
A model for the evolution of high frequencies of males in an androdioecious plant based on a...
10
A model for the evolution of high frequencies of males in an androdioecious plant based on a cross-compatibility advantage of males CHRISTINE VASSILIADIS , MYRIAM VALERO* , PIERRE SAUMITOU-LAPRADE & BERNARD GODELLEà 1 Laboratoire de Ge ´ne ´tique et Evolution des Populations Ve ´ge ´tales, ESA-CNRS 8016, Ba ˆ t SN2, Universite ´ de Lille 1, F-59655 Villeneuve d’Ascq cedex, France and àLaboratoire d’Evolution et Syste ´matique des Ve ´ge ´taux, Universite ´ Paris Sud, Orsay, France Lloyd’s (1975) and Charlesworth & Charlesworth’s (1978) phenotypic selection models for the maintenance of androdioecy predict that males (female-sterile individuals) must have an advantage in fertility (K) of at least two in order to invade a hermaphroditic population, and that their equilibrium frequency (x eq (K ) 2)/2(K ) 1)) is always less than 0.5. In this paper, we develop a model in which male fertility is frequency-dependent, a situation not investigated in the previous models, to explore the conditions under which a high frequency of males (i.e. more than 50%) could be maintained at equilibrium. We demonstrate that a gametophytic self-incompatibility (GSI) locus linked to a nuclear sex determination locus can favour rare alleles through male function, by causing frequency- dependent selection. Thus, the spread of a female-sterility allele in a hermaphroditic population may be induced. In contrast with the previous models, our model can explain male frequencies greater than 50% in a functionally androdioecious species, as long as there is (i) dominance of female-sterility at the sex locus, and (ii) a few alleles at the self-incompatibility locus, even if the advantage in fertility of male phenotype is lower than two. Keywords: androdioecy, evolution, female-sterility, reproduction, self-incompatibility. Introduction The two symmetrical sexual dimorphisms: gynodioecy (co-occurrence of female with hermaphroditic plants) and androdioecy (males and hermaphrodites) are both encountered in Angiosperms. Gynodioecy is the second most common plant breeding system after hermaphro- ditism (7% and 72%, respectively, see Delannay, 1978; Richards, 1997), whereas androdioecy is remarkably rare (Yampolsky & Yampolsky, 1922; Charlesworth, 1984; Anderson & Symon, 1989; Vassiliadis, 1999). Lloyd (1975) and Charlesworth & Charlesworth (1978) developed phenotypic selection models (Lloyd, 1977) to understand the maintenance of these two sexual dimor- phisms in the context of evolution towards dioecy. These authors demonstrated that unisexual plants can- not invade a hermaphroditic population unless they have an advantage in fertility (K). Specifically, assuming nuclear inheritance of sex and a large panmictic popu- lation, Lloyd (1975) and Charlesworth & Charlesworth (1978) have demonstrated that unisexual plants must be at least twice as fit as hermaphrodites (K ‡ 2) and that their equilibrium frequency (x) is always less than 0.5: x K 2 2 K 1 : 1 Under these assumptions, as the same conditions hold for both male and female morphs, these phenotypic selection models or compensation models cannot explain why gynodioecy is more frequent than androdioecy. In a partially selfing hermaphroditic population, selective pressures for the spread of a unisexual mutant are no longer identical for male and female plants (Lloyd, 1975; Charlesworth & Charlesworth, 1978; Charlesworth, 1984). Female plants (always outcrossed) may be favoured when selfed hermaphrodites suer from inbreeding depression. On the other hand, in the case of a female-sterile mutant, the occurrence of selfing lowers male plant fertility as a result of the reduction in *Correspondence. E-mail: [email protected] 1 Present address: Laboratoire Ge´ nome, Populations, Interactions UMR CNS 500; case 063, F-34095 Montpellier cedex 05, France Heredity 85 (2000) 413–422 Received 10 December 1999, accepted 16 May 2000 Ó 2000 The Genetical Society of Great Britain. 413
-
Upload
paris-sorbonne -
Category
Documents
-
view
3 -
download
0
Transcript of A model for the evolution of high frequencies of males in an androdioecious plant based on a...

A model for the evolution of high frequenciesof males in an androdioecious plant based on
a cross-compatibility advantage of males
CHRISTINE VASSILIADIS , MYRIAM VALERO* , PIERRE SAUMITOU-LAPRADE & BERNARD GODELLEà1
Laboratoire de GeÂneÂtique et Evolution des Populations VeÂgeÂtales, ESA-CNRS 8016, Baà t SN2, Universite de Lille 1,F-59655 Villeneuve d'Ascq cedex, France and àLaboratoire d'Evolution et SysteÂmatique des VeÂgeÂtaux,
Universite Paris Sud, Orsay, France
Lloyd's (1975) and Charlesworth & Charlesworth's (1978) phenotypic selection models for themaintenance of androdioecy predict that males (female-sterile individuals) must have an advantage infertility (K) of at least two in order to invade a hermaphroditic population, and that their equilibriumfrequency (xeq � (K ) 2)/2(K ) 1)) is always less than 0.5. In this paper, we develop a model in whichmale fertility is frequency-dependent, a situation not investigated in the previous models, to explorethe conditions under which a high frequency of males (i.e. more than 50%) could be maintained atequilibrium. We demonstrate that a gametophytic self-incompatibility (GSI) locus linked to a nuclearsex determination locus can favour rare alleles through male function, by causing frequency-dependent selection. Thus, the spread of a female-sterility allele in a hermaphroditic population maybe induced. In contrast with the previous models, our model can explain male frequencies greater than50% in a functionally androdioecious species, as long as there is (i) dominance of female-sterility atthe sex locus, and (ii) a few alleles at the self-incompatibility locus, even if the advantage in fertility ofmale phenotype is lower than two.
Keywords: androdioecy, evolution, female-sterility, reproduction, self-incompatibility.
