
A New Miniaturized Lizard From the Late Eocene of France and Spain
11
A New Miniaturized Lizard From the Late Eocene of France and Spain ARNAU BOLET 1 * AND MARC AUG E 2 1 Institut Catal a de Paleontologia Miquel Crusafont, Grup de Recerca de Faunes del Mesozoic/Universitat Aut onoma de Barcelona, Carrer Escola Industrial 23, Sabadell, E-08201, Catalonia, Spain 2 Mus eum national d’Histoire naturelle, D epartement Histoire de la Terre, UMR 7207, CNRS, rue Buffon, F-75231, Paris, France Abstract We report here a new lizard genus and species shared by two late Eocene localities situated at both versants of the present Pyrenees (South-Western Europe), one located in France (Escamps, MP19), and the other in Catalonia, Spain (Soss ıs, MP17a). The recovered specimens are remarkable because of their small size and peculiar morphology. Features of the dentary are interpreted as adaptations to a fossorial or semi- fossorial lifestyle, although such modifications obscure the exact phyloge- netic relationships of the new taxon. We suggest that it might represent a further example of scincoid lizard that independently achieved adapta- tions for burrowing or surface-dwelling. This taxon reinforces the hypoth- eses that link the Southern Pyrenean assemblages to those from France rather than to those of the rest of the Iberian Peninsula, which are sup- posed to be somehow isolated and endemic to a certain degree during the middle and late Eocene, forming part of the so-called Western Iberian Bioprovince. Anat Rec, 297:505–515, 2014. V C 2014 Wiley Periodicals, Inc. Key words: squamata; late Eocene; Scincoidea; dental mor- phology; miniaturization; paleobiogeography The classical French localities at the Phosphorites du Quercy (Fig. 1) contain the best-known Paleogene sequence from Europe, mainly regarding the time span from the late Eocene to early Oligocene. They have yielded a diverse and rich herpetofauna, including the best-known Eocene lizard assemblages from Europe (see Aug e, 2005). The lizards from Escamps (MP19, Phos- phorites du Quercy, France) were described in Aug e (2005) and the assemblage is quite similar to that of other French localities of the MP17-19 interval. Lizard- bearing localities in Southern Europe, however, are scarcely known, and thus still have a great potential regarding the paleobiogeographical and paleoenviron- mental information that can be obtained from their study. The Iberian lizard assemblage from Soss ıs (MP17a, Catalonia, Spain; Fig. 1) shares many taxa with several French contemporaneous localities (Bolet and Evans, 2013; see also Table 1). This similarity, also reported for mammalian assemblages (Casanovas-Cla- dellas and Santaf e-Llopis, 1987; Cuesta, 1991; Badiola et al., 2009), suggests that a connection existed between the two regions at the earliest late Eocene. In contrast, a certain degree of endemicity has been detected for mammalian taxa from localities situated in Central and Western Iberian Peninsula, allowing the definition of the Western Iberian Bioprovince (Cuesta, 1991). Examples Grant sponsor: FPI Grant; Grant number: BES-2009-026731; Grant sponsor: Ministerio de Ciencia e Innovaci on, Spain; Grant number: CGL2008-06533-C03-01/BTE; Grant sponsor: Ministerio de Econom ıa y Competitividad, Spain; Grant num- bers: CGL2011-30069-C02-01, CGL2011-27343; Grant sponsor: Ministerio de Ciencia E Innovaci on, Spain. *Correspondence to: Arnau Bolet, Institut Catal a de Paleonto- logia Miquel Crusafont/Universitat Aut onoma de Barcelona, Carrer Escola Industrial 23, Sabadell, E-08201, Catalonia, Spain. Fax: 00-34-937276641. E-mail: [email protected] Received 26 February 2013; Accepted 11 June 2013. DOI 10.1002/ar.22855 Published online 31 January 2014 in Wiley Online Library (wileyonlinelibrary.com). THE ANATOMICAL RECORD 297:505–515 (2014) V V C 2014 WILEY PERIODICALS, INC.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A New Miniaturized Lizard From the Late Eocene of France and Spain

A New Miniaturized Lizard From theLate Eocene of France and Spain
ARNAU BOLET1* AND MARC AUG�E2
1Institut Catal�a de Paleontologia Miquel Crusafont, Grup de Recerca de Faunes delMesozoic/Universitat Aut�onoma de Barcelona, Carrer Escola Industrial 23,
Sabadell, E-08201, Catalonia, Spain2Mus�eum national d’Histoire naturelle, D�epartement Histoire de la Terre, UMR 7207,
CNRS, rue Buffon, F-75231, Paris, France
AbstractWe report here a new lizard genus and species shared by two late
Eocene localities situated at both versants of the present Pyrenees(South-Western Europe), one located in France (Escamps, MP19), and theother in Catalonia, Spain (Soss�ıs, MP17a). The recovered specimens areremarkable because of their small size and peculiar morphology. Featuresof the dentary are interpreted as adaptations to a fossorial or semi-fossorial lifestyle, although such modifications obscure the exact phyloge-netic relationships of the new taxon. We suggest that it might represent afurther example of scincoid lizard that independently achieved adapta-tions for burrowing or surface-dwelling. This taxon reinforces the hypoth-eses that link the Southern Pyrenean assemblages to those from Francerather than to those of the rest of the Iberian Peninsula, which are sup-posed to be somehow isolated and endemic to a certain degree during themiddle and late Eocene, forming part of the so-called Western IberianBioprovince. Anat Rec, 297:505–515, 2014. VC 2014 Wiley Periodicals,Inc.
Key words: squamata; late Eocene; Scincoidea; dental mor-phology; miniaturization; paleobiogeography
The classical French localities at the Phosphorites duQuercy (Fig. 1) contain the best-known Paleogenesequence from Europe, mainly regarding the time spanfrom the late Eocene to early Oligocene. They haveyielded a diverse and rich herpetofauna, including thebest-known Eocene lizard assemblages from Europe (seeAug�e, 2005). The lizards from Escamps (MP19, Phos-phorites du Quercy, France) were described in Aug�e(2005) and the assemblage is quite similar to that ofother French localities of the MP17-19 interval. Lizard-bearing localities in Southern Europe, however, arescarcely known, and thus still have a great potentialregarding the paleobiogeographical and paleoenviron-mental information that can be obtained from theirstudy. The Iberian lizard assemblage from Soss�ıs(MP17a, Catalonia, Spain; Fig. 1) shares many taxawith several French contemporaneous localities (Boletand Evans, 2013; see also Table 1). This similarity, alsoreported for mammalian assemblages (Casanovas-Cla-dellas and Santaf�e-Llopis, 1987; Cuesta, 1991; Badiolaet al., 2009), suggests that a connection existed between
the two regions at the earliest late Eocene. In contrast,a certain degree of endemicity has been detected formammalian taxa from localities situated in Central andWestern Iberian Peninsula, allowing the definition of theWestern Iberian Bioprovince (Cuesta, 1991). Examples
Grant sponsor: FPI Grant; Grant number: BES-2009-026731;Grant sponsor: Ministerio de Ciencia e Innovaci�on, Spain;Grant number: CGL2008-06533-C03-01/BTE; Grant sponsor:Ministerio de Econom�ıa y Competitividad, Spain; Grant num-bers: CGL2011-30069-C02-01, CGL2011-27343; Grant sponsor:Ministerio de Ciencia E Innovaci�on, Spain.
