
A habitat model for brown bear conservation and land use planning in the central Apennines
10
A habitat model for brown bear conservation and land use planning in the central Apennines Mario Posillico a, * , Alberto Meriggi b , Elisa Pagnin c , Sandro Lovari c , Luigi Russo d a Gestione ex Azienda di Stato Foreste Demaniali, 67031 Castel di Sangro (AQ), Italy b Dipartimento di Biologia Animale, Universit a di Pavia, 27100 Pavia, Italy c Dipartimento di Scienze Ambientali, Sezione di Ecologia Comportamentale, Etologia e Gestione della Fauna, Universit a di Siena, 53100 Siena, Italy d Parco Naturale Regionale Monti Simbruini, 00020 Jenne (RM), Italy Received 23 November 2002; received in revised form 23 July 2003; accepted 29 July 2003 Abstract We investigated the brown bear habitat suitability in an 8000 km 2 study area encompassing Abruzzo, Latium, and Molise regions in central-southern Italy. Based on long-term field surveys and published records, we classified bear habitat as occupied or unoc- cupied in 92 out of 320 sample squares (5 5 km). For each sample square 36 habitat variables were measured from topographic maps and Corine land-cover III level digital maps. The influence of habitat features on bear presence was investigated by multi- variate and one-way analyses of variance and by logistic regression analysis. The logistic model correctly classified 95.5% of sample squares of bear presence and 93.8% of those where bears were absent. Average altitude, deciduous woodlands and ecotone length, showed a positive relationship with bear presence, whereas vineyard-olive groves and shrublands were negatively correlated with bear presence. No specific land management guidelines or strategies exist for bear conservation in central Italy, based on knowledge of habitat–population relationships. The landscape scale habitat model we developed could be useful to predict bear occurrence, to identify critical areas for a brown bear conservation strategy, and to enhance the arrangement of the protected areas network for the conservation of this species. Ó 2003 Elsevier Ltd. All rights reserved. Keywords: Ursus arctos; Distribution; Habitat modelling; Management; Central Italy 1. Introduction Worldwide, especially throughout densely populated landscapes like those in central-southern Europe, hu- man activities have often deprived wildlife of the most suitable habitat, confining large carnivores, e.g. bears, to remote or less accessible areas (Swenson et al., 2000). Once distributed along most of the Apennine mountain chain, the brown bear (Ursus arctos) range in peninsular Italy was gradually reduced because of habitat loss and direct persecution (Febbo and Pellegrini, 1990). Today, brown bears in central Italy exist as a remnant, isolated population inhabiting 3000–5000 km 2 from Molise to Marche and Umbria regions (Fig. 1). However, the most densely and steadily occupied area is about 1500–2500 km 2 wide, in the Apennine mountains of southern Ab- ruzzo, Latium and Molise regions, surrounding the Abruzzo National Park (Meriggi et al., 2001). Minimum bear population size, estimated by snow tracking in 1985 over 600 km 2 (Abruzzo National Park + 200 km 2 of its buffer area), was 38–39 bears (Boscagli, 1990). No comprehensive estimates of population size have been carried out, but a preliminary evaluation of average bear density has been made by two DNA fingerprinting studies through a part of the ‘‘core’’ distribution range. These suggest a population density of 1 bear/50–80 km 2 (Lorenzini and Posillico, 2000; Forest Service, unpub- lished data). At such densities, a small bear population (e.g. 60 bears) might be distributed over a 3000–4800 km 2 area. This fully protected, threatened bear Biological Conservation 118 (2004) 141–150 www.elsevier.com/locate/biocon BIOLOGICAL CONSERVATION * Corresponding author. Present address: Dipartimento di Scienze Ambientali, Sezione di Ecologia Comportamentale, Etologia e Gesti- one della Fauna, Universit a di Siena, via P.A. Mattioli 4, 53100 Siena, Italy. Tel.: +39-0577-23-2953/4; fax: +39-0577-23-2825. E-mail address: [email protected] (M. Posillico). 0006-3207/$ - see front matter Ó 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocon.2003.07.017
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A habitat model for brown bear conservation and land use planning in the central Apennines

BIOLOGICAL
CONSERVATION
Biological Conservation 118 (2004) 141–150
www.elsevier.com/locate/biocon
A habitat model for brown bear conservation andland use planning in the central Apennines
Mario Posillico a,*, Alberto Meriggi b, Elisa Pagnin c, Sandro Lovari c, Luigi Russo d
a Gestione ex Azienda di Stato Foreste Demaniali, 67031 Castel di Sangro (AQ), Italyb Dipartimento di Biologia Animale, Universit�a di Pavia, 27100 Pavia, Italy
c Dipartimento di Scienze Ambientali, Sezione di Ecologia Comportamentale, Etologia e Gestione della Fauna, Universit�a di Siena, 53100 Siena, Italyd Parco Naturale Regionale Monti Simbruini, 00020 Jenne (RM), Italy
Received 23 November 2002; received in revised form 23 July 2003; accepted 29 July 2003
Abstract
We investigated the brown bear habitat suitability in an 8000 km2 study area encompassing Abruzzo, Latium, and Molise regions
in central-southern Italy. Based on long-term field surveys and published records, we classified bear habitat as occupied or unoc-
cupied in 92 out of 320 sample squares (5� 5 km). For each sample square 36 habitat variables were measured from topographic
maps and Corine land-cover III level digital maps. The influence of habitat features on bear presence was investigated by multi-
variate and one-way analyses of variance and by logistic regression analysis. The logistic model correctly classified 95.5% of sample
squares of bear presence and 93.8% of those where bears were absent. Average altitude, deciduous woodlands and ecotone length,
showed a positive relationship with bear presence, whereas vineyard-olive groves and shrublands were negatively correlated with
bear presence. No specific land management guidelines or strategies exist for bear conservation in central Italy, based on knowledge
of habitat–population relationships. The landscape scale habitat model we developed could be useful to predict bear occurrence, to
identify critical areas for a brown bear conservation strategy, and to enhance the arrangement of the protected areas network for the
conservation of this species.