Introduction
The two symmetrical sexual dimorphisms: gynodioecy(co-occurrence of female with hermaphroditic plants)and androdioecy (males and hermaphrodites) are bothencountered in Angiosperms. Gynodioecy is the secondmost common plant breeding system after hermaphro-ditism (7% and 72%, respectively, see Delannay, 1978;Richards, 1997), whereas androdioecy is remarkablyrare (Yampolsky & Yampolsky, 1922; Charlesworth,1984; Anderson & Symon, 1989; Vassiliadis, 1999).Lloyd (1975) and Charlesworth & Charlesworth (1978)developed phenotypic selection models (Lloyd, 1977) tounderstand the maintenance of these two sexual dimor-phisms in the context of evolution towards dioecy.These authors demonstrated that unisexual plants can-not invade a hermaphroditic population unless theyhave an advantage in fertility (K). Speci®cally, assuming
nuclear inheritance of sex and a large panmictic popu-lation, Lloyd (1975) and Charlesworth & Charlesworth(1978) have demonstrated that unisexual plants must beat least twice as ®t as hermaphrodites (K ³ 2) and thattheir equilibrium frequency (x) is always less than 0.5:
x � K ÿ 2
2 K ÿ 1� � : �1�
Under these assumptions, as the same conditions holdfor both male and female morphs, these phenotypicselection models or compensation models cannot explainwhy gynodioecy is more frequent than androdioecy.In a partially sel®ng hermaphroditic population,
selective pressures for the spread of a unisexual mutantare no longer identical for male and female plants(Lloyd, 1975; Charlesworth & Charlesworth, 1978;Charlesworth, 1984). Female plants (always outcrossed)may be favoured when selfed hermaphrodites su�erfrom inbreeding depression. On the other hand, in thecase of a female-sterile mutant, the occurrence of sel®nglowers male plant fertility as a result of the reduction in
*Correspondence. E-mail: [email protected] address: Laboratoire Ge nome, Populations, Interactions
UMR CNS 500; case 063, F-34095 Montpellier cedex 05, France
Heredity 85 (2000) 413±422 Received 10 December 1999, accepted 16 May 2000
Ó 2000 The Genetical Society of Great Britain. 413

the number of available female gametes. Consequently,gynodioecy may be maintained even if female advantagein ®tness is less than two. But for androdioecy to bemaintained, the fertility advantage ofmalesmust bemorethan double that of hermaphrodites. This could explainthe rarity of androdioecy compared with gynodioecy.
Another possible explanation of the greater frequencyof gynodioecy in nature concerns sex determination insuch species. In many known gynodioecious species,expression of the sexual phenotype is the result of aninteraction between nuclear and cytoplasmic genes(Couvet et al., 1990), whereas in the only two well-studied androdioecious species, sex inheritance appearsto be nuclear. In Mercurialis annua there is onedominant gene for male phenotype (Pannell, 1997b);and in Datisca glomerata two recessive genes areinvolved (Wolf et al., 1997). Various models assuminga nucleo-cytoplasmic inheritance of sex (e.g. Lewis,1941; Couvet et al., 1986; Frank, 1989) have demon-strated that conditions for the maintenance of gyno-dioecy are less restrictive compared with simple nucleargenetic determination. Moreover, these models estab-lished that female plants under some conditions canreach equilibrium frequencies greater than 50%, partic-ularly when stochastic factors in¯uence the spatialdistribution of sex-determining genes (Couvet et al.,1998). High female frequencies are indeed oftenobserved in gynodioecious species (e.g. Domme e et al.,1983; Cuguen et al., 1994).
The existence of functional androdioecy (i.e. a speciesin which hermaphrodites have been proved to havee�ective male function) has been veri®ed in only sevenplant species so far (Oxalis suksdor®i, Orndu�, 1972;Datisca glomerata, Liston et al., 1990; Fritsch & Riese-berg, 1992; Mercurialis annua, Pannell, 1997c,d; Fraxi-nus lanuginosa, Ishida & Hiura, 1998; Schizopeponbryoniaefolius, Akimoto et al., 1999; Phillyrea angusti-folia, Lepart & Domme e, 1992; Vassiliadis et al. 2000;Fraxinus ornus, Domme e et al., 1999). Datisca glomerataand M. annua are characterized by a male advantagein fertility more than double that of hermaphrodites;they exhibit high outcrossing rates but are not self-incompatible, and male frequencies within populationsare low (<30%). In these species the maintenance ofmale individuals may be explicable according to Lloyd's(1975) and Charlesworth & Charlesworth's (1978)models. Contrary to theoretical expectations, hermaph-rodites of S. bryoniaefolius are highly endogamous.However, in this species, the outcrossing rate has beenreported to vary with male frequency (Akimoto et al.,1999) and male fertility is thus frequency-dependent, acondition not considered in previous models.
In P. angustifolia, male frequencies are close to 50%in most of the populations that have been studied in
southern France (review in Vassiliadis, 1999). This sexratio could suggest a functional dioecy. However, inprevious controlled-crosses experiments (Vassiliadiset al., 2000), we have shown that hermaphrodites didreproduce via their male function. Moreover, theestimated male advantage in siring success was only1.93 compared with hermaphrodites. In this previousstudy, hermaphrodites were shown to be self-incompat-ible and crossing success between hermaphrodites to bemore variable than between males and hermaphrodites,perhaps because of cross-incompatibility amonghermaphrodites. The male fertility of hermaphroditeshas also been examined in a natural population usingpaternity analysis (Vassiliadis, 1999), and is not close tozero: in P. angustifolia hermaphrodites are functionaland not cryptic females.
In the present paper, we explore how self-incompat-ibility may interact with sex determination in anandrodioecious species to favour the male plants andthus increase their frequency at equilibrium above thatpredicted by previous models. A model is developed inwhich a sex determination locus is completely genetic-ally linked to a self-incompatibility locus. The in¯uencesof three key parameters are discussed: dominance vs.recessiveness of female-sterility alleles, male advantagein fertility (K), and number of self-incompatibilityalleles (n).
Model assumptions and parameters
The model developed in this study considers one in®nitepopulation, with no pollen limitation, no overlappinggenerations, and no mutations. The initial populationcontains only hermaphroditic individuals (initial fre-quency of males x � 0). The model considers a gameto-phytic self-incompatibility (GSI) system which involvesone S-locus, with `n' alleles: S1, S2,¼,Sn. Under such asystem, Si pollen is compatible with any recipient sporo-phyte Sj/Sk (with j and k ¹ i). In the initial population,there are n(n ) 1)/2 di�erent diploid genotypes which areall heterozygotes as a result of the incompatibility system(Wright, 1939). At equilibrium, these genotypes are atequal frequencies, because of the negative frequency-dependent selection caused by self-incompatibilitysystems. Consequently, each Si allele is present at afrequency of 1/n in the population (Wright, 1939).
Starting from this equilibrium situation for self-incompatibility alleles, a second nuclear locus, notedA, governing the sexual phenotype is introduced. Twoalleles are possible at this locus. The allelic form Am
encodes for female-sterility (male phenotype) and Ah
causes female-fertility (hermaphroditic phenotype). Thetwo loci, S (self-incompatibility) and A (female-sterility),are in complete linkage disequilibrium.
414 C. VASSILIADIS ET AL.
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.

