
A comparison of the synaptic proteome in human chronic schizophrenia and rat ketamine psychosis...
19
ORIGINAL ARTICLE A comparison of the synaptic proteome in human chronic schizophrenia and rat ketamine psychosis suggest that prohibitin is involved in the synaptic pathology of schizophrenia K-H Smalla 1 , M Mikhaylova 2 , J Sahin 2 , H-G Bernstein 3 , B Bogerts 3 , A Schmitt 4 , R van der Schors 5 , AB Smit 5 , KW Li 5 , ED Gundelfinger 6 and MR Kreutz 2 1 Special Lab Molecular Biological Techniques, Leibniz Institute for Neurobiology, Magdeburg, Germany; 2 Project Group Neuroplasticity, Leibniz Institute for Neurobiology, Magdeburg, Germany; 3 Department of Psychiatry, Medical Faculty, Otto-von Guericke University, Magdeburg, Germany; 4 Department of Psychiatry, Medical Faculty, Georg-August University Go ¨ ttingen, Germany; 5 Department of Molecular and Cellular Neurobiology, Center for Neurogenomics and Cognitive Research, Vrije Universiteit, Amsterdam, The Netherlands and 6 Department of Neurochemistry/Molecular Biology, Leibniz Institute for Neurobiology, Magdeburg, Germany Many studies in recent years suggest that schizophrenia is a synaptic disease that crucially involves a hypofunction of N-methyl-D-aspartate receptor-mediated signaling. However, at present it is unclear how these pathological processes are reflected in the protein content of the synapse. We have employed two-dimensional gel electrophoresis in conjunction with mass spectrometry to characterize and compare the synaptic proteomes of the human left dorsolateral prefrontal cortex in chronic schizophrenia and of the cerebral cortex of rats treated subchronically with ketamine. We found consistent changes in the synaptic proteomes of human schizophrenics and in rats with induced ketamine psychosis compared to controls. However, commonly regulated proteins between both groups were very limited and only prohibitin was found upregulated in both chronic schizophrenia and the rat ketamine model. Prohibitin, however, could be a new potential marker for the synaptic pathology of schizophrenia and might be causally involved in the disease process. Molecular Psychiatry (2008) 13, 878–896; doi:10.1038/mp.2008.60; published online 27 May 2008 Keywords: proteomics; chronic schizophrenia; postsynaptic density; left dorsolateral prefrontal cortex; synapse; prohibitin Introduction The etiology of most neuropsychiatric disorders is incompletely understood. In recent years the concept has emerged that for several disorders, including schizophrenia, their pivotal cause might be a dysre- gulation in synapto-dendritic morphogenesis during development. 1,2 There is accumulating evidence that disturbances during neuronal differentiation and maturation processes in development can have profound effects on neuronal information processing in the adult despite the relatively mild morphological phenotypes in the affected brain regions. The most prevailing evidence for altered synapto-dendritic microcircuitry exists in schizophrenia (for reviews see Eastwood, 2004; 3 Lewis and Levitt, 2002; 4 McGlashan and Hoffman, 2000; 5 Woods, 1998 6 ). Interestingly, the aberrant wiring patterns observed in patients with chronic paranoid schizophrenia seem to be particularly prominent only in certain brain regions such as the prefrontal cortex. 3,4,7 The structur- al anomalies in schizophrenic patients include a region- and layer-specific reduction of spine synapses that concomitantly frequently exhibit an aberrant morphology, 8–15 smaller dendritic fields and a decreased number of basilar dendrites in conjunction with simplified dendritic branching patterns. 8,16,17 In essence, the available data suggest that the wiring of circuitry is altered in schizophrenia 3,5,6,15,17,18 and that the synaptic pathology might be causally related to abnormalities in dendrite morphology. At present, it is not clear how these alterations were initially brought about during development and why no phenotypic correction occurred until adulthood. As the synaptic and dendritic aberrations in schizo- phrenia are relatively mild and not present in all brain regions, it is reasonable to assume that local changes at a limited number of synapses and Received 26 October 2007; revised 23 April 2008; accepted 25 April 2008; published online 27 May 2008 Correspondence: Dr MR Kreutz, Project Group Neuroplasticity, Leibniz Institute for Neurobiology, Brenneckestrasse 6, Magdeburg, Saxony-Anhalt 39118, Germany. E-mail: [email protected] Molecular Psychiatry (2008) 13, 878–896 & 2008 Nature Publishing Group All rights reserved 1359-4184/08 $30.00 www.nature.com/mp
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A comparison of the synaptic proteome in human chronic schizophrenia and rat ketamine psychosis...

ORIGINAL ARTICLE
A comparison of the synaptic proteome in human chronicschizophrenia and rat ketamine psychosis suggest thatprohibitin is involved in the synaptic pathology ofschizophreniaK-H Smalla1, M Mikhaylova2, J Sahin2, H-G Bernstein3, B Bogerts3, A Schmitt4, R van der Schors5,
AB Smit5, KW Li5, ED Gundelfinger6 and MR Kreutz2
1Special Lab Molecular Biological Techniques, Leibniz Institute for Neurobiology, Magdeburg, Germany; 2Project GroupNeuroplasticity, Leibniz Institute for Neurobiology, Magdeburg, Germany; 3Department of Psychiatry, Medical Faculty, Otto-vonGuericke University, Magdeburg, Germany; 4Department of Psychiatry, Medical Faculty, Georg-August University Gottingen,Germany; 5Department of Molecular and Cellular Neurobiology, Center for Neurogenomics and Cognitive Research, VrijeUniversiteit, Amsterdam, The Netherlands and 6Department of Neurochemistry/Molecular Biology, Leibniz Institute forNeurobiology, Magdeburg, Germany
Many studies in recent years suggest that schizophrenia is a synaptic disease that cruciallyinvolves a hypofunction of N-methyl-D-aspartate receptor-mediated signaling. However, atpresent it is unclear how these pathological processes are reflected in the protein content ofthe synapse. We have employed two-dimensional gel electrophoresis in conjunction withmass spectrometry to characterize and compare the synaptic proteomes of the human leftdorsolateral prefrontal cortex in chronic schizophrenia and of the cerebral cortex of ratstreated subchronically with ketamine. We found consistent changes in the synaptic proteomesof human schizophrenics and in rats with induced ketamine psychosis compared to controls.However, commonly regulated proteins between both groups were very limited and onlyprohibitin was found upregulated in both chronic schizophrenia and the rat ketamine model.Prohibitin, however, could be a new potential marker for the synaptic pathology ofschizophrenia and might be causally involved in the disease process.Molecular Psychiatry (2008) 13, 878–896; doi:10.1038/mp.2008.60; published online 27 May 2008
Keywords: proteomics; chronic schizophrenia; postsynaptic density; left dorsolateral prefrontalcortex; synapse; prohibitin
Introduction
The etiology of most neuropsychiatric disorders isincompletely understood. In recent years the concepthas emerged that for several disorders, includingschizophrenia, their pivotal cause might be a dysre-gulation in synapto-dendritic morphogenesis duringdevelopment.1,2 There is accumulating evidence thatdisturbances during neuronal differentiation andmaturation processes in development can haveprofound effects on neuronal information processingin the adult despite the relatively mild morphologicalphenotypes in the affected brain regions. The mostprevailing evidence for altered synapto-dendriticmicrocircuitry exists in schizophrenia (for reviewssee Eastwood, 2004;3 Lewis and Levitt, 2002;4
McGlashan and Hoffman, 2000;5 Woods, 19986).Interestingly, the aberrant wiring patterns observedin patients with chronic paranoid schizophrenia seemto be particularly prominent only in certain brainregions such as the prefrontal cortex.3,4,7 The structur-al anomalies in schizophrenic patients include aregion- and layer-specific reduction of spine synapsesthat concomitantly frequently exhibit an aberrantmorphology,8–15 smaller dendritic fields and adecreased number of basilar dendrites in conjunctionwith simplified dendritic branching patterns.8,16,17 Inessence, the available data suggest that the wiring ofcircuitry is altered in schizophrenia3,5,6,15,17,18 and thatthe synaptic pathology might be causally related toabnormalities in dendrite morphology.
At present, it is not clear how these alterations wereinitially brought about during development and whyno phenotypic correction occurred until adulthood.As the synaptic and dendritic aberrations in schizo-phrenia are relatively mild and not present inall brain regions, it is reasonable to assume thatlocal changes at a limited number of synapses and
Received 26 October 2007; revised 23 April 2008; accepted 25April 2008; published online 27 May 2008
Correspondence: Dr MR Kreutz, Project Group Neuroplasticity,Leibniz Institute for Neurobiology, Brenneckestrasse 6, Magdeburg,Saxony-Anhalt 39118, Germany.E-mail: [email protected]
Molecular Psychiatry (2008) 13, 878–896& 2008 Nature Publishing Group All rights reserved 1359-4184/08 $30.00
www.nature.com/mp

dendrites are sufficient to trigger changes leading todisease development. According to preclinical andclinical data, an imbalance of dopamine and/orglutamate transmission in the striatum and/or theprefrontal cortex appears crucial in schizophre-nia.19,20 The N-methyl-D-aspartate (NMDA) receptoris anchored to the postsynaptic density (PSD), amultiprotein meshwork of surface receptors, scaffold-ing proteins and intracellular effectors. Moreover, thePSD is a site of integration of glutamate and dopaminesignaling by mechanisms of transactivation or con-vergence21,22 of intracellular pathways. Indeed, sev-eral PSD proteins have been associated withschizophrenia although the nature of their role islargely unclear. Interestingly, ketamine, a noncompe-titive antagonist of NMDA receptors, is known toinduce psychotomimetic effects in healthy volunteersand to re-exacerbate psychotic symptoms in chronicschizophrenic patients (Kristiansen et al.23 andreferences therein). Due to the blockade of the NMDAreceptor ion channel, ketamine is predicted to inducea state of NMDA receptor hypofunction. In this study,we analyzed systematically the synaptic proteome inthe subchronic rat ketamine model of psychosis.24,25
To further validate the animal model we subsequentlycompared the data with a proteomic screen of asynaptic membrane fraction of the left dorsolateralprefrontal cortex (DPLFC) of patients suffering fromchronic schizophrenia. To validate the approach wefinally examined in more detail the expression of oneprotein, prohibitin, which was found to be dysregu-lated under both conditions.
Materials and methods
Human subjectsCase recruitment, acquisition of personal data,autopsy and handling of autoptic material has beenapproved by the responsible ethical committee. Forprohibitin immunostainings post-mortem brains ofeight patients with schizophrenia according to Diag-nostic and Statistical Manual of Mental Disorders,2nd edn, revised (five men, three women) and sevenmatched control cases (five men, two women) wereinvestigated (Supplementary Table 1). All brains wereobtained from the New Magdeburg brain collection.Patients with schizophrenia and psychically healthypatients died between years 1988 and 1993. Ageranged from 38 to 63 years. A lifetime psychiatricdiagnosis of schizophrenia was established accordingto DSM-III-R (American Psychiatric Association,1987) by using the Diagnostic Instrument for brainstudies,26 a structured instrument for the collection ofclinical, pharmacological and other relevant informa-tion from case histories. All patients with schizo-phrenia had histories of inpatient hospitalization.Only patients with well-documented and extensiveclinical records were selected for the present study.Demographic data are summarized in SupplementaryTable 1. The mean duration of illness was 18.3 yearswith a range from 8 to 34 years. The subtypes of
schizophrenia according to DSM-III-R and otherclinical characteristics are also presented in Supple-mentary Table 1. Brain volumes were calculated bythe weight method using fresh whole brain weightand brain density.27 There were no significantdifferences (Student’s t-test, w2-analysis) betweenpatients with schizophrenia and controls with regardto age, gender and storage delay. For quantitativeimmunoblotting experiments, six shock-frozen tissuesamples of the DPLFC were analyzed from schizo-phrenic and control patients, respectively (for detailssee Bernstein et al.28). Two-dimensional (2D) gelelectrophoresis was performed with DPLFC tissuesamples from 10 schizophrenic and 15 control cases(for patient and treatment details, see SupplementaryTable 2). All schizophrenic patients received anti-psychotic drug medication during the course of theirdisease. Control brains of patients without a history ofneuropsychiatric disorders were obtained from thesame pathological institutes or medical examiner’soffice. Brains with lifetime reports of abuse of alcohol,drugs, dementia, neurological illness, trauma, chronicterminal disease known to affect the brain wereexcluded. In addition, quantitative neuropathologicalchanges due to neurodegenerative disorders wereruled out by an experienced neuropathologist.29
Animal experiments
All experimental procedures were performed accord-ing to the National Institutes of Health guidelines forthe care and use of laboratory animals and Germanlaw of animal welfare; they were approved by thelocal authorities. All rats were 12 weeks old when theexperiments were started (weight: 250–300 g). Ani-mals were kept on a 12/12 h dark–light cycle at arelative humidity of 50–60% and 22 1C with food andwater available ad libitum. A total of eight rats wereused for each experimental group. Ketamine psycho-sis was induced by the subchronic administration ofthe drug (30 mg kg�1) in male Sprague–Dawley rats(local breeding facilities of the Leibniz Institute forNeurobiology) as previously described.24 Ketamine(Sigma-Aldrich, Munich, Germany ) was dissolved in0.9% NaCl solution and administered intraperitone-ally on 5 consecutive days. This treatment regime hasbeen shown to induce psychosis-like behavior in ratsfor several weeks.25 Control animals received thesame number of saline injections under the same timeregime as ketamine-treated rats. To control forpotential effects of antipsychotic medication onprohibitin levels rats were subchronically adminis-tered with clozapine (Sigma-Aldrich). The rats re-ceived daily injections of the drug (5 mg per kg) for 14days. Control rats were injected the correspondingvolume of vehicle. Fourteen days after termination ofketamine or directly after termination of clozapinetreatment, rats were anesthetized with halothane anddecapitated. From the brains the cortical tissue wasdissected and then further subjected to biochemicalfractionation.
Prohibitin in schizophreniaK-H Smalla et al
879
Molecular Psychiatry

Biochemical fractionation and 2D gel electrophoresisIn order to obtain a fraction enriched for synapticstructures a protocol established previously30 wasused. Briefly, cortical tissue was homogenized in10 mM Tris-HCl, pH 8.1 containing 0.5% TritonX-100 and protease inhibitors (Complete; Roche,Mannheim, Germany), incubated for 1 h and spun at100 000 g for 1 h (Optima Ultracentrifuge; BeckmanCoulter, Fullerton, CA, USA). The resulting pellet wasrehomogenized in the same buffer and centrifugedagain for 1 h at 100 000 g. Pellets were stored at �80 1C.
