
A Comparison of Abundance and Diversity of Epiphytic Microalgal Assemblages on the Leaves of the...
11
Research Article A Comparison of Abundance and Diversity of Epiphytic Microalgal Assemblages on the Leaves of the Seagrasses Posidonia oceanica (L.) and Cymodocea nodosa (Ucria) Asch in Eastern Tunisia Lotfi Mabrouk, 1,2 Mounir Ben Brahim, 1,2 Asma Hamza, 2 Mabrouka Mahfoudhi, 2 and Med Najmeddine Bradai 2 1 Faculty of Science of Sfax, Route de la Soukra km 4, BP 802, 3038 Sfax, Tunisia 2 National Institute of Sciences and Technology of the Sea, BP 1035, 3018 Sfax, Tunisia Correspondence should be addressed to Lotfi Mabrouk; lotfi[email protected] Received 23 February 2014; Revised 20 April 2014; Accepted 6 May 2014; Published 3 June 2014 Academic Editor: Andrew McMinn Copyright © 2014 Lotfi Mabrouk et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. We studied spatial patterns in assemblages of epiphytic microalgae on the leaves of two seagrass species with different morphologies and longevity, Cymodocea nodosa and Posidonia oceanica, which cooccur in Chebba in Eastern Tunisia. Epiphyte assemblages were described for each species in summer. Epiphyte microalgal assemblages were more abundant on the leaves of C. nodosa but more diversified on the leaves of P. oceanica. We suggest that the differences in species composition and abundance between those seagrass species may reflect an interaction of timescales of seagrass longevity with timescales of algal reproductive biology. Short-lived C. nodosa was dominated by fast growing species such as the cyanobacteria species Oscillatoria sp., while P. oceanica leaves were colonized by more mature and diversified species such as Prorocentrales. Local environmental conditions (hydrodynamics, light penetration), host characteristics (meadow type, shapes forms of leaves, life span, and growth rate), and grazing effect seem also to be responsible for these dissimilarities in epiphytic microalgae communities. 1. Introduction e habitats of marine Magnoliophyta Posidonia and Cymod- ocea are, with the coral, the most important marine ecosys- tems of the Mediterranean. eir leaves provide substrates suitable for the establishment and growth of a number of micro- and macrocolonizers which form laminates assem- blages characterized by high species diversity [1]. Previous studies showed that epiphytic community struc- ture is influenced by biotic factors such as leaf age, seasonal cycle of the host, and grazing pressure by herbivores [2–5], as well as by abiotic factors such as light, temperature, nutrients, and water motion [6–8]. e density of the seagrass canopy and shoot size can significantly modify epiphytic biomass, presumably due to effects of light penetration [9, 10], whereas shoot morphology, leaf, and stem ages can influence epiphyte distribution and abundance due to differences in the surface area that is available for epiphyte settlement [11, 12]. Epiphytic microalgae include many toxic species [13–16]. Knowledge of the epiphytes and potentially toxic species of microalgae is necessary to assess the influence of those species on fisheries exploitation and human health. Moreover, various studies have documented a good correlation between the abundance of those toxic species in the water column and their abundance on macrophytes [15, 17]. Although a good number of surveys have focused on the epiphytic community in the north basin of the Mediterranean Sea, the south part has received little attention. e purpose of this study is to investigate the diversity and species abundance of leaf epiphytic microalgae on Posidonia oceanica (L.) and Cymodocea nodosa (Ucria) Asch in eastern Tunisia where several fish farms have recently been installed. Hindawi Publishing Corporation Journal of Marine Biology Volume 2014, Article ID 275305, 10 pages http://dx.doi.org/10.1155/2014/275305
Transcript of A Comparison of Abundance and Diversity of Epiphytic Microalgal Assemblages on the Leaves of the...

Research ArticleA Comparison of Abundance and Diversity of EpiphyticMicroalgal Assemblages on the Leaves of the SeagrassesPosidonia oceanica (L) and Cymodocea nodosa (Ucria)Asch in Eastern Tunisia
Lotfi Mabrouk12 Mounir Ben Brahim12 Asma Hamza2
Mabrouka Mahfoudhi2 and Med Najmeddine Bradai2
1 Faculty of Science of Sfax Route de la Soukra km 4 BP 802 3038 Sfax Tunisia2 National Institute of Sciences and Technology of the Sea BP 1035 3018 Sfax Tunisia
Correspondence should be addressed to Lotfi Mabrouk lotfi2328yahoofr
Received 23 February 2014 Revised 20 April 2014 Accepted 6 May 2014 Published 3 June 2014
Academic Editor Andrew McMinn
Copyright copy 2014 Lotfi Mabrouk et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
We studied spatial patterns in assemblages of epiphyticmicroalgae on the leaves of two seagrass species with differentmorphologiesand longevity Cymodocea nodosa and Posidonia oceanica which cooccur in Chebba in Eastern Tunisia Epiphyte assemblages weredescribed for each species in summer Epiphyte microalgal assemblages were more abundant on the leaves of C nodosa but morediversified on the leaves of P oceanicaWe suggest that the differences in species composition and abundance between those seagrassspecies may reflect an interaction of timescales of seagrass longevity with timescales of algal reproductive biology Short-lived Cnodosa was dominated by fast growing species such as the cyanobacteria species Oscillatoria sp while P oceanica leaves werecolonized by more mature and diversified species such as Prorocentrales Local environmental conditions (hydrodynamics lightpenetration) host characteristics (meadow type shapes forms of leaves life span and growth rate) and grazing effect seem also tobe responsible for these dissimilarities in epiphytic microalgae communities
1 Introduction
Thehabitats of marineMagnoliophyta Posidonia andCymod-ocea are with the coral the most important marine ecosys-tems of the Mediterranean Their leaves provide substratessuitable for the establishment and growth of a number ofmicro- and macrocolonizers which form laminates assem-blages characterized by high species diversity [1]
Previous studies showed that epiphytic community struc-ture is influenced by biotic factors such as leaf age seasonalcycle of the host and grazing pressure by herbivores [2ndash5] aswell as by abiotic factors such as light temperature nutrientsand water motion [6ndash8] The density of the seagrass canopyand shoot size can significantly modify epiphytic biomasspresumably due to effects of light penetration [9 10] whereasshoot morphology leaf and stem ages can influence epiphyte
distribution and abundance due to differences in the surfacearea that is available for epiphyte settlement [11 12]
Epiphytic microalgae include many toxic species [13ndash16]Knowledge of the epiphytes and potentially toxic speciesof microalgae is necessary to assess the influence of thosespecies onfisheries exploitation andhumanhealthMoreovervarious studies have documented a good correlation betweenthe abundance of those toxic species in the water column andtheir abundance on macrophytes [15 17] Although a goodnumber of surveys have focused on the epiphytic communityin the north basin of the Mediterranean Sea the south parthas received little attention
Thepurpose of this study is to investigate the diversity andspecies abundance of leaf epiphytic microalgae on Posidoniaoceanica (L) and Cymodocea nodosa (Ucria) Asch in easternTunisia where several fish farms have recently been installed
Hindawi Publishing CorporationJournal of Marine BiologyVolume 2014 Article ID 275305 10 pageshttpdxdoiorg1011552014275305
2 Journal of Marine Biology
Chebba
Tunisia
8∘20
9984009∘10
99840010
∘00
99840010
∘50
99840011
∘40
99840012
∘30
998400
36∘40
998400
35∘50
998400
35∘00
998400
34∘10
998400
33∘20
998400
32∘30
998400
Figure 1 Map of the study area showing the sampling stations
Our aim was to examine the following hypothesis (1) dueto their large leaves epiphytic microalgae on the leaves ofPosidonia oceanicamay be more diverse and more abundantthan those on Cymodocea nodosa leaves (2) variation of thephenological parameters of the host plant may explain thepattern of microepiphyte abundance (3) some potentiallytoxic species are likely to be found among the epiphyticcommunity on thePosidonia oceanica andCymodocea nodosaleaves
2 Materiel and Methods
21 Study Area This study was conducted in the locality ofOued Lafrann (35∘1510158401810158401015840 N 110710158402810158401015840 E) in the region ofChebba (east of Tunisia) where 32 aquaculture cages of seabeam have been recently installed The climate is semiarid(average precipitation 350mm per year) and sunny withstrong northerly winds This area is under consideration asa marine protected area (submitted in Italo-Tunisian projectiTunES) because of its biocenotic richness and its distinc-tive ecosystems especially macrophytes mainly Cymodoceanodosa that occur near Posidonia oceanica bed Halophilastipulacea and many macroalgae
Surveying and sampling were conducted during Septem-ber 2013 by SCUBA diving in two stations about 1 km apart(Figure 1) Posidonia oceanica meadow (P) and Cymodoceanodosa meadow (C) To reduce habitat variability in thepresent study all samples were collected at the same depthof 5m Sampling was conducted in the same day watertemperature was 22∘C water salinity was 382 plusmn 02 gsdotLminus1(mean plusmn SD) and wind speed was about 20 kmh Becauseof the proximity of sampling stations measurements ofphysicochemical parameters are not carried out
3 Sampling Treatment
To study the epiphytic microalgae on Posidonia oceanicaand Cymodocea nodosa leaves shoots were collected using a
metal square quadrat with a 40 cm inner edge The quadratwas randomly placed over the shoots which were all care-fully collected At each station six replicate quadrats weresampled The shoot density of P oceanica and C nodosawas measured in six replicates of 025m2 and expressed asnumber of shootssdotmminus2
In the laboratory from each quadrate leaves wereremoved from 15 shoots in distichous order of insertion andseparated into the various categories defined by Giraud [18]For each shoot the following leaf traits were scored (1) totalnumber of standing leaves (2) total number and (3) lengthof adult and intermediate leaves and (4) leaf width Leafarea index (LAI m2sdotmminus2) was determined as product of leafsurface area (total leaf lengthtimesmean leaf width cm2sdotshootminus2)and shoot density
To detach the leaf microepiphytic communities fromeach quadrat leaves were detached from their sheet andweighed to 100 g with an electronic precision balance [19]The epiphytes were then separated from the weighted leavesby vigorous shaking and washing with filtered seawater Thisprocedure was repeated several times to ensure that most ofthe attached organismswere separatedThismethodwas usedby previous researchers to removemicroepiphytic species [1520ndash23]Thefilteredmaterial was thenpassed through 250 and100 120583m mesh sieves to remove large particles and was fixedwith Lugolrsquos solution and finally preserved in 5 formalinand its volume (119881) was noted All filtered materials were keptin the dark at ambient temperature until their microscopicobservation Settling long glass tubes used for sedimentationprocedure were 2 cm wide by 21 cm long and have a baseplate that contains a coverslip into which the algae settle Tomix the sample the bottle was gently tilted back and forth 10times before pouring A 50mL subsample was poured intothe settling chamber and left to settle for 24 h Subsampleswere examined in an inverted microscope at medium (times200)magnification by scanning the entire surface of the settlingchamber to enumerate epiphytic microalgae [24] The totalnumber of microalgae individuals (119873) contained in 100 g offresh weight Posidonia (expressed as number of individualper 100 g of fresh weight of Posidonia (fwm)) is obtained bythe following conversion119873 = (119899 times119881)V with 119899 = number ofindividuals counted 119881 = volume of the filtered material andV = volume of the sedimentation chamber (50mL)The iden-tified taxa were divided into groups (diatoms dinoflagellatesand cyanobacteria)