*Correspondence to: Arnau Bolet, Institut Catal�a de Paleonto-logia Miquel Crusafont/Universitat Aut�onoma de Barcelona,Carrer Escola Industrial 23, Sabadell, E-08201, Catalonia,Spain. Fax: 00-34-937276641. E-mail: [email protected]
Received 26 February 2013; Accepted 11 June 2013.
DOI 10.1002/ar.22855Published online 31 January 2014 in Wiley Online Library(wileyonlinelibrary.com).
THE ANATOMICAL RECORD 297:505–515 (2014)
VVC 2014 WILEY PERIODICALS, INC.

of shared taxa with localities from Central Europe exist(e.g., artiodactyls; Badiola, 2004), but the presence of ahigh number of endemic taxa pertaining to differentgroups (perissodactyls, rodents, primates; e.g., Badiolaand Cuesta, 2008; Marig�o et al., 2010; Minwer-Barakatet al., 2012) points to a certain degree of isolation of agreat part of the Iberian Peninsula (the Southern Pyre-nean basins excluded). The lack of described lizardassemblages from the Western Iberian Bioprovince pre-cludes comparison with Southern Pyrenean and Frenchfaunas, although the study of available material shouldaddress this issue in the close future (A. Bolet in prepa-ration). In this contribution, an additional example of ashared taxon between the Southern Pyrenean andFrench late Eocene faunas is presented, supporting thesimilarity of the assemblages from these two regions.
MATERIALS AND METHODS
The available material comprises three specimensfrom the late Eocene of France and Spain. The Frenchmaterial belongs to the collection of the Universit�e desSciences et Techniques du Languedoc (USTL), EscampsB locality (UM-ECB), from the reference level MP19,and consists of a tiny, almost complete and well-preserved right dentary. The two additional specimensare from the collections of the Institut Catal�a de Paleon-tologia (ICP), formerly Institut de Paleontologia de Saba-dell (IPS), Soss�ıs locality (MP17a, Catalonia, Spain), andconsist of one left and one right small dentary frag-ments, which could belong to the same individual.Images were taken with the Environmental ScanningElectron Microscope (ESEM) Quanta 200 from the Uni-versitat de Barcelona (UB), and drawings of the typespecimen were made with a camera lucida attached to astereoscopic binocular. Data on lizards from selectedlocalities were taken from Aug�e (2005), Aug�e (2007) andBolet and Evans (2013).
RESULTS
Systematic Paleontology
Reptilia Linnaeus, 1758Squamata Oppel, 1811Scincoidea Oppel, 1811?Scincidae Gray, 1825Genus Pyrenasaurus nov.Type species: Pyrenasaurus evansae sp. nov.Etymology: From Pyrena, the latin root of Pyrenaeum,
the mountains that now separate Catalonia from France,the Pyrenees (Pyr�en�ees in French, Pirineus in Catalan,Pirineos in Spanish); and saurus, meaning lizard inGreek.
Diagnosis: As for the type and only species.Pyrenasaurus evansae sp. nov.Figures 2 and 3A,BEtymology: Dedicated to Prof. Susan E. Evans for her
efforts in the study on paleoherpetofauna in general,and on lepidosaurs specifically, and for her help and sup-port to the senior author through the elaboration of hisPh.D. dissertation.
Holotype: UM-ECB 1703 (Fig. 2A–E), complete rightdentary with seven preserved teeth (eleven tooth posi-tions in total).
Type locality: Escamps B (Phosphorites du Quercy,France).
Type horizon: Escamps Standard level, MP19.Geographical and stratigraphical distribution: Known
from the type locality of Escamps (MP19 of the Phos-phorites du Quercy, France) and from Soss�ıs (MP17a,Central Southern Pyrenean Basin, also called Graus-Tremp basin, Catalonia, Spain).
Referred material: The additional specimens referredto the same taxon come from the locality of Soss�ıs(MP17a, Central Southern Pyrenean Basin, Catalonia,Spain). They were referred to ?Scincoidea incertae sedis,unnamed new genus and species in Bolet and Evans(2013): IPS 56061 (Fig. 2F,G,I), partial left dentary withtwo teeth; IPS 56168, fragment of right dentary withone preserved tooth (Fig. 2H,J).
Diagnosis: Very small lizard (length of dentary equals3.4 mm as preserved) characterized by the followingcombination of characters: a low tooth count (eleventooth positions in total), a closed and fused Meckeliancanal, a straight dentary ventral border, a moderatelyhigh coronoid process of the dentary (higher than thetallest tooth crown), and a highly distinctive tooth mor-phology: anterior teeth (first to fifth positions) wherepreserved, small and slightly posteriorly recurved, show-ing the typical scincoid pattern (sensu Richter, 1994);posterior teeth (ninth to eleventh positions) larger, labio-lingually compressed, and with a characteristic deflec-tion in the convexity of the posterior and anterior (lessdeveloped) shafts close to the tip; medial teeth (sixth toeighth positions) intermediate in shape, height, androbustness. Regarding the posterior teeth, striations onthe lingual side are restricted to the upper third of thecrowns. Pyrenasaurus can be differentiated from mostknown lizards by its low tooth count, exceptions includ-ing all amphisbaenians, some scincids, some lacertids,some anguimorphs, xantusiids, pygopods and some fossilforms probably related to scincoids and teiioids (see dis-cussion). It can be easily differenciated from all these
Fig. 1. Map of Western Europe with Paleogene localities mentionedin the text.; B: Berru; Bo: Boutersem; H: Hainin; M: Messel; Pr:Pr�emontr�e. Escamps is one of many Phosporites du Quercy localities.The star serves to situate Malp�eri�e, Le Bretou and Les Pradigues.
506 BOLET AND AUG�E

TABLE 1. List of taxa from selected late Eocene localities (data from Aug�e, 2005 and Bolet and Evans, 2013)
SOSS�IS MP17A MALP�ERI�E MP17 ESCAMPS MP19
IguaniaIguanidae
Geiseltaliellus lamandini Geiseltaliellus lamandiniGeiseltaliellus
pradiguensisGeiseltaliellus pradiguensis
Geiseltaliellus sp.Cadurciguana hoffstetteri Cadurciguana hoffstetteriPseudolacerta mucronata Pseudolacerta mucronata
Pseudolacerta sp.Gekkota
Cadurcogekko piveteaui Cadurcogekko piveteauiGekkota indet. Form AGekkota indet. Form BGekkota indet. Form C
LacertiformesLacertidae
Plesiolacerta lydekkeri Plesiolacerta lydekkeriQuercycerta maxima Quercycerta maxima
Gracilicerta sindexi Gracilicerta sindexi Gracilicerta sindexiEscampcerta amblyodonta
Dormaalisaurusgirardoti
Dormaalisaurusrossmanni
?Lacertidae indet.(amblyodont form)
?TeiidaeBrevisaurus smithi
ScincoideaScincoidea indet. Form Aa
Scincoidea indet. Form B Scincoidea indet. Form BScincidae
Orthoscincus malperiensis?Scincidae
Pyrenasaurus evansaeb Pyrenasaurus evansae?Scincidae indet.
CordyliformesCordyliformes indet.
?CordylidaeEocordyla mathisi
AnguimorphaAnguidae
GlyptosaurinaePlacosaurus sp.
Glyptosaurinae indetGlyptosaurini indet. Glyptosaurini indet.
AnguinaeHelvetisaurus picteti
Anguis sp. Anguis sp.Anguinae indet.
Form AAnguinae indet.