For 2D gel electrophoresis, samples werehomogenized in lysis buffer (8 M urea, 4% 3[(3-cholamidopropyl)dimethylammonio]-propanesulfonicacid (CHAPS), 10 mM Tris-HCl pH 7.5, 1% dithio-threitol (DTT)), lysed for 2 h and spun at 20 000 g for20 min. Protein concentrations of the supernatantwere determined using a modified Bradford assay(Bio-Rad, Munich, Germany). Samples containing400mg protein were adjusted to a total volume of400ml, complemented with 2 ml immobilized pHgradient (IPG) buffer 3–10 and loaded on an IPGStrip(Immobiline 18 cm DryStrip 3-11NL; GE Healthcare,Freiburg, Germany). Strips were rehydrated at 30 V for12 h. Focusing was achieved by increasing the voltageto 8000 V with a maximum current of 50 mA per stripfor a total of 75 000 V hours.
After focusing strips were stored at �80 1C, forSDS–polyacrylamide gel electrophoresis (PAGE)strips were equilibrated first in 6 M urea, 2% SDS,1% DTT, 30% glycerol in 50 mM Tris-HCl pH 8.4 for15 min and afterward for 15 min in the same bufferbut containing 2.5% iodoacetamide instead of DTT.SDS–PAGE was run overnight at 20 1C in a HoeferDALT system using 1.5 mm 11% gels (Duracryl,Genomic solutions; gel size 24� 19 cm) at 80 V.Proteins were visualized by silver staining. Foridentification of proteins a preparative 2D gel wasrun in parallel with an 800mg protein sample andsubsequently stained with Coomassie brilliant blue.
Computer-aided gel analysis and quantificationGels were digitized on a GS800 scanner (Bio-Rad) forfurther data processing using PDQuest 2D gel analysissoftware (version 8.0. 035; Bio-Rad) for spot detection,gel matching and spot quantification. For evaluationof differential protein regulation a two-tailed Stu-dent’s t-test was applied to optical density data forcomparison between the two groups of samples. Onlydifferences for P < 0.05 were considered as significantand were independently verified by visual inspectionfor correct matching. Exclusively for statisticallyrelevant data ratios were calculated from the meanoptical densities of a protein spot within correspond-ing groups.
Sample preparation for mass spectrometry and MALDI-TOFProtein spots were excised manually with a diameterof 2 mm and digested with trypsin (Promega Germany,Mannheim), Ziptip purified (Millipore Germany,
Schwalbach) and loaded onto a matrix-assisted laserdesorption/ionization (MALDI) target plate as de-scribed.31 For protein analysis an Applied Biosystems4700 Proteomics Analyzer with TOF/TOF Optics wasused as described earlier.32 For peptide mass finger-print analysis a mass tolerance of 20 p.p.m. withinternal calibration and one missing cleavage sitewere allowed. Mass spectra were searched using theMascot engine against the SwissProt human database.As criteria for correct identification the Mascotprobability score (typically > 95%) and the compar-ison of theoretical and experimental values formolecular mass and isoelectric point were used. Thisway we could clearly define the identity of theprotein although sometimes more than one proteinwas suggested by the database search.
Subcellular fractionation, immunoblotting andimmunocytochemistryQuantitative immunoblot analysis was performed aspreviously described.33,34 In brief, centrifugation for1 h at 100 000 g resulted in the soluble fractions. Afterwashing the pellets twice in the same buffer (1 h100 000 g) aliquots of the particulate fractions (P) wereused for western blots. The remaining pellets wererehomogenized in lysis buffer (0.5 mM NaHCO3,2.5 mM Tris-HCl, 0.5% Triton X-100, pH 8.1), incu-bated for 1 h at 4 1C and subsequently spun for 1 h at100 000 g. Pellets were washed once with lysis buffer(1 h 100 000 g) and finally resuspended in gel-loadingbuffer. Proteins (20mg per lane) were separated byelectrophoresis on 5–20% polyacrylamide gels underfully denaturing conditions (SDS–PAGE) and trans-ferred onto nitrocellulose. For immunodetectionwestern blots were incubated overnight with amonoclonal prohibitin antibody (Abcam UK,Cambridge; dilution 1:1000). After extensive washingblots were incubated with horseradish peroxidase(HRP)-conjugated secondary antibodies (anti-mouseimmunoglobulin (IgG) and anti-guinea pig IgG fromDAKO (dilution 1:5000, Hamburg, Germany; anti-rabbit IgG from Cell Signalling Germany, Frankfurt,dilution 1:5000) for 2 h and then developed using theECL detection system (GE Healthcare). Subcellularfractionation of rat brain followed published proto-cols.34 Quantification was performed with the Quan-tity One software from Bio-Rad.
Immunocytochemical stainings were performed aspreviously described.35 Cortical primary neurons after21 days in vitro (DIV) were fixed in 4% formaldehydein phosphate-buffered saline (PBS) for 10 min at37 1C. Cultures were then permeabilized with 0.25%Triton X-100 for 10 min, blocked for 1.5 h with 5%bovine serum albumin buffer and then incubatedovernight with primary antibody (rabbit anti-prohibi-tin (1:100; Abcam), anti-ProSAP235 guinea pig (1:800)and mouse anti-Bassoon antibody36 (1:1000). Cover-slips were then rinsed in PBS and incubated withfluorescence-labeled antibody (Alexa-488, Alexa-568,Cy5; Molecular Probes, Leiden, the Netherlands). Tovisualize mitochondria cells were incubated with
Prohibitin in schizophreniaK-H Smalla et al
880
Molecular Psychiatry

mitotracker 568 (Invitrogen, Karlsruhe, Germany) at adilution of 1:2500 for 20 min at 37 1C. Thereafter cellswere fixed and stained with prohibitin and ProSAP2antibodies. The fluorescence intensity of the synapticprohibitin staining was analyzed as the mean grayvalue within the selected area. This is the sum of thegray values of all the pixels in the selection dividedby the number of pixels (http://rsb.info.nih.gov/ij/).The gray scale (gray values) ranges from 0 to 255.
Prohibitin overexpression in cortical primary culturesTransfection of primary neurons and analysis ofsynapto-dendritic phenotypes were performed essen-tially as previously described.37,38 Neurons weretransfected at DIV7 and fixed with 4% paraformalde-hyde (PFA) at DIV21. Stainings with synaptic anddendritic markers were performed as described above.Dendrites were analyzed with Sholl analysis usingImageJ (http://rsb.info.nih.gov/ij/). For dendritic-spinemeasurements, the maximum length and head widthof each spine present on the dendrites was manuallytraced and automatically measured by computer. Thegreen fluorescent protein (GFP) channel was taken as areadout for spine morphology because GFP-prohibitinis like GFP highly soluble and filled all spines asjudged by ProSAP2 immunocytochemistry. Spineshapes were classified according to categories basedon confocal laserscan pictures. Capturing of confocalimages and their subsequent analysis was performedindependently by two different researchers (MM, JS)unaware of the group structure. Filopodia-like andthin spines were not further differentiated.
Tissue processing for immunoblot analysis of humantissueFor western blots, frozen tissue from the DPLFC of sixcontrol cases and schizophrenics was used (fordemographic details see Bernstein et al.28). Shock-frozen tissue samples from the DPLFC were homo-genized in 20 mM Tris buffer, pH 7.4, containing aprotease inhibitor mixture (Roche). Detection wasperformed with HRP-conjugated anti-mouse secondaryantibody and the ECL detection kit (GE Healthcare).
Tissue processing for immunohistochemistry of humanbrainThe tissue preparation was performed as describedearlier.39 Briefly, brains were fixed in toto in 8%phosphate-buffered formaldehyde (pH 7.0) for at least2 months. After embedding of the brains in Paraplast,serial coronal 20-mm-thick sections were cut on amicrotome and mounted. Frontal and occipital poleswere separated by coronal cuts anterior to the genuand posterior to the splenium of the corpus callosum.Frontal poles were used throughout this study. Every50th section was stained according to the combinedNissl (cresyl violet) and myelin (Heidenhain-Wolcke)procedure. The actual thickness of each section wasthen determined with a � 100 oil immersion objec-tive, because it is known that section thickness isdecreased after histological processing. The thickness
of the section was determined by focusing the upperand lower surfaces of the section and then subtractingthe z axis coordinate of the lower surface from that ofthe upper surface. The movements in the z axis weremeasured with a microcator as an integral part of theLeica DM RB microscope (Leica, Gie�en, Germany).The section thickness after the histological proce-dures was 18.9±1.0 mm (mean±s.d.)
Prohibitin immunoreactivity was detected with amonoclonal antibody (ab-1836; Abcam) using thenickel-enhanced avidin–biotin technique as de-scribed.40 For immunohistochemical staining sevenwhole brain frontal sections from the prefrontal cortexof each case were used. Three of these sections weresubjected to prohibitin immunohistochemistry. Thesampling of the sections was performed system-atically at intervals of about 2 mm in an area situatedbetween 3.5 and 2 cm rostral of the genu of the corpuscallosum. The staining protocol was performed asdescribed earlier.41 After preincubation of the sectionswith methanol/H2O2 to block endogenous peroxi-dases and repeated washing with PBS, the antibodywas applied at a working dilution of 1:50 in PBS.Thereafter the sections were processed with theavidin–biotin method (Vectastain-peroxidase kit; Vec-tor, Burlingame, CA, USA) and the reaction productwas visualized with 3,30-diaminobenzidine. The colorreaction was enhanced by adding 2 ml of a 0.5% (v/v)nickel ammonium sulfate solution to the diamino-benzidine.42 Specificity of the immunoreaction wascontrolled by incubation with buffer and omitting theantiserum. No specific staining was observed afterperforming control reactions.
Morphometrical analysisAll slides were coded so that the investigators (H-G B,S Funke) were blind to diagnosis during the analysis.The section thickness after the histological proce-dures was 18.9±1.0 mm (mean±s.d.). A counting gridwas used to define a 3D box within the thickness ofthe section as described earlier,39 allowing at least4-mm guard zones at the top and bottom of the section,and to apply a direct, 3D counting method. Prohibi-tin-immunoreactive neurons were counted separatelyin a linear probe of stacked counting boxes extendingfrom the pial surface to the underlying white matter.7
Laminar boundaries, which were most clearly visibleat low power (� 6.3), were marked on the linearprobes by switching back and forth between �6.3and �40 objectives during the analysis so that celldensity for each of the six layers could be calculated.Total length of the linear probe provided a measure-ment of cortical thickness. Values from the five linearprobes were averaged to obtain a mean value ofneuronal density. Eight boxes per cortical layer werecounted. For selected cortical brain areas (DPLFC leftand right, orbitofrontal cortex right), we examined thedensities of all neurons to clarify if changes in thenumber of prohibitin-immunoreactive neurons arepossibly due to alterations in the overall number ofneurons.
Prohibitin in schizophreniaK-H Smalla et al
881
Molecular Psychiatry
Statistical analysisStatistical analysis of immunoblotting experimentswas performed with Student’s t-test. Cell densities ofthe six cortical layers were analyzed using repeatedmeasures ANOVA. A Bonferroni correction wasapplied to take into account the large number ofcomparisons. A P-value of < 0.05 was regardedsignificant. The interrater reliability as determinedwith a Spearman’s correlation was 0.89.
Results
Subchronic ketamine changes levels of a few proteinsof the rat synaptic proteomeOn the basis of the hypothesis that schizophreniainvolves a hypofunction of the glutamatergic systemin the brain we choose the rat ketamine model ofschizophrenia to analyze alterations in cortical pro-tein levels with a proteomic approach. A PSD-enriched fraction was isolated biochemically andproteins were resolved using 2D gel electrophoresis.The normalized protein profiles of either controlanimals or the ketamine-treated group obtained fromeight gels for each group showed approximately 1000separate protein spots. A comparison among gels wasnot limited by the ability to match spots between casegroups and normalized gels because more than 80%of all spots were matched to the reference gel allowingexcellent comparison across all case groups.
