A genetic variation located in the promoter region of the UPAR (CD87) gene is associated with the...
10
ARTHRITIS & RHEUMATISM Vol. 63, No. 1, January 2011, pp 247–256 DOI 10.1002/art.30101 © 2011, American College of Rheumatology A Genetic Variation Located in the Promoter Region of the UPAR (CD87) Gene Is Associated With the Vascular Complications of Systemic Sclerosis Mirko Manetti, 1 Yannick Allanore, 2 Lucile Revillod, 2 Cinzia Fatini, 3 Serena Guiducci, 1 Giovanna Cuomo, 4 Claudia Bonino, 5 Valeria Riccieri, 6 Laura Bazzichi, 7 Vasiliki Liakouli, 8 Paola Cipriani, 8 Roberto Giacomelli, 8 Rosanna Abbate, 3 Stefano Bombardieri, 7 Guido Valesini, 6 Carlomaurizio Montecucco, 5 Gabriele Valentini, 4 Lidia Ibba-Manneschi, 3 and Marco Matucci-Cerinic 1 Objective. The UPAR gene encodes a pleiotropic receptor (urokinase-type plasminogen activator recep- tor [uPAR]) involved in fibrosis, immunity, angiogene- sis, and vascular remodeling. Previous studies have implicated uPAR in systemic sclerosis (SSc) vasculopa- thy and impaired angiogenesis. We undertook this study to investigate whether UPAR gene promoter polymor- phisms might be associated with SSc phenotypes in the European Caucasian population. Methods. We studied a total population of 1,339 individuals. The Italian discovery cohort comprised 388 SSc patients and 391 healthy controls. The French replication cohort consisted of 344 SSc patients and 216 healthy controls. The UPAR rs344781 and rs4251805 single-nucleotide polymorphisms were genotyped by polymerase chain reaction–restriction fragment length polymorphism assay. Results. In the Italian cohort, the rs344781 G allele was associated with SSc-related digital ulceration (odds ratio [OR] 1.39), pulmonary arterial hyperten- sion (PAH) (OR 1.81), anticentromere antibody (ACA) positivity (OR 1.45), and limited cutaneous SSc (lcSSc) (OR 1.37). The rs344781 GG genotype was associated with SSc-related (OR 3.79), ACA-positive SSc (OR 2.17), and lcSSc (OR 1.96). Allelic and genotypic asso- ciations with SSc-related digital ulceration and ACA- positive SSc were replicated in the French sample. Combined analyses showed an association of the rs344781 G allele and GG genotype with SSc-related digital ulceration (allele OR 1.41, genotype OR 2.15), SSc-related PAH (allele OR 1.65, genotype OR 3.16), ACA-positive SSc (allele OR 1.47, genotype OR 2.40), and lcSSc (allele OR 1.34, genotype OR 1.77). In a multivariate logistic regression analysis model including the above associated phenotypes of SSc pa- tients, the rs344781 GG genotype remained an indepen- dent risk factor for SSc-related digital ulceration (OR 1.96) and SSc-related PAH (OR 2.68). Conclusion. The UPAR rs344781 gene variant is associated with the SSc vascular phenotype. Systemic sclerosis (SSc; scleroderma) is charac- terized by early microangiopathy, defective angiogene- sis, immunologic abnormalities, and progressive fibrosis Supported by grants from the Ministero dell’Istruzione, dell’Universita ` e della Ricerca, the University of Florence (to Drs. Ibba-Manneschi and Matucci-Cerinic), and the Associazione per lo studio della Sclerosi Sistemica e delle Malattie Fibrosanti. Dr. All- anore and Ms Revillod’s work was supported by INSERM and the Agence Nationale pour la Recherche (grant R07094KS). 1 Mirko Manetti, PhD, Serena Guiducci, MD, Marco Matucci- Cerinic, MD, PhD: University of Florence and Excellence Centre for Research, Transfer and High Education on Chronic, Inflammatory, Degenerative and Neoplastic Disorders for the Development of Novel Therapies, Florence, Italy; 2 Yannick Allanore, MD, PhD, Lucile Revillod: Universite ´ Paris Descartes, INSERM U781, Ho ˆpital Necker, and Ho ˆpital Cochin, Paris, France; 3 Cinzia Fatini, MD, PhD, Rosanna Abbate, MD, Lidia Ibba-Manneschi, MD: University of Florence, Florence, Italy; 4 Giovanna Cuomo, MD, Gabriele Valentini, MD: University of Naples, Naples, Italy; 5 Claudia Bonino, MD, Carlomau- rizio Montecucco, MD: IRCCS Policlinico San Matteo and University of Pavia, Pavia, Italy; 6 Valeria Riccieri, MD, Guido Valesini, MD: University of Rome La Sapienza, Rome, Italy; 7 Laura Bazzichi, MD, Stefano Bombardieri, MD: University of Pisa, Pisa, Italy; 8 Vasiliki Liakouli, MD, Paola Cipriani, MD, Roberto Giacomelli, MD, PhD: University of L’Aquila, L’Aquila, Italy. Address correspondence to Mirko Manetti, PhD, Department of Anatomy, Histology and Forensic Medicine, University of Florence, Viale G. B. Morgagni 85, I-50134 Florence, Italy. E-mail: [email protected]. Submitted for publication May 7, 2010; accepted in revised form October 12, 2010. 247
Transcript of A genetic variation located in the promoter region of the UPAR (CD87) gene is associated with the...

ARTHRITIS & RHEUMATISMVol. 63, No. 1, January 2011, pp 247–256DOI 10.1002/art.30101© 2011, American College of Rheumatology
A Genetic Variation Located in the Promoter Region ofthe UPAR (CD87) Gene Is Associated With
the Vascular Complications of Systemic Sclerosis
Mirko Manetti,1 Yannick Allanore,2 Lucile Revillod,2 Cinzia Fatini,3 Serena Guiducci,1
Giovanna Cuomo,4 Claudia Bonino,5 Valeria Riccieri,6 Laura Bazzichi,7 Vasiliki Liakouli,8
Paola Cipriani,8 Roberto Giacomelli,8 Rosanna Abbate,3 Stefano Bombardieri,7
Guido Valesini,6 Carlomaurizio Montecucco,5 Gabriele Valentini,4 Lidia Ibba-Manneschi,3
and Marco Matucci-Cerinic1
Objective. The UPAR gene encodes a pleiotropicreceptor (urokinase-type plasminogen activator recep-tor [uPAR]) involved in fibrosis, immunity, angiogene-sis, and vascular remodeling. Previous studies haveimplicated uPAR in systemic sclerosis (SSc) vasculopa-thy and impaired angiogenesis. We undertook this studyto investigate whether UPAR gene promoter polymor-phisms might be associated with SSc phenotypes in theEuropean Caucasian population.
Methods. We studied a total population of 1,339individuals. The Italian discovery cohort comprised 388
SSc patients and 391 healthy controls. The Frenchreplication cohort consisted of 344 SSc patients and 216healthy controls. The UPAR rs344781 and rs4251805single-nucleotide polymorphisms were genotyped bypolymerase chain reaction–restriction fragment lengthpolymorphism assay.
Results. In the Italian cohort, the rs344781 Gallele was associated with SSc-related digital ulceration(odds ratio [OR] 1.39), pulmonary arterial hyperten-sion (PAH) (OR 1.81), anticentromere antibody (ACA)positivity (OR 1.45), and limited cutaneous SSc (lcSSc)(OR 1.37). The rs344781 GG genotype was associatedwith SSc-related (OR 3.79), ACA-positive SSc (OR2.17), and lcSSc (OR 1.96). Allelic and genotypic asso-ciations with SSc-related digital ulceration and ACA-positive SSc were replicated in the French sample.Combined analyses showed an association of thers344781 G allele and GG genotype with SSc-relateddigital ulceration (allele OR 1.41, genotype OR2.15), SSc-related PAH (allele OR 1.65, genotypeOR 3.16), ACA-positive SSc (allele OR 1.47, genotypeOR 2.40), and lcSSc (allele OR 1.34, genotype OR 1.77).In a multivariate logistic regression analysis modelincluding the above associated phenotypes of SSc pa-tients, the rs344781 GG genotype remained an indepen-dent risk factor for SSc-related digital ulceration(OR 1.96) and SSc-related PAH (OR 2.68).