4 Data Analysis
Data were tested for normality using the Kolmogorov-Smirnov goodness of fit test and for heterogeneity usingCochranrsquos C Test and then transformed if necessary [25]
Analyses of similarity (ANOSIM) randomization tests(with untransformed data) were used to test for differencesin species abundance among sampling stations [26] If dif-ferences were found using ANOSIM then SIMPER analysiswas used for identifying which species primarily accountedfor observed differences in epiphytic assemblages betweensampling stations SIMPER generates a ranking of the percent
Journal of Marine Biology 3
contribution of the species that are most important to thesignificant differences These analyses used a matrix com-posed of Bray-Curtis similarity coefficients generated withlog(119909+1)-transformed dataThe calculationswere performedusing the statistical software PRIMER 50 [27]
Univariate indices species number (119878) and diversity(Shannon-Weaver index 1198671015840) were calculated for each sam-pling station The one-way ANOVA was used to test foroverall differences between these indices and the Tukey HSDmultiple comparison tests were used in pairwise comparisonsbetween sampling stations One-way analysis of variance(ANOVA) was used to test the hypothesis that the abundanceof each of the most abundant groups varies among samplingstations Tukey test HSDwas employed for a posteriori multi-ple comparisons of means The calculations were performedusing the statistical software STATISTICA v10 (StatSoft Inc)
Relationships between epiphytic species abundance andbiometric parameters (shoot density leaf area index andaverage length of adult and intermediate leaves) were exam-ined using the RELATE procedure in PRIMER RELATEis the equivalent of a nonparametric Mantel test [28] itassesses the degree of correspondence between matrices andvia a randomization test it provides a measure of statisticalsignificance of the relationship [28]Thematrix of similaritiesbetween epiphytic species abundance (based on the Bray-Curtis coefficient from untransformed data) was comparedwith a matrix of the similarities between biometric parame-ters (based on Euclidean distance from untransformed data)The significance of any correlation between matrices wasassessed with a randomization test
Canonical correspondence analysis (CCA) a direct gra-dient analysis technique [29] was used to investigate the rela-tionship between epiphytic species and biometric parametersof the two sampling macrophytes Epiphyte abundances datawere log(119909+1)-transformed Downweighting for rare specieswas performed We used CANOCO 45 (Scientia Software)
5 Results
Cymodocea nodosa exhibits the highest shoot density whilePosidonia oceanica has the highest leaf area index (LAI)(Figure 2) The average leaf number was 721 plusmn 213 (meanplusmn standard deviation) and 366 plusmn 112 for P oceanica and Cnodosa respectively The highest mean value of leaf length(4857 plusmn 787 cm) was recorded for P oceanica whereas thelowest mean value (163 plusmn 487 cm) was observed for Cnodosa
A total of 52 taxa of epiphytic microalgae were identified(Table 1) The highest number of collected species wasdiatoms (24) followed by dinoflagellates (22) and cyanobac-teria (6) Species number and 1198671015840 index of microepiphyticassemblage were high on Posidonia oceanica leaves (Figure 3)
Analysis of similarity (ANOSIM) of epiphytic microalgaelog(119883+1)-transformed specie abundances showed significantdifference (119877 = 0987 119875 = 002) between the P oceanicastation and C nodosa station
Analyses of similarity percentage (SIMPER) showed thatthe average dissimilarity between P oceanica and C nodosa
0
200
400
600
800
1000
1200
1400
1600
Posidonia Cymodocea0
5
10
15
20
25
30
35
40
DensityLAI
LAI(
m2m
2)
Den
sity
(sho
ots
m2)
Figure 2 Average leaf area index (LAI) and shoot density of Poceanica and Cymodocea nodosa
epiphytes groups is high (6444) This procedure alsoallowed us to determine the species that contribute to thisdissimilarity they are Oscillatoria sp Fragilaria Pseudan-abaena sp andAmphiprora constricta (Table 2) that are moreabundant on C nodosa leaves
Dinoflagellates diatoms and cyanobacteria were com-mon epiphytes on leaves (Figure 4) Abundances of thosegroups were included in the univariate analyses of variancesDinoflagellates did not differ significantly between stations(119865(110)
= 131 119875 = 0279) Significant differences weredetected for Gymnodiniales and Peridiniales among stations(119865(110)
= 286842 119875 lt 0001 119865(110)
= 166678 119875 lt 0001resp) with high abundance on leaves of Cymodocea Incontrast Prorocentrales were more abundant on the leavesof Posidonia (119865
(110)= 1579 119875 = 0003 Tukey test) Epiphytic
diatoms were more abundant on Posidonia leaves (119865(110)
=1364 119875 = 0004 Tukey test) In contrast cyanobacteriawere higher on Cymodocea leaves (119865
(110)= 174926 119875 lt
0001 Tukey test) When the abundances of toxic speciesare grouped significant differences were detected amongstations (119865
(110)= 5763 119875 lt 0001) with high abundance
on Posidonia leaves Significant differences were detected forspecies number and for1198671015840 index (119865
(110)= 34005 119875 lt 0001
119865(110)
= 1414 119875 = 0004) 119878 and1198671015840 of P oceanica were higherthan those of C nodosa (Table 3)
The results of RELATE tests indicated that there is acorrelation between the biometric parameters (shoot densityLAI and average adult and intermediate leaves) and speciesabundance (Spearman rank correlation statistic Rho = 0684119875 = 004)
Canonical correspondence analysis (CCA) indicates thatthe axis I and axis II expressed high cumulative variancespecies-biometric parameters (981 Table 4) Stations Pand C are placed separately on the left and the right of thediagrams respectively Species placed closely with samples
4 Journal of Marine Biology
Table 1 ldquoPresence-absencerdquo species list of epiphyticmicroalgae identified on the leaves of P oceanica and ofC nodosa in prospected stations
Species Posidonia CymodoceaDinoflagellates
Amphidinium carteraeHulburt 0 1Ceratium furca (Ehrenberg) Claparede and Lachmann 1 0Ceratium tripos (Muller) 1 0Coolia monotisMeunier 0 1Cyst of dinoflagellates 0 1Gonyaulax spinifera 0 1Gymnodinium sp 0 1Ostreopsis ovata (Fukuyo) 0 1Polykrikos sp 0 1Prorocentrum concavum (Fukuyo) 1 0Prorocentrum gracile (Schutt) 1 0Prorocentrum lima (Ehrenberg) Dodge 1 0Prorocentrum micans Ehrenberg 1 0Prorocentrum minimum (Pavillard) J Schiller 1 0Prorocentrum rathymum (Loeblich) Shirley and Schmidt 1 0Prorocentrum triestinum (Schiller) 1 0Protoperidinium curtipes (Jorgensen) Balech 1 0Protoperidinium depressum (Bailey) Balech 1 0Protoperidinium divergens (Ehrenberg) Balech 1 0Protoperidinium mite (Pavillard) Balech 1 0Protoperidinium ovum (Schiller) Balech 1 0Protoperidinium steinii (Joslashrgensen) Balech 1 0Protoperidiniumsp 0 1Scrippsiella sp 1 1
DiatomsAmphiprora constricta Ehrenberg 1 0Amphora sp 1 1Asterionellopsis glacialis (Castracane) Round 1 0Asteromphalus flabellatus (Brebisson) Greville 1 0Bacillariasp 1 0Budelphia sp 1 0Chaetoceros sp 1 0Climacosphenia moniligera (Ehrenberg) 1 0Coscinodiscus sp 0 1Dactyliosolen sp 1 0Diploneis sp 1 0Fragilaria sp 1 0Grammatophora sp 1 0Guinardia sp 1 1Licmophora sp 1 1Navicula sp 1 1Nitzschia fonticola Grunow 1 0Pinnularia sp 1 0Pleurosigma sp 1 0Rhizosolenia styliformis Brightwell 0 1Striatella unipunctata (Lyngbye) C Agardh 1 0Surirella sp 1 0
Journal of Marine Biology 5
Table 1 Continued
Species Posidonia CymodoceaCyanobacteria
Anabaena sp 1 1Lyngbya sp 0 1Merismopedia sp 0 1Micrococcus sp 0 1Oscillatoria sp 1 1Pseudanabaena sp 1 0
05
1015202530354045
Posidonia Cymodocea0020406081121416182
SH998400
Figure 3 Mean diversity index (1198671015840) and species number (119878) onleaves in prospected stations
010002000300040005000600070008000
Dinoflagellates Diatoms CyanobacteriaGroup
PosidoniaCymodocea
(ind100
g FW
M)
Figure 4 Average abundance of microepiphytes on leaves inprospected stations
of P oceanica (P) station belong mainly to ProrocentralesSpecies closely placed near samples of C nodosa stationbelong mainly to cyanobacteria (Figure 5)
6 Discussion
Differences between leaf epiphytic microalgae communitieswere found between C nodosa and P oceanica in our studyarea (East of Tunisia) leaf microepiphytic communities ofPosidonia were more diverse than those of C nodosa (119878 and
1198671015840 Table 3) The same results were found by Mazzella et al
[30] in Adriatic Sea and by Turki [19] in Northern TunisiaContrary to what was expected total abundance of epiphyticmicroalgae was higher on C nodosa A possible hypothesisto explain this result is that P oceanica develops much largerleaf area index values thanC nodosawhich contribute to lightattenuation within the seagrass canopy by leaf self-shading[31] Unfortunately lightmeasurementswere not taken in thisstudy but previous studies have shown that light attenuationby the canopy is reduced with decreased leaf area index[32] Therefore the higher light availability that may occurin C nodosa meadows may explain the higher abundance ofepiphytic microalgae found in these meadows
Ours results showed that microepiphytic abundances arecorrelated with shoot density and phenological parametersof the host plant (RELATE procedure) prorocentrales abun-dance was related to leaf length and leaf area index whilecyanobacteria was related to Cymodocea shootdensity (CCAprocedure Figure 5) Correlation between microepiphyticspecies and leaf phenological parameters of the host plant hasbeen also found by previous studies for P oceanica [5] andfor Zostera marina [33] Indeed differences between leavesshapes could explain epiphytic species variability betweenP oceanica and C nodosa The leaves of P oceanica areribbon-shaped (40 to 140 cm long 7 to 11mmwide) gatheredtogether in fascicles (from 5 to 8 leaves) at the tips of thestems and have a life of between 5 and 13 months [34] Theleaves of C nodosa are also ribbon-shaped but shorter thanthose of Posidonia oceanica (10 to 30 cm long 2 to 4mmwide) gathered together in smaller fascicles (from 2 to 5leaves) and have a life of between 2 and 8 months [35 36]Cymodocea nodosa has the higher value of specific growthrate (up to 47) than of P oceanica (15) [31] The shortlife span C nodosa could explain domination of fast growingspecies such as Oscillatoria sp [37] and other cyanobacteriaspecies Posidonia oceanica leaves are older they offer moretime for the epiphytic species to settle and the epiphyticcommunity will be composed of a more mature and morediverse community
The epiphytic composition is influenced by the interac-tion between the lifespan of the host and the reproductivelifespan of the epiphytes [38] Where the host is long-livedas for Posidonia local recruitment from existing epiphyteswith fast reproductive strategies can continually reinforcethe local composition For instance the density of diatomsincreased relatively quickly and became dominant (after 7 to10 weeks) in the older parts of the sheet [1 22] Cyanobacteria
6 Journal of Marine Biology
Table 2 SIMPER results
Species Group Posidonia Group Cymodocea Av Diss DissSD Contrib CumAv Abund Av Abund