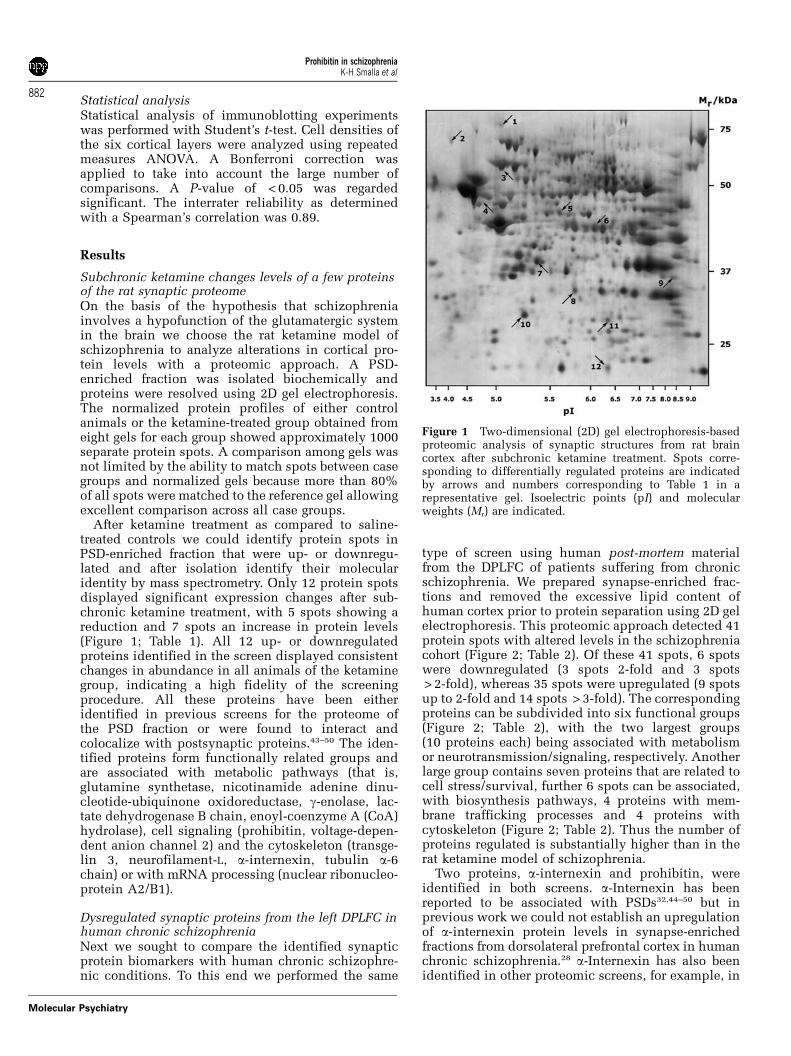
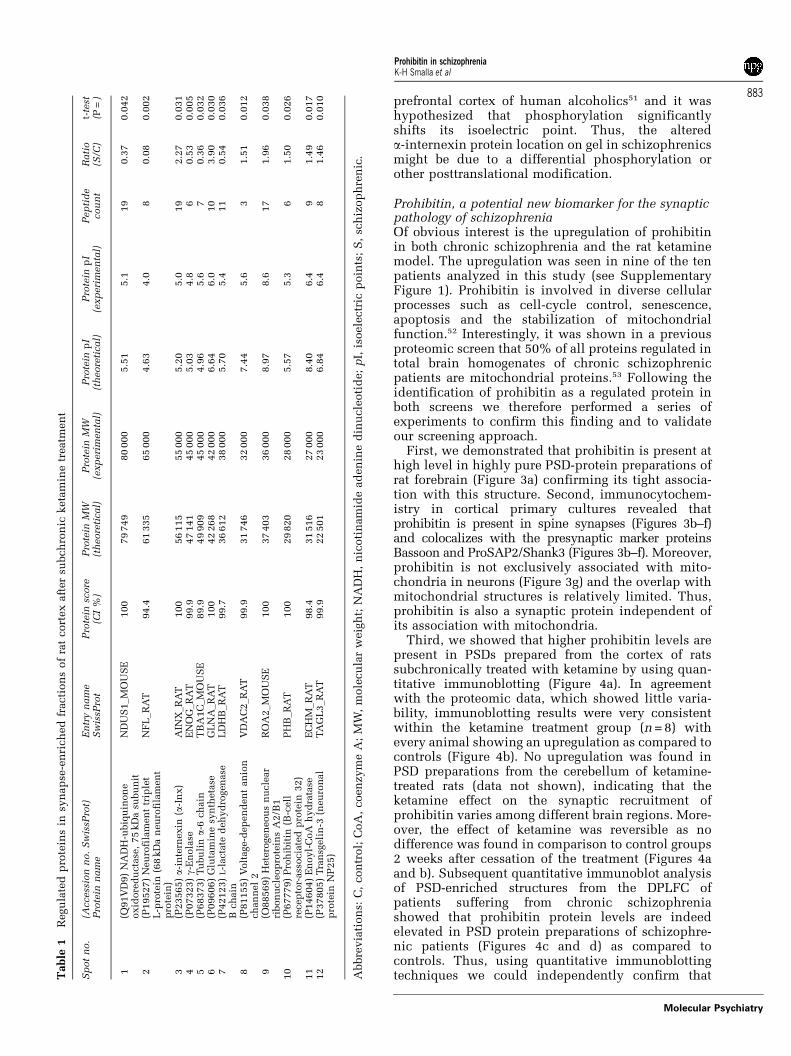
After ketamine treatment as compared to saline-treated controls we could identify protein spots inPSD-enriched fraction that were up- or downregu-lated and after isolation identify their molecularidentity by mass spectrometry. Only 12 protein spotsdisplayed significant expression changes after sub-chronic ketamine treatment, with 5 spots showing areduction and 7 spots an increase in protein levels(Figure 1; Table 1). All 12 up- or downregulatedproteins identified in the screen displayed consistentchanges in abundance in all animals of the ketaminegroup, indicating a high fidelity of the screeningprocedure. All these proteins have been eitheridentified in previous screens for the proteome ofthe PSD fraction or were found to interact andcolocalize with postsynaptic proteins.43–50 The iden-tified proteins form functionally related groups andare associated with metabolic pathways (that is,glutamine synthetase, nicotinamide adenine dinu-cleotide-ubiquinone oxidoreductase, g-enolase, lac-tate dehydrogenase B chain, enoyl-coenzyme A (CoA)hydrolase), cell signaling (prohibitin, voltage-depen-dent anion channel 2) and the cytoskeleton (transge-lin 3, neurofilament-L, a-internexin, tubulin a-6chain) or with mRNA processing (nuclear ribonucleo-protein A2/B1).
Dysregulated synaptic proteins from the left DPLFC inhuman chronic schizophreniaNext we sought to compare the identified synapticprotein biomarkers with human chronic schizophre-nic conditions. To this end we performed the same
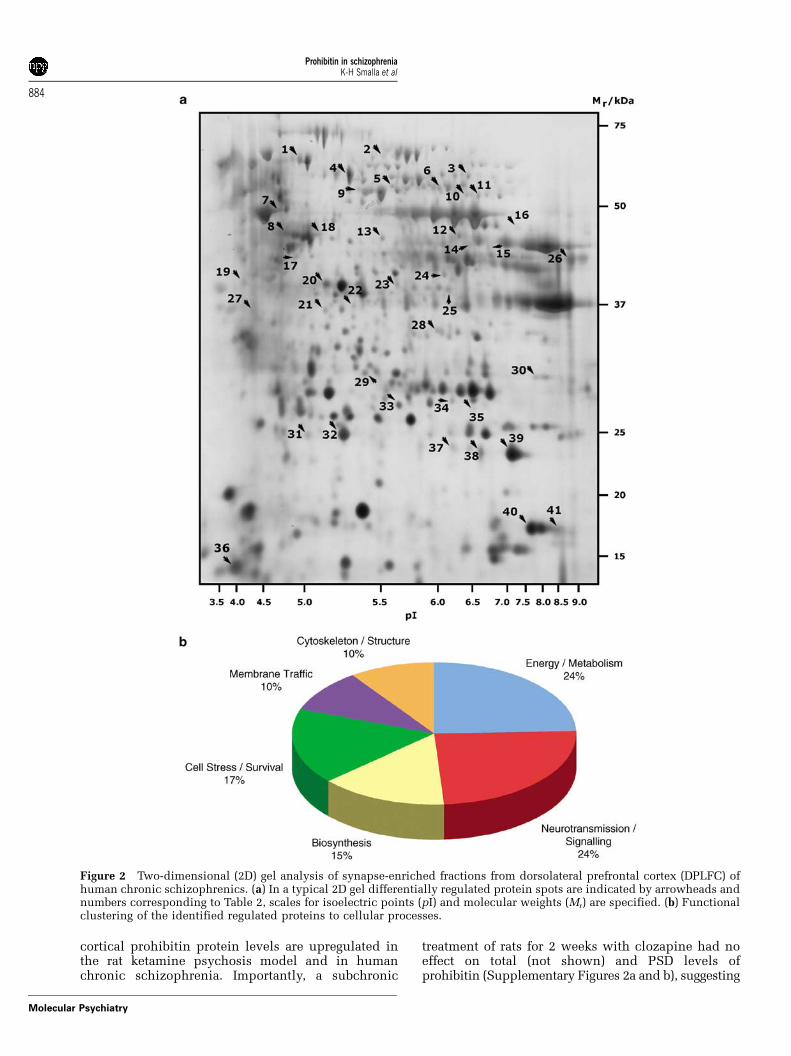
type of screen using human post-mortem materialfrom the DPLFC of patients suffering from chronicschizophrenia. We prepared synapse-enriched frac-tions and removed the excessive lipid content ofhuman cortex prior to protein separation using 2D gelelectrophoresis. This proteomic approach detected 41protein spots with altered levels in the schizophreniacohort (Figure 2; Table 2). Of these 41 spots, 6 spotswere downregulated (3 spots 2-fold and 3 spots> 2-fold), whereas 35 spots were upregulated (9 spotsup to 2-fold and 14 spots > 3-fold). The correspondingproteins can be subdivided into six functional groups(Figure 2; Table 2), with the two largest groups(10 proteins each) being associated with metabolismor neurotransmission/signaling, respectively. Anotherlarge group contains seven proteins that are related tocell stress/survival, further 6 spots can be associated,with biosynthesis pathways, 4 proteins with mem-brane trafficking processes and 4 proteins withcytoskeleton (Figure 2; Table 2). Thus the number ofproteins regulated is substantially higher than in therat ketamine model of schizophrenia.
Two proteins, a-internexin and prohibitin, wereidentified in both screens. a-Internexin has beenreported to be associated with PSDs32,44–50 but inprevious work we could not establish an upregulationof a-internexin protein levels in synapse-enrichedfractions from dorsolateral prefrontal cortex in humanchronic schizophrenia.28 a-Internexin has also beenidentified in other proteomic screens, for example, in
Figure 1 Two-dimensional (2D) gel electrophoresis-basedproteomic analysis of synaptic structures from rat braincortex after subchronic ketamine treatment. Spots corre-sponding to differentially regulated proteins are indicatedby arrows and numbers corresponding to Table 1 in arepresentative gel. Isoelectric points (pI) and molecularweights (Mr) are indicated.
Prohibitin in schizophreniaK-H Smalla et al
882
Molecular Psychiatry
prefrontal cortex of human alcoholics51 and it washypothesized that phosphorylation significantlyshifts its isoelectric point. Thus, the altereda-internexin protein location on gel in schizophrenicsmight be due to a differential phosphorylation orother posttranslational modification.
Prohibitin, a potential new biomarker for the synapticpathology of schizophreniaOf obvious interest is the upregulation of prohibitinin both chronic schizophrenia and the rat ketaminemodel. The upregulation was seen in nine of the tenpatients analyzed in this study (see SupplementaryFigure 1). Prohibitin is involved in diverse cellularprocesses such as cell-cycle control, senescence,apoptosis and the stabilization of mitochondrialfunction.52 Interestingly, it was shown in a previousproteomic screen that 50% of all proteins regulated intotal brain homogenates of chronic schizophrenicpatients are mitochondrial proteins.53 Following theidentification of prohibitin as a regulated protein inboth screens we therefore performed a series ofexperiments to confirm this finding and to validateour screening approach.
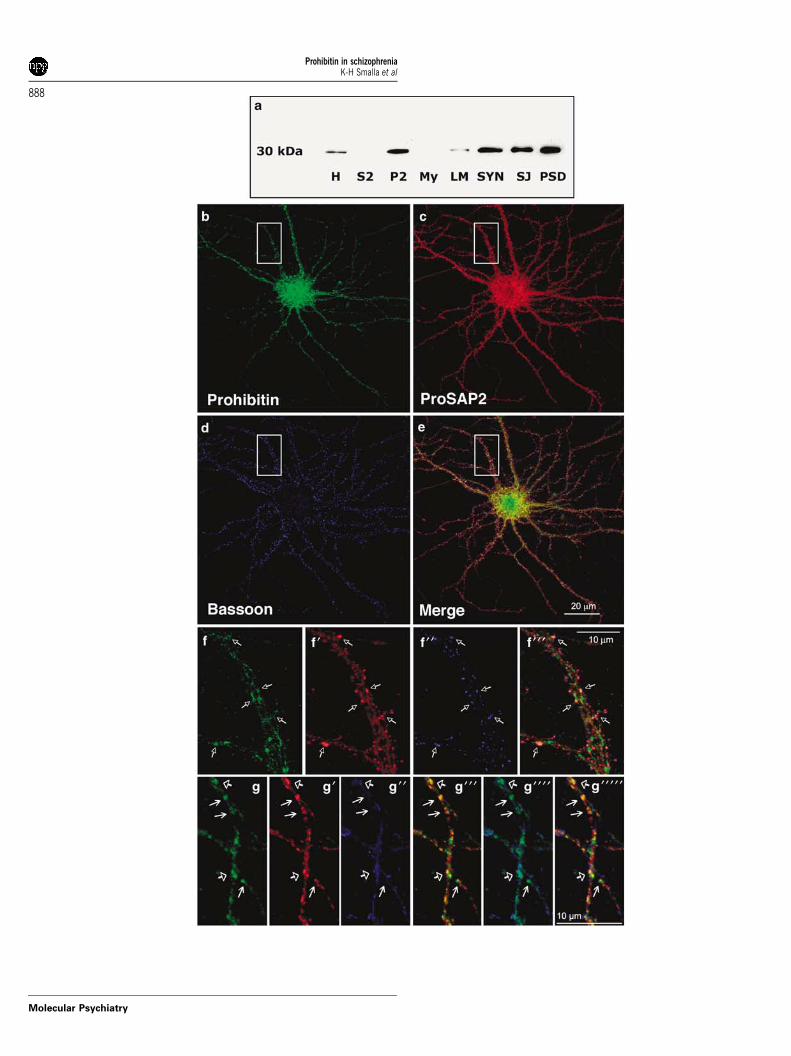
First, we demonstrated that prohibitin is present athigh level in highly pure PSD-protein preparations ofrat forebrain (Figure 3a) confirming its tight associa-tion with this structure. Second, immunocytochem-istry in cortical primary cultures revealed thatprohibitin is present in spine synapses (Figures 3b–f)and colocalizes with the presynaptic marker proteinsBassoon and ProSAP2/Shank3 (Figures 3b–f). Moreover,prohibitin is not exclusively associated with mito-chondria in neurons (Figure 3g) and the overlap withmitochondrial structures is relatively limited. Thus,prohibitin is also a synaptic protein independent ofits association with mitochondria.
