![[Revised] Revisiting Verb Aspect in T'boli](https://static.fdokumen.com/doc/165x107/631ef9e50ff042c6110c9f71/revised-revisiting-verb-aspect-in-tboli.jpg)
A novel aspect of lindane testicular toxicity: in vitro effects on peritubular myoid cells
11
A novel aspect of lindane testicular toxicity: in vitro effects on peritubular myoid cells Leopoldo Silvestroni*, Fabio Rossi, Massimo Magnanti, Carla Lubrano, Vittorio Santiemma, Simonetta Palleschi Department of Medical Physiopathology, University of Rome ‘La Sapienza’, Viale del Policlinico, 00161, Rome, Italy Received 17 April 1999; accepted 17 June 1999 Abstract The in vitro effects of the insecticide lindane have been investigated in rat testis peritubular myoid cells (PMCs). Upon PMC exposure to lindane, polarity increase and decrease of dipole dynamics were seen at the membrane level (EC50 20 mM), leading to a partial dissipation of the membrane intrinsic dipole potential. The initial membrane depolarization was increased by Cl 2 efflux and limited by Ca 21 -activated repolarizing currents. Concomitantly, lindane produced an increase in [Ca 21 ] i (EC50 125 mM) resulting from both Ca 21 release from an inositol 1,4,5-trisphosphate-sensitive intracellular store and a voltage-dependent Ca 21 influx from the extracellular medium. Of particular interest from a toxicologic point of view, insecticide concentrations well below those effective in altering ion homeostasis potently inhibited both [Ca 21 ] i increase and contraction induced by the natural agonists vasopressin and endothelin-1 (IC50s , 10 mM). These data demonstrate that PMCs are highly susceptible to lindane and suggest that the insecticide may exert testicular toxicity by interfering with hormone-regulated PMC function. © 1999 Elsevier Science Inc. All rights reserved. Keywords: Lindane; Membrane fluidity; Intracellular calcium; Membrane potential; Peritubular myoid cells; Contraction; Vasopressin; Endothelin-1 1. Introduction It has been known for decades that some synthetic chem- icals can alter reproductive function. This property is shared by many toxicants such as pesticides [1], dioxins [2], poly- chlorinated biphenyls [3], phthalates [4], alkylphenols [5], and bisphenol A [6]. Different reproductive toxicants act in cells through different mechanisms. Some bind to steroid receptors and alter the cell responses to natural agonists [7], others bind to the cytoplasmic arhylhydrocarbon receptor [8] and activate the dioxin responsive element on DNA [9], still others activate phospholipases [10] or mimic inositol- 1,4,5-trisphosphate (IP 3 ) [11], thereby activating compo- nents of the intracellular signal transduction pathways. Even low doses of some agents during the fetal life may be detrimental to male fertility in adulthood [12]. Based on a meta analysis applied to a large number of human semen data, a 50% reduction in sperm production over the last 50 years has been reported [13], supposedly accounted for in part by prenatal exposure to adverse environmental factors [14]. However, the reported evidence that semen quality is decreasing has been criticized for inconsistency with other data [15,16] or methodologic inadequacies [17]. Lindane, the g-isomer of hexachlorocyclohexane, is an organohalogenated pesticide. The parasiticidal activity of lindane lies in its blocking effect on GABA receptor-linked chloride channels [18]. Chronic lindane treatment with 10 to 40 mg/kg exerts antiestrogenic activity in female rats as evidenced by delayed vaginal opening, disrupted estrous cycling, and reduced uterine weight [19]. In the mouse ovary, the insecticide affects cholesterol side-chain cleav- age as judged by decreased conversion of cholesterol to progesterone [20], while in yeast expressing human proges- terone receptor, lindane inhibits the transcriptional activity induced by progesterone [21]. The male gonad is highly susceptible to lindane. After oral administration to adult rats of 6 mg/kg b.wt. for 5 d or 30 mg/kg b.wt. as a single dose, lindane crosses the blood-testis barrier and depresses sper- matogenesis, as evidenced by a numeric reduction in sper- matids and fragmentation of Sertoli cells [22]. In the same * Corresponding author. Tel.: 139-06-49970710; fax: 139-06-490530. E-mail address: [email protected] (L. Silvestroni) Reproductive Toxicology 13 (1999) 431– 441 0890-6238/99/$ – see front matter © 1999 Elsevier Science Inc. All rights reserved. PII: S0890-6238(99)00040-4
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A novel aspect of lindane testicular toxicity: in vitro effects on peritubular myoid cells

A novel aspect of lindane testicular toxicity: in vitro effects onperitubular myoid cells
Leopoldo Silvestroni*, Fabio Rossi, Massimo Magnanti, Carla Lubrano, Vittorio Santiemma,Simonetta Palleschi
Department of Medical Physiopathology, University of Rome ‘La Sapienza’, Viale del Policlinico, 00161, Rome, Italy
Received 17 April 1999; accepted 17 June 1999
Abstract
The in vitro effects of the insecticide lindane have been investigated in rat testis peritubular myoid cells (PMCs). Upon PMC exposureto lindane, polarity increase and decrease of dipole dynamics were seen at the membrane level (EC50 20mM), leading to a partial dissipationof the membrane intrinsic dipole potential. The initial membrane depolarization was increased by Cl2 efflux and limited by Ca21-activatedrepolarizing currents. Concomitantly, lindane produced an increase in [Ca21]i (EC50 125mM) resulting from both Ca21 release from aninositol 1,4,5-trisphosphate-sensitive intracellular store and a voltage-dependent Ca21 influx from the extracellular medium. Of particularinterest from a toxicologic point of view, insecticide concentrations well below those effective in altering ion homeostasis potently inhibitedboth [Ca21]i increase and contraction induced by the natural agonists vasopressin and endothelin-1 (IC50s, 10 mM). These datademonstrate that PMCs are highly susceptible to lindane and suggest that the insecticide may exert testicular toxicity by interfering withhormone-regulated PMC function. © 1999 Elsevier Science Inc. All rights reserved.
Keywords:Lindane; Membrane fluidity; Intracellular calcium; Membrane potential; Peritubular myoid cells; Contraction; Vasopressin; Endothelin-1
1. Introduction
It has been known for decades that some synthetic chem-icals can alter reproductive function. This property is sharedby many toxicants such as pesticides [1], dioxins [2], poly-chlorinated biphenyls [3], phthalates [4], alkylphenols [5],and bisphenol A [6]. Different reproductive toxicants act incells through different mechanisms. Some bind to steroidreceptors and alter the cell responses to natural agonists [7],others bind to the cytoplasmic arhylhydrocarbon receptor[8] and activate the dioxin responsive element on DNA [9],still others activate phospholipases [10] or mimic inositol-1,4,5-trisphosphate (IP3) [11], thereby activating compo-nents of the intracellular signal transduction pathways. Evenlow doses of some agents during the fetal life may bedetrimental to male fertility in adulthood [12]. Based on ameta analysis applied to a large number of human semendata, a 50% reduction in sperm production over the last 50
years has been reported [13], supposedly accounted for inpart by prenatal exposure to adverse environmental factors[14]. However, the reported evidence that semen quality isdecreasing has been criticized for inconsistency with otherdata [15,16] or methodologic inadequacies [17].
