
Duration of breastfeeding is associated with leptin (LEP) DNA ...
Upload
independentCategory
view
2download
0
A cutaneous gene therapy approach to human leptindeficiencies: correction of the murine ob/obphenotype using leptin-targeted keratinocyte grafts
FERNANDO LARCHER,*,†,1,2 MARCELA DEL RIO,*,†,1 FERNANDO SERRANO,‡
JOSE CARLOS SEGOVIA,*,† ANGEL RAMIREZ,* ALVARO MEANA,§
ANGUSTIAS PAGE,* JOSE LUIS ABAD,‡ MANUEL A. GONZALEZ,‡
JUAN BUEREN,*,† ANTONIO BERNAD,‡ AND JOSE LUIS JORCANO*,†
*Project of Cell and Molecular Biology and Gene Therapy. CIEMAT. Avenida Complutense 22,28040 Madrid, Spain; †Fundacion Marcelino Botın for Gene Therapy, ‡Department of Immunologyand Oncology, Centro Nacional de Biotecnologıa, Madrid, Spain; and §Centro de Transfusiones delPrincipado de Asturias, Oviedo, Spain
ABSTRACT Leptin deficiency produces a phenotypeof obesity, diabetes, and infertility in the ob/ob mouse.In humans, leptin deficiency occurs in some cases ofcongenital obesity and in lipodystrophic disorders char-acterized by reduced adipose tissue and insulin resis-tance. Cutaneous gene therapy is considered an attrac-tive potential method to correct circulating proteindeficiencies, since gene-transferred human keratino-cytes can produce and secrete gene products withsystemic action. However, no studies showing correc-tion of a systemic defect have been reported. We reportthe successful correction of leptin deficiency usingcutaneous gene therapy in the ob/ob mouse model. As afeasibility approach, skin explants from transgenicmice overexpressing leptin were grafted on immunode-ficient ob/ob mice. One month later, recipient micereached body weight values of lean animals. Otherbiochemical and clinical parameters were also normal-ized. In a second human gene therapy approach, aretroviral vector encoding both leptin and EGFPcDNAs was used to transduce HK and, epithelial graftsenriched in high leptin-producing HK were trans-planted to immunosuppressed ob/ob mice. HK-derivedleptin induced body weight reduction after a drop inblood glucose and food intake. Leptin replacementthrough genetically engineered HK grafts provides avaluable therapeutic alternative for permanent treat-ment of human leptin deficiency conditions.—Larcher,F., Del Rio, M., Serrano, F., Segovia, J. C., Ramırez, A.,Meana, A., Page, A., Abad, J. L., Gonzalez, M. A.,Bueren, J., Bernad, A., Jorcano, J. L. A cutaneous genetherapy approach to human leptin deficiencies: correc-tion of the murine ob/ob phenotype using leptin-tar-geted keratinocyte grafts. FASEB J. 15, 1529–1538 (2001)
Key Words: skin z lipodistrophy z diabetes z leptin replacement
Leptin, the 16 kDa product of the OB gene, is anessential hormone secreted mainly by adipocytes thatregulates food intake and neuroendocrine function via
specific receptors in the hypothalamus (1, 2). Experi-mental evidence suggests that leptin may also exertdirect effects at the level of gene expression or cellularfunction on peripheral tissues including hematopoieticcells, T cells, the endocrine pancreas, pituitary, ovary,adipocytes, skeletal muscle, and hepatocytes (3, 4). Inaddition, a role for leptin has recently been reported inangiogenesis and bone formation (5–7).
Congenital leptin deficiency has been identified inhumans and is associated with a rare, severe early-onsetobesity (8, 9). Recombinant leptin therapy is shown tolead to a sustained reduction in body weight in one ofthese patients, predominantly as a result of fat loss (10).
Decreased circulating leptin levels are also reportedin patients with different forms of generalized lipoatro-phic diabetes (7). Lipoatrophic diabetes designates agroup of syndromes characterized by extreme paucityof adipose tissue, hyperglycemia, severe insulin resis-tance, hypoleptinemia, and voracious appetite (11, 12).Other serious clinical features in the congenital form ofthe disease include an anabolic syndrome with organo-megaly and hypertrophic cardiomyopathy. Since nocausal therapy has yet been identified, a prophylacticapproach is used in these patients based on a rigidcontrol of calorie consumption and anabolic syndromein order to delay diabetic complications and progres-sion of hypertrophic cardiomyopathy (11).
Leptin administration has recently been shown toincrease glucose metabolism and restore insulin sensi-tivity in two transgenic mouse models of congenitalgeneralized lipodystrophy (13–16), suggesting a criticalrole for leptin in the physiopathology of these disordersand its therapeutic potential for the treatment ofhuman lipoatrophic diabetes.
Protein injection is frequently used to treat peptidehormone deficiencies. Although leptin has been deliv-
1 These authors contributed equally to these studies.2 Correspondence: CIEMAT, Avenida Complutense 22 Edifi-
cio 7, 28040 Madrid, Spain. E-mail: [email protected]
15290892-6638/01/0015-1529 © FASEB

ered by this route with good results in a patientsuffering from leptin-deficient inherited obesity (10),the treatment may be compromised by its short dura-tion of action and the need for repetitive daily dosing.Gene therapy is thus an attractive alternative approachfor leptin delivery and has received recent experimen-tal support (17, 19)
On the basis of their proliferative capacity and theproven value of cultured keratinocytes for skin woundcoverage, autologous grafts of genetically engineeredkeratinocytes have been proposed as a suitable vehicleto correct for deficiencies in circulating proteins (20).Previous studies have shown that the product of exog-enous genes such as GH, factor IX, interleukin 6 (IL-6),etc., reach the blood circulation after being synthe-sized, processed, and secreted by gene-transferred hu-man epidermal cells (21–24). In addition to these exvivo approaches, transgenic mouse studies stronglysupport the concept of the epidermis as an efficientsecretory bioreactor (25–27).
A further boost to cutaneous gene therapy for sys-temic disorders has recently been provided. Partialcorrection of hemophilia A in factor VIII-deficient micewas achieved after grafting of factor VIII-expressingmouse skin from a loricrin factor VIII transgenic mouse(27). Injection of plasmid DNA coding for IL-10 intorat skin also led to transient inhibition of contacthypersensitivity at a distant area of the skin (28). To ourknowledge, however, use of genetically modified hu-man keratinocytes to correct for a systemic defect hasnot been described to date.
Here we report the successful correction of a leptindeficiency-derived metabolic disorder through cutaneousgene therapy using both murine and human keratino-cytes. For this study, we used the obese ob/ob mutantmouse (29, 30) as a model of leptin deficiency. In theseanimals, the absence of leptin accounts for a dramaticphenotype characterized by extreme obesity due to in-creased food intake and low energy expenditure. Otherclinical features include decreased immune function,infertility, and impaired wound healing. At the biochem-ical level, the ob/ob mice present hyperglycemia, hyperin-sulinemia, and reduced levels of gonadotrophins (30).