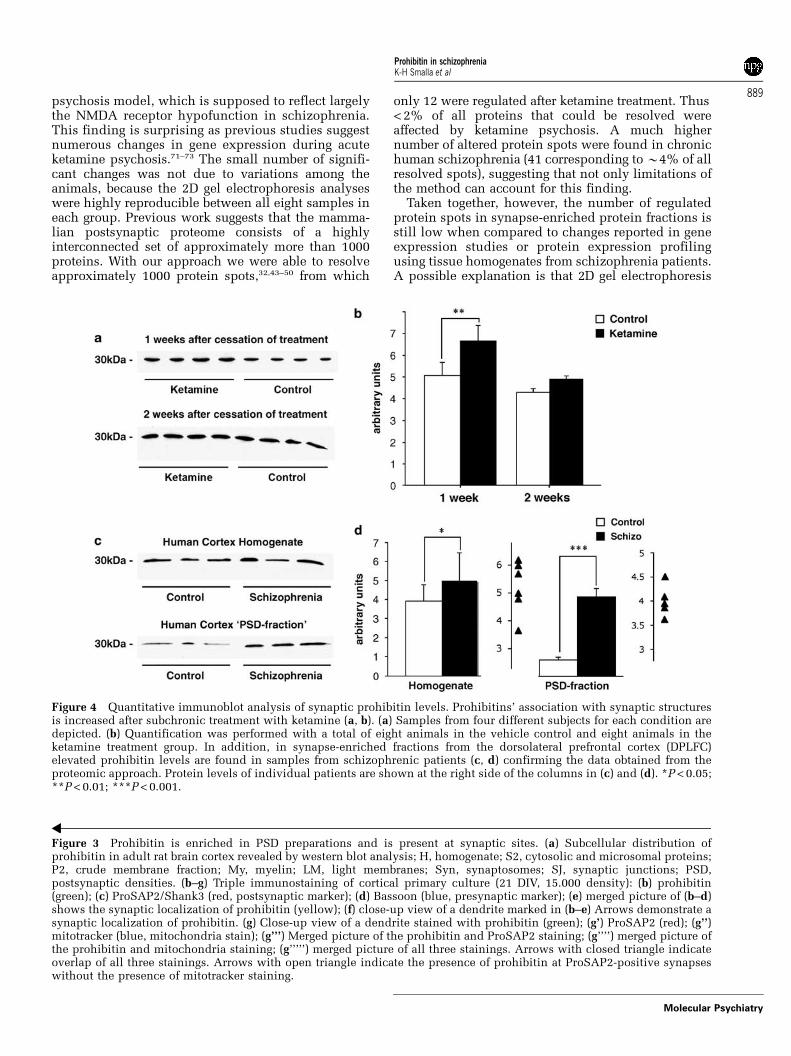
Third, we showed that higher prohibitin levels arepresent in PSDs prepared from the cortex of ratssubchronically treated with ketamine by using quan-titative immunoblotting (Figure 4a). In agreementwith the proteomic data, which showed little varia-bility, immunoblotting results were very consistentwithin the ketamine treatment group (n = 8) withevery animal showing an upregulation as compared tocontrols (Figure 4b). No upregulation was found inPSD preparations from the cerebellum of ketamine-treated rats (data not shown), indicating that theketamine effect on the synaptic recruitment ofprohibitin varies among different brain regions. More-over, the effect of ketamine was reversible as nodifference was found in comparison to control groups2 weeks after cessation of the treatment (Figures 4aand b). Subsequent quantitative immunoblot analysisof PSD-enriched structures from the DPLFC ofpatients suffering from chronic schizophreniashowed that prohibitin protein levels are indeedelevated in PSD protein preparations of schizophre-nic patients (Figures 4c and d) as compared tocontrols. Thus, using quantitative immunoblottingtechniques we could independently confirm thatT
able
1R
egu
late
dp
rote
ins
insy
nap
se-e
nri
ch
ed
fracti
on
sof
rat
cort
ex
aft
er
subch
ron
icketa
min
etr
eatm
en
t
Sp
ot
no.
(Access
ion
no.
Sw
issP
rot)
Pro
tein
nam
eE
ntr
yn
am
eS
wis
sPro
tP
rote
insc
ore
(CI
%)
Pro
tein
MW
(th
eore
tical)
Pro
tein
MW
(exp
eri
men
tal)
Pro
tein
pI
(th
eore
tical)
Pro
tein
pI
(exp
eri
men
tal)
Pep
tid
ecou
nt
Rati
o(S
/C)
t-te
st(P
=)
1(Q
91V
D9)
NA
DH
-ubiq
uin
on
eoxid
ore
du
cta
se,
75
kD
asu
bu
nit
ND
US
1_M
OU
SE
100
79
749
80
000
5.5
15.1
19
0.3
70.0
42
2(P
19527)
Neu
rofi
lam
en
ttr
iple
tL
-pro
tein
(68
kD
an
eu
rofi
lam
en
tp
rote
in)
NF
L_R
AT
94.4
61
335
65
000
4.6
34.0
80.0
80.0
02
3(P
23565)a-
inte
rnexin
(a-I
nx)
AIN
X_R
AT
100
56
115
55
000
5.2
05.0
19
2.2
70.0
31
4(P
07323)g-
En
ola
seE
NO
G_R
AT
99.9
47
141
45
000
5.0
34.8
60.5
30.0
05
5(P
68373)
Tu
bu
lina-
6ch
ain
TB
A1C
_M
OU
SE
89.9
49
909
45
000
4.9
65.6
70.3
60.0
32
6(P
09606)
Glu
tam
ine
syn
theta
seG
LN
A_R
AT
100
42
268
42
000
6.6
46.0
10
3.9
00.0
30
7(P
42123)
L-l
acta
ted
eh
yd
rogen
ase
Bch
ain
LD
HB
_R
AT
99.7
36
612
38
000
5.7
05.4
11
0.5
40.0
36
8(P
81155)
Volt
age-d
ep
en
den
tan
ion
ch
an
nel
2V
DA
C2_R
AT
99.9
31
746
32
000
7.4
45.6
31.5
10.0
12
9(O
88569)
Hete
rogen
eou
sn
ucle
ar
ribon
ucle
op
rote
ins
A2/B
1R
OA
2_M
OU
SE
100
37
403
36
000
8.9
78.6
17
1.9
60.0
38
10
(P67779)
Pro
hib
itin
(B-c
ell
recep
tor-
ass
ocia
ted
pro
tein
32)
PH
B_R
AT
100
29
820
28
000
5.5
75.3
61.5
00.0
26
11
(P14604)
En
oyl-
CoA
hyd
rata
seE
CH
M_R
AT
98.4
31
516
27
000
8.4
06.4
91.4
90.0
17
12
(P37805)
Tra
nsg
eli
n-3
(neu
ron
al
pro
tein
NP
25)
TA
GL
3_R
AT
99.9
22
501
23
000
6.8
46.4
81.4
60.0
10
Abbre
via
tion
s:C
,con
trol;
CoA
,coen
zym
eA
;M
W,
mole
cu
lar
weig
ht;
NA
DH
,n
icoti
nam
ide
ad
en
ine
din
ucle
oti
de;
pI,
isoele
ctr
icp
oin
ts;
S,
sch
izop
hre
nic
.
Prohibitin in schizophreniaK-H Smalla et al
883
Molecular Psychiatry
cortical prohibitin protein levels are upregulated inthe rat ketamine psychosis model and in humanchronic schizophrenia. Importantly, a subchronic
treatment of rats for 2 weeks with clozapine had noeffect on total (not shown) and PSD levels ofprohibitin (Supplementary Figures 2a and b), suggesting
Figure 2 Two-dimensional (2D) gel analysis of synapse-enriched fractions from dorsolateral prefrontal cortex (DPLFC) ofhuman chronic schizophrenics. (a) In a typical 2D gel differentially regulated protein spots are indicated by arrowheads andnumbers corresponding to Table 2, scales for isoelectric points (pI) and molecular weights (Mr) are specified. (b) Functionalclustering of the identified regulated proteins to cellular processes.
Prohibitin in schizophreniaK-H Smalla et al
884
Molecular Psychiatry
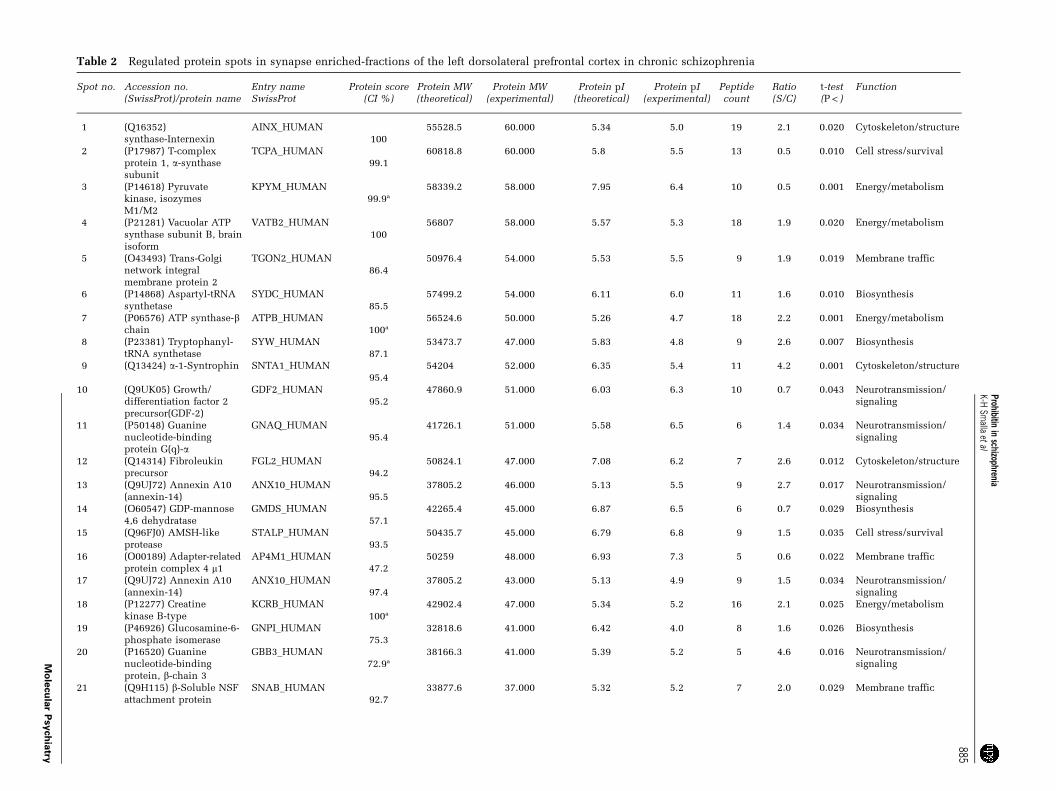
Table 2 Regulated protein spots in synapse enriched-fractions of the left dorsolateral prefrontal cortex in chronic schizophrenia
Spot no. Accession no.(SwissProt)/protein name
Entry nameSwissProt
Protein score(CI %)
Protein MW(theoretical)
Protein MW(experimental)
Protein pI(theoretical)
Protein pI(experimental)
Peptidecount
Ratio(S/C)
t-test(P < )
Function
1 (Q16352)synthase-Internexin
AINX_HUMAN100
55528.5 60.000 5.34 5.0 19 2.1 0.020 Cytoskeleton/structure
2 (P17987) T-complexprotein 1, a-synthasesubunit
TCPA_HUMAN99.1
60818.8 60.000 5.8 5.5 13 0.5 0.010 Cell stress/survival
3 (P14618) Pyruvatekinase, isozymesM1/M2
KPYM_HUMAN99.9a
58339.2 58.000 7.95 6.4 10 0.5 0.001 Energy/metabolism
4 (P21281) Vacuolar ATPsynthase subunit B, brainisoform
VATB2_HUMAN100
56807 58.000 5.57 5.3 18 1.9 0.020 Energy/metabolism
5 (O43493) Trans-Golginetwork integralmembrane protein 2
TGON2_HUMAN86.4
50976.4 54.000 5.53 5.5 9 1.9 0.019 Membrane traffic
6 (P14868) Aspartyl-tRNAsynthetase
SYDC_HUMAN85.5
57499.2 54.000 6.11 6.0 11 1.6 0.010 Biosynthesis
7 (P06576) ATP synthase-bchain
ATPB_HUMAN100a
56524.6 50.000 5.26 4.7 18 2.2 0.001 Energy/metabolism
8 (P23381) Tryptophanyl-tRNA synthetase
SYW_HUMAN87.1
53473.7 47.000 5.83 4.8 9 2.6 0.007 Biosynthesis
9 (Q13424) a-1-Syntrophin SNTA1_HUMAN95.4
54204 52.000 6.35 5.4 11 4.2 0.001 Cytoskeleton/structure
10 (Q9UK05) Growth/differentiation factor 2precursor(GDF-2)
GDF2_HUMAN95.2
47860.9 51.000 6.03 6.3 10 0.7 0.043 Neurotransmission/signaling
11 (P50148) Guaninenucleotide-bindingprotein G(q)-a
GNAQ_HUMAN95.4
41726.1 51.000 5.58 6.5 6 1.4 0.034 Neurotransmission/signaling
12 (Q14314) Fibroleukinprecursor
FGL2_HUMAN94.2
50824.1 47.000 7.08 6.2 7 2.6 0.012 Cytoskeleton/structure
13 (Q9UJ72) Annexin A10(annexin-14)
ANX10_HUMAN95.5
37805.2 46.000 5.13 5.5 9 2.7 0.017 Neurotransmission/signaling
14 (O60547) GDP-mannose4,6 dehydratase
GMDS_HUMAN57.1
42265.4 45.000 6.87 6.5 6 0.7 0.029 Biosynthesis
15 (Q96FJ0) AMSH-likeprotease
STALP_HUMAN93.5
50435.7 45.000 6.79 6.8 9 1.5 0.035 Cell stress/survival
16 (O00189) Adapter-relatedprotein complex 4 m1
AP4M1_HUMAN47.2
50259 48.000 6.93 7.3 5 0.6 0.022 Membrane traffic
17 (Q9UJ72) Annexin A10(annexin-14)
ANX10_HUMAN97.4
37805.2 43.000 5.13 4.9 9 1.5 0.034 Neurotransmission/signaling
18 (P12277) Creatinekinase B-type
KCRB_HUMAN100a
42902.4 47.000 5.34 5.2 16 2.1 0.025 Energy/metabolism
19 (P46926) Glucosamine-6-phosphate isomerase
GNPI_HUMAN75.3
32818.6 41.000 6.42 4.0 8 1.6 0.026 Biosynthesis
20 (P16520) Guaninenucleotide-bindingprotein, b-chain 3
GBB3_HUMAN72.9a
38166.3 41.000 5.39 5.2 5 4.6 0.016 Neurotransmission/signaling
21 (Q9H115) b-Soluble NSFattachment protein
SNAB_HUMAN92.7
33877.6 37.000 5.32 5.2 7 2.0 0.029 Membrane traffic
Prohibitinin
schizophreniaK-H
Smalla
etal
885
Mo
lecu
lar
Psych
iatry
Table 2 Continued
Spot no. Accession no.(SwissProt)/protein name
Entry nameSwissProt
Protein score(CI %)
Protein MW(theoretical)
Protein MW(experimental)
Protein pI(theoretical)
Protein pI(experimental)
Peptidecount
Ratio(S/C)
t-test(P < )
Function
22 (Q9H2U2) Inorganicpyrophosphatase 2
IPYR2_HUMAN70.3
38451.2 37.000 7.07 5.4 8 1.8 0.035 Energy/metabolism
23 (Q9Y5K3) Choline-phosphatecytidylyltransferase B
PCY1B_HUMAN90.8
42198.9 41.000 5.99 5.7 8 1.8 0.029 Biosynthesis
24 (P09972) Fructose-bisphosphate aldolase C
ALDOC_HUMAN100
39699.4 42.000 6.46 6.1 13 1.9 0.021 Energy/metabolism
25 (Q9H2U2) Inorganicpyrophosphatase 2
IPYR2_HUMAN97.2
38451.2 40.000 7.07 6.1 9 1.6 0.033 Energy/metabolism
26 (P07355) Annexin A2 ANXA2_HUMAN94.4
38676.9 44.000 7.56 8.6 10 0.7 0.019 Neurotransmission/signaling
27 (P51858) Hepatoma-derived growth factor(HDGF)
HDGF_HUMAN97.2
26885.9 36.000 4.7 4.3 6 1.5 0.016 Neurotransmission/signaling
28 (Q15773) Myeloidleukemia factor 2
MLF2_HUMAN99.4a
28186.2 35.000 6.4 6.0 7 2.9 0.020 Neurotransmission/signaling
29 (P35232) Prohibitin PHB_HUMAN97.8
29804.1 30.000 5.57 5.4 7 1.6 0.002 Multiple, biosynthesis
30 (P27144) Adenylatekinase isoenzyme 4
KAD4_HUMAN99.9
25366.2 29.000 8.47 7.8 10 2.2 0.034 Energy/metabolism
31 (O75947) ATP synthaseD chain
ATP5H_HUMAN100a
18405.5 25.000 5.22 5.0 12 2.2 0.013 Energy/metabolism
32 (P32119)Peroxiredoxin 2
PRDX2_HUMAN93.1a
21918.3 25.000 5.67 5.3 4 2.0 0.027 Cell stress/survival
33 (P30048)Peroxiredoxin 3,
PRDX3_HUMAN100a
28017.3 27.000 7.67 5.7 6 1.6 0.016 Cell stress/survival
34 (P21796) Voltage-dependent anion-selectivechannel 1
VDAC1_HUMAN91.8a
30736.6 28.000 8.63 6.2 4 1.4 0.011 Energy/metabolism
35 (P01213) b-Neoendorphin-dynorphin precursor
PDYN_HUMAN93.5
28766.3 25.000 6.07 6.4 7 2.0 0.012 Neurotransmission/signaling
36 (P20472) Parvalbumin-a PRVA_HUMAN99.6
12058.8 14.000 4.98 3.7 8 1.6 0.025 Cell stress/survival
37 (P02511) a-Crystallin Bchain (a(B)-crystallin)
CRYAB_HUMAN99.9a
20146.4 24.000 6.76 6.2 9 2.0 0.001 Cell stress/survival
38 (P02511) a-Crystallin Bchain (a(B)-crystallin)
CRYAB_HUMAN100a
20146.4 23.000 6.76 6.5 9 1.8 0.016 Cell stress/survival
39 (P30086) Phosphatidy-lethanolamine-bindingprotein
PEBP1_HUMAN100a
21026.7 24.000 7.42 7.2 9 1.8 0.040 Neurotransmission/signaling
40 (P23528) Cofilin-1 COF1_HUMAN99.9
18587.7 18.000 8.26 7.7 9 1.5 0.022 Cytoskeleton/structure
41 (O60493) Sorting nexin-3(SDP3 protein)
SNX3_HUMAN100a
18676.7 18.000 8.73 8.5 8 1.6 0.007 Membrane traffic
Abbreviations: AMSH, associated molecule with the SH3 domain; C, control; MW, molecular weight; NSF, N-ethylmaleimide-sensitive factor; pI, isoelectric points;S, schizophrenic.aMS/MS identification with total CI % ion > 90%.