Form BPlatynota
Necrosaurus eucarinatus Necrosaurus eucarinatusNecrosaurus cayluxi Necrosaurus cayluxi
HelodermatidaeEurheloderma gallicum Eurheloderma gallicum
AmphisbaeniaBlanidae
cf. Blanidae indet. Blanidae indet. Blanidae indet.Squamata indet.
Squamata indet.c
aScincomorpha indet. Form A in Aug�e (2005).b?Scincoidea incertae sedis, unnamed new genus and species in Bolet and Evans (2013).cLacertilia incertae sedis in Aug�e (2005).
LIZARD FROM THE EOCENE OF FRANCE AND SPAIN 507
taxa with reduced tooth counts by the characteristicshape of its posterior teeth, as well as an unusual den-tary morphology, with a closed and fused Meckeliancanal, a straight ventral border, a symphysis that doesnot form a sudden angle with it, and a moderately highcoronoid process.
Description
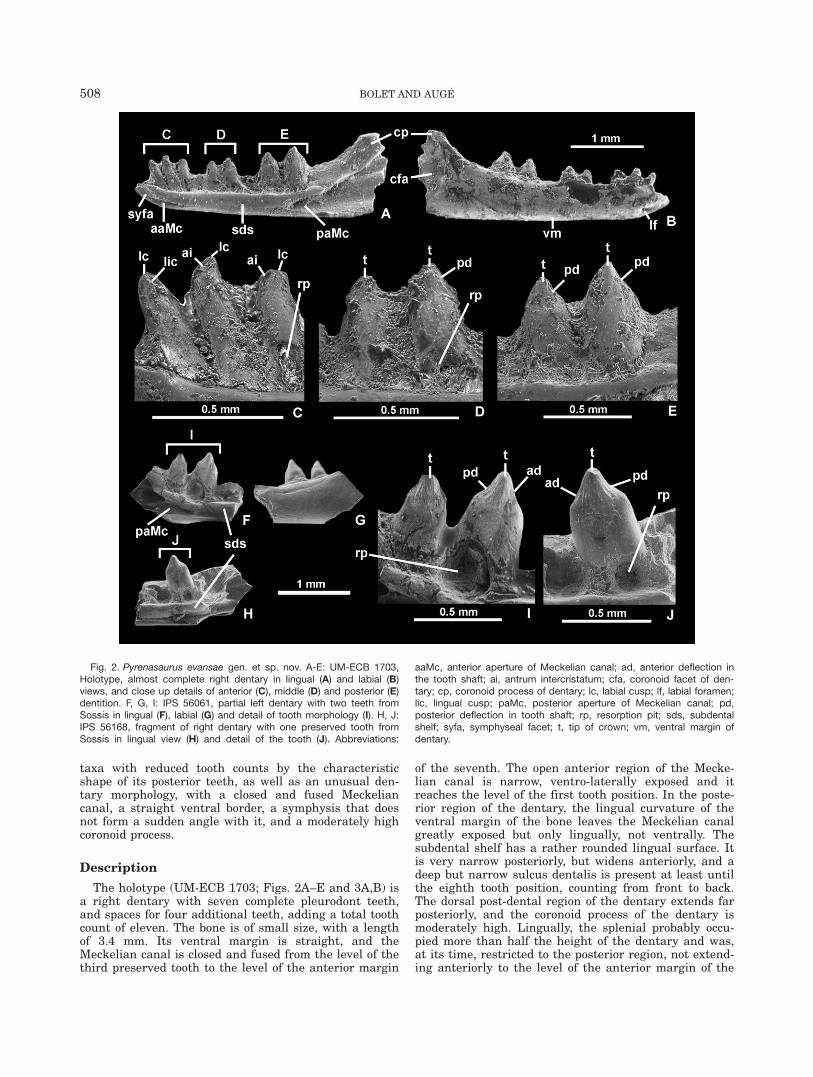
The holotype (UM-ECB 1703; Figs. 2A–E and 3A,B) isa right dentary with seven complete pleurodont teeth,and spaces for four additional teeth, adding a total toothcount of eleven. The bone is of small size, with a lengthof 3.4 mm. Its ventral margin is straight, and theMeckelian canal is closed and fused from the level of thethird preserved tooth to the level of the anterior margin
of the seventh. The open anterior region of the Mecke-lian canal is narrow, ventro-laterally exposed and itreaches the level of the first tooth position. In the poste-rior region of the dentary, the lingual curvature of theventral margin of the bone leaves the Meckelian canalgreatly exposed but only lingually, not ventrally. Thesubdental shelf has a rather rounded lingual surface. Itis very narrow posteriorly, but widens anteriorly, and adeep but narrow sulcus dentalis is present at least untilthe eighth tooth position, counting from front to back.The dorsal post-dental region of the dentary extends farposteriorly, and the coronoid process of the dentary ismoderately high. Lingually, the splenial probably occu-pied more than half the height of the dentary and was,at its time, restricted to the posterior region, not extend-ing anteriorly to the level of the anterior margin of the
Fig. 2. Pyrenasaurus evansae gen. et sp. nov. A-E: UM-ECB 1703,Holotype, almost complete right dentary in lingual (A) and labial (B)views, and close up details of anterior (C), middle (D) and posterior (E)dentition. F, G, I: IPS 56061, partial left dentary with two teeth fromSoss�ıs in lingual (F), labial (G) and detail of tooth morphology (I). H, J:IPS 56168, fragment of right dentary with one preserved tooth fromSoss�ıs in lingual view (H) and detail of the tooth (J). Abbreviations:
aaMc, anterior aperture of Meckelian canal; ad, anterior deflection inthe tooth shaft; ai, antrum intercristatum; cfa, coronoid facet of den-tary; cp, coronoid process of dentary; lc, labial cusp; lf, labial foramen;lic, lingual cusp; paMc, posterior aperture of Meckelian canal; pd,posterior deflection in tooth shaft; rp, resorption pit; sds, subdentalshelf; syfa, symphyseal facet; t, tip of crown; vm, ventral margin ofdentary.
508 BOLET AND AUG�E

penultimate tooth. The lingual anterior process of thecoronoid was probably restricted to a position far behindthe tooth row (no facet is preserved in the lingual sur-face of the conoroid process of the dentary). The exten-sion of the posteroventral process of the dentary cannotbe ascertained because it is broken. In labial view, thereis one large foramen situated at the level of the thirdtooth position (the first preserved), and the presence ofadditional posteriorly positioned foramina cannot be dis-carded (not clear because the surface is not completelyclean). The large triangular facet for the anterolateralprocess of the coronoid is restricted to a posterior posi-tion, far from the posteriormost-situated tooth.