We examined two dominance relationships: female-sterility is either dominant over female-fertility (Am >Ah) or recessive (Am < Ah). (Analysis for the recessivecase is presented in the Appendix.)In the dominant case, female-sterile individuals are
heterozygotes and the female-sterility allele Am isassociated with only one self-incompatibility allele, S1.All other self-incompatibility alleles (S2¼Sn) play thesame role in the population for obvious reasons ofsymmetry. We will thus consider their frequencies to beidentical.Under these assumptions, in a population containing
n incompatibility alleles, each `regular' pollen genotype(not associated with female-sterility) will be incompat-ible if its incompatibility allele corresponds to one of thetwo alleles of the hermaphrodite recipient; these aredi�erent from each other and are both di�erent from S1
[probability � 2/(n ) 1)]. Because all non-male geno-types are present at the same frequency, all other`regular' pollen grains are thus compatible with aprobability of 1 ) (2/(n ) 1)) or (n ) 3)/(n ) 1) (see
Table 1). In the model presented in this paper, we de®nethe male advantage in fertility (K) as the amount ofpollen produced by a male divided by the amount ofpollen produced by a hermaphrodite. Moreover, weassume that no ®tness variation occurs within eachsexual phenotype.All of the di�erent possible genotypes and their
associated phenotypes, under the assumption of dom-inant female-sterility, are presented in Table 1(a), alongwith their relative male-fertilities and frequencies. Thecrossing table (Table 1b) among pollen donors andhermaphrodite recipients in generation t gives thecompatibility conditions for each cross, the frequencyof compatible pollen genotypes and the genotypesproduced in the next generation (t + 1). Not all thepollen types contribute to the next generation. There-fore, the frequency of a progeny genotype must beweighted by the frequency of compatible pollen overeach hermaphrodite recipient. This allows the calcula-tion of the male frequency in the generation t + 1 (x¢),given in the following recursion equation:
x0 �Kx2
Kx2� nÿ3
nÿ1ÿ �
Kx2� 1ÿ x
ÿ �� Kx2
Kx nÿ2nÿ1ÿ �� 1ÿ x� � nÿ3
nÿ1ÿ � : �2�
Results
Dominant model
The equilibrium male frequencies (xeq) were easily foundby solving the recursion eqn (2), for x¢ � x. The solutionof the resulting ®rst-degree equation is given in thefollowing equation:
xeq �K nÿ1
nÿ3ÿ �ÿ 2
2 K nÿ2nÿ3ÿ �ÿ 1
� � �3�
Table 1 Characteristics of crosses and progeny in thedominant model (Am > Ah): only one `male' haplotype isfound (S1Am) in the population that contains `n'incompatibility alleles
(a) Genotypes and associated phenotypes, relative malefertility and frequencies of individuals (there must be atleast three self-incompatibility alleles)
GenotypeSexual
phenotypeRelative
male fertility Frequency
S1Am=SiAh Male K xi ¹ 1SiAh=SjAh Hermaphrodite 1 1 ) xi ¹ 1, j ¹ 1, i ¹ j
(b) Crossing table among pollen donors and hermaphrodite recipients
Recipient genotype(frequency)
Pollen genotype(frequency)
SiAh/SjAh S1Am SkAh
(1) (Kx/2)* (Kx/2* + (1 ) x)**)
Compatibility conditions Always compatible k ¹ i and k ¹ j
Compatibility probability 1 1 ) 2/(n ) 1) = (n ) 3)/(n ) 1)
Progeny genotype S1Am/SiAh SiAh/SkAh
i ¹ 1 i ¹ 1, k ¹ 1, i ¹ k
Progeny frequency (Kx/2)/W ((Kx/2 + (1 ) x)) (n ) 3)/(n ) 1))/W
*Pollen produced by male; ** pollen produced by hermaphrodite; W: weighting factor because all pollen types do not contribute to the nextgeneration.
ANDRODIOECY REVISITED THROUGH INCOMPATIBILITY 415
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.

This result looks very similar to the equilibriumfrequency obtained by Lloyd (1975) and Charlesworth& Charlesworth (1978) given in eqn (1): x � Kÿ2
2 Kÿ1� �, withK in the denominator replaced by K 0 � K nÿ2
nÿ3ÿ �
> K andin the numerator by K nÿ1
nÿ3ÿ �
. Replacing K with K¢ makessense because the ratio nÿ2
nÿ3ÿ �
> 1 corresponds to thecross-compatibility advantage of males compared withhermaphrodites, as we shall demonstrate in the follow-ing paragraph.
The genotype of a male is S1Am/SiAh (i ¹ 1). Thus itproduces half S1Am and half SiAh pollen. The genotypeof a hermaphrodite is SjAh/SkAh (with j ¹ k, j and k ¹ 1;for instance S2Ah/S3Ah). S1 pollen grains are alwayscompatible with hermaphrodites, whereas Si pollengrains have a probability nÿ3
nÿ1 of being compatible:indeed, out of the n ) 1 possible alleles (i � 2¼n),n ) 3 alleles are compatible (i � 4¼n in our example).The mean probability that a pollen grain produced by amale will be compatible on a given stigma is thus:
1
2� nÿ 3
2 nÿ 1� � �nÿ 2
nÿ 1:
On the same stigma (say S2Ah/S3Ah), the mean prob-ability that a pollen grain produced by a hermaphroditewill be compatible is nÿ3
nÿ1.The ratio ��nÿ 2�=�nÿ 1��=��nÿ 3�=�nÿ 1�� � nÿ2
nÿ3 > 1is thus the cross-compatibility advantage of malesrelative to hermaphrodites. Such a di�erence betweenmales and hermaphrodites does not exist when S1 allelescan be either associated with the Ah allele or with Am
allele: in this case, the equilibrium male frequency is theone obtained with the compensation model, correspond-ing to eqn (1). When the alleles S1 andAm are in completelinkage disequilibrium, the cross-compatibility advan-tage alone cannot account for the di�erences in maleequilibrium frequency between our model and thecompensation model. Taking into account just thecross-compatibility advantage would lead to the follow-ing male frequency:
K nÿ2nÿ3ÿ �ÿ 2
2 K nÿ2nÿ3ÿ �ÿ 1
� � < K nÿ1nÿ3ÿ �ÿ 2
2 K nÿ2nÿ3ÿ �ÿ 1
� � � xeq:
The di�erence from the numerator in eqn (3) (given herein xeq) and indicated in bold characters can be explainedby comparing the recursion equation for x obtained inour model (eqn 2), rewritten using K 0 � K nÿ2
nÿ3ÿ �
:
x0 � 1
2
nÿ 1
nÿ 2
� �K 0x
K 0x� 1ÿ x� � ; �4�
with the recursion equation corresponding to the com-pensation model:
x0 � 1
2
K 0xK 0x� 1ÿ x� � : �5�
Equation (5) gives the expected proportion of males x¢in generation t + 1, when males represent a fraction xof the population at generation t, and produce K¢ moremale gametes than hermaphrodites. The factor 1/2represents the fact that half of male o�spring are males.In our model, this is no longer true. Males indeedproduce half S1Am gametes, and half SiAh gametes. Buton a given stigma, the probability of compatibility ofSiAh pollen grains is only nÿ3
nÿ1. The mean proportion ofo�spring bearing allele S1Am which will be male is thus12 =
12� 1
2nÿ3nÿ1ÿ �� � � 1
2nÿ1nÿ2ÿ �
which is greater than 1/2.
