
A near-infrared narrow-waveband ratio to determine Leaf Area Index in orchards
11
A near-infrared narrow-waveband ratio to determine Leaf Area Index in orchards S. Delalieux a, ⁎, B. Somers a , S. Hereijgers a , W.W. Verstraeten a , W. Keulemans b , P. Coppin a a Department of Biosystems, M3-BIORES, Katholieke Universiteit Leuven, Celestijnenlaan 200E, 3001 Heverlee, Belgium b Department of Biosystems, Crop Bio-engineering, Katholieke Universiteit Leuven, W. de Croylaan 42, 3001 Heverlee, Belgium ABSTRACT ARTICLE INFO Article history: Received 22 February 2008 Received in revised form 23 May 2008 Accepted 24 May 2008 Available online xxxx Keywords: Hyperspectral data LAI Spectral index PROSPECT ACRM Orchards An innovative index for LAI estimation over orchard canopies based on the combination of two near-infrared narrow wavebands (1050 nm and 1250 nm) is presented. The index is insensitive to physiological changes on the leaf level even under changing pheneological developmental stage. In the meantime, the index is highly sensitive for variations at the canopy level. Values of LAI estimated by means of this newly developed index, called sLAIDI (standardized LAI Determining Index), resulted in excellent agreements with in-situ measured LAI values of citrus trees, with determination coefficients (R 2 ) of 0.83. Based on in-situ measured data and simulated data (PROSPECT and ACRM), the newly developed sLAIDI was also found to perform outstanding at high LAI values in contrary to well-known LAI related indices such as the Normalized Difference Vegetation Index (NDVI) and the Modified Chlorophyll Ratio Index 2 (MCARI2) that saturate at LAI values of 5 and 8 respectively. Moreover, the sLAIDI index was independent of changing pigment contents. Consequently, this index was regarded a valuable tool to more accurately extract biochemical parameters after normalizing the considered spectral data for LAI. By implementing sLAIDI in existing chlorophyll-related indices (e.g., GM index), the performance to estimate the leaf chlorophyll content from spectral citrus and peach canopy level data is increased (R 2 increases from 0.68 to 0.77). Moreover, a correction factor was proposed to enable the application of sLAIDI on fields with strong variations of water content. © 2008 Elsevier Inc. All rights reserved. 1. Introduction The Leaf Area Index (LAI) is a widely used input parameter in many biophysical models in the field of hydrological, ecological, geo- morphological, and climate modeling (Wulder et al.,1998). In addition, LAI is used for growth and yield prediction (Rasmussen, 1997), and plays an important role in monitoring change detection in canopy structure due to, for example, biotic or abiotic stresses (Coppin et al., 2004). Accurate estimates of LAI are required because the exposed area of living leaves is imperative in various biophysical processes such as plant transpiration and CO 2 exchange. Consequently, in capital- intensive orchard systems, accurate LAI estimates can contribute to a better management (e.g., pruning, precision farming). In the remote sensing community LAI is known as one of the major factors in radiative transfer models to upscale leaf level spectral data to canopy- level (Asner,1998; Haboudane et al., 2002; Zarco-Tejada et al., 2004). Although direct measurements of LAI are considered to be the most accurate, they are time-consuming, expensive, and often unfeasible for remote locations (Jonckheere et al., 2004). Thus, using remote sensing data may be an attractive solution to estimate LAI. Methods to estimate LAI from remote sensing data can generally be classified in two groups, i.e., a model-based and an empirical approach (Tang et al., 2007). The model-based approach is founded on inversion of physically based canopy reflectance models by determining a set of canopy biophysical variables that minimizes the difference between simulated and observed spectral reflectance (Kuusk & Nilson, 2000, Jacquemoud et al., 1995). The traditional iterative method is time-consuming and often requires a simplification of the models when processing large datasets. This results in a decrease of the inversion accuracy. Therefore look-up table (LUT) methods were developed which could partly overcome this drawback. They operate through a database of simulated spectral canopy reflectances variable in structural and radiometric properties. However, LUTcreation can be complicated and requires an extensive set of reliable field measurements. Several recent studies proposed the use of artificial neural networks (Gong et al., 1999; Jensen et al., 1999; Weiss & Baret, 1999; Atzberger, 2004; Schlerf & Atzberger, 2006) and genetic algorithms (Fang et al., 2003) in the forward and inverse modeling of radiative transfer models for retrieving biophysical variables. Application of those models, however, is often limited by the requirements of fairly large homogeneous areas and/or additional unknown input data (Fang et al., 2003). The empirical approach based on the use of vegetation indices is therefore often preferred for the retrieval of LAI since no previous site- specific information is required and the computation time is short. Numerous leaf-level research studies have focused on the prediction of crop physiological status through the extraction of leaf biochem- ical parameters, such as the concentration of chlorophyll a + b (C ab ), carotenoids, water (C w ), and dry matter (C m ) using hyperspectral vegetation indices (Carter, 1994; Carter & Knapp, 2001; Gitelson & Remote Sensing of Environment xxx (2008) xxx-xxx ⁎ Corresponding author. Tel.: +32 16 32 9749; fax: +32 16 32 97 60. E-mail address: [email protected] (S. Delalieux). RSE-07186; No of Pages 11 0034-4257/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.rse.2008.05.003 Contents lists available at ScienceDirect Remote Sensing of Environment journal homepage: www.elsevier.com/locate/rse ARTICLE IN PRESS Please cite this article as: Delalieux, S., et al., A near-infrared narrow-waveband ratio to determine Leaf Area Index in orchards, Remote Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
Transcript of A near-infrared narrow-waveband ratio to determine Leaf Area Index in orchards

Remote Sensing of Environment xxx (2008) xxx-xxx
RSE-07186; No of Pages 11
Contents lists available at ScienceDirect
Remote Sensing of Environment
j ourna l homepage: www.e lsev ie r.com/ locate / rse
ARTICLE IN PRESS
A near-infrared narrow-waveband ratio to determine Leaf Area Index in orchards
S. Delalieux a,⁎, B. Somers a, S. Hereijgers a, W.W. Verstraeten a, W. Keulemans b, P. Coppin a
a Department of Biosystems, M3-BIORES, Katholieke Universiteit Leuven, Celestijnenlaan 200E, 3001 Heverlee, Belgiumb Department of Biosystems, Crop Bio-engineering, Katholieke Universiteit Leuven, W. de Croylaan 42, 3001 Heverlee, Belgium
⁎ Corresponding author. Tel.: +32 16 32 97 49; fax: +3E-mail address: [email protected]
0034-4257/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.rse.2008.05.003
Please cite this article as: Delalieux, S., etSensing of Environment (2008), doi:10.1016/
A B S T R A C T
A R T I C L E I N F OArticle history:
An innovative index for LAI Received 22 February 2008Received in revised form 23 May 2008Accepted 24 May 2008Available online xxxxKeywords:Hyperspectral dataLAISpectral indexPROSPECTACRMOrchards
estimation over orchard canopies based on the combination of two near-infrarednarrow wavebands (1050 nm and 1250 nm) is presented. The index is insensitive to physiological changes onthe leaf level even under changing pheneological developmental stage. In the meantime, the index is highlysensitive for variations at the canopy level. Values of LAI estimated by means of this newly developed index,called sLAIDI (standardized LAI Determining Index), resulted in excellent agreements with in-situ measuredLAI values of citrus trees, with determination coefficients (R2) of 0.83. Based on in-situ measured data andsimulated data (PROSPECT and ACRM), the newly developed sLAIDI was also found to perform outstanding athigh LAI values in contrary to well-known LAI related indices such as the Normalized Difference VegetationIndex (NDVI) and the Modified Chlorophyll Ratio Index 2 (MCARI2) that saturate at LAI values of 5 and 8respectively. Moreover, the sLAIDI index was independent of changing pigment contents. Consequently, thisindex was regarded a valuable tool to more accurately extract biochemical parameters after normalizingthe considered spectral data for LAI. By implementing sLAIDI in existing chlorophyll-related indices (e.g.,GM index), the performance to estimate the leaf chlorophyll content from spectral citrus and peach canopylevel data is increased (R2 increases from 0.68 to 0.77). Moreover, a correction factor was proposed to enablethe application of sLAIDI on fields with strong variations of water content.
© 2008 Elsevier Inc. All rights reserved.
1. Introduction
The Leaf Area Index (LAI) is a widely used input parameter in manybiophysical models in the field of hydrological, ecological, geo-morphological, and climatemodeling (Wulder et al.,1998). In addition,LAI is used for growth and yield prediction (Rasmussen, 1997), andplays an important role in monitoring change detection in canopystructure due to, for example, biotic or abiotic stresses (Coppin et al.,2004). Accurate estimates of LAI are required because the exposed areaof living leaves is imperative in various biophysical processes such asplant transpiration and CO2 exchange. Consequently, in capital-intensive orchard systems, accurate LAI estimates can contribute to abetter management (e.g., pruning, precision farming). In the remotesensing community LAI is known as one of the major factors inradiative transfer models to upscale leaf level spectral data to canopy-level (Asner, 1998; Haboudane et al., 2002; Zarco-Tejada et al., 2004).Although direct measurements of LAI are considered to be the mostaccurate, they are time-consuming, expensive, and often unfeasible forremote locations (Jonckheere et al., 2004). Thus, using remote sensingdatamay be an attractive solution to estimate LAI.Methods to estimateLAI from remote sensing data can generally be classified in two groups,i.e., a model-based and an empirical approach (Tang et al., 2007). Themodel-based approach is founded on inversion of physically based
2 16 32 97 60.e (S. Delalieux).
l rights reserved.
al., A near-infrared narrow-j.rse.2008.05.003
canopy reflectance models by determining a set of canopy biophysicalvariables that minimizes the difference between simulated andobserved spectral reflectance (Kuusk & Nilson, 2000, Jacquemoudet al., 1995). The traditional iterative method is time-consuming andoften requires a simplification of the models when processing largedatasets. This results in a decrease of the inversion accuracy. Thereforelook-up table (LUT) methods were developed which could partlyovercome this drawback. They operate through a database ofsimulated spectral canopy reflectances variable in structural andradiometric properties. However, LUTcreation can be complicated andrequires an extensive set of reliable fieldmeasurements. Several recentstudies proposed the use of artificial neural networks (Gong et al.,1999; Jensen et al., 1999; Weiss & Baret, 1999; Atzberger, 2004; Schlerf& Atzberger, 2006) and genetic algorithms (Fang et al., 2003) in theforward and inverse modeling of radiative transfer models forretrieving biophysical variables. Application of thosemodels, however,is often limited by the requirements of fairly large homogeneous areasand/or additional unknown input data (Fang et al., 2003).
