to the conventional in vivo rodentassay for antidotes to haemorrhagic venoms. Toxicon 1997;35:1413-21.
22. Rizzo V, Kim D, Duran WN, DeFouw DO. Ontogeny of micro-vascular permeability to macromolecules in the chick cho-rioallantoic membrane during normal angiogenesis. MicrovascRes 1995;49:49-63.
23. Melkonian G, Munoz N, Chung J, Tong C, Marr R, Talbot P.Capillary plexus development in the day five to day six chickchorioallantoic membrane is inhibited by cytochalasin D andsuramin. J Exp Zool 2002;292:241-54.
24. Danesi R, Del BD, Soldani P, Campagni A, La RRV, Myers CE,et al. Suramin inhibits bFGF-induced endothelial cell prolifera-tion and angiogenesis in the chick chorioallantoic membrane.Br J Cancer 1993;68:932-8.
25. Ausprunk DH. Distribution of hyaluronic acid and sulfated gly-cosaminoglycans during blood-vessel development in the chickchorioallantoic membrane. Am J Anat 1986;177:313-31.
26. Veikkola T, Karkkainen M, Claesson-Welsh L, Alitalo K. Reg-ulation of angiogenesis via vascular endothelial growth factorreceptors. Cancer Res 2000;60:203-12.
27. Wilting J, Eichmann A, Christ B. Expression of the avian VEGFreceptor homologues Quek1 and Quek2 in blood-vascular andlymphatic endothelial and non-endothelial cells during quailembryonic development. Cell Tissue Res 1997;288:207-23.
28. Schmidt M, Flamme I. The in vivo activity of vascular endothe-lial growth factor isoforms in the avian embryo. Growth Factors1998;15:183-97.
29. Flamme I, Schulze-Osthoff K, Jacob HJ. Mitogenic activity ofchicken chorioallantoic fluid is temporally correlated to vasculargrowth in the chorioallantoic membrane and related to fibroblastgrowth factors. Development 1991;111:683-90.
30. Ribatti D, Urbinati C, Nico B, Rusnati M, Roncali L, Presta M.Endogenous basic fibroblast growth factor is implicated in thevascularization of the chick embryo chorioallantoic membrane.Dev Biol 1995;170:39-49.
31. Parsons-Wingerter P, Elliot KE, Clark JI, Farr AG. Fibroblastgrowth factor-2 selectively stimulates angiogenesis of small ves-sels in arterial tree. Arterioscler Thromb Vasc Biol 2000;20:1250-6.
32. Bikfalvi A, Klein S, Pintucci G, Rifkin DB. Biological roles offibroblast growth factor-2. Endocr Rev 1997;18:26-45.
33. Olivo M, Bhardwaj R, Schulze-Osthoff K, Sorg C, Jacob HJ,Flamme I. A comparative study on the effects of tumor necrosisfactor-a (TNF-a), human angiogenic factor (h-AF) and basicfibroblast growth factor (bFGF) on the chorioallantoic membraneof the chick embryo. Anat Rec 1992;234:105-15.
34. Ribatti D, Vacca A, Giacchetta F, Cesaretti S, Anichini M,Roncali L, et al. Lipoprotein (a) induces angiogenesis on thechick embryo chorioallantoic membrane. Eur J Clin Invest 1998;28:533-7.
35. Bernardini G, Spinetti G, Ribatti D, Camarda G, Morbidelli L,Ziche M, et al. I-309 binds to and activates endothelial cellfunctions and acts as an angiogenic molecule in vivo. Blood2000;96:4039-45.
36. Dusseau JW, Hutchins PM, Malbasa DS. Stimulation of angio-genesis by adenosine on the chick chorioallantoic membrane.Circ Res 1986;59:163-70.
37. Adams RH, Wilkinson GA, Weiss C, Diella F, Gale NW, Deut-sch U, et al. Roles of ephrinB ligands and EphB receptors incardiovascular development: demarcation of arterial/venous do-mains, vascular morphogenesis, and sprouting angiogenesis.Gene Dev 1999;13:295-306.
38. Wang HU, Chen Z-F, Anderson DJ. Molecular distinction andangiogenic interaction between embryonic arteries and veinsrevealed by ephrin-B2 and its receptor Eph-B4. Cell 1998;93:741-53.
39. Herzog Y, Kalcheim C, Kahane N, Reshef R, Neufeld G. Dif-ferential expression of neuropilin-1 and neuropilin-2 in arteriesand veins. Mech Dev 2001;109:115-9.
40. Mukouyama Y, Shin D, Britsch S, Taniguchi M, Anderson DJ.Sensory nerves determine the pattern of arterial differentiationand blood vessel branching in the skin. Cell 2002;109:693-705.
41. Guyton AC. Physics of blood: blood flow, and pressure: Hemo-dynamics. In: Guyton AC, Hall JE, eds. Textbook of medicalphysiology. Philadelphia: Saunders, 1981:206-18.
42. Meyers MO, Gagliardi AR, Flattmann GJ, Su JL, Wang Y-Z,Woltering EA. Suramin analogs inhibit human angiogenesis inVitro. J Surg Res 2000;91:130-4.
43. Alliegro MC, Alliegro MA, Cragoe EJ Jr, Glaser BM. Amilorideinhibition of angiogenesis in vitro. J Exp Zool 1993;267:245-52.
J Lab Clin Med228 Hazel March 2003




