The combined e�ects of the cross-compatibilityadvantage �K 0 � K nÿ2
nÿ3ÿ �
> K� and of the biased sex-ratio in male o�spring 1
2nÿ1nÿ2ÿ �
> 12 can lead to very high
male frequencies (Fig. 1). For n � 3, the male frequencyreaches unity, leading to population extinction. Forhigher values of n, the male frequency is always higherthan the value (K ) 2)/2(K ) 1) predicted by the com-pensation model with a simple nuclear sex determin-ation (Fig. 1), although it approaches this limit forhigher values of n. Recall that for low values of maleadvantage, K (between 1 and 2, not shown), malescannot be maintained in the compensation model.In contrast, they can be maintained at rather higherfrequencies using our model. For instance, with K � 1.5and with six self-incompatibility alleles, eqn (2) showsthat 25% males can be maintained at equilibrium. Thisrequires a much higher male advantage (K � 3) under
Fig. 1 Male frequency at equilibrium in our dominant model(upper nappe, eqn 2) and in the compensation model (lower
nappe, eqn 1), as a function of the number of S-alleles (n, from3 to 12) and the relative male advantage in fertility (K, from 2to 11).
416 C. VASSILIADIS ET AL.
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.

the compensation model (eqn 1). In our model, eqn (3)shows that the frequency of males is higher than 0.5whenever K > n ) 3.In this model, hermaphrodites are not simply cryptic
females, because their pollen grains can fertilize theovules of other hermaphrodites. For instance, whenK � 2 and n � 5, there are 5 ´ (5 ) 1)/2 � 10 possibleplant genotypes that are all heterozygous at the self-incompatibility locus (Table 2). Among these 10 geno-types, four bear the dominant female-sterility allele(S1Am) and one of the other alleles (S2, S3, S4 or S5; seeTable 2). Considering a given hermaphroditic genotype(say S2S3, see Table 3), the proportion of S1Am pollengrains that is compatible is 0.5, which ensures that halfof the progeny are males. Of the remaining 50%compatible pollen grains (non-S1Am), half are producedby males, and half by ®ve di�erent types of hermaph-rodites, which therefore function as true hermaphroditesand not cryptic females. In the compensation model, in apopulation with 50% males (as half of male o�springare males), there cannot be any true hermaphroditesbecause the latter do not contribute to the subsequentgeneration via male gametes. However, in our model, itis possible to observe a population made up of 50%males and 50% hermaphrodites which all demonstrate areal male function. This is possible because of the biasedsex ratio in the progeny of males (as shown above).
Recessive model
When assuming recessive female-sterility, it is notpossible to solve the recursion equations for x¢ � xanalytically to ®nd the equilibrium frequency of males.We thus iterated the recursion equations numerically forvarious parameter sets (eqns A1, A2, A3 and A4) usingMATHEMATICAMATHEMATICA v.3.0 software (Wolfram, 1996). Becausethe female-sterility allele is recessive, males can only behomozygotes for this allele, and the only possiblegenotype for males will be S1Am/S2Am (Table A1a).The equilibrium male frequency is in general smaller
than in the compensation model, except for small K andsmall n (Fig. 2).
Discussion
Our model demonstrates that an incompatibilitysystem completely linked with a sex determinationlocus causes a frequency-dependent selection, and cantherefore favour rare alleles through male function.Thus, it may induce the spread of the female-sterilityallele (rare) within a hermaphroditic population.Contrary to the results predicted by Lloyd (1975)and Charlesworth & Charlesworth (1978), this modelcan explain male frequencies greater than 0.5 in a T
able
2Amountofpollen
grainsproducedin
apopulationatequilibrium
withatw
o-fold
relativefertilityadvantageformalesand®veself-incompatibility
alleles,the®fthallele(denotedS1Am)causingdominantfemale-sterility.In
thiscase,theequilibrium
frequency
ofmalesis0.5.HalfoftheS2pollen
grainsare
producedbyhermaphrodites
(3´1/24),halfare
producedbymales(1/8).AllS1Ampollen
grainsare
producedbymales
Hermaphrodites:totalfrequency
=0.5
Males:totalfrequency
=0.5
Genotype
S2Ah/S
3Ah
S2Ah/S
4Ah
S2Ah/S
5Ah
S3Ah/S
4Ah
S3Ah/S
5Ah
S4Ah/S
5Ah
S1Am/S
2Ah
S1Am/S
3Ah
S1Am/S
4Ah
S1Am/S
5Ah
Frequency
1/12
1/12
1/12
1/12
1/12
1/12
1/8
1/8
1/8
1/8
Male
fertility
11
11
11
22
22
S2Ah:1/24
S2Ah:1/24
S2Ah:1/24
S2Ah:1/8
S3Ah:1/24
S3Ah:1/24
S3Ah:1/24
S3Ah:1/8
Amountofpollen
S4Ah:1/24
S4Ah:1/24
S4Ah:1/24
S4Ah:1/8
grainsproduced
S5Ah:1/24
S5Ah:1/24
S5Ah:1/24
S5Ah:1/8
S1Am:1/8
S1Am:1/8
S1Am:1/8
S1Am:1/8
ANDRODIOECY REVISITED THROUGH INCOMPATIBILITY 417
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.
functionally androdioecious species. However, theequilibrium frequency of male phenotypes is highlydependent on three key parameters: the dominance offemale-sterility at the sex locus, the number of allelesat the self-incompatibility locus, and the ®tnessadvantage of the male morph.
In our dominant model, all Am alleles are found inS1Am haplotypes, which is expected if the female-sterility mutation arises only once, and the S1 allele isassociated only with Am: the S1Ah haplotype does notexist. Two explanations would lead to this case. In the®rst explanation, when the female-sterility mutation ®rstappears, S1 alleles are as common as any other S allele,and associated with the Ah allele in hermaphrodites.However, even if the S1Ah haplotype is initially quite
common, there is a selective process that reduces itsfrequency. As the S1Am male haplotype becomes com-mon, as a result of the male advantage coupled with thecompatibility advantage, the S1 allele will be at adisadvantage in hermaphrodites. Through this selectiveprocess, S1Am frequency increases and the S1Ah haplo-type, although not completely eliminated at equilibrium(result not shown), may be eliminated in small popula-tions by random drift, so that S1 remains only in theS1Am haplotype. In the second explanation, when thefemale-sterility mutation ®rst appears, this new allele,Am, has a pleiotropic e�ect on the self-incompatibilitylocus, which creates a new S-allele called S1. This S1
allele is then only associated with Am.We chose to model a GSI rather than a sporophytic
self-incompatibility for several reasons. First, GSI ismore widespread than SSI in angiosperms (Charles-worth, 1985). Secondly, GSI is simpler to implement andthus ideal for an initial approach. Thirdly, GSI is theonly system in which maleness can be associated with asingle S-allele. In the SSI case, the female-sterilehaplotype would never bene®t from an absolute cross-compatibility advantage.