Conclusion. The UPAR rs344781 gene variant isassociated with the SSc vascular phenotype.
Systemic sclerosis (SSc; scleroderma) is charac-terized by early microangiopathy, defective angiogene-sis, immunologic abnormalities, and progressive fibrosis
Supported by grants from the Ministero dell’Istruzione,dell’Universita e della Ricerca, the University of Florence (to Drs.Ibba-Manneschi and Matucci-Cerinic), and the Associazione per lostudio della Sclerosi Sistemica e delle Malattie Fibrosanti. Dr. All-anore and Ms Revillod’s work was supported by INSERM and theAgence Nationale pour la Recherche (grant R07094KS).
1Mirko Manetti, PhD, Serena Guiducci, MD, Marco Matucci-Cerinic, MD, PhD: University of Florence and Excellence Centre forResearch, Transfer and High Education on Chronic, Inflammatory,Degenerative and Neoplastic Disorders for the Development of NovelTherapies, Florence, Italy; 2Yannick Allanore, MD, PhD, LucileRevillod: Universite Paris Descartes, INSERM U781, Hopital Necker,and Hopital Cochin, Paris, France; 3Cinzia Fatini, MD, PhD, RosannaAbbate, MD, Lidia Ibba-Manneschi, MD: University of Florence,Florence, Italy; 4Giovanna Cuomo, MD, Gabriele Valentini, MD:University of Naples, Naples, Italy; 5Claudia Bonino, MD, Carlomau-rizio Montecucco, MD: IRCCS Policlinico San Matteo and Universityof Pavia, Pavia, Italy; 6Valeria Riccieri, MD, Guido Valesini, MD:University of Rome La Sapienza, Rome, Italy; 7Laura Bazzichi, MD,Stefano Bombardieri, MD: University of Pisa, Pisa, Italy; 8VasilikiLiakouli, MD, Paola Cipriani, MD, Roberto Giacomelli, MD, PhD:University of L’Aquila, L’Aquila, Italy.
Address correspondence to Mirko Manetti, PhD, Departmentof Anatomy, Histology and Forensic Medicine, University of Florence,Viale G. B. Morgagni 85, I-50134 Florence, Italy. E-mail:[email protected].
Submitted for publication May 7, 2010; accepted in revisedform October 12, 2010.
247
of the skin and internal organs (1,2). In SSc, organdysfunction is ultimately caused by vascular failure andoverproduction and deposition of extracellular matrix(ECM) components in the affected tissues (1,2). Vascu-lopathy is a critical issue, and vascular complications,including digital ulceration, pulmonary arterial hyper-tension (PAH), and scleroderma renal crisis, jeopardizepatients’ quality of life and have a very poor prognosis(3).
Clinical and experimental data suggest that thepathogenesis of SSc is multifactorial, involving bothgenetic and environmental factors (4). The search fornew genetic components may help to identify markerspredicting specific vascular, fibrotic, or autoimmunemanifestations, thus suggesting new targeted treatmentstrategies.
Urokinase-type plasminogen activator receptor(uPAR; CD87 antigen) is a key component of theplasminogen activation system, a well-characterized sys-tem of serine proteases (5). The targeted gene disrup-tion of the plasminogen activation system componentshas shown their crucial involvement in several patho-physiologic conditions, including thrombolysis, inflam-
mation, immunity, fertility, tissue and vascular remodel-ing, cancer invasion, and vascularization (5). Urokinase-type plasminogen activator receptor is a glycosylphosphatidylinositol–anchored 3–extracellular domainreceptor expressed on cells of different lineages, such asmonocytes, neutrophils, activated T cells, fibroblasts,endothelial cells, and vascular smooth muscle cells (5,6).Urokinase-type plasminogen activator receptor concen-trates the serine protease activity of its ligand, uPA, atthe cell–matrix interface and promotes ECM and vascu-lar remodeling (6). Moreover, uPAR not only functionsas a uPA receptor, but also interacts with the ECMprotein vitronectin and intracellular signaling mediators,such as integrin family members and the G protein–coupled chemotactic receptor formyl peptide receptor–like 1 (6). Therefore, uPAR is a critical regulator of avariety of cell functions, including growth factor activa-tion, cell adhesion, migration, differentiation, and pro-liferation (5,6).
Recent studies have shown that the uPAR pro-tein is implicated in SSc-related microvascular abnor-malities and impaired angiogenesis (7,8). In particular, ithas been demonstrated that uPAR is overexpressed in
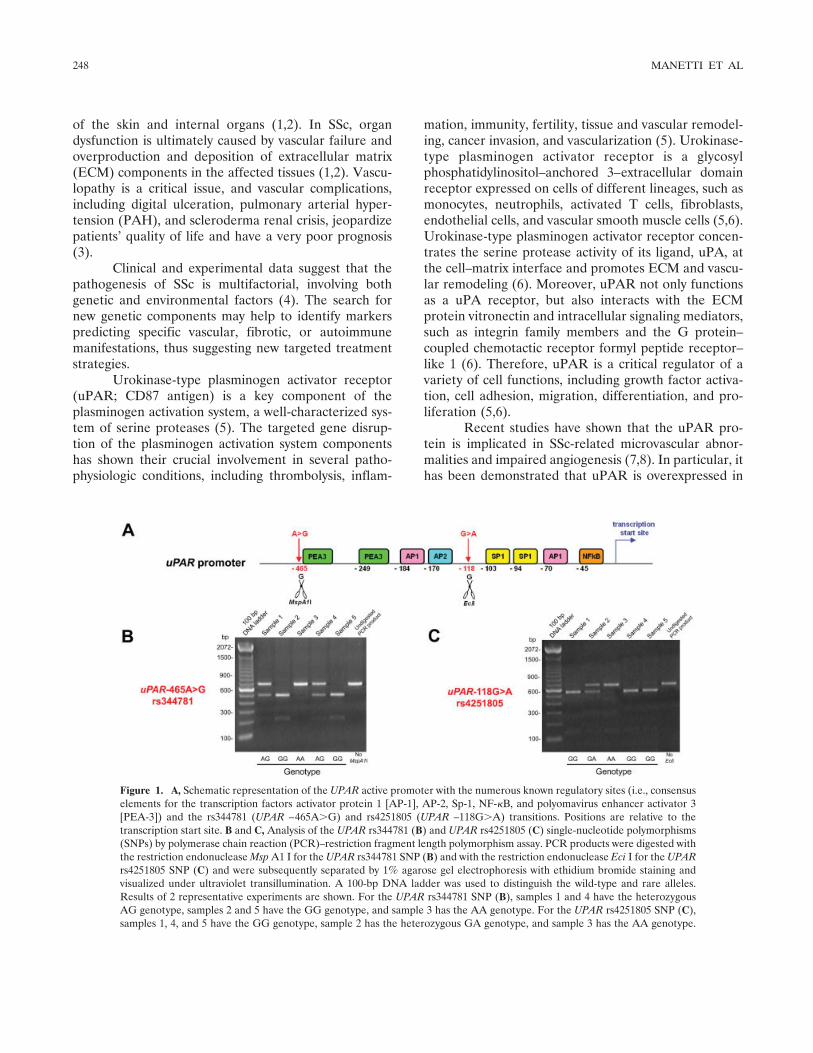
Figure 1. A, Schematic representation of the UPAR active promoter with the numerous known regulatory sites (i.e., consensuselements for the transcription factors activator protein 1 [AP-1], AP-2, Sp-1, NF-�B, and polyomavirus enhancer activator 3[PEA-3]) and the rs344781 (UPAR –465A�G) and rs4251805 (UPAR –118G�A) transitions. Positions are relative to thetranscription start site. B and C, Analysis of the UPAR rs344781 (B) and UPAR rs4251805 (C) single-nucleotide polymorphisms(SNPs) by polymerase chain reaction (PCR)–restriction fragment length polymorphism assay. PCR products were digested withthe restriction endonuclease Msp A1 I for the UPAR rs344781 SNP (B) and with the restriction endonuclease Eci I for the UPARrs4251805 SNP (C) and were subsequently separated by 1% agarose gel electrophoresis with ethidium bromide staining andvisualized under ultraviolet transillumination. A 100-bp DNA ladder was used to distinguish the wild-type and rare alleles.Results of 2 representative experiments are shown. For the UPAR rs344781 SNP (B), samples 1 and 4 have the heterozygousAG genotype, samples 2 and 5 have the GG genotype, and sample 3 has the AA genotype. For the UPAR rs4251805 SNP (C),samples 1, 4, and 5 have the GG genotype, sample 2 has the heterozygous GA genotype, and sample 3 has the AA genotype.