Oscillatoria sp 500 6000 3019 62 4685 4685Navicula sp 393333 3850 1336 207 2074 6759Fragilaria sp 625 0 334 632 519 7278Pseudanabaena sp 600 0 331 649 513 7791Amphiprora constricta 450 0 248 574 385 8176Guinardia sp 3333 300 147 51 229 8405Anabaenasp 300 4167 142 579 221 8626Pinnularia sp 250 0 141 281 218 8844Amphora sp 100 300 109 425 169 9013
Table 3 Analysis of variance ANOVA of the abundances of major groups of microalgae and cyanobacteria in prospected stations
Source Df MS F P Tukey testTotal abundance
Habitat 1 0050 15555 0003 C gt PResiduals 10 0003
DinoflagellatesHabitat 1 0031 1312 0279 n sResiduals 10 0023
DiatomsHabitat 1 0067 13643 0004 C lt PResiduals 10 0005
CyanobacteriaHabitat 1 1310 1749266 lt0001 C gt PResiduals 10 0001
Toxic speciesHabitat 1 1255 57635 lt0001 C lt PResiduals 10 0022
Species number (S)Habitat 1 1452000 34005 lt0001 C lt PResiduals 10 42700
H1015840
Habitat 1 0032 14143 0004 C lt PResiduals 10 0002
C Cymodocea nodosameadow P Posidonia oceanicameadow n s not significant Data were log(119909 + 1)-transformed after significant Cochran C test
include some fast growing species such asOscillatoria sp [37]Borowitzka et al [38] suggest that seagrass epiphytes can beclassified into groups based on their seasonal distribution (a)epiphytes occurring throughout the year (b) epiphytes with adistinct seasonal pattern in their occurrence and (c) transientcolonizers
Difference between epiphytic compositions on the leavesof seagrass species has been studied over the world andshowed that more persistent and structurally complex sea-grass species tend to havemore diverse epiphyte assemblagesShort-lived seagrass species such as Heterozostera sp andHalodule sp are likely to be relatively depauperate in epiphytespecies richness compared to persistent seagrass species such
as Amphibolis sp [38] The findings of Aligizaki and Niko-laidis [15] showed that themaximumabundances of epiphyticdinoflagellates were recorded in branched macroalgae whilethose abundances were remarkably lower in the less leafymarine phanerogame such as Cymodocea
The two species of macrophytes made different habitatsdepending on local environmental conditions (nutrient sub-stratum current ect) our prospected Posidonia meadow isa monospecific fringing reef whereas sampling C nodosais a continuous extensive bed Differences of hydrodynamicforces between those two habitats and wave exposure canalso be responsible for species variability of the epiphyticcommunities Indeed seagrasses are able to modify current
Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 Journal of Marine Biology
Chebba
Tunisia
8∘20
9984009∘10
99840010
∘00
99840010
∘50
99840011
∘40
99840012
∘30
998400
36∘40
998400
35∘50
998400
35∘00
998400
34∘10
998400
33∘20
998400
32∘30
998400
Figure 1 Map of the study area showing the sampling stations
Our aim was to examine the following hypothesis (1) dueto their large leaves epiphytic microalgae on the leaves ofPosidonia oceanicamay be more diverse and more abundantthan those on Cymodocea nodosa leaves (2) variation of thephenological parameters of the host plant may explain thepattern of microepiphyte abundance (3) some potentiallytoxic species are likely to be found among the epiphyticcommunity on thePosidonia oceanica andCymodocea nodosaleaves
2 Materiel and Methods
21 Study Area This study was conducted in the locality ofOued Lafrann (35∘1510158401810158401015840 N 110710158402810158401015840 E) in the region ofChebba (east of Tunisia) where 32 aquaculture cages of seabeam have been recently installed The climate is semiarid(average precipitation 350mm per year) and sunny withstrong northerly winds This area is under consideration asa marine protected area (submitted in Italo-Tunisian projectiTunES) because of its biocenotic richness and its distinc-tive ecosystems especially macrophytes mainly Cymodoceanodosa that occur near Posidonia oceanica bed Halophilastipulacea and many macroalgae
Surveying and sampling were conducted during Septem-ber 2013 by SCUBA diving in two stations about 1 km apart(Figure 1) Posidonia oceanica meadow (P) and Cymodoceanodosa meadow (C) To reduce habitat variability in thepresent study all samples were collected at the same depthof 5m Sampling was conducted in the same day watertemperature was 22∘C water salinity was 382 plusmn 02 gsdotLminus1(mean plusmn SD) and wind speed was about 20 kmh Becauseof the proximity of sampling stations measurements ofphysicochemical parameters are not carried out
3 Sampling Treatment
To study the epiphytic microalgae on Posidonia oceanicaand Cymodocea nodosa leaves shoots were collected using a
metal square quadrat with a 40 cm inner edge The quadratwas randomly placed over the shoots which were all care-fully collected At each station six replicate quadrats weresampled The shoot density of P oceanica and C nodosawas measured in six replicates of 025m2 and expressed asnumber of shootssdotmminus2
In the laboratory from each quadrate leaves wereremoved from 15 shoots in distichous order of insertion andseparated into the various categories defined by Giraud [18]For each shoot the following leaf traits were scored (1) totalnumber of standing leaves (2) total number and (3) lengthof adult and intermediate leaves and (4) leaf width Leafarea index (LAI m2sdotmminus2) was determined as product of leafsurface area (total leaf lengthtimesmean leaf width cm2sdotshootminus2)and shoot density
To detach the leaf microepiphytic communities fromeach quadrat leaves were detached from their sheet andweighed to 100 g with an electronic precision balance [19]The epiphytes were then separated from the weighted leavesby vigorous shaking and washing with filtered seawater Thisprocedure was repeated several times to ensure that most ofthe attached organismswere separatedThismethodwas usedby previous researchers to removemicroepiphytic species [1520ndash23]Thefilteredmaterial was thenpassed through 250 and100 120583m mesh sieves to remove large particles and was fixedwith Lugolrsquos solution and finally preserved in 5 formalinand its volume (119881) was noted All filtered materials were keptin the dark at ambient temperature until their microscopicobservation Settling long glass tubes used for sedimentationprocedure were 2 cm wide by 21 cm long and have a baseplate that contains a coverslip into which the algae settle Tomix the sample the bottle was gently tilted back and forth 10times before pouring A 50mL subsample was poured intothe settling chamber and left to settle for 24 h Subsampleswere examined in an inverted microscope at medium (times200)magnification by scanning the entire surface of the settlingchamber to enumerate epiphytic microalgae [24] The totalnumber of microalgae individuals (119873) contained in 100 g offresh weight Posidonia (expressed as number of individualper 100 g of fresh weight of Posidonia (fwm)) is obtained bythe following conversion119873 = (119899 times119881)V with 119899 = number ofindividuals counted 119881 = volume of the filtered material andV = volume of the sedimentation chamber (50mL)The iden-tified taxa were divided into groups (diatoms dinoflagellatesand cyanobacteria)
4 Data Analysis
Data were tested for normality using the Kolmogorov-Smirnov goodness of fit test and for heterogeneity usingCochranrsquos C Test and then transformed if necessary [25]
Analyses of similarity (ANOSIM) randomization tests(with untransformed data) were used to test for differencesin species abundance among sampling stations [26] If dif-ferences were found using ANOSIM then SIMPER analysiswas used for identifying which species primarily accountedfor observed differences in epiphytic assemblages betweensampling stations SIMPER generates a ranking of the percent
Journal of Marine Biology 3
contribution of the species that are most important to thesignificant differences These analyses used a matrix com-posed of Bray-Curtis similarity coefficients generated withlog(119909+1)-transformed dataThe calculationswere performedusing the statistical software PRIMER 50 [27]
Univariate indices species number (119878) and diversity(Shannon-Weaver index 1198671015840) were calculated for each sam-pling station The one-way ANOVA was used to test foroverall differences between these indices and the Tukey HSDmultiple comparison tests were used in pairwise comparisonsbetween sampling stations One-way analysis of variance(ANOVA) was used to test the hypothesis that the abundanceof each of the most abundant groups varies among samplingstations Tukey test HSDwas employed for a posteriori multi-ple comparisons of means The calculations were performedusing the statistical software STATISTICA v10 (StatSoft Inc)
Relationships between epiphytic species abundance andbiometric parameters (shoot density leaf area index andaverage length of adult and intermediate leaves) were exam-ined using the RELATE procedure in PRIMER RELATEis the equivalent of a nonparametric Mantel test [28] itassesses the degree of correspondence between matrices andvia a randomization test it provides a measure of statisticalsignificance of the relationship [28]Thematrix of similaritiesbetween epiphytic species abundance (based on the Bray-Curtis coefficient from untransformed data) was comparedwith a matrix of the similarities between biometric parame-ters (based on Euclidean distance from untransformed data)The significance of any correlation between matrices wasassessed with a randomization test
Canonical correspondence analysis (CCA) a direct gra-dient analysis technique [29] was used to investigate the rela-tionship between epiphytic species and biometric parametersof the two sampling macrophytes Epiphyte abundances datawere log(119909+1)-transformed Downweighting for rare specieswas performed We used CANOCO 45 (Scientia Software)
5 Results
Cymodocea nodosa exhibits the highest shoot density whilePosidonia oceanica has the highest leaf area index (LAI)(Figure 2) The average leaf number was 721 plusmn 213 (meanplusmn standard deviation) and 366 plusmn 112 for P oceanica and Cnodosa respectively The highest mean value of leaf length(4857 plusmn 787 cm) was recorded for P oceanica whereas thelowest mean value (163 plusmn 487 cm) was observed for Cnodosa
A total of 52 taxa of epiphytic microalgae were identified(Table 1) The highest number of collected species wasdiatoms (24) followed by dinoflagellates (22) and cyanobac-teria (6) Species number and 1198671015840 index of microepiphyticassemblage were high on Posidonia oceanica leaves (Figure 3)
Analysis of similarity (ANOSIM) of epiphytic microalgaelog(119883+1)-transformed specie abundances showed significantdifference (119877 = 0987 119875 = 002) between the P oceanicastation and C nodosa station
Analyses of similarity percentage (SIMPER) showed thatthe average dissimilarity between P oceanica and C nodosa
0
200
400
600
800
1000
1200
1400
1600
Posidonia Cymodocea0
5
10
15
20
25
30
35
40
DensityLAI
LAI(
m2m
2)
Den
sity
(sho
ots
m2)
Figure 2 Average leaf area index (LAI) and shoot density of Poceanica and Cymodocea nodosa
epiphytes groups is high (6444) This procedure alsoallowed us to determine the species that contribute to thisdissimilarity they are Oscillatoria sp Fragilaria Pseudan-abaena sp andAmphiprora constricta (Table 2) that are moreabundant on C nodosa leaves
Dinoflagellates diatoms and cyanobacteria were com-mon epiphytes on leaves (Figure 4) Abundances of thosegroups were included in the univariate analyses of variancesDinoflagellates did not differ significantly between stations(119865(110)
= 131 119875 = 0279) Significant differences weredetected for Gymnodiniales and Peridiniales among stations(119865(110)
= 286842 119875 lt 0001 119865(110)
= 166678 119875 lt 0001resp) with high abundance on leaves of Cymodocea Incontrast Prorocentrales were more abundant on the leavesof Posidonia (119865