The dentition gradually increases its robustness, frommoderate (anterior region) to very robust (posteriorregion). The height of the preserved teeth slightlyincreases from the anterior to the posterior region, withthe crowns projecting above the parapet of the dentaryapproximately one half of the tooth height. The anteriorteeth are slighlty more anteriorly inclined than the pos-terior ones. The first three teeth (Fig. 2C; their morphol-
ogy is best seen in the second one) are cylindrical, witha slightly posterolingually recurved tip. The morphologyof the anterior teeth is not clear because it has not beenpossible to clean the matrix that covers them, but thesecond and probably the third tooth present the typicalscincoid pattern (sensu Richter, 1994). Although it is notpossible to observe the morphology in detail, it is clearthat, apart from the lingual cusp, there is a second labi-ally positioned cusp (cuspis labialis), and also a well-developed antrum intercristatum. There is an increasein the size of the middle (Fig. 2D) and posterior (Fig.2E) teeth, with the posteriormost tooth being the largestone. The last preserved teeth present a distinctiveshape: they are labiolingually compressed, with a flat-tened lingual surface (Fig. 2E). The crowns are badlyworn in this specimen, so the characteristic shape of theposterior dentition of the taxon is mainly based on obser-vations made on other specimens (Fig. 2F–J, see below).Allowing for the worn and partially covered surface ofsuch teeth, the crowns appear to present a deflection inthe convexity of at least the posterior margin of the
Fig. 3. Drawings of fossil (A–E) and extant (F–J) selected taxa forcomparison purposes. A, B: Pyrenasaurus evansae gen. et sp. nov.from Escamps (late Eocene, France); C: Scincoidea indet. Form Bfrom the late Eocene of Malp�eri�e (late Eocene, France); D: Orthoscin-cus malperiensis (Scincidae) from Malp�eri�e; E: Tiliqua pusilla (Scinci-dae, Lygosominae) from Riversleigh (Miocene, Australia). F: Dibamus
noveaguineae (Dibamidae); G: Blanus strauchi (Blanidae); H: Vanzo-saura rubricauda (Gymnophthalmidae); I: Acontias percivali (Scincidae,Acontinae); J: Cyclodomorphus branchialis (Scincidae, Lygosominae).A, B and G original. Others redrawn from: C, D, Aug�e (2005); E, Sheaand Hutchinson (1992); F, Greer (1985); H, Guerra and Montero (2009);I, Kosma (2004); J, Estes and Williams (1984). E, F, H and J reversed.
LIZARD FROM THE EOCENE OF FRANCE AND SPAIN 509

tooth. It is not discarded that the anterior margin wasalso originally deflected, but poorly preserved and cov-ered by matrix. This gives the crowns the characteristicresemblance to a short obelisk, more clearly observablein the Soss�ıs specimens (Fig. 2I,J). The presence of stria-tions in the lingual side of the holotype teeth cannot beascertained, but this could be related to a certain degreeof wear. Large, posteriorly preserved resorption pits arepreserved at the bases of some tooth positions, mainlythe fifth and eighth.
Referred Specimens
The referred posterior part of dentary fragments (IPS56168 and IPS 56061; Fig. 2F–J) present exactly thesame bone and tooth morphology, and it is possible thatthey represent both sides of a single individual. Theteeth are stout and labiolingually compressed, and showa very characteristic outline in lateral view. The anteriorand posterior margins converge at an angle of 16� (closeto parallel) in their bases, but there is an abrupt changeto a more obtuse angle (80�) passed the mid shaft of thetooth. This gives a short obelisk profile, except that inthis region, the anterior and posterior margins present adeflection in their convexity, rather than being straight(Fig. 2I,J). The tip is slightly lingually recurved. Thereare striations only in the lingual side of the tip, and onlyoccupying the upper third of the crown. The Meckeliancanal is open in the preserved region, but its marginsapproach each other (Fig. 2F,H) and may have been incontact or fused in a more anterior region (not pre-served). A resorption pit is preserved at the base of thepenultimate tooth, being large and posterolinguallypositioned.
The French and Spanish specimens are indistinguish-able and they are referred to the same species P. evan-sae. Admittedly, the taxon is known only fromfragmentary material at Soss�ıs. Hence, it is possible thatthe specimens from Soss�ıs represent a distinct speciesbut until more fossils appear, showing possible signifi-cant differences, we decline to name a new species.
DISCUSSION
Morphology and Systematics
The holotype is very distinctive with respect to anyother known lizard, fossil or extant, and the two referredspecimens provide detail on the unique tooth morphol-ogy of the posterior dentition. We are aware thatdescribing a new genus based on dentary material aloneoften generates problems regarding the referral of poten-tial new material. However, it is clear that the dentitionof Pyrenasaurus is so unique that it will be easily recog-nizable even among very fragmentary material. In fact,attribution to the same taxon of additional materialfrom a second locality (Soss�ıs) appears secure even beingbased on very fragmentary specimens. The peculiar den-tition of this taxon might be useful to recognize addi-tional material from other tooth bearing bones (i.e.,maxilla and perhaps the premaxilla). Regarding theaffinities of the new taxon, while its status as a newgenus of lizard is easy to prove, its exact relationshipswithin Squamata are more difficult to elucidate. Theobservable combination of characters is not characteris-tic of any concrete lizard group, but the potential affin-
ities of the taxon can be discussed in terms of theirpresence or absence in such groups. A reduced toothcount in the dentary is unknown for iguanians, and thepresence of a sulcus dentalis, the high coronoid processof the dentary, the presence of a right angle between theparapet of the dentary and the sudbental shelf all argueagainst anguimorphan affinities. A reduced tooth countis rare among gekkotans (an exception would be Apra-sia; Parker, 1956), and this group presents reduced sym-physeal facets in comparison to those of Pyrenasaurus.Dibamids are interesting in presenting a jaw configura-tion similar to that of Pyrenasaurus, although theirteeth are much higher and pointed (Fig. 3F), and thesymphysis is clearly different (Meckelian canal open inthis part of the dentary of Pyrenasaurus). Amphisbae-nians are strictly burrowing forms probably related tolacertoids (e.g., Wiens et al., 2012), strongly differingfrom Pyrenasaurus in tooth morphology and jaw configu-ration. The maximum dentary tooth count is nine amongextant forms, but two new taxa from the Paleocene ofEurope referred to Amphisbaenia (Folie et al., 2013) pos-sess 10–12 teeth. However, their exact affinities willhave to wait a complete description of the material andthe tooth shape is clearly different from that of Pyrena-saurus, the teeth being, as well as the dentary, muchmore robust in the former (A. Bolet, personal observa-tion). The general morphology of later (Early Eoceneonwards) European amphisbaenian dentaries is distinc-tive and different from Pyrenasaurus, mainly regardingthe slender shape of the latter in contrast to more robustdentaries in European amphisbaenians (e.g., Blanus,Fig. 3G), which otherwise present a characteristic shapeof the symphysis, as well as an open Meckelian canal forall their length (closed and fused in Pyrenasaurus).Again, the tooth shape of Pyrenasaurus is quite differentfrom known amphisbaenians. Cryptolacerta, a furtherlimb-reduced form from the middle Eocene of Germany(M€uller et al., 2011), is clearly different from Pyrenasau-rus in terms of size, dentary length, robustness andshape, and tooth count and morphology, among othercharacters. This leaves “Scincomorpha” (Scin-coids1Lacertiforms) as the group in which the taxon ismost easily accommodated. Note, however, that thistaxon has been regarded as a paraphyletic assemblagein most molecular analyses (e.g., Vidal and Hedges,2005; Wiens et al., 2012; Pyron et al., 2013). Among lac-ertiforms, lacertids present certain homogeneity withjaws characterized by arched dentaries with an openMeckelian canal, a lateral facet for the articulation ofthe coronoid and with posterior bicuspid or tricuspidteeth. Some lacertids, however, present strong feedingadaptations including reduction of tooth count, such asDracaenosaurus (from the Eocene/Oligocene of France;e.g., M€uller, 2004; Aug�e, 2005), but the shape of the den-tary still shows a clearly lacertid morphology, and inthis case its massive teeth represent an adaptation to adurophagous diet, and no adaptations to fossoriality aresupposed. Lacertiform teiids show a great variabilityregarding tooth shapes, but a reduced tooth count isvery rare, except in some Late Cretaceous forms classi-fied as polyglyphanodontids (e.g., Adamisaurus; Sulim-ski, 1972), and the lack of cementum at the base of theteeth of Pyrenasaurus, among other characters, arguesagainst a referral to Teiioidea (Estes et al., 1988). Sometupinambid teiids would represent modern examples of
510 BOLET AND AUG�E

durophagous taxa, and again their reduced dentarytooth count (e.g., 12 in Dracaena; Kosma, 2004) is notrelated to fossoriality (Montero et al., 2004). Xantusiidspresent a reduced tooth-count (12–14 in Xantusia,Hecht, 1956), but they differ from Pyrenasaurus in pre-senting a Meckelian canal that is fused to a greaterextent (both anteriorly and posteriorly), as well as avery different tooth morphology and configuration of theposterior region of the dentary.
