
A darkling beetle population in West Texas during the 1997–1998 El Niño
11
Journal of Arid Environments (2001) 49: 711}721 doi:10.1006/jare.2001.0821, available online at http://www.idealibrary.com on A darkling beetle population in West Texas during the 1997+1998 El Nin 8 o Richard J. Deslippe, Jesus R. Salazar & Yu-Jie Guo Department of Biological Sciences, Texas Tech University, Lubbock, TX 79409-3131 U.S.A. (Received 16 October 2000, accepted 14 March 2001, published electronically 3 September 2001) We examined the abundance and distribution of Eleodes extricata (Teneb- rionidae) in a semi-arid region of West Texas. During the 2-year study, climatic conditions were extreme but opposite, with 1997 being unusually wet and mild, and 1998 being unusually dry and hot. These conditions were associated with striking differences in the population structure of the beetles. Under identical pitfall-sampling efforts, we captured 1562 and 671 individuals in 1997 and 1998, respectively. Capture rates were lowest in grassland, and highest in sand shinnery oak, and within the latter, they were lowest in blowouts, intermediate along the edges of blowouts, and highest under vegetative cover. Sex ratios were consistently female-biased, and the degree of bias increased from spring through fall in both years. These increases were more pronounced in 1998, with the sex ratios of early and late sampling periods differing by over four-fold, from 1 )15 to 5)00. Females were heavier than males, and the dry masses of both sexes increased from spring though fall in both years. As with sex ratios, these increases were more pronounced in 1998. Only the largest beetles survived the drought, illustrating how extreme conditions can exert especially strong directional selection, and affect sex ratios in populations with sexual size dimorphism. ( 2001 Academic Press Keywords: abundance; body mass; climate; grassland; distribution; Eleodes extricata; sand shinnery oak; sex ratios Introduction Darkling beetles (Tenebrionidae) of the genus Eleodes are common and conspicuous insects of arid and semi-arid ecosystems. About 140 species live in North America, distributed mainly throughout the western United States and northern Mexico (Tanner, 1961). Adults are typically large, black, flightless, long-lived, highly mobile, and can be important generalist consumers, feeding on dead and living vegetation, seeds, and soft-bodied insects (Allsopp, 1980; Rogers et al., 1988). Their numbers tend to be stable across years (Calkins & Kirk, 1975; Wise, 1981a; Sheldon & Rogers, 1984), but pronounced changes in precipitation and temperature can make for exceptions (Ahearn, 1971; Thomas, 1979; Sheldon & Rogers, 1984). The El Nin 8 o of 1997–1998 has been described as the climate event of the century (Changnon, 2000). In the region encompassing our study sites, it resulted in 1997 being unusually wet and mild, and 1998 being unusually dry and hot. These extreme but 0140-1963/01/120711#11 $35.00/0 ( 2001 Academic Press
Transcript of A darkling beetle population in West Texas during the 1997–1998 El Niño

Journal of Arid Environments (2001) 49: 711}721doi:10.1006/jare.2001.0821, available online at http://www.idealibrary.com on
A darkling beetle population in West Texas duringthe 1997+1998 El Nin8 o
Richard J. Deslippe, Jesus R. Salazar & Yu-Jie Guo
Department of Biological Sciences, Texas Tech University, Lubbock,TX 79409-3131 U.S.A.
(Received 16 October 2000, accepted 14 March 2001, published electronically 3 September 2001)
We examined the abundance and distribution of Eleodes extricata (Teneb-rionidae) in a semi-arid region of West Texas. During the 2-year study,climatic conditions were extreme but opposite, with 1997 being unusually wetand mild, and 1998 being unusually dry and hot. These conditions wereassociated with striking differences in the population structure of thebeetles. Under identical pitfall-sampling efforts, we captured 1562 and671 individuals in 1997 and 1998, respectively. Capture rates were lowest ingrassland, and highest in sand shinnery oak, and within the latter, they werelowest in blowouts, intermediate along the edges of blowouts, and highestunder vegetative cover. Sex ratios were consistently female-biased, and thedegree of bias increased from spring through fall in both years. These increaseswere more pronounced in 1998, with the sex ratios of early and late samplingperiods differing by over four-fold, from 1)15 to 5)00. Females wereheavier than males, and the dry masses of both sexes increased from springthough fall in both years. As with sex ratios, these increases were morepronounced in 1998. Only the largest beetles survived the drought, illustratinghow extreme conditions can exert especially strong directional selection, andaffect sex ratios in populations with sexual size dimorphism.