� 2003 Elsevier Ltd. All rights reserved.
Keywords: Ursus arctos; Distribution; Habitat modelling; Management; Central Italy
1. Introduction
Worldwide, especially throughout densely populated
landscapes like those in central-southern Europe, hu-
man activities have often deprived wildlife of the mostsuitable habitat, confining large carnivores, e.g. bears, to
remote or less accessible areas (Swenson et al., 2000).
Once distributed along most of the Apennine mountain
chain, the brown bear (Ursus arctos) range in peninsular
Italy was gradually reduced because of habitat loss and
direct persecution (Febbo and Pellegrini, 1990). Today,
brown bears in central Italy exist as a remnant, isolated
* Corresponding author. Present address: Dipartimento di Scienze
Ambientali, Sezione di Ecologia Comportamentale, Etologia e Gesti-
one della Fauna, Universit�a di Siena, via P.A. Mattioli 4, 53100 Siena,
Italy. Tel.: +39-0577-23-2953/4; fax: +39-0577-23-2825.
E-mail address: [email protected] (M. Posillico).
0006-3207/$ - see front matter � 2003 Elsevier Ltd. All rights reserved.
doi:10.1016/j.biocon.2003.07.017
population inhabiting 3000–5000 km2 from Molise to
Marche and Umbria regions (Fig. 1). However, the most
densely and steadily occupied area is about 1500–2500
km2 wide, in the Apennine mountains of southern Ab-
ruzzo, Latium and Molise regions, surrounding theAbruzzo National Park (Meriggi et al., 2001). Minimum
bear population size, estimated by snow tracking in 1985
over 600 km2 (Abruzzo National Park+ 200 km2 of its
buffer area), was 38–39 bears (Boscagli, 1990). No
comprehensive estimates of population size have been
carried out, but a preliminary evaluation of average bear
density has been made by two DNA fingerprinting
studies through a part of the ‘‘core’’ distribution range.These suggest a population density of 1 bear/50–80 km2
(Lorenzini and Posillico, 2000; Forest Service, unpub-
lished data). At such densities, a small bear population
(e.g. 60 bears) might be distributed over a 3000–4800
km2 area. This fully protected, threatened bear

142 M. Posillico et al. / Biological Conservation 118 (2004) 141–150
population (Servheen, 1999) requires timely conserva-
tion strategies and a proactive management approach
(Peyton et al., 1999).
Active bear conservation is only scheduled within
protected areas and usually without common guidelines.Furthermore, no wide scale habitat management strat-
egies exist to accomplish bear conservation in central
Italy based on habitat–population relationships. The
available budget to conduct long-term research and time
are often limiting factors for bear conservation. Thus,
quantitative models could represent useful tools to as-
semble and synthesize existing knowledge to combine
research with conservation (Walters, 1986; Starfield,1997; Williams, 1997; Merrill et al., 1999; Gross and
Miller, 2001).
Our goal was to produce a model of bear–habitat
relationships to identify habitat variables (sensu Hall et
al., 1997) affecting bear presence at the home range level,
and to evaluate the effectiveness of the national and
regional networks of protected areas for bear conser-
vation. Brown bear occurrence through the eastern andnorthern parts of the former range is gradually in-
creasing in central Italy. Thus, our model could help to
predict the potential distribution of brown bears to
spread through their previous historical range, i.e. its
range during 16th–18th centuries, the earliest period for
which bear distribution is reasonably known (Febbo
and Pellegrini, 1990) and to identify critical areas for the
expansion of this population.
2. Methods
2.1. Study area
Our study area (8000 km2) was located in central
Italy, from Latium Apennines (west) to the Adriatic sea(east) (central coordinates: 41�510 North; 12�270 East)(Fig. 1). This area included the core bear range and
encompassed Latium, Abruzzo and Molise regions.
Land use was mainly deciduous forests (35%) and crops
(27%). Pastures (16%) and rocks (2%) were present
mainly on the ridges of the Apennine chain and olive-
tree groves, orchards and vineyards (4%) at the lowest
altitudes or along valley bottoms. Shrubland (11%)resulted mainly from re-colonisation of abandoned
pastures and cultivations and from forest degradation.
The climate ranges from Mediterranean type near the
sea coast to meso-Mediterranean and sub-Mediterra-
nean, with the increase of altitude. Urban areas and
villages (2%) were mainly located at low altitudes and
along valley bottoms. Lakes and rivers, wetlands and
perennial snowfields occupy the rest of the study area.These habitat features vary widely across the study
area, along north–south, east–west and altitudinal
gradients.
2.2. Habitat analysis
Using a Universal Transverse Mercator grid system
(UTM) we identified 320 sample squares of 25 km2 each
on a 5 km spaced grid. The size of the sample squareswas based on the average size of annual home ranges of
adult female brown bears. Because no data about home
range size of the Apennine bear population were avail-
able, we used data from Croatia where the estimated
female mean home range size was 107 km2 (SD ¼ 35.7;
range ¼ 78–147; N ¼ 3) (Huber and Roth, 1993). In
agreement with Laymon and Barrett (1986) and Schulz
and Joyce (1992), the sample square size we adopted wasroughly equal to 15–20% of the estimated home range.
For each sample square we measured 36 habitat vari-
ables of which 19 concerned land cover types, seven
landscape structure (McGarigal and Marks, 1995), five
human pressure, and five physical characteristics (Table
1). Percentage cover of each vegetation or landcover type
was measured from Corine land-cover III level vector
maps, issuedby theCentro Interregionale diCartografia –Ministry of the Environment. Corine land-covermaps are
based on a supervised, computer-aided classification of
Landsat Thematic Mapper imagery (pixel size
0.09 ha), providing a nationwide homogeneous landcover
database at a 1:100,000 scale. The interpretation
of Landsat imagery has been checked and enhanced us-
ing high resolution aerial images (1:70,000), allowing
a minimum mapping resolution finer than 6.3 ha (250�250m). Length of railways, sealed and unsealed roads and
trails were measured from vector data layers developed
from topographic maps (scale 1:100,000 and 1:25,000
respectively, Istituto Geografico Militare). We digitised
elevation contours and mapped elevation points (moun-
tain tops, springs, valley bottoms, etc.) from 1:25,000
topographic maps to build a digital terrain model (DTM)
in raster format. We calculated altitude variables (mean,maximum, minimum, range and standard deviation in m
a.s.l.) assigning an altitude value to a set of randompoints
(n ¼ 1000 per sample square) overlaid on the study area�sDTMand summarizing altitude values by sample square.