248 MANETTI ET AL

microvascular endothelial cells isolated from SSc skin(7). However, in SSc endothelial cells, uPAR domain 1undergoes a matrix metalloproteinase 12 (MMP-12)–dependent cleavage, resulting in the impairment of cellmotility and invasion and loss of angiogenic capacity(7,8). Furthermore, increased plasma levels of solubleuPAR have been found in SSc patients (9). Interestingly,experimental evidence suggests that Upar gene defi-ciency is implicated in the pathogenesis and progressionof dermal and renal fibrosis in mice (10,11). In addition,the regulation of cell surface uPAR expression wasshown to be a crucial step in the transdifferentiation offibroblasts into myofibroblasts, which play a major rolein fibrotic disorders by excessive synthesis and contrac-tion of ECM (12).
The human UPAR gene (also known as PLAURor CD87) has been mapped to the long arm of chromo-some 19 (19q13.2) and contains 7 exons and 6 introns(6). Two single-nucleotide polymorphisms (SNPs) havebeen identified in the active promoter region of theUPAR gene: an A-to-G transition at position –465(UPAR–465A�G, rs344781) and a G-to-A transition atposition –118 (UPAR –118G�A, rs4251805) upstreamof the transcription start site (13). These SNPs arelocated within a region that contains a number ofregulatory sites, including consensus elements for the
transcription factors activator protein 1 (AP-1), AP-2,Sp-1, NF-�B, and polyomavirus enhancer activator 3,and they may therefore influence UPAR gene expression(13) (Figure 1).
In the present study, we considered the UPARgene as a potential candidate gene for SSc, and weinvestigated whether UPAR rs344781 and rs4251805gene polymorphisms might be associated with SSc sus-ceptibility and clinical phenotypes in 2 European Cau-casian populations. Given the pivotal role of uPAR invascular biology, we also analyzed the possible associa-tion of UPAR SNPs with the major vascular complica-tions of SSc, such as digital ulceration and PAH.
PATIENTS AND METHODS
Study population and study design. Using a hypothe-sis-driven candidate gene approach, we performed a case–control association study, with a replication strategy, thatincluded a total of 1,339 individuals from 2 European Cauca-sian populations. The Italian discovery cohort consisted of 388consecutive SSc patients and 391 control subjects. The Frenchreplication cohort comprised 344 consecutive SSc patients and216 control subjects. All SSc patients and control subjects inthe discovery cohort were of Italian European Caucasianancestry, which was defined as all 4 grandparents being ItalianCaucasian. Similarly, all study subjects in the replication cohortwere of French European Caucasian ancestry (i.e., all 4
Table 1. Demographic, clinical, and serologic characteristics of the 732 patients with SSc*
CharacteristicItalian cohort
(n � 388)†French cohort
(n � 344)‡
Combinedcohorts
(n � 732)
Age, mean � SD years 57.5 � 12.4 56.8 � 12.7 57.2 � 12.6Disease duration, mean � SD years 8.6 � 7.4 11.0 � 8.4 9.7 � 7.9Sex
Male 51 (13.1) 63 (18.3) 114 (15.6)Female 337 (86.9) 281 (81.7) 618 (84.4)
Disease subsetlcSSc 248 (63.9) 202 (65.4) 450 (64.6)dcSSc 140 (36.1) 107 (34.6) 247 (35.4)
Autoantibody positiveANA 346 (89.2) 251 (84.8) 597 (87.3)Anti–topo I 125 (32.2) 73 (26.2) 198 (29.7)ACA 131 (34.5) 94 (33.5) 225 (34.0)
Vascular complicationsDigital ulceration (past and/or current) 145 (45.6) 113 (40.8) 258 (43.4)PAH (confirmed by right-sided heart catheterization) 65 (16.8) 27 (8.3) 92 (12.9)
ILD 101 (28.9) 141 (45.6) 242 (36.7)
* Except where indicated otherwise, values are the number (%) of patients. SSc � systemic sclerosis.† Data were missing for anticentromere antibody (ACA) positivity (8 patients), past and/or current digitalulceration (70 patients), pulmonary arterial hypertension (PAH) (2 patients), and interstitial lung disease(ILD) on high-resolution computed tomography scan (38 patients).‡ Data were missing for limited cutaneous SSc (lcSSc) and diffuse cutaneous SSc (dcSSc) subsets (35patients), antinuclear antibody (ANA) positivity (48 patients), anti–topoisomerase I (anti–topo I)positivity (65 patients), ACA positivity (63 patients), past and/or current digital ulceration (67 patients),PAH (18 patients), and ILD on high-resolution computed tomography scan (35 patients).
ASSOCIATION BETWEEN UPAR GENE VARIANT AND SSc VASCULAR PHENOTYPE 249

grandparents being French Caucasian). SSc patients and con-trols were distributed throughout Italy and France. The 2control groups consisted of healthy, unrelated blood donors orvolunteers who were matched to the SSc patients by age, sex,ethnicity, and geographic region. The study was approved byall local institutional review boards, and written informedconsent was obtained from all study subjects.
Clinical assessment. At enrollment, a detailed clinicalhistory was established for the patients with SSc by reviewingtheir medical records. Patients with a concomitant auto-immune or connective tissue disease (overlap syndromes) wereexcluded from the study. Limited cutaneous SSc (lcSSc) anddiffuse cutaneous SSc (dcSSc) subsets were defined accordingto the criteria described by LeRoy et al (14). For classificationaccording to cutaneous subset, the maximum extent of skininvolvement during the disease course was taken into account.Disease duration was calculated since the first non–Raynaud’sphenomenon symptom of SSc.
Patients with SSc were phenotypically assessed aspreviously recommended (15). Past and/or present digitalulceration (i.e., ulceration on the fingertips and other fingerareas) was recorded. Patients with a systolic pulmonary arterypressure of �40 mm Hg at echocardiography underwentright-sided heart catheterization to confirm the presence ofPAH. Confirmed PAH was defined as a mean pulmonaryartery pressure of �25 mm Hg at rest or �30 mm Hg duringexercise, with normal pulmonary capillary wedge pressure(�15 mm Hg). Interstitial lung disease was studied withhigh-resolution computed tomography scanning. Patients withSSc were tested for antinuclear antibodies using indirectimmunofluorescence and HEp-2 cells as antigen substrate(Antibodies Inc.). Anticentromere antibodies (ACAs) weredetermined by their distinctive indirect immunofluorescencepattern on HEp-2 cells. Anti–topoisomerase I (anti–topo I)antibodies were determined by passive immunodiffusionagainst calf thymus extract (Inova Diagnostics) or by counter-immunoelectrophoresis. Demographic, clinical, and serologiccharacteristics of the SSc patients in both the Italian discoverycohort and the French replication cohort are summarized inTable 1.