Gymnophthalmids sometimes present a closed andfused Meckelian canal, but their tooth counts are higherthan in Pyrenasaurus, and their long lower jaws presenta very different configuration (e.g., Vanzosaura, seeGuerra and Montero, 2009 and Fig. 3H) regarding, forexample, the extent of the lingual anterior process of thecoronoid. Among “scincomorphans,” scincoids (scin-cids1cordylids), and specially scincids, are the groupthat shows a greater variability in the tooth shape andthe one in which tooth count reduction is more wide-spread. Extant representatives of Scincidae are tooabundant and diverse to be discussed here in detail, butit is worth mentioning that a closed and fused Meckeliancanal is widespread among acontine (e.g., Acontias, Fig.3I) and lygosomine (e.g., Tiliqua, Fig. 3E; Cyclodomor-phus, Fig. 3J) scincids. The only example of a toothshape that vaguely recalls the posterior dentition of Pyr-enasaurus is that of the lygosomine Egernia stokesii, asfigured in Kosma (2004, Pl. V, Fig. 11), but similarity isrestricted to the outline of the tooth, and none of thecharacteristic features of the posteriormost teeth of Pyre-nasaurus are observable in this taxon suggesting thatthey are not closely related forms.
The overall shape of the dentary of Pyrenasaurus(Figs. 2A,B and 3A,B), the great reduction of the toothcount, as well as the presence of a well-developedantrum intercristatum in the anterior teeth (Fig. 2C),together with a characteristic tooth shape in the poste-rior region (Fig. 2D–F,I,J), all suggest that Pyrenasaurusis more likely a scincid than a cordylid, because thesefeatures are not known among the latter. Most scincidspresent a generalized scincoid dentition (sensu Richter,1994), although the presence of characteristic teeth insome forms usually highlights adaptations to eating agiven food item (e.g., durophagy, see Estes and Williams,1984). The Paleogene scincoids are generaly regarded asbasal (e.g., Folie et al., 2005), but without testing thephylogenetic position of Pyrenasaurus, we cannot be pos-itive about its affinities with any clade. These forms canbe easily differentiated by their higher tooth count,apart from the lack of the distinctive posterior teeth ofPyrenasaurus, and include Scincoideus haininensis Folieet al., 2005 (Hainin, MP1-5, Belgium) and Ornatocepha-lus metzleri Weber, 2004 (Messel, MP11, Germany). Thenamed Paleogene European scincids are Orthoscincusmalperiensis Aug�e, 2005 (Malp�eri�e, MP17, France; Fig.3D) and Axonoscincus sabatieri Aug�e, 2003 (Pr�emontr�e,MP10, France); and a possible cordylid is Eocordylamathisi Aug�e, 2005 (Le Bretou, MP16 and Malp�eri�e andLes Pradigues, MP17, France). Foliesaurus boutersemen-sis Aug�e and Smith, 2009 (Boutersem, MP21, Belgium)is probably a scincid, and Berruva louisi Aug�e, 2005(Berru, MP6, France), described as a scincid, has alter-natively considered as a basal scincoid (Aug�e and Smith,2009). All of them possess the typical scincoid tooth pat-tern (sensu Richter, 1994). Admittedly, the shape of the
posterior teeth of Pyrenasaurus is not the paradigm forscincids or scincoids. However, a great number of formswith specialized dentitions are known among scincoids(e.g., Kosma, 2004; Aug�e, 2005), and the anterior teethmorphology fits well with this group. The most similarof the taxa listed above, regarding the overall shape andsize of the dentary, is the contemporaneous Orthoscincus(Fig. 3D). Among the characters common in both formsare the short dentary, the reduced tooth count and theelevated coronoid process of the dentary. However, Pyre-nasaurus differs from Orthoscincus in having a lowertooth count (11 vs. 14–15 in Orthoscincus), enlarged pos-terior teeth (homodont and generally smaller dentitionin Orthoscincus), a distinctive tooth crown (contra typi-cal scincoid shape in Orthoscincus) in the posteriorregion, a closed Meckelian canal (open in Orthoscincus),and a straight ventral border of the dentary (slightlyarched in Orthoscincus). Other specimens referred toindeterminated “scincomorphs” are present in PaleogeneFrench localities (see Aug�e, 2005), but one among themis particularly interesting for comparison with Pyrena-saurus. This specimen, USTL MAL 605, is fromMalp�eri�e (MP17, France, Fig. 3C), was referred to“Scincomorpha ind�etermin�e Forme B” in Aug�e (2005)and, despite some differences, it shares with Pyrenasau-rus, among other characters, a similar overall morphol-ogy of the dentary, a low tooth count, the enlargement ofthe posterior teeth, and an unusual posterior toothshape where the tip is somehow isolated by two lateralridges. The subdental shelf has a high lingual surface,which gradually diminishes after the eighth dental posi-tion. In this aspect, it shares with Pyrenasaurus the clo-sure of the Meckelian canal, with a restricted andmostly ventral opening situated in the anterior part ofthe dentary, and a wide mostly lateral exposure in itsposterior part. However, a subtle suture that suggeststhat the fusion was not complete, still remains. The lackof the posterior end of the dentary precludes any infor-mation regarding the extent of the coronoid process ofthe dentary. The main difference with the holotype ofPyrenasaurus is the shape of the posteriormost teeth,with the tip being more clearly isolated, giving them atricuspid appearance. Moreover, the tooth implantationseems to be less deep. The morphological similaritiesnoted above suggest affinities between Pyrenasaurusand USTL, MAL 605, but at the same time, the noteddifferences, mainly regarding the dentition, denote thatthey may represent not only different species, but alsoprobably different (although possibly related) genera. Anunnamed cordyliform and a possible scincid, both differ-ent from Pyrenasaurus on the basis of tooth shape andopen Meckelian canal, were reported from Soss�ıs (Boletand Evans, 2013), suggesting that scincoids were notrare in the late Eocene of the Southern PyreneanBasins, and the same could be applied to the Phosphor-ites du Quercy (see Aug�e, 2005).
Lifestyle
Some features of the dentary of Pyrenasaurus suggestit could represent a burrowing or surface-dwelling liz-ard. Among the characters that support this are minia-turization, the short length of the bone with a lownumber of teeth, and the high coronoid process of thedentary. Although the fusion of the Meckelian groove
LIZARD FROM THE EOCENE OF FRANCE AND SPAIN 511

appears in some non-fossorial lizards, this character sug-gests burrowing habits when it appears in combinationwith miniaturization and decrease of tooth number (Bro-schinski and Sigogneau-Russell, 1996). Adaptations tofossoriality appear early in the fossil record, with exam-ples of burrowing forms reported from the Late Jurassic(e.g., Saurillodon from Guimarota, Portugal; Kosma,2004) and the Early Cretaceous (e.g., Tarratosaurusfrom Anoual, Morocco; Broschinski and Sigogneau-Russell, 1996). Jucaraseps, a tiny lizard from the EarlyCretaceous of Spain, has been interpreted as an earlyexample of a surface-dweller lizard (Bolet and Evans,2012), and Chalcidosaurus from Guimarota as a semi-fossorial form (Kosma, 2004). Because the fossil recordonly rarely preserves complete specimens, it is often notpossible to discern between true burrowing forms andsurface-dwellers, because relative length of the limbsand tail cannot be measured. This is the case of Pyrena-saurus, which, although it presents some jaw adapta-tions for a fossorial or semifossorial lifestyle, it isdifficult to state to which of the two groups it belongedto in absence of postcranial material.