The empirical approach based on the use of vegetation indices istherefore often preferred for the retrieval of LAI since no previous site-specific information is required and the computation time is short.
Numerous leaf-level research studies have focused on the predictionof crop physiological status through the extraction of leaf biochem-ical parameters, such as the concentration of chlorophyll a + b (Cab),carotenoids, water (Cw), and dry matter (Cm) using hyperspectralvegetation indices (Carter, 1994; Carter & Knapp, 2001; Gitelson &
waveband ratio to determine Leaf Area Index in orchards, Remote

2 S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
Merzlyak,1996; Peñuelas et al.,1994; Sims&Gamon, 2002; Zarco-Tejadaet al., 2001, 2004). For large scale application, it should however bemoreinteresting to extract leaf biochemical parameters, and as such cropphysiological status directly from canopy reflectance measurements.However, the simultaneous interaction of LAI and chlorophyll on thevegetation spectrumhampers this action. Therefore, it can be concludedthat there is certainly a need for an efficient and accurate extractionof LAI.
Already in 1969 a strong correlation was found between a red tonear-infrared (NIR) spectral ratio and LAI (Jordan, 1969). Probably themost studied and implemented vegetation index ever to predict LAI isbased on these spectral regions and is called the NormalizedDifference Vegetation Index (NDVI) (Tucker, 1979; Turner et al.,1999; Qi et al., 2000). Later studies attempted to improve this NDVI byreducing the influence of soil on canopy reflectance (Baret & Guyot,1991; Huete, 1988, Rondeaux et al., 1996). Notwithstanding the manyremote sensing studies conducted in this research field, the potentialto predict LAI with hyperspectral remote sensing has not been fullyrealized as it still fails to address the following issues:
• A recognized drawback of vegetation indices designed to predict LAIfrom remotely sensed data is that they saturate when applied indense canopies. The sensitivity of the NDVI is known to asympto-tically approach a saturation level when LAI exceeds 2 to 5 (Birky,2001; Gao, 1996). However, LAI of for example citrus canopies canyield values up to 11 (Kalma & Stanhill, 1969; Turrell, 1961; Cohen &Fuchs, 1987).
• Another eminent drawback of the LAI predicting indices is that thusfar only few studies have focused on the prediction of LAI from avegetation spectrum without interference of chlorophyll effects. Theencountered problem is caused by the fact that LAI and chlorophyllcontent have similar effects on canopy reflectance from the greenregion (~ 550 nm) to the red-edge (~ 750 nm) region of the electro-magnetic spectrum. Vegetation indices based on the red and NIRspectral features will therefore overestimate the LAI when dealingwith a canopy having high chlorophyll content, while they will un-derestimate the LAI of canopies with low chlorophyll content.Haboudane et al. (2004) introduced the indices MTVI2 (ModifiedTriangular Vegetation Index 2) and MCARI2 (Modified ChlorophyllAbsorption Ratio Index 2) as indices which were less sensitive tochlorophyll concentration variations. However, also these LAI predict-ing vegetation indices rely on NIR as well as the red wavelengths,which point out a possible interaction between LAI and chlorophyll.
To address these issues, the current study presents the ‘standar-dized LAI Determining Index’ or ‘sLAIDI’. The sLAIDI is a standardizedratio index consisting of two wavelengths (1050 nm and 1250 nm)situated in the NIR spectral domain thereby minimizing the influenceof variations in chlorophyll concentration. This index was obtained asa result of a thorough leaf-level study and upscaling to canopy level.The general approach of this study is that indices which are invariantat leaf level, even under varying biochemical properties, and variant atcanopy level, will provide a good measure for canopy structure, withLAI being one of the major elements determining canopy structure.
This paper aims at investigating the capability of sLAIDI foraccurately estimating LAI of fruit orchards by applying the index onsimulated and in in-situ measured canopy spectral datasets. Thesimulated datasets were gathered via the radiative transfer models‘PROSPECT’ (Jacquemoud et al., 1996) and ‘ACRM’ (Kuusk, 2001). Thesedatasets made it possible to test the sensitivity of the developed indexto changing water and chlorophyll content and to high LAI values (cfr.mentioned drawbacks of existing indices).
The ‘sLAIDI’ is not only thought to estimate LAI for large areas oforchard trees in an accurate, non-destructive, and time-efficient way,but furthermore opens up new perspectives in the framework ofaccurate extracting leaf biochemical parameters from canopy level data.Compared to leaf-level measurements, predicting leaf chlorophyll
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
content from reflectance measurements at the canopy scale has provento be difficult due to unknown LAI values. Therefore, it was tested if amore accurate extraction of leaf chlorophyll content was achievablewhen combining an existing chlorophyll-related index with sLAIDI. Inthis case, sLAIDI has to be considered as a normalizing factor for LAI.
This paper first outlines the experimental setup (Section 2). Themethodology (Section 3) briefly discusses the way the new index isachieved and how the robustness of this index is tested to estimateLAI. Results are presented and discussed in Section 4. The last part ofthis paper (Section 5) intends to combine a chlorophyll-related indexto the new index for a more accurate extraction of chlorophyll contentof in- situ measured datasets.
2. Materials
2.1. Measurements in a controlled greenhouse environment
A temporal leaf-level hyperspectral dataset was collected in acontrolled greenhouse environment. The two youngest leaves of 25apple plants (Malus x domestica cv Rewena chip budded on M9rootstock)weremeasuredwith the plant probe attached to a FieldSpecPro JR spectroradiometer with a spectral range of 350–2500 nm. Thesampling interval across the 350–1050 nm range is 1.4 nm with aspectral resolution of 3 nm. The sampling interval and the spectralresolution are approximately 2 nm and 30 nm, respectively, for the1050–2500 nm range (Analytical Spectral Devices Inc., Boulder, USA).Resultant data afterwards are interpolated by the ASD software toproduce values at each nanometer interval. A white Spectralonreference panel (Labsphere, Inc., North Sutton, NH, USA) was usedfor reflectance calibration. The measurements started at the first dayafter leaf unfolding andwere repeated at short time intervals (1, 2, 4, 5,8, 10, 12, 15, 18, 22, and 26 days later). Each time, two measurementpoints at different positions were taken from each leaf in order toobtain an adequate representation of the whole leaf. The assumptionwas made that leaves unfurling at the beginning of this experiment,were fully expanded after 20 days (MacHardy, 1996).
2.2. Field measurements
2.2.1. Study areas and site descriptionThe study areas were located near Zaragoza, Spain (Lat: 41.28°N,
Long: 1.22°W), and near Wellington, South Africa (33.35°S, 18.55°E).Peach canopies (Prunus persica L. Batsch) were grown in Spain on acalcareous soil with a clay–loamy texture. The 6500m2 peach orchardarea enclosed 205 trees (4 m by 5 m plant distance, average canopyheight of 2.8 m) covering a dynamic range of chlorophyll concentrationdue to iron chlorosis induction in 48 trees. These latter were selected forfurther measurements. A 3600 m2 citrus orchard offered the 38 citrustrees for the trial in SouthAfrica. Tenyears old ‘MidknightValencia’ citrus(Citrus sinensis L.) trees, bud grafted onto ‘Carrizo’ citrange rootstockwere grown on an Albic Luvisol soil. Plant distance was 4.5 m by 2.5 mwith an average tree height of 2.9 m. Uniform fertilizing and manage-ment practices were implemented for all trees (i.e., timing and amountof fertilization, amount of irrigation, pest and disease control, etc.).
2.2.2. Hyperspectral dataSpectral reflectancemeasurements were taken on clear sunny days
between 11 AM and 2 PM, local time to minimize the effects ofchanges in the solar angle. Canopy as well as leaf-level hyperspectralmeasurements were collected with a portable FieldSpec Pro FRspectroradiometer (Analytical Spectral Devices Inc., Boulder, USA)with a spectral range of 350–2500 nm. While the sampling intervaland spectral resolution across the 350–1050 nm are similar to that ofthe FieldSpec Pro JRmodel, the values for the 1050–2500 nm range areapproximately 2 nm and 10 nm. Canopy data were gathered at 2 mabove the canopies with a FOV of 25°, resulting in a footprint of 88 cm
waveband ratio to determine Leaf Area Index in orchards, Remote

3S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
at the top of the canopy. Background interferences were as sucheliminated. Reflectance spectra of leaves, picked randomly from theupper hemisphere of the canopy, were collected bymeasuring spots of10 mm diameter using a plant probe containing a 100 W halogenreflectorized lamp. Leaf transmittance measurements were collectedusing the RTS-3ZC integrating sphere, designed for use with the ASDFieldSpec spectroradiometer. A Fieldspec Pro JR was used for thosemeasurements (see Section 2.1).