Genotyping. Genomic DNA was purified from peri-pheral blood mononuclear cells according to standard pro-cedures. The UPAR rs344781 and rs4251805 SNPs were geno-typed by polymerase chain reaction–restriction fragmentlength polymorphism (PCR-RFLP) assay using the forwardprimer 5�-CCACTACGCCCGGCTAATTT-3� and the reverseprimer 5�-GTCCCTGCACGTCTTCTCTC-3�, which were de-signed on the basis of the UPAR gene promoter sequencepublished in the National Institutes of Health GenBank data-base (Homo sapiens PLAUR gene, promoter region and partialCoding DNA Sequence; accession no. AY620436). PCR wasperformed in a total reaction volume of 25 �l containing 2 �lof diluted genomic DNA, using the AccuPrime SuperMix IIsystem (Invitrogen). The amplification protocol was as follows:initial denaturation at 94°C for 2 minutes, 35 cycles of 94°C for30 seconds (denaturation), 62°C for 30 seconds (annealing),and 68°C for 1 minute (extension), with a final extension at68°C for 5 minutes, using a TC-512 thermocycler (Techne).
PCR products were digested with the Msp A1 I restric-tion enzyme for UPAR rs344781 SNP genotyping and the EciI restriction enzyme for UPAR rs4251805 SNP genotyping
(both from New England Biolabs) (Figure 1). Msp A1 Irestriction analysis was carried out by adding 2 units of Msp A1I to 10 �l of PCR product with 1� NEBuffer 4 (New EnglandBiolabs) and 100 �g/ml bovine serum albumin (BSA) in a totalreaction volume of 15 �l. The UPAR rs344781 wild-type A andrs344781 rare G alleles could be differentiated due to arestriction site between basepairs –466 and –467 in the rare Gallele. Eci I restriction analysis was done by adding 1.5 units ofEci I to 10 �l of PCR product with 1� NEBuffer 2 and 100�g/ml BSA in a total reaction volume of 15 �l. By Eci Iendonuclease fragmentation, the UPAR rs4251805 wild-type Gallele could be distinguished from the rs4251805 rare A alleleby cutting of the G allele between basepairs –106 and –107.
Following incubation at 37°C for 2 hours, PCR-RFLPproducts were separated by 1% agarose gel electrophoresiswith ethidium bromide staining and a 100-bp DNA ladder(Invitrogen), and visualized under ultraviolet transillumination(Figure 1). Genotyping was performed under blinded condi-tions with regard to the disease status of subjects. PCR-RFLPresults were analyzed by 2 independent assessors with consen-sus agreement. In the case of discordant results (�2% of allsamples), the particular samples were genotyped again inorder to avoid any errors in genotype assessment. Controlsamples of predetermined genotypes and negative controls(i.e., undigested PCR products, PCR amplification withoutgenomic DNA template) were included in each sample set, and�20% of all samples were randomly selected for confirmation,showing �99% accuracy.
Statistical analysis. Statistical analyses were per-formed using SPSS software for Windows, version 13.0. Cate-gorical and quantitative variables were described as numbersand percentages and as the mean � SD, respectively. The levelof significance for all tests was set to a Type I error rate of � �5% (P � 0.05). A chi-square test with 1 degree of freedom wasused to compare the observed numbers of each genotype incontrol subjects with those expected for a population inHardy-Weinberg equilibrium. Allele frequency of the UPARrs344781 and rs4251805 SNPs was obtained by direct count.Genotype distribution and allele frequency of the UPARrs344781 and rs4251805 SNPs in both patients and healthycontrols were compared by chi-square analysis. The associationbetween the UPAR rs344781 SNP and SSc or clinical pheno-types (cutaneous subset, autoantibody status, and vascularcomplications) was assessed using a standard univariate logisticregression analysis. Odds ratios (ORs) with 95% confidenceintervals (95% CIs) were determined. When an association wasdetected in the Italian Caucasian discovery cohort, we per-formed a replication study in the French Caucasian cohort tofurther strengthen the power of the results, and subsequentlythe analysis was performed in the combined European Cauca-sian populations in order to increase the power of detection.
In the univariate analysis, a significant association wasfound between the UPAR rs344781 SNP and the clinicalphenotypes of lcSSc, ACA-positive SSc, SSc with digital ulcer-ation, and SSc with PAH. Therefore, we used a multivariatelogistic regression analysis model including these associatedSSc phenotypes as the dependent variables, in order to assessthe independent effect of the UPAR rs344781 SNP on thoseclinical phenotypes.
Power calculations were driven through an asymptom-atic noncentral chi-square approach. The combined set of 732
250 MANETTI ET AL
SSc cases and 607 controls provided a power of �80% todetect an association between SSc and the UPAR rs344781 Gallele with an OR of 1.30 at the 5% significance level.
RESULTS
Italian Caucasian population. Genotype fre-quencies were in Hardy-Weinberg equilibrium in theItalian Caucasian control population for the UPARrs344781 and rs4251805 SNPs. Moreover, both thers344781 and rs4251805 allele and genotype frequenciesobserved in healthy controls corresponded well with theEuropean Caucasian population frequencies reported inthe International HapMap project database.
The case–control association study performed inthe discovery sample (388 SSc patients and 391 controls)did not reveal any role of the UPAR rs4251805 SNP ininfluencing the predisposition to SSc or clinical pheno-type. Indeed, both genotype distribution and allelefrequency for the UPAR rs4251805 polymorphism weresimilar in SSc patients and healthy controls (genotypes
84.0% GG, 14.8% GA, 1.2% AA versus 88.8% GG,10.0% GA, 1.2% AA, respectively; P � 0.3) (minorallele frequency 0.09 versus 0.06, respectively; P � 0.1).
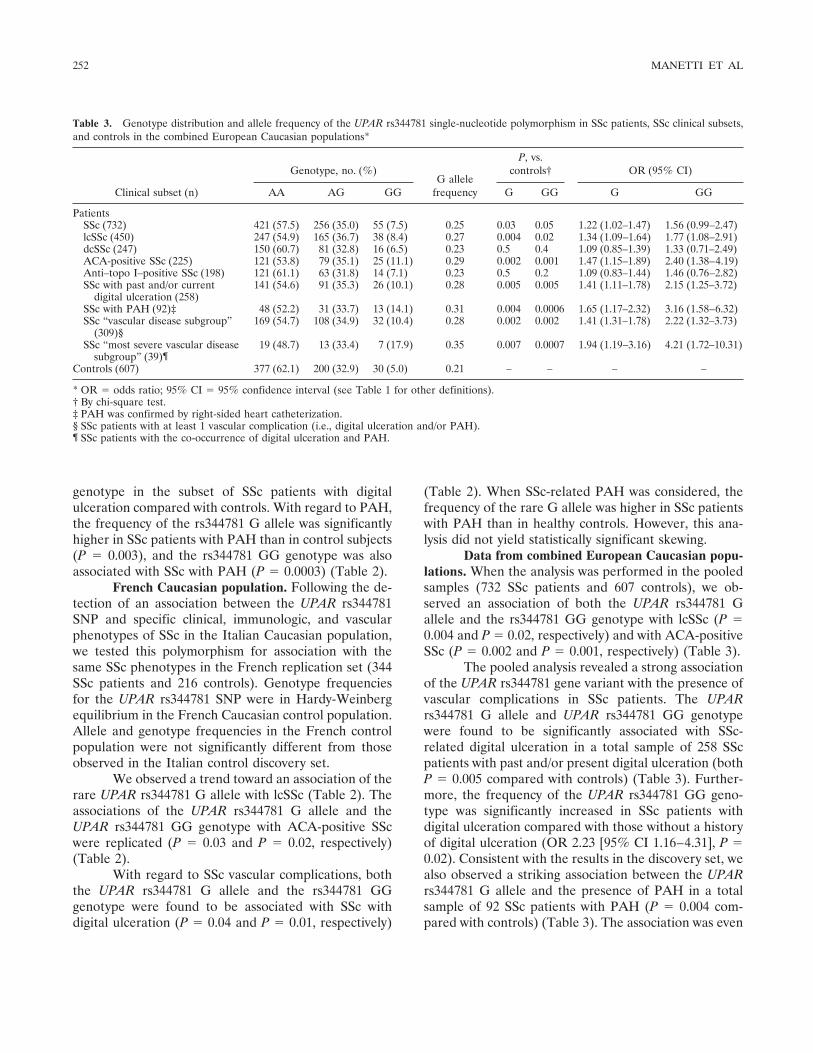
For the UPAR rs344781 SNP, the frequency ofthe rare G allele was significantly higher in SSc patientsthan in control subjects (P � 0.03) (Table 2). When theSSc clinical subsets were analyzed, the UPAR rs344781G allele and homozygous GG genotype were found tobe associated with lcSSc (both P � 0.02), but not withdcSSc (Table 2). In addition, both the UPAR rs344781 Gallele and the UPAR rs344781 GG genotype were sig-nificantly associated with ACA-positive SSc (both P �0.02) (Table 2). No association with anti–topo I antibod-ies was found.