Lindane, theg-isomer of hexachlorocyclohexane, is anorganohalogenated pesticide. The parasiticidal activity oflindane lies in its blocking effect on GABA receptor-linkedchloride channels [18]. Chronic lindane treatment with 10 to40 mg/kg exerts antiestrogenic activity in female rats asevidenced by delayed vaginal opening, disrupted estrouscycling, and reduced uterine weight [19]. In the mouseovary, the insecticide affects cholesterol side-chain cleav-age as judged by decreased conversion of cholesterol toprogesterone [20], while in yeast expressing human proges-terone receptor, lindane inhibits the transcriptional activityinduced by progesterone [21]. The male gonad is highlysusceptible to lindane. After oral administration to adult ratsof 6 mg/kg b.wt. for 5 d or 30 mg/kg b.wt. as a single dose,lindane crosses the blood-testis barrier and depresses sper-matogenesis, as evidenced by a numeric reduction in sper-matids and fragmentation of Sertoli cells [22]. In the same
* Corresponding author. Tel.:139-06-49970710; fax:139-06-490530.E-mail address:[email protected] (L. Silvestroni)
Reproductive Toxicology 13 (1999) 431–441
0890-6238/99/$ – see front matter © 1999 Elsevier Science Inc. All rights reserved.PII: S0890-6238(99)00040-4

animal model, a 50% reduction in circulating testosteronehas been reported in adults exposed to lindane during lac-tation [23], suggesting that the perturbation of intratesticularsteroidogenesis by lindane is minimally or not reversible.
Lindane produces several nonreproductive effects. In fi-broblasts and hepatocytes cultured with lindane at 1.7 to 170mM for 24 h, there is enhanced free radical productionultimately leading to cell transformation and oxidativestress [24]. In rat brain cortex [25], renal tubule cultures[10], and mouse peritoneal macrophages [26], 50 to 500mMlindane acutely stimulates IP3 and arachidonic acid produc-tion by activating membrane phospholipases. In myometrialmyocytes exposure to lindane at 10mM for 4 min theinsecticide severs gap-junctional communications [27] andat 1 to 200mM for 6 min alters Ca21 homeostasis and mayperturb Ca21-regulated stimulus-contraction coupling invivo, leading to an increased probability of delayed ordysfunctional labor in exposed individuals [28]. Interest-ingly, due to its structural similarity to IP3, lindane mayrelease Ca21 from IP3-sensitive intracellular stores and dis-regulate Ca21-dependent events [29].
A high degree of hydrophobicity allows lindane to easilyintercalate into the phospholipid bilayer [30]. When thebiophysical aspects of lindane-cell interaction were inves-tigated, increased membrane fluidity was measured in cellmembranes from kidney cortex/tubules and ventral prostate[31], whereas no membrane perturbation was detected in ratenterocytes [32]. Quite different results were obtained insynthetic bilayers in the fluid phase, where lindane wasfound to increase the rigidity of lipid acyl chains [33]. Inagreement with the latter finding, we recently found thatlindane lowers the molecular dynamics in the membrane ofthe uppermost region of living human spermatozoa [34]. Inthe same report, evidence was provided that lindane altersthe intrinsic membrane dipole potential and induces calciuminflux through voltage-operated Ca21 channels, an array ofseemingly nonreceptor-mediated effects likely stemmingfrom lindane partitioning in the sperm surface.
To get more insight into the mechanism of lindane go-nadal toxicity, the present investigation was carried out toevaluate lindane effects on peritubular myoid cells (PMCs),the smooth muscle cell layer embedded in the interstitialmatrix surrounding the seminiferous tubules. In the testis,peristaltic PMC contraction participates in the outwardsperm progression within the tubular lumen. A number ofnatural agonists (vasopressin, TGF-b, PDGF, endothelin-1)acting through receptor-mediated mechanisms cooperate inregulating PMC motor activity [35]. Central to stimulus-contraction coupling, a transient increase in [Ca21]i drivessignal transduction for all the peptides cited. For example,endothelin-1 produces contraction in PMCs by increasing[Ca21]i through a Ca21 influx from extracellular mediumcoupled to a Ca21 release from inositol-3-phosphate-sensi-tive intracellular stores [36].
Recent in vitro studies demonstrated that PMCs secrete anumber of extracellular matrix components (e.g., type I and
IV collagens, fibronectin, and proteoglycans) and growthfactors (e.g., activin-A, IGF-I, and PModS) [35], some ofwhich are known to affect Sertoli cell differentiation [37]and function [38]. Hence, PMCs can rightly be regarded asintratesticular regulators of spermatogenesis [39].
We report here that lindane interaction with PMCs bringsabout an array of effects consisting of an increase of polarityand a decrease of dipole dynamics of the bilayer, membranedepolarization, and an increase in [Ca21]i, and that sucheffects occur at insecticide doses similar to intratesticularconcentrations measured in lindane exposed rats [22]. Ofparticular toxicologic interest, in the present study doses oflindane too low to evoke apparent effects in PMCs, may behigh enough to transiently inhibit [Ca21]i and contractioninduced by vasopressin (AVP) and endothelin-1 (ET-1),two key hormones involved in the intragonadal control ofspermatogenesis.
2. Materials and methods
2.1. Chemicals
Prodan (2-dimethylamino-6-propionylnaphthalene) wasobtained from Molecular Probes (Eugene, OR, USA). Fura-2-AM, bisoxonol, ET-1, AVP, lindane (g-isomer of hexa-chlorocyclohexane, 99% purity), calf skin type I collagen,HEPES, EGTA, digitonin, neomycin, 4-bromophenacylbromide (BPB), nicardipine, and porcine pancreatic trypsinwere purchased from Sigma Chemical Co. (St. Louis, MO,USA). Type A collagenase (lot 83687025) was from Boehr-inger (Mannheim, Germany). DMEM, F12, and fetal calfserum (FCS) were purchased from Gibco (Grand Island,NY, USA). All other reagents were of the highest puritycommercially available. The buffer used in this study con-sisted of 30 mM HEPES, 1.19 mM MgSO4z7H2O, 5 mMKCl, 122 mM NaCl, 1.5 mM CaCl2, and 5.5 mMD-glucose,pH 7.4 (regular buffer). Na1-free and Cl2-free buffers wereprepared by equimolar substitution of sodium and chloridesalts with choline (choline buffer) and gluconate (gluconatebuffer) salts, respectively. Ca21-free buffers were preparedby omitting CaCl2 (Ca21-free buffer) and adding 2.5 mMEGTA to Ca21-free buffer (EGTA buffer).