2.2.3. Biochemical and physiological dataThe chlorophyll content for each selected leaf in the peach (15
leaves per tree; 48 trees) and citrus (20 leaves per tree; 38 trees)orchard was determined. For the peach leaves, a SPAD-502 (MinoltaCamera Co., Ltd, Japan) Chlorophyll Meter was used, while the citrusleaves were measured with a CCM-200 (Opti-Sciences Inc., Hudson,NH, USA) device. Mean values were calculated per tree to comparewith parameters obtained at the crown level. The relative measure-ments were calibrated to obtain absolute chlorophyll concentration bycomparing the digital chlorophyll meter readings with concentrationsderived from destructive chemical analysis in the laboratory for asubset of leaf samples (five per tree). Chlorophyll concentration wasdetermined with the acetone (80%) extraction method (Arnon, 1994).Coefficients of determination of 0.86 and 0.90 were obtained for thepeach and citrus data respectively.
Water content and drymatter content weremeasured byweighingthe selected fresh and dried citrus and peach leaves respectively.Equivalent water thickness (Cw) defined as the ratio between thequantity of water and the leaf area was obtained after measuring themean leaf areawith the LI-3000A Leaf AreaMeter (precision of 1mm2).
2.2.4. Leaf Area Index (LAI)Leaf area index (LAI) values of the 38 Midknight Valencia trees
were measured on cloudy days under totally diffuse light. The LAIvalues were determined by measuring the PAR (photosyntheticallyactive radiation) two times above and twelve times below the canopywith a ceptometer using 80 individual sensors on a probe (AccuPARLP-80, Decagon Devices Inc., Pullman, WA, USA). The light scatteringmodel suggested by Norman and Campbell (1989) was inverted todetermine the LAI of the canopies. This resulted for overcast days inthe following equation:
LAI ¼ ln τ=A ð1Þ
With τ being the fraction of transmitted PAR, and A representing thearea of the soil surface [m2].
This indirect non-contact LAI determination was chosen for itsconvenience. Nevertheless, direct methods for LAI determination aremore accurate and therefore a destructive calibration experiment wascarried out on three trees in the same orchard. LAI values wererecorded four times per tree with the AccuPAR LP-80 while graduallyharvesting and counting the leaves.
In this calibration experiment the LAI values were calculated fromthe total leaf area and the area of soil surface beneath the tree. Thetotal leaf area is the product of the number of leaves multiplied by themean surface area per leaf which was determined by measuring thesurface areas of 10% of the picked leaves with the area meter LI-3000A(LI-COR, Lincoln NE, USA). The ceptometer readings were comparedwith the LAI values derived from destructive analysis. A linear fit witha coefficient of determination of 0.95 was obtained.
3. Methods
To clarify the subsequent steps described in this section, it is usefulto recall the principle of the present study. A prerequisite for a spectralalgorithm to monitor LAI is that its values change only when the totalleaf area alters. I.e., a modification in the biophysical composition and/
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
or developmental stage of the leaves may not alter the results of thealgorithm. As a result, this section starts with the description of theselection of invariant spectral features at leaf-level, before testingthem at canopy level.
3.1. Selection of invariant spectral features at leaf-level
The spectral behaviour of apple leaves is investigated throughouttime from unfurling until total expansion. Temporal spectral changesare monitored to define wavelengths or wavelength ratios that re-main stable during leaf development. The development of a leaf ischaracterized by a variety of biophysical changes, e.g., size, weight,increase in pigments, water content and structural changes. Theamount of proportional alteration of the averaged spectral reflectancefor each wavelength over the period of interest (26 days) is examinedand themost robust features are extracted (Delalieuxet al., 2007a). Themost robust feature was selected based on its insensitivity to differentspecies and conditions by applying it to the gathered apple, peachand citrus leaf dataset. In a next step, this selected feature is im-plemented in a canopy-level study to test their invariance in morecomplex scenarios.
3.2. Testing invariance of selected index at canopy level
3.2.1. In-situ datasetCanopy spectra differ from leaf spectra in that variations of LAI and
background contribution alter the reflectance properties of canopies.Consequently it is not straightforward to upscale findings at leaf scaleto the canopy scale. Therefore, only the selectedmost invariant featureat the leaf-level study (Section 3.1) is applied on the measured citruscanopy spectra to test whether it stays invariant at this more complexlevel.
3.2.2. Simulated datasetRadiative transfer simulationswere performed to test the robustness
of the selected feature to changing leaf chlorophyll and water contentand tovarying LAI. Therefore, canopy reflectance spectrawere generatedfor LAI values ranging from1 to 11with leaf chlorophyll contents varyingfrom 2.50 to 100.00 μg/cm2. These ranges were chosen to mimic theranges of the parametersmeasured on the field. Additional variations inleaf water content ranging from 5.00 to 28.60 μg/cm2 were simulatedbased on the range available in the LOPEX (Leaf Optical PropertiesExperiment) dataset. This dataset included leaf measurements ofdifferent biochemical constituents from a large sample of tree species.The LOPEX data were acquired during an experiment conducted by theJoint Research Centre of the European Commission (Ispra, Italy); theLOPEX database was made available by the JRC through the FTP siteftp://ftp-gvm.jrc.it/verstmi/lopex/.
The canopy radiative transfer model chosen in this study is called‘ACRM’ andhas beendevelopedbyKuusk (2001). It is a two-layer canopyreflectance model which simulates the directional reflectance of ahomogeneous layer of vegetation and a thin layer of vegetation on theground surface in the 400–2400 nm optical domain of radiation with aspectral resolution of 1 nm. The model accounts for the non-lambertiansoil reflectance, specular reflection of direct sun radiation on leaves, hot-spot effect and a two-parameter leaf angle distribution (Kuusk, 2001).It requires information on the spectral variability of soil reflectanceas a function of four vectors according to Price (1990), illuminationconditions and a set of plant specific parameters, such as, leaf area index(LAI), leaf angle distribution (LAD), leaf size, and an accurate descriptionof leaf optical properties. The latterweremeasured in–situ (Section 2.2),as well as simulated with the PROSPECT model (version 3.01) to obtaina larger variability in leaf biochemistry. PROSPECT is a simple buteffective radiative transfer model that calculates the leaf transmittanceand reflectance with a limited number of input parameters: the leafinternal structure parameter N, the chlorophyll content Cab, (μg cm− 2),
waveband ratio to determine Leaf Area Index in orchards, Remote
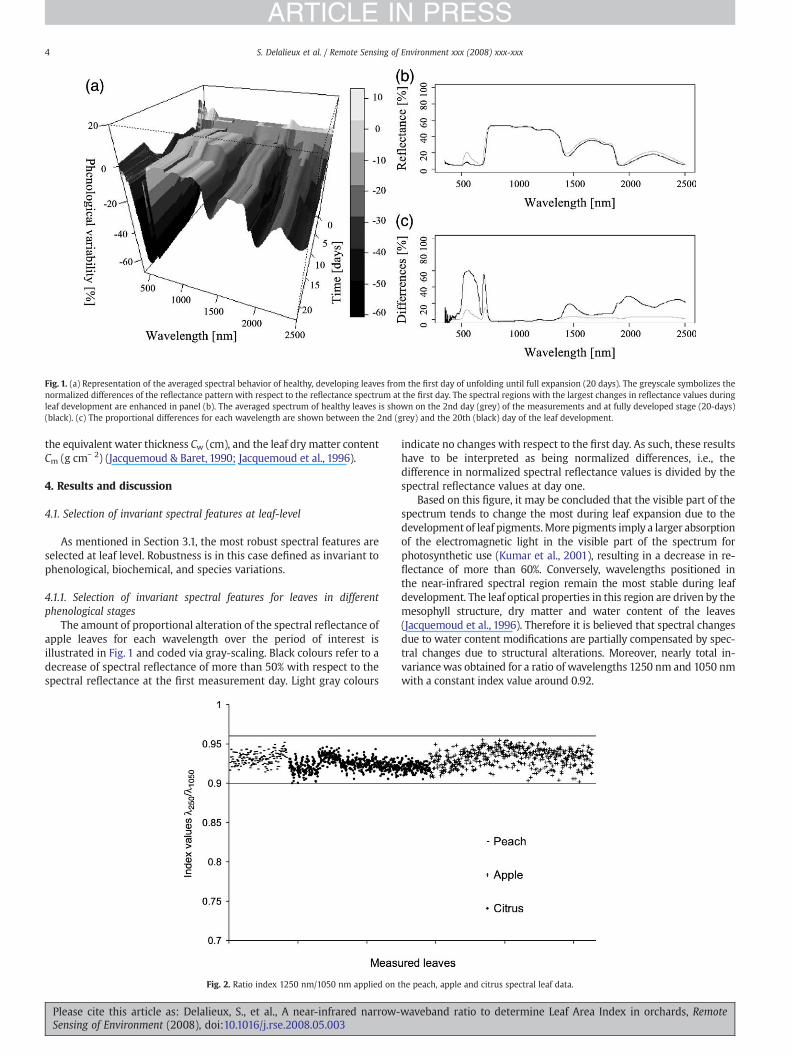
Fig. 1. (a) Representation of the averaged spectral behavior of healthy, developing leaves from the first day of unfolding until full expansion (20 days). The greyscale symbolizes thenormalized differences of the reflectance pattern with respect to the reflectance spectrum at the first day. The spectral regions with the largest changes in reflectance values duringleaf development are enhanced in panel (b). The averaged spectrum of healthy leaves is shown on the 2nd day (grey) of the measurements and at fully developed stage (20-days)(black). (c) The proportional differences for each wavelength are shown between the 2nd (grey) and the 20th (black) day of the leaf development.