In our study, the results of models are highlydependent on the dominance relation at the sex locus.If female-sterility is dominant, a high frequency of males(>50%) can be maintained within populations evenwhen their advantage in ®tness is low (K £ 2), as long asthe number of S-alleles is also low (n < 6) (Fig. 1).On the other hand, if female-sterility is recessive,equilibrium male frequency is smaller than or equal to0.5 (Fig. 2). When female-sterility is dominant, onlyone SI allele is associated with female-sterility (S1Am)and it is exclusively paternally inherited. This haplotypeis more frequently transmitted than others becauseit bene®ts from (i) a cross-compatibility advantage
Table 3 What happens in an S2Ah/S3Ah ¯ower. Only S4Ah, S5Ah and S1Am pollen grains germinate. Half of the ovules arefertilized by S1Am pollen grains, produced only by males. Half of the ovules are fertilized by S4 or S5 pollen grains, half ofwhich are produced by males, and the remaining half by hermaphrodites, which are thus not cryptic females (even though themale proportion in the population is 0.5). Males thus contribute all S1Am alleles and 1/2 of non-S1Am alleles, whereashermaphrodites contribute 1/2 of non-S1Am alleles
Pollen grains genotypes S2Ah S3Ah S4Ah S5Ah S1Am
Available amount issued from:Hermaphrodites 1/2 1/2 1/2 1/2 0Males 1/2 1/2 1/2 1/2 1
Pollen grain ®tness 0 0 1 1 1
Proportion of ovules sired 0 0 1/4 1/4 1/2
Pollen-donors contributionHermaphrodites 1/8 1/8 0Males 1/8 1/8 1/2
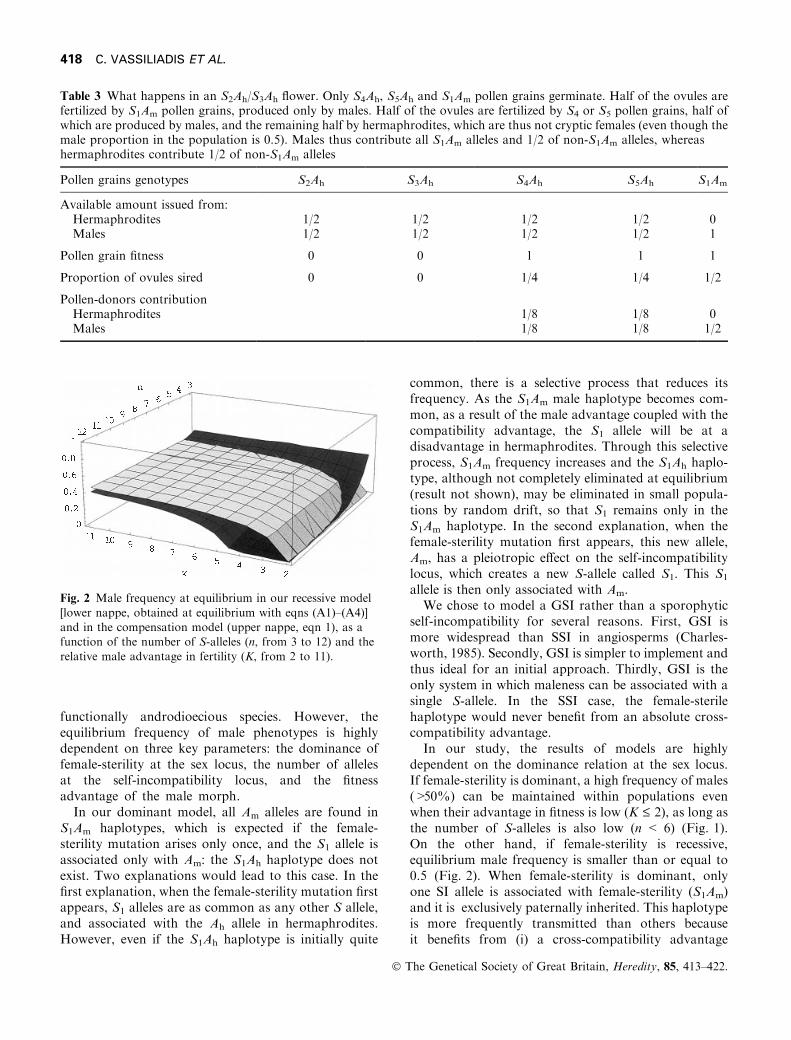
Fig. 2 Male frequency at equilibrium in our recessive model
[lower nappe, obtained at equilibrium with eqns (A1)±(A4)]and in the compensation model (upper nappe, eqn 1), as afunction of the number of S-alleles (n, from 3 to 12) and the
relative male advantage in fertility (K, from 2 to 11).
418 C. VASSILIADIS ET AL.
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.

nÿ2nÿ3 > 1 resulting from the allele S1 [males are alwayscompatible (fully or half-compatible) with all herma-phroditic genotypes]; (ii) a male-biased paternal sexratio 1
2nÿ1nÿ2ÿ �
> 12 resulting from a greater cross-compati-
bility of male-determining alleles; and (iii) a male ®tnessadvantage (K). The ®rst two phenomena are a result ofthe fact that males can interact with n ) 1 partners(di�erent genotypes), whereas hermaphrodites have onlyn ) 2 possible partners. Consequently, a small n causes ahigher male advantage than a large n, and the formersituation should be encountered more frequently insmall, recently founded populations (colonizationevent). However, as the number of SI alleles (n)increases, incompatibility among hermaphroditesdecreases and the male cross-compatibility advantagebecomes negligible. This can occur through mutation or,more probably, migration from other populations,especially in wind-pollinated species such as Phillyrea,Fraxinus, and Mercurialis. Consequently, when n ishigh, the di�erence in ®tness between males andhermaphrodites depends only on their relative advan-tage in male ®tness, and the equilibrium male frequencymatches the results of the compensation model.If female-sterility is recessive (see Appendix), the two
male haplotypes (S1Am and S2Am) can be found in aheterozygous state in hermaphroditic individuals so thatno haplotype is exclusively associated with males andpaternally inherited. Consequently, males bene®t fromonly a weak cross-compatibility advantage. Paradoxi-cally, we ®nd that the male advantage in ®tness (K)favours the frequency of hermaphrodites: as K increases,the frequency of S1Am and S2Am haplotypes increases inthe pollen pool, and providing that n is large enough,more hermaphrodite phenotypes (heterozygotes forS1Am or S2Am haplotypes) will be produced than malephenotypes. Consequently, when female-sterility isrecessive, the equilibrium male frequency is even smallerthan in the compensation model for large values of nand K (Fig. 2). The genetics of sex determination hasbeen studied in only two species; female-sterility isdominant in one species (Pannell, 1997b) and recessivein the other (Wolf et al., 1997). Therefore, it is notpossible to draw any conclusion about the dominance offemale-sterility.The number of S-alleles (n) that are maintained at
mutation±selection equilibrium within a ®nite popula-tion depends on population size N in a gametophyticself-incompatibility system (Wright, 1939; Vekemans &Slatkin, 1994; Schierup, 1998; Vekemans et al., 1998),and may be less than six when N < 50 (Vekemans &Slatkin, 1994; Schierup, 1998). This small value of nabove which a high frequency of males cannot bemaintained in our dominant model is thus plausible insmall populations.