2.2. PMC preparation
Peritubular myoid cells were prepared and cultured asdescribed previously [36]. Briefly, decapsulated testes from18-d old Sprague–Dawley rats (Charles River Italia, Calco,Italy) were sequentially digested with 0.25% trypsin and0.1% collagenase. PMCs were purified by differential cen-trifugation on a Percoll discontinuous gradient, washedtwice, and plated. Cell viability and purity were evaluatedby using a trypan blue exclusion method [40] and indirectimmunofluorescence labeling with desmine and vimentine[41], respectively. Cell viability and purity were always
432 L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

.95% and did not vary to a significant extent in the courseof the experiments. Primary cultures of PMCs were main-tained at 32°C in a 5% CO2 atmosphere in DMEM/F12(1 : 1) containing 10% FCS. For [Ca21]i and membranepotential measurements, suspensions of PMCs cultured for3 to 5 d were prepared by a brief trypsin/EDTA treatment(0.025% trypsin, 0.02% EDTA), washed twice, and main-tained at room temperature in regular buffer. Membranebiophysical properties were evaluated in PMCs grown onglass coverslips. Just before measurements, the coverslipswere freed of culture medium by dipping twice in regularbuffer.
2.3. Fluorescence spectroscopy
Fluorescence measurements were carried out with acomputer-driven L-type spectrofluorometer (KontronSFM25) upon continuous stirring (200 rev./min) at 37°C(60.1C). Reading cuvettes (acryl 67.755, Sarstedt) con-tained 1.5 mL buffer. Output signals were collected, stored,and elaborated by using dedicated computer programs de-veloped by one of us (S.P.). Membrane polarity and molec-ular dynamics were investigated by exploiting the spectro-scopic properties of Prodan, a lipophilic fluorescentmembrane probe extremely susceptible to both the numberand kinetics of surrounding water molecules [42]. [Ca21]i
and membrane potential were evaluated by using the fluo-rescent probes fura-2 and bisoxonol, respectively. For inhi-bition experiments, PMCs were preincubated for 5 min inthe presence of either 4mM BPB, a phospholipase A2inhibitor [43], or 2 mM neomycin, a phosphatidylinositolbreakdown inhibitor [44], 50mM picrotoxin, a blocker ofGABA receptor-linked chloride channels [45] or 5mMnicardipine, an inhibitor of voltage-operated calcium chan-nels (VOCC) [46]. Fluorescence readings were performedin the continuous presence of the selected inhibitor. Prelim-inary experiments showed that the above substances did notinterfere with either cell viability or spectroscopic readings.Lindane from stock solutions in DMSO was added to PMCsuspensions to obtain the selected final concentrations(DMSO never exceeded 0.2%). Best line fits to the exper-imental data were obtained by using the computer programTableCurve2D (Jandel Scientific, Erkrath, Germany), andthe biologically relevant parameters were estimated visuallyfrom the graphs.
2.4. Membrane polarity and molecular dynamicsevaluation.
PMCs were loaded (37°C, 20 min) with Prodan by add-ing 1 mL of probe stock solution (0.175 mM in DMSO,0.117 mM final concentration). Fluorescence excitation(from 320 nm to 420 nm) and emission (from 410 nm to 540nm) spectra were collected at an emission of 490 nm and atan excitation of 370 nm, respectively, over a 30 min periodat 2 min intervals and elaborated in terms of generalized
polarization (GPem and GPex) as previously described [42].Lindane from a stock solution in DMSO was added tolabeled PMCs at various concentrations (from 10mM to 200mM; final DMSO never exceeded 0.1%). Control samples(no lindane added) were treated with 0.1% DMSO.
2.5. [Ca21] i determination.
PMCs suspended in regular buffer (53 106 cells/mL)were loaded with the fluorescent calcium chelator fura-2-AM (1.5 mL/mL of a 2 mMstock solution in DMSO; finalprobe concentration 3mM) for 45 min at 37°C. After wash-ing, probe de-esterification was allowed to proceed for 15min at 37°C; afterwards, the cells were washed and main-tained in regular buffer at room temperature in the dark untiluse (max 2.5 h from loading). In order to reduce fura-2leakage and compartmentalization in loaded cells, the ex-periments were conducted in the continuous presence of 250mM sulfinpyrazone, an inhibitor of organic anion transport[47]. Just before measurements, 100-mL aliquots (63 105
cells) were washed once in the selected buffer and trans-ferred to the reading cuvette. Fluorescence measurementswere carried out at 340/505 nm excitation/emission andconverted into nM Ca21 [48] assuming an equilibrium dis-sociation constant of 224 nM for the Ca21-fura-2 complex.Minimal and maximal fluorescence were obtained in theabsence of free Ca21 (addition of 5 mM EGTA after celllysis with 30mM digitonin) and in the presence of saturat-ing Ca21 (addition of 10 mM CaCl2), respectively. Thelindane effect on AVP- (100 nM) and ET-1- (4 nM) induced[Ca21]i increase was evaluated by preincubating PMCs inthe presence of 0 (solvent alone) to 50mM lindane for 30min; fluorescence readings were collected in the continuouspresence of the lindane and Student’s unpairedt-test wasused for the statistical analysis of data. Concentrations wereselected to produce maximal Ca21 response, as assessed inpreliminary experiments (data not shown).
2.6. Membrane potential
The potential sensitive fluorescent dye bisoxonol wasadded to the cell suspension (63 105 cells) into the fluoro-metric cuvette at 200 nM final concentration (1.5mL of a200 mM stock solution in DMSO). After 2 to 4 min incu-bation, the time necessary to allow transmembrane probeequilibration, readings were collected at 540/580 nm exci-tation/emission and fluorescence intensity values were ex-pressed in arbitrary units.
2.7. PMC contraction assay.
PMC contraction was evaluated by using a method pre-viously described [49], with slight modifications. Briefly,after 6 to 8 d of culture, cells were detached by trypsiniza-tion (see PMC preparation) and suspended in MEM con-taining 1.6 mg/mL type I collagen at a final density of 43
433L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

105 cells/mL. Aliquots of 0.5 mL were transferred into24-multiwell tissue culture plates previously coated with0.67% agarose and cultured overnight (37°C in a 5% CO2
atmosphere). Lindane from prewarmed 53 stock solutionsin MEM was added by partial substitution of PMC incuba-tion medium to obtain 0 (solvent alone), 1, 5, 25, and 50mMfinal concentrations. After 30 min, ET-1 or AVP were addedfrom 5003 stock solutions in water to obtain 4 and 100 nMfinal concentrations, respectively. Nothing was added tocontrol wells. Collagen contraction was evaluated after 3 hincubation, a time necessary for the phenomenon to becompleted. The plates were videotaped (Screen Machine II,FAST Multimedia AG, Munchen, Germany), the areas ofthe collagen discs were calculated from the digitized im-ages, (SigmaScan Pro, Jandel Scientific, Erkrath, Germany)and contraction (c) computed according to the formula:
c 5~ A 2 a!
A(1)
wherea is the area of the collagen disc andA is the well’sarea. Student’s unpairedt-test was used for the statisticalanalysis of data.