( 2001 Academic Press
Keywords: abundance; body mass; climate; grassland; distribution; Eleodesextricata; sand shinnery oak; sex ratios
Introduction
Darkling beetles (Tenebrionidae) of the genus Eleodes are common and conspicuousinsects of arid and semi-arid ecosystems. About 140 species live in North America,distributed mainly throughout the western United States and northern Mexico (Tanner,1961). Adults are typically large, black, flightless, long-lived, highly mobile, and can beimportant generalist consumers, feeding on dead and living vegetation, seeds, andsoft-bodied insects (Allsopp, 1980; Rogers et al., 1988). Their numbers tend to be stableacross years (Calkins & Kirk, 1975; Wise, 1981a; Sheldon & Rogers, 1984), butpronounced changes in precipitation and temperature can make for exceptions (Ahearn,1971; Thomas, 1979; Sheldon & Rogers, 1984).
The El Nin8 o of 1997–1998 has been described as the climate event of the century(Changnon, 2000). In the region encompassing our study sites, it resulted in 1997 beingunusually wet and mild, and 1998 being unusually dry and hot. These extreme but
0140-1963/01/120711#11 $35.00/0 ( 2001 Academic Press

712 R. J. DESLIPPE ET AL.
opposite conditions afforded a rare context in which to examine the influence ofclimate on darkling beetles. We here report on the spatial and temporal variations in theabundance and distribution of a locally common but little studied beetle, Eleodesextricata. We also compare sex ratios during the El Nin8 o period to determine whetherthey changed within and between years, as one might expect when survivorship dif-fers between the sexes.
Materials and methods
Study area
We trapped E. extricata over 2 years in a semi-arid region of West Texas where thelandscape is a mosaic of agricultural land, grassland, and sand shinnery oak. The farmedlands are used primarily to cultivate cotton, and the grasslands are mainly convertedagricultural lands of the Conservation Reserve Program (CRP). Sand shinnery oak isnative habitat that spanned some 2)3 million ha in North America before becoming thetarget of extensive eradication efforts (Fig. 1; Dillion et al., 1994). It is dominatedby Quercus havardii, a clonal deciduous shrub rarely exceeding a meter in height (Mayeset al., 1998).
Three study sites were established at 1050 m elevation in CRP grassland and sandshinnery oak. They were located within a 5-km radius in Yoakum County, about 40 kmfrom the eastern border of New Mexico (Fig. 1). The grassland site contained abundantgrowth of blue grama (Bouteloua gracilis), side-oats grama (B. curtipendula) and old-world bluestem (Bothriochloa ischaemum) in compacted clay loam (Fig. 2). The shinneryoak sites were either occasionally or never assessable to cattle, and we refer to them as
Figure 1. Map showing the past distribution of sand shinnery oak ( ), and the location of studysite in Yoakum County (33325@N; 102350@W) .

Figure 2. Sharp transitions occur between the grassland (a), [foreground] and sand shinnery oakhabitats. The latter contains many open wind-swept blowouts, like the one featured in (b).
DARKLING BEETLES DURING AN EL NIN3 O 713
grazed and ungrazed, respectively. In contrast to the hard soil of the grassland, thesesites had deep sandy soil overlying sandy clay loam. They featured dense patches ofQ. havardii interrupted by wind-swept areas devoid of vegetation (blowouts) (Fig. 2).