The linkage between the two loci is a key assumptionin our model. Indeed, if the linkage between self-incompatibility and sex determination is not complete,our results coincide with those of the compensationmodel (results not shown). However, the recombinanthaplotype for the S-allele originally associated withfemale-fertility (S1Ah in the dominant model), will havea higher probability of being lost by drift than thenonrecombinant male haplotype (S1Am), provided thatn is small enough (see above discussion).The male haplotype (S1Am) can be regarded as a
sel®sh gene, which favours its own transmission to thedetriment of the others. Indeed, this exclusively pater-nally inherited haplotype not only suppresses the femalefunction, but also biases the sex ratio of its owno�spring and thus acts as a sex-ratio distorter. Selectionacts on the haplotype as a whole, and so recombinationbetween the two loci might be selected against. Suchsel®sh haplotypes, implying two loci that never recom-bine (carried in an inversion loop, for example), havealready been described (e.g. sex-ratio distorters; seereview in Werren & Beukeboom, 1998).In this study, we have modelled the evolution of male
frequency considering that androdioecy evolves fromhermaphroditism, as in all previous theoretical studies(Lloyd's (1975), Charlesworth & Charlesworth's (1978)phenotypic models as well as Pannell's (1997a) meta-population genetic model). However, the only clear datafavour androdioecy as a reversion from dioecy (Mercu-rialis annua, Pannell, 1997d). Because experimental dataare so scarce, we still cannot reject the hypothesis of anevolution from hermaphrodism. Moreover, in the caseof Phillyrea angustifolia, even if phylogenetic data(Wallander & Albert 2000) do not provide a su�cientresolution to determine whether androdioecy evolvedfrom dioecy or hermaphrodism, the presence of a relictpistil in male ¯owers, as the only signi®cant morpho-logical di�erence between male and hermaphrodite¯owers (Lepart & Domme e, 1992), strongly suggeststhat androdioecy has evolved from an hermaphroditeancestor.In conclusion, a gametophytic self-incompatibility
system linked to a nuclear sex determinism, by causing afrequency-dependent selection, may explain the highfrequencies of males recently reported in some andro-dioecious species (these include the functional andro-dioecious species P. angustifolia (Vassiliadis, 1999) andin two other potentially androdioecious species, Fraxinuslanuginosa (10±49.7% males in populations) (Ishida &Hiura, 1998) and Fraxinus ornus (50% males in popu-lations) (Domme e et al., 1999), for which the pollen ofhermaphrodites has been proven potentially fertile). Theconditions required by the model to explain the main-tenance of high male frequency within a hermaphroditic
ANDRODIOECY REVISITED THROUGH INCOMPATIBILITY 419
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.

population are dominance of female-sterility (Am > Ah)with a complete linkage between Am and S1 alleles, anda few S-alleles. Controlled crosses can easily test bothpredictions of this model.
Acknowledgements
We thank Xavier Vekemans, Carolyn R. Engel andTean J. Mitchell for their suggestions on improving themanuscript, and two anonymous reviewers for theirdetailed critical reviews. The study was supported byC. Vassilidis.
References
AKIMOTOAKIMOTO, J.J., FUKUHARAFUKUHARA, T.T. ANDAND KIKUZAWAKIKUZAWA, K.K. 1999. Sex ratiosand genetic variation in a functionally androdioecious
species, Schizopepon bryoniaefolius (Cucurbitaceae). Am. J.Bot., 86, 880±886.
ANDERSONANDERSON, G. J.G. J. ANDAND SYMONSYMON, D. E.D. E. 1989. Functional dioecy and
andromonoecy in Solanum. Evolution, 43, 204±219.CHARLESWORTHCHARLESWORTH, B.B. ANDAND CHARLESWORTHCHARLESWORTH, D.D. 1978. A model forthe evolution of dioecy and gynodioecy. Am. Nat., 112,
975±997.CHARLESWORTHCHARLESWORTH, D.D. 1984. Androdioecy and the evolution ofdioecy. Biol. J. Linn. Soc., 23, 333±348.
CHARLESWORTHCHARLESWORTH, D.D. 1985. Distribution of dioecy and self-incompatibility in angiosperms. In: Greenwood, P. J.,Harvey, P. H. and Slatkin, M. (eds) Essays in Honour ofJohn Maynard Smith, pp. 237±268. Cambridge University
Press, Cambridge.COUVETCOUVET, D.D., BONNEMAISONBONNEMAISON, F.F. ANDAND GOUYONGOUYON, P. H.P. H. 1986. Themaintenance of females among hermaphrodites: the impor-
tance of nuclear-cytoplasmic interactions. Heredity, 57,325±330.
COUVETCOUVET, D.D., RONCERONCE, O.O. ANDAND GLIDDONGLIDDON, C.C. 1998. The maintenance
of nucleocytoplasmic polymorphism in a metapopulation:the case of gynodioecy. Am. Nat., 152, 59±70.
COUVETCOUVET, D.D., ATLANATLAN, A.A., BELHASSENBELHASSEN, E.E., GLIDDONGLIDDON, C. J.C. J., GOUYONGOUYON,
P.-H.P.-H. ANDAND KJELLBERGKJELLBERG, F.F. 1990. Coevolution between twosymbionts: the case of cytoplasmic male sterility in higherplants. Oxford Surv. Evol. Biol., 7, 225±249.
CUGUENCUGUEN, J.J., WATTIERWATTIER, R.R., SAUMITOUSAUMITOU--LAPRADELAPRADE, P.P., FORCIOLIFORCIOLI, D.D.,
MORCHENMOÈ RCHEN, M.M., VANVAN DIJKDIJK, H.H. AND VERNET, P.AND VERNET, P. 1994. Gynodioecyand mtDNA polymorphism in natural populations of thegynodioecious species Beta vulgaris ssp. maritima. GeÂneÂt.
SeÂl. EÂvol., 26, 87±101.DELANNAYDELANNAY, X.X. 1978. La gynodioecie chez les angiospermes.Nat. Belges, 59, 223±235.
DOMMEEDOMME E, B.B., GUILLERMGUILLERM, J. L.J. L. ANDAND VALDEYRONVALDEYRON, G.G. 1983. Re gimede reproduction et he te rozygotie des populations de Thymusvulgaris L., dans une succession postculturale. C. r. Acad.Sci. Paris, 296, 111±114.
DOMMEEDOMMEÂ E, B.B., GESLOTGESLOT, A.A., THOMPSONTHOMPSON, J. D.J. D., REILLEREILLE, M.M. ANDAND
DENELLEDENELLE, N.N. 1999. Androdioecy in the entomophilous treeFraxinus ornus (Oleaceae). New Phytol., 143, 419±426.
FRANKFRANK, S. A.S. A. 1989. The evolutionary dynamics of cytoplasmic
male sterility. Am. Nat., 133, 345±376.FRITSCHFRITSCH, P.P. ANDAND RIESEBERGRIESEBERG, L. H.L. H. 1992. High outcrossing ratesmaintain male and hermaphrodite individuals in popula-
tions of the ¯owering plant Datisca glomerata. Nature, 359,633±636.