3. Results
3.1. Effects of lindane on membrane biophysicalproperties
Prodan was taken up by PMCs in a time-dependentmanner, with an apparent steady state of uptake attained at20 min. Prodan fluorescence excitation and emission spectracollected in unstimulated PMCs are shown in Fig. 1. Theexcitation spectrum shows a single emission peak centeredat 370 nm, and the emission spectrum shows two emission
peaks centered at 435 nm and 490 nm, respectively. Thisspectroscopic behavior is indicative of a highly orderedliquid-crystalline lipid phase and is typical of cholesterol-rich phospholipid bilayers above their transition tempera-ture [50]. From the curves, GPex and GPemvalues of 0.4060.02 and 0.216 0.03 were derived, respectively. Lindaneincreased both GPex and GPem, with maximal values at-tained about 7 min after addition (Fig. 2). From the dose-response curves, EC50s of 20 and 30mM were calculated forGPex and GPem, respectively (Fig. 3).PMC preincubationwith BPB or neomycin had no effect on lindane-induced GPmodification (not shown), meaning that the observed vari-ations of membrane fluidity did not depend upon phospho-lipid degradation by either PLA2 or PLC, respectively.From these findings it may be inferred that the reduction ofdipole kinetics (as mirrored by the GPex increase) and theincrease of membrane polarity (as mirrored by the GPem
increase) primarily depend upon lindane partitioning at thewater-lipid interface of the bilayer.
Fig. 1. Fluorescence excitation and emission spectra of Prodan-loadedPMCs. Tracings are the best line fits to the experimental data and arerepresentative of at least four experiments. a.u.5 arbitrary units.
Fig. 2. Time course of GPem (panel a) and GPex (panel b) values ofProdan-loaded PMCs in the absence (only DMSO, open symbols) orpresence of 150mM lindane (closed symbols). The traces are the best linefits to the experimental data and are representative of at least four consis-tent experiments. Arrow5 lindane or DMSO addition.
434 L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441
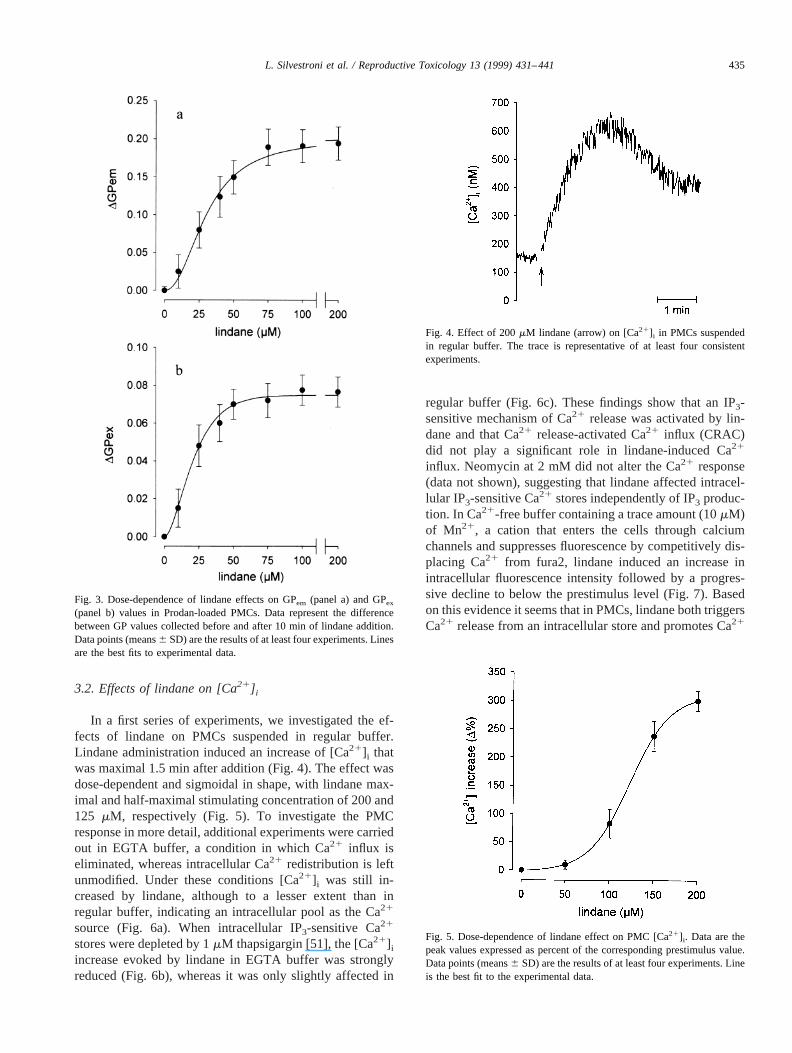
3.2. Effects of lindane on [Ca21] i
In a first series of experiments, we investigated the ef-fects of lindane on PMCs suspended in regular buffer.Lindane administration induced an increase of [Ca21]i thatwas maximal 1.5 min after addition (Fig. 4). The effect wasdose-dependent and sigmoidal in shape, with lindane max-imal and half-maximal stimulating concentration of 200 and125 mM, respectively (Fig. 5). To investigate the PMCresponse in more detail, additional experiments were carriedout in EGTA buffer, a condition in which Ca21 influx iseliminated, whereas intracellular Ca21 redistribution is leftunmodified. Under these conditions [Ca21]i was still in-creased by lindane, although to a lesser extent than inregular buffer, indicating an intracellular pool as the Ca21
source (Fig. 6a). When intracellular IP3-sensitive Ca21
stores were depleted by 1mM thapsigargin [51], the [Ca21]i
increase evoked by lindane in EGTA buffer was stronglyreduced (Fig. 6b), whereas it was only slightly affected in
regular buffer (Fig. 6c). These findings show that an IP3-sensitive mechanism of Ca21 release was activated by lin-dane and that Ca21 release-activated Ca21 influx (CRAC)did not play a significant role in lindane-induced Ca21
influx. Neomycin at 2 mM did not alter the Ca21 response(data not shown), suggesting that lindane affected intracel-lular IP3-sensitive Ca21 stores independently of IP3 produc-tion. In Ca21-free buffer containing a trace amount (10mM)of Mn21, a cation that enters the cells through calciumchannels and suppresses fluorescence by competitively dis-placing Ca21 from fura2, lindane induced an increase inintracellular fluorescence intensity followed by a progres-sive decline to below the prestimulus level (Fig. 7). Basedon this evidence it seems that in PMCs, lindane both triggersCa21 release from an intracellular store and promotes Ca21
Fig. 3. Dose-dependence of lindane effects on GPem (panel a) and GPex
(panel b) values in Prodan-loaded PMCs. Data represent the differencebetween GP values collected before and after 10 min of lindane addition.Data points (means6 SD) are the results of at least four experiments. Linesare the best fits to experimental data.