Prohibitinin
schizophreniaK-H
Smalla
etal
886
Mo
lecu
lar
Psych
iatry

that the treatment with neuroleptics is not causallyrelated to the regulation of prohibitin protein levels.
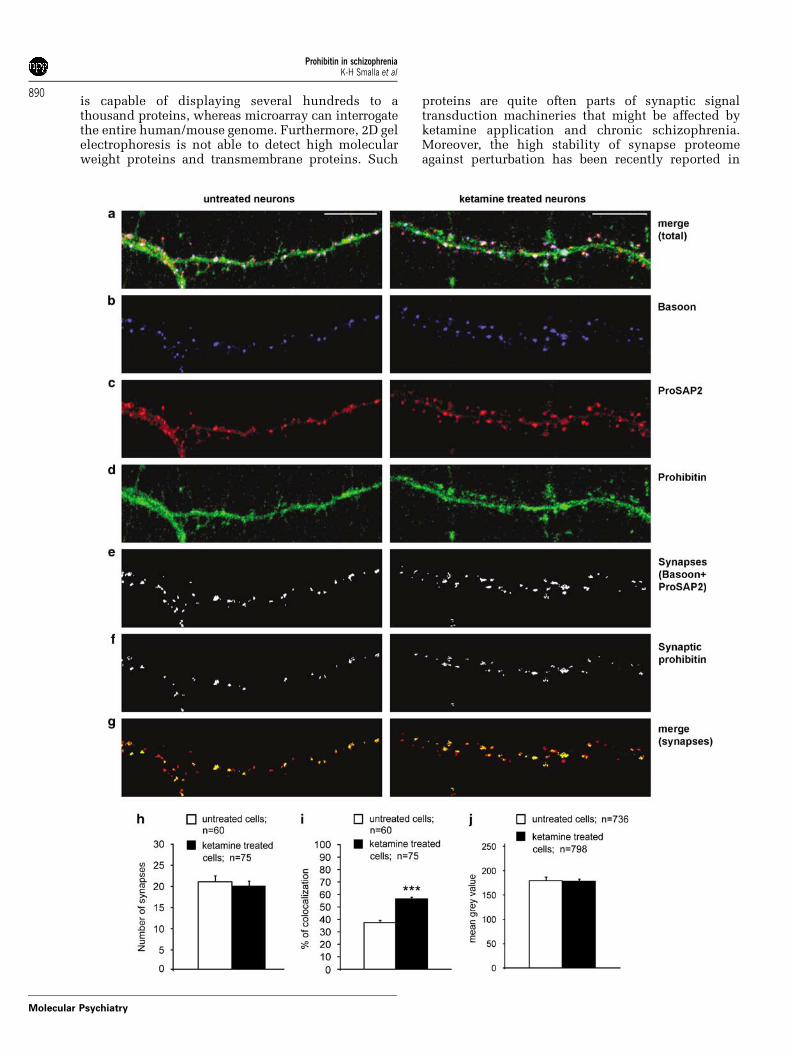
Prohibitin in cortical primary culturesFurther support for the hypothesis that prohibitinassociation with the synapse depends upon NMDAreceptor activity was found in studies using corticalprimary cultures. Application of ketamine to themedia with subsequent blockage of NMDA receptorsfor 48 h resulted in profoundly increased synapticprohibitin immunofluorescence as compared tocontrols (Figures 5a–g). This was due to the increasednumber of synapses containing prohibitin (Figure 5i),whereas the total synaptic staining intensity forprohibitin was unchanged (Figure 5j). Ketaminetreatment had no influence on the number ofsynapses in cortical primary cultures (Figure 5h).
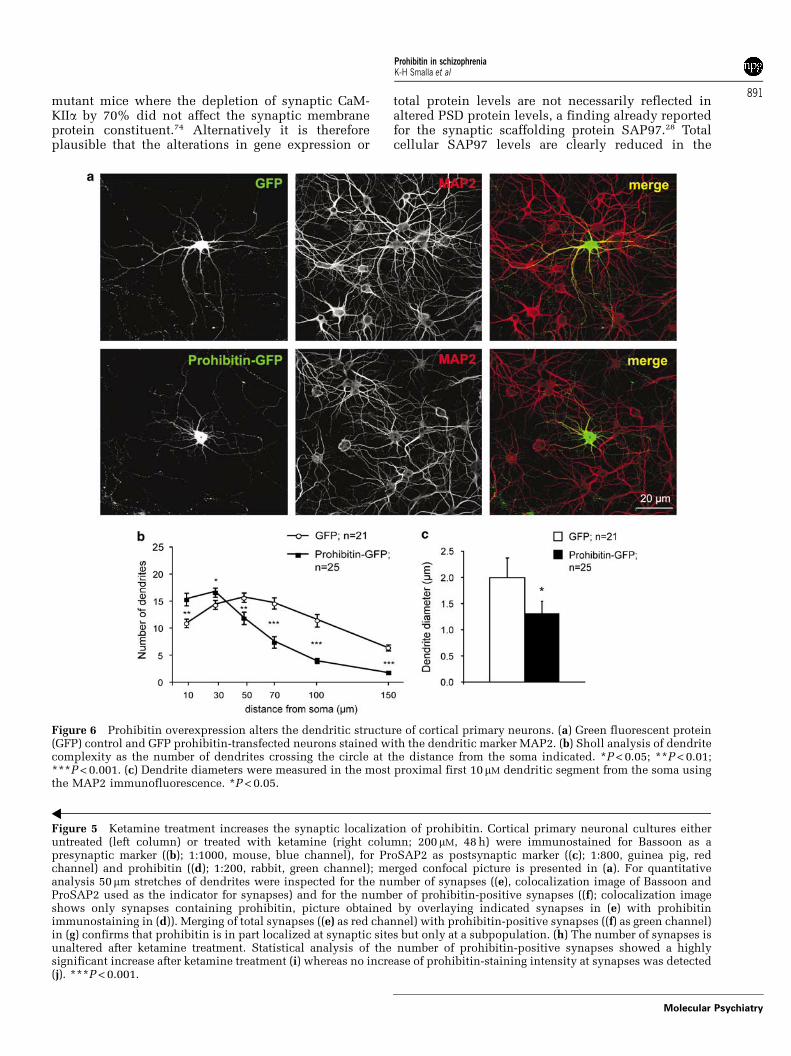
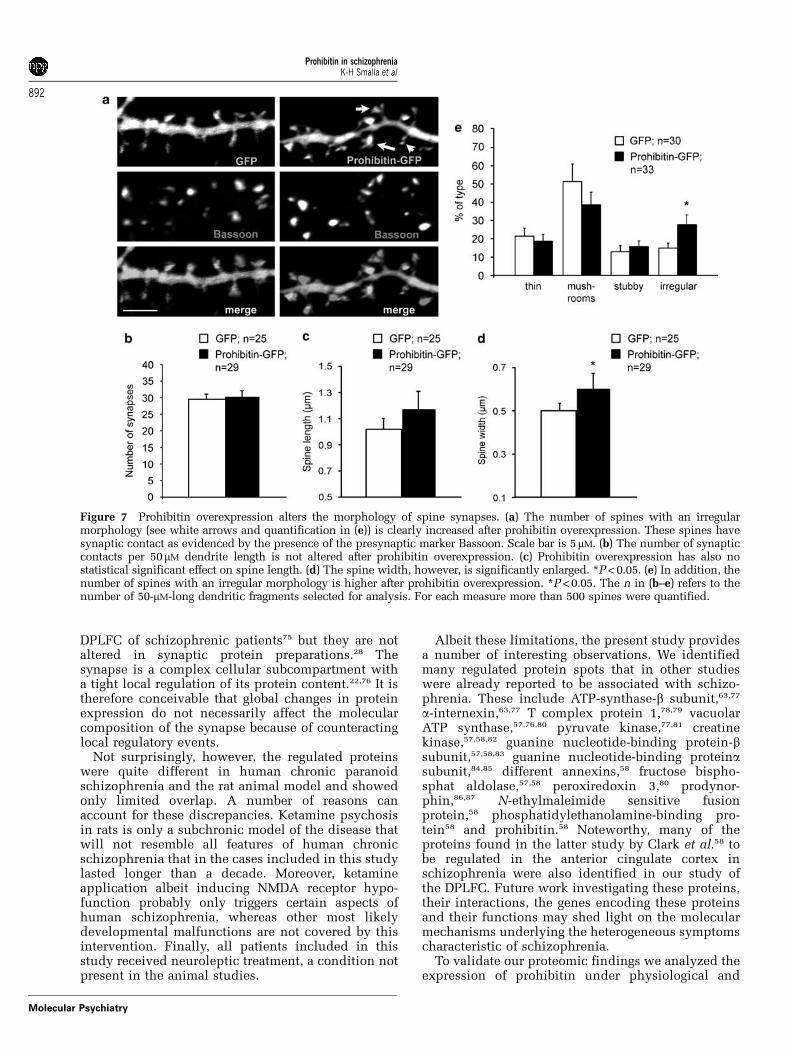
To address the question whether elevated prohibi-tin levels might induce synapto-dendritic phenotypesthat resemble alterations seen in schizophrenia wethen induced overexpression of a GFP–prohibitinfusion protein during development of corticalprimary neurons in culture. Subsequent Sholl analy-sis revealed that dendrites from GFP-prohibitintransfected neurons have shorter dendrites thancorresponding control neurons (Figures 6a and b).Following prohibitin overexpression proximal den-drites are also much thinner than in GFP-controlneurons (Figure 6c). Interestingly, albeit shorter andthinner these dendrites are in their proximal segmentvery bushy and complex (Figures 6a and b). Theaverage dendritic synapse density is unchanged inGFP-prohibitin transfected neurons as compared toGFP controls (Figures 7a and b) and also the spinelength was not statistically significant altered (Figure 7c).However, the average spine width as evidenced bythe GFP fluorescence is significantly enlarged(Figure 7d). This enlargement is correlated with anincreased number of spines with an irregular shape(Figure 7e). Taken together these data indicate thatsome features of the synapto-dendritic alterations incertain brain regions in chronic schizophrenia areresembled by the overexpression of prohibitin inprimary neurons.