Skin grafts from a leptin-overexpressing skin transgenicmouse proved highly efficient for the correction of theob/ob mouse systemic defect. This result led us to thedevelopment of a human cutaneous gene therapy-rele-vant approach. Thus, ex vivo leptin-transduced humankeratinocyte (HK) grafts comprising less than 10% of thebody surface corrected the leptin deficiency of ob/obmouse. This is also the first study demonstrating success-ful replacement of a missing circulating protein usinggenetically modified human keratinocyte grafts.
MATERIALS AND METHODS
Animals
BALB/c CB17-Prkdcscid and C57BL/6Job/ob mice were pur-chased from IFFA-Credo-Charles River (St. Aulin-les-Elbeuf,
France) and maintained at the CIEMAT Laboratory AnimalsFacility (Spanish registration number 28079–21 A). Animalswere housed in individually ventilated type II cages, maxi-mum 4 mice each with 25 air changes per hour and 10 KGygamma irradiated soft wood pellets as bedding. All handlingwas done under sterile conditions. All experimental proce-dures were carried out according to European and Spanishlaws and regulations (European convention 123 about the useand protection of vertebrate mammals used in experimenta-tion and other scientific purposes. Spanish R.D 223/88 andO.M. 13–10-89 of the Ministry of Agricultural, Food andFisheries about the protection and use of animals in scientificresearch and internal biosafety and bioethics guidelines).
Generation of K5-leptin transgenic mice
A SmaI/NotI fragment from plasmid pOB1 containing themouse leptin cDNA (a generous gift of Dr. M. Reitman,NIDDK, NIH) was blunted and inserted into a cassettecontaining the 5.2 kb bovine regulatory sequences, the 59rabbit b-globin intron 2, and the 39 polyadenylation se-quences (31, 32). The construct was designated pK5-leptin.The transgene was excised from the plasmid pK5-leptin withNotI, purified by low melting point agarose electrophoresisand Elutip columns (Schleicher and Schuell, Keene, NH),adjusted to a final concentration of ;2 mg/ml, and microin-jected into (C57BL/6J3DBA/2)F2 mouse embryos as de-scribed (31, 32). Founder mice were identified by Southernblot analysis of tail DNA using the 5.2 kb fragment corre-sponding to the bovine keratin K5 promoter as a probe. Adetailed description of the characterization of K5-leptin trans-genic mice will be reported elsewhere (F. Larcher et al.,unpublished results).
Generation of graft recipient immunodeficient ob/ob mice
Fertile heterozygous ob/1 males were first mated with ho-mozygous severe combined immunodeficiency SCID/SCIDfemale mice (BALB/c CB-17 strain) to obtain double het-erozygous SCID/1, ob/1 animals. ob/1 mice were identifiedusing the one-step PCR-based genotyping method describedby Namae et al. (33) Double heterozygous SCID/1, ob/1males were crossed with female homozygous SCID/SCIDmice to obtain immunodeficient ob/1 breeding pairs. Im-munodeficiency was assessed by the absence of B and Tlymphocytes in blood by FACS analysis using FITC- and PEfluorochrome-conjugated antibodies specific for murine Bcells (anti-B220; Caltag, Burlingame, CA) and T cells (anti-CD3; BD-PharMingen, San Diego, CA), respectively, as de-scribed (34). Finally, male and female immunodeficientheterozygous ob/1 mice pairs were bred to obtain immuno-deficient ob/ob graft recipient obese mice, which developed atnormal mendelian rates. A similar breeding strategy wasattempted to obtain immunodeficient ob/ob mice on a nu/nubackground with low efficiency.
Mouse skin grafts
Dorsal full-thickness skin pieces of 2–2.5 cm2 were obtainedfrom 5- to 7-wk-old K5-leptin transgenic mice or controllittermates. Shaved donor skin pieces were grafted onto awound created by removing a similar-sized piece of full-thickness back skin in female immunodeficient obese ob/obrecipient mice. Graft and host skin edges were joined usingsurgical silk suture and the grafted area was covered with athin layer of NewSkin (Medtech, Jackson, WY) as the onlyprotective dressing. This procedure allows graft-take monitor-ing and produced normal-haired donor skin.
1530 Vol. 15 July 2001 LARCHER ET AL.The FASEB Journal

Keratinocyte cell cultures and retroviral gene transfer
Human primary keratinocytes were obtained from newbornforeskins by repetitive trypsin incubation, then seeded onlethally irradiated 3T3-J2 cells (a gift from Dr. J. Garlick,SUNY) as described (35). Retroviral infection was performedon small-sized cell colonies (8–16 cells) of first-passage pri-mary keratinocytes. Cells were incubated with pLZRS-leptin-IRES-EGFP or pLZRS-IRES-EGFP vector supernatants at atiter of 1 to 5 3 106 CFU/ml for 4 h in the presence ofpolybrene (8 mg/ml) on 2 consecutive days. Keratinocytes at60–80% confluence were trypsinized, resuspended inPBS/2% FCS, analyzed for EGFP expression and sorted byfluorescence-activated cell sorting on a FACStar PLUS flowcytometer (Becton Dickinson, San Jose, CA) as described(36). Transduced, EGFP-sorted keratinocytes were seeded onlive human fibroblast-containing fibrin gels at a density of 1 3104 cells/cm2. Fibrin-fibroblast gels were prepared accordingto Del Rio et al. (36).
Mouse primary keratinocyte cultures from newborn K5-leptin transgenic animals or control littermates were obtainedas described (32).
Human keratinocyte grafts and immunosuppressionof ob/ob mice
After reaching confluence, transduced human keratinocytecultures on fibrin-fibroblast gels were manually detached,divided into 2–2.5 cm2 squares, and grafted onto the dorsalregion of female cyclosporin-A immunosuppressed ob/ob mice(one square/mouse) using the flap method described byBarrandon et al. (37). Leptin- and control-transduced, EGFP-sorted human keratinocyte composite cultures were graftedin five immunosuppressed mice, respectively. Mice werehoused in pathogen-free conditions for the duration of theexperiment. Grafting experiments were also performed inob/ob mice with no immunosuppressive treatment.
Human-to-mouse xenografts were maintained in cyclo-sporin-A-treated ob/ob mice according to Compton et al., withminor modifications (38). Animals received daily intraperito-neal injections of 25 mg/kg of cyclosporin-A (Sandimmum 50mg/ml, Novartis, Summit, NJ) in a sterile olive oil suspension(200 ml/mouse), starting 1 day before grafting and for theduration of the experiment (16 days).
Retroviral vector design and production
Retroviral expression vectors were constructed using theLZRS backbone vector (39, 40). To construct the pLZRS-IRES-EGFP vector, a XhoI/NotI fragment containing the IRESand the EGFP gene was obtained from plasmid pIRES2-EGFP(Clontech, Palo Alto, CA) and cloned into the XhoI/NotI sitesof plasmid pL3 to capture a BamHI site. The resultingconstruct was digested with BamHI and NotI, and the frag-ment of interest cloned into the BamHI/NotI sites of theretroviral vector backbone pLZRS-CMV-EGFP (kindly pro-vided by Dr. G.P. Nolan, Stanford University, CA). To con-struct the pLZRS-leptin-IRES-EGFP vector, a BamHI/EcoRIfragment containing the mouse leptin cDNA obtained fromplasmid pK5-leptin was cloned into the BamHI/EcoRI sites ofpLZRS-IRES-EGFP vector plasmid. Defective retroviruseswere generated through transient transfection of 293-T cellswith packaging and retroviral vector plasmids according toYang et al. (40).