4 S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
the equivalent water thickness Cw (cm), and the leaf dry matter contentCm (g cm− 2) (Jacquemoud & Baret, 1990; Jacquemoud et al., 1996).
4. Results and discussion
4.1. Selection of invariant spectral features at leaf-level
As mentioned in Section 3.1, the most robust spectral features areselected at leaf level. Robustness is in this case defined as invariant tophenological, biochemical, and species variations.
4.1.1. Selection of invariant spectral features for leaves in differentphenological stages
The amount of proportional alteration of the spectral reflectance ofapple leaves for each wavelength over the period of interest isillustrated in Fig. 1 and coded via gray-scaling. Black colours refer to adecrease of spectral reflectance of more than 50% with respect to thespectral reflectance at the first measurement day. Light gray colours
Fig. 2. Ratio index 1250 nm/1050 nm applied on t
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
indicate no changes with respect to the first day. As such, these resultshave to be interpreted as being normalized differences, i.e., thedifference in normalized spectral reflectance values is divided by thespectral reflectance values at day one.
Based on this figure, it may be concluded that the visible part of thespectrum tends to change the most during leaf expansion due to thedevelopment of leaf pigments.More pigments imply a larger absorptionof the electromagnetic light in the visible part of the spectrum forphotosynthetic use (Kumar et al., 2001), resulting in a decrease in re-flectance of more than 60%. Conversely, wavelengths positioned inthe near-infrared spectral region remain the most stable during leafdevelopment. The leaf optical properties in this region are driven by themesophyll structure, dry matter and water content of the leaves(Jacquemoud et al., 1996). Therefore it is believed that spectral changesdue to water content modifications are partially compensated by spec-tral changes due to structural alterations. Moreover, nearly total in-variancewas obtained for a ratio of wavelengths 1250 nm and 1050 nmwith a constant index value around 0.92.
he peach, apple and citrus spectral leaf data.
waveband ratio to determine Leaf Area Index in orchards, Remote
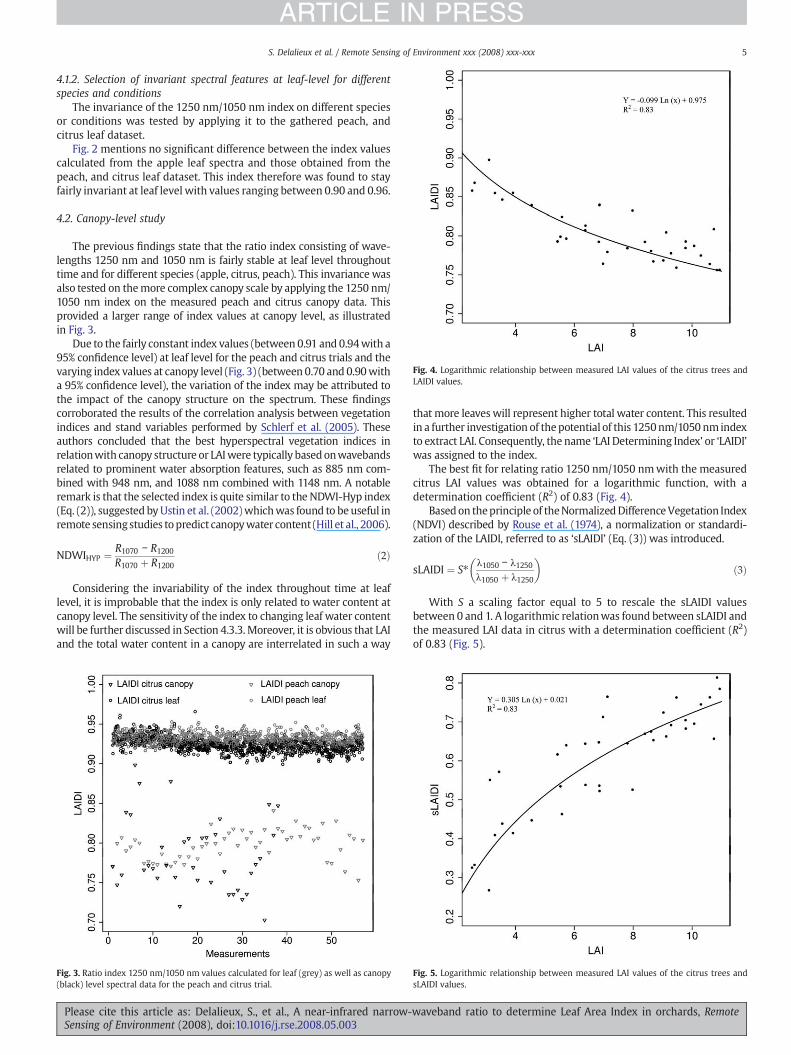
Fig. 4. Logarithmic relationship between measured LAI values of the citrus trees andLAIDI values.
5S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
4.1.2. Selection of invariant spectral features at leaf-level for differentspecies and conditions
The invariance of the 1250 nm/1050 nm index on different speciesor conditions was tested by applying it to the gathered peach, andcitrus leaf dataset.
Fig. 2 mentions no significant difference between the index valuescalculated from the apple leaf spectra and those obtained from thepeach, and citrus leaf dataset. This index therefore was found to stayfairly invariant at leaf level with values ranging between 0.90 and 0.96.
4.2. Canopy-level study
The previous findings state that the ratio index consisting of wave-lengths 1250 nm and 1050 nm is fairly stable at leaf level throughouttime and for different species (apple, citrus, peach). This invariance wasalso tested on themore complex canopy scale by applying the 1250 nm/1050 nm index on the measured peach and citrus canopy data. Thisprovided a larger range of index values at canopy level, as illustratedin Fig. 3.
Due to the fairly constant index values (between 0.91 and 0.94with a95% confidence level) at leaf level for the peach and citrus trials and thevarying index values at canopy level (Fig. 3) (between 0.70 and 0.90witha 95% confidence level), the variation of the index may be attributed tothe impact of the canopy structure on the spectrum. These findingscorroborated the results of the correlation analysis between vegetationindices and stand variables performed by Schlerf et al. (2005). Theseauthors concluded that the best hyperspectral vegetation indices inrelationwith canopy structure or LAIwere typically basedonwavebandsrelated to prominent water absorption features, such as 885 nm com-bined with 948 nm, and 1088 nm combined with 1148 nm. A notableremark is that the selected index is quite similar to the NDWI-Hyp index(Eq. (2)), suggested byUstin et al. (2002)whichwas found to beuseful inremote sensingstudies topredict canopywater content (Hill et al., 2006).
NDWIHYP ¼ R1070 − R1200
R1070 þ R1200ð2Þ
Considering the invariability of the index throughout time at leaflevel, it is improbable that the index is only related to water content atcanopy level. The sensitivity of the index to changing leaf water contentwill be further discussed in Section 4.3.3.Moreover, it is obvious that LAIand the total water content in a canopy are interrelated in such a way
Fig. 3. Ratio index 1250 nm/1050 nm values calculated for leaf (grey) as well as canopy(black) level spectral data for the peach and citrus trial.
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
that more leaves will represent higher total water content. This resultedin a further investigation of thepotential of this 1250nm/1050 nm indexto extract LAI. Consequently, the name ‘LAI Determining Index’ or ‘LAIDI’was assigned to the index.
The best fit for relating ratio 1250 nm/1050 nmwith the measuredcitrus LAI values was obtained for a logarithmic function, with adetermination coefficient (R2) of 0.83 (Fig. 4).
Basedon theprinciple of theNormalizedDifferenceVegetation Index(NDVI) described by Rouse et al. (1974), a normalization or standardi-zation of the LAIDI, referred to as ‘sLAIDI’ (Eq. (3)) was introduced.
sLAIDI ¼ S⁎λ1050 − λ1250λ1050 þ λ1250
� �ð3Þ
With S a scaling factor equal to 5 to rescale the sLAIDI valuesbetween 0 and 1. A logarithmic relationwas found between sLAIDI andthe measured LAI data in citrus with a determination coefficient (R2)of 0.83 (Fig. 5).
Fig. 5. Logarithmic relationship between measured LAI values of the citrus trees andsLAIDI values.