(110)= 1579 119875 = 0003 Tukey test) Epiphytic
diatoms were more abundant on Posidonia leaves (119865(110)
=1364 119875 = 0004 Tukey test) In contrast cyanobacteriawere higher on Cymodocea leaves (119865
(110)= 174926 119875 lt
0001 Tukey test) When the abundances of toxic speciesare grouped significant differences were detected amongstations (119865
(110)= 5763 119875 lt 0001) with high abundance
on Posidonia leaves Significant differences were detected forspecies number and for1198671015840 index (119865
(110)= 34005 119875 lt 0001
119865(110)
= 1414 119875 = 0004) 119878 and1198671015840 of P oceanica were higherthan those of C nodosa (Table 3)
The results of RELATE tests indicated that there is acorrelation between the biometric parameters (shoot densityLAI and average adult and intermediate leaves) and speciesabundance (Spearman rank correlation statistic Rho = 0684119875 = 004)
Canonical correspondence analysis (CCA) indicates thatthe axis I and axis II expressed high cumulative variancespecies-biometric parameters (981 Table 4) Stations Pand C are placed separately on the left and the right of thediagrams respectively Species placed closely with samples
4 Journal of Marine Biology
Table 1 ldquoPresence-absencerdquo species list of epiphyticmicroalgae identified on the leaves of P oceanica and ofC nodosa in prospected stations
Species Posidonia CymodoceaDinoflagellates
Amphidinium carteraeHulburt 0 1Ceratium furca (Ehrenberg) Claparede and Lachmann 1 0Ceratium tripos (Muller) 1 0Coolia monotisMeunier 0 1Cyst of dinoflagellates 0 1Gonyaulax spinifera 0 1Gymnodinium sp 0 1Ostreopsis ovata (Fukuyo) 0 1Polykrikos sp 0 1Prorocentrum concavum (Fukuyo) 1 0Prorocentrum gracile (Schutt) 1 0Prorocentrum lima (Ehrenberg) Dodge 1 0Prorocentrum micans Ehrenberg 1 0Prorocentrum minimum (Pavillard) J Schiller 1 0Prorocentrum rathymum (Loeblich) Shirley and Schmidt 1 0Prorocentrum triestinum (Schiller) 1 0Protoperidinium curtipes (Jorgensen) Balech 1 0Protoperidinium depressum (Bailey) Balech 1 0Protoperidinium divergens (Ehrenberg) Balech 1 0Protoperidinium mite (Pavillard) Balech 1 0Protoperidinium ovum (Schiller) Balech 1 0Protoperidinium steinii (Joslashrgensen) Balech 1 0Protoperidiniumsp 0 1Scrippsiella sp 1 1
DiatomsAmphiprora constricta Ehrenberg 1 0Amphora sp 1 1Asterionellopsis glacialis (Castracane) Round 1 0Asteromphalus flabellatus (Brebisson) Greville 1 0Bacillariasp 1 0Budelphia sp 1 0Chaetoceros sp 1 0Climacosphenia moniligera (Ehrenberg) 1 0Coscinodiscus sp 0 1Dactyliosolen sp 1 0Diploneis sp 1 0Fragilaria sp 1 0Grammatophora sp 1 0Guinardia sp 1 1Licmophora sp 1 1Navicula sp 1 1Nitzschia fonticola Grunow 1 0Pinnularia sp 1 0Pleurosigma sp 1 0Rhizosolenia styliformis Brightwell 0 1Striatella unipunctata (Lyngbye) C Agardh 1 0Surirella sp 1 0
Journal of Marine Biology 5
Table 1 Continued
Species Posidonia CymodoceaCyanobacteria
Anabaena sp 1 1Lyngbya sp 0 1Merismopedia sp 0 1Micrococcus sp 0 1Oscillatoria sp 1 1Pseudanabaena sp 1 0
05
1015202530354045
Posidonia Cymodocea0020406081121416182
SH998400
Figure 3 Mean diversity index (1198671015840) and species number (119878) onleaves in prospected stations
010002000300040005000600070008000
Dinoflagellates Diatoms CyanobacteriaGroup
PosidoniaCymodocea
(ind100
g FW
M)
Figure 4 Average abundance of microepiphytes on leaves inprospected stations
of P oceanica (P) station belong mainly to ProrocentralesSpecies closely placed near samples of C nodosa stationbelong mainly to cyanobacteria (Figure 5)
6 Discussion
Differences between leaf epiphytic microalgae communitieswere found between C nodosa and P oceanica in our studyarea (East of Tunisia) leaf microepiphytic communities ofPosidonia were more diverse than those of C nodosa (119878 and
1198671015840 Table 3) The same results were found by Mazzella et al
[30] in Adriatic Sea and by Turki [19] in Northern TunisiaContrary to what was expected total abundance of epiphyticmicroalgae was higher on C nodosa A possible hypothesisto explain this result is that P oceanica develops much largerleaf area index values thanC nodosawhich contribute to lightattenuation within the seagrass canopy by leaf self-shading[31] Unfortunately lightmeasurementswere not taken in thisstudy but previous studies have shown that light attenuationby the canopy is reduced with decreased leaf area index[32] Therefore the higher light availability that may occurin C nodosa meadows may explain the higher abundance ofepiphytic microalgae found in these meadows
Ours results showed that microepiphytic abundances arecorrelated with shoot density and phenological parametersof the host plant (RELATE procedure) prorocentrales abun-dance was related to leaf length and leaf area index whilecyanobacteria was related to Cymodocea shootdensity (CCAprocedure Figure 5) Correlation between microepiphyticspecies and leaf phenological parameters of the host plant hasbeen also found by previous studies for P oceanica [5] andfor Zostera marina [33] Indeed differences between leavesshapes could explain epiphytic species variability betweenP oceanica and C nodosa The leaves of P oceanica areribbon-shaped (40 to 140 cm long 7 to 11mmwide) gatheredtogether in fascicles (from 5 to 8 leaves) at the tips of thestems and have a life of between 5 and 13 months [34] Theleaves of C nodosa are also ribbon-shaped but shorter thanthose of Posidonia oceanica (10 to 30 cm long 2 to 4mmwide) gathered together in smaller fascicles (from 2 to 5leaves) and have a life of between 2 and 8 months [35 36]Cymodocea nodosa has the higher value of specific growthrate (up to 47) than of P oceanica (15) [31] The shortlife span C nodosa could explain domination of fast growingspecies such as Oscillatoria sp [37] and other cyanobacteriaspecies Posidonia oceanica leaves are older they offer moretime for the epiphytic species to settle and the epiphyticcommunity will be composed of a more mature and morediverse community
The epiphytic composition is influenced by the interac-tion between the lifespan of the host and the reproductivelifespan of the epiphytes [38] Where the host is long-livedas for Posidonia local recruitment from existing epiphyteswith fast reproductive strategies can continually reinforcethe local composition For instance the density of diatomsincreased relatively quickly and became dominant (after 7 to10 weeks) in the older parts of the sheet [1 22] Cyanobacteria
6 Journal of Marine Biology
Table 2 SIMPER results
Species Group Posidonia Group Cymodocea Av Diss DissSD Contrib CumAv Abund Av Abund
Oscillatoria sp 500 6000 3019 62 4685 4685Navicula sp 393333 3850 1336 207 2074 6759Fragilaria sp 625 0 334 632 519 7278Pseudanabaena sp 600 0 331 649 513 7791Amphiprora constricta 450 0 248 574 385 8176Guinardia sp 3333 300 147 51 229 8405Anabaenasp 300 4167 142 579 221 8626Pinnularia sp 250 0 141 281 218 8844Amphora sp 100 300 109 425 169 9013
Table 3 Analysis of variance ANOVA of the abundances of major groups of microalgae and cyanobacteria in prospected stations
Source Df MS F P Tukey testTotal abundance
Habitat 1 0050 15555 0003 C gt PResiduals 10 0003
DinoflagellatesHabitat 1 0031 1312 0279 n sResiduals 10 0023
DiatomsHabitat 1 0067 13643 0004 C lt PResiduals 10 0005
CyanobacteriaHabitat 1 1310 1749266 lt0001 C gt PResiduals 10 0001
Toxic speciesHabitat 1 1255 57635 lt0001 C lt PResiduals 10 0022
Species number (S)Habitat 1 1452000 34005 lt0001 C lt PResiduals 10 42700
H1015840
Habitat 1 0032 14143 0004 C lt PResiduals 10 0002
C Cymodocea nodosameadow P Posidonia oceanicameadow n s not significant Data were log(119909 + 1)-transformed after significant Cochran C test
include some fast growing species such asOscillatoria sp [37]Borowitzka et al [38] suggest that seagrass epiphytes can beclassified into groups based on their seasonal distribution (a)epiphytes occurring throughout the year (b) epiphytes with adistinct seasonal pattern in their occurrence and (c) transientcolonizers
Difference between epiphytic compositions on the leavesof seagrass species has been studied over the world andshowed that more persistent and structurally complex sea-grass species tend to havemore diverse epiphyte assemblagesShort-lived seagrass species such as Heterozostera sp andHalodule sp are likely to be relatively depauperate in epiphytespecies richness compared to persistent seagrass species such
as Amphibolis sp [38] The findings of Aligizaki and Niko-laidis [15] showed that themaximumabundances of epiphyticdinoflagellates were recorded in branched macroalgae whilethose abundances were remarkably lower in the less leafymarine phanerogame such as Cymodocea
The two species of macrophytes made different habitatsdepending on local environmental conditions (nutrient sub-stratum current ect) our prospected Posidonia meadow isa monospecific fringing reef whereas sampling C nodosais a continuous extensive bed Differences of hydrodynamicforces between those two habitats and wave exposure canalso be responsible for species variability of the epiphyticcommunities Indeed seagrasses are able to modify current
Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
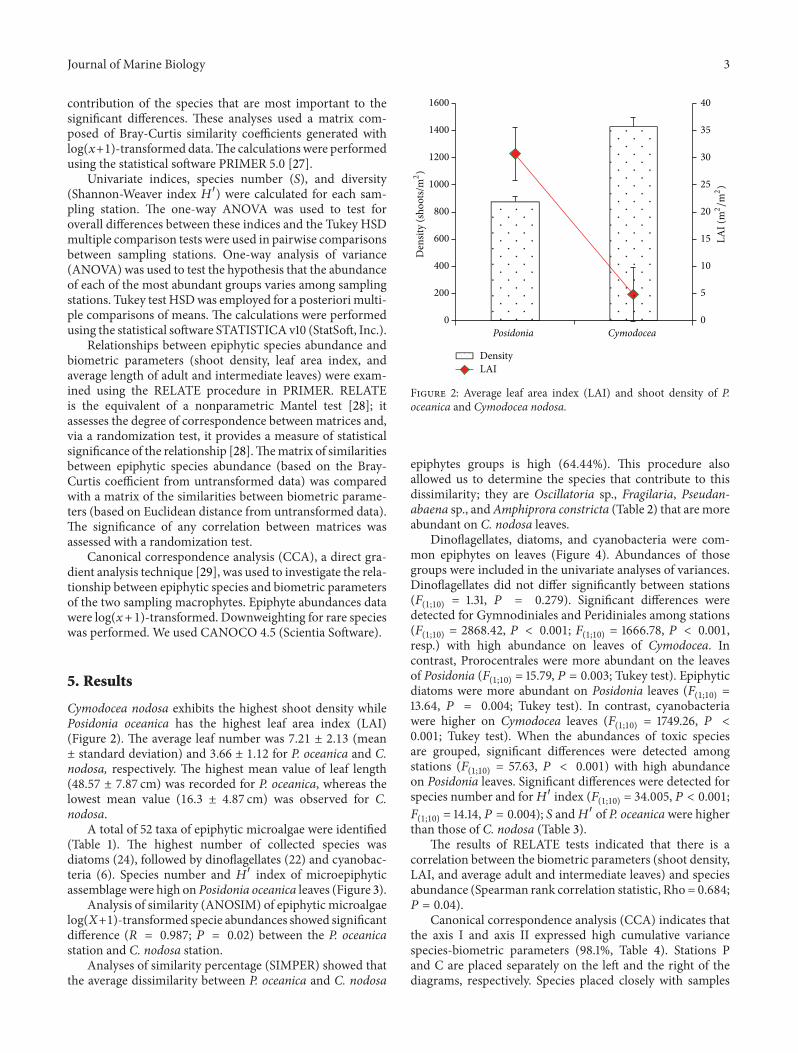
Journal of Marine Biology 3
contribution of the species that are most important to thesignificant differences These analyses used a matrix com-posed of Bray-Curtis similarity coefficients generated withlog(119909+1)-transformed dataThe calculationswere performedusing the statistical software PRIMER 50 [27]
Univariate indices species number (119878) and diversity(Shannon-Weaver index 1198671015840) were calculated for each sam-pling station The one-way ANOVA was used to test foroverall differences between these indices and the Tukey HSDmultiple comparison tests were used in pairwise comparisonsbetween sampling stations One-way analysis of variance(ANOVA) was used to test the hypothesis that the abundanceof each of the most abundant groups varies among samplingstations Tukey test HSDwas employed for a posteriori multi-ple comparisons of means The calculations were performedusing the statistical software STATISTICA v10 (StatSoft Inc)
Relationships between epiphytic species abundance andbiometric parameters (shoot density leaf area index andaverage length of adult and intermediate leaves) were exam-ined using the RELATE procedure in PRIMER RELATEis the equivalent of a nonparametric Mantel test [28] itassesses the degree of correspondence between matrices andvia a randomization test it provides a measure of statisticalsignificance of the relationship [28]Thematrix of similaritiesbetween epiphytic species abundance (based on the Bray-Curtis coefficient from untransformed data) was comparedwith a matrix of the similarities between biometric parame-ters (based on Euclidean distance from untransformed data)The significance of any correlation between matrices wasassessed with a randomization test
Canonical correspondence analysis (CCA) a direct gra-dient analysis technique [29] was used to investigate the rela-tionship between epiphytic species and biometric parametersof the two sampling macrophytes Epiphyte abundances datawere log(119909+1)-transformed Downweighting for rare specieswas performed We used CANOCO 45 (Scientia Software)
5 Results
Cymodocea nodosa exhibits the highest shoot density whilePosidonia oceanica has the highest leaf area index (LAI)(Figure 2) The average leaf number was 721 plusmn 213 (meanplusmn standard deviation) and 366 plusmn 112 for P oceanica and Cnodosa respectively The highest mean value of leaf length(4857 plusmn 787 cm) was recorded for P oceanica whereas thelowest mean value (163 plusmn 487 cm) was observed for Cnodosa
A total of 52 taxa of epiphytic microalgae were identified(Table 1) The highest number of collected species wasdiatoms (24) followed by dinoflagellates (22) and cyanobac-teria (6) Species number and 1198671015840 index of microepiphyticassemblage were high on Posidonia oceanica leaves (Figure 3)