Glucose and leptin measurements
Blood samples were collected by tail incision. Blood glucoselevels were determined between 9 and 10 am on nonfasted
mice using a Gluco-touch digital monitor (Lifescan, Johnson& Johnson, Burnaby, B.C., Canada). Both serum and culturedkeratinocyte-derived mouse leptin were determined using theQuantikine murine leptin ELISA kit (R&D Systems, Abing-don, Oxon, UK) following the manufacturer’s instructionswith minor modifications. Serum samples were assayed at1:5–1:10 dilutions.
Animal weight and food intake
Body weight was measured every other day at the same timeof day. For feeding studies, mice were placed in individualcages and acclimated to autoclaved standard rodent chowdiet (A04, Panlab, Barcelona, Spain). A preweighed amountof diet was given per animal, and the difference in foodweight after each time period provided a measure of foodconsumption.
Histology
Human keratinocyte graft recipient animals were killed at day16 postgrafting. The graft-containing flap area was excisedand tissue samples were formalin-fixed/paraffin-embeddedfor routine hematoxylin/eosin staining or frozen in OCT.Human involucrin immunostaining was performed on 5 mmparaffin sections using the human-specific mouse monoclo-nal anti-involucrin antibody SY5 (Sigma, St. Louis, MO) at a1:100 dilution. The ABC elite peroxidase kit (Dako, Carpin-teria, CA) was used for antibody detection.
EGFP fluorescence was analyzed in unfixed 7 mm frozensections under the fluorescence microscope. Sections weremounted with Mowiol (Hoechst, Somerville, NJ) containingDAPI to stain nuclei.
Statistical analysis
Statistical significance was assessed by ANOVA with repeatedmeasures analysis and Student’s t test. All values are expressedas the mean 6 se.
RESULTS
Correction of the ob/ob phenotype using skin graftsfrom K5-leptin transgenic mice
To test the feasibility of using skin grafts containinggenetically manipulated keratinocytes to correct theob/ob mouse defect, we first developed transgenic miceoverexpressing mouse leptin cDNA under the controlof the bovine keratin K5 gene regulatory elements. Aspreviously shown by our group and others, the 5.2 kbbovine K5 gene promoter enables high constitutivebasal layer-specific expression of a wide variety oftransgenes in the epidermis (31, 32, 41, 42). In thiscase, transgenic epidermal cells were able to efficientlyexpress, process, and secrete leptin resulting in veryhigh serum leptin levels (105613 ng/ml), leading to amore marked lean phenotype than that of the trans-genic skinny mouse described by Ogawa et al. (43). Adetailed description of K5-leptin mice will be reportedelsewhere (F. Larcher et al., unpublished results).Transplantation of 2–2.5 cm2 shaved back skin piecesfrom K5-leptin mice to immunodeficient ob/ob mice
1531SKIN GENE THERAPY FOR LEPTIN DEFICIENCY
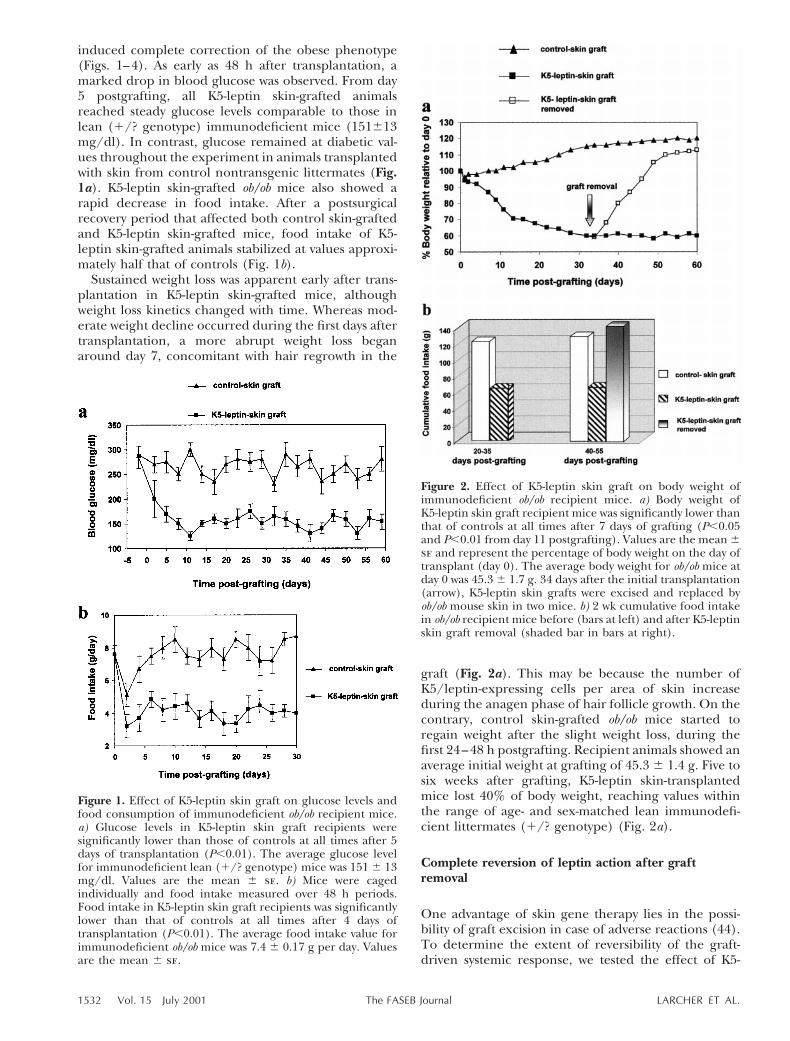
induced complete correction of the obese phenotype(Figs. 1–4). As early as 48 h after transplantation, amarked drop in blood glucose was observed. From day5 postgrafting, all K5-leptin skin-grafted animalsreached steady glucose levels comparable to those inlean (1/? genotype) immunodeficient mice (151613mg/dl). In contrast, glucose remained at diabetic val-ues throughout the experiment in animals transplantedwith skin from control nontransgenic littermates (Fig.1a). K5-leptin skin-grafted ob/ob mice also showed arapid decrease in food intake. After a postsurgicalrecovery period that affected both control skin-graftedand K5-leptin skin-grafted mice, food intake of K5-leptin skin-grafted animals stabilized at values approxi-mately half that of controls (Fig. 1b).
Sustained weight loss was apparent early after trans-plantation in K5-leptin skin-grafted mice, althoughweight loss kinetics changed with time. Whereas mod-erate weight decline occurred during the first days aftertransplantation, a more abrupt weight loss beganaround day 7, concomitant with hair regrowth in the
graft (Fig. 2a). This may be because the number ofK5/leptin-expressing cells per area of skin increaseduring the anagen phase of hair follicle growth. On thecontrary, control skin-grafted ob/ob mice started toregain weight after the slight weight loss, during thefirst 24–48 h postgrafting. Recipient animals showed anaverage initial weight at grafting of 45.3 6 1.4 g. Five tosix weeks after grafting, K5-leptin skin-transplantedmice lost 40% of body weight, reaching values withinthe range of age- and sex-matched lean immunodefi-cient littermates (1/? genotype) (Fig. 2a).