Although small lizards weighting less than one gramare said to lack the strength to burrow (Benesch andWithers, 2002), the dentary of Pyrenasaurus is of similarsize to that of recent Blanus, a fully burrowing form,and thus it cannot be discarded that Pyrenasaurus wasa burrower only based on its size. An additional charac-ter that has been related to fossoriality is the presenceof an angle at the symphysis of the dentary, as presentin most amphisbaenians (e.g., Gans, 1974). However, itis important to note that Gans (1974) already statedthat not all amphisbaenians present it, and examples ofextant (e.g., Amphisbaena ridleyi; Pregill, 1984) and fos-sil (e.g., Palaeoblanus; Schleich, 1988; B€ohme, 1999)taxa lacking this character exist. Moreover, some lizardsdescribed as fossorial forms (e.g., the gymnophthalmidsCalyptommatus nicterus, Scriptosaura catimbau, andNothobachia ablephara) also seem to lack this character(see figures in Roscito and Rodriges, 2010). Pyrenasau-rus presents a subtle angle at the level of the symphysis,but it is not comparable to that existing in most amphis-baenians. However, because of the exceptions notedabove, this is not an argument against an interpretationof Pyrenasaurus as a fossorial lizard. At least half of theorigins of the burrowing morph occur in one group, theScincidae (Wiens et al., 2006).
The peculiar tooth shape of Pyrenasaurus suggests anadaptation to a determinate kind of food. However, sincethis shape is unique among known taxa, it is not possi-ble to ascertain to which items it can be related. Thescincid genus Acontias (Fig. 3I), has a jaw configurationthat presents overall similarities to that of Pyrenasaurus(see Kosma, 2004), including the low tooth count (ten inA. percivali), the closed and fused Meckelian canal(although it is closed for a longer way) and the great dis-tance between the last tooth and the coronoid process ofthe dentary. However, differences in tooth morphologyand the shape of the symphysis and the dentary in gen-eral suggest that both genera are not related, and super-ficial similarities should be regarded as convergencesdue to a similar lifestyle. Thus, the observations madefor Acontias percivali as a surface-dweller in fallenleaves and feeding on insects (Rogner, 1994), mightapply to Pyrenasaurus. Most species of the genus feed
on beetle larvae and termites (Broadley and Greer,1969), but two of the species feed, in addition to differentkinds of insects, on other squamates. Acontias plumbeusconsumes vertebrates such as other burrowing skinksand frogs (Broadley and Greer, 1969) and A. meleagrisfeeds upon worm snakes (Typhlops sp.) and juveniles ofits own species (Hauschild and Gabner, 1995). It is clearthat the overall similarities between Pyrenasaurus andAcontias are not enough to warrant the same lifestyleand, due to its small size, Pyrenasaurus would havebeen more easily a prey than a predator, at least regard-ing other vertebrates.
Pyrenasaurus was not the only squamate presentingadaptations to fossoriality found at Soss�ıs and Escamps.Amphisbaenians (most probably blanids) have beenreported from both localities (Aug�e, 2012; Bolet andEvans, 2013). The slender shape of the dentary suggeststhat the taxon was not so strongly adapted for fossorial-ity as, for example, an amphisbaenian. Curiously, theposition of the last labial foramen, just at the level ofthe anterior edge of the last tooth, was reported as anunusual feature present in an indeterminated scincoidfrom Malp�eri�e and Escamps (Scincoidea indet. Form Bof Aug�e, 2005), and this unnamed taxon also possesses acharacteristic posterior dentition, consisting in this caseof enlarged spatulated teeth. It seems that the Eocenewas a period during which scincoids radiated in Europe(see Aug�e, 2005) resulting in a variate collection ofbizarre forms. These forms disappear at the GrandeCoupure, and scincoids that appear after the Eocene–Oligocene limit (e.g., Foliesaurus from the Early Oligo-cene of Boutersem, Belgium; Aug�e and Smith, 2009) areclearly different from them, and are closer to the typicalscincoid pattern regarding the shape of both the denti-tion and the dentary. Because Pyrenasaurus is notclearly related to any known fossorial squamate, itseems to represent a further example of a lizard thathas independently achieved adaptations for burrowing/surface-dwelling (see Table 2 for characters seen in someselected fossorial lizards). Because many fossil forms aregood candidates to represent independent adaptations tofossoriality, their identification is important in order toestimate the plasticity of this treat among squamates,and will probably show that these independent lineagesare more widespread than what we have been able torealize up to now. It is clear, however, that the lack ofarticulated specimens (specimens from Messel being anotable exception) or at least associated postcranial ele-ments in most Paleogene localities clearly hamper dis-cussion on this issue.
Comparison of the Soss�ıs Lizard Assemblage(Spain) With Those From Malp�eri�e andEscamps (France)
To assess if the Southern Pyrenean basins (repre-sented by Soss�ıs as the single late Eocene lizard assem-blage described from the Iberian Peninsula so far, seeBolet and Evans, 2013) present a similar faunal compo-sition to contemporaneous French localities, two repre-sentative French lizard assemblages have been chosen:Malp�eri�e (MP17), and Escamps (MP19). As seen in theresulting table (Table 1), both French localities share agreat number of taxa, some of them even at the specieslevel. Pleurodont iguanians are represented by three
512 BOLET AND AUG�E

different genera at the Phosphorites du Quercy: Cadur-ciguana, Geiseltaliellus and Pseudolacerta. Of them,only Cadurciguana seems to be absent from Soss�ıs,while Geiseltaliellus and Pseudolacerta are clearly pres-ent (Bolet and Evans, 2013). Gekkotans are representedin Malp�eri�e and Escamps only by Cadurcogekko. Threegekkotan taxa have been identified from Soss�ıs, one ofthem possibly related to (or at least not necessarily dif-ferent from) Cadurcogekko (see Bolet and Evans, 2013).Regarding the lacertids, the persistance of Dormaalisau-rus up to the MP17 at Soss�ıs can be interpreted as alate occurrence of the genus in contrast to a previousdisappearance of the taxon from more northern assemb-lages, as stated by its absence from all post-MP16French localities (Aug�e, 2005). Plesiolacerta seems to beabsent from Soss�ıs. Quercycerta and Escampcerta areboth absent from Soss�ıs, but they are taxa with arestricted distribution even within the Phosphorites duQuercy (only reported from Malp�eri�e and Escampsrespectively). Gracilicerta sindexi, however, is a taxonshared by the three localities. Regarding the scincoids,the scincid Orthoscincus has been only reported fromMalp�eri�e, while cordyliforms are represented by the?cordyloid Eocordyla at Malp�eri�e (Aug�e, 2005) and aprobably different cordyliform and a possible scincidfrom Soss�ıs (Bolet and Evans, 2013).