The Shannon–Wiener diversity index (HD)wasmeasured
as
HD ¼ �Xn
i¼1
ðPi � ln PiÞ;
where Pi is the proportion of the sample square occupied
by patch type i. All variables were measured by PatchAnalyst software (Elkie et al., 1999) and Spatial Analyst
(v. 2.0a) extension of Arcview� GIS (v. 3.2a, ESRI,
Redlands, CA, USA).
We determined the bear range within the study area
from literature and from data collected by several au-
thors (Russo, 1990; Boscagli et al., 1995; Posillico, 1996;
Posillico and Sammarone, 1997). Data on bear presence

Table 1
Average values (SE) of habitat variables with significant differences between presence and absence sample squares (one-way ANOVA)
Habitat variables Bear absence n ¼ 48 Bear presence n ¼ 42 F P
Sealed roads length 16.82 (1.37) 5.93 (0.93) 41.85 <0.0001
Unsealed roads length 13.18 (1.54) 3.72 (0.63) 30.27 <0.0001
Settlements 2.70 (0.43) 0.62 (0.18) 18.87 <0.0001
Crops 19.24 (3.39) 1.06 (0.61) 25.63 <0.0001
Vineyards and olive tree groves 6.15 (1.56) 0.16 (0.13) 13.50 <0.0001
Heterogeneous and patchy cultivated areas 20.53 (2.57) 2.11 (0.78) 43.68 <0.0001
Deciduous woodlands 22.79 (3.00) 55.41 (2.90) 60.61 <0.0001
Rocks 1.18 (0.81) 4.15 (1.14) 4.63 0.034
Steppes (high altitude discontinuous grassland) 1.71 (0.47) 5.31 (1.41) 6.24 0.014
Shannon–Wiener diversity index 1.53 (0.05) 1.14 (0.05) 31.51 <0.0001
Average altitude 704 (52.53) 1409 (37.98) 114.90 <0.0001
Altitude standard deviation 145.99 (10.43) 234.32 (13.91) 26.36 <0.0001
Minimum altitude 420 (36.34) 839 (53.49) 43.07 <0.0001
Maximum altitude 1095 (69.84) 1928 (39.96) 102.24 <0.0001
Altitude range 675 (53.95) 1089 (60.01) 26.56 <0.0001
Patch numbera 21.10 (0.97) 15.09 (1.05) 17.59 <0.0001
Mean patch size (ha)b 10.80 (1.77) 41.82 (4.70) 40.76 <0.0001
Ecotonal length (km)c 437.1 (33.23) 829.94 (52.37) 41.51 0.001
Mean size of woodland patches (ha)d 10.58 (19.0) 41.02 (29.1) 35.77 <0.0001
Number of woodland patchese 41 (2.48) 2.27 (1.53) 17.81 <0.0001
Woodland isolation index (km)f 668.27 (685.5) 161.52 (168.7) 22.76 <0.0001
a Total number of CORINE polygons/sample square.bMean size of CORINE polygons/sample square.c Sum of perimeter of CORINE polygons/sample square.dMean size of woodland polygons/sample square.e Total number of woodland polygons/sample square.fMean distance from woodland edge of 100 points/sample square. Distance for points inside wood patches has been set to 0.
M. Posillico et al. / Biological Conservation 118 (2004) 141–150 143
were collected monthly from 1989 to 2000 along trails
(900 km) by trained observers over 100 sample squares
(2500 km2). These represented 43% of the 231 sample
squares within the historical distribution range of the
brown bear in our study area and were located across
the Abruzzo National Park, Majella National Park,
Sirente-Velino Regional Park, Roveto valley, southern–
eastern–northern buffer of the Abruzzo National Park,and several State Forests and Reserves. Accordingly, we
were confident that the sampled area should be repre-
sentative of bear–habitat relationships in central Italy
within a broad spectrum of habitat conditions. Bear
occurrence information within the northern part of its
distribution range, outside the study area, were not al-
ways adequate in terms of sampling scheme and effort (if
any) to provide useful data. Sampling effort (total traillength/study area surface) was 0.36 km per km2, and was
greater than the ratio recommended by Kendall et al.
(1992) and Clevenger and Purroy (1996) to detect sig-
nificant changes in relative bear abundance (0.08–0.14
km per km2) across the whole bear distribution range.
Along the trails all signs of bear presence were recorded,
but we used only scats (>90% of all bear signs collected)
as an index of bear presence as the detection of scats isthe least influenced by habitat characteristics and their
attribution to bears is less subject to uncertainty. A
sample square was classified as occupied only if bear
presence (i.e. at least 1 scat/year) was confirmed for a
period P5 years. The minimum number of scats re-
corded within sample squares in which bears occurred
was 15 for a P5 year period.
2.3. Data analysis
According to our field sampling, we found evidence
of long-term bear presence in 44 sample squares whereasabsence was documented for 56 out of 100 sample
squares. However, within eight sample squares (out of
the 56 within which bear has been classified as absent),
shepherds, hikers, mushroom and berry pickers reported
bear occurrence throughout the sampling period (<3
times/square). As field data collection and ad hoc sur-
veys did not validate such reports, nor we could be
confident about the reliability of reporting people, wediscarded these eight squares from the analyses. The
remaining 92 squares were used to formulate a predic-
tive model of bear habitat suitability that was then ap-
plied to the whole study area and to the historical bear
range (Fig. 1).