Complete reversion of leptin action after graftremoval
One advantage of skin gene therapy lies in the possi-bility of graft excision in case of adverse reactions (44).To determine the extent of reversibility of the graft-driven systemic response, we tested the effect of K5-
Figure 1. Effect of K5-leptin skin graft on glucose levels andfood consumption of immunodeficient ob/ob recipient mice.a) Glucose levels in K5-leptin skin graft recipients weresignificantly lower than those of controls at all times after 5days of transplantation (P,0.01). The average glucose levelfor immunodeficient lean (1/? genotype) mice was 151 6 13mg/dl. Values are the mean 6 se. b) Mice were cagedindividually and food intake measured over 48 h periods.Food intake in K5-leptin skin graft recipients was significantlylower than that of controls at all times after 4 days oftransplantation (P,0.01). The average food intake value forimmunodeficient ob/ob mice was 7.4 6 0.17 g per day. Valuesare the mean 6 se.
Figure 2. Effect of K5-leptin skin graft on body weight ofimmunodeficient ob/ob recipient mice. a) Body weight ofK5-leptin skin graft recipient mice was significantly lower thanthat of controls at all times after 7 days of grafting (P,0.05and P,0.01 from day 11 postgrafting). Values are the mean 6se and represent the percentage of body weight on the day oftransplant (day 0). The average body weight for ob/ob mice atday 0 was 45.3 6 1.7 g. 34 days after the initial transplantation(arrow), K5-leptin skin grafts were excised and replaced byob/ob mouse skin in two mice. b) 2 wk cumulative food intakein ob/ob recipient mice before (bars at left) and after K5-leptinskin graft removal (shaded bar in bars at right).
1532 Vol. 15 July 2001 LARCHER ET AL.The FASEB Journal

leptin skin graft removal on ‘slimmed’ immunodefi-cient ob/ob mice. The K5-leptin skin graft was replacedwith a size-matched skin piece from an ob/ob mousedonor. A sudden drop in circulating leptin after graftexcision appeared to act as a strong starvation signal,and rapid weight gain was observed concomitant withthe recovery of food intake that reached control skin-grafted immunodeficient ob/ob values (Fig. 2a, b). Incontrast, immunodeficient ob/ob animals that conservedthe K5-leptin skin graft remained within lean animalvalues (90 days after grafting, corresponding to thelatest records; data not shown).
K5-leptin skin grafts produce stable serum leptinlevels
Serum leptin was determined at different times post-grafting. K5-leptin skin-grafted mice showed valuesranging from 3.1 to 4.5 ng/ml, a range within thephysiological levels found in age-matched lean mice(Fig. 3). The presence of stable serum leptin through-out the experiment indicates that the K5 promoterremains active in the graft. Serum leptin was undetect-able in control skin-grafted mice. Thus, a contributionof the subcutaneous (s.c.) adipose tissue from thecontrol skin graft to circulating leptin appears negligi-ble. As predicted, animals that underwent K5-leptinskin graft removal showed an absence of serum leptin,as determined 5 days after graft excision (data notshown). Both a long-lasting corrective effect and areversible effect on graft removal were the hallmarks ofthis transgenic approach. A representative sequence ofexperimental steps and changes in physical appearanceis illustrated in Fig. 4.
Correction of the ob/ob phenotype through ex vivoleptin-transduced human keratinocyte grafts
Having achieved efficient systemic leptin delivery fromtransgenic mouse keratinocytes, we sought a relevant exvivo approach for human gene therapy.
Efficient leptin gene transfer into primary HKs wascarried out with high-titer replication-defective ret-roviruses obtained through transient transfection in293T cells (39, 40). The retroviral construct (Fig. 5a)contained an internal ribosome entry site (IRES) tocoexpress leptin with EGFP as a selectable marker.The same construct lacking leptin cDNA was used totransduce control keratinocytes. Use of the IRESsequence in retroviral vectors for keratinocyte target-ing has been described (45, 46). To produce popu-lations of entirely genetically engineered humankeratinocytes before grafting, transduced cells wereselected by EGFP sorting. This technique, previouslyused to select EGFP gene-transferred pig keratino-cytes (36), renders viable HK cells without affectingtheir capacity for growth and differentiation (M. Del
Figure 3. Circulating leptin levels in K5-leptin skin-graftedimmunodeficient ob/ob recipient mice. Serum was collectedfrom mice at the times indicated and leptin levels weremeasured using a commercial murine leptin ELISA kit.Values are the mean 6 se. Average serum leptin of age-matched lean mice (1/? genotype) was 3.4 6 1.6 ng/ml.Serum leptin in control skin-grafted animals was undetectableat all time points measured.
Figure 4. K5-leptin skin graft-mediated correction of obesityin immunodeficient ob/ob mice. a) Physical appearance of adonor K5-leptin transgenic mouse (right) compared to acontrol littermate (left). The dashed square represents thedonor skin piece to be grafted. b) Physical appearance of animmunodeficient ob/ob mouse immediately after transplanta-tion (day 0) with K5-leptin skin graft (left). An immunodefi-cient lean mouse (1/? genotype) is shown on the right.c) Representative body shape of a corrected immunodeficientob/ob mouse 35 days postgrafting (right). A comparisonbetween age-matched immunodeficient ob/ob (middle) andimmunocompetent C57BL/6J ob/ob mouse (left) is alsoshown.
1533SKIN GENE THERAPY FOR LEPTIN DEFICIENCY
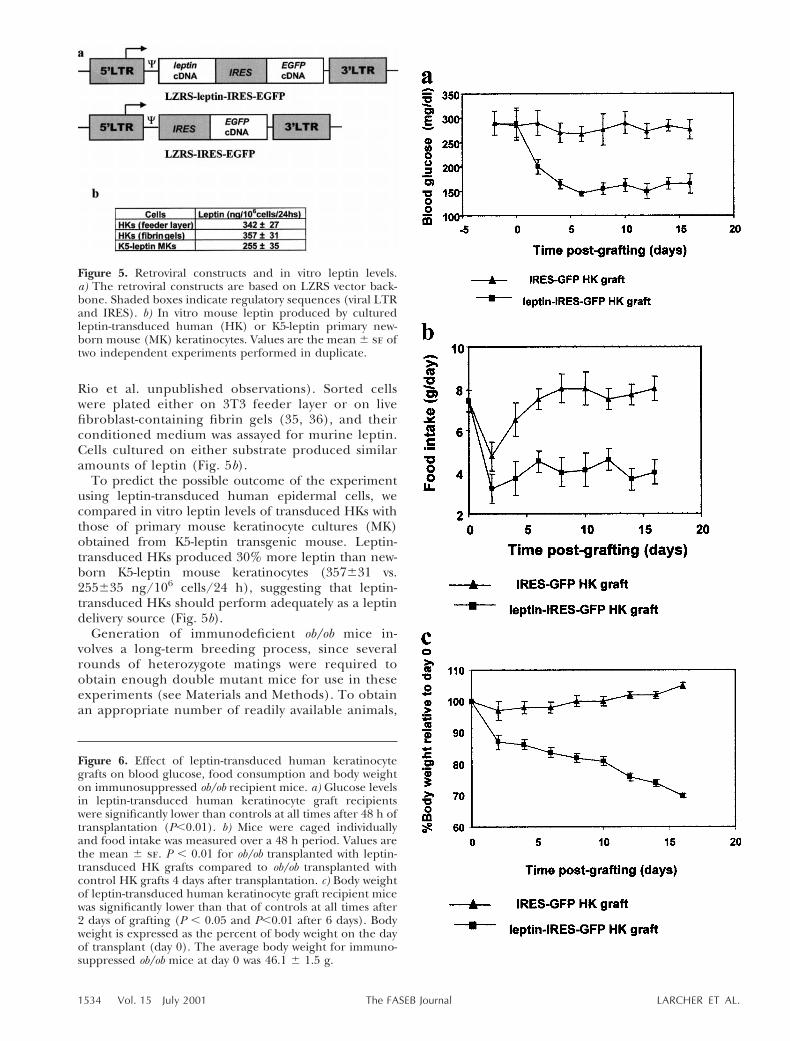
Rio et al. unpublished observations). Sorted cellswere plated either on 3T3 feeder layer or on livefibroblast-containing fibrin gels (35, 36), and theirconditioned medium was assayed for murine leptin.Cells cultured on either substrate produced similaramounts of leptin (Fig. 5b).