Even if we take the contemporaneous localities fromthe Phosphorites du Quercy, we find some variations inthe composition of the lizard assemblages, with sometaxa (even at the genus level) only reported from thetype locality or at most restricted to a few sites. Theabsence of such taxa at Soss�ıs cannot be argued to repre-sent an actual difference between the French localitiesand those of the Southern Pyrenean basins because theyare variably present even among the Phosphorites duQuercy localities. These taxa include Escampcerta, Quer-cycerta, Orthoscincus, Eocordyla, an indeterminate“scincomorph” (here referred to Scincoidea indet. FormA) and an indeterminate lizard (referred to Lacertiliaincertae sedis in Aug�e, 2005). Among the genera sharedby Soss�ıs and at least one of the two French selectedlocalities are Geiseltaliellus, Pseudolacerta, a gekkotan
(if it actually represents Cadurcogekko) and Gracilicerta.Among the taxa not identified at the genus level butthat could still represent shared taxa between Soss�ısand at least one of the two selected French localities arean indeterminate amphisbaenian (probably referable toBlanidae), a glyptosaurine, and an anguine. If we exam-ine which taxa are shared by the two French localitiesbut not by Soss�ıs, this is reduced to Cadurciguana, Ple-siolacerta, Eurheloderma, and Necrosaurus. Note that,with the exception of Cadurciguana, they correspond tomainly large taxa, and such large lizards were inter-preted as underrepresented at Soss�ıs (Bolet and Evans,2013), although the reason is not clear. Pyrenasaurus is,then, another shared taxon between Soss�ıs and one ofthe French localities belonging to the Phosphorites duQuercy. The future description of middle and late Eocenelizard assemblages from the different basins of the Ibe-rian Peninsula, and their comparison with those fromSoss�ıs and the Phosphorites du Quercy, are intended toclarify if these similarities between the assemblagesfrom North and South of the Pyrenees are extensive tothe rest of the Iberian Peninsula or, if as suggested bymammals, they prove to be different enough to argue infavour of the existence of the so-called Western IberianBioprovince.
CONCLUSIONS
The new taxon here described is an example of ahighly apomorphic form that, despite being representedby few isolated specimens, adds information to taxonom-ical, biogeographical, and ecomorphological discussionson Eocene squamates from Europe. Pyrenasaurus is situ-ated at both versants of the Pyrenees, supporting thehypotheses that link the Southern Pyreneean basins fau-nas to those from France, although herpetological proofsin favour or against the presence of an endemic WesternIberian Bioprovince need to wait until the description ofCentral and Western Spanish assemblages. We suggestit could also represent a further example of a lineagethat has independently adapted to a fossorial lifestyle,either as a burrowing form or as a surface-dweller, as
TABLE 2. Comparisons between selected miniaturized lizards
TaxonDentary
tooth countTooth shape
(anterior/posterior) SymphysisMeckelian
canalSize (dentary
length) Lifestyle
Dibamus novaguineae(Dibamidae)
10 Pointed, stronglyrecurved, unicuspid
Angled Closed 4 mm Fossorial
Blanus strauchi(Amphisbaenia,Blanidae)
7–9 Pointed, slightlyrecurved, unicuspid
Angled Open 3–4 mm Fossorial
Vanzosaura rubricauda(Gymnophthalmidae)
19–21 Unicuspid Angled Closed 3 mm Fossorial
Acontias percivali(Scincidae, Acontinae)
10 Unicuspid, scincoidmorphology
Angled Closed ca. 6 mm Surface-dwelling
Cyclodomorphusbranchialis(Scincidae,Lygosominae)
12 Unicuspid, robust,scincoidmorphology
Rounded Closed >9 mm Surface-dwelling
Amphisbaena ridleyi(Amphisbaenia,Amphisbaenidae)
6–7 Pointed, unicuspid/amblyodont
Barely angled Closed 7.5 mm Semi-fossorial
Pyrenasaurus evansae(Scincoidea,?Scincidae)
11 Unicuspid, scincoidmorphology/obeliskprofile
Barely angled Closed 3.4 mm ?Fossorial
LIZARD FROM THE EOCENE OF FRANCE AND SPAIN 513

suggested by the shape of its dentary and its low toothcount. We tentatively refer this new taxon to Scincoideaand suggest scincid affinities, although its phylogeneticrelationships are far from being clear.
ACKNOWLEDGEMENTS
The authors thank B. Marandat (USTL, France) foraccess to the material from Escamps. They also thankS. Moy�a-Sol�a (ICP, Catalonia), for providing access tothe material from Soss�ıs, and J. Marig�o and R. Minwer-Barakat (ICP) for information on Eocene material andlocalities. J.-C. Rage (Mus�eum national d’Histoirenaturelle, France) kindly assisted A. Bolet in his visit atcollections, and provided important information regard-ing French material and localities. The comments ofJ.D. Daza (Villanova University, USA), A. �Cer�nansk�y(Geological Institute, Slovak Academy of Sciences, Slo-vak Republic), Annelise Folie (Royal Belgian Institute ofNatural Sciences, Belgium) and S.C. Miller (Universityof Utah School of Medicine, USA) greatly improved themanuscript.
LITERATURE CITED
Aug�e M. 2003. La faune de Lacertilia (Reptilia, Squamata) del’�Eocene inf�erieur de Pr�emontr�e (Bassin de Paris, France). Geodi-versitas 25:539–574.
Aug�e M. 2005. �Evolution des l�ezards du Pal�eogene en Europe. MemMus Natl Hist Nat 1–369, CD-ROM.
Aug�e M. 2007. Past and present distribution of iguanid lizards. ArqMus Nac (Rio de Janeiro) 65:403–416.
Aug�e M. 2012. Amphisbaenians from the European Eocene: a bio-geographical review. Palaeobio Palaeonv 92:425–443.
Aug�e M, Smith R. 2009. An assemblage of early oligocene lizards(Squamata) from the locality of Boutersem (Belgium), with com-ments on the Eocene-Oligocene transition. Zool J Linn Soc 155:148–170.
Badiola A. 2004. Estudio paleontol�ogico del yacimiento del EocenoSuperior de Zambrana (�Alava, regi�on Vasco-Cant�abrica).Tafonom�ıa, paleobiolog�ıa de mam�ıferos e implicaciones biocro-nol�ogicas. PhD Dissertation. Universidad del Pa�ıs Vasco, Espa~na.
Badiola A, Cuesta MA. 2008. New endemic eocene equoids from theIberian Peninsula (Western Europe). J Vertebr Paleontol 28:1149–1161.
Badiola A, Checa L, Cuesta MA, Quer R, Hooker JJ, Astibia H.2009. The role of new Iberian finds in understanding EuropeanEocene mammalian paleobiogeography. Geol Acta 7:243–258.
Benesch AR, Withers PC. 2002. Burrowing performance and therole of limb reduction in Lerista (Scincidae, Lacertilia). SenckenbLethaea 82:107–114.
B€ohme M. 1999. Doppelschleichen (Sauria, Amphisbaenidae) ausdem Untermioz€an von Subersheim 3 (S€uddeutschland). MittBayer Staatssaml Pal€aont Hist Geol 39:85–90.
Bolet A, Evans SE. 2012. A tiny lizard (Lepidosauria, Squamata)from the Lower Cretaceous of Spain. Palaeontology 55:491–500.
Bolet A, Evans SE. 2013. Lizards and amphisbaenians (Reptilia,Squamata) from the late Eocene of Soss�ıs (Catalonia, Spain).Palaeontol Electron 16.1.8A.
Broadley DG, Greer AE. 1969. A revision of the genus AcontiasCuvier (Sauria: Scincidae). Arnoldia 36:1–20.