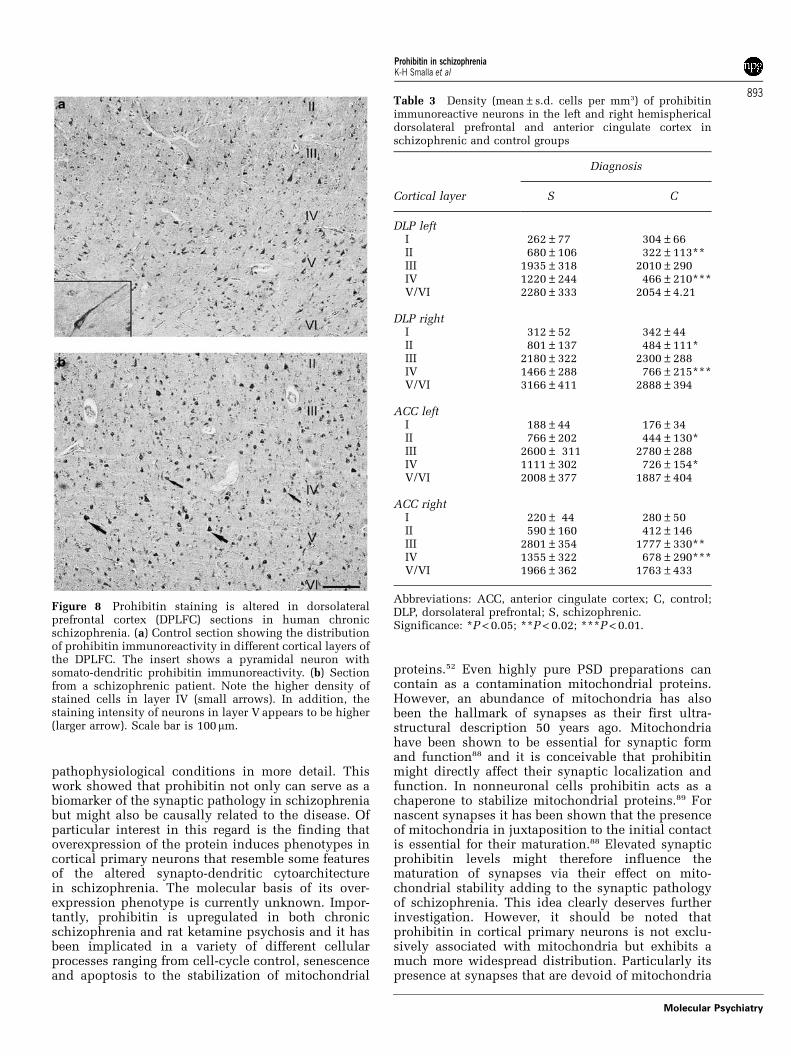
Prohibitin in post-mortem brainsIn the next series of experiments we examined thedistribution of prohibitin immunoreactivity in paraf-fin-embedded human brain tissue sections from theDPLFC. It should be emphasized that paraffinsections usually do not allow to detect a synapticlocalization of a given protein and we could indeedonly observe a somato-dendritic staining pattern.Immunostaining of human cortical sections revealedabundant localization of prohibitin to neuronal cellbodies and dendrites in all layers of the cerebralcortex but predominantly in layers III and V (Figure 8).Large pyramidal and fusiform neurons wereprominently labeled. Interneurons situated in layersII, IV and VI were less frequently stained (Figure 8). Inbrains of schizophrenics, we observed a different
staining pattern in that the relative paucity ofimmunostained neurons in layer IV typically seen incontrols was not detectable (Figure 8). Moreover,staining intensity layer V pyramidal neurons ap-peared to be increased (Figure 8). This qualitativedifference was substantiated by layer-specific quanti-tative assessment of neuronal prohibitin-immunopo-sitive neurons. Quantitative morphometric analysisrevealed that changes particularly occurred in layerIV of tissue sections from schizophrenic brains(Figure 8; Table 3). No significant changes were foundin the number of cells in the different cortical layersbetween control and schizophrenic brains (Table 4).Although it is at present unclear how this altereddistribution relates to the synaptic localization ofprohibitin at the cellular level it points to a redis-tribution of protein levels within the DPLFC.
Discussion
The present report is concerned with the first screenof the synaptic proteome in human chronic schizo-phrenia and the rat animal model of ketamine-induced psychosis. Subtle structural alterations inthe thalamus, hippocampus and cortex have beenassociated with schizophrenia and particularly theleft dorsolateral prefrontal cortex was reported toexhibit more prominent morphological abnormal-ities.8,13,16,54–56 To gain better insight into the mole-cular basis of schizophrenia a number of proteomicscreens on post-mortem brain in schizophrenia thereforehave been performed using these brain regions.57–62
In addition, corresponding screens in animal modelsof psychosis have been published.63 Complementarywork has been carried out with gene expressionprofiling of schizophrenic human brain and corre-sponding animal models.64–68 A major shortcoming ofthese approaches is that although they focus onregion-specific differences, they detect rather globalchanges in brain protein composition rather thandrawing attention to the organelles that are mostlikely affected in schizophrenia. Thus, proteomicscreens of schizophrenia-related protein alterationsin protein homogenates are corrupted by the fact thatthe majority of cells that will be looked at are glialcells and not neurons. In particular, the importantprocesses related to the synaptic pathology of schizo-phrenia might have been overlooked so far.
The PSD is a specialized protein matrix observed inprimarily excitatory synapses and is crucially in-volved in (1) the anchoring of glutamate receptors aswell as synaptic cell adhesion molecules, (2) thetopological organization of signal transduction path-ways and finally (3) the dynamics of synapsemorphology by its connection to the spine cytoskele-ton.21,22 Thus, the PSD has to be considered as apotential target of structural alterations in schizo-phrenia that will have fundamental functional con-sequences. In light of the reported synaptic pathologyof schizophrenia3,10,69,70 we were surprised to findonly a few regulated spots in the rat ketamine
Prohibitin in schizophreniaK-H Smalla et al
887
Molecular Psychiatry
Prohibitin in schizophreniaK-H Smalla et al
888
Molecular Psychiatry
psychosis model, which is supposed to reflect largelythe NMDA receptor hypofunction in schizophrenia.This finding is surprising as previous studies suggestnumerous changes in gene expression during acuteketamine psychosis.71–73 The small number of signifi-cant changes was not due to variations among theanimals, because the 2D gel electrophoresis analyseswere highly reproducible between all eight samples ineach group. Previous work suggests that the mamma-lian postsynaptic proteome consists of a highlyinterconnected set of approximately more than 1000proteins. With our approach we were able to resolveapproximately 1000 protein spots,32,43–50 from which
only 12 were regulated after ketamine treatment. Thus< 2% of all proteins that could be resolved wereaffected by ketamine psychosis. A much highernumber of altered protein spots were found in chronichuman schizophrenia (41 corresponding to B4% of allresolved spots), suggesting that not only limitations ofthe method can account for this finding.
Taken together, however, the number of regulatedprotein spots in synapse-enriched protein fractions isstill low when compared to changes reported in geneexpression studies or protein expression profilingusing tissue homogenates from schizophrenia patients.A possible explanation is that 2D gel electrophoresis
Figure 3 Prohibitin is enriched in PSD preparations and is present at synaptic sites. (a) Subcellular distribution ofprohibitin in adult rat brain cortex revealed by western blot analysis; H, homogenate; S2, cytosolic and microsomal proteins;P2, crude membrane fraction; My, myelin; LM, light membranes; Syn, synaptosomes; SJ, synaptic junctions; PSD,postsynaptic densities. (b–g) Triple immunostaining of cortical primary culture (21 DIV, 15.000 density): (b) prohibitin(green); (c) ProSAP2/Shank3 (red, postsynaptic marker); (d) Bassoon (blue, presynaptic marker); (e) merged picture of (b–d)shows the synaptic localization of prohibitin (yellow); (f) close-up view of a dendrite marked in (b–e) Arrows demonstrate asynaptic localization of prohibitin. (g) Close-up view of a dendrite stained with prohibitin (green); (g’) ProSAP2 (red); (g’’)mitotracker (blue, mitochondria stain); (g’’’) Merged picture of the prohibitin and ProSAP2 staining; (g’’’’) merged picture ofthe prohibitin and mitochondria staining; (g’’’’’) merged picture of all three stainings. Arrows with closed triangle indicateoverlap of all three stainings. Arrows with open triangle indicate the presence of prohibitin at ProSAP2-positive synapseswithout the presence of mitotracker staining.
Figure 4 Quantitative immunoblot analysis of synaptic prohibitin levels. Prohibitins’ association with synaptic structuresis increased after subchronic treatment with ketamine (a, b). (a) Samples from four different subjects for each condition aredepicted. (b) Quantification was performed with a total of eight animals in the vehicle control and eight animals in theketamine treatment group. In addition, in synapse-enriched fractions from the dorsolateral prefrontal cortex (DPLFC)elevated prohibitin levels are found in samples from schizophrenic patients (c, d) confirming the data obtained from theproteomic approach. Protein levels of individual patients are shown at the right side of the columns in (c) and (d). *P < 0.05;**P < 0.01; ***P < 0.001.
Prohibitin in schizophreniaK-H Smalla et al
889
Molecular Psychiatry
is capable of displaying several hundreds to athousand proteins, whereas microarray can interrogatethe entire human/mouse genome. Furthermore, 2D gelelectrophoresis is not able to detect high molecularweight proteins and transmembrane proteins. Such
proteins are quite often parts of synaptic signaltransduction machineries that might be affected byketamine application and chronic schizophrenia.Moreover, the high stability of synapse proteomeagainst perturbation has been recently reported in
Prohibitin in schizophreniaK-H Smalla et al
890
Molecular Psychiatry
mutant mice where the depletion of synaptic CaM-KIIa by 70% did not affect the synaptic membraneprotein constituent.74 Alternatively it is thereforeplausible that the alterations in gene expression or
total protein levels are not necessarily reflected inaltered PSD protein levels, a finding already reportedfor the synaptic scaffolding protein SAP97.28 Totalcellular SAP97 levels are clearly reduced in the
Figure 5 Ketamine treatment increases the synaptic localization of prohibitin. Cortical primary neuronal cultures eitheruntreated (left column) or treated with ketamine (right column; 200mM, 48 h) were immunostained for Bassoon as apresynaptic marker ((b); 1:1000, mouse, blue channel), for ProSAP2 as postsynaptic marker ((c); 1:800, guinea pig, redchannel) and prohibitin ((d); 1:200, rabbit, green channel); merged confocal picture is presented in (a). For quantitativeanalysis 50mm stretches of dendrites were inspected for the number of synapses ((e), colocalization image of Bassoon andProSAP2 used as the indicator for synapses) and for the number of prohibitin-positive synapses ((f); colocalization imageshows only synapses containing prohibitin, picture obtained by overlaying indicated synapses in (e) with prohibitinimmunostaining in (d)). Merging of total synapses ((e) as red channel) with prohibitin-positive synapses ((f) as green channel)in (g) confirms that prohibitin is in part localized at synaptic sites but only at a subpopulation. (h) The number of synapses isunaltered after ketamine treatment. Statistical analysis of the number of prohibitin-positive synapses showed a highlysignificant increase after ketamine treatment (i) whereas no increase of prohibitin-staining intensity at synapses was detected(j). ***P < 0.001.
Figure 6 Prohibitin overexpression alters the dendritic structure of cortical primary neurons. (a) Green fluorescent protein(GFP) control and GFP prohibitin-transfected neurons stained with the dendritic marker MAP2. (b) Sholl analysis of dendritecomplexity as the number of dendrites crossing the circle at the distance from the soma indicated. *P < 0.05; **P < 0.01;***P < 0.001. (c) Dendrite diameters were measured in the most proximal first 10mM dendritic segment from the soma usingthe MAP2 immunofluorescence. *P < 0.05.
Prohibitin in schizophreniaK-H Smalla et al
891
Molecular Psychiatry
DPLFC of schizophrenic patients75 but they are notaltered in synaptic protein preparations.28 Thesynapse is a complex cellular subcompartment witha tight local regulation of its protein content.22,76 It istherefore conceivable that global changes in proteinexpression do not necessarily affect the molecularcomposition of the synapse because of counteractinglocal regulatory events.
Not surprisingly, however, the regulated proteinswere quite different in human chronic paranoidschizophrenia and the rat animal model and showedonly limited overlap. A number of reasons canaccount for these discrepancies. Ketamine psychosisin rats is only a subchronic model of the disease thatwill not resemble all features of human chronicschizophrenia that in the cases included in this studylasted longer than a decade. Moreover, ketamineapplication albeit inducing NMDA receptor hypo-function probably only triggers certain aspects ofhuman schizophrenia, whereas other most likelydevelopmental malfunctions are not covered by thisintervention. Finally, all patients included in thisstudy received neuroleptic treatment, a condition notpresent in the animal studies.
Albeit these limitations, the present study providesa number of interesting observations. We identifiedmany regulated protein spots that in other studieswere already reported to be associated with schizo-phrenia. These include ATP-synthase-b subunit,63,77
a-internexin,63,77 T complex protein 1,78,79 vacuolarATP synthase,57,76,80 pyruvate kinase,77,81 creatinekinase,57,58,82 guanine nucleotide-binding protein-bsubunit,57,58,83 guanine nucleotide-binding proteinasubunit,84,85 different annexins,58 fructose bispho-sphat aldolase,57,58 peroxiredoxin 3,80 prodynor-phin,86,87 N-ethylmaleimide sensitive fusionprotein,58 phosphatidylethanolamine-binding pro-tein58 and prohibitin.58 Noteworthy, many of theproteins found in the latter study by Clark et al.58 tobe regulated in the anterior cingulate cortex inschizophrenia were also identified in our study ofthe DPLFC. Future work investigating these proteins,their interactions, the genes encoding these proteinsand their functions may shed light on the molecularmechanisms underlying the heterogeneous symptomscharacteristic of schizophrenia.
To validate our proteomic findings we analyzed theexpression of prohibitin under physiological and
Figure 7 Prohibitin overexpression alters the morphology of spine synapses. (a) The number of spines with an irregularmorphology (see white arrows and quantification in (e)) is clearly increased after prohibitin overexpression. These spines havesynaptic contact as evidenced by the presence of the presynaptic marker Bassoon. Scale bar is 5mM. (b) The number of synapticcontacts per 50mM dendrite length is not altered after prohibitin overexpression. (c) Prohibitin overexpression has also nostatistical significant effect on spine length. (d) The spine width, however, is significantly enlarged. *P < 0.05. (e) In addition, thenumber of spines with an irregular morphology is higher after prohibitin overexpression. *P < 0.05. The n in (b–e) refers to thenumber of 50-mM-long dendritic fragments selected for analysis. For each measure more than 500 spines were quantified.