Fig. 4. Effect of 200mM lindane (arrow) on [Ca21]i in PMCs suspendedin regular buffer. The trace is representative of at least four consistentexperiments.
Fig. 5. Dose-dependence of lindane effect on PMC [Ca21]i. Data are thepeak values expressed as percent of the corresponding prestimulus value.Data points (means6 SD) are the results of at least four experiments. Lineis the best fit to the experimental data.
435L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

influx from the extracellular medium. By using the VOCCorganic blocker nicardipine, the [Ca21]i increase producedby the xenobiotic (Fig. 8) was clearly reduced with respectto control (Fig. 4), suggesting the voltage dependence of thelindane-induced calcium influx.
3.3. Effects of lindane on membrane potential
The effect of lindane on PMCs loaded with the voltage-sensitive dye bisoxonol is shown in Fig. 9a. The addition of
lindane induced membrane depolarization consisting of afirst ultra-rapid phase followed by a second, slower one; thelatter phase reached a maximum about 1.5 min after addi-tion with only a small tendency to return to the basal level.The phenomenon was dose-dependent, with maximal andhalf-maximal effects attained at 200 and 100mM lindane,respectively (Fig. 10). In the absence of either extracellularNa1, Ca21, or Cl2, the first phase of depolarization did notchange, meaning that the lindane induced rapid membranedepolarization independently of transmembrane ion fluxes(Fig. 9b, c, d). As for the second phase of depolarization,choline substitution for extracellular Na1 did not modify toa significant extent the cell response obtained in regularbuffer (Fig. 9b), whereas a manifest increase was producedwhen either Cl2 or Ca21 were omitted from the extra-cellular medium. These findings indicate that an outwardchloride current participates in the lindane-induced depolar-
Fig. 6. Effect of 200mM lindane on PMC [Ca21]i suspended in EGTAbuffer (panel a), in EGTA buffer after addition of 1mM thapsigargin (panelb), and in regular buffer after addition of 1mM thapsigargin (panel c). Thetraces are representative of at least four consistent experiments. Shortarrow 5 lindane addition; long arrow5 thapsigargin addition.
Fig. 7. Effect of 200mM lindane (short arrow) on [Ca21]i in PMCssuspended in Ca21-free buffer to which was added 10mM Mn21 (longarrow). The trace is representative of three consistent experiments. a.u.5arbitrary units.
Fig. 8. Effect of 200mM lindane (arrow) on [Ca21]i in PMCs suspendedin regular buffer in the presence of the VOCC inhibitor nicardipine (5mM).The trace is representative of three consistent experiments.
436 L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

ization and that the extent of this depolarization was limitedby the activation of Ca21-dependent repolarizing mecha-nisms (likely, Ca21-activated K1 channels). Neither 0.5 to50 mM GABA nor 50 mM picrotoxin had an effect on themembrane resting potential or lindane-induced depolariza-tion (data not shown); based on this evidence, involvementof GABA receptor/Cl2 channels in lindane effects on PMCscan reasonably be ruled out.
3.4. Effect of lindane on PMC responses to AVP andET-1
As expected [36], ET-1 induced a transient [Ca21]i in-crease in PMCs suspended in regular buffer (Fig. 11a), anda comparable calcium response was also evoked by AVP(Fig. 11b). In the presence of lindane, the above responseswere reduced in magnitude without significant changes intiming and curve shape. The effect was concentration-de-pendent, with 100 and 60% maximal inhibition produced by50 mM lindane and an IC50 of 8 and 6mM lindane for ET-1and AVP, respectively (Fig. 12a). Remarkably, maximalinhibition of [Ca21]i increase was produced by lindanedoses well below those effective in depleting intracellularcalcium stores. In accordance with literature data [52], bothpeptides elicited PMC contraction (c 5 0.75 6 0.04 and0.65 6 0.03 for ET-1 and AVP, respectively). The effectwas dose-dependently prevented by lindane, with 100 and60% inhibition produced by 50mM lindane and an approx-imate IC50 of 8 and 4 mM lindane for ET-1 and AVP,respectively (Fig. 12b).
4. Discussion
The aim of the present investigation was to examine thein vitro effects of lindane on the smooth muscular compo-nent of the rat testis. Concentrations of lindane in thepresent work were selected on the basis of previous studiesin which other biochemical processes were investigated.Prodan generalized polarization (GP) showed that the in-
Fig. 9. Effect of 200mM lindane (arrow) on membrane potential in PMCs suspended in regular buffer (panel a), Na21-free buffer (panel b), EGTA buffer(panel c), and Cl2-free buffer (panel d). The vertical bar represents the fluorescence increase produced by 30 mM KCl in a parallel batch of cells and refersto all the traces presented in the figure. Each trace is representative of at least three consistent experiments.
Fig. 10. Dose-dependence of lindane effect on PMC membrane potential.The data correspond to the fluorescence values 1.5 min after addition andare expressed as percent of the prestimulus value. Data points (means6SD) are the results of at least four experiments. Line is the best fit to theexperimental data.
437L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

secticide intercalates with phospholipids at the lipid/waterinterface, a finding in good correlation with the high lipid/water partition coefficient of lindane [53]. The observedlindane-induced increment of both GPem and GPex supportsan increase of membrane polarity accompanied by increasedmembrane order.
Inhibition experiments ruled out that modifications ofmembrane phospholipid composition by PLA2 and PLC,two lindane-activatable membrane lipases [10], play a sig-nificant role in the phenomenon. Instead, the effect observedlikely resulted from a progressive increase in intramem-brane lindane concentration. Because the asymmetrical dis-tribution of chlorine atoms in the lindane molecule givesthis compound a significant dipole moment, its membranepartition likely produces both the increase of membranedipole number (i.e., bilayer polarity increase) and a decreasein dipole kinetics (i.e., membrane order increase).
Previous papers reported that lindane increases mem-brane fluidity in plasma membranes isolated from rat ventralprostate [54] and renal cortex [55], whereas no effect wasshown on membrane fluidity in rat enterocytes [32] andintracellular membranes [56]. The above studies were car-ried out by using DPH, a probe sensing the deepest region
of the bilayer; hence, comparing those findings with ours isnot feasible. However, by using 1,3-di(2-pyrenyl)propane, aprobe that populates the same membrane region where Pro-dan molecules localize, lindane was found to exert an or-dering effect on membrane lipids [33], a result consistentalso with a recent finding from our lab showing that theinsecticide reduces the molecular dynamics at the spermsurface [34]. In cells, membrane biophysical properties in-fluence the spatial arrangement of membrane-embeddedproteins, by which effect protein functions are modulated[57]. Then, because the membrane environment is signifi-cantly altered by lindane, this compound, and perhaps otherstructurally related chemicals as well, may be expected tointerfere with membrane-located biologic processes (e.g.,receptor recognition, ion permeability, substrate transport)even in the absence of direct lindane-protein interaction.This assumption is in keeping with a previous study report-
Fig. 11. Representative tracings of ET-1 (4 nM, panel a) and AVP (100nM, panel b) effects on [Ca21]i in PMCs suspended in regular buffer.Arrow 5 hormone addition.