waveband ratio to determine Leaf Area Index in orchards, Remote
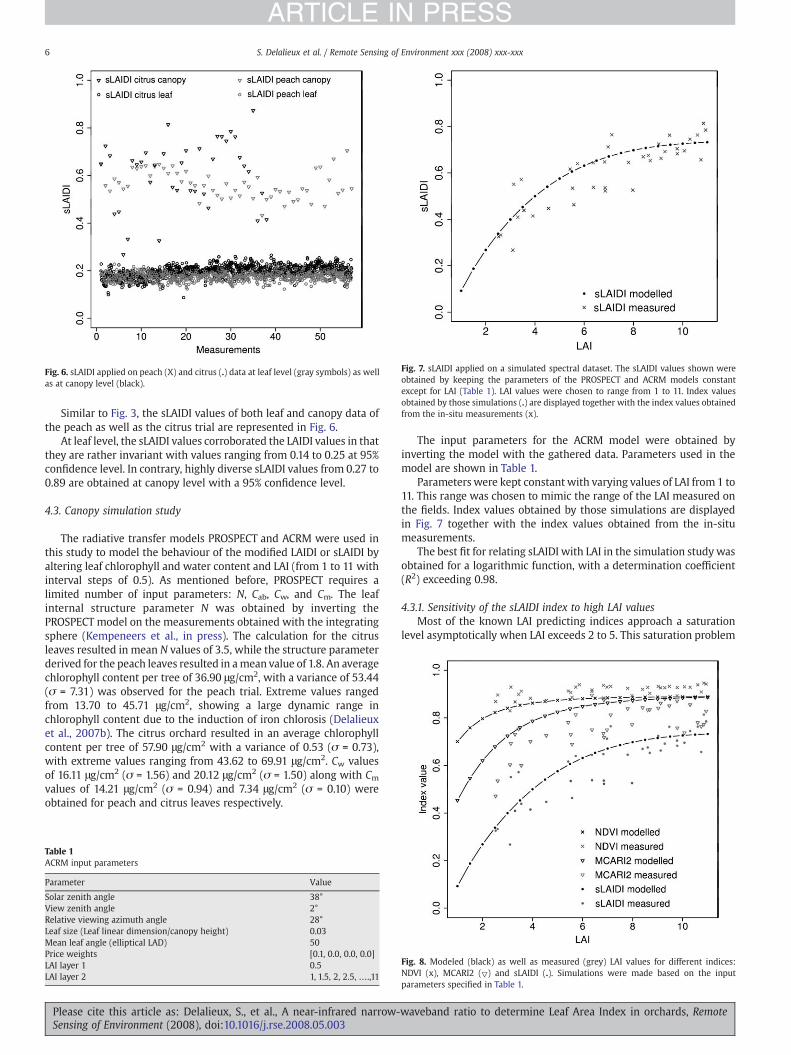
Fig. 6. sLAIDI applied on peach (X) and citrus (.) data at leaf level (gray symbols) as wellas at canopy level (black).
Fig. 7. sLAIDI applied on a simulated spectral dataset. The sLAIDI values shown wereobtained by keeping the parameters of the PROSPECT and ACRM models constantexcept for LAI (Table 1). LAI values were chosen to range from 1 to 11. Index valuesobtained by those simulations (.) are displayed together with the index values obtainedfrom the in-situ measurements (x).
6 S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
Similar to Fig. 3, the sLAIDI values of both leaf and canopy data ofthe peach as well as the citrus trial are represented in Fig. 6.
At leaf level, the sLAIDI values corroborated the LAIDI values in thatthey are rather invariant with values ranging from 0.14 to 0.25 at 95%confidence level. In contrary, highly diverse sLAIDI values from 0.27 to0.89 are obtained at canopy level with a 95% confidence level.
4.3. Canopy simulation study
The radiative transfer models PROSPECT and ACRM were used inthis study to model the behaviour of the modified LAIDI or sLAIDI byaltering leaf chlorophyll and water content and LAI (from 1 to 11 withinterval steps of 0.5). As mentioned before, PROSPECT requires alimited number of input parameters: N, Cab, Cw, and Cm. The leafinternal structure parameter N was obtained by inverting thePROSPECT model on the measurements obtained with the integratingsphere (Kempeneers et al., in press). The calculation for the citrusleaves resulted in mean N values of 3.5, while the structure parameterderived for the peach leaves resulted in amean value of 1.8. An averagechlorophyll content per tree of 36.90 μg/cm2, with a variance of 53.44(σ = 7.31) was observed for the peach trial. Extreme values rangedfrom 13.70 to 45.71 μg/cm2, showing a large dynamic range inchlorophyll content due to the induction of iron chlorosis (Delalieuxet al., 2007b). The citrus orchard resulted in an average chlorophyllcontent per tree of 57.90 μg/cm2 with a variance of 0.53 (σ = 0.73),with extreme values ranging from 43.62 to 69.91 μg/cm2. Cw valuesof 16.11 μg/cm2 (σ = 1.56) and 20.12 μg/cm2 (σ = 1.50) along with Cmvalues of 14.21 μg/cm2 (σ = 0.94) and 7.34 μg/cm2 (σ = 0.10) wereobtained for peach and citrus leaves respectively.
Table 1ACRM input parameters
Parameter Value
Solar zenith angle 38°View zenith angle 2°Relative viewing azimuth angle 28°Leaf size (Leaf linear dimension/canopy height) 0.03Mean leaf angle (elliptical LAD) 50Price weights [0.1, 0.0, 0.0, 0.0]LAI layer 1 0.5LAI layer 2 1, 1.5, 2, 2.5, ….,11
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
The input parameters for the ACRM model were obtained byinverting the model with the gathered data. Parameters used in themodel are shown in Table 1.
Parameters were kept constant with varying values of LAI from 1 to11. This range was chosen to mimic the range of the LAI measured onthe fields. Index values obtained by those simulations are displayedin Fig. 7 together with the index values obtained from the in-situmeasurements.
The best fit for relating sLAIDI with LAI in the simulation study wasobtained for a logarithmic function, with a determination coefficient(R2) exceeding 0.98.
4.3.1. Sensitivity of the sLAIDI index to high LAI valuesMost of the known LAI predicting indices approach a saturation
level asymptotically when LAI exceeds 2 to 5. This saturation problem
Fig. 8. Modeled (black) as well as measured (grey) LAI values for different indices:NDVI (x), MCARI2 (▽) and sLAIDI (.). Simulations were made based on the inputparameters specified in Table 1.
waveband ratio to determine Leaf Area Index in orchards, Remote
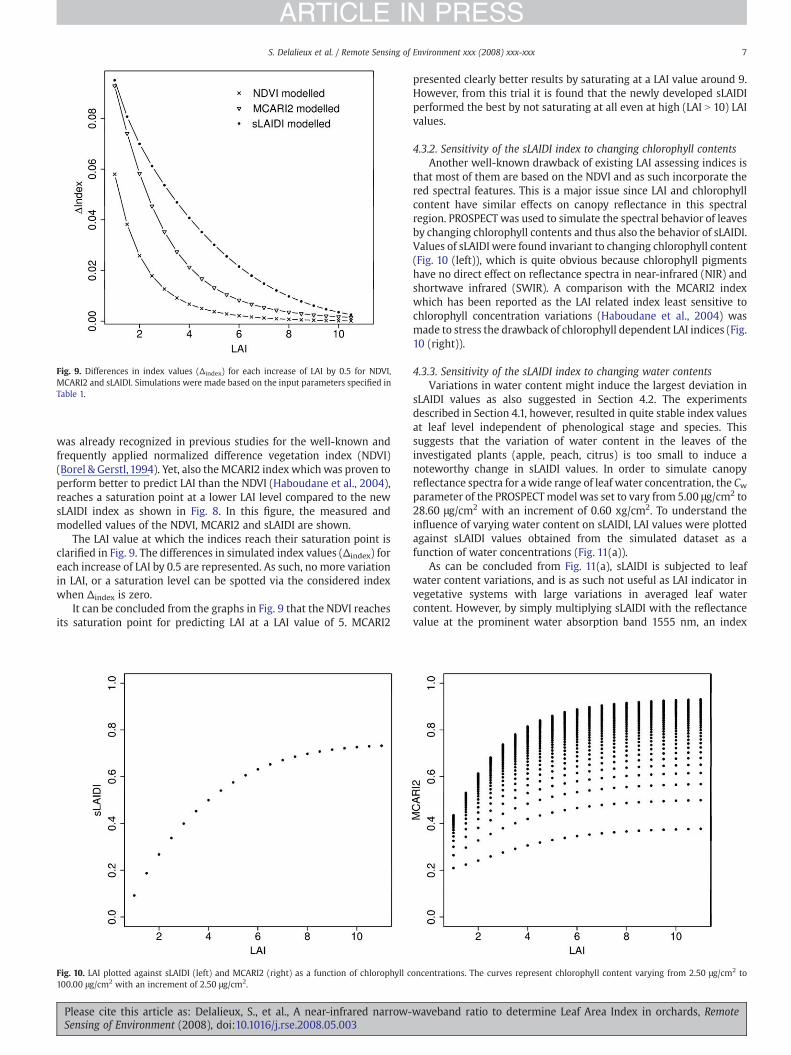
Fig. 9. Differences in index values (Δindex) for each increase of LAI by 0.5 for NDVI,MCARI2 and sLAIDI. Simulations were made based on the input parameters specified inTable 1.
7S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
was already recognized in previous studies for the well-known andfrequently applied normalized difference vegetation index (NDVI)(Borel & Gerstl, 1994). Yet, also theMCARI2 indexwhich was proven toperform better to predict LAI than the NDVI (Haboudane et al., 2004),reaches a saturation point at a lower LAI level compared to the newsLAIDI index as shown in Fig. 8. In this figure, the measured andmodelled values of the NDVI, MCARI2 and sLAIDI are shown.
The LAI value at which the indices reach their saturation point isclarified in Fig. 9. The differences in simulated index values (Δindex) foreach increase of LAI by 0.5 are represented. As such, no more variationin LAI, or a saturation level can be spotted via the considered indexwhen Δindex is zero.
It can be concluded from the graphs in Fig. 9 that the NDVI reachesits saturation point for predicting LAI at a LAI value of 5. MCARI2
Fig. 10. LAI plotted against sLAIDI (left) and MCARI2 (right) as a function of chlorophyll c100.00 μg/cm2 with an increment of 2.50 μg/cm2.
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
presented clearly better results by saturating at a LAI value around 9.However, from this trial it is found that the newly developed sLAIDIperformed the best by not saturating at all even at high (LAI N 10) LAIvalues.