Wildlife–habitat relationships are multidimensional
in nature, so we used multivariate analytical techniques
to describe habitat–bear population relationships and topredict habitat suitability for bears (Morrison et al.,
1992; Clark et al., 1993). We used multivariate analysis
of variance (MANOVA) to test the global differences of
the average values of habitat variables between sample

Fig. 1. Historical distribution range (grey polygon, after Boscagli, 1999; Febbo and Pellegrini, 1990) and present range (dark grey circles, after
Meriggi et al., 2001) of the brown bear in central Italy, showing boundaries of the study area (square grid) and main administrative regions. Each
black circle outside the study area often represents one sign of presence between 1990 and 2000. The frequency of these signs should not be read as an
index of local bear abundance, as many of them are likely to refer to the same bear.
144 M. Posillico et al. / Biological Conservation 118 (2004) 141–150
squares where bears were present or not, and one-way
ANOVA to identify the most important variables that
contributed to the differences. As the dependent variable
(bear presence/absence) was binomial, we chose di-
chotomous logistic regression analysis (DLgR) to assessthe probability of bear occurrence in the sample squares
(Hosmer and Lemeshow, 1989; Trexler and Travis,
1993; Massolo and Meriggi, 1998). This technique is
appropriate for non-normally distributed data and it
provides results which are statistically robust to viola-
tion of assumptions of parametric statistics (Trexler and
Travis, 1993; Brito et al., 1999). Equation parameters
were estimated by a stepwise forward selection proce-dure (maximum likelihood method) to reduce the
number of independent variables actually entered into
the model. The goodness-of-fit with all variables was
tested by the model chi-square test, while the improve-
ment test was used to compare the change of log-like-
lihood between successive steps in model building.
Nagelkerke�s R2 was interpreted as the proportion of
explained variance of the dependent variable (Norusis,1992). We used the Wald statistics to test the null hy-
pothesis that regression coefficients were equal to 0 and
R statistic to estimate partial correlations between the
dependent variable and each independent variable en-
tered into the model. Hosmer and Lemeshow (1989)
goodness-of-fit statistics was used to test for differences
between expected and observed values of the predicted
probability. We performed a curve-fit linear regression
analysis to assess the influence of entered independent
variables on the probability of bear presence predicted
by the DLgR (Pompilio and Meriggi, 2001).
Model testing and validation were performed on thebasis of two subsequent steps: (1) the percentage of the
cases correctly classified by the model in the two groups
was used to evaluate the capability of the model to de-
scribe the initial data; (2) predicted probabilities of bear
occurrence within sample squares that were not included
in the formulation of the logistic regression model were
compared with the bear distribution resulting from lit-
erature and agency information (n ¼ 67 sample squareswithin the study area). Although not always collected
according to our sampling scheme, these are the only
other reliable and available data on bear occurrence.
This allowed us to assess the ability of the model to
describe independent data.
All statistical analyses were performed with the SPSS
PCþ 10.0 package (SPSS Inc., Chicago, IL, USA).
2.4. Mapping the habitat model
To be a useful tool, the model output should allow a
ready identification of potential bear habitat and of
areas relevant to conservation and management. How-
ever, direct transformation of probability scores as-
signed to each sample square into a bear–habitat

M. Posillico et al. / Biological Conservation 118 (2004) 141–150 145
management map is unrealistic. First, the probability
score – and consequently management prescriptions –
assigned to every point within a sample square is influ-
enced by the position of the sampling grid used to
generate the model. The use of another – shifted – UTMsampling grid to calculate the probability of bear pres-
ence could produce different values for the same point,
with management options and priorities changing ac-
cordingly. Second, the model does not account for the
effect of adjacent probability scores on the value as-
signed to a given sample square or point. Thus, we built
15 additional sample square grids that were shifted 1.25
and 2.5 km east, west, north and south of the originalgrid. For the resulting 5120 sample squares (16 total
grids� 320 cells each) we ran a dichotomous logistic
regression model incorporating only the variables en-
tered in the original habitat model. The spatial inter-
section of the overlapping 16 grids produced 1.56 km2
pixels (1.25 km side), which was the mapping resolution
of our model. This is consistent with brown bear home
range size and with the spatial resolution of the Corinelandcover maps. The probability value assigned to each
pixel equalled the sum of the probability scores of the 16
overlapping squares divided by the number of overlap-
ping grids.
3. Results
We found significant broad differences between the
average values of habitat variables of squares with bear
presence and absence (Wilks� Lambda ¼ 0.2, d.f. ¼ 35,
P < 0:001). Twenty-one habitat variables significantly
contributed to these broad differences (Table 1). Vari-
ables related to human pressure were significantly lower
in the sample squares of bear presence, with the excep-
tion of railroad and trail length. Bear presence wassignificantly and positively associated with the extension
of deciduous woodlands and negatively associated to
woodland isolation index and woodland patch number
(Table 1). Crops, vineyard-olive groves, and heteroge-
neous cultivated areas showed average values signifi-
cantly lower in sample squares where bears were present.
The presence of the brown bear was significantly related
to rocks and steppe (i.e. high altitude discontinuous
Table 2
Parameter estimate of the logistic regression model of brown bear presence–
Habitat variables B SE
Vineyard-olive tree groves )0.40 0.32
Deciduous woodland 0.042 0.02
Shrubland )0.22 0.09
Average altitude 0.07 0.001
Ecotonal length 0.03 0.001
Constant 9.81 2.50
Model v2 ¼ 97:99, d.f. ¼ 5, P < 0:001; )2 log likelihood ¼ 29.37.
grassland), to a low number of habitat patches and to a
low landscape diversity (Table 1).