Pitfall trapping
Two sampling regimes were used to collect E. extricata with pitfall traps. First, weestablished traps in grassland, and in grazed and ungrazed sand shinnery oak to assess

714 R. J. DESLIPPE ET AL.
habitat use at a broad scale. Traps were set for 1 week in May, July and September of1997 and 1998, and were separated by 10 m in a 7]5 array. Second, we establishedtraps in ungrazed sand shinnery oak to assess habitat use at a narrower scale. Traps wereset for 2 weeks in June, August and October of 1997 and 1998, and for each of 20blowouts, a single trap was placed in the center and edge of the blowout, and 5 m fromthe blowout in vegetation. In both regimes, the traps were circular plastic containers(8 cm in diameter; 11cm deep), they were buried flush with the ground surface, andwere filled with a mixture of ethylene glycol and water. Of the 990 traps, 18 weredestroyed by mammals or became filled with blowing sand, and were thus excludedfrom analyses.
Laboratory procedures
All E. extricata captures were transferred to vials and stored in 70% ethanol. They werelater enumerated, sexed, oven-dried at 1403C for 24 h, and weighed. Abundancesacross habitats were analysed with descriptive statistics at the broader scale, and withKruskal}Wallis tests at the narrower scale. As the sampling designs and locationsalternated each month, we standardized comparisons of both sex ratios and bodymasses by pooling results over 2-month intervals. Sex ratios were analysed withlog-likelihood tests, and body masses were analysed with ANOVAs. For all statisticaltests, the statistical software program used was SAS (1998). Temperature and precipi-tation records were obtained from a weather station located within 35 km of the fieldsites.
Results
The 1997–1998 El Nin8 o affected climate considerably in the study region, as 1997was atypically wet and cool, and 1998 was atypically dry and hot (Fig. 3). Totalprecipitation and mean temperature in 1997 were 5)3 cm greater and 0)43C lower thanthe 40-year mean. The area received 40)7 cm of rain from April to September, anamount accounting for 77)3% of total precipitation for the year. In sharp contrast to1997, annual precipitation and mean temperature in 1998 were 10)1 cm lower and 1)53Cgreater than the 40-year mean. Only 16)7 cm of rain fell in the area from April toSeptember, 45)0% of the meager total precipitation for 1998, and 59)0% less than thesame period of the previous year.
Abundance of E. extricata differed dramatically both yearly and seasonally(Figs 4 & 5). Under identical sampling efforts, we captured 1562 individuals in1997 and 671 individuals in 1998. Of the 2233 beetles, only 104 (4)7%) were trapped inMay, and two (0)1%) were trapped in October. In 1997, captures were high from Junethrough September, with numbers peaking in July and August. In contrast, captures in1998 were low in all months except June.
Abundance of E. extricata also differed substantially among habitats at both scalesexamined (Figs 4 & 5). At the broader scale, overall captures were fewest in thegrassland site [(22/817)*100"2)7%], intermediate in the ungrazed sand shinneryoak site [(273/817)*100"33)4%], and greatest in the grazed sand shinnery oak site[(522/817)*100"63)9%]. This ranking pattern was identical in every samplingperiod (Fig. 4). At the narrower scale, overall captures were fewest in blowouts[(154/1416)*100"10)9%], intermediate along the edges of blowouts [(576/1416)*100"40)7%], and greatest in sand shinnery oak [(686/1416)*100"48)4%]. However, thisoverall pattern was not consistent between years, for in 1998, unlike in 1997, there weremore captures along edges of blowouts than in sand shinnery oak (Fig. 5). Capture ratesacross the habitats differed significantly in some months (June 1997: H"29)9,

Figure 3. Total annual precipitation and mean annual temperature during the El Nin8 o periodcompared to the 40-year mean. Whereas 1997 was unusually wet and mild, 1998 was unusuallydry and hot.