To predict the possible outcome of the experimentusing leptin-transduced human epidermal cells, wecompared in vitro leptin levels of transduced HKs withthose of primary mouse keratinocyte cultures (MK)obtained from K5-leptin transgenic mouse. Leptin-transduced HKs produced 30% more leptin than new-born K5-leptin mouse keratinocytes (357631 vs.255635 ng/106 cells/24 h), suggesting that leptin-transduced HKs should perform adequately as a leptindelivery source (Fig. 5b).
Generation of immunodeficient ob/ob mice in-volves a long-term breeding process, since severalrounds of heterozygote matings were required toobtain enough double mutant mice for use in theseexperiments (see Materials and Methods). To obtainan appropriate number of readily available animals,
Figure 6. Effect of leptin-transduced human keratinocytegrafts on blood glucose, food consumption and body weighton immunosuppressed ob/ob recipient mice. a) Glucose levelsin leptin-transduced human keratinocyte graft recipientswere significantly lower than controls at all times after 48 h oftransplantation (P,0.01). b) Mice were caged individuallyand food intake was measured over a 48 h period. Values arethe mean 6 se. P , 0.01 for ob/ob transplanted with leptin-transduced HK grafts compared to ob/ob transplanted withcontrol HK grafts 4 days after transplantation. c) Body weightof leptin-transduced human keratinocyte graft recipient micewas significantly lower than that of controls at all times after2 days of grafting (P , 0.05 and P,0.01 after 6 days). Bodyweight is expressed as the percent of body weight on the dayof transplant (day 0). The average body weight for immuno-suppressed ob/ob mice at day 0 was 46.1 6 1.5 g.
Figure 5. Retroviral constructs and in vitro leptin levels.a) The retroviral constructs are based on LZRS vector back-bone. Shaded boxes indicate regulatory sequences (viral LTRand IRES). b) In vitro mouse leptin produced by culturedleptin-transduced human (HK) or K5-leptin primary new-born mouse (MK) keratinocytes. Values are the mean 6 se oftwo independent experiments performed in duplicate.
1534 Vol. 15 July 2001 LARCHER ET AL.The FASEB Journal

we performed the human xenotransplants on cyclos-porin-A immunosuppressed ob/ob mice. Previousstudies showed that cyclosporin-treated C57BL/6Jmice accepted human keratinocyte xenografts for anaverage of 16 days (38), a period when a clearcorrective response was observed with the K5-leptinskin grafts. Both control (IRES-EGFP) and leptin(leptin-IRES-EGFP)-transduced keratinocytes grownto confluence on fibrin-fibroblast gels were trans-planted using an s.c. flap technique (36, 37) incyclosporin-A immunosuppressed C57BL/6J ob/obmice. A study of serum glucose levels and food intakeshowed a rapid corrective effect in the leptin-HKtransplanted mice (Fig. 6a, b). A drop in glucosewithin 48 h of transplantation was the first sign ofkeratinocyte-derived leptin action (Fig. 6a). Foodintake was markedly reduced in leptin-HK trans-planted mice, and food intake measurements showeda 50% reduction compared with control-HK trans-planted mice (Fig. 6b). The rate of weight loss wasrapid and sustained. At the end of the experiment(16 days after grafting), leptin-HK transplanted micehad lost an average of 30% of starting weight. Incontrast, control HK-transplanted mice showed amoderate weight gain (Fig. 6c). Serum leptin wasdetermined at three different times throughout theexperiment. Leptin levels were stable and rangedfrom 1.9 to 2.5 ng/ml (mean 2.2060.24 ng/ml, n55at day five; 2.1160.16 ng/ml at day 10, n54;2.1060.21 ng/ml at day 15, n54; Table 1). At day 16postgrafting, animals were killed to analyze grafthistopathology. Histological evaluation revealed awell-developed, healthy stratified epithelium of hu-man origin, as determined by hematoxylin/eosinstaining and specific human involucrin immunostain-ing (Fig. 7a, b). Active gene expression was con-firmed by EGFP fluorescence in the graft (Fig. 7c).One of the leptin-HK-grafted animals showed a pro-gressive loss of the phenotypic corrective effect start-ing around day 8 postgrafting (data not shown).Histological examination of the grafted area in thisanimal was consistent with T cell-mediated rejection(Fig. 7d), indicating premature loss of the immuno-suppressive treatment efficacy.
As also found for the K5-leptin grafts, a 2–2.5 cm2
ex vivo genetically engineered HK graft sufficed toreplace physiological leptin levels, which in turnreversed the ob/ob syndrome in a short-term experi-ment.
DISCUSSION
The majority of systemic protein deficiencies (i.e., typeI diabetes, hemophilia, dwarfism) are currently treatedby multiple injections of the deficient polypeptide. Safeadministration procedures that would eliminate thisunpleasant duty on the part of the patient would behighly desirable (19). Somatic gene therapy offers anattractive alternative for permanent protein delivery.
Numerous ex vivo, in vivo, and transgenic studieshave established the potential therapeutic use of genet-ically manipulated keratinocytes as a noninvasive vehi-cle of circulatory systemic proteins (20–27).
Most studies of systemic keratinocyte gene therapyhave emphasized critical issues such as duration oftransgene expression and serum levels of the modifiedkeratinocyte-derived proteins (47–49). Less work hasaddressed the correction of circulating protein defi-ciencies. To date, only a partial corrective effect, in amouse model of hemophilia A, has been achievedthrough grafts of involucrin factor VIII transgenicmouse skin (27).