Analysis of similarity (ANOSIM) of epiphytic microalgaelog(119883+1)-transformed specie abundances showed significantdifference (119877 = 0987 119875 = 002) between the P oceanicastation and C nodosa station
Analyses of similarity percentage (SIMPER) showed thatthe average dissimilarity between P oceanica and C nodosa
0
200
400
600
800
1000
1200
1400
1600
Posidonia Cymodocea0
5
10
15
20
25
30
35
40
DensityLAI
LAI(
m2m
2)
Den
sity
(sho
ots
m2)
Figure 2 Average leaf area index (LAI) and shoot density of Poceanica and Cymodocea nodosa
epiphytes groups is high (6444) This procedure alsoallowed us to determine the species that contribute to thisdissimilarity they are Oscillatoria sp Fragilaria Pseudan-abaena sp andAmphiprora constricta (Table 2) that are moreabundant on C nodosa leaves
Dinoflagellates diatoms and cyanobacteria were com-mon epiphytes on leaves (Figure 4) Abundances of thosegroups were included in the univariate analyses of variancesDinoflagellates did not differ significantly between stations(119865(110)
= 131 119875 = 0279) Significant differences weredetected for Gymnodiniales and Peridiniales among stations(119865(110)
= 286842 119875 lt 0001 119865(110)
= 166678 119875 lt 0001resp) with high abundance on leaves of Cymodocea Incontrast Prorocentrales were more abundant on the leavesof Posidonia (119865
(110)= 1579 119875 = 0003 Tukey test) Epiphytic
diatoms were more abundant on Posidonia leaves (119865(110)
=1364 119875 = 0004 Tukey test) In contrast cyanobacteriawere higher on Cymodocea leaves (119865
(110)= 174926 119875 lt
0001 Tukey test) When the abundances of toxic speciesare grouped significant differences were detected amongstations (119865
(110)= 5763 119875 lt 0001) with high abundance
on Posidonia leaves Significant differences were detected forspecies number and for1198671015840 index (119865
(110)= 34005 119875 lt 0001
119865(110)
= 1414 119875 = 0004) 119878 and1198671015840 of P oceanica were higherthan those of C nodosa (Table 3)
The results of RELATE tests indicated that there is acorrelation between the biometric parameters (shoot densityLAI and average adult and intermediate leaves) and speciesabundance (Spearman rank correlation statistic Rho = 0684119875 = 004)
Canonical correspondence analysis (CCA) indicates thatthe axis I and axis II expressed high cumulative variancespecies-biometric parameters (981 Table 4) Stations Pand C are placed separately on the left and the right of thediagrams respectively Species placed closely with samples
4 Journal of Marine Biology
Table 1 ldquoPresence-absencerdquo species list of epiphyticmicroalgae identified on the leaves of P oceanica and ofC nodosa in prospected stations
Species Posidonia CymodoceaDinoflagellates
Amphidinium carteraeHulburt 0 1Ceratium furca (Ehrenberg) Claparede and Lachmann 1 0Ceratium tripos (Muller) 1 0Coolia monotisMeunier 0 1Cyst of dinoflagellates 0 1Gonyaulax spinifera 0 1Gymnodinium sp 0 1Ostreopsis ovata (Fukuyo) 0 1Polykrikos sp 0 1Prorocentrum concavum (Fukuyo) 1 0Prorocentrum gracile (Schutt) 1 0Prorocentrum lima (Ehrenberg) Dodge 1 0Prorocentrum micans Ehrenberg 1 0Prorocentrum minimum (Pavillard) J Schiller 1 0Prorocentrum rathymum (Loeblich) Shirley and Schmidt 1 0Prorocentrum triestinum (Schiller) 1 0Protoperidinium curtipes (Jorgensen) Balech 1 0Protoperidinium depressum (Bailey) Balech 1 0Protoperidinium divergens (Ehrenberg) Balech 1 0Protoperidinium mite (Pavillard) Balech 1 0Protoperidinium ovum (Schiller) Balech 1 0Protoperidinium steinii (Joslashrgensen) Balech 1 0Protoperidiniumsp 0 1Scrippsiella sp 1 1
DiatomsAmphiprora constricta Ehrenberg 1 0Amphora sp 1 1Asterionellopsis glacialis (Castracane) Round 1 0Asteromphalus flabellatus (Brebisson) Greville 1 0Bacillariasp 1 0Budelphia sp 1 0Chaetoceros sp 1 0Climacosphenia moniligera (Ehrenberg) 1 0Coscinodiscus sp 0 1Dactyliosolen sp 1 0Diploneis sp 1 0Fragilaria sp 1 0Grammatophora sp 1 0Guinardia sp 1 1Licmophora sp 1 1Navicula sp 1 1Nitzschia fonticola Grunow 1 0Pinnularia sp 1 0Pleurosigma sp 1 0Rhizosolenia styliformis Brightwell 0 1Striatella unipunctata (Lyngbye) C Agardh 1 0Surirella sp 1 0
Journal of Marine Biology 5
Table 1 Continued
Species Posidonia CymodoceaCyanobacteria
Anabaena sp 1 1Lyngbya sp 0 1Merismopedia sp 0 1Micrococcus sp 0 1Oscillatoria sp 1 1Pseudanabaena sp 1 0
05
1015202530354045
Posidonia Cymodocea0020406081121416182
SH998400
Figure 3 Mean diversity index (1198671015840) and species number (119878) onleaves in prospected stations
010002000300040005000600070008000
Dinoflagellates Diatoms CyanobacteriaGroup
PosidoniaCymodocea
(ind100
g FW
M)
Figure 4 Average abundance of microepiphytes on leaves inprospected stations
of P oceanica (P) station belong mainly to ProrocentralesSpecies closely placed near samples of C nodosa stationbelong mainly to cyanobacteria (Figure 5)
6 Discussion
Differences between leaf epiphytic microalgae communitieswere found between C nodosa and P oceanica in our studyarea (East of Tunisia) leaf microepiphytic communities ofPosidonia were more diverse than those of C nodosa (119878 and
1198671015840 Table 3) The same results were found by Mazzella et al
[30] in Adriatic Sea and by Turki [19] in Northern TunisiaContrary to what was expected total abundance of epiphyticmicroalgae was higher on C nodosa A possible hypothesisto explain this result is that P oceanica develops much largerleaf area index values thanC nodosawhich contribute to lightattenuation within the seagrass canopy by leaf self-shading[31] Unfortunately lightmeasurementswere not taken in thisstudy but previous studies have shown that light attenuationby the canopy is reduced with decreased leaf area index[32] Therefore the higher light availability that may occurin C nodosa meadows may explain the higher abundance ofepiphytic microalgae found in these meadows
Ours results showed that microepiphytic abundances arecorrelated with shoot density and phenological parametersof the host plant (RELATE procedure) prorocentrales abun-dance was related to leaf length and leaf area index whilecyanobacteria was related to Cymodocea shootdensity (CCAprocedure Figure 5) Correlation between microepiphyticspecies and leaf phenological parameters of the host plant hasbeen also found by previous studies for P oceanica [5] andfor Zostera marina [33] Indeed differences between leavesshapes could explain epiphytic species variability betweenP oceanica and C nodosa The leaves of P oceanica areribbon-shaped (40 to 140 cm long 7 to 11mmwide) gatheredtogether in fascicles (from 5 to 8 leaves) at the tips of thestems and have a life of between 5 and 13 months [34] Theleaves of C nodosa are also ribbon-shaped but shorter thanthose of Posidonia oceanica (10 to 30 cm long 2 to 4mmwide) gathered together in smaller fascicles (from 2 to 5leaves) and have a life of between 2 and 8 months [35 36]Cymodocea nodosa has the higher value of specific growthrate (up to 47) than of P oceanica (15) [31] The shortlife span C nodosa could explain domination of fast growingspecies such as Oscillatoria sp [37] and other cyanobacteriaspecies Posidonia oceanica leaves are older they offer moretime for the epiphytic species to settle and the epiphyticcommunity will be composed of a more mature and morediverse community
The epiphytic composition is influenced by the interac-tion between the lifespan of the host and the reproductivelifespan of the epiphytes [38] Where the host is long-livedas for Posidonia local recruitment from existing epiphyteswith fast reproductive strategies can continually reinforcethe local composition For instance the density of diatomsincreased relatively quickly and became dominant (after 7 to10 weeks) in the older parts of the sheet [1 22] Cyanobacteria
6 Journal of Marine Biology
Table 2 SIMPER results
Species Group Posidonia Group Cymodocea Av Diss DissSD Contrib CumAv Abund Av Abund
Oscillatoria sp 500 6000 3019 62 4685 4685Navicula sp 393333 3850 1336 207 2074 6759Fragilaria sp 625 0 334 632 519 7278Pseudanabaena sp 600 0 331 649 513 7791Amphiprora constricta 450 0 248 574 385 8176Guinardia sp 3333 300 147 51 229 8405Anabaenasp 300 4167 142 579 221 8626Pinnularia sp 250 0 141 281 218 8844Amphora sp 100 300 109 425 169 9013
Table 3 Analysis of variance ANOVA of the abundances of major groups of microalgae and cyanobacteria in prospected stations
Source Df MS F P Tukey testTotal abundance
Habitat 1 0050 15555 0003 C gt PResiduals 10 0003
DinoflagellatesHabitat 1 0031 1312 0279 n sResiduals 10 0023
DiatomsHabitat 1 0067 13643 0004 C lt PResiduals 10 0005
CyanobacteriaHabitat 1 1310 1749266 lt0001 C gt PResiduals 10 0001
Toxic speciesHabitat 1 1255 57635 lt0001 C lt PResiduals 10 0022
Species number (S)Habitat 1 1452000 34005 lt0001 C lt PResiduals 10 42700
H1015840
Habitat 1 0032 14143 0004 C lt PResiduals 10 0002
C Cymodocea nodosameadow P Posidonia oceanicameadow n s not significant Data were log(119909 + 1)-transformed after significant Cochran C test
include some fast growing species such asOscillatoria sp [37]Borowitzka et al [38] suggest that seagrass epiphytes can beclassified into groups based on their seasonal distribution (a)epiphytes occurring throughout the year (b) epiphytes with adistinct seasonal pattern in their occurrence and (c) transientcolonizers
Difference between epiphytic compositions on the leavesof seagrass species has been studied over the world andshowed that more persistent and structurally complex sea-grass species tend to havemore diverse epiphyte assemblagesShort-lived seagrass species such as Heterozostera sp andHalodule sp are likely to be relatively depauperate in epiphytespecies richness compared to persistent seagrass species such
as Amphibolis sp [38] The findings of Aligizaki and Niko-laidis [15] showed that themaximumabundances of epiphyticdinoflagellates were recorded in branched macroalgae whilethose abundances were remarkably lower in the less leafymarine phanerogame such as Cymodocea
The two species of macrophytes made different habitatsdepending on local environmental conditions (nutrient sub-stratum current ect) our prospected Posidonia meadow isa monospecific fringing reef whereas sampling C nodosais a continuous extensive bed Differences of hydrodynamicforces between those two habitats and wave exposure canalso be responsible for species variability of the epiphyticcommunities Indeed seagrasses are able to modify current
Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 Journal of Marine Biology
Table 1 ldquoPresence-absencerdquo species list of epiphyticmicroalgae identified on the leaves of P oceanica and ofC nodosa in prospected stations
Species Posidonia CymodoceaDinoflagellates
Amphidinium carteraeHulburt 0 1Ceratium furca (Ehrenberg) Claparede and Lachmann 1 0Ceratium tripos (Muller) 1 0Coolia monotisMeunier 0 1Cyst of dinoflagellates 0 1Gonyaulax spinifera 0 1Gymnodinium sp 0 1Ostreopsis ovata (Fukuyo) 0 1Polykrikos sp 0 1Prorocentrum concavum (Fukuyo) 1 0Prorocentrum gracile (Schutt) 1 0Prorocentrum lima (Ehrenberg) Dodge 1 0Prorocentrum micans Ehrenberg 1 0Prorocentrum minimum (Pavillard) J Schiller 1 0Prorocentrum rathymum (Loeblich) Shirley and Schmidt 1 0Prorocentrum triestinum (Schiller) 1 0Protoperidinium curtipes (Jorgensen) Balech 1 0Protoperidinium depressum (Bailey) Balech 1 0Protoperidinium divergens (Ehrenberg) Balech 1 0Protoperidinium mite (Pavillard) Balech 1 0Protoperidinium ovum (Schiller) Balech 1 0Protoperidinium steinii (Joslashrgensen) Balech 1 0Protoperidiniumsp 0 1Scrippsiella sp 1 1