4.3.2. Sensitivity of the sLAIDI index to changing chlorophyll contentsAnother well-known drawback of existing LAI assessing indices is
that most of them are based on the NDVI and as such incorporate thered spectral features. This is a major issue since LAI and chlorophyllcontent have similar effects on canopy reflectance in this spectralregion. PROSPECT was used to simulate the spectral behavior of leavesby changing chlorophyll contents and thus also the behavior of sLAIDI.Values of sLAIDI were found invariant to changing chlorophyll content(Fig. 10 (left)), which is quite obvious because chlorophyll pigmentshave no direct effect on reflectance spectra in near-infrared (NIR) andshortwave infrared (SWIR). A comparison with the MCARI2 indexwhich has been reported as the LAI related index least sensitive tochlorophyll concentration variations (Haboudane et al., 2004) wasmade to stress the drawback of chlorophyll dependent LAI indices (Fig.10 (right)).
4.3.3. Sensitivity of the sLAIDI index to changing water contentsVariations in water content might induce the largest deviation in
sLAIDI values as also suggested in Section 4.2. The experimentsdescribed in Section 4.1, however, resulted in quite stable index valuesat leaf level independent of phenological stage and species. Thissuggests that the variation of water content in the leaves of theinvestigated plants (apple, peach, citrus) is too small to induce anoteworthy change in sLAIDI values. In order to simulate canopyreflectance spectra for awide range of leaf water concentration, the Cwparameter of the PROSPECTmodel was set to vary from 5.00 μg/cm2 to28.60 μg/cm2 with an increment of 0.60 xg/cm2. To understand theinfluence of varying water content on sLAIDI, LAI values were plottedagainst sLAIDI values obtained from the simulated dataset as afunction of water concentrations (Fig. 11(a)).
As can be concluded from Fig. 11(a), sLAIDI is subjected to leafwater content variations, and is as such not useful as LAI indicator invegetative systems with large variations in averaged leaf watercontent. However, by simply multiplying sLAIDI with the reflectancevalue at the prominent water absorption band 1555 nm, an index
oncentrations. The curves represent chlorophyll content varying from 2.50 μg/cm2 to
waveband ratio to determine Leaf Area Index in orchards, Remote
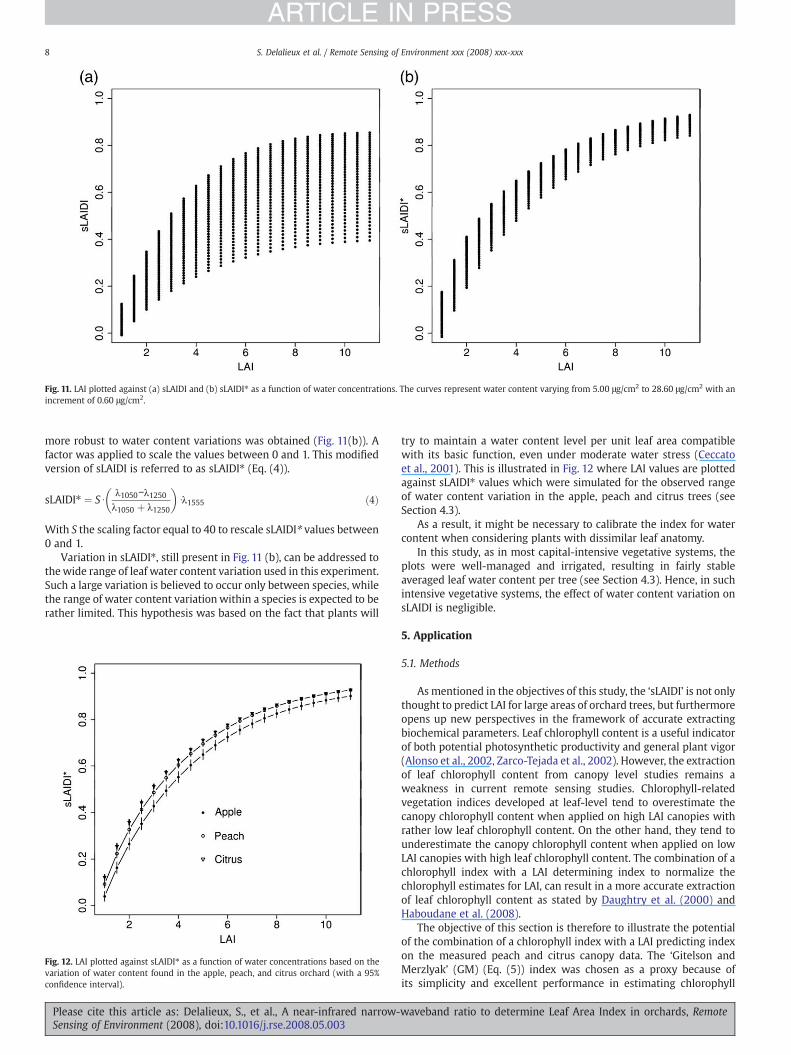
Fig. 11. LAI plotted against (a) sLAIDI and (b) sLAIDI⁎ as a function of water concentrations. The curves represent water content varying from 5.00 μg/cm2 to 28.60 μg/cm2 with anincrement of 0.60 μg/cm2.
8 S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
more robust to water content variations was obtained (Fig. 11(b)). Afactor was applied to scale the values between 0 and 1. This modifiedversion of sLAIDI is referred to as sLAIDI⁎ (Eq. (4)).
sLAIDI⁎ ¼ Sdλ1050−λ1250λ1050 þ λ1250
� �dλ1555 ð4Þ
With S the scaling factor equal to 40 to rescale sLAIDI⁎ values between0 and 1.
Variation in sLAIDI⁎, still present in Fig. 11 (b), can be addressed tothewide range of leaf water content variation used in this experiment.Such a large variation is believed to occur only between species, whilethe range of water content variationwithin a species is expected to berather limited. This hypothesis was based on the fact that plants will
Fig. 12. LAI plotted against sLAIDI⁎ as a function of water concentrations based on thevariation of water content found in the apple, peach, and citrus orchard (with a 95%confidence interval).
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
try to maintain a water content level per unit leaf area compatiblewith its basic function, even under moderate water stress (Ceccatoet al., 2001). This is illustrated in Fig. 12 where LAI values are plottedagainst sLAIDI⁎ values which were simulated for the observed rangeof water content variation in the apple, peach and citrus trees (seeSection 4.3).
As a result, it might be necessary to calibrate the index for watercontent when considering plants with dissimilar leaf anatomy.
In this study, as in most capital-intensive vegetative systems, theplots were well-managed and irrigated, resulting in fairly stableaveraged leaf water content per tree (see Section 4.3). Hence, in suchintensive vegetative systems, the effect of water content variation onsLAIDI is negligible.
5. Application
5.1. Methods
Asmentioned in the objectives of this study, the ‘sLAIDI’ is not onlythought to predict LAI for large areas of orchard trees, but furthermoreopens up new perspectives in the framework of accurate extractingbiochemical parameters. Leaf chlorophyll content is a useful indicatorof both potential photosynthetic productivity and general plant vigor(Alonso et al., 2002, Zarco-Tejada et al., 2002). However, the extractionof leaf chlorophyll content from canopy level studies remains aweakness in current remote sensing studies. Chlorophyll-relatedvegetation indices developed at leaf-level tend to overestimate thecanopy chlorophyll content when applied on high LAI canopies withrather low leaf chlorophyll content. On the other hand, they tend tounderestimate the canopy chlorophyll content when applied on lowLAI canopies with high leaf chlorophyll content. The combination of achlorophyll index with a LAI determining index to normalize thechlorophyll estimates for LAI, can result in a more accurate extractionof leaf chlorophyll content as stated by Daughtry et al. (2000) andHaboudane et al. (2008).
The objective of this section is therefore to illustrate the potentialof the combination of a chlorophyll index with a LAI predicting indexon the measured peach and citrus canopy data. The ‘Gitelson andMerzlyak’ (GM) (Eq. (5)) index was chosen as a proxy because ofits simplicity and excellent performance in estimating chlorophyll
waveband ratio to determine Leaf Area Index in orchards, Remote
9S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
content at leaf level (Delalieux et al., 2007b). This index is linearlyrelated to leaf chlorophyll content (Gitelson & Merzlyak, 1997).
GM ¼ λ750λ550
ð5Þ
The potential of this GM index to estimate leaf chlorophyll from acanopy dataset is tested by relating the obtained index values with theleaf chlorophyll content of the peach and citrus leaves obtained bydestructive chemical analysis. In a next step, the performance of theindex combined with the sLAIDI is evaluated to test whether thiscombined index performs better in predicting leaf chlorophyll contentfrom a canopy dataset.
5.2. Results and discussion
TheGM index iswell known as a good estimator for leaf chlorophyllcontent from spectral leaf data. However, this index would be muchmore useful if it could estimate leaf chlorophyll content from spectralcanopy data. The GM index was therefore applied on the peach andcitrus datasets to test its potential for predicting leaf chlorophyllcontent from canopy datasets. As described in Section 4.3, the peachdataset consisted of canopies with similar LAI values and distinctleaf chlorophyll contents as opposed to the citrus dataset which wascharacterized by fairly constant chlorophyll contents and strongvarying LAI values.