In the present study, we have been able to fullycorrect the phenotype of the leptin-deficient ob/obmouse. When K5-leptin transgenic skin patches weregrafted onto immunodeficient ob/ob mice, a rapid,lasting corrective effect was achieved as the clinical andbiochemical parameters related to the obese pheno-type returned to normal values. High serum leptinprotein levels resulted from the basal cell-specific ker-atin K5 gene promoter activity. High hGH proteinlevels were similarly found in the blood of transgenicmice in which an hGH cDNA was under the control ofthe keratin K14 gene promoter (26). The long-lastingeffect achieved with the K5-leptin transgenic skin graftsreinforces the idea that the use of strong epidermalpromoters would be a major asset to skin gene therapy.To be able to use gene constructs similar to those usedin the generation of transgenic mice, however, im-proved nonviral gene transfer methods for humankeratinocytes are still required (36).
Transgenic approaches, although not of direct clini-cal relevance, allow us to address issues critical tomodeling a therapeutic human ex vivo system, such as1) whether the product of transferred genes can bedelivered from epidermis to blood circulation, 2) tocompare therapeutic protein concentrations in epider-mal cell culture, 3) to make a rough estimate of thegraft size needed, and 4) to assess potential undesiredeffects.
We used the K5-leptin transgenic skin grafting stud-
TABLE 1. In vivo leptin levelsa
Days after grafting 0 5 10 15
Serum leptin levels inleptin HK mice (ng/ml) 0 6 0.0 (n55) 2.20 6 0.24 (n55) 2.11 6 0.16 (n54) 2.10 6 0.21 (n54)
a Blood samples were collected and mouse serum leptin was determined by ELISA at the times indicated. Control HK grafted micepresented undetectable leptin levels at all time points. Values are the mean 6 se.
1535SKIN GENE THERAPY FOR LEPTIN DEFICIENCY

ies to predict whether the ex vivo approach usinggenetically engineered human keratinocytes would befeasible. Differences in promoter specificity (viral LTRsvs. K5) and/or the presence of epidermal appendages,such as hair follicles in the transgenic mouse skin grafts,may lead to substantial variations in keratinocyte-de-rived leptin per area unit of grafted skin in vivo. In fact,although transduced and selected human keratinocytesproduced 30% more bioactive leptin than keratinocytesderived from K5-leptin transgenic mice in vitro, serumleptin levels obtained with leptin HKs in vivo wereslightly lower than predicted. Nevertheless, when graftsof leptin-producing human keratinocytes of similar sizeto those from K5-leptin mice were used, a rapid correc-tive effect was also achieved. Both the transgenic and exvivo cutaneous gene therapy approaches reported hereappear to be as efficient as another long-term deliverysystem in which a leptin-carrying adeno-associated virusis targeted to muscle in ob/ob mice (18). Skin accessi-bility, ex vivo keratinocyte manipulation, and the pos-sibility of graft removal highlight the feasibility as wellas the safety advantages of this procedure over othergene therapy methods for leptin delivery. Both the
immunosuppressive protocol and the grafting tech-nique used allowed only a short-term study. We havenonetheless been able to show that ex vivo, geneticallymodified human keratinocyte grafts comprising lessthan 10% of total body surface were suitable for cor-rection of leptin deficiency in mice. Some clinical andbiochemical parameters of the ob/ob phenotype werenot monitored during our corrective study. Based onprevious studies of recombinant leptin administrationand the fact that the serum leptin levels reached eitherafter transgenic mouse skin or human keratinocytegrafts were within values found in lean animals, glu-cose, weight, and food intake appear to be representa-tive markers of the complete ob/ob syndrome (50).Experiments are under way to determine whetherorthotopic grafting of genetically engineered leptin-expressing human keratinocytes are able to producelong-term correction in immunodeficient ob/ob mice.This goal appears within reach, considering the recentsuccess in achieving sustainable gene expression ingenetically engineered human keratinocytes (24, 49; M.Del Rio et al., unpublished results).
Obesity in humans is a major chronic disorder with a
Figure 7. Histological appearance of leptin-transduced human skin graft in immunodeficient ob/ob mice. a) Human involucrinimmunostaining of a section of the grafted area on the inner surface of the recipient mouse dorsal region demonstratingpositive staining in the leptin-transduced human epithelium (arrowhead). b) Hematoxylin/eosin staining of leptin-HK graft,showing the presence of a well-differentiated epithelium. c) Detection of transduced gene expression through EGFP-fluorescence in an unfixed frozen section of the leptin-HK graft. Nuclei were visualized by DAPI staining (blue fluorescence).d) Hematoxylin/eosin staining of a graft section from a leptin-HK-grafted mouse that lost the leptin-corrective effect. Note thepresence of lymphocyte infiltrate consistent with T cell-mediated rejection of the graft. 340 (a) or 3400 (b–d).
1536 Vol. 15 July 2001 LARCHER ET AL.The FASEB Journal

high incidence of morbidity; its molecular pathogenesisis the subject of intense study. Most obese individualsappear to be resistant to leptin, showing higher thannormal levels of the hormone. In fact, inherited leptindeficiency appears to be a rare cause of extreme obesityin humans (8, 9). Recombinant human leptin has beentested successfully on one such leptin-deficient patient(10). Nonetheless, since leptin is a short half-life pro-tein susceptible to breakdown and inactivation by thegastrointestinal system, daily s.c. injections need to beadministered. It has become apparent that a group ofpatients suffering from some forms of lipodystrophyshow reduced leptin levels (12). Alleviation of severeinsulin resistance after leptin administration has re-cently been shown in two transgenic mouse models ofcongenital generalized lipodystrophy (CGL) (13–16).CGL patients would thus probably benefit from leptinreplacement. It is conceivable that, under experimentalconditions equivalent to those described here, a singleoperation replacing less than 10% of the epidermalsurface with leptin-producing keratinocyte sheetswould suffice to restore normal leptin levels in patientssuffering from leptin deficiency.
Epidermal regeneration obtained with cultured ker-atinocytes has been shown to be long-lasting as well aslife-saving in cases of severe wounds (51, 52). In the lastfew years, cultured epidermal sheets bearing melano-cytes have also been used to treat stable vitiligo withencouraging results; more important, new surgical pro-cedures have been demonstrated that allow graftingwithout scarring (53). These technical advantages andthe feasibility of stable transduction of epidermal stemcells (24, 47, 48) lead us to predict a promising futurefor keratinocyte-mediated gene therapy for leptin andperhaps for other protein deficiencies.
This work was supported by grants PM98–0039 and SAF98–0047 from the Spanish Ministry for Science and Technol-ogy. We wish to thank Dr. Alberto Alvarez and Israel Ormanfor FACS analysis, Cathy Mark for editorial revision of themanuscript, Isabel de los Santos for technical help, Dr. ClaraMartin for surgical assistance, Jesus Martinez for animal care,and Dr. Fernando Benavidez for helpful comments on animalbreeding strategies.