When SSc vascular complications were consid-ered, the rs344781 G allele frequency was found to be0.29 in SSc patients with digital ulceration, comparedwith 0.23 in controls (P � 0.03) (Table 2). Furthermore,we observed an increased, but not significantly different,percentage of subjects carrying the homozygous GG
Table 2. Genotype distribution and allele frequency of the UPAR rs344781 single-nucleotide polymorphism in SSc patients, SSc clinical subsets,and controls in the Italian and French Caucasian populations*
Population,clinical subset (n)
Genotype, no. (%)G allele
frequency
P, vs.controls† OR (95% CI)
AA AG GG G GG G GG
Italian CaucasianPatients
SSc (388) 210 (54.1) 143 (36.9) 35 (9.0) 0.27 0.03 0.07 1.29 (1.03–1.63) 1.66 (0.96–2.89)lcSSc (248) 132 (53.2) 90 (36.3) 26 (10.5) 0.29 0.02 0.02 1.37 (1.06–1.77) 1.96 (1.09–3.55)dcSSc (140) 78 (55.7) 53 (37.9) 9 (6.4) 0.25 0.3 0.7 1.16 (0.85–1.59) 1.15 (0.52–2.57)ACA-positive SSc (131) 68 (51.9) 48 (36.6) 15 (11.5) 0.30 0.02 0.02 1.45 (1.06–1.98) 2.17 (1.09–4.32)Anti–topo I–positive
SSc (125)72 (57.6) 45 (36.0) 8 (6.4) 0.24 0.6 0.7 1.10 (0.79–1.54) 1.15 (0.49–2.64)
SSc with past and/orcurrent digitalulceration (145)
75 (51.7) 56 (38.6) 14 (9.7) 0.29 0.03 0.09 1.39 (1.03–1.89) 1.79 (0.89–3.61)
SSc with PAH (65)‡ 32 (49.2) 21 (32.3) 12 (18.5) 0.35 0.003 0.0003 1.81 (1.21–2.69) 3.79 (1.78–8.12)Controls (391) 236 (60.4) 133 (34.0) 22 (5.6) 0.23 – – – –
French CaucasianPatients
SSc (344) 211 (61.3) 113 (32.9) 20 (5.8) 0.22 0.2 0.2 1.20 (0.89–1.62) 1.67 (0.72–3.90)lcSSc (202) 115 (56.9) 75 (37.1) 12 (6.0) 0.25 0.06 0.3 1.36 (0.98–1.89) 1.64 (0.66–4.11)dcSSc (107) 72 (67.3) 28 (26.2) 7 (6.5) 0.20 0.9 0.2 1.03 (0.68–1.55) 1.82 (0.64–5.16)ACA-positive SSc (94) 53 (56.4) 31 (33.0) 10 (10.6) 0.27 0.03 0.02 1.57 (1.05–2.34) 3.09 (1.18–8.12)Anti–topo I–positive
SSc (73)49 (67.1) 18 (24.7) 6 (8.2) 0.21 0.7 0.1 1.09 (0.68–1.74) 2.33 (0.78–6.95)
SSc with past and/orcurrent digitalulceration (113)
66 (58.4) 35 (31.0) 12 (10.6) 0.26 0.04 0.01 1.49 (1.01–2.17) 3.20 (1.25–8.21)
SSc with PAH (27)‡ 16 (59.3) 10 (37.0) 1 (3.7) 0.22 0.6 0.9 1.20 (0.61–2.40) 1.10 (0.13–9.38)Controls (216) 141 (65.3) 67 (31.0) 8 (3.7) 0.19 – – – –
* OR � odds ratio; 95% CI � 95% confidence interval (see Table 1 for other definitions).† By chi-square test.‡ PAH was confirmed by right-sided heart catheterization.
ASSOCIATION BETWEEN UPAR GENE VARIANT AND SSc VASCULAR PHENOTYPE 251
genotype in the subset of SSc patients with digitalulceration compared with controls. With regard to PAH,the frequency of the rs344781 G allele was significantlyhigher in SSc patients with PAH than in control subjects(P � 0.003), and the rs344781 GG genotype was alsoassociated with SSc with PAH (P � 0.0003) (Table 2).
French Caucasian population. Following the de-tection of an association between the UPAR rs344781SNP and specific clinical, immunologic, and vascularphenotypes of SSc in the Italian Caucasian population,we tested this polymorphism for association with thesame SSc phenotypes in the French replication set (344SSc patients and 216 controls). Genotype frequenciesfor the UPAR rs344781 SNP were in Hardy-Weinbergequilibrium in the French Caucasian control population.Allele and genotype frequencies in the French controlpopulation were not significantly different from thoseobserved in the Italian control discovery set.
We observed a trend toward an association of therare UPAR rs344781 G allele with lcSSc (Table 2). Theassociations of the UPAR rs344781 G allele and theUPAR rs344781 GG genotype with ACA-positive SScwere replicated (P � 0.03 and P � 0.02, respectively)(Table 2).
With regard to SSc vascular complications, boththe UPAR rs344781 G allele and the rs344781 GGgenotype were found to be associated with SSc withdigital ulceration (P � 0.04 and P � 0.01, respectively)
(Table 2). When SSc-related PAH was considered, thefrequency of the rare G allele was higher in SSc patientswith PAH than in healthy controls. However, this ana-lysis did not yield statistically significant skewing.
Data from combined European Caucasian popu-lations. When the analysis was performed in the pooledsamples (732 SSc patients and 607 controls), we ob-served an association of both the UPAR rs344781 Gallele and the rs344781 GG genotype with lcSSc (P �0.004 and P � 0.02, respectively) and with ACA-positiveSSc (P � 0.002 and P � 0.001, respectively) (Table 3).