DiatomsAmphiprora constricta Ehrenberg 1 0Amphora sp 1 1Asterionellopsis glacialis (Castracane) Round 1 0Asteromphalus flabellatus (Brebisson) Greville 1 0Bacillariasp 1 0Budelphia sp 1 0Chaetoceros sp 1 0Climacosphenia moniligera (Ehrenberg) 1 0Coscinodiscus sp 0 1Dactyliosolen sp 1 0Diploneis sp 1 0Fragilaria sp 1 0Grammatophora sp 1 0Guinardia sp 1 1Licmophora sp 1 1Navicula sp 1 1Nitzschia fonticola Grunow 1 0Pinnularia sp 1 0Pleurosigma sp 1 0Rhizosolenia styliformis Brightwell 0 1Striatella unipunctata (Lyngbye) C Agardh 1 0Surirella sp 1 0
Journal of Marine Biology 5
Table 1 Continued
Species Posidonia CymodoceaCyanobacteria
Anabaena sp 1 1Lyngbya sp 0 1Merismopedia sp 0 1Micrococcus sp 0 1Oscillatoria sp 1 1Pseudanabaena sp 1 0
05
1015202530354045
Posidonia Cymodocea0020406081121416182
SH998400
Figure 3 Mean diversity index (1198671015840) and species number (119878) onleaves in prospected stations
010002000300040005000600070008000
Dinoflagellates Diatoms CyanobacteriaGroup
PosidoniaCymodocea
(ind100
g FW
M)
Figure 4 Average abundance of microepiphytes on leaves inprospected stations
of P oceanica (P) station belong mainly to ProrocentralesSpecies closely placed near samples of C nodosa stationbelong mainly to cyanobacteria (Figure 5)
6 Discussion
Differences between leaf epiphytic microalgae communitieswere found between C nodosa and P oceanica in our studyarea (East of Tunisia) leaf microepiphytic communities ofPosidonia were more diverse than those of C nodosa (119878 and
1198671015840 Table 3) The same results were found by Mazzella et al
[30] in Adriatic Sea and by Turki [19] in Northern TunisiaContrary to what was expected total abundance of epiphyticmicroalgae was higher on C nodosa A possible hypothesisto explain this result is that P oceanica develops much largerleaf area index values thanC nodosawhich contribute to lightattenuation within the seagrass canopy by leaf self-shading[31] Unfortunately lightmeasurementswere not taken in thisstudy but previous studies have shown that light attenuationby the canopy is reduced with decreased leaf area index[32] Therefore the higher light availability that may occurin C nodosa meadows may explain the higher abundance ofepiphytic microalgae found in these meadows
Ours results showed that microepiphytic abundances arecorrelated with shoot density and phenological parametersof the host plant (RELATE procedure) prorocentrales abun-dance was related to leaf length and leaf area index whilecyanobacteria was related to Cymodocea shootdensity (CCAprocedure Figure 5) Correlation between microepiphyticspecies and leaf phenological parameters of the host plant hasbeen also found by previous studies for P oceanica [5] andfor Zostera marina [33] Indeed differences between leavesshapes could explain epiphytic species variability betweenP oceanica and C nodosa The leaves of P oceanica areribbon-shaped (40 to 140 cm long 7 to 11mmwide) gatheredtogether in fascicles (from 5 to 8 leaves) at the tips of thestems and have a life of between 5 and 13 months [34] Theleaves of C nodosa are also ribbon-shaped but shorter thanthose of Posidonia oceanica (10 to 30 cm long 2 to 4mmwide) gathered together in smaller fascicles (from 2 to 5leaves) and have a life of between 2 and 8 months [35 36]Cymodocea nodosa has the higher value of specific growthrate (up to 47) than of P oceanica (15) [31] The shortlife span C nodosa could explain domination of fast growingspecies such as Oscillatoria sp [37] and other cyanobacteriaspecies Posidonia oceanica leaves are older they offer moretime for the epiphytic species to settle and the epiphyticcommunity will be composed of a more mature and morediverse community
The epiphytic composition is influenced by the interac-tion between the lifespan of the host and the reproductivelifespan of the epiphytes [38] Where the host is long-livedas for Posidonia local recruitment from existing epiphyteswith fast reproductive strategies can continually reinforcethe local composition For instance the density of diatomsincreased relatively quickly and became dominant (after 7 to10 weeks) in the older parts of the sheet [1 22] Cyanobacteria
6 Journal of Marine Biology
Table 2 SIMPER results
Species Group Posidonia Group Cymodocea Av Diss DissSD Contrib CumAv Abund Av Abund
Oscillatoria sp 500 6000 3019 62 4685 4685Navicula sp 393333 3850 1336 207 2074 6759Fragilaria sp 625 0 334 632 519 7278Pseudanabaena sp 600 0 331 649 513 7791Amphiprora constricta 450 0 248 574 385 8176Guinardia sp 3333 300 147 51 229 8405Anabaenasp 300 4167 142 579 221 8626Pinnularia sp 250 0 141 281 218 8844Amphora sp 100 300 109 425 169 9013
Table 3 Analysis of variance ANOVA of the abundances of major groups of microalgae and cyanobacteria in prospected stations
Source Df MS F P Tukey testTotal abundance
Habitat 1 0050 15555 0003 C gt PResiduals 10 0003
DinoflagellatesHabitat 1 0031 1312 0279 n sResiduals 10 0023
DiatomsHabitat 1 0067 13643 0004 C lt PResiduals 10 0005
CyanobacteriaHabitat 1 1310 1749266 lt0001 C gt PResiduals 10 0001
Toxic speciesHabitat 1 1255 57635 lt0001 C lt PResiduals 10 0022
Species number (S)Habitat 1 1452000 34005 lt0001 C lt PResiduals 10 42700
H1015840
Habitat 1 0032 14143 0004 C lt PResiduals 10 0002
C Cymodocea nodosameadow P Posidonia oceanicameadow n s not significant Data were log(119909 + 1)-transformed after significant Cochran C test
include some fast growing species such asOscillatoria sp [37]Borowitzka et al [38] suggest that seagrass epiphytes can beclassified into groups based on their seasonal distribution (a)epiphytes occurring throughout the year (b) epiphytes with adistinct seasonal pattern in their occurrence and (c) transientcolonizers
Difference between epiphytic compositions on the leavesof seagrass species has been studied over the world andshowed that more persistent and structurally complex sea-grass species tend to havemore diverse epiphyte assemblagesShort-lived seagrass species such as Heterozostera sp andHalodule sp are likely to be relatively depauperate in epiphytespecies richness compared to persistent seagrass species such
as Amphibolis sp [38] The findings of Aligizaki and Niko-laidis [15] showed that themaximumabundances of epiphyticdinoflagellates were recorded in branched macroalgae whilethose abundances were remarkably lower in the less leafymarine phanerogame such as Cymodocea
The two species of macrophytes made different habitatsdepending on local environmental conditions (nutrient sub-stratum current ect) our prospected Posidonia meadow isa monospecific fringing reef whereas sampling C nodosais a continuous extensive bed Differences of hydrodynamicforces between those two habitats and wave exposure canalso be responsible for species variability of the epiphyticcommunities Indeed seagrasses are able to modify current
Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Journal of Marine Biology 5
Table 1 Continued
Species Posidonia CymodoceaCyanobacteria
Anabaena sp 1 1Lyngbya sp 0 1Merismopedia sp 0 1Micrococcus sp 0 1Oscillatoria sp 1 1Pseudanabaena sp 1 0
05
1015202530354045
Posidonia Cymodocea0020406081121416182
SH998400
Figure 3 Mean diversity index (1198671015840) and species number (119878) onleaves in prospected stations
010002000300040005000600070008000
Dinoflagellates Diatoms CyanobacteriaGroup
PosidoniaCymodocea
(ind100
g FW
M)
Figure 4 Average abundance of microepiphytes on leaves inprospected stations
of P oceanica (P) station belong mainly to ProrocentralesSpecies closely placed near samples of C nodosa stationbelong mainly to cyanobacteria (Figure 5)
6 Discussion
Differences between leaf epiphytic microalgae communitieswere found between C nodosa and P oceanica in our studyarea (East of Tunisia) leaf microepiphytic communities ofPosidonia were more diverse than those of C nodosa (119878 and
1198671015840 Table 3) The same results were found by Mazzella et al
[30] in Adriatic Sea and by Turki [19] in Northern TunisiaContrary to what was expected total abundance of epiphyticmicroalgae was higher on C nodosa A possible hypothesisto explain this result is that P oceanica develops much largerleaf area index values thanC nodosawhich contribute to lightattenuation within the seagrass canopy by leaf self-shading[31] Unfortunately lightmeasurementswere not taken in thisstudy but previous studies have shown that light attenuationby the canopy is reduced with decreased leaf area index[32] Therefore the higher light availability that may occurin C nodosa meadows may explain the higher abundance ofepiphytic microalgae found in these meadows
Ours results showed that microepiphytic abundances arecorrelated with shoot density and phenological parametersof the host plant (RELATE procedure) prorocentrales abun-dance was related to leaf length and leaf area index whilecyanobacteria was related to Cymodocea shootdensity (CCAprocedure Figure 5) Correlation between microepiphyticspecies and leaf phenological parameters of the host plant hasbeen also found by previous studies for P oceanica [5] andfor Zostera marina [33] Indeed differences between leavesshapes could explain epiphytic species variability betweenP oceanica and C nodosa The leaves of P oceanica areribbon-shaped (40 to 140 cm long 7 to 11mmwide) gatheredtogether in fascicles (from 5 to 8 leaves) at the tips of thestems and have a life of between 5 and 13 months [34] Theleaves of C nodosa are also ribbon-shaped but shorter thanthose of Posidonia oceanica (10 to 30 cm long 2 to 4mmwide) gathered together in smaller fascicles (from 2 to 5leaves) and have a life of between 2 and 8 months [35 36]Cymodocea nodosa has the higher value of specific growthrate (up to 47) than of P oceanica (15) [31] The shortlife span C nodosa could explain domination of fast growingspecies such as Oscillatoria sp [37] and other cyanobacteriaspecies Posidonia oceanica leaves are older they offer moretime for the epiphytic species to settle and the epiphyticcommunity will be composed of a more mature and morediverse community
The epiphytic composition is influenced by the interac-tion between the lifespan of the host and the reproductivelifespan of the epiphytes [38] Where the host is long-livedas for Posidonia local recruitment from existing epiphyteswith fast reproductive strategies can continually reinforcethe local composition For instance the density of diatomsincreased relatively quickly and became dominant (after 7 to10 weeks) in the older parts of the sheet [1 22] Cyanobacteria
6 Journal of Marine Biology
Table 2 SIMPER results
Species Group Posidonia Group Cymodocea Av Diss DissSD Contrib CumAv Abund Av Abund
Oscillatoria sp 500 6000 3019 62 4685 4685Navicula sp 393333 3850 1336 207 2074 6759Fragilaria sp 625 0 334 632 519 7278Pseudanabaena sp 600 0 331 649 513 7791Amphiprora constricta 450 0 248 574 385 8176Guinardia sp 3333 300 147 51 229 8405Anabaenasp 300 4167 142 579 221 8626Pinnularia sp 250 0 141 281 218 8844Amphora sp 100 300 109 425 169 9013
Table 3 Analysis of variance ANOVA of the abundances of major groups of microalgae and cyanobacteria in prospected stations
Source Df MS F P Tukey testTotal abundance
Habitat 1 0050 15555 0003 C gt PResiduals 10 0003
DinoflagellatesHabitat 1 0031 1312 0279 n sResiduals 10 0023
DiatomsHabitat 1 0067 13643 0004 C lt PResiduals 10 0005
CyanobacteriaHabitat 1 1310 1749266 lt0001 C gt PResiduals 10 0001
Toxic speciesHabitat 1 1255 57635 lt0001 C lt PResiduals 10 0022
Species number (S)Habitat 1 1452000 34005 lt0001 C lt PResiduals 10 42700
H1015840
Habitat 1 0032 14143 0004 C lt PResiduals 10 0002
C Cymodocea nodosameadow P Posidonia oceanicameadow n s not significant Data were log(119909 + 1)-transformed after significant Cochran C test
include some fast growing species such asOscillatoria sp [37]Borowitzka et al [38] suggest that seagrass epiphytes can beclassified into groups based on their seasonal distribution (a)epiphytes occurring throughout the year (b) epiphytes with adistinct seasonal pattern in their occurrence and (c) transientcolonizers