A logistic regression analysis performed on the 92
sample squares correctly classified 95.5% of presence
squares and 93.8% of absence ones. Five habitat vari-ables entered the model, of which average altitude,
shrubland and deciduous woodland had significant re-
gression coefficients and made the greatest contribution
(Table 2). The probability of bear presence increased as
altitude and percentage of deciduous woods increased,
and percentage of shrubland decreased. As evidenced by
curve-fit analyses, the mean altitude of most sample
squares with predicted occurrence values >0.75 is >1100m a.s.l., while most sample squares above 900 m a.s.l.
had probability values <0.25, with a probability thresh-
old around 1000 m. A similar pattern was observed for
the extension of deciduous woodlands, with a threshold
around 50%. The model explained 87.4% of the variance
of the dependent variable (Nagelkerke�s R2) with no
significant differences between observed and expected
probability values of bear presence (Hosmer–Lemeshowv2 ¼ 5:08; P ¼ 0:75).
The validation procedure across 67 sample squares
(for which independent data on bear presence were
available) showed that the model was able to correctly
classify 93% and 87% of absence and presence squares,
respectively. Out of the residual 72 sample squares
within which bear occurrence was uncertain (8) or not
investigated (64), 40% were classified within the presencegroup.
Within the whole study area, our model classified
39% of the sample squares (2800 km2) as >0.5 proba-
bility of bear presence and 61% (5200 km2) as <0.5.
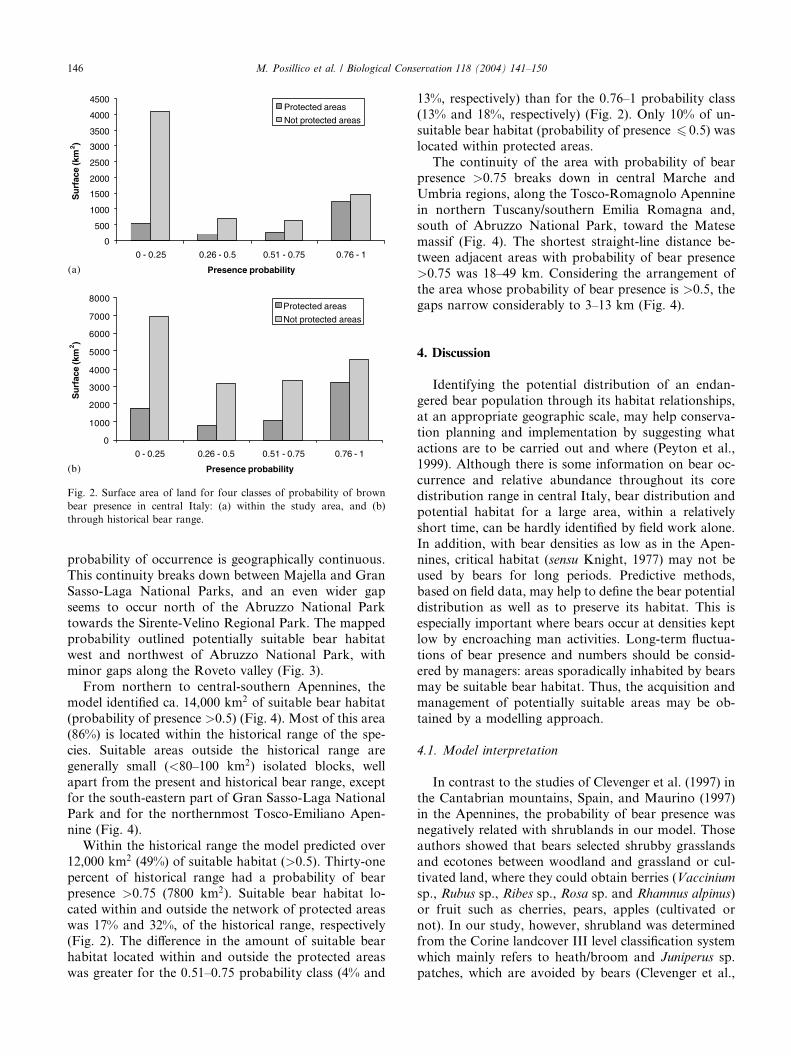
Moreover, 71% of the squares had probability values
6 0.25 or >0.75 (Fig. 2). Most of the sample squares
with a predicted probability of bear presence 60.25
(52% of the study area) were located outside the pro-tected area network (i.e. National and Regional Parks,
State Forests and Reserves). The areas with a predicted
probability of bear occurrence >0.50 were not equally
distributed between protected and non-protected areas,
but accounted for 16% and 23% of the total area, re-
spectively (Fig. 2).
With respect to the present bear distribution and the
arrangement of protected land, the area with a high
absence vs. habitat variables in central Italy
Wald P R
1.5 0.22 0.0
3.4 0.06 0.1
5.7 0.02 )0.212.5 <0.001 0.3
3.8 0.05 0.01
15.4 <0.001
0
500
1000
1500
2000
2500
3000
3500
4000
4500
0 - 0.25 0.26 - 0.5 0.51 - 0.75 0.76 - 1
Presence probability
Su
rfac
e (k
m2 )
Protected areas
Not protected areas
0
1000
2000
3000
4000
5000
6000
7000
8000
0 - 0.25 0.26 - 0.5 0.51 - 0.75 0.76 - 1
Presence probability
Su
rfac
e (k
m2 )
Protected areas
Not protected areas
(a)
(b)
Fig. 2. Surface area of land for four classes of probability of brown
bear presence in central Italy: (a) within the study area, and (b)
through historical bear range.
146 M. Posillico et al. / Biological Conservation 118 (2004) 141–150
probability of occurrence is geographically continuous.
This continuity breaks down between Majella and Gran
Sasso-Laga National Parks, and an even wider gap
seems to occur north of the Abruzzo National Park
towards the Sirente-Velino Regional Park. The mapped
probability outlined potentially suitable bear habitat
west and northwest of Abruzzo National Park, withminor gaps along the Roveto valley (Fig. 3).