REFERENCES
1. Friedman, J. M., and Halaas, J. L. (1998) Leptin and theregulation of body weight in mammals. Nature (London) 395,763–770
2. Ahima, R. S., and Flier, J. S. (2000) Leptin. Annu. Rev. Physiol.62, 413–437
3. Wang, M. Y., Koyama, K., Shimabukuro, M., Newgard, C. B., andUnger, R. H. (1998) OB-Rb gene transfer to leptin-resistantislets reverses diabetogenic phenotype. Proc. Natl. Acad. Sci. USA95, 714–718
4. Kim, Y. B., Uotani,, S., Pierroz, D. D., Flier, J. S., and Kahn, B. B.(2000) In vivo administration of leptin activates signal transduc-tion directly in insulin-sensitive tissues: overlapping but distinctpathways from insulin. Endocrinology 141, 2328–2339
5. Ducy, P., Amling, M., Takeda, S., Priemel, M., Schilling, A. F.,Beil, F. T., Shen, J., Vinson, C., Rueger, J. M., and Karsenty, G.(2000) Leptin inhibits bone formation through a hypothalamicrelay: a central control of bone mass. Cell 100, 197–207
6. Bouloumie, A., Drexler, H. C., Lafontan, M., and Busse, R.(1998) Leptin, the product of Ob gene, promotes angiogenesis.Circ. Res. 83, 1059–1066
7. Sierra-Honigmann, M. R., Nath, A. K., Murakami, C., Garcia-Cardena, G., Papapetropoulos, A., Sessa, W. C., Madge, L. A.,Schechner, J. S., Schwabb, M. B., Polverini, P., et al. (1998)Biological action of leptin as an angiogenic factor. Science 281,1683–1686
8. Montague, C. T., Farooqi, I. S., Whitehead, J. P., Soos, M. A.,Rau, H., Wareham, N. J., Sewter, C. P., Digby, J. E., Mohammed,S. N., Hurst, J. A., Cheetham, C., Earley, A., Barnett, A., Prins, J.,and O’Rahilly, S. (1997) Congenital leptin deficiency is associ-ated with severe early-onset obesity in humans. Nature (London)387, 903–908
9. Strobel, A., Issad, T., Camoin, L., Ozata, M., and Strosberg, A. D.(1998) A leptin missense mutation associated with hypogonad-ism and morbid obesity. Nat. Genet. 3, 213–215
10. Farooqi, I. S., Jebb, S. A., Langmack, G., Lawrence, E.,Cheetham, C. H., Prentice, A. M., Hughes, I. A., McCamish,M. A., and O’Rahilly, S. (1999) Effects of recombinant leptintherapy in a child with congenital leptin deficiency. N. Engl.J. Med. 341, 879–884
11. Seip, M., and Trygstad, O. (1996) Generalized lipodystrophy,congenital and acquired (lipoatrophy). Acta Paediatr. Suppl. 13,2–28
12. Pardini, V. C., Victoria, I. M., Rocha, S. M., Andrade, D. G.,Rocha, A. M., Pieroni, F. B., Milagres, G., Purisch, S., and Velho,G. (1998) Leptin levels, beta-cell function, and insulin sensitivityin families with congenital and acquired generalized lipoatropicdiabetes. J. Clin. Endocrinol. Metab. 83, 503–508
13. Shimomura, I., Hammer, R. E., Richardson, J. A., Ikemoto, S.,Bashmakov, Y., Goldstein, J. L., and Brown, M. S. (1998) Insulinresistance and diabetes mellitus in transgenic mice expressingnuclear SREBP-1c in adipose tissue: model for congenital gen-eralized lipodystrophy. Genes Dev. 12, 3182–3194
14. Moitra, J., Mason, M. M., Olive, M., Krylov, D., Gavrilova, O.,Marcus-Samuels, B., Feigenbaum, L., Lee, E., Aoyama, T.,Eckhaus, M., Reitman, M., and Vinson, C. (1998) Life withoutwhite fat: a transgenic mouse. Genes Dev. 12, 3168–3181
15. Shimomura, I., Hammer, R. E., Ikemoto, S., Brown, M. S., andGoldstein, J. L. (1999) Leptin reverses insulin resistance anddiabetes mellitus in mice with congenital lipodystrophy. Nature(London) 401, 73–76
16. Ebihara, K., Ogawa, Y., Masuzaki, H., Miyanaga, F., Shintani, M.,Abe, M., Hayashi, T., Hosoda, K., Inoue, G., Yoshimasa, Y.,Reitman, M., and Nakao, K. (2000) Increased glucose metabo-lism and insulin sensitivity in a mouse model of lipoatrophicdiabetes by transgenic overexpression of leptin. Diabetes 49,Suppl. A210 (abstr.)
17. Muzzin, P., Eisensmith, R. C., Copeland, K. C., and Woo, S. L.(1996) Correction of obesity and diabetes in genetically obesemice by leptin gene therapy. Proc. Natl. Acad. Sci. USA 93,14804–14808
18. Murphy, J. E., Zhou, S., Giese, K., Williams, L. T., Escobedo,J. A., and Dwarki, V. J. (1997) Long-term correction of obesityand diabetes in genetically obese mice by a single intramuscularinjection of recombinant adeno-associated virus encodingmouse leptin. Proc. Natl. Acad. Sci. USA 94, 13921–13926
19. Morsy, M. A., Gu, M. C., Zhao, J. Z., Holder, D. J., Rogers, I. T.,Pouch, W. J., Motzel, S. L., Klein, H. J., Gupta, S. K., Liang, X.,et al. (1998) Leptin gene therapy and daily protein administra-tion: a comparative study in the ob/ob mouse. Gene Ther. 5,8–18
20. Fenjves, E. S., Smith, J., Zaradic, S., and Taichman, L. B. (1994)Systemic delivery of secreted protein by grafts of epidermalkeratinocytes: prospects for keratinocyte gene therapy. Hum.Gene Ther. 5, 1241–1248
21. Morgan, J. R., Barrandon, Y., Green, H., and Mulligan, R. C.(1987) Expression of an exogenous growth hormone gene bytransplantable human epidermal cells. Science 237, 1476 –1479
22. Teumer, J., Lindahl, A., and Green, H. (1990) Human growthhormone in the blood of athymic mice grafted with cultures ofhormone-secreting human keratinocytes. FASEB J. 4, 3245–3250
23. Gerrard, A. J., Hudson, D. L., Brownlee, G. G., and Watt, F. M.(1993) Towards gene therapy for haemophilia B using primaryhuman keratinocytes. Nat. Genet. 3, 180–183
1537SKIN GENE THERAPY FOR LEPTIN DEFICIENCY

24. Mathor, M. B., Ferrari, G., Dellambra, E., Cilli, M., Mavilio, F.,Cancedda, R., and De Luca, M. (1996) Clonal analysis of stablytransduced human epidermal stem cells in culture. Proc. Natl.Acad. Sci. USA 93, 10371–10376
25. Alexander, M. Y., Bidichandani, S. I., Cousins, F. M., Robinson,C. J., Duffie, E., and Akhurst, R. J. (1995) Circulating humanfactor IX produced in keratin-promoter transgenic mice: afeasibility study for gene therapy of haemophilia B. Hum. Mol.Genet. 4, 993–999
26. Wang, X., Zinkel, S., Polonsky, K., and Fuchs, E. (1997) Trans-genic studies with a keratin promoter-driven growth hormonetransgene: prospects for gene therapy. Proc. Natl. Acad. Sci. USA94, 219–226
27. Fakharzadeh, S. S., Zhang, Y., Sarkar, R., and Kazazian, H. H., Jr.(2000) Correction of the coagulation defect in hemophilia Amice through factor VIII expression in skin. Blood. 95, 2799–2805