The pooled analysis revealed a strong associationof the UPAR rs344781 gene variant with the presence ofvascular complications in SSc patients. The UPARrs344781 G allele and UPAR rs344781 GG genotypewere found to be significantly associated with SSc-related digital ulceration in a total sample of 258 SScpatients with past and/or present digital ulceration (bothP � 0.005 compared with controls) (Table 3). Further-more, the frequency of the UPAR rs344781 GG geno-type was significantly increased in SSc patients withdigital ulceration compared with those without a historyof digital ulceration (OR 2.23 [95% CI 1.16–4.31], P �0.02). Consistent with the results in the discovery set, wealso observed a striking association between the UPARrs344781 G allele and the presence of PAH in a totalsample of 92 SSc patients with PAH (P � 0.004 com-pared with controls) (Table 3). The association was even
Table 3. Genotype distribution and allele frequency of the UPAR rs344781 single-nucleotide polymorphism in SSc patients, SSc clinical subsets,and controls in the combined European Caucasian populations*
Clinical subset (n)
Genotype, no. (%)G allele
frequency
P, vs.controls† OR (95% CI)
AA AG GG G GG G GG
PatientsSSc (732) 421 (57.5) 256 (35.0) 55 (7.5) 0.25 0.03 0.05 1.22 (1.02–1.47) 1.56 (0.99–2.47)lcSSc (450) 247 (54.9) 165 (36.7) 38 (8.4) 0.27 0.004 0.02 1.34 (1.09–1.64) 1.77 (1.08–2.91)dcSSc (247) 150 (60.7) 81 (32.8) 16 (6.5) 0.23 0.5 0.4 1.09 (0.85–1.39) 1.33 (0.71–2.49)ACA-positive SSc (225) 121 (53.8) 79 (35.1) 25 (11.1) 0.29 0.002 0.001 1.47 (1.15–1.89) 2.40 (1.38–4.19)Anti–topo I–positive SSc (198) 121 (61.1) 63 (31.8) 14 (7.1) 0.23 0.5 0.2 1.09 (0.83–1.44) 1.46 (0.76–2.82)SSc with past and/or current
digital ulceration (258)141 (54.6) 91 (35.3) 26 (10.1) 0.28 0.005 0.005 1.41 (1.11–1.78) 2.15 (1.25–3.72)
SSc with PAH (92)‡ 48 (52.2) 31 (33.7) 13 (14.1) 0.31 0.004 0.0006 1.65 (1.17–2.32) 3.16 (1.58–6.32)SSc “vascular disease subgroup”
(309)§169 (54.7) 108 (34.9) 32 (10.4) 0.28 0.002 0.002 1.41 (1.31–1.78) 2.22 (1.32–3.73)
SSc “most severe vascular diseasesubgroup” (39)¶
19 (48.7) 13 (33.4) 7 (17.9) 0.35 0.007 0.0007 1.94 (1.19–3.16) 4.21 (1.72–10.31)
Controls (607) 377 (62.1) 200 (32.9) 30 (5.0) 0.21 – – – –
* OR � odds ratio; 95% CI � 95% confidence interval (see Table 1 for other definitions).† By chi-square test.‡ PAH was confirmed by right-sided heart catheterization.§ SSc patients with at least 1 vascular complication (i.e., digital ulceration and/or PAH).¶ SSc patients with the co-occurrence of digital ulceration and PAH.
252 MANETTI ET AL

stronger when the GG genotype was considered (P �0.0006). In addition, both the UPAR rs344781 G alleleand the rs344781 homozygous GG genotype were foundto be significantly associated with PAH when comparingSSc patients with PAH and those without PAH (alleleOR 1.40 [95% CI 1.01–1.96], P � 0.04; genotype OR2.32 [95% CI 1.19–4.53], P � 0.01). In a multivariatelogistic regression analysis model including the subset ofpatients with lcSSc, autoantibody status, and the vascularcomplications of SSc (SSc with digital ulceration and SScwith PAH), the UPAR rs344781 GG genotype remainedan independent risk factor both for SSc with digitalulceration (OR 1.96 [95% CI 1.01–3.85], P � 0.04) andfor SSc with PAH (OR 2.68 [95% CI 1.25–5.75],P � 0.01).
In order to further investigate the role of theUPAR rs344781 gene variant in influencing the severityof SSc vascular phenotype, we analyzed the rs344781genotype distribution and allele frequency in the “vas-cular disease subgroup” of SSc patients, which com-prised patients with at least 1 vascular complication (i.e.,digital ulceration and/or PAH), as well as in the “mostsevere vascular disease subgroup,” including SSc pa-tients with co-occurrence of digital ulceration and PAH.The frequency of the UPAR rs344781 G allele wassignificantly higher in both the “vascular disease sub-group” and the “most severe vascular disease subgroup”of SSc patients than in control subjects (P � 0.002 andP � 0.007, respectively) (Table 3). A significant associ-ation with homozygosity for the G allele was also foundin the “vascular disease subgroup” (P � 0.002), and amore evident association in the “most severe vasculardisease subgroup” (P � 0.0007) was observed. In con-trast, no association was found between the UPARrs344781 SNP and SSc without vascular complications.Concordant with these results, the frequencies of boththe UPAR rs344781 G allele and the GG genotype weresignificantly higher in the SSc “vascular disease sub-group” compared with SSc patients without any vascularcomplication (allele OR 1.28 [95% CI 1.01–1.63], P �0.04; genotype OR 2.02 [95% CI 1.15–3.55], P � 0.01).
DISCUSSION
In this hypothesis-driven candidate gene associa-tion study, we analyzed for the first time the role of theUPAR gene in influencing the predisposition to SSc andits clinical phenotypes. Our findings show that the UPARrs344781 gene promoter variant is significantly associ-ated with defined SSc phenotypes and may thereforecontribute to the typical heterogeneity of the disease.
Indeed, we observed an association of both the UPARrs344781 G allele and the GG genotype with SSc, whichwas restricted to the subsets of patients with lcSSc andACA positivity, as well as to the subsets of SSc patientswith major vascular complications, such as digital ulcer-ation and PAH. However, it is well known that certainsubgroups of SSc patients, such as those with lcSSc andACAs, are at high risk of developing PAH (16) andperipheral vascular occlusive disease complicated bydigital ulceration, gangrene, and digital loss (17,18).Therefore, we performed multivariate logistic regressionanalyses, which highlighted that, among the cutaneous,autoimmune, and vascular phenotypes, the UPARrs344781 SNP was most significantly and independentlyassociated with SSc-related digital ulceration and PAH.Moreover, when we compared SSc patients with at least1 vascular complication (i.e., digital ulceration and/orPAH) and SSc patients with the co-occurrence of digitalulceration and PAH, we also observed an effect of thers344781 SNP in influencing the severity of the SScvascular phenotype.
SSc belongs to a group of complex disorders inwhich multiple genetic and environmental componentsinteract in modulating disease pathogenesis and evolu-tion (4,19). It appears that different genetic variants indifferent candidate genes may influence and predict thedevelopment of different SSc clinical subsets and majororgan manifestations, such as pulmonary fibrosis andPAH (4,20). Candidate genes for SSc include genesencoding proteins involved in autoimmunity, fibrosis,and vasculopathy, fully reflecting disease heterogeneityand complexity (4). In this study, we considered UPARas a potential candidate gene for SSc, since it encodesfor a pleiotropic cell surface receptor involved in ECMturnover, innate and adaptive immunity, angiogenesis,and vascular remodeling (5–7,10–12,21). Moreover, datafrom our previous studies showed involvement of theuPAR protein in SSc microvascular abnormalities andimpaired angiogenesis (7–9).
Our data, which provide evidence for an associ-ation of the UPAR rs344781 gene variant with definedcutaneous, autoimmune, and vascular phenotypes ofSSc, are concordant with previous findings demonstrat-ing the role of uPAR in exerting pleiotropic functions,which are crucial for different pathophysiologic pro-cesses (21,22). Urokinase-type plasminogen activatorreceptor participates in the initiation of the innate andadaptive immune responses by regulating mononuclearcell adhesion and migration in peripheral tissues, and byfavoring T cell activation, proliferation, and differentia-tion in peripheral lymphoid organs (21). Indeed, uPAR
ASSOCIATION BETWEEN UPAR GENE VARIANT AND SSc VASCULAR PHENOTYPE 253

modulates several steps in the inflammatory cascade, israpidly up-regulated during T cell activation, and isexpressed by antigen-presenting cells, suggesting its par-ticipation in T cell priming (21). In addition, uPAR mayhave a crucial role in fibrosis, since targeted deletion ofthe Upar gene in mice up-regulates fibrosis-related geneexpression and attenuates the production and activity ofMMPs, resulting in reduced ECM turnover and collagenaccumulation (10).