From northern to central-southern Apennines, the
model identified ca. 14,000 km2 of suitable bear habitat
(probability of presence >0.5) (Fig. 4). Most of this area
(86%) is located within the historical range of the spe-
cies. Suitable areas outside the historical range are
generally small (<80–100 km2) isolated blocks, well
apart from the present and historical bear range, exceptfor the south-eastern part of Gran Sasso-Laga National
Park and for the northernmost Tosco-Emiliano Apen-
nine (Fig. 4).
Within the historical range the model predicted over
12,000 km2 (49%) of suitable habitat (>0.5). Thirty-one
percent of historical range had a probability of bear
presence >0.75 (7800 km2). Suitable bear habitat lo-
cated within and outside the network of protected areaswas 17% and 32%, of the historical range, respectively
(Fig. 2). The difference in the amount of suitable bear
habitat located within and outside the protected areas
was greater for the 0.51–0.75 probability class (4% and
13%, respectively) than for the 0.76–1 probability class
(13% and 18%, respectively) (Fig. 2). Only 10% of un-
suitable bear habitat (probability of presence 6 0.5) was
located within protected areas.
The continuity of the area with probability of bearpresence >0.75 breaks down in central Marche and
Umbria regions, along the Tosco-Romagnolo Apennine
in northern Tuscany/southern Emilia Romagna and,
south of Abruzzo National Park, toward the Matese
massif (Fig. 4). The shortest straight-line distance be-
tween adjacent areas with probability of bear presence
>0.75 was 18–49 km. Considering the arrangement of
the area whose probability of bear presence is >0.5, thegaps narrow considerably to 3–13 km (Fig. 4).
4. Discussion
Identifying the potential distribution of an endan-
gered bear population through its habitat relationships,
at an appropriate geographic scale, may help conserva-tion planning and implementation by suggesting what
actions are to be carried out and where (Peyton et al.,
1999). Although there is some information on bear oc-
currence and relative abundance throughout its core
distribution range in central Italy, bear distribution and
potential habitat for a large area, within a relatively
short time, can be hardly identified by field work alone.
In addition, with bear densities as low as in the Apen-nines, critical habitat (sensu Knight, 1977) may not be
used by bears for long periods. Predictive methods,
based on field data, may help to define the bear potential
distribution as well as to preserve its habitat. This is
especially important where bears occur at densities kept
low by encroaching man activities. Long-term fluctua-
tions of bear presence and numbers should be consid-
ered by managers: areas sporadically inhabited by bearsmay be suitable bear habitat. Thus, the acquisition and
management of potentially suitable areas may be ob-
tained by a modelling approach.
4.1. Model interpretation
In contrast to the studies of Clevenger et al. (1997) in
the Cantabrian mountains, Spain, and Maurino (1997)in the Apennines, the probability of bear presence was
negatively related with shrublands in our model. Those
authors showed that bears selected shrubby grasslands
and ecotones between woodland and grassland or cul-
tivated land, where they could obtain berries (Vaccinium
sp., Rubus sp., Ribes sp., Rosa sp. and Rhamnus alpinus)
or fruit such as cherries, pears, apples (cultivated or
not). In our study, however, shrubland was determinedfrom the Corine landcover III level classification system
which mainly refers to heath/broom and Juniperus sp.
patches, which are avoided by bears (Clevenger et al.,

Fig. 3. Potential brown bear distribution in the study area. Habitat model sample square size: 25 km2; size of probability scores� pixels: 1.56 km2.
National and Regional Parks area shown as dotted polygons; thick-solid lines represent boundaries of main administrative regions.
M. Posillico et al. / Biological Conservation 118 (2004) 141–150 147
1997; Maurino, 1997). Ecotonal length, selected by the
dichotomous logistic regression model, may better rep-
resent the ecotone shrubland described by Maurino
(1997).
Beech (Fagus sylvatica) and oak (Quercus cerris)
woodlands, as well as their ecotonal interface, providemost of the forest-related bear foods (Zunino and Her-
rero, 1972; Fabbri, 1987; Klahre and Roth, 1996; Lovari
et al., 1996). The distribution of these forest types starts
from 800–1000 m to timberline (1800–1900 m). Al-
though other acorn-producing oak woodland exists at
lower altitudes, we suggest that bear occurrence there is
negatively influenced by the reduced total cover of
woodland, by its increased isolation and because of theincreased human presence, which raises disturbance,
decreases habitat suitability and increases vulnerability
to illegal hunting (Elgmork, 1978; McLellan and
Shackleton, 1989; Mace and Waller, 1996).
These results suggest that, at an altitude of �800–
1000 m, the probability of bear presence could be neg-
atively affected by both the higher levels of human
presence and by the related landscape modifications,affecting woodland total cover and spatial structure.
Some high altitude sample squares had a high pre-
dicted probability of bear occurrence despite the absence
of forest cover. This could be explained by the remote-
ness of such areas with respect to human disturbance
and by the active search for locally abundant Rhamnus
alpinus patches and late-season grasses. Moreover, many
sheep flocks are herded onto such high altitude pastures,
representing one of the main sources of animal proteinfor bears (Lovari et al., 1996). Even if guarded by
shepherds and dogs, sheep are taken almost only from
the enclosures where they are kept at night. Regional
authorities compensate for the damages to livestock by
bear, wolf and dog, but bears are only responsible for
4.8% of livestock depredation (Cozza et al., 1996) and
they are seldom killed (illegally) to protect livestock.
4.2. Implications for conservation
Our model showed a good overlap between the sys-
tem of protected areas and habitat suitability, as well as
present bear distribution, in the central Apennines.
However, suitable landscape extends far outside pro-
tected areas. Resident bears regularly occur in a large
contiguous block of potential bear habitat, extendingP 200 km west and northwest of the Abruzzo National
Park (Simbruini Regional Park, unpublished data;

Fig. 4. Potential distribution of the brown bear through its historical range and across central-southern Apennines.