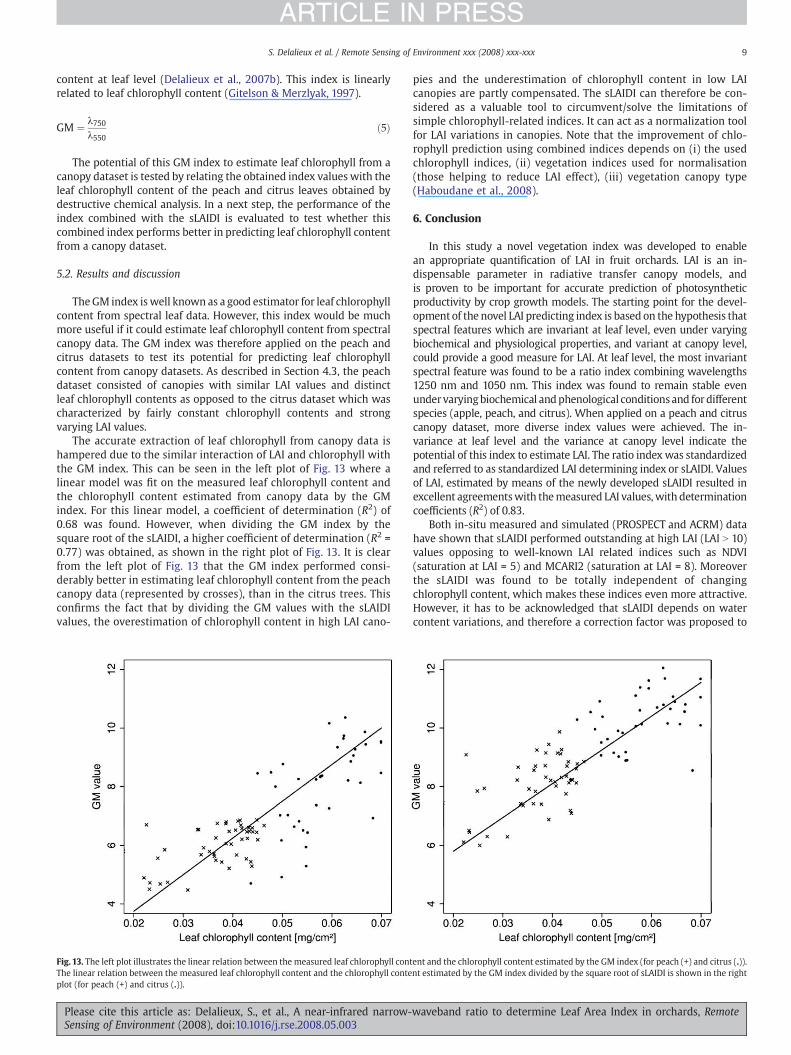
The accurate extraction of leaf chlorophyll from canopy data ishampered due to the similar interaction of LAI and chlorophyll withthe GM index. This can be seen in the left plot of Fig. 13 where alinear model was fit on the measured leaf chlorophyll content andthe chlorophyll content estimated from canopy data by the GMindex. For this linear model, a coefficient of determination (R2) of0.68 was found. However, when dividing the GM index by thesquare root of the sLAIDI, a higher coefficient of determination (R2 =0.77) was obtained, as shown in the right plot of Fig. 13. It is clearfrom the left plot of Fig. 13 that the GM index performed consi-derably better in estimating leaf chlorophyll content from the peachcanopy data (represented by crosses), than in the citrus trees. Thisconfirms the fact that by dividing the GM values with the sLAIDIvalues, the overestimation of chlorophyll content in high LAI cano-
Fig. 13. The left plot illustrates the linear relation between the measured leaf chlorophyll contThe linear relation between the measured leaf chlorophyll content and the chlorophyll conteplot (for peach (+) and citrus (.)).
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
pies and the underestimation of chlorophyll content in low LAIcanopies are partly compensated. The sLAIDI can therefore be con-sidered as a valuable tool to circumvent/solve the limitations ofsimple chlorophyll-related indices. It can act as a normalization toolfor LAI variations in canopies. Note that the improvement of chlo-rophyll prediction using combined indices depends on (i) the usedchlorophyll indices, (ii) vegetation indices used for normalisation(those helping to reduce LAI effect), (iii) vegetation canopy type(Haboudane et al., 2008).
6. Conclusion
In this study a novel vegetation index was developed to enablean appropriate quantification of LAI in fruit orchards. LAI is an in-dispensable parameter in radiative transfer canopy models, andis proven to be important for accurate prediction of photosyntheticproductivity by crop growth models. The starting point for the devel-opment of the novel LAI predicting index is based on the hypothesis thatspectral features which are invariant at leaf level, even under varyingbiochemical and physiological properties, and variant at canopy level,could provide a good measure for LAI. At leaf level, the most invariantspectral feature was found to be a ratio index combining wavelengths1250 nm and 1050 nm. This index was found to remain stable evenundervaryingbiochemical andphenological conditionsand fordifferentspecies (apple, peach, and citrus). When applied on a peach and citruscanopy dataset, more diverse index values were achieved. The in-variance at leaf level and the variance at canopy level indicate thepotential of this index to estimate LAI. The ratio indexwas standardizedand referred to as standardized LAI determining index or sLAIDI. Valuesof LAI, estimated by means of the newly developed sLAIDI resulted inexcellent agreementswith themeasured LAI values, with determinationcoefficients (R2) of 0.83.
Both in-situ measured and simulated (PROSPECT and ACRM) datahave shown that sLAIDI performed outstanding at high LAI (LAI N 10)values opposing to well-known LAI related indices such as NDVI(saturation at LAI = 5) and MCARI2 (saturation at LAI = 8). Moreoverthe sLAIDI was found to be totally independent of changingchlorophyll content, which makes these indices even more attractive.However, it has to be acknowledged that sLAIDI depends on watercontent variations, and therefore a correction factor was proposed to
ent and the chlorophyll content estimated by the GM index (for peach (+) and citrus (.)).nt estimated by the GM index divided by the square root of sLAIDI is shown in the right
waveband ratio to determine Leaf Area Index in orchards, Remote

10 S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
enable the application of this index on fields with a lot of watervariation in the plants. Due to the fact that the experimental plots inthis research study were well-managed and irrigated, no additionalcorrection factor was needed.
Results have demonstrated that combining a LAI predicting indexwith a chlorophyll-related index offers a more accurate prediction ofchlorophyll content. The performance of the newly developed LAIdetermining index was tested on fruit orchards, but we recognize theneed of testing the sLAIDI on other plant production systems before awidespread operational implementation can be aimed at.
Acknowledgements
This project was funded by the Katholieke Universiteit Leuven,Department of Biosystems. The authors further would like toacknowledge funding for preliminary research (Hypercrunch andHyperpeach project) provided by the Belgian Science Policy Office. Wealso acknowledge Dr. A. Kuusk and Dr. S. Jacquemoud for providing theACRM and PROSPECT code respectively.
References
Alonso, M., Rozados, M. J., Vega, J. A., Pérez-Gorostiaga, P., Cuiñas, P., Fontúrbel, M. T., &Fernández, C. (2002). Biochemical responses of Pinus pinaster trees to fire-inducedtrunk girdling and crown scorch: Secondary metabolites and pigments as needlechemical indicators. Journal of Chemical Ecology, 28, 687−700.
Arnon, D. I. (1994). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Betavulgaris. Plant Physiology, 24, 1−15.
Asner, G. P. (1998). Biophysical and biochemical sources of variability in canopy re-flectance. Remote Sensing of Environment, 64, 234−253.
Atzberger, C. (2004). Object-based retrieval of biophysical canopy variables usingartificial neural nets and radiative transfer models. Remote Sensing of Environment,93, 53−67.
Baret, F., & Guyot, G. (1991). Potentials and limits of vegetation indices for LAI and APARassessment. Remote Sensing of Environment, 35, 161−173.
Birky, A. K. (2001). NDVI and a simple model of deciduous forest seasonal dynamics.Ecological Modelling, 143, 43−58.
Borel, C. C., & Gerstl, S. A. W. (1994). Nonlinear spectral mixing models for vegetativeand soil surfaces. Remote Sensing of Environment, 47, 403−416.
Carter, G. A. (1994). Ratios of leaf reflectances in narrow wavebands as indicators ofplant stress. International Journal of Remote Sensing, 15, 697−703.
Carter, G. A., & Knapp, A. K. (2001). Leaf optical properties in higher plants: Linkingspectral characteristics to stress and chlorophyll concentration. American Journal ofBotany, 88, 677−684.
Ceccato, P., Flasse, S., Tarantola, S., Jacquemoud, S., & Grégoire, J. (2001). Detectingvegetation leaf water content using reflectance in the optical domain. RemoteSensing of Environment, 77, 22−33.
Cohen, S., & Fuchs, M. (1987). The distribution of leaf area, radiation photosynthesis andtranspiration in a shamouti orange hedgeroworchard Part I. Leaf area and radiation.Agricultural and Forest Meteorology, 40, 123−144.
Coppin, P., Jonckheere, I., Lambin, E., Nackaerts, K., & Muys, B. (2004). Methods innatural ecosystem monitoring: A review. International Journal of Remote Sensing,25, 1565−1569.
Daughtry, C. S. T., Walthall, C. L., Kim, M. S., Brown de Colstoun, E. McMurtrey III, J.E.(2000). Estimating Corn Leaf Chlorophyll Concentration from Leaf and CanopyReflectance. Remote Sensing of Environment, 74, 229−239.
Delalieux, S., van Aardt, J. A. N., Keulemans, W., Schrevens, E., & Coppin, P. (2007).Detection of biotic stress (Venturia inaequalis) in apple trees using hyperspectraldata: Non-parametric statistical approaches and physiological implications. Euro-pean Journal of Agronomy, 27, 130−143.
Delalieux, S., van Aardt, J. A. N., Zarco-Tejada, P. J., Kempeneers, P., Verstraeten, W. W., &Coppin, P. (2007). Development of robust hyperspectral indices for the detection ofdeviations of normal plant state. EARSeL eProceedings, 6, 82−93.
Fang, H., Liang, S., & Kuusk, A. (2003). Retrieving leaf area index using a geneticalgorithm with a canopy radiative transfer model. Remote Sensing of Environment,85, 257−270.