28. Meng, X., Sawamura, D., Tamai, K., Hanada, K., Ishida, H., andHashimoto, I. (1998) Keratinocyte gene therapy for systemicdiseases. Circulating interleukin 10 released from gene-trans-ferred keratinocytes inhibits contact hypersensitivity at distantareas of the skin. J. Clin. Invest. 101, 1462–1467
29. Coleman, D. L. (1978) Obese and diabetes: two mutant genescausing diabetes-obesity syndromes in mice. Diabetologia 14,141–148
30. Leiter, E. H. (1992) Variation in the severity and duration ofhyperglycemia in the C57BL/6-ob/ob (obese) mouse. JAX Notes451
31. Ramirez, A., Bravo, A., Jorcano, J. L., and Vidal, M. (1994)Sequences 59 of the bovine keratin 5 gene direct tissue- and celltype-specific expression of a lacZ gene in the adult and duringdevelopment. Differentiation 58, 53–64
32. Murillas, R., Larcher, F., Conti, C. J., Santos, M., Ullrich, A., andJorcano, J. L. (1995) Expression of a dominant negative mutantof epidermal growth factor receptor in the epidermis of trans-genic mice elicits striking alterations in hair follicle develop-ment and skin structure. EMBO J. 14, 5216–5223
33. Namae, M., Mori, Y., Yasuda, K., Kadowaki, T., Kanazawa, Y., andKomeda, K. (1998) New method for genotyping the mouseLep(ob) mutation, using a polymerase chain reaction assay. Lab.Anim. Sci. 48, 103–105
34. Albella, B., Segovia, J. C., Guenechea, G., Pragnell, I. B., andBueren, J. A. (1999) Preserved long-term repopulation anddifferentiation properties of hematopoietic grafts subjected toex vivo expansion with stem cell factor and interleukin 11.Transplantation 67, 1348–1357
35. Meana, A., Iglesia, J., Del Rio, M., Larcher, F., Madrigal, B.,Fresno, M. F., Martin, C., San Roman, F., and Tevar, F. (1998)Large surface of cultured human epithelium obtained on adermal matrix based on live fibroblast-containing fibrin gels.Burns 7, 621–630
36. Del Rio, M., Larcher, F., Meana, A., Segovia, J., and Jorcano, J.(1999) Non-viral transfer of genes to pig primary keratinocytes.Induction of angiogenesis by composite grafts of modifiedkeratinocytes overexpressing VEGF driven by a keratin pro-moter. Gene Ther. 6, 1734–1741
37. Barrandon, Y., Li, V., and Green, H. (1988) New techniques forthe grafting of cultured human epidermal cells onto athymicanimals. J. Invest. Dermatol. 91, 315–318
38. Compton, C. C., Rizk, I., Regauer, S., Burd, E., Holaday, J., andKenner, J. (1994) The effect of bromocryptine-induced hypo-prolactinemia on xenogeneic and allogeneic skin graft survivalin a mouse model. J. Burn Care Rehabil. 15, 393–400
39. Kinsella, T. M., and Nolan, G. P. (1996) Episomal vectors rapidlyand stably produce high-titer recombinant retrovirus. Hum. GeneTher. 7, 1405–1413
40. Yang, S., Delgado, R., King, S., Woffendin, C., Barker, C., Yang,Z., Xu, L., Nolan, G., and Nabel, G. (1999) Generation ofretroviral vector for clinical studies using transient transfection.Hum. Gene Ther. 10, 123–132
41. Robles, A. I., Larcher, F., Whalin, R. B., Murillas, R., Richie, E.,Gimenez-Conti, I. B., Jorcano, J. L., and Conti, C. J. (1996)Expression of cyclin D1 in epithelial tissues of transgenic miceresults in epidermal hyperproliferation and severe thymic hy-perplasia. Proc. Natl. Acad. Sci. USA 93, 7634–7638
42. DiGiovanni, J., Bol, D. K., Wilker, E., Beltran, L., Carbajal, S.,Moats, S., Ramirez, A., Jorcano, J., and Kiguchi, K. (2000)Constitutive expression of insulin-like growth factor-1 in epider-mal basal cells of transgenic mice leads to spontaneous tumorpromotion. Cancer Res. 60, 1561–1570
43. Ogawa, Y., Masuzaki, H., Hosoda, K., Aizawa-Abe, M., Suga, J.,Suda, M., Ebihara, K., Iwai, H., Matsuoka, N., Satoh, N., et al.(1999) Increased glucose metabolism and insulin sensitivity intransgenic skinny mice overexpressing leptin. Diabetes 48, 1822–1829
44. Taichman, L. (1994) Epithelial gene therapy. In The KeratinocyteHandbook (Leight, I., Lane, B., and Watt, F., eds) pp. 543–551,Cambridge Univ. Press
45. Deng, H., Choate, K. A., Lin, Q., and Khavari, P. A. (1998)High-efficiency gene transfer and pharmacologic selection ofgenetically engineered human keratinocytes. BioTechniques 25,274–280
46. Pfutzner, W., Hengge, U. R., Joari, M. A., Foster, R. A., andVogel, J. C. (1999) Selection of keratinocytes transduced withthe multidrug resistance gene in an in vitro skin model presentsa strategy for enhancing gene expression in vivo. Hum. GeneTher. 10, 2811–2821
47. Deng, H., Lin, Q., and Khavari, P. A. (1997) Sustainablecutaneous gene delivery. Nat. Biotechnol. 15, 1388–1391
48. De Luca, M., and Pellegrini, G. (1997) The importance ofepidermal stem cells in keratinocyte-mediated gene therapy.Gene Ther. 4, 381–383
49. Kolodka, T., Garlick, J., and Taichman, L. (1998) Evidence forkeratinocyte stem cells in vitro: long term engraftment andpersistence of transgene expression from retrovirus-transducedkeratinocytes. Proc. Natl. Acad. Sci. USA 95, 4356–4361
50. Ioffe, E., Moon, B., Connolly, E., and Friedman, J. M. (1998)Abnormal regulation of the leptin gene in the pathogenesis ofobesity. Proc. Natl. Acad. Sci. USA 95, 11852–11857
51. Gallico, G. G., O’Connor, N. E., Compton, C. C., Kehinde, O.,and Green, H. (1984) Permanent coverage of large burnwounds with autologous cultured human epithelium. N. Engl.J. Med. 311, 448–451
52. Pellegrini, G., Bondanza, S., Guerra, L., and De Luca, M.(1998) Cultivation of human keratinocyte stem cells: currentand future clinical applications. Med. Biol. Eng. Comput. 36,778 –790
53. Guerra, L., Capurro, S., Melchi, F., Primavera, G., Bondanza, S.,Cancedda, R., Luci, A., De Luca, M., and Pellegrini, G. (2000)Treatment of ‘stable’ vitiligo by timed surgery and transplanta-tion of cultured epidermal autografts. Arch. Dermatol. 136,1380–1389
Received for publication February 12, 2001.Accepted for publication March 26, 2001.
1538 Vol. 15 July 2001 LARCHER ET AL.The FASEB Journal
Copyright © 2022 FDOKUMEN




