Difference between epiphytic compositions on the leavesof seagrass species has been studied over the world andshowed that more persistent and structurally complex sea-grass species tend to havemore diverse epiphyte assemblagesShort-lived seagrass species such as Heterozostera sp andHalodule sp are likely to be relatively depauperate in epiphytespecies richness compared to persistent seagrass species such
as Amphibolis sp [38] The findings of Aligizaki and Niko-laidis [15] showed that themaximumabundances of epiphyticdinoflagellates were recorded in branched macroalgae whilethose abundances were remarkably lower in the less leafymarine phanerogame such as Cymodocea
The two species of macrophytes made different habitatsdepending on local environmental conditions (nutrient sub-stratum current ect) our prospected Posidonia meadow isa monospecific fringing reef whereas sampling C nodosais a continuous extensive bed Differences of hydrodynamicforces between those two habitats and wave exposure canalso be responsible for species variability of the epiphyticcommunities Indeed seagrasses are able to modify current
Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 Journal of Marine Biology
Table 2 SIMPER results
Species Group Posidonia Group Cymodocea Av Diss DissSD Contrib CumAv Abund Av Abund
Oscillatoria sp 500 6000 3019 62 4685 4685Navicula sp 393333 3850 1336 207 2074 6759Fragilaria sp 625 0 334 632 519 7278Pseudanabaena sp 600 0 331 649 513 7791Amphiprora constricta 450 0 248 574 385 8176Guinardia sp 3333 300 147 51 229 8405Anabaenasp 300 4167 142 579 221 8626Pinnularia sp 250 0 141 281 218 8844Amphora sp 100 300 109 425 169 9013
Table 3 Analysis of variance ANOVA of the abundances of major groups of microalgae and cyanobacteria in prospected stations
Source Df MS F P Tukey testTotal abundance
Habitat 1 0050 15555 0003 C gt PResiduals 10 0003
DinoflagellatesHabitat 1 0031 1312 0279 n sResiduals 10 0023
DiatomsHabitat 1 0067 13643 0004 C lt PResiduals 10 0005
CyanobacteriaHabitat 1 1310 1749266 lt0001 C gt PResiduals 10 0001
Toxic speciesHabitat 1 1255 57635 lt0001 C lt PResiduals 10 0022
Species number (S)Habitat 1 1452000 34005 lt0001 C lt PResiduals 10 42700
H1015840
Habitat 1 0032 14143 0004 C lt PResiduals 10 0002
C Cymodocea nodosameadow P Posidonia oceanicameadow n s not significant Data were log(119909 + 1)-transformed after significant Cochran C test
include some fast growing species such asOscillatoria sp [37]Borowitzka et al [38] suggest that seagrass epiphytes can beclassified into groups based on their seasonal distribution (a)epiphytes occurring throughout the year (b) epiphytes with adistinct seasonal pattern in their occurrence and (c) transientcolonizers
Difference between epiphytic compositions on the leavesof seagrass species has been studied over the world andshowed that more persistent and structurally complex sea-grass species tend to havemore diverse epiphyte assemblagesShort-lived seagrass species such as Heterozostera sp andHalodule sp are likely to be relatively depauperate in epiphytespecies richness compared to persistent seagrass species such
as Amphibolis sp [38] The findings of Aligizaki and Niko-laidis [15] showed that themaximumabundances of epiphyticdinoflagellates were recorded in branched macroalgae whilethose abundances were remarkably lower in the less leafymarine phanerogame such as Cymodocea
The two species of macrophytes made different habitatsdepending on local environmental conditions (nutrient sub-stratum current ect) our prospected Posidonia meadow isa monospecific fringing reef whereas sampling C nodosais a continuous extensive bed Differences of hydrodynamicforces between those two habitats and wave exposure canalso be responsible for species variability of the epiphyticcommunities Indeed seagrasses are able to modify current
Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Journal of Marine Biology 7
Table 4 Summary of canonical correspondence analysis (CCA)of the three data matrices (epiphytic microalgae stations andbiometric parametersvariables)
Axis 1 Axis 2Eigenvalues 0564 0020Species-environment correlations 0992 0760Cumulative percentage variance ofspecies data 696 720
Cumulative percentage variance ofspecies-environment relation 948 981
Sum of all eigenvalues 0810Sum of all canonical eigenvalues 0595Explained variance by CCA 7345Intraset correlations of variables with axes
Density 09581 minus01771Average of adult leaf length minus09645 minus00258LAI minus09591 minus01494
flow and this depending on species [39] For instance waveattenuation is the highest when seagrasses occupy a largeportion of the water column such as Posidonia oceanica bed[40] As a result bed with high canopy is slower than low-canopy seagrass bed [41 42] The canopy of P oceanica tendsto attenuate currents and waves thereby reducing the forcesexerted on individual shoots [42] Even at the edge of thecanopy seagrass shoots may be sheltered to a certain extentby the presence of adjacent shoots [43] Natural fluctuationsin water flow also affect the epiphytic community If anepiphytic community develops during relatively calm con-ditions species with high drag (ie large area exposed tothe flow) may become dominant but if the flow increasesover a short period of time (eg storms) these epiphytes arethen removed [44] Our study is carried out in September(summer season) which is fairly quiet period since thestorms intensified especially in autumn in the easternTunisiaConsequently the effect of the removal of epiphytes by stronghydrodynamics would be less important
The epiphyte-grazer interaction also plays an importantrole in controlling the abundance and diversity of epiphytesIndeed the epiphytes of marine macrophytes are a foodsource for a range of grazers and in turn they influencethe diversity and abundance of epiphytes by removing thesubstrate and biomass of the host plant [38] The effect ofgrazing is also an important factor that contributes to thevariation between the two macrophytes species as manyspecies feed on epiphytes of Posidonia including the echinoidParacentrotus lividus some decapods and amphipods [4546] In addition about 50 species of fish are encountered in Poceanica beds [47]
Grazers can be highly selective and thus may have astrong effect on the spatial pattern of the periphytic com-munity [48] Some grazers (scrapers) feed preferentially ontightly attached diatoms [46] whereas others (surfers) favoroverstory diatoms [49] Neckles et al [50] have shown thatnumber of diatoms decreased in the presence of grazers Inaddition strong currents (andor high waves) may eliminate
10
minus15minus15 15
P
PP
P
P
P
CC
C C
C
C
11
171626
22
19
21
28
10
15
12
27
23
30
24
8
2
25
133729
6
45
18
LAI
Lm
D
Figure 5 Diagram of canonical correspondence analysis (CCA)showing the effects of biometric parameters of the host plant onepiphytic microalgae species and their ordination according to thefirst and second axes Species whose coverage and frequency areless than 10 were eliminated Species that were grouped withPosidonia station (P) are 10 Prorocentrum concavum 11 Prorocen-trum minimum 12 Prorocentrum lima 15 Nitzschia fonticola 16Fragilaria sp 17 Surirella sp 19 Diploneis sp 21 Pleurosigma sp23 Pinnularia sp 26 Amphiprora constricta 27 Anabaena sp and28 Pseudanabaena sp Species that are grouped with Cymodocea(C) station are 2 Gymnodinium sp 3 Amphidinium carterae 4Gonyaulax spinifera 5 Cyst of dinoflagellates 6 Protoperidiniumsp 8 Coolia monotis 13 Rhizosolenia styliformis 18 Guinardia sp24 Amphora sp 25 Nitzschia sp 29 Coscinodiscus sp and 30Oscillatoria sp
grazers allowing for more epiphytes to grow [51] Cyanobac-teria have different mechanisms to reduce the grazing pres-sure by large-bodied cladocerans They can form colonieswith sizes beyond the ingestion capacity cyanobacteria mayproduce grazing deterrents and potent endotoxins that arereleased upon digestion while cells that are embedded inmucus could be resistant to digestion [52]
Diatom abundance was higher in P oceanica they aredominated by the order of naviculales which includes themost abundant species present on the leaves ofP oceanica [5354] Various studies have suggested that diatoms and bacteriaare among the first organisms to colonize the submergedobjects or organisms and they shape a biofilm influencing thesettlement of some invertebrate larvae [53] The diatoms alsoconstitute a food source for various organisms living in thePosidonia canopy [53ndash55]
Dinoflagellates were more abundant on the leaves of Poceanica especially the order of prorocentrales Abundanceof potentially toxic dinoflagellates on Posidonia leaves washigher than those onC nodosa leavesThese findings supportthose of Romdhane et al [56] and Armi et al [57] in Tunisiaand those of Aligizaki and Nikolaidis [15] in the northof the Mediterranean Sea They are Amphidinium carteraeCoolia monatis Ostreopsis ovata Prorocentrum concavumP minimum P rathymum P triestinum P micans andP lima Those species are potential toxin producers [58]
8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 Journal of Marine Biology
For instance epiphytic Prorocentrum species are mainlyassociated with okadaic acid and the production of analogues[59] These results are particularly useful in this area with therecent establishment of several fish farms because epiphyticdinoflagellates are easily resuspended in the water column[8] and that abundance of phytoplankton in vegetated areasis higher than in the unvegetated station [16] MoreoverMarr et al [60] concluded that the underestimation oftoxic dinoflagellates associated with a toxic event may bedue in part to the lack of sampling of the benthic andepiphytic communities Finally it is necessary to improve amanagement program to protect seagrass species and theirassociated epiphytes in eastern Tunisia and ovoid installationof aquaculture farm around seagrass beds whenever possiblenot only to preserve this vulnerable ecosystems [35] but alsoto ovoid a possible contamination by epiphytic potentiallytoxic species
7 Conclusion
Abundance and species composition of epiphytic microalgaewere different on the leaves of P oceanica and C nodosaLocal environmental conditions (hydrodynamics light pen-etration) host characteristics (meadow type shape formsof leaves life span and growth rate) and grazing effectseem to be responsible for these dissimilarities in epiphyticmicroalgae communities
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by the research project EBHARldquoEtat du Benthos et des Habitats Remarquablesrdquo of theNational Institute of Sciences and Technology of the Sea(INSTM) The authors thank Dr Sarra Mabrouk of NationalInstitute of Sciences for help The authors wish to acknowl-edge use of the MapTool program for graphics in this paperThey also thank ProfessorAndrewMcMinn and reviewers fortheir comments which improved the quality of the paper
References
[1] L Mazzella and G F Russo ldquoGrazing effect of two Gibbulaspecies (Mollusca Archaeogastropoda) on the epiphytic com-munity of Posidonia oceanica leavesrdquo Aquatic Botany vol 35no 3-4 pp 357ndash373 1989
[2] L Mazzella M C Buia and L Spinoccia ldquoBiodiversity ofepiphytic diatom community on leaves of Posidonia oceanicardquoin Proceedings of the 13th Diatom Symposium DMarino andMMontresor Eds Biopress Bristol UK 1994
[3] M C Gambi M Lorenti G F Russo M B Scipione and VZupo ldquoDepth and seasonal distribution of some groups of thevagile fauna of the Posidonia oceanica leaf stratum structuraland trophic analysesrdquo Marine Ecology vol 13 no 1 pp 17ndash391992