Gao, B. (1996). NDWI—A normalized difference water index for remote sensing ofvegetation water from space. Remote Sensing of Environment, 58, 257−266.
Gitelson, A. A., & Merzlyak, M. N. (1996). Signature analysis of leaf reflectance spectra:Algorithm development for remote sensing of chlorophyll. Journal of PlantPhysiology, 148, 494−500.
Gitelson, A., & Merzlyak, M. (1997). Remote estimation of chlorophyll content in higherplant leaves. International Journal of Remote Sensing, 18, 291−298.
Gong, P., Wang, D., & Liang, S. (1999). Inverting a canopy reflectance model using aneural network. International Journal of Remote Sensing, 20, 111−122.
Haboudane, D., Miller, J. R., Tremblay, N., Zarco-Tejada, P. J., & Dextraze, L. (2002).Integrated narrow-band vegetation indices for prediction of crop chlorophyllcontent for application to precision agriculture. Remote Sensing of Environment, 81,416−426.
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
Haboudane, D., Miller, J. R., Pattey, E., Zarco-Tejada, P. J., & Strachan, I. B. (2004).Hyperspectral vegetation indices and novel algorithms for predicting green LAI ofcrop canopies: Modeling and validation in the context of precision agriculture.Remote Sensing of Environment, 90, 337−352.
Haboudane, D., Tremblay, N., Miller, J. R., & Vigneault, P. (2008). Remote estimation ofcrop chlorophyll content using spectral indices derived from hyperspectral data.IEEE Transactions on Geosciences and remote sensing, 46, 423−437.
Hill, M. J., Held, A. A., Leuning, R., Coops, N. C., Hughes, D., & Cleugh, H. A. (2006). MODISspectral signals at a flux tower site: Relationships with high-resolution data, andCO2 flux and light use efficiency measurements. Remote Sensing of Environment,103, 351−368.
Huete, A. R. (1988). A soil vegetation adjusted index (SAVI). Remote Sening of Environment,25, 295−309.
Jacquemoud, S., & Baret, F. (1990). PROSPECT: A model of leaf optical properties spectra.Remote Sensing of Environment, 34, 75−91.
Jacquemoud, S., Verdebout, J., Schmück, G., Andreoli, G., & Hosgood, B. (1995).Investigation of leaf biochemistry by statistics. Remote Sensing of Environment, 54,180−188.
Jacquemoud, S., Ustin, S. L., Verdebout, J., Schmuck, G., Andreoli, G., & Hosgood, B.(1996). Estimating leaf biochemistry using the PROSPECT leaf optical propertiesmodel. Remote Sensing of Environment, 56, 194−202.
Jensen, J., Qiu, F., & Jim, M. (1999). Predictive modeling of coniferous forest age usingstatistical and artificial neural network approaches applied to remote sensor data.International Journal of Remote Sensing, 20, 2805−2822.
Jonckheere, I., Fleck, S., Nackaerts, K., Muys, B., Coppin, P., Weiss, M., & Baret, F. (2004).Reviewofmethods for in situ leaf area index determination. Part I. Theories, sensorsand hemispherical photography. Agricultural and Forest Meteorology, 121, 19−35.
Jordan, C. F. (1969). Deviation of leaf-area index from quality of light on the forest floor.Ecology, 50, 663−666.
Kalma, J. D., & Stanhill, G. (1969). The radiation climate of an irrigated orange plantation.Solar Energy, 12, 491−508.
Kempeneers, P., Zarco-Tejada, P. J., North, P. R. J., De Backer, S., Delalieux, S., Sepulcre-Canto,G., Morales, F., van Aardt, J. A. N., Sagardoy, R., Coppin, P. & Scheunders, P., (in press).Model inversion for chlorophyll estimation in open canopies from hyperspectralimagery. International Journal of Remote Sensing (2008). doi: 10.1080/0143116YYxx.
Kumar, L., Schmidt, K. S., Dury, S., & Skidmore, A. K. (2001). Imaging spectrometry andvegetation science (pp. 111–155) Dordrecht: Kluwer Academic Press.
Kuusk, A., & Nilson, T. (2000). A directional multispectral forest reflectance model.Remote Sensing of Environment, 72, 244−252.
Kuusk, A. (2001). A two-layer canopy reflectance model. Journal of QuantitativeSpectroscopy & Radiative Transfer, 71, 1−9.
MacHardy, W. E. (1996). Physiological responses to infections. Apple scab: Biology,epidemiology, and management (pp. 152−165). Minnesota: APS Press.
Norman, J. M., & Campbell, G. S. (1989). In R.W. Pearcy, J. R. Ehleringer, H. A. Mooney, &P. W. Rundel (Eds.), Plant physiological ecology: Field methods and instrumentation(pp. 301−325). London: Chapman & Hall.
Peñuelas, J., Gamon, J. A., Fredeen, A. L., Merino, J., & Field, C. B. (1994). Reflectanceindices associated with physiological changes in nitrogen- and water-limitedsunflower leaves. Remote Sensing of Environment, 48, 135−146.
Price, J. C. (1990). On the information content of soil reflectance spectra. Remote Sensingof Environment, 33, 113−121.
Qi, J., Kerr, Y. H., Moran, M. S., Weltz, M., Huete, A. R., Sorooshian, S., & Bryant, R. (2000).Leaf Area Index estimates using remotely sensed data and BRDF models in asemiarid region. Remote Sensing of Environment, 73, 18−30.
Rasmussen, M. (1997). Operational yield forecast using AVHRR NDVI data: Reduction ofenvironmental and inter-annual variability. International Journal of Remote Sensing,18, 1059−1077.
Rondeaux, G., Steven, M., & Baret, F. (1996). Optimization of soil-adjusted vegetationindices. Remote Sensing of Environment, 55, 95−107.
Rouse, J. W., Haas, R. H., Schelle, J. A., Deering, D. W., & Harlan, J. C. (1974). Monitoringthe vernal advancement and retrogradation (green wave effect) of naturalvegetation. Type III Final Report, NASA Goddard Space Flight Center, Green belt,Maryland.
Schlerf, M., Atzberger, C., & Hill, J. (2005). Remote sensing of forest biophysical variablesusing HyMap imaging spectrometer data. Remote Sensing of Environment, 95,177−194.
Schlerf, M., & Atzberger, C. (2006). Inversion of a forest reflectance model to estimatestructural canopy variables from hyperspectral remote sensing data. RemoteSensing of Environment, 100, 281−294.
Sims, D. A., & Gamon, J. A. (2002). Relationships between leaf pigment content andspectral reflectance across a wide range of species, leaf structures and develop-mental stages. Remote Sensing of Environment, 81, 337−354.
Tang, S., Chen, J. M., Zhu, Q., Li, X., Chen, M., Sun, R., Zhou, Y., Deng, F., & Xie, D. (2007).LAI inversion algorithm based on directional reflectance kernels. Journal ofEnvironmental Management, 85, 638−648.
Tucker, C. J. (1979). Red and photographic infrared linear combinations for monitoringvegetation. Remote Sensing of Environment, 8, 127−150.
Turner, D. P., Cohen, W. B., Kennedy, R. E., Fassnacht, K. S., & Briggs, J. M. (1999).Relationships between leaf area index and Landsat TM spectral vegetation indicesacross three temperate zone sites. Remote Sensing of Environment, 70, 52−68.
Turrell, F. M. (1961). Growth of the photosynthetic area of citrus. Botanical Gazette, 122,285−298.
Ustin, S. L., Roberts, D. A., Gardner, M., & Dennison, P. (2002). Evaluation of thepotential of Hyperion data to estimate wildfire hazard in the Santa Ynez FrontRange, Santa Barbara, California. In T. L. Stein (Ed.), International Geoscience andRemote Sensing Symposium 2002, Proceedings, and 24th Canadian Symposium
waveband ratio to determine Leaf Area Index in orchards, Remote

11S. Delalieux et al. / Remote Sensing of Environment xxx (2008) xxx-xxx
ARTICLE IN PRESS
on Remote Sensing, Toronto, Canada, 24–28 June 2002, IEEE, Piscataway, NJ (2002)(pp. 796−798).
Weiss, M., & Baret, F. (1999). Evaluation of canopy biophysical variable retrievalperformances from the accumulation of large swath satellite data. Remote Sensingof Environment, 70, 293−306.
Wulder, M. A., Le Drew, E. F., Franklin, S. E., & Lavigne, M. B. (1998). Aerial image textureinformation in the estimation of the northern deciduous and mixed wood forestleaf area index (LAI). Remote Sensing of Environment, 64, 64−76.
Zarco-Tejada, P. J., Miller, J. R., Noland, T. L., Mohammed, G. H., & Sampson, P. H. (2001).Scaling-up and model inversion methods with narrow-band optical indices for
Please cite this article as: Delalieux, S., et al., A near-infrared narrow-Sensing of Environment (2008), doi:10.1016/j.rse.2008.05.003
chlorophyll content estimation in closed forest canopies with hyperspectral data.IEEE Transactions on Geoscience and Remote Sensing, 39, 1491−1507.
Zarco-Tejada, P. J., Miller, J. R., Mohammed, G. H., Noland, T. L., & Sampson, P. H. (2002).Vegetation stress detection through chlorophyll a + b estimation and fluorescenceeffects on hyperspectral imagery. Journal of Environment Quality, 31, 1433−1441.
Zarco-Tejada, P. J., Miller, J. R., Morales, A., Berjon, A., & Agüera, J. (2004). Hyperspectralindices and model simulation for chlorophyll estimation in open canopy tree crops.Remote Sensing of Environment, 90, 463−476.
waveband ratio to determine Leaf Area Index in orchards, Remote




