DARKLING BEETLES DURING AN EL NIN3 O 715
p(0)0001; August 1997: H"31)7, p(0)0001; June 1998: H"9)7, p(0)008), but notin others (October 1997: H"3)7, p"0)160; August 1998: H"5)3, p"0)072; Octo-ber 1998: H"0)0, p"1)000).
Sex ratios were consistently female-biased, and the degree of bias increased fromspring through fall in both years. These increases were more pronounced in 1998, withthe sex ratios of early and late sampling periods differing by over four fold, from1)15 to 5)00 (Fig. 6). Sex ratios differed significantly from 1 : 1 in some samplingperiods, but not in others both in 1997 (May–June: G"6)1; p"0)014; July–August:G"53)9; p(0)0001; September–October: G"21)6; p(0)0001) and in 1998(May–June: G"1)1; p"0)302; July–August: G"3)62; p"0)057; September–October: G"6)0; p"0)014).
Females were also heavier than males, and the mean dry masses of both sexesincreased from spring through fall in both years. As with sex ratios, these increases

Figure 4. Abundance of Eleodes extricata in pitfall traps set across habitats in May, July andSeptember of (a) 1998 and (b) 1997. Bars are standard errors, and numbers above bars aresample sizes. Grassland ( ); sand shinnery oak-grazed ( ); sand shinnery oak-ungrazed ( ).
716 R. J. DESLIPPE ET AL.
were more pronounced in 1998 (Fig. 7). In a three-factor ANOVA, sex, month andyear, and the interactions of sex by month, and month by year were each significant(Table 1).
Discussion
Climatic extremes may impact populations for better or worse (Grant, 1986; Langley& Shure, 1988; Wright et al., 1999; Anderson, 2000), and they often have an especiallystrong influence in biomes characterized by physically harsh conditions like deserts andtundra (Lima et al., 1999; Sala et al., 2000). In this study, climatic extremes ina semi-arid environment were associated with striking differences between years inthe population structure of E. extricata. The wet and mild year was highly favorable,whereas the dry and hot year was highly unfavorable for the beetles. Either reducedactivity or reduced survivorship of individuals may explain the fewer captures in thedrought year. We favor the latter explanation as the main reason, for seasonal changes insex ratios and body mass were particularly pronounced in the drought year, and thepatterns were consistent with differential survivorship based on body size. Further,one might expect increased not reduced activity by the beetles in the drought year due tofood deprivation (McIntyre & Vaughn, 1997).

Figure 5. Abundance of Eleodes extricata in pitfall traps set about blowouts in June, August andOctober of (a) 1998 and (b) 1997. Bars are standard errors, and numbers above bars are samplesizes. Blowout ( ); edge of blowout ( ); sand shinnery oak ( ).
DARKLING BEETLES DURING AN EL NIN3 O 717
The drought led to reduced plant cover, as plant growth was severely restricted. Eventhe normally robust Q. havardii withered and, in some places, died. Consequently, thedistinction between sites of vegetative cover and edges of blowouts within the sandshinnery oak habitat diminished. At the broader scale, we regularly detected a strongassociation of the beetles with habitat type, a result consistent with patterns of habitat useof E. extricata in Colorado (Stapp, 1997) and Wyoming (Parmenter et al., 1989a).Whicker & Tracy (1987) determined that the highest body temperatures of sevenEleodes spp. were maintained by E. extricata. Similarly, Parmenter et al. (1989b) in theirstudy of four Eleodes spp. found that E. extricata was the one most frequently en-countered in sunny, sparsely vegetated areas, and it was the species with the greatesttolerance of high temperature. A preference of the beetles for warmer sites may explainwhy we captured them more frequently in grazed versus ungrazed sand shinnery oak, asthe former habitat has less plant cover. At the narrower scale, we also regularly detecteda strong association with habitat, as E. extricata was trapped significantly more oftenboth in vegetative cover and along edges of blowouts than in the blowouts themselves.However, patterns of captures differed between years in vegetative cover and alongedges of blowouts, reflecting the effect of the drought on Q. havardii.