148 M. Posillico et al. / Biological Conservation 118 (2004) 141–150
Zompo Lo Schioppo Nature Reserve, unpublished
data). The role of this area for the expansion of the bear
population is likely important. Thus, we recommend
that particular attention should be paid locally to con-
servation-oriented actions for bears and to habitatmanagement of eastern and northern parts of the pres-
ent bear range, from Simbruini Regional Park to Gran
Sasso-Laga and Sibillini National Parks. We suggest a
careful and timely planning of conservation actions
there. Reliable estimates of bear occurrence and the
enforcement of habitat conservation measures, as well as
the environmental impact evaluation of land manage-
ment, will help bear recovery and persistence in thislarge, contiguous block of potential habitat. Given the
currently small population size, a sound management of
potential bear range is important for long-term brown
bear survival in the Apennines.
Conservation actions should be devised and coordi-
nated on a wide geographic scale suitable for bears
(Schoen, 1990). No effective bear conservation policy
may be implemented without the cooperation of theagencies responsible for land management. Some hu-
man-related factors (land use planning, habitat loss and
fragmentation, hunting, poaching, changes in regula-
tions, etc.) are also important but could not be included
in our model. A coordinated habitat policy with bio-
logically sound aims and clear objectives is essential for
long-term bear conservation. This also requires an as-
sessment of socio-economic implications related to
habitat preservation.
The need for a biologically-based regulation to assessand limit the impact of natural resource exploitation on
bear occurrence and bear habitat has been recently re-
cognised as a priority task by several agencies (parks,
ministries, forest service, NGOs). These also asked for
the establishment of a technical committee, on behalf of
the Ministry of the Environment, to formulate a bear
conservation plan and to identify urgent conservation
actions. Therefore, our model for potential bear occur-rence may help to proactively manage habitat for bears,
providing the area within which the impact of harmful
activities to bear habitat should be assessed on a multi-
level management scale. The impact of all detrimen-
tal human activities should be critically considered
for sample squares having a >0.5 probability of bear
presence and for developing an environmental impact
assessment procedure. We recommend that the envi-ronmental impact assessment should explicitly consider
the existence and status of resources important to bears
(food, cover, refuge, microclimate) as well as human
impact and habitat loss and fragmentation (at local
and regional scales). In this way, suitable areas will be

M. Posillico et al. / Biological Conservation 118 (2004) 141–150 149
preserved and population expansion and recovery of
bear could occur.
Acknowledgements
Our work has been endorsed and/or funded by the
Italian Ministry of the Environment (Nature Conser-
vation Direction) and by the Italian Ministry of Forests
and Agriculture (through a European Union grant,
LIFE99NAT/IT/006244). We thank P. Verucci, A. Ca-
rucci, B. Ragni, M. Spinetti and M. Dell�Orso for data
on recent bear presence within Latium, Umbria, Ab-ruzzo and Marche Regions. E. Dupr�e provided useful
suggestions as to GIS processing. A. Massolo provided
useful suggestions in statistics and habitat modelling.
We thank A. Cumer and I. Chiucchiarelli, who provided
Corine coverages and technical insight about Corine
landcover maps. We are particularly grateful to F. Van
Manen, N. Owen-Smith, F. Cassola and A. Clevenger
for revising our paper, their stimulating criticism as wellas for providing significant improvements.
References
Boscagli, G., 1990. Marsican brown bear population in Central Italy –
status report. Aquilo Series in Zoology 27, 81–83.
Boscagli, G., 1999. Status and management of the brown bear in
Central Italy (Abruzzo). In: Servheen, C., Herrero, S., Peyton, B.
(Eds.), Bears. Status Survey and Conservation Action Plan. IUCN/
SSC Bear and Polar Bear Specialist Group. IUCN, Gland
Switzerland, pp. 81–84.
Boscagli, G., Pellegrini, M., Febbo, D., Pellegrini, M., Cal�o, C.M.,
Castellucci, C., 1995. Distribuzione storica recente (1900–1991)
dell�orso bruno marsicano all�esterno del Parco Nazionale d�Ab-
ruzzo. Atti Societ�a Italiana di Scienze Naturali, Museo Civico di
Storia Naturale di Milano 134/1993, pp. 46–84.
Brito, J.C., Crespo, E.G., Paulo, O.S., 1999. Modelling wildlife
distributions: logistic multiple regression vs. overlap analysis.
Ecography 22, 251–260.
Clark, J.D., Dunn, J.E., Smith, K.G., 1993. A multivariate model of
black bear habitat use for a geographic information system.
Journal of Wildlife Management 57, 519–526.
Clevenger, A.P., Purroy, F.J., 1996. Sign surveys for estimating trend
of a remnant brown bear population in northern Spain. Wildlife
Biology 2, 275–281.
Clevenger, A.P., Purroy, F.J., Campos, M.A., 1997. Habitat assess-
ment of a relict brown bear population in Northern Spain.
Biological Conservation 80, 17–22.
Cozza, K., Fico, R., Battistini, M.L., Rogers, E., 1996. The damage–
conservation interface illustrated by predation on domestic live-
stock in central Italy. Biological Conservation 78, 329–336.
Elgmork, K., 1978. Human impact on a brown bear population.
Biological Conservation 13, 81–103.
Elkie, P.C., Rempel, R.S., Carr, A.P., 1999. Patch Analyst user�smanual. A tool for quantifying landscape structure. Northwest
Science & Technology, NWST Technical Manual TM-002. Ontario
Ministry for Natural Resources. Thunder Bay, Ontario, Canada.
Fabbri, M., 1987. Le abitudini alimentari dell�orso bruno nel Parco
Nazionale d�Abruzzo. M. Sc. (Laurea) Thesis, University of Parma,
Parma, Italy.
Febbo, D., Pellegrini, M., 1990. The historical presence of the
brown bear in the Apennines. Aquilo Series in Zoology 27,
85–88.
Gross, J.E., Miller, M.W., 2001. Chronic wasting disease in mule deer:
disease dynamics and control. Journal of Wildlife Management 65,
205–215.
Hall, L.S., Krausman, P.R., Morrison, M.L., 1997. The habitat
concept and a plea for standard terminology. Wildlife Society
Bulletin 25, 173–182.