Fig. 12. Dose-dependence of lindane effect on ET-1-(4 nM, closed sym-bols) and AVP- (100 nM, open symbols) induced [Ca21]i increase (panela) and contraction (panel b). Data are normalized assuming maximalresponses (no lindane added)5 100. Data points (means6 SE) are theresults of four experiments, each run in triplicate. Lines are the best fits toexperimental data. *P , 0.05; **P , 0.01.
438 L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

ing that perturbation of membrane fluidity by lindane haseffects on adenylate cyclase activity and glucose transport[31].
Concerning the lindane effect on [Ca21]i, both a releasefrom internal stores and an influx from the extracellularmedium contributed to the induced [Ca21]i. Pharmacologicmanipulations showed that IP3-sensitive Ca21 stores weredepleted by lindane independently of de novo synthesis ofIP3. These results were not unexpected, because the struc-tural similarity between IP3 and lindane makes the latterable to interact with and activate sarcoplasmic IP3 receptors[27]. However, the evidence that [Ca21]i did not return tothe basal level after lindane addition and the known abilityof lindane to inhibit Ca21-ATPases [58,59] and to interferewith calmodulin mRNA expression [60], suggest that mul-tiple mechanisms of [Ca21]i increase may be important.Finally, a supplementary [Ca21]i increase, mostly contrib-uted by a voltage-operated influx sensitive to the VOCCorganic blocker nicardipine, was triggered by lindane. Thisfinding was produced in the absence of apparent cytotoxic-ity, as assessed by the trypan blue exclusion method, and isin agreement with previous reports showing that lindaneaffects voltage-dependent Ca21 currents in several celltypes both in vitro [34,26] and in vivo [61].
The opening of voltage-dependent mechanisms of Ca21
influx implies that lindane alters the membrane potential. Infact, we found that lindane depolarized PMCs and that twocomponents were incorporated into this depolarization: anultra-fast component, independent of transmembrane ionfluxes, and a larger, slower one dependent on an efflux ofchloride. Concerning the first component, besides the netcharge separation across the bilayer, membrane potential isdetermined by the density of fixed charges at the lipid/waterinterface (surface potential) and by the arrangement andnumber of intrabilayer dipoles (dipole potential). Becausethe lindane molecule is electroneutral, it cannot modify thesurface potential; instead, the high lindane dipole momentmakes it able to alter the membrane dipole potential [34].By taking into account the biophysical findings discussedabove, a reasonable conclusion is that lindane partition inthe PMC surface alters the membrane dipole potential byeither increasing intramembrane dipole concentration or re-orienting local dipoles. The finding that the second compo-nent of membrane depolarization was coupled to a chlorideefflux was unexpected because lindane is known to inhibitrather than activate both GABA-operated and voltage-de-pendent chloride currents [18]. In fact, in contrast with othercell models, neither GABA nor picrotoxin had an apparenteffect on the resting membrane potential or lindane-induceddepolarization in PMCs. However, it should be rememberedthat at least two additional routes of chloride efflux mayoperate in cells, i.e., Ca21-activated and cyclic AMP-depen-dent Cl2 channels. Our present data do not allow a conclu-sive interpretation, and additional work is warranted todistinguish among these possibilities.
Both AVP- and ET-1-inducible [Ca21]i increase and
contraction were strongly inhibited by lindane. The effectwas evident at lindane concentrations well below thoseeffective in altering basal [Ca21]i, a result that makes un-likely a role for Ca21 store depletion in the observed inhi-bition. This finding is in agreement with a previous reportshowing that in rat myometrial smooth muscle cells, lindaneconcentrations as low as 1mM, although scarcely effectivein altering basal [Ca21]i, dramatically inhibited the Ca21
response to charbacol [28]. The evidence that lindane didnot dampen the hormone-induced [Ca21]i increase by de-pleting intracellular Ca21 stores suggests that earlier ele-ments in the hormone signal transduction pathways may beaffected. Among these, receptor-coupled GTPases andphosphinositidase C, as well as the receptors themselves,are membrane-embedded proteins involved in AVP- andET-1-triggered Ca21 signaling [62], and represent potentialtargets for lindane action. Although the inhibitory effect oflindane could have also been contributed by its interactionwith hydrophobic domains of membrane proteins, an alter-native possibility can be proposed. It is known that confor-mation and functioning of membrane-inserted proteins aresignificantly influenced by intramembrane dipoles [63,64].This knowledge and the evidence that lindane alters thenumber and dynamics of membrane dipoles (this study),raise the plausible hypothesis that the observed impairmentof PMC responses to AVP and ET-1 may have resultedfrom membrane environment perturbation by lindane. Ourhypothesis is also in keeping with previous reports in whichlindane interference with function of intrinsic proteins wasshown, supposedly accounted for by membrane perturba-tion by the insecticide [10,31].
In vivo studies demonstrated that adult male rats exposedto lindane suffered from hypospermiogenesis and Sertolicell ballooning and fragmentation that, after perinatal expo-sure, were accompanied by testicular weight reduction [22,23].
By considering that PMCs produce paracrine factorsmodulating Sertoli cell functions essential for spermatogen-esis, it may be that the in vivo effects of lindane on testicularfunction depend in part on lindane action on PMCs. Thatlindane concentrations measured in vivo overlap those wefound to be effective in inhibiting PMC responses to phys-iologic agonists in vitro lends good support to the aboveconsideration.
Great caution is needed in extrapolating in vitro obser-vations to real life; however, by remembering that PMC-secreted factors strongly modulate Sertoli cell differentia-tion [65], and considering also that it is the Sertoli cellnumber that determines the number of spermatogenic cells,impairment of PMC function during gonadal developmentcould account for the reduced testicular size observed in ratsperinatally exposed to lindane. Further studies are beingcarried out in our laboratory aiming to evaluate the effectsof lindane and other structurally related xenobiotics onPModS and activin-A secretion in PMCs.
439L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

Acknowledgments
This research was supported by grants from the MedicalFaculty of the University of Rome ‘La Sapienza’ (to L.S.and V.S.) and funded in part by the Istituto Superiore diSanita Research Project ‘Prevention of Risk Factors ofChild and Maternal Health’.
References
[1] Soto AM, Chung KL, Sonnenschein C. The pesticides endosulfan,toxaphene, and dieldrin have estrogenic effects on human estrogen-sensitive cells. Environ Health Perspect 1994;102:380–3.
[2] Mably TA, Bjerke DL, Moore RW, Gendron–Fitzpatrick A, PetersonRE. In utero and lactational exposure of male rats to 2,3,7,8-tetra-chlorodibenzo-p-dioxin. 3. Effects on spermatogenesis and reproduc-tive capability. Toxicol Appl Pharmacol 1992;114:118–26.