The most intriguing results of this study were the clear seasonal and yearly differ-ences in sex ratios and body masses. As conditions became both drier and warmer, sexratios became increasingly female-biased, and the mean body masses of both males and

Figure 6. Sex ratios from spring through fall during the El Nin8 o period (a, 1998; b, 1997).Numbers within bars indicate sample sizes.
718 R. J. DESLIPPE ET AL.
females increased. Because these patterns were consistent between years, and wereespecially pronounced during the drought, they indicate that the smallest individuals inthe population did not survive the stress. The larvae presumably completed theirdevelopment, as adults were similarly abundant in the spring of each year. However,they were apparently severely food deprived, as the majority of both sexes wereextremely small in the spring of 1998. With a notable exception (Wise, 1981a, b), therehas been little attempt to determine sex ratios in ecological investigations of darklingbeetles. In sharp contrast to our results, Wise (1981a, b) determined that sex ratios wereclearly male biased in E. obscurus, and were about 1 : 1 in E. fusiformis, E. sponsus andE. tricostatus.
Our study provides an example of how a drought may exert directional selection onbody size and promote mortality. The drastic reductions in the size and abundance ofadults likely resulted in fewer eggs being laid which likely led, in turn, to a loweredabundance of the soil-dwelling larvae. As larvae damage some growing plants and
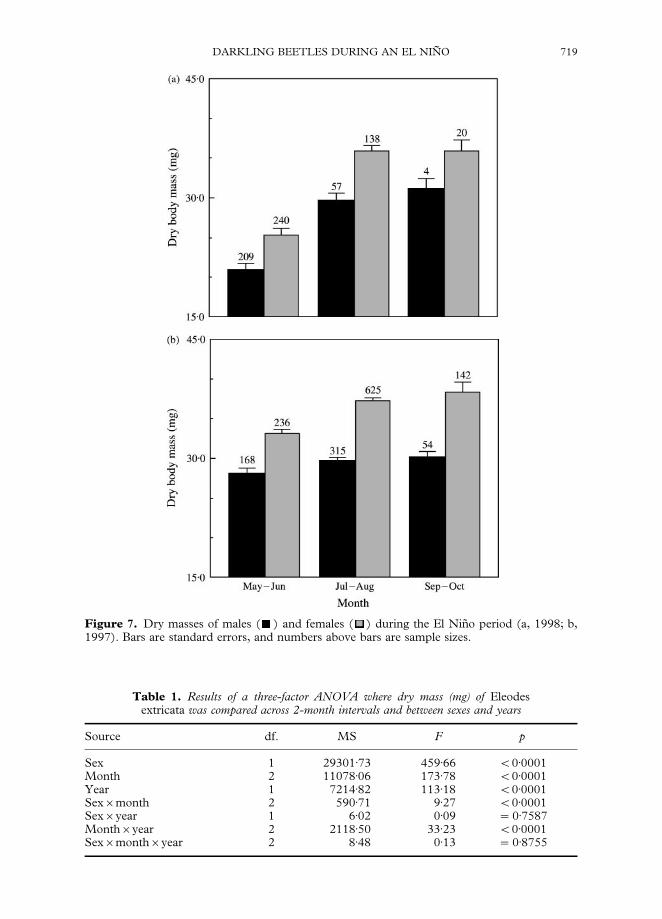
Figure 7. Dry masses of males ( ) and females ( ) during the El Nin8 o period (a, 1998; b,1997). Bars are standard errors, and numbers above bars are sample sizes.