Broschinski A, Sigogneau-Russell D. 1996. Remarkable lizardremains from the lower Cretaceous of Anoual (Morocco). AnnPaleontol (Vert-Invert) 82:147–175.
Casanovas-Cladellas ML, Santaf�e-Llopis JV. 1987. Cantabrotheriumtruyolsi nov. gen. nov. sp. (Palaeotheriidae, Perissodactyla) unexemple d’end�emisme dans le Pal�eogene ib�erique. M€unchner Geo-wiss Abh R A: Geol Pal€aontol 10:243–252.
Cuesta Ru�ız-Colmenares MA. 1991. Perisod�actilos del Eoceno de laCuenca del Duero. PhD Dissertation. Universidad de Salamanca,Espa~na.
Estes R, Williams EE. 1984. Ontogenetic variation in the molari-form teeth of lizards. J Vertebr Paleontol 4:96–107.
Folie A, Sig�e B, Smith T. 2005. A new scincomorph lizard from thePalaeocene of Belgium and the origin of Scincoidea in Europe.Naturwissenschaften 92:542–546.
Folie A, Smith R, Smith T. 2012. New basal amphisbaenian lizardsfrom the Middle Paleocene of Hainin and its implication in theevolution of European amphisbaenians. 4th International Geolog-ica Belgica Meeting 2012. Moving Plates and Melting Icecaps.Processes and Forcing Factors in Geology. Brussels: GeologicaBelgica abstracts. S8.1.
Folie A, Smith R, Smith T. 2013. New amphisbaenian lizards fromthe Early Paleogene of Europe and their implications for the earlyevolution of modern amphisbaenians. Geol Belg 16:227–235.
Gans C. 1974. Biomechanics, an approach to vertebrate biology.Philadelphia: J.B. Lippincott Company.
Gray JE. 1825. A synopsis of the genera of reptiles and amphibia,with a description of some new species. Ann Phil 10:193–217.
Greer AE. 1985. The relationships of the Lizard genera Anelytropsisand Dibamus. J Herpetol 19:116–156.
Guerra C, Montero R. 2009. The skull of Vanzosaura rubricauda(Squamata: Gymnophthalmidae). Acta Zool-Stockholm 90:359–371.
Hauschild A, Gabner P. 1995. Skinke im Terrarium. Hannover: Lan-buch-Verlag.
Hecht MK. 1956. A new xantusiid lizard from the Eocene of Wyom-ing. Am Mus Novit 1774:1–8.
Kosma R. 2004. The dentitions of recent and fossil scincomorphan liz-ards (Lacertilia, Squamata)—systematics, functional morphology,palecology. PhD Dissertation, University of Hannover, Germany.
Linnaeus C. 1758. Tomus I. Systema naturae per regna trianaturae, secundum classes, ordines, genera, species cum charac-teribus, differentiis, synonymis, locis. Editio decima, reformata.Stockholm: Laurentii Salvii.
Marig�o J, Minwer-Barakat R, Moy�a-Sol�a S. 2010. A new Anchomo-myni (Adapoidea, Primates) from the Mazater�on Middle Eocenelocality (Almaz�an Basin, Soria, Spain). J Hum Evol 58:353–361.
Minwer-Barakat R, Marig�o J, Moy�a-Sol�a S. 2012. Pseudoloris cues-tai, a new microchoerine (Primates, Omomyidae) from the MiddleEocene of the Iberian Peninsula. J Vertebr Paleontol 32:407–418.
Montero R, Abdala V, Moro S, Gallardo G. 2004. Atlas de Tupinam-bis rufescens (Squamata: Teiidae). Anatom�ıa externa, osteolog�ıa ybibliograf�ıa. Cuad Herpetol 18:17–32.
M€uller J. 2004. Cranial osteology of Dracaenosaurus croizeti, a lac-ertid lizard from the Oligocene of France (Reptilia, Squamata).Neu Jahr Geol Pal€aon Abhand 232:253–266.
M€uller J, Hipsley CA, Head JJ, Kardjilov N, Hilger A, Wuttke M,Reisz RR. 2011. Eocene lizard from Germany reveals amphisbae-nian origins. Nature 473:364–367.
Oppel M. 1811. Die Ordnung, Familien und Gattungen der Reptilienals Prodrom einer Naturgeschichte derselben. M€unchen: Lindauer.
Parker HW. 1956. The lizard genus Aprasia: its taxonomy andtemperature-correlated variation. Bull Br Mus (Nat Hist) Zool 3:365–385.
Pregill G. 1984. Durophagous feeding adaptations in an amphisbae-nid. J Herpetol 18:186–191.
Pyron RA, Burbrink FT, JJ Wiens. 2013. A phylogeny and updatedclassification of squamata, including 4161 species of lizards andsnakes. BMC Evol Biol 13:93.
Richter A. 1994. Lacertilia aus Unteren Kreide von U~na und Galve(Spanien) und Anoual (Marokko). Berl Geowiss Abh A 14:1–147.
Rogner M. 1994. Echsen. Bd. 2: Warane, Skinke und andere Echsensowie Br€uckenechsen und Krokodile. Stuttgart: Ulmer-Verlag.
Roscito JG, Rodrigues M. 2010. Comparative cranial osteology offossorial lizards from the tribe Gymnophthalmini (Squamata,Gymnophthalmidae). J Morphol 271:1352–1365.
Schleich HH. 1988. Neue Reptilienfunde aus dem Terti€ar Deutsch-lands 8. Palaeoblanus tobieni n. gen., n. sp.—neue Doppelschlei-chen aus dem Terti€ar Deutschlands. Pal€aont Z 62:95–105.
514 BOLET AND AUG�E

Shea GM, Hutchinson MN. 1992. A new species of lizard (Tiliqua)from the Miocene of Riversleigh, Queensland. Mem Qd Mus 32:303–310.
Sulimski A. 1972. Adamisaurus magnidentatus n. gen. et n. sp.(Sauria) from the upper cretaceous of Mongolia. Palaeontol Pol27:33–40.
Vidal N, Hedges, SB. 2005. The phylogeny of squamate reptiles (liz-ards, snakes, and amphisbaenians) inferred from nine nuclearprotein-coding genes. C R Biol 328:1000–1008.
Weber S. 2004. Ornatocephalus metzleri gen. et spec. nov (Lacerti-lia, Scincoida)—taxonomy and Paleobiology of a basal ScincoidLizard from the Messel Formation (Middle Eocene: basal Lute-
tian, Geiseltalium), Germany. Abh Senckenberg Naturforsch Ges561:1–159.
Wiens JJ, Brandley MC, Reeder TW. 2006. Why does a trait evolvemultiple times within a clade? Repeated evolution of snake-likebody form in squamate reptiles. Evolution 61:123–141.
Wiens JJ, Kuczynski CA, Townsend T, Reeder TW, Mulcahy DG,Sites JW, Jr. 2010. Combining phylogenomics and fossils inhigher-level squamate reptile phylogeny: molecular data changethe placement of fossil taxa. Syst Biol 59:674–688.
Wiens JJ, Hutter CR, Mulcahy DG, Noonan, BP, Townsend TM,Sites JW, Jr, Reeder TW. 2012. Resolving the phyogeny of lizardsand snakes (Squamata) with extensive sampling of genes and spe-cies. Biol Lett 8:1043–1046.
LIZARD FROM THE EOCENE OF FRANCE AND SPAIN 515




