ISHIDAISHIDA, K.K. ANDAND HIURAHIURA, T.T. 1998. Pollen fertility and ¯owering
phenology in an androdioecious tree, Fraxinus lanuginosa(Oleaceae), in Hokkaido, Japan. Int. J. Pl. Sci., 159, 941±947.
LEPARTLEPART, J.J. ANDAND DOMMEEDOMMEÂ E, B.B. 1992. Is Phillyrea angustifolia L.(Oleaceae) an androdioecious species? Bot. J. Linn. Soc.,
108, 375±387.LEWISLEWIS, D.D. 1941. Male-sterility in natural populations ofhermaphrodite plants. New Phytol., 40, 56±63.
LISTONLISTON, A.A., RIESEBERGRIESEBERG, L. H.L. H. ANDAND ELIASELIAS, T. S.T. S. 1990. Functionalandrodioecy in the ¯owering plant Datisca glomerata.Nature, 343, 641±642.
LLOYDLLOYD, D. G.D. G. 1975. The maintenance of gynodioecy andandrodioecy in angiosperms. Genetica, 45, 325±339.
LLOYDLLOYD, D. G.D. G. 1977. Genetic and phenotypic models of naturalselection. J. Theor. Biol., 69, 543±560.
ORNDUFF, R.ORNDUFF, R. 1972. The breakdown of trimorphic incompat-ibility in Oxalis section Corniculatae. Evolution, 26, 52±65.
PANNELLPANNELL, J.J. 1997a. The maintenance of gynodioecy and
androdioecy in a metapopulation. Evolution, 51, 10±20.PANNELLPANNELL, J.J. 1997b. Mixed genetic and environmental sexdetermination in an androdioecious population of Mercu-
rialis annua. Heredity, 78, 50±56.PANNELLPANNELL, J.J. 1997c. Variation in sex ratios and sex allocation inandrodioecious Mercurialis annua. J. Ecol., 85, 57±69.
PANNELLPANNELL, J.J. 1997d. Widespread functional androdioecy inMercurialis annua L. (Euphorbiaceae). Biol. J. Linn. Soc.,61, 95±116.
RICHARDSRICHARDS, A. J.A. J. 1997. Plant Breeding Systems, 2nd edn.
Chapman & Hall, London.SCHIERUPSCHIERUP, M. H.M. H. 1998. The number of self-incompatibilityalleles in a ®nite, subdivided population. Genetics, 149,
1153±1162.VASSILIADISVASSILIADIS, C.C. 1999. Evolution et maintien de l'androdioeÂcie.Etude theÂorique et approches expeÂrimentales chez Phillyrea
angustifolia L. Ph.D. Thesis. Universite des Sciences etTechnologies de Lille, Lille.
VASSILIADISVASSILIADIS, C.C., LEPARTLEPART, J.J., SAUMITOUSAUMITOU--LAPRADELAPRADE, P.P. ANDAND
VERNETVERNET, P., P. 2000. Self-incompatibility and male fertilisation
success in Phillyrea angustifolia (Oleaceae). Int. J. Pl. Sci.,161, 393±402.
VEKEMANSVEKEMANS, X.X. ANDAND SLATKINSLATKIN, M.M. 1994. Gene and allelic
genealogies at a gametophytic self-incompatibility locus.Genetics, 137, 1157±1165.
VEKEMANSVEKEMANS, X.X., SCHIERUPSCHIERUP, M. H.M. H. ANDAND CHRISTIANSENCHRISTIANSEN, F. B.F. B. 1998.
Mate availability and fecundity selection in multi-allelic self-incompatibility systems in plants. Evolution, 52, 19±29.
WALLANDERWALLANDER, E.E. ANDAND ALBERTALBERT, V. A.V. A. 2000. Phylogeny and
classi®cation of Oleaceae based on RPS16 and TRNL-Fsequence data. Am. J. Bot., 87, 1827±1841.
WERRENWERREN, J. H.J. H. ANDAND BEUKEBOOMBEUKEBOOM, L. W.L. W. 1998. Sex determination,sex ratios, and genetic con¯ict. Ann. Rev. Ecol. Syst., 29,
233±261.
420 C. VASSILIADIS ET AL.
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.

WOLFWOLF, D. E.D. E., RIESEBERGRIESEBERG, L. H.L. H. ANDAND SPENCERSPENCER, S. C.S. C. 1997. The
genetic mechanism of sex determination in the androdioe-cious ¯owering plant, Datisca glomerata (Datiscaceae).Heredity, 78, 190±204.
WOLFRAMWOLFRAM, S.S. 1996. The MATHEMATICAATHEMATICA Book. Wolfram Media/Cambridge University Press, Cambridge.
WRIGHTWRIGHT, S.S. 1939. The distribution of self-sterility alleles in
populations. Genetics, 24, 538±552.YAMPOLSKYYAMPOLSKY, C.C. ANDAND YAMPOLSKYYAMPOLSKY, H.H. 1922. Distribution of sexforms in the phanerogamic ¯ora. Bibl. Genet., 3, 1±62.
Appendix: recessive female sterility
If female-sterility is recessive (Am < Ah), males must behomozygous for this allele, which would never occur ifonly one SI allele is associated with the sterility allele.We thus assumed that two incompatibility alleles (S1
and S2) are in complete linkage disequilibrium with thesterility allele (Am). All other self-incompatibility alleles(S3¼Sn) play the same role in the population forobvious symmetry reasons, and we will thus considertheir frequencies to be identical.A `regular' pollen grain (neither S1 nor S2) will be
incompatible if its incompatibility allele corresponds toone of the two alleles of the recipient which are di�erentfrom each other, and both di�erent from S1 and S2
(probability=2/(n ) 2)). All other pollen genotypes arecompatible with a probability of 1 ± (2/(n ) 2)) or(n ) 4)/(n ) 2) (see Table A1). In addition, situationswhere a `regular' pollen grain arrives on the stigma of ahermaphrodite bearing one S-allele associated with thefemale-sterility allele (i.e. S1 or S2), the other being anyother S-allele must be considered. The compatibilityprobability in that case is (n ) 3)/(n ) 2) (see Table A1).