[3] Gray LE Jr, Ostby J, Marshall R, Andrews, J. Reproductive andthyroid effects of low-level polychlorinated biphenyl (Aroclor 1254)exposure. Fundam Appl Toxicol 1993;20:288–94.
[4] Murature DA, Tang SY, Steinhardt G, Dougherty RC. Phtalate estersand semen quality parameters. Biomed Env Mass Spectr 1987;14:473–7.
[5] White R, Jobling S, Sumpter JP, Parker MG. Environmentally per-sistent alkylphenolic compounds are estrogenic. Endocrinology 1994;135:175–82.
[6] Steinmetz R, Brown NG, Allen DL, Bigsby M, Ben.Jonathan N. Theenvironmental estrogen bisphenol A stimulates prolactin release andin vivo. Endocrinology 1997;138:1780–6.
[7] Danzo BJ. Environmental xenobiotics may disrupt normal endocrinefunction by interfering with the binding of physiological ligands tosteroid receptors and binding proteins. Env Health Persp 1997;105:294–301.
[8] Wen LP, Koeiman N, Whitlock JP. Dioxin-inducible, Ah receptor-dependent transcription. Proc Natl Acad Sci USA 1990;87:8545–9.
[9] Denison MS, Fisher JM, Whitlock JP Jr. The DNA recognition sitefor the dioxin-Ah receptor complex. Nucleotide sequence and func-tional analysis. J Biol Chem 1988;263:17221–4.
[10] Puente–Fraga JC, Lopez–Aparicio P, Senar S, Recio MN, Perez–Albarsanz MA. Hexachlorocyclohexanes affect the arachidonic acidrelease from phosphatidylinositol but not from other phospholipidclasses in tubular cell cultures. Biosci Rep 1995;15:191–9.
[11] Mohr FC, Alojipan SV, Dunston SK, Pessah IN. The delta isomer ofhexachlorocyclohexane induces rapid release of the myo-inositol-1,4,5-trisphosphate-sensitive Ca21 store and blocks capacitative Ca21
entry in rat basophilic leukemia cells. Mol Pharmacol 1995;48:512–22.
[12] Eskenazi B, Kimmel G. Workshop on perinatal exposure to dioxin-like compounds. II. Reproductive effects. Environ Health Perspect1995;103:143–5.
[13] Carlsen E, Giwercman A, Keiding N, Skakkebaek NE. Evidence fordecreasing quality of semen during past 50 years. Br Med J 1992;305:609–13.
[14] Sharpe RM, Skakkebaek NE. Are oestrogens involved in fallingsperm counts and disorders of the male reproductive tract? Lancet1993;341:1392–5.
[15] Fisch H, Goluboff ET. Geographic variations in sperm counts: apotential cause of bias in studies of semen quality. Fertil Steril1996;65:1044–6.
[16] Berling S, Wolner–Hanssen P. No evidence of deteriorating semenquality among men in infertile relationships during the last decade: astudy of males from Southern Sweden. Hum Reprod 1997;12:1002–5.
[17] Olsen GW, Bodner KM, Ramlow JM, Ross CE, Lipshultz LI. Havesperm counts be reduced 50% in 50 years? A statistical modelrevisited. Fertil Steril 1995;63:887–93.
[18] Eldefrawi AT, Eldefrawi ME. Receptors for gamma-aminobutyricacid and voltage-dependent chloride channels as targets for drugs andtoxicants. FASEB J 1987;1:262–71.
[19] Cooper RL, Chadwick RW, Rehnberg GL, Goldman JM, Booth KC,Hein JF, McElroy WK. Effect of lindane on hormonal control ofreproductive function in the female rat. Toxicol Appl Pharmacol1989;99:384–94.
[20] Sircar S, Lahiri P. Effect of lindane on mitochondrial side-chaincleavage of cholesterol in mice. Toxicology 1990;61:41–6.
[21] Jin L, Tran DQ, Ide CF, McLachlan JA, Arnold SF. Several syntheticchemicals inhibit progesterone receptor-mediated transactivation inyeast. Biochem Biophys Res Commun 1997;233:139–46.
[22] Dalsenter PR, Faqi AS, Webb J, Merker HJ, Chahoud I. Reproductivetoxicity and tissue concentrations of lindane in adult male rats. HumExp Toxicol 1996;15:406–10.
[23] Dalsenter PR, Faqi AS, Webb J, Merker HJ, Chahoud I. Reproductivetoxicity and toxicokinetics of lindane in the male offspring of ratsexposed during lactation. Hum Exp Toxicol 1997;15:146–53.
[24] Descampiaux B, Leroux JM, Peucelle C, Erb F. Assay of free-radicaltoxicity and antioxidant effect on the Hep 3B cell line: a test surveyusing lindane. Cell Biol Toxicol 1996;12:19–28.
[25] del Hoyo N, Pulido JA, Perez–Albarsanz MA. Characterization ofphosphoinositide hydrolysis products induced by hexachlorocyclo-hexane isomers in rat brain cortex. Biosci Rep 1993;13:119–26.
[26] Pinelli E, Cambon C, Tronchere H, Chap H, Teissie J, Pipy B.Ca(21)-dependent activation of phospholipases C and D from mouseperitoneal macrophages by a selective trigger of Ca21 influx, gamma-hexachlorocyclohexane. Biochem Biophys Res Commun 1994;199:699–705.
[27] Criswell KA, Loch–Caruso R, Stuenkel EL. Lindane inhibition of gapjunctional communication in myometrial myocytes is partially depen-dent on phosphoinositide-generated second messengers. ToxicolAppl Pharmacol 1995;130:280–93.
[28] Criswell KA, Stuenkel EL, Loch–Caruso R. Lindane increases intra-cellular calcium in rat myometrial smooth muscle cells through mod-ulation of inositol 1,4,5-trisphosphate-sensitive stores. J PharmacolExp Ther 1994;270:1015–24.
[29] Anand M, Meera P, Kumar R, Gupta GS, Tripathi O, Srimal R.Possible role of calcium in the cardiovascular effects of prolongedadministration of gamma-HCH (lindane) in rats. J Appl Toxicol1995;15:245–8.
[30] Omann GM, Lakowicz JR. Interactions of chlorinated hydrocarboninsecticides with membranes. Biochim Biophys Acta 1982;684:83–95.
[31] Lopez–Aparicio P, Recio MN, Prieto JC, Perez–Albarsanz MA. Roleof lindane in membranes. Effects on membrane fluidity and activity ofmembrane-bound proteins. Biosci Rep 1994;14:131–8.
[32] Carrero I, Rodriguez–Henche N, Guijarro LG, Recio MN, Perez–Albarsanz MA, Prieto JC. Lindane decreases forskolin-stimulatedcyclic AMP accumulation but does not modify Gs in rat enterocytes.Cell Signal 1993;5:453–62.
[33] Antunes–Madeira MC, Almeida LM, Madeira VM. Effects of lindaneon membrane fluidity: intramolecular excimerization of a pyrenederivative and polarization of diphenylhexatriene. Biochim BiophysActa 1990;1022:110–14.