[4] P Prado T Alcoverro B Martınez-Crego A Verges M Perezand J Romero ldquoMacrograzers strongly influence patterns ofepiphytic assemblages in seagrass meadowsrdquo Journal of Experi-mentalMarine Biology and Ecology vol 350 no 1-2 pp 130ndash1432007
[5] LMabrouk A HamzaM B Brahim andM-N Bradai ldquoTem-poral and depth distribution of microepiphytes on Posidoniaoceanica (L) Delile leaves in a meadow off Tunisiardquo MarineEcology vol 32 no 2 pp 148ndash161 2011
[6] P S Lavery T Reid G A Hyndes and B R Van Elven ldquoEffectof leaf movement on epiphytic algal biomass of seagrass leavesrdquoMarine Ecology Progress Series vol 338 pp 97ndash106 2007
[7] K-S Lee S R Park and Y K Kim ldquoEffects of irradiancetemperature and nutrients on growth dynamics of seagrassesa reviewrdquo Journal of Experimental Marine Biology and Ecologyvol 350 no 1-2 pp 144ndash175 2007
[8] L Mabrouk A Hamza M Mahfoudi and M-N Bradai ldquoSpa-tial and temporal variations of epiphytic Ostreopsis siamensison Posidonia oceanica (L) Delile leaves in Mahdia (Tunisia)rdquoCahiers de Biologie Marine vol 53 no 4 pp 419ndash427 2012
[9] T J B Carruthers Leaf production canopy structure and lightclimate in a density-manipulated Amphibolis griffithii meadow[MS thesis] University of Western Australia 1994
[10] I Castejon-Silvo and J Terrados ldquoPatterns of spatial variationof nutrient content epiphyte load and shoot size of Posidoniaoceanica seagrass meadows (Mediterranean Sea)rdquoMarine Ecol-ogy vol 33 no 2 pp 165ndash175 2012
[11] D A Bulthuis and W J Woelkerling ldquoBiomass accumulationand shading effects of epiphytes on leaves of the seagrassHeterozostera tasmanica inVictoria AustraliardquoAquatic Botanyvol 16 no 2 pp 137ndash148 1983
[12] M A Borowitzka R C Lethbridge and L Charlton ldquoSpeciesrichness spatial distribution and colonisation pattern of algaland invertebrate epiphytes on the seagrassAmphiholis grifiithiirdquoMarine Ecology Progress Series vol 64 pp 281ndash291 1990
[13] M A Faust ldquoMorphology of ciguatera-causing Prorocentrumlima (Pyrrhophyta) from widely differing sitesrdquo Journal ofPhycology vol 27 pp 642ndash648 1991
[14] J Foden D A Purdie S Morris and S Nascimento ldquoEpiphyticabundance and toxicity of Prorocentrum lima populations in theFleet Lagoon UKrdquo Harmful Algae vol 4 no 6 pp 1063ndash10742005
[15] K Aligizaki and G Nikolaidis ldquoThe presence of the potentiallytoxic genera Ostreopsis and Coolia (Dinophyceae) in the NorthAegean Sea Greecerdquo Harmful Algae vol 5 no 6 pp 717ndash7302006
[16] L Mabrouk A Hamza and M N Bradai ldquoVariability inthe structure of planktonic microalgae assemblages in watercolumn associated with Posidonia oceanica (L) bed in TunisiardquoJournal of Marine Biology vol 2014 Article ID 621238 7 pages2014
[17] M Vila E Garces and M Maso ldquoPotentially toxic epiphyticdinoflagellate assemblages on macroalgae in the NW Mediter-raneanrdquoAquaticMicrobial Ecology vol 26 no 1 pp 51ndash60 2001
[18] G Giraud ldquoSur une methode de mesure et de comptage desstructures foliaires de Posidonia oceanica (Linnaeus) DelilerdquoBulletin Museum drsquoHistoire Naturelle de Marseille vol 39 pp33ndash39 1979
[19] S Turki ldquoDistribution of toxic dinoflagellates along the leavesof seagrass Posidonia oceanica and Cymodocea nodosa from theGulf of Tunisrdquo Cahiers de Biologie Marine vol 46 no 1 pp 29ndash34 2005
Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Journal of Marine Biology 9
[20] P A Penhale ldquoMacrophyte-epiphyte biomass and productivityin an eelgrass (Zostera marina L) communityrdquo Journal ofExperimentalMarine Biology and Ecology vol 26 no 2 pp 211ndash224 1977
[21] F T Short ldquoPrimary elemental constituentsrdquo in SeagrassResearchMethods R C Phillips and C P McRoy Eds pp 105ndash110 UNESCO Paris France 1990
[22] P Dauby and M Poulicek ldquoMethods for removing epiphytesfrom seagrasses SEM observations on treated leavesrdquo AquaticBotany vol 52 no 3 pp 217ndash228 1995
[23] G A Kendrick and P S Lavery ldquoAssessing biomass assemblagestructure and productivity of algal epiphytes on seagrassesrdquo inGlogal Seagrass Research Methods F T Short and R G ColesEds Elsevier Amsterdam The Netherlands 2001
[24] H Utermohl ldquoZur vervollkommung der quantitativenPhytomicroorganisms-Methodik Mitteilungenrdquo Verhandlu-ngen der Internationalen Vereinigung fur Theoretische undAngewandte Limnologie vol 9 pp 1ndash38 1958
[25] J H Zar Biostatistical Analysis Prentice Hall Upper SaddleRiver NJ USA 4th edition 1999
[26] K R Clarke ldquoNon-parametric multivariate analyses of changesin community structurerdquo Australian Journal of Ecology vol 18no 1 pp 117ndash143 1993
[27] K R Clarke and R M Warwick Change in Marine Commu-nities An Approach to Statistical Analysis and InterpretationPRIMER-E Plymouth UK 2001
[28] P J Somerfield K R Clarke and F Olsgard ldquoA comparisonof the power of categorical and correlational tests applied tocommunity ecology data from gradient studiesrdquo Journal ofAnimal Ecology vol 71 no 4 pp 581ndash593 2002
[29] C J Ter Braak and P F Verdonschot ldquoCanonical correspon-dence analysis and related multivariate methods in aquaticecologyrdquo Aquatic Sciences vol 57 no 3 pp 255ndash289 1995
[30] L Mazzella P Guidetti M Lorenti et al ldquoBiomass partitioninginAdriatic seagrass ecosystems (Posidonia oceanicaCymodoceanodosa Zostera marina)rdquo Rapport Commission InternationalMer Mediterranee vol 35 pp 562ndash563 1998
[31] D Lobelle E J Kenyon K J Cook and J C Bull ldquoLocalcompetition and metapopulation processes drive long-termseagrass-epiphyte population dynamicsrdquo PLoS ONE vol 8 no2 Article ID e57072 2013
[32] S Enrıquez and N I Pantoja-Reyes ldquoForm-function analysisof the effect of canopy morphology on leaf self-shading in theseagrass Thalassia testudinumrdquo Oecologia vol 145 no 2 pp235ndash243 2005
[33] M P Johnson M Edwards F Bunker and C A Maggs ldquoAlgalepiphytes of Zostera marina variation in assemblage structurefrom individual leaves to regional scalerdquoAquatic Botany vol 82no 1 pp 12ndash26 2005
[34] G Pergent and C Pergent-Martini ldquoSome applications of lep-idochronological analysis in the seagrass Posidonia oceanicardquoBotanica Marina vol 33 pp 299ndash310 1990
[35] E P Green and F T Short World Atlas of Seagrasses UIMEPWorld Conservation Monitoring Centre University of Califor-nia Press Berkeley Calif USA 2003
[36] J Reyes and M Sanson ldquoTemporal distribution and reproduc-tive phenology of the epiphytes on Cymodocea nodosa leaves inthe Canary IslandsrdquoBotanicaMarina vol 40 no 3 pp 193ndash2011997
[37] M Prabakaran ldquoInvitro antimicrobial potentials ofmarineOsc-illatoria speciesrdquo Asian Journal of Plant Science and Researchvol 1 no 3 pp 58ndash64 2011
[38] M A Borowitzka P Lavery and M Keulen ldquoEpiphytes ofseagrassesrdquo in Seagrasses Biology Ecology and Conservation AW D Larkum R J Orth and C M Duarte Eds SpringerAmsterdam The Netherlands 2006
[39] T Seaborn ldquoLimpets and their algal epibionts costs and benefitsof Acrosiphonia spp and ulva lactuca growthrdquo Journal of MarineBiology vol 2014 Article ID 891943 7 pages 2014
[40] M S Fonseca and J A Cahalan ldquoA preliminary evaluation ofwave attenuation by four species of seagrassrdquo Estuarine Coastaland Shelf Science vol 35 no 6 pp 565ndash576 1992
[41] M C Gambi A R M Nowell and P A Jumars ldquoFlumeobservations on flow dynamics in Zostera marina (eelgrass)bedsrdquoMarine Ecology Progress Series vol 61 pp 159ndash169 1990
[42] E W Koch J D Ackerman J Verduin and M van KeulenldquoFluid dynamics in seagrass ecologyrdquo in Seagrasses BiologyEcology and Conservation A W D Larkum R J Orth and CM Duarte Eds Springer Amsterdam The Netherlands 2006
[43] T C Granata T Serra J Colomer X Casamitjana C MDuarte and E Gacia ldquoFlow and particle distributions in anearshore seagrass meadow before and after a stormrdquo MarineEcology Progress Series vol 218 pp 95ndash106 2001
[44] B J F Biggs ldquoHydraulic habitat of plants in streamsrdquo RegulatedRivers Research and Management vol 12 no 2-3 pp 131ndash1441996
[45] R Zakhama-Sraieb Y R Sghaier and F Charfi-CheikhrouhaldquoCommunity structure of amphipods on shallow Posidoniaoceanica meadows off Tunisian coastsrdquo Helgoland MarineResearch vol 65 no 2 pp 203ndash209 2011
[46] W Belgacem H Langar G Pergent and O K Ben Has-sine ldquoAssociated mollusc communities of a Posidonia oceanicameadow inCap Zebib (offNorth East Tunisia)rdquoAquatic Botanyvol 104 pp 170ndash175 2013
[47] M L Harmelin-Vivien ldquoIchtyofaune des herbiers de posidoniesdu parc naturel regional deCorserdquo in InternationalWorkshop onPosidonia Beds C F Boudouresque AMeinesz E Fresi and VGravez Eds GIS Posidonie Marseille France 1989
[48] O Sarnelle K W Kratz and S D Cooper ldquoEffects of aninvertebrate grazer on the spatial arrangement of a benthicmicrohabitatrdquo Oecologia vol 96 no 2 pp 208ndash218 1993
[49] L Tall A Cattaneo L Cloutier S Dray and P LegendreldquoResource partitioning in a grazer guild feeding on a multilayerdiatom matrdquo Journal of the North American BenthologicalSociety vol 25 no 4 pp 800ndash810 2006
[50] H A Neckles E T Koepfler L W Haas R L Wetzel andR J Orth ldquoDynamics of epiphytic photoautotrophs and het-erotrophs in Zostera marina (Eelgrass) microcosms responsesto nutrient enrichment and grazingrdquo Estuaries vol 17 no 3 pp597ndash605 1994
[51] A Schanz P Polte and H Asmus ldquoCascading effects of hydro-dynamics on an epiphyte-grazer system in intertidal seagrassbeds of theWadden SeardquoMarine Biology vol 141 no 2 pp 287ndash297 2002
[52] I M Hamisi T J Lyimo andM H S Muruke ldquoCyanobacterialoccurrence and diversity in seagrass meadows in coastal tanza-niardquoWestern Indian Ocean Journal of Marine Science vol 3 no2 pp 113ndash122 2004
[53] R Novak ldquoA study in ultra-ecology microorganisms on theseagrass Posidonia oceanica (L) Delilerdquo Marine Ecology vol 5no 2 pp 143ndash190 1984
[54] L Mazzella M B Scipione and M C Buia ldquoSpatio-temporaldistribution of algal and animal communities in a Posidonia
10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 Journal of Marine Biology
oceanica meadowrdquo Marine Ecology vol 10 no 2 pp 107ndash1291989
[55] T Ribes H Salvado J Romero and M D P GracialdquoForaminiferal colonization on artificial seagrass leavesrdquo Jour-nal of Foraminiferal Research vol 30 no 3 pp 192ndash201 2000
[56] M S Romdhane H C Eilertsen O K D Yahia and M ND Yahia ldquoToxic dinoflagellate blooms in Tunisian lagoonscauses and consequences for aquaculturerdquo inHarmful Algae BReguera J Blanco M L Fernandez and T Wyatt Eds Xuntade Galicia and Intergovernmental Oceanographic Commissionof UNESCO Vigo Spain 1998
[57] Z Armi S Turki E Trabelsi and N Ben Maiz ldquoFirst recordedproliferation of Coolia monotis (Meunier 1919) in the NorthLake of Tunis (Tunisia) correlation with environmental factorsrdquoEnvironmental Monitoring and Assessment vol 164 no 1ndash4 pp423ndash433 2010
[58] I Nakajima Y Oshima and T Yasumoto ldquoToxicity of benthicDinoflagellates in Okinawardquo Bulletin of the Japanese Society Forthe Science of Fish vol 47 pp 1029ndash1033 1981
[59] S M Nascimento D A Purdie and S Morris ldquoMorphologytoxin composition and pigment content of Prorocentrum limastrains isolated from a coastal lagoon in southern UKrdquo Toxiconvol 45 no 5 pp 633ndash649 2005
[60] J C Marr A E Jackson and J L McLachlan ldquoOccurrenceof Prorocentrum lima a DSP toxin-producing species from theAtlantic coast of Canadardquo Journal of Applied Phycology vol 4no 1 pp 17ndash24 1992
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology



![MANGROVES. SEAGRASSES AND CORALS [A simple layman-type field guide]](https://static.fdokumen.com/doc/165x107/6321610a0c12e1161503c4a8/mangroves-seagrasses-and-corals-a-simple-layman-type-field-guide.jpg)