The di�erent genotypes of the individuals under thisassumption, their associated phenotypes, relative malefertility, and frequencies are presented in Table A1(a).The frequency of compatibility of a pollen genotypewith a hermaphrodite recipient SjAh/SkAh (j ¹ k, j andk ¹ 1), and the phenotype of the progeny produced areshown in Table A1(b). This allows the calculation of themale frequency in generation t + 1 (x¢), according tothe four following recursion equations:
x0 � y2y � z
Kx� yKx� y � 2 nÿ3
nÿ2ÿ �
y � z� �
" #�A1�
y0 � 1
2 2y � z� � y � z Kx� y� �Kx� y � nÿ4
nÿ2ÿ �
y � z� �
" #�A2�
t0 � y0 �A3�
z0 � y � z2y � z
2y nÿ3nÿ2ÿ �
Kx� y � nÿ3nÿ2ÿ �
y � z� � �z nÿ4
nÿ2ÿ �
Kx� y � nÿ4nÿ2ÿ �
y � z� �
" #�A4�
Table A1: Characteristics of crossesand progeny in the recessive model(Ah > Am): two `male' haplotypes arefound (S1Am and S2Am) in thepopulation which contains nincompatibility alleles
(a) Genotypes and associatedphenotypes, relative male fertility andfrequencies of individuals (there mustbe at least three self-incompatibilityalleles)
GenotypeSexual
phenotypeRelative male
fertilityFrequency
x + y + z + t = 1
S1Am=S2Am Male K x
S1Am=SiAh Hermaphrodite 1 y
i ¹ 1 and 2
S2Am=SiAh Hermaphrodite 1 t*
i ¹ 1 and 2
SiAh=SjAh Hermaphrodite 1 z
i and j ¹ 1 and 2, i ¹ j
*For obvious reasons of symmetry, t = y.
ANDRODIOECY REVISITED THROUGH INCOMPATIBILITY 421
Ó The Genetical Society of Great Britain, Heredity, 85, 413±422.
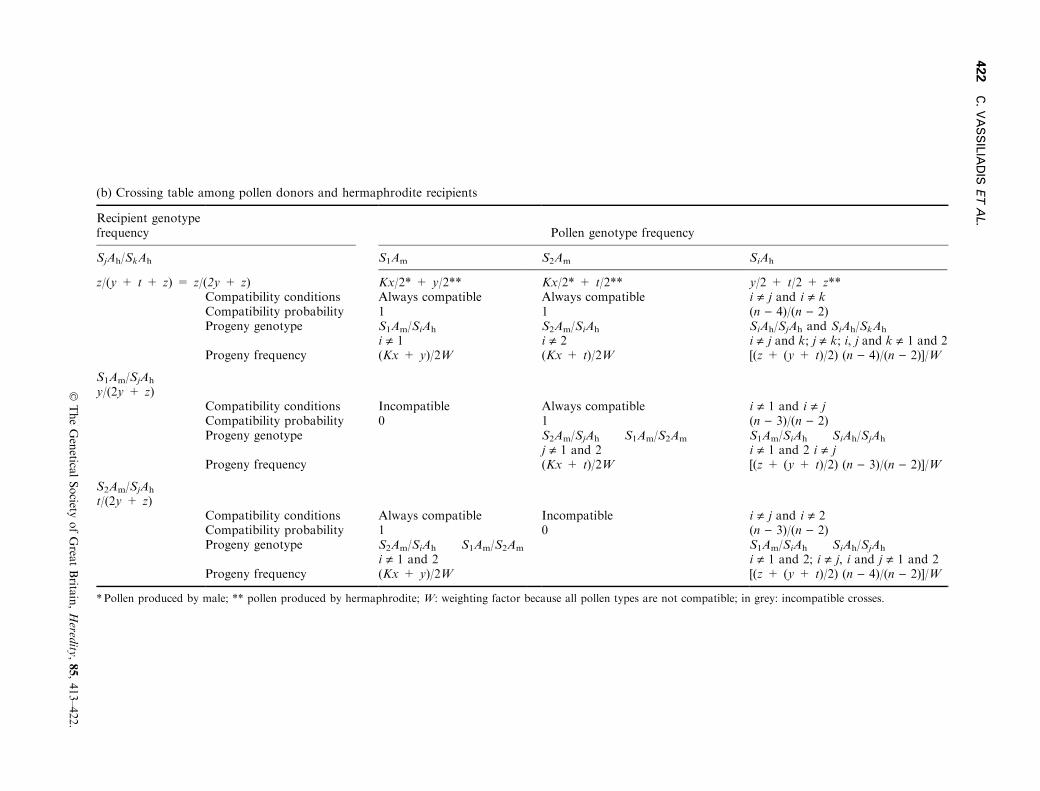
(b) Crossing table among pollen donors and hermaphrodite recipients
Recipient genotypefrequency Pollen genotype frequency
SjAh/SkAh S1Am S2Am SiAh
z/(y + t + z) = z/(2y + z) Kx/2* + y/2** Kx/2* + t/2** y/2 + t/2 + z**Compatibility conditions Always compatible Always compatible i ¹ j and i ¹ kCompatibility probability 1 1 (n ) 4)/(n ) 2)Progeny genotype S1Am/SiAh S2Am/SiAh SiAh/SjAh and SiAh/SkAh
i ¹ 1 i ¹ 2 i ¹ j and k; j ¹ k; i, j and k ¹ 1 and 2Progeny frequency (Kx + y)/2W (Kx + t)/2W [(z + (y + t)/2) (n ) 4)/(n ) 2)]/W
S1Am/SjAh
y/(2y + z)Compatibility conditions Incompatible Always compatible i ¹ 1 and i ¹ jCompatibility probability 0 1 (n ) 3)/(n ) 2)Progeny genotype S2Am/SjAh S1Am/S2Am S1Am/SiAh SiAh/SjAh
j ¹ 1 and 2 i ¹ 1 and 2 i ¹ jProgeny frequency (Kx + t)/2W [(z + (y + t)/2) (n ) 3)/(n ) 2)]/W
S2Am/SjAh
t/(2y + z)Compatibility conditions Always compatible Incompatible i ¹ j and i ¹ 2Compatibility probability 1 0 (n ) 3)/(n ) 2)Progeny genotype S2Am/SiAh S1Am/S2Am S1Am/SiAh SiAh/SjAh
i ¹ 1 and 2 i ¹ 1 and 2; i ¹ j, i and j ¹ 1 and 2Progeny frequency (Kx + y)/2W [(z + (y + t)/2) (n ) 4)/(n ) 2)]/W
* Pollen produced by male; ** pollen produced by hermaphrodite; W: weighting factor because all pollen types are not compatible; in grey: incompatible crosses.
422
C.
VA
SS
ILIA
DIS
ET
AL.
ÓTheGenetica
lSociety
ofGrea
tBrita
in,Hered
ity,85,413±422.

![Dr T Malakoutian.ppt [Compatibility Mode]](https://static.fdokumen.com/doc/165x107/63364bfed2b7284203084459/dr-t-malakoutianppt-compatibility-mode.jpg)

![0. Otwarcie Konferencji_[Compatibility Mode] - DAFA](https://static.fdokumen.com/doc/165x107/6326d44d6d480576770cfa67/0-otwarcie-konferencjicompatibility-mode-dafa.jpg)


![Ruth Knight presentation.ppt [Compatibility Mode]](https://static.fdokumen.com/doc/165x107/631d5d013ba403638902baaf/ruth-knight-presentationppt-compatibility-mode.jpg)











![Q1_FY_2012_final_290711.ppt [Read-Only] [Compatibility ... - Infinity](https://static.fdokumen.com/doc/165x107/63207975e9691360fe01ce09/q1fy2012final290711ppt-read-only-compatibility-infinity.jpg)