[34] Silvestroni L, Fiorini R, Palleschi S. Partition of the organochlorineinsecticide lindane into the human sperm surface induces membranedepolarization and Ca21 influx. Biochem J 1997;321:691–8.
[35] Gnessi L, Fabbri A, Spera G. Gonadal peptides as mediators ofdevelopment and functional control of the testis: an integrated systemwith hormones and local environment. Endoc Rev 1997;18:541–609.
[36] Santiemma V, Beligotti F, Magnanti M., Palleschi S, Silvestroni L,Fabbrini A. Endothelin-1 stimulates deoxyribonucleic acid synthesis
440 L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441

and contraction in testicular peritubular myoid cells. Biol Reprod1996;54:583–90.
[37] Norton JN, Vigne JL, Skinner MK. Regulation of Sertoli cell differ-entiation by the testicular paracrine factor PModS: analysis of com-mon signal transduction pathways. Endocrinology 1994;134:149–57.
[38] Maekawa M, Kamimura K, Nagano T. Peritubular myoid cells in thetestis: their structure and function. Arch Histol Cytol 1996;59:1–13.
[39] Skinner MK. Cell-cell interactions in the testis. Endoc Rev 1991;12:45–77.
[40] Walum E, Peterson A, Erkell LJ. Photometric recording of cellviability using trypan blue in perfused cell cultures. Xenobiotica1985;15:701–4.
[41] Anthony CT, Skinner MK. Cytochemical and biochemical character-ization of testicular peritubular myoid cells. Biol Reprod 1989;40:811–23.
[42] Parasassi T, Giusti AM, Gratton E, Monaco E, Raimondi M, Ravag-nan G, Sapora O. Evidence for an increase in water concentration inbilayer after oxidative damage of phospholipids induced by ionizingradiation. Int J Radiat Biol 1994;65:329–34.
[43] Shaw JO, Roberts MF, Ulevitch RJ, Henson P, Dennis EA. Phospho-lipase A2 contamination of cobra venom factor preparations. Biologicrole in complement-dependent in vivo reaction and inactivation withP-bromophenylacyl bromide. Am J Pathol 1978;91:517–30.
[44] Schwertz DW, Halberson JB, Palmer JW, Feinberg H. Characteriza-tion of phospholipase C-mediated phospatidylinositol degradation inrat heart ventricle. Arch Biochem Biophys 1987;253:388–98.
[45] Olsen RW, Yang J, King RG, Dilber A, Stauber GB, Ransom RW.Barbiturate and benzodiazepine modulation of GABA receptor bind-ing and function. Life Sci 1976;39:1969–76.
[46] Godfraind T, Miller R, Wibo M. Calcium antagonism and calciumentry blockade. Pharmacol Rev 1986;38:321–416.
[47] Di Virgilio F, Steinberg TH, Silverstein SC. Inhibition of Fura-2sequestration and secretion with organic anion transport blockers.Cell Calcium 1990;11:57–62.
[48] Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca21
indicators with greatly improved fluorescence properties. J Biol Chem1985;260:3440–50.
[49] Tung PS, Fritz IB. Transforming growth factor-b and platelet-derivedgrowth factor synergistically stimulate contraction by testicular peri-tubular cells in culture in serum-free medium. J Cell Physiol 1991;146:386–93.
[50] Parasassi T, Loiero M, Raimondi M, Ravagnan G, Gratton E.Absence of lipid gel-phase domains in seven mammalian cell linesand in four primary cell types. Biochem Biophys Acta 1993;1153:143–54.
[51] Thastrup O, Dawson AP, Scharff O, Foder B, Cullen PJ, Drobak BK,Bjerrum PJ, Christensen SB, Hanley MR. Thapsigargin, a novel
molecular probe for studying intracellular calcium release and stor-age. Agents and Actions 1994;43:187–93.
[52] Tripiciano A, Filippini A, Giustiniani Q, Palombi F. Direct visual-ization of rat peritubular myoid cell contraction in response to endo-thelin. Biol Reprod 1996;55:25–31.
[53] Lakowicz JR, Hogen D, Omann G. Diffusion and partitioning of apesticide, lindane, into phosphatidylcholine bilayers. A new fluores-cence quenching method to study chlorinated hydrocarbon-membraneinteractions. Biochim Biophys Acta 1977;471:401–11.
[54] Senar S, Gutierrez–Ocana MT, Perez–Albarsanz MA, Recio MN.Influence of lindane on the fluidity of the rat ventral prostate mem-branes. Biosci Rep 1991;11:101–10.
[55] Perez–Albarsanz MA, Lopez–Aparicio P, Senar S, Recio MN. Effectsof lindane on fluidity and lipid composition in rat renal cortex mem-branes. Biochim Biophys Acta 1991;1066:124–30.
[56] Antunes–Madeira MC, Madeira VMC. Membrane fluidity as affectedby the insecticide lindane. Biochim Biophys Acta 1989;982:161–6.
[57] Lee AG, Michelangeli F, East JM. Tests for the importance of fluidityfor the function of membrane proteins. Biochem Soc Trans 1989;17:962–4.
[58] Srivastava SC, Kumar R, Prasad AK, Srivastava SP. Effect of hexa-chlorocyclohexane (HCH) on testicular plasma membrane of rat.Toxicol Lett 1995;75:153–7.
[59] Jones OT, Froud RJ, Lee AG. Interactions of hexachlorocyclohexaneswith the (Ca21-Mg21)-ATPase from sarcoplasmic reticulum. Bio-chim. Biophys Acta 1985;812:740–51.
[60] Barron S, Tusell JM, Serratosa J. Effect of hexachlorocyclohexaneisomers on calmodulin mRNA expression in the central nervoussystem. Brain Res Mol Brain Res 1995;20:279–86.
[61] Tusell JM, Barron S, Serratosa J. Anticonvulsant activity of cal-modulin antagonist W-7 in convulsions induced by lindane andBayK-8644: effects in c-fos expression. Neurotoxicology 1994;15:751– 6.
[62] Howl J, Rudge SA, Lavis RA, Davies AR, Parslow RA, Hughes PJ,Kirk CJ, Michell RH, Wheatley M. Rat testicular myoid cells expressvasopressin receptors: receptor structure, signal transduction, anddevelopmental regulation. Endocrinology 1995;136(5):2206–13.
[63] Cladera J, O’Shea P. Intramembrane molecular dipoles affect themembrane insertion and folding of a model amphiphilic peptide.Biophys J 1998;74(5):2434–42.
[64] Bevan JA, Bevan RD, Shreeve SM. Variable receptor affinity hy-pothesis. FASEB J 1989;3:1696–704.
[65] Chaudhary J, Skinner MK. Transcriptional regulation of Sertoli celldifferentiation (transferrin promoter activation) during testicular de-velopment. Dev Genet 1995;16:114–8.
441L. Silvestroni et al. / Reproductive Toxicology 13 (1999) 431–441




