Prohibitin in schizophreniaK-H Smalla et al
892
Molecular Psychiatry
pathophysiological conditions in more detail. Thiswork showed that prohibitin not only can serve as abiomarker of the synaptic pathology in schizophreniabut might also be causally related to the disease. Ofparticular interest in this regard is the finding thatoverexpression of the protein induces phenotypes incortical primary neurons that resemble some featuresof the altered synapto-dendritic cytoarchitecturein schizophrenia. The molecular basis of its over-expression phenotype is currently unknown. Impor-tantly, prohibitin is upregulated in both chronicschizophrenia and rat ketamine psychosis and it hasbeen implicated in a variety of different cellularprocesses ranging from cell-cycle control, senescenceand apoptosis to the stabilization of mitochondrial
proteins.52 Even highly pure PSD preparations cancontain as a contamination mitochondrial proteins.However, an abundance of mitochondria has alsobeen the hallmark of synapses as their first ultra-structural description 50 years ago. Mitochondriahave been shown to be essential for synaptic formand function88 and it is conceivable that prohibitinmight directly affect their synaptic localization andfunction. In nonneuronal cells prohibitin acts as achaperone to stabilize mitochondrial proteins.89 Fornascent synapses it has been shown that the presenceof mitochondria in juxtaposition to the initial contactis essential for their maturation.88 Elevated synapticprohibitin levels might therefore influence thematuration of synapses via their effect on mito-chondrial stability adding to the synaptic pathologyof schizophrenia. This idea clearly deserves furtherinvestigation. However, it should be noted thatprohibitin in cortical primary neurons is not exclu-sively associated with mitochondria but exhibits amuch more widespread distribution. Particularly itspresence at synapses that are devoid of mitochondria
Figure 8 Prohibitin staining is altered in dorsolateralprefrontal cortex (DPLFC) sections in human chronicschizophrenia. (a) Control section showing the distributionof prohibitin immunoreactivity in different cortical layers ofthe DPLFC. The insert shows a pyramidal neuron withsomato-dendritic prohibitin immunoreactivity. (b) Sectionfrom a schizophrenic patient. Note the higher density ofstained cells in layer IV (small arrows). In addition, thestaining intensity of neurons in layer V appears to be higher(larger arrow). Scale bar is 100mm.
Table 3 Density (mean±s.d. cells per mm3) of prohibitinimmunoreactive neurons in the left and right hemisphericaldorsolateral prefrontal and anterior cingulate cortex inschizophrenic and control groups
Diagnosis
Cortical layer S C
DLP leftI 262±77 304±66II 680±106 322±113**III 1935±318 2010±290IV 1220±244 466±210***V/VI 2280±333 2054±4.21
DLP rightI 312±52 342±44II 801±137 484±111*III 2180±322 2300±288IV 1466±288 766±215***V/VI 3166±411 2888±394
ACC leftI 188±44 176±34II 766±202 444±130*III 2600± 311 2780±288IV 1111±302 726±154*V/VI 2008±377 1887±404
ACC rightI 220± 44 280±50II 590±160 412±146III 2801±354 1777±330**IV 1355±322 678±290***V/VI 1966±362 1763±433
Abbreviations: ACC, anterior cingulate cortex; C, control;DLP, dorsolateral prefrontal; S, schizophrenic.Significance: *P < 0.05; **P < 0.02; ***P < 0.01.
Prohibitin in schizophreniaK-H Smalla et al
893
Molecular Psychiatry

suggests that the protein fulfills functions in synapticprocesses independent of its role as a mitochondrialchaperone.
Although a specific role of prohibitin in synapseshas not yet been established, a recent report byRajalingam et al.90 has shown that prohibitin isdirectly involved in the localization of the adhesioncomplex proteins cadherin and �-catenin to theplasma membrane. In the absence of prohibitin,cadherin and �-catenin stabilize adherens junctionsand cell migration.90 Adherens junctions are specia-lized zones of cell–cell contact that resemble inseveral aspects the connection of the pre- andpostsynapse. In a neuronal context one might there-fore speculate that prohibitin is a regulator of synapticcell–cell contacts and dendrite or spine growth andlimits synapse number and dendrite complexity bylocal signaling events.
Conclusions
Proteomic analysis of the DPLFC in human chronicschizophrenia and of the cortex in rat ketamine-induced psychosis revealed consistent changes in thesynaptic proteome under both conditions. However,
there was only limited overlap between both screensquestioning the face validity of the rat model. Despitethis limited overlap subsequent analysis confirmsthat prohibitins’ association with synapses couldestablish a new biomarker for schizophrenia. Weshow that prohibitins’ association with the synapseincreases with decreased NMDA receptor activityinduced by ketamine, which is in line with theobserved increased levels of the protein in PSDpreparations of patients suffering from chronic schi-zophrenia. This is consistent with the hypothesis thata hypofunction of NMDA receptor signaling iscausally related to the disease. Taken together, thecurrent study in conjunction with previous work58
establishes prohibitin as a novel and promisingcandidate molecule for the pathogenesis of schizo-phrenia.
Acknowledgments
This study was supported by grants from the BMBF(01GZ0307/ 01GA0505), DFG (SFB 779 TP B8þB9),LSA (N1/TP4, N2 TP5), Schram Foundation, LeibnizSociety (Pakt fur Forschung) and Fonds der Che-mischen Industrie to EDG and MRK, and Center forMedical Systems Biology (CMSB) to KWL and ABS.JS is supported by a stipend from the DFG graduateprogram (GRK 1167). We gratefully acknowledge theprofessional technical assistance of C Borutzki, HDobrowolny, S Funke, M Marunde and K Pohlmann.
References
1 Harrison PJ. The neuropathology of schizophrenia. A critical reviewof the data and their interpretation. Brain 1999; 122: 593–624.
2 Kaufmann WE, Moser HW. Dendritic anomalies in disordersassociated with mental retardation. Cereb Cortex 2000; 10: 981–991.
3 Eastwood SL. The synaptic pathology of schizophrenia: is aberrantneurodevelopment and plasticity to blame? Int Rev Neurobiol2004; 59: 47–72.
4 Lewis DA, Levitt P. Schizophrenia as a disorder of neurodevelop-ment. Annu Rev Neurosci 2002; 25: 409–432.
5 McGlashan TH, Hoffman RE. Schizophrenia as a disorder ofdevelopmentally reduced synaptic connectivity. Arch Gen Psy-chiatry 2000; 57: 637–648.
6 Woods BT. Is schizophrenia a progressive neurodevelopmentaldisorder? Toward a unitary pathogenetic mechanism. Am JPsychiatry 1998; 155: 1661–1670.
7 Selemon LD, Mrzljak J, Kleinman JE, Herman MM, Goldman-RakicPS. Regional specificity in the neuropathologic substrates ofschizophrenia: a morphometric analysis of Broca’s area 44 andarea 9. Arch Gen Psychiatry 2003; 60: 69–77.
8 Black JE, Kodish IM, Grossman AW, Klintsova AY, Orlovskaya D,Vostrikov V et al. Pathology of layer V pyramidal neurons in theprefrontal cortex of patients with schizophrenia. Am J Psychiatry2004; 161: 742–744.
9 Garey LJ, Ong WY, Patel TS, Kanani M, Davis A, Mortimer AMet al. Reduced dendritic spine density on cerebral corticalpyramidal neurons in schizophrenia. J Neurol NeurosurgPsychiatry 1998; 65: 446–453.
10 Glantz LA, Gilmore JH, Lieberman JA, Jarskog LF. Apoptoticmechanisms and the synaptic pathology of schizophrenia. Schi-zophr Res 2006; 81: 47–63.
11 Glantz LA, Lewis DA. Decreased dendritic spine density onprefrontal cortical pyramidal neurons in schizophrenia. Arch GenPsychiatry 2000; 57: 65–73.
Table 4 Density (mean±s.d. cells� 103 mm�3) of neurons inthe left and right DLP/ACC of schizophrenia and controlgroups
Cortical layer Diagnosis
S C
DLP leftI 6.9±1.9 8.1±2.3II 22.6±8.0 25.3±8.9III 31.2±6.7 35.2±6.6IV 37.0±9.0 42.1±7.8V/VI 33.7±5.4 34.4±4.1
DLP rightI 7.4±1.1 8.3±1.3II 24.7±7.8 24.3±7.6III 37.6±7.1 36.3±4.9IV 43.2±8.6 40.5±6.7V/VI 29.3±5.0 31.3±5.6
ACC leftI 7.7±1.8 7.9±1.6II 69.0±9.4 66.5±9.0III 52.2±7.0 52.1± 6.3IV 54.4±4.0 51.3±4.7V/VI 48.9±8.0 47.4±7.1
ACC rightI 8.6±2.2 7.9±1.9II 57.1±11.1 56.4±9.6III 54.0±7.7 51.1±6.8IV 52.7±3.8 50.3±5.5V/VI 46.9±8.8 47.2±7.3
Abbreviations: ACC, anterior cingulate cortex; C, control;DLP, dorsolateral prefrontal; S, schizophrenic.
Prohibitin in schizophreniaK-H Smalla et al
894
Molecular Psychiatry

12 Hill SK, Ragland JD, Gur RC, Gur RE. Neuropsychologicaldifferences among empirically derived clinical subtypes ofschizophrenia. Neuropsychology 2001; 15: 492–501.
13 Kolluri N, Sun Z, Sampson AR, Lewis DA. Lamina-specificreductions in dendritic spine density in the prefrontal cortex ofsubjects with schizophrenia. Am J Psychiatry 2005; 162: 1200–1202.
14 Kolomeets NS, Orlovskaya DD, Rachmanova VI, Uranova NA.Ultrastructural alterations in hippocampal mossy fiber synapses inschizophrenia: a postmortem morphometric study. Synapse 2005;57: 47–55.
15 Stephan KE, Baldeweg T, Friston KJ. Synaptic plasticity anddysconnection in schizophrenia. Biol Psychiatry 2006; 59: 929–939.
16 Cullen TJ, Walker MA, Eastwood SL, Esiri MM, Harrison PJ, Crow TJ.Anomalies of asymmetry of pyramidal cell density and structurein dorsolateral prefrontal cortex in schizophrenia. Br J Psychiatry2006; 188: 26–31.
17 Selemon LD, Goldman-Rakic PS. The reduced neuropil hypoth-esis: a circuit based model of schizophrenia. Biol Psychiatry 1999;45: 17–25.
18 Selemon LD. Regionally diverse cortical pathology in schizophre-nia: clues to the etiology of the disease. Schizophr Bull 2001; 27:349–377.
19 Moghaddam B. Bringing order to the glutamate chaos in schizo-phrenia. Neuron 2003; 40: 881–884.
20 Tsai G, Coyle JT. Glutamatergic mechanisms in schizophrenia.Annu Rev Pharmacol Toxicol 2002; 42: 165–179.
21 Kennedy MB, Beale HC, Carlisle HJ, Washburn LR. Integration ofbiochemical signalling in spines. Nat Rev Neurosci 2005; 6: 423–434.
22 Okabe S. Molecular anatomy of the postsynaptic density. Mol CellNeurosci 2007; 34: 503–518.
23 Kristiansen LV, Huerta I, Beneyto M, Meador-Woodruff JH. NMDAreceptors and schizophrenia. Curr Opin Pharmacol 2007; 7: 48–55.
24 Bernstein HG, Becker A, Keilhoff G, Spilker C, Gorczyca WA,Braunewell KH et al. Brain region-specific changes in theexpression of calcium sensor proteins after repeated applicationsof ketamine to rats. Neurosci Lett 2003; 339: 95–98.
25 Keilhoff G, Becker A, Grecksch G, Wolf G, Bernstein HG. Repeatedapplication of ketamine to rats induces changes in the hippocam-pal expression of parvalbumin, neuronal nitric oxide synthase andcFOS similar to those found in human schizophrenia. Neu-roscience 2004; 126: 591–598.
26 Dean B, Crook JM, Pavey G, Opeskin K, Copolov DL. Muscarinic 1and 2 receptor mRNA in the human caudate-putamen: no changein m1 mRNA in schizophrenia. Mol Psychiatry 2000; 5: 203–207.
27 Yamada S, Esaki Y, Mizutani T. Intracranial cavity volume can beaccurately estimated from the weights of intracranial contents:confirmation by the dental plaster casting method. NeuropatholAppl Neurobiol 1999; 25: 341–344.
28 Bernstein HG, Sahin J, Smalla KH, Gundelfinger ED, Bogerts B,Kreutz MR. A reduced number of cortical neurons show increasedcaldendrin protein levels in chronic schizophrenia. Schizophr Res2007; 96: 246–256.
29 Danos P, Baumann B, Kramer A, Bernstein HG, Stauch R, Krell Det al. Volumes of association thalamic nuclei in schizophrenia: apostmortem study. Schizophr Res 2003; 60: 141–155.
30 Smalla KH, Matthies H, Langnase K, Shabir S, Bockers TM,Wyneken U et al. The synaptic glycoprotein neuroplastin isinvolved in long-term potentiation at hippocampal CA1 synapses.Proc Natl Acad Sci USA 2000; 97: 4327–4332.
31 Van den Oever MC, Spijker S, Li KW, Jimenez CR, Koya E,Van der Schors RC et al. A proteomics approach to identifylong-term molecular changes in rat medial prefrontal cortexresulting from sucrose self-administration. J Proteome Res 2006;5: 147–154.
32 Li KW, Hornshaw MP, Van Der Schors RC, Watson R, Tate S,Casetta B et al. Proteomics analysis of rat brain postsynapticdensity. Implications of the diverse protein functional groupsfor the integration of synaptic physiology. J Biol Chem 2004; 279:987–1002.
33 Bernstein HG, Smalla KH, Bogerts B, Gordon-Weeks PR,Beesley PW, Gundelfinger ED et al. The immunolocalization ofthe synaptic glycoprotein neuroplastin differs substantiallybetween the human and the rodent brain. Brain Res 2007; 1134:107–112.
34 Smalla KH, Seidenbecher CI, Tischmeyer W, Schicknick H,Wyneken U, Bockers TM et al. Kainate-induced epileptic seizuresinduce a recruitment of caldendrin to the postsynaptic density inrat brain. Brain Res Mol Brain Res 2003; 116: 159–162.