Even more important is the fact that uPAR playskey roles in angiogenesis, cardiovascular disorders, andvascular remodeling by regulating multiple endothelialcell and vascular smooth muscle cell functions, such asadhesion, migration, invasion, and proliferation (7,8,22).Indeed, vasculopathy in SSc consists of a progressive lossof capillaries not compensated by angiogenesis, as wellas intimal proliferation and structural changes in multi-ple vascular beds leading to vessel narrowing and oblit-eration with consequent chronic tissue ischemia andsevere organ complications (2,3). Together with ourprevious findings implicating uPAR in SSc endothelialcell defects and impaired angiogenesis (7,8), the asso-ciation between the UPAR rs344781 polymorphism andSSc-related digital ulceration and PAH further supportsthe notion that uPAR plays a role in the vascularpathophysiology of SSc.
Among the vascular manifestations of SSc, digitalulceration is a major and common clinical problemassociated with substantial morbidity (reduced quality oflife, pain, and disability), and it can evolve into gangreneand result in amputation (18,23). PAH has emerged asthe most devastating vascular complication of SSc, witha dramatic impact on prognosis and survival, being thesingle most common cause of disease-related death (24).In order to identify patients at high risk of developingdigital ulceration and PAH, and thus to allow earlydiagnosis and improve outcomes in SSc, it is mostimportant to find predictors of these major vascularcomplications. In particular, predicting PAH appears tobe very critical for increasing the survival rate in SSc. Inthis scenario, our results add the UPAR gene to anemerging pool of vasculopathy risk factors, including theKCNA5, TNFAIP3, IL23R, SDF-1/CXCL12, ENG,NOS2, and CX3CR1 genes, that could be used in thefuture as powerful indicators of the development ofSSc-related PAH (4,25–31). Moreover, future gene–gene interaction investigations will contribute to a betterunderstanding of the genetic predisposition to SSc-related vasculopathy.
Methodologic limitations of genetic studies mustalways be considered. Appropriate sample sizes and
homogeneity in the genetic background of the studypopulation are critical to avoid result bias by low statis-tical power and population stratification. The presentstrategy, which included a large sample size of pheno-typically well-characterized populations of similar eth-nicity with replication in an independent cohort,emphasizes the relevance of our results. This workconsequently provides a basis for further studies of theassociation of the UPAR rs344781 SNP with SSc pheno-types in different ethnic populations.
It should be noted that we did not apply Bonfer-roni correction for multiple testing, since the clinicalfeatures tested were not truly independent. In addition,there is no clear consensus about the best way to correct,and application of this statistical methodology seems tooconservative when studying rare subgroups of very het-erogeneous disorders, such as SSc (32). Nevertheless,we performed multivariate logistic regression analysesto evaluate the independence of the observed associa-tions between the UPAR rs344781 SNP and differentclinical phenotypes. Because of the relatively low prev-alence of PAH in SSc (8–17% of patients), the associa-tion will need to be confirmed in larger cohorts of SScpatients with PAH. In addition, since the PAH pheno-type can change during the disease course, a prospectivefollowup study of the 2 cohorts will improve vascularphenotype analyses. However, our population was oneof the largest used to date for a genetic association studyrelated to SSc with digital ulceration and SSc with PAH,and, notably, PAH was diagnosed using the gold stan-dard method of catheterization.
In the present study, we used a conventionalhypothesis-driven candidate gene approach that hasbeen employed in most of the genetic studies of SSccarried out to date (20). However, the genome-wideassociation study approach, which makes it possible toscreen SNPs across the entire genome without previousknowledge of candidate regions or genes, has yieldedrecognition of a wealth of new genetic susceptibility loci,leading to the identification of new pathogenetic mech-anisms of complex genetic disorders (20). A first largegenome-wide association study of SSc was recently re-ported (33). That study has identified CD247 as a newsusceptibility locus for SSc and has also confirmed therole of different autoimmunity-associated gene regions,such as HLA class II, and genes, such as IRF5 andSTAT4, as SSc genetic risk factors (33). To the best ofour knowledge, the UPAR gene and other genes thathave been previously associated with SSc vascular phe-notypes, but not with SSc per se, were not highlighted inthat genome-wide association study (25–31,33). There-
254 MANETTI ET AL

fore, large genome-wide association studies with precisevascular phenotyping will be required in order to firmlyestablish the role of those genes as SSc-related vascu-lopathy risk factors.
Moreover, it should be noted that in the presentstudy we selected, as genetic markers, 2 variants locatedwithin the active UPAR promoter region which maymodulate UPAR transcriptional activity in an allele-specific manner, although the functional consequencesremain unclear (13). However, we cannot exclude thepossibility that linkage disequilibrium between thers344781 SNP and other variants that have been recentlyidentified in the UPAR gene (34) may influence SScsusceptibility and vascular phenotype. Further studiesare needed to address this important question. Further-more, it remains to be determined whether the UPARrs344781 SNP may be associated with other SSc vascularcomplications (e.g., scleroderma renal crisis). Finally,further functional studies are necessary to elucidate theexact molecular mechanisms by which the UPARrs344781 SNP may be implicated in different aspects ofSSc vascular phenotype, such as lack of angiogenesis andproliferative vasculopathy. In fact, digital ulceration andPAH are two different vascular complications of SScthat may share some pathogenetic mechanisms but thatmay also have different etiologies. In this context, theassociation of the UPAR rs344781 SNP with both SSc-related digital ulceration and SSc-related PAH could beexplained by the well-known pleiotropic function ofuPAR in vascular biology (22).
In conclusion, our data provide the first geneticevidence of a defined vascular phenotype of SSc taggedby the UPAR rs344781 gene variant, which might be usedas a biomarker in the future. This study provides newinsight into the pathogenesis of SSc, including clues tothe mechanisms leading to the development of severevascular complications, and generates new perspectiveswith regard to potential therapies.
AUTHOR CONTRIBUTIONS
All authors were involved in drafting the article or revising itcritically for important intellectual content, and all authors approvedthe final version to be published. Dr. Manetti had full access to all ofthe data in the study and takes responsibility for the integrity of thedata and the accuracy of the data analysis.Study conception and design. Manetti, Allanore, Fatini, Ibba-Manneschi, Matucci-Cerinic.Acquisition of data. Manetti, Allanore, Revillod, Guiducci, Cuomo,Bonino, Riccieri, Bazzichi, Liakouli, Cipriani, Giacomelli, Bombar-dieri, Valesini, Montecucco, Valentini, Ibba-Manneschi, Matucci-Cerinic.Analysis and interpretation of data. Manetti, Allanore, Fatini, Abbate,Ibba-Manneschi, Matucci-Cerinic.
REFERENCES
1. Varga J, Abraham D. Systemic sclerosis: a prototypic multisystemfibrotic disorder. J Clin Invest 2007;117:557–67.
2. Manetti M, Guiducci S, Ibba-Manneschi L, Matucci-Cerinic M.Mechanisms in the loss of capillaries in systemic sclerosis: angio-genesis versus vasculogenesis. J Cell Mol Med 2010;14:1241–54.
3. Guiducci S, Giacomelli R, Matucci-Cerinic M. Vascular compli-cations of scleroderma. Autoimmun Rev 2007;6:520–3.
4. Allanore Y, Wipff J, Kahan A, Boileau C. Genetic basis forsystemic sclerosis. Joint Bone Spine 2007;74:577–83.
5. Blasi F, Carmeliet P. uPAR: a versatile signalling orchestrator. NatRev Mol Cell Biol 2002;3:932–43.
6. Ragno P. The urokinase receptor: a ligand or a receptor? Story ofa sociable molecule. Cell Mol Life Sci 2006;63:1028–37.