Table 1. Results of a three-factor ANOVA where dry mass (mg) of Eleodesextricata was compared across 2-month intervals and between sexes and years
Source df. MS F p
Sex 1 29301)73 459)66 (0)0001Month 2 11078)06 173)78 (0)0001Year 1 7214)82 113)18 (0)0001Sex]month 2 590)71 9)27 (0)0001Sex]year 1 6)02 0)09 "0)7587Month]year 2 2118)50 33)23 (0)0001Sex]month]year 2 8)48 0)13 "0)8755
DARKLING BEETLES DURING AN EL NIN3 O 719

720 R. J. DESLIPPE ET AL.
germinating seeds but not others (Allsopp, 1980), their reduction probably impactsplant community structure and ecosystem recovery. The extent of their impact isunclear, however, for they have been poorly studied especially compared to adults andother detritivores of arid and semi-arid environments (Crawford, 1979).
Our study also highlights that differential survivorship associated with body sizecan affect sex ratios in populations with sexual size dimorphism. Climatic extremesdo not always affect sex ratios of populations significantly (Smith, 1996; Brown& Brown, 1998), but, like our study, sometimes they do (Moran & Hurd, 1994; Asakura1995; Van Horne et al. 1997), reinforcing the notion that sex ratios can evolve in variableenvironments through natural selection independent of maternal strategies of sex alloca-tion. Short-term survival through transient, acute stress can be critical in shapingpopulations for many subsequent generations.
We thank Mike Abel, Richard Cardenas, Andrew Moore and Leahann Peterson for field help, andthank Julio Ferrer and Richard Fagerlund for identifying the beetles. The research was supportedin part by a Howard Hughes Medical Institute grant through the Undergraduate BiologicalSciences Education Program to Texas Tech University. Voucher specimens have been depositedin the Swedish Museum of Natural History.
References
Ahearn, G.A. (1971). Ecological factors affecting population sampling of desert tenebrionidbeetles. American Midland Naturalist, 86: 385–406.
Allsopp, P.G. (1980). The biology of false wireworms and their adults (soil-inhabiting Teneb-rionidae) (Coleoptera): a review. Bulletin of Entomological Research, 70: 343–379.
Anderson, O.R. (2000). Abundance of terrestrial Gymnamoebae at a northeastern U.S. site:a four-year study, including the El Nin8 o of 1997–1998. Journal of Eukaryotic Microbiology, 47:148–155.
Asakura, A. (1995). Sexual differences in life history and resource utilization by the hermitcrab. Ecology, 76: 2295–2313.
Brown, C.R. & Brown, M.B. (1998). Intense natural selection on body size and wing and tailasymmetry in cliff swallows during severe weather. Evolution, 52: 1461–1475.
Calkins, C.O. & Kirk, V.M. (1975). Distribution of false wireworms (Coleoptera: Tenebrionidae)in relation to soil texture. Environmental Entomology, 4: 373–374.
Changnon, S.A. (2000). El NinJ o 1997–1998: The Climate Event of the Century. New York, NY:Oxford University Press. 215 pp.
Crawford, C.S. (1979). Desert detritivores: a review of life history patterns and trophic roles.Journal of Arid Environments, 2: 31–42.
Dillion, S.S., McGinley, M.A., Friese, C.F. & Zak, J.C. (1994). Construction of sand shinnery oakcommunities of the Llano Estacado: animal disturbances, plant community structure, andrestoration. Restoration Ecology, 2: 51–60.
Grant, P.R. (1986). Interspecific competition in fluctuating environments. In: Diamond, J.& Case, T.J. (Eds), Community Ecology, pp. 173–191. New York: Harper & Row. 665 pp.
Langley, A.K. & Shure Jr, D.J. (1988). The impact of climatic extremes on cotton rat (Sigmodonhispidus) populations. American Midland Naturalist, 120: 136–143.
Lima, M., Marquet, P.A. & Jaksic, F.M. (1999). El Nin8 o events, precipitation patterns, and rodentoutbreaks are statistically associated in semiarid Chile. Ecography, 22: 213–218.
McIntyre, N.E. & Vaughn, T.T. (1997). Effects of food deprivation and olfactory and visualcues on movement patterns of two Eleodes species (Coleoptera: Tenebrionidae) in a windtunnel. Annals of the Entomological Society of America, 90: 260–265.