35 Wendholt D, Spilker C, Schmitt A, Dolnik A, Smalla KH, Proepper Cet al. ProSAP-interacting protein 1 (ProSAPiP1), a novel protein ofthe postsynaptic density that links the spine-associated Rap-Gap(SPAR) to the scaffolding protein ProSAP2/Shank3. J Biol Chem2006; 281: 13805–13816.
36 tom Dieck S, Sanmarti-Vila L, Langnaese K, Richter K, Kindler S,Soyke A et al. Bassoon, a novel zinc-finger CAG/glutamine-repeatprotein selectively localized at the active zone of presynapticnerve terminals. J Cell Biol 1998; 142: 499–509.
37 Dieterich DC, Karpova A, Mikhaylova M, Zdobnova I, Koenig I,Landwehr M et al. Caldendrin–Jacob: a protein liaison thatcouples NMDA receptor signalling to the nucleus. PLoS Biol2008; 6: 286–306.
38 Spilker C, Acuna Sanhueza GA, Bockers TM, Kreutz MR,Gundelfinger ED. SPAR2, a novel SPAR-related proteinwith GAP activity for Rap1 and Rap2. J Neurochem 2008; 104:187–201.
39 Bernstein HG, Stanarius A, Baumann B, Henning H, Krell D,Danos P et al. Nitric oxide synthase-containing neurons in thehuman hypothalamus: reduced number of immunoreactive cellsin the paraventricular nucleus of depressive patients and schizo-phrenics. Neuroscience 1998; 83: 867–875.
40 Bernstein HG, Baumann B, Danos P, Diekmann S, Bogerts B,Gundelfinger ED et al. Regional and cellular distribution of neuralvisinin-like protein immunoreactivities (VILIP-1 and VILIP-3) inhuman brain. J Neurocytol 1999; 28: 655–662.
41 Bernstein HG, Seidenbecher CI, Smalla KH, Gundelfinger ED,Bogerts B, Kreutz MR. Distribution and cellular localization ofcaldendrin immunoreactivity in adult human forebrain. J Histo-chem Cytochem 2003; 51: 1109–1112.
42 Hsu SM, Soban E. Color modification of diaminobenzidine(DAB) precipitation by metallic ions and its application fordouble immunohistochemistry. J Histochem Cytochem 1982; 30:1079–1082.
43 Cheng D, Hoogenraad CC, Rush J, Ramm E, Schlager MA, Duong DMet al. Relative and absolute quantification of postsynaptic densityproteome isolated from rat forebrain and cerebellum. Mol CellProteomics 2006; 5: 1158–1170.
44 Collins MO, Husi H, Yu L, Brandon JM, Anderson CN, BlackstockWP et al. Molecular characterization and comparison of thecomponents and multiprotein complexes in the postsynapticproteome. J Neurochem 2006; 97: 16–23.
45 Dosemeci A, Tao-Cheng JH, Vinade L, Jaffe H. Preparation ofpostsynaptic density fraction from hippocampal slices and proteo-mic analysis. Biochem Biophys Res Commun 2006; 339: 687–694.
46 Jordan BA, Fernholz BD, Boussac M, Xu C, Grigorean G, Ziff EBet al. Identification and verification of novel rodent postsynapticdensity proteins. Mol Cell Proteomics 2004; 3: 857–871.
47 Li K, Hornshaw MP, van Minnen J, Smalla KH, Gundelfinger ED,Smit AB. Organelle proteomics of rat synaptic proteins: correlation-profiling by isotope-coded affinity tagging in conjunction withliquid chromatography-tandem mass spectrometry to reveal post-synaptic density specific proteins. J Proteome Res 2005; 4: 725–733.
48 Peng J, Kim MJ, Cheng D, Duong DM, Gygi SP, Sheng M.Semiquantitative proteomic analysis of rat forebrain postsynapticdensity fractions by mass spectrometry. J Biol Chem 2004; 279:21003–21011.
49 Trinidad JC, Specht CG, Thalhammer A, Schoepfer R, Burlingame AL.Comprehensive identification of phosphorylation sites inpostsynaptic density preparations. Mol Cell Proteomics 2006; 5:914–922.
50 Yoshimura Y, Yamauchi Y, Shinkawa T, Taoka M, Donai H,Takahashi N et al. Molecular constituents of the postsynapticdensity fraction revealed by proteomic analysis using multi-dimensional liquid chromatography-tandem mass spectrometry.J Neurochem 2004; 88: 759–768.
51 Alexander-Kaufman K, James G, Sheedy D, Harper C, Matsumoto I.Differential protein expression in the prefrontal white matter ofhuman alcoholics: a proteomics study. Mol Psychiatry 2006; 11:56–65.
Prohibitin in schizophreniaK-H Smalla et al
895
Molecular Psychiatry

52 Mishra S, Murphy LC, Nyomba BL, Murphy LJ. Prohibitin: apotential target for new therapeutics. Trends Mol Med 2005; 11:192–197.
53 Prabakaran S, Swatton JE, Ryan MM, Huffaker SJ, Huang JT, Griffin JLet al. Mitochondrial dysfunction in schizophrenia: evidencefor compromised brain metabolism and oxidative stress. MolPsychiatry 2004; 9: 684–697.
54 Kalus P, Muller TJ, Zuschratter W, Senitz D. The dendriticarchitecture of prefrontal pyramidal neurons in schizophrenicpatients. Neuroreport 2000; 11: 3621–3625.
55 Manoach DS, Gollub RL, Benson ES, Searl MM, Goff DC, HalpernE et al. Schizophrenic subjects show aberrant fMRI activation ofdorsolateral prefrontal cortex and basal ganglia during workingmemory performance. Biol Psychiatry 2000; 48: 99–109.
56 Meyer-Lindenberg AS, Olsen RK, Kohn PD, Brown T, Egan MF,Weinberger DR et al. Regionally specific disturbance of dorsolat-eral prefrontal-hippocampal functional connectivity in schizo-phrenia. Arch Gen Psychiatry 2005; 62: 379–386.
57 Beasley CL, Pennington K, Behan A, Wait R, Dunn MJ, Cotter D.Proteomic analysis of the anterior cingulate cortex in the majorpsychiatric disorders: evidence for disease-associated changes.Proteomics 2006; 6: 3414–3425.
58 Clark D, Dedova I, Cordwell S, Matsumoto I. A proteome analysisof the anterior cingulate cortex gray matter in schizophrenia. MolPsychiatry 2006; 11: 459–470.
59 Johnston-Wilson NL, Sims CD, Hofmann JP, Anderson L, Shore AD,Torrey EF, et al., The Stanley Neuropathology Consortium.Disease-specific alterations in frontal cortex brain proteins inschizophrenia, bipolar disorder, and major depressive disorder.Mol Psychiatry 2000; 5: 142–149.
60 Mei J, Kolbin D, Kao HT, Porton B. Protein expression profiling ofpostmortem brain in schizophrenia. Schizophr Res 2006; 84:204–213.
61 Novikova SI, He F, Cutrufello NJ, Lidow MS. Identification ofprotein biomarkers for schizophrenia and bipolar disorder in thepostmortem prefrontal cortex using SELDI-TOF-MS ProteinChipprofiling combined with MALDI-TOF-PSD-MS analysis. Neuro-biol Dis 2006; 23: 61–76.
62 Pennington K, Beasley CL, Dicker P, Fagan A, English J, Pariante CMet al. Prominent synaptic and metabolic abnormalities revealed byproteomic analysis of the dorsolateral prefrontal cortex inschizophrenia and bipolar disorder. Mol Psychiatry, publishedonline 16 October 2007; e-pub ahead of print.
63 Paulson L, Martin P, Persson A, Nilsson CL, Ljung E, Westman-Brinkmalm A et al. Comparative genome- and proteome analysisof cerebral cortex from MK-801-treated rats. J Neurosci Res 2003;71: 526–533.
64 Arion D, Unger T, Lewis DA, Levitt P, Mirnics K. Molecularevidence for increased expression of genes related to immune andchaperone function in the prefrontal cortex in schizophrenia. BiolPsychiatry 2007; 63: 711–721.
65 Dean B, Keriakous D, Scarr E, Thomas EA. Gene expressionprofiling in Brodmann’s area 46 from subjects with schizophrenia.Aust N Z J Psychiatry 2007; 41: 308–320.
66 DeLisi LE, Fleischhaker W. Schizophrenia research in the era ofthe genome. Curr Opin Psychiatry 2007; 20: 109–110.
67 Saito A, Fujikura-Ouchi Y, Kuramasu A, Shimoda K, Akiyama K,Matsuoka H et al. Association study of putative promoterpolymorphisms in the neuroplastin gene and schizophrenia.Neurosci Lett 2007; 411: 168–173.
68 Sivagnanasundaram S, Fletcher D, Hubank M, Illingworth E,Skuse D, Scambler P. Differential gene expression in thehippocampus of the Df1/þ mice: a model for 22q11.2 deletionsyndrome and schizophrenia. Brain Res 2007; 1139: 48–59.
69 Eastwood SL, Harrison PJ. Hippocampal synaptic pathology inschizophrenia, bipolar disorder and major depression: a study ofcomplexin mRNAs. Mol Psychiatry 2000; 5: 425–432.
70 Harrison PJ, Eastwood SL. Preferential involvement of excitatoryneurons in medial temporal lobe in schizophrenia. Lancet 1998;352: 1669–1673.
71 Di Maria E, Gulli R, Begni S, De Luca A, Bignotti S, Pasini A et al.Variations in the NMDA receptor subunit 2B gene (GRIN2B) andschizophrenia: a case-control study. Am J Med Genet B Neurop-sychiatr Genet 2004; 128: 27–29.
72 Iasevoli F, Polese D, Ambesi-Impiombato A, Muscettola G,de Bartolomeis A. Ketamine-related expression of glutamatergicpostsynaptic density genes: possible implications in psychosis.Neurosci Lett 2007; 416: 1–5.
73 Malhotra AK, Breier A, Goldman D, Picken L, Pickar D. Theapolipoprotein E epsilon 4 allele is associated with blunting ofketamine-induced psychosis in schizophrenia. A preliminaryreport. Neuropsychopharmacology 1998; 19: 445–448.
74 Li KW, Miller S, Klychnikov O, Loos M, Stahl-Zeng J, Spijker Set al. Quantitative proteomics and protein network analysis ofhippocampal synapses of CaMKIIalpha mutant mice. J ProteomeRes 2007; 6: 3127–3133.
75 Toyooka K, Iritani S, Makifuchi T, Shirakawa O, Kitamura N,Maeda K et al. Selective reduction of a PDZ protein, SAP-97, in theprefrontal cortex of patients with chronic schizophrenia.J Neurochem 2002; 83: 797–806.
76 Sutton MA, Schuman EM. Dendritic protein synthesis, synapticplasticity, and memory. Cell 2006; 127: 49–58.
77 Carboni L, Vighini M, Piubelli C, Castelletti L, Milli A, DomeniciE. Proteomic analysis of rat hippocampus and frontal cortex afterchronic treatment with fluoxetine or putative novel antidepres-sants: CRF1 and NK1 receptor antagonists. Eur Neuropsychophar-macol 2006; 16: 521–537.
78 Vercauteren FG, Bergeron JJ, Vandesande F, Arckens L, Quirion R.Proteomic approaches in brain research and neuropharmacology.Eur J Pharmacol 2004; 500: 385–398.
79 Yang MS, Yu L, Guo TW, Zhu SM, Liu HJ, Shi YY et al. Evidencefor association between single nucleotide polymorphisms in Tcomplex protein 1 gene and schizophrenia in the Chinese Hanpopulation. J Med Genet 2004; 41: e63.
80 La Y, Wan C, Zhu H, Yang Y, Chen Y, Pan Y et al. Hippocampusprotein profiling reveals aberration of malate dehydrogenase inchlorpromazine/clozapine treated rats. Neurosci Lett 2006; 408:29–34.
81 Stone WS, Faraone SV, Su J, Tarbox SI, Van Eerdewegh P, Tsuang MT.Evidence for linkage between regulatory enzymes in glycolysisand schizophrenia in a multiplex sample. Am J Med Genet BNeuropsychiatr Genet 2004; 127: 5–10.
82 Burbaeva G, Savushkina OK, Boksha IS. Creatine kinase BBin brain in schizophrenia. World J Biol Psychiatry 2003; 4: 177–183.
83 Kunugi H, Kato T, Fukuda R, Tatsumi M, Sakai T, Nanko S.Association study of C825T polymorphism of the G-protein b3subunit gene with schizophrenia and mood disorders. J NeuralTransm 2002; 109: 213–218.
84 Minoretti P, Politi P, Coen E, Di Vito C, Bertona M, Bianchi M et al.The T393C polymorphism of the GNAS1 gene is associated withdeficit schizophrenia in an Italian population sample. NeurosciLett 2006; 397: 159–163.
85 Tani M, Mui K, Minami Y, Kiriike N. Association of a GTP-bindingprotein Go alpha subunit mutation with schizophrenia. MolPsychiatry 2001; 6: 359.
86 Ventriglia M, Bocchio Chiavetto L, Bonvicini C, Tura GB, Bignotti S,Racagni G et al. Allelic variation in the human prodynorphin genepromoter and schizophrenia. Neuropsychobiology 2002; 46: 17–21.
87 Zhang CS, Tan Z, Lu L, Wu SN, He Y, Gu NF et al. Polymorphismof Prodynorphin promoter is associated with schizophrenia inChinese population. Acta Pharmacol Sin 2004; 25: 1022–1026.
88 Li Z, Okamoto K, Hayashi Y, Sheng M. The importance ofdendritic mitochondria in the morphogenesis and plasticity ofspines and synapses. Cell 2004; 119: 873–887.
89 Nijtmans LG, de Jong L, Artal Sanz M, Coates PJ, Berden JA, Back JWet al. Prohibitins act as a membrane-bound chaperone for thestabilization of mitochondrial proteins. EMBO J 2000; 19: 2444–2451.
90 Rajalingam K, Wunder C, Brinkmann V, Churin Y, Hekman M, SieversC et al. Prohibitin is required for Ras-induced Raf-MEK-ERK activationand epithelial cell migration. Nat Cell Biol 2005; 7: 837–843.
Supplementary Information accompanies the paper on the Molecular Psychiatry website (http://www.nature.com/mp)
Prohibitin in schizophreniaK-H Smalla et al
896
Molecular Psychiatry



