7. D’Alessio S, Fibbi G, Cinelli M, Guiducci S, Del Rosso A,Margheri F, et al. Matrix metalloproteinase 12–dependent cleav-age of urokinase receptor in systemic sclerosis microvascularendothelial cells results in impaired angiogenesis. Arthritis Rheum2004;50:3275–85.
8. Margheri F, Manetti M, Serrati S, Nosi D, Pucci M, Matucci-Cerinic M, et al. Domain 1 of the urokinase-type plasminogenactivator receptor is required for its morphologic and functional,�2 integrin–mediated connection with actin cytoskeleton in humanmicrovascular endothelial cells: failure of association in systemicsclerosis endothelial cells. Arthritis Rheum 2006;54:3926–38.
9. Bandinelli F, Bartoli F, Perfetto E, Del Rosso A, Moggi-PignoneA, Guiducci S, et al. The fibrinolytic system components areincreased in systemic sclerosis and modulated by Alprostadil (�1ciclodestryn). Clin Exp Rheumatol 2005;23:671–7.
10. Kanno Y, Kaneiwa A, Minamida M, Kanno M, Tomogane K,Takeuchi K, et al. The absence of uPAR is associated with theprogression of dermal fibrosis. J Invest Dermatol 2008;128:2792–7.
11. Zhang G, Kim H, Cai X, Lopez-Guisa JM, Alpers CE, Liu Y, et al.Urokinase receptor deficiency accelerates renal fibrosis in obstruc-tive nephropathy. J Am Soc Nephrol 2003;14:1254–71.
12. Bernstein AM, Twining SS, Warejcka DJ, Tall E, Masur SK.Urokinase receptor cleavage: a crucial step in fibroblast-to-myo-fibroblast differentiation. Mol Biol Cell 2007;18:2716–27.
13. Schneider UV, Nielsen RL, Pedersen C, Eugen-Olsen J. Theprognostic value of the suPARnostic ELISA in HIV-1 infectedindividuals is not affected by uPAR promoter polymorphisms.BMC Infect Dis 2007;7:134.
14. LeRoy EC, Black C, Fleischmajer R, Jablonska S, Krieg T,Medsger TA Jr, et al. Scleroderma (systemic sclerosis): classifica-tion, subsets and pathogenesis. J Rheumatol 1988;15:202–5.
15. Valentini G, Matucci Cerinic M. Disease-specific quality indica-tors, guidelines and outcome measures in scleroderma. Clin ExpRheumatol 2007;25:159–62.
16. Steen VD. Autoantibodies in systemic sclerosis. Semin ArthritisRheum 2005;35:35–42.
17. Wigley FM, Wise RA, Miller R, Needleman BW, Spence RJ.Anticentromere antibody as a predictor of digital ischemic loss inpatients with systemic sclerosis. Arthritis Rheum 1992;35:688–93.
18. Steen V, Denton CP, Pope JE, Matucci-Cerinic M. Digital ulcers:overt vascular disease in systemic sclerosis. Rheumatology (Ox-ford) 2009;48 Suppl 3:iii19–24.
19. Agarwal SK, Tan FK, Arnett FC. Genetics and genomic studies inscleroderma (systemic sclerosis). Rheum Dis Clin North Am2008;34:17–40.
20. Allanore Y, Dieude P, Boileau C. Updating the genetics ofsystemic sclerosis. Curr Opin Rheumatol 2010;22:665–70.
21. Mondino A, Blasi F. uPA and uPAR in fibrinolysis, immunity andpathology. Trends Immunol 2004;25:450–5.
22. Binder BR, Mihaly J, Prager GW. uPAR–uPA–PAI-1 interactionsand signaling: a vascular biologist’s view. Thromb Haemost 2007;97:336–42.
ASSOCIATION BETWEEN UPAR GENE VARIANT AND SSc VASCULAR PHENOTYPE 255

23. Amanzi L, Braschi F, Fiori G, Galluccio F, Miniati I, Guiducci S,et al. Digital ulcers in scleroderma: staging, characteristics andsub-setting through observation of 1614 digital lesions. Rheuma-tology (Oxford) 2010;49:1374–82.
24. McLaughlin V, Humbert M, Coghlan G, Nash P, Steen V.Pulmonary arterial hypertension: the most devastating vascularcomplication of systemic sclerosis. Rheumatology (Oxford) 2009;48 Suppl 3:iii25–31.
25. Wipff J, Dieude P, Guedj M, Ruiz B, Riemekasten G, CracowskiJL, et al. Association of a KCNA5 gene polymorphism withsystemic sclerosis–associated pulmonary arterial hypertension inthe European Caucasian population. Arthritis Rheum 2010;62:3093–100.
26. Dieude P, Guedj M, Wipff J, Ruiz B, Riemekasten G, Matucci-Cerinic M, et al. Association of the TNFAIP3 rs5029939 variantwith systemic sclerosis in the European Caucasian population.Ann Rheum Dis 2010;69:1958–64.
27. Agarwal SK, Gourh P, Shete S, Paz G, Divecha D, Reveille JD, etal. Association of interleukin 23 receptor polymorphisms withanti-topoisomerase-I positivity and pulmonary hypertension insystemic sclerosis [published erratum appears in J Rheumatol2010;37:1979]. J Rheumatol 2009;36:2715–23.
28. Manetti M, Liakouli V, Fatini C, Cipriani P, Bonino C, Vettori S,et al. Association between a stromal cell-derived factor 1 (SDF-1/
CXCL12) gene polymorphism and microvascular disease in sys-temic sclerosis. Ann Rheum Dis 2009;68:408–11.
29. Wipff J, Kahan A, Hachulla E, Sibilia J, Cabane J, Meyer O, et al.Association between an endoglin gene polymorphism and systemicsclerosis-related pulmonary arterial hypertension. Rheumatology(Oxford) 2007;46:622–5.
30. Kawaguchi Y, Tochimoto A, Hara M, Kawamoto M, Sugiura T,Katsumata Y, et al. NOS2 polymorphisms associated with thesusceptibility to pulmonary arterial hypertension with systemicsclerosis: contribution to the transcriptional activity. Arthritis ResTher 2006;8:R104.
31. Marasini B, Cossutta R, Selmi C, Pozzi MR, Gardinali M,Massarotti M, et al. Polymorphism of the fractalkine receptorCX3CR1 and systemic sclerosis-associated pulmonary arterialhypertension. Clin Dev Immunol 2005;12:275–9.
32. Perneger TV. What’s wrong with Bonferroni adjustments. BMJ1998;316:1236–8.
33. Radstake TR, Gorlova O, Rueda B, Martin JE, Alizadeh BZ,Palomino-Morales R, et al. Genome-wide association study ofsystemic sclerosis identifies CD247 as a new susceptibility locus.Nat Genet 2010;42:426–9.
34. Stewart CE, Hall IP, Parker SG, Moffat MF, Wardlaw AJ,Connolly MJ, et al. PLAUR polymorphisms and lung function inUK smokers. BMC Med Genet 2009;10:112.
DOI 10.1002/art.30205
Erratum
In the article by Rogerson et al in the November 2010 issue of Arthritis & Rheumatism (pages 3365–3373), anincorrect quantity of soluble interleukin-6 receptor was reported in the “Isolation of primary chondrocytesand treatment with cytokines” section under Materials and Methods. The fourth sentence of the second fullparagraph in the right column on page 3366 should have read, “In the case of IL-6, 100 ng/ml sIL-6R wasadded.”
We regret the error.
256 MANETTI ET AL