Mayes, S.G., McGinley, M.A. & Werth, C.R. (1998). Clonal population structure and geneticvariation in sand-shinnery oak, Quercus havardii (Fagaceae). American Journal of Botany, 85:1609–1617.
Moran, M.D. & Hurd, L.E. (1994). Environmentally determined male-biased sex ratio ina praying mantid. American Midland Naturalist, 132: 205–208.

DARKLING BEETLES DURING AN EL NIN3 O 721
Parmenter, R.R., Parmenter, C.A. & Cheney, C.D. (1989a). Factors influencing microhabitatpartitioning among coexisting species of arid-land darkling beetles (Tenebrionidae): Behavioralresponses to vegetation architecture. The Southwestern Naturalist, 34: 319–329.
Parmenter, R.R., Parmenter, C.A. & Cheney, C.D. (1989b). Factors influencing microhabitatpartitioning in arid-land darkling beetles (Tenebrionidae): temperature and water conservation.Journal of Arid Environments, 17: 57–67.
Rogers, L.E., Woodley, N.E., Sheldon, J.K. & Beedlow, P.A. (1988). Diets of darkling beetles(Coleoptera: Tenebrionidae) within a shrub-steppe ecosystem. Annals of the EntomologicalSociety of America, 81: 782–791.
Sala, O.E., Chapin III, F.S., Armesto, J.J., Berlow, E., Bloomfield, J., Dirzo, R., Huber-Sanwald,E., Huenneke, L.F., Jackson, R.B., Kinzig, A., Leemans, R., Lodge, D.M., Mooney, H.A.,Oesterheld, M., Poff, N.L., Sykes, M.T., Walker, B.H., Walker, M. & Wall, D.H. (2000).Global biodiversity scenarios for the year 2100. Science, 287: 1770–1774.
SAS Institute. (1998). SAS/STAT User’s Guide, Version 7.0. SAS Institute, Cary, NC.Sheldon, J.K. & Rogers, L.E. (1984). Seasonal and habitat distribution of tenebrionid beetles in
shrub-steppe communities of the Hanford site in eastern Washington. Environmental Entomo-logy, 13: 214–220.
Smith, G.R. (1996). Annual life-history variation in the striped plateau lizard, Sceloporus virgatus.Canadian Journal of Zoology, 74: 2025–2030.
Stapp, P. (1997). Microhabitat use and community structure of darkling beetles (Coleoptera:Tenebrionidae) in shortgrass prairie: effects of season, shrub cover and soil type. AmericanMidland Naturalist, 137: 298–311.
Tanner, V.M. (1961). A checklist of the species of Eleodes and descriptions of new species(Coleoptera: Tenebrionidae). The Great Basin Naturalist, 21: 55–78.
Thomas, D.B. (1979). Patterns in the abundance of some tenebrionid beetles in the MojaveDesert. Environmental Entomology, 8: 568–574.
Van Horne, B., Olson, G.S., Schooley, R.L., Corn, J.G. & Burnham, K.P. (1997). Effects ofdrought and prolonged winter on Townsend’s ground squirrel demography in shrubsteppehabitats. Ecological Monographs, 67: 295–315.
Whicker, A.D. & Tracy, C.R. (1987). Tenebrionid beetles in the shortgrass prairie: daily andseasonal patterns of activity and temperature. Ecological Entomology, 12: 97–108.
Wise, D.H. (1981a). Seasonal and yearly patterns in the densities of darkling beetles (Coleoptera:Tenebrionidae) in a montane community. Environmental Entomology, 10: 350–358.
Wise, D.H. (1981b). A removal experiment with darkling beetles: lack of evidence for interspecificcompetition. Ecology, 62: 727–738.
Wright, S.J., Carrasco, C., Calderon, O. & Paton, S. (1999). The El Nin8 o–Southern Oscillation,variable fruit production, and famine in a tropical forest. Ecology, 80: 1632–1647.




