Hosmer Jr., D.W., Lemeshow, S., 1989. Applied Logistic regression.
John Wiley and Sons, New York.
Huber, D., Roth, H.U., 1993. Movements of European brown bears in
Croatia. Acta Theriologica 38, 151–159.
Kendall, K.C., Metzgar, L.H., Patterson, D.A., Steele, B.M., 1992.
Power of sign surveys to monitor population trends. Ecological
Applications 2, 422–430.
Klahre, J., Roth, H.U., 1996. Nutritional preferences of Abruzzo
brown bears before hibernation. Abstracts. First Symposium of the
Italian Theriological Association, Perugia, October 1996.
Knight, R.R., 1977. Biological considerations in the delineation of
critical habitat. International Conference on Bear Research and
Management 3, 1–3.
Laymon, S.A., Barrett, R.H., 1986. Developing and testing habitat-
capability models: pitfalls and recommendations. In: Verner, J.,
Morrison, M.L., Ralph, C.J. (Eds.), Wildlife 2000. Modelling
Habitat Relationships of Terrestrial Vertebrates. University of
Wisconsin Press, Madison, WI, pp. 87–92.
Lorenzini, R., Posillico, M., 2000. DNA fingerprinting and brown bear
conservation in Abruzzo – central Italy. Italian Journal of Zoology
67, 326–327.
Lovari, S., Posillico, M., Varriale, M., Burrini, L., 1996. L�orso bruno
negli Altopiani maggiori d�Abruzzo. Final report to the Ministry of
Agriculture and Forests, Rome, Italy.
Mace, R.D., Waller, J.S., 1996. Grizzly bear distribution and human
conflicts in Jewel Basin Hiking Area, Swan Mountains, Montana.
Wildlife Society Bulletin 24, 461–467.
Massolo, A., Meriggi, A., 1998. Factors affecting habitat occupancy by
wolves in the northern Apennines (Northern Italy): a model of
habitat suitability. Ecography 21, 97–107.
Maurino, L., 1997. Uso dello habitat nell�orso bruno. M.Sc. (Laurea)
Thesis, University of Siena, Siena, Italy.
McGarigal, K., Marks B., 1995. FRAGSTATS: spatial pattern
analysis program for quantifying landscape structure. Gen Tech.
Rep. PNW-GTR-351. US Department of Agriculture, Forest
Service, Pacific Northwest Research Station, Portland, OR,
USA.
McLellan, B.N., Shackleton, D.M., 1989. Immediate reaction of
grizzly bears to human activities. Wildlife Society Bulletin 17, 269–
274.
Meriggi, A., Sacchi, O., Ziliani, U., Posillico, M., 2001. Definizione
dell�areale potenziale di cervo sardo, muflone e orso bruno. In:
Lovari, S. (Ed.), Progetto di monitoraggio dello stato di conserv-
azione di alcuni mammiferi particolarmente a rischio della fauna
italiana. Ministry of the Environment, Department for Nature
Conservation, Rome, Italy.
Merrill, T., Mattson, D.J., Wright, G.R., Quigley, H.B., 1999.
Defining landscapes suitable for the restoration of grizzly bears in
Idaho. Biological Conservation 87, 231–248.
Morrison, M.L., Marcot, G.B., Mannan, R.W., 1992. Wildlife–
Habitat Relationships: Concepts and Applications. University of
Wisconsin Press, Madison, WI.
Norusis M.J., 1992. SPSS/PC+ Advanced statistics, Ver. 5.0, Chicago.
Peyton, B., Servheen C., Herrero, S., 1999. An overview of bear
conservation planning and implementation. In: Servheen, C.,
Herrero, S., Peyton, B. (Eds.), Bears. Status Survey and Conser-
vation Action Plan. IUCN/SSC Bear and Polar Bear Specialist
Group, IUCN, Gland Switzerland, pp. 8–24.

150 M. Posillico et al. / Biological Conservation 118 (2004) 141–150
Pompilio, L.,Meriggi, A., 2001.Modelling wild ungulate distribution in
Alpine habitat: a case study. Italian Journal of Zoology 68, 281–289.
Posillico, M., 1996. Brown bear presence in State Forests and
neighbour areas in Central Italy. Journal of Wildlife Research 1,
250–252.
Posillico, M., Sammarone, L., 1997. Relazione conclusiva del progetto
LIFE per la conservazione dell�orso bruno 1993–1997. Ministry of
Agriculture and Forests, State Forest Service, Rome, Italy.
Russo, L., 1990. L�orso bruno marsicano: dati preliminari dall�analisidelle schede faunistiche del Parco Nazionale d�Abruzzo. Bollettino
della Societ�a dei Naturalisti in Napoli 98–99, 107–122.
Servheen, C., 1999. Conservation of small bear populations through
strategic planning. Ursus 11, 67–74.
Schoen, J.W., 1990. Bear habitat management: a review and future
perspectives. International Conference on Bear Research and
Management 8, 143–154.
Schulz, T.T., Joyce, L.A., 1992. A spatial application of a marten
habitat model. Wildlife Society Bulletin 20, 74–83.
Starfield, A.M., 1997. A pragmatic approach to modelling for
wildlife management. Journal of Wildlife Management 61, 261–
270.
Swenson, J.E., Gerstl, N., Dahle, B., Zedrosser, A., 2000. Action Plan
for the Conservation of the Brown Bear in Europe. WWF
International, Gland, Switzerland.
Trexler, J.C., Travis, J., 1993. Non traditional regression analyses.
Ecology 74, 1629–1637.
Walters, C., 1986. Adaptive management of renewable resources.
Macmillan, New York.
Williams, B.K., 1997. Logic and science in wildlife biology. Journal of
Wildlife Management 61, 1007–1015.
Zunino, F., Herrero, S., 1972. The status of the brown bear in Abruzzo
National Park, 1971. Biological Conservation 4, 263–272.




















