
Growth plasticity in Cymodocea nodosa stands: the importance of nutrient supply
16
Aquatic Botany, 47 (1994) 249-264 249 0304-3770/94/$07.00 © 1994 - Elsevier Science B.V. All rights reserved Growth plasticity in Cymodocea nodosa stands: the importance of nutrient supply Marta Perez *,a, Carlos M. Duarte b, Javier Romero a, Kaj Sand-Jensen c, Teresa Alcoverro a "Departament d'Ecoiogta, Universitatde Barcelona, Diagonal 645, 08028 Barcelona, Spain bCentre d'Estudis Avaneats de Blanes, Camino de Santa B,~rbara, 17300 Blanes, Girona, Spain CFreshwaterBiologicalLaboratory, Universityof Copenhagen, 51Helsingorsgade, 3400 Hillerod, Denmark (Accepted 27 October 1993 ) Abstract We examine here the growth plasticity (as the variability in leaf growth and production, and shoot mortality rate) and stand structure (rhizome, root and shoot biomasses and nutrient contents, and shoot age structure), of the Mediterranean seagrass Cymodocea nodosa (Ucria) Aschers. growing across an existing gradient of nutrient supply in shallow water (from 2/JM soluble reactive phospho- rus at nutrient-rich, to 0.08/JM at nutrient-poor sites). Phosphorus concentrations in plant tissues increased strongly (three-fold in leaves, and ten-fold in rhizomes and roots) from nutrient-poor to nutrient-rich sites. Leaf phosphorus concentrations at nutrient-poor sites were only half of those re- quired for balanced seagrass growth, indicating the importance of phosphorus in limiting C. nodosa growth at the nutrient-poor sites. Phosphorus-limited plants allocated a greater proportion of the lim- iting resource (32% of the P standing stock, compared with about 10% at nutrient-sufficient sites), and a greater proportion of the produced biomass (about 40e of the biomass, compared with about 15% of the biomass at nutrient-sufficient sites), to root development, thereby increasing their capac- ity to acquire phosphorus from the sediment at the expense of impaired shoot growth and size. Re- sources were relocated as nutrient shortage ameliorates towards nutrient-rich sites, yielding a reduc- tion in the relative allocation to below-ground biomass (which represented about 80% and 50% of the biomass at nutrient-poor and nutrient-rich sites, respectively). Shoot growth at the nutrient-rich site exceeded that at sites receiving intermediate and low nutrient loadings by two-fold and three-fold, respectively, yet the biomass of plants growing under high nutrient supply was almost half of that of the plants receiving intermediate nutrient loading. This was due to reduced shoot density at the nu- trient-rich site, resulting from higher shoot mortality at higher nutrient concentrations, which suggests a shift from phosphorus as the limiting resource towards limitation by light or oxygen supply to the roots. Despite their lower biomass, the fast growth of plants at the nutrient-rich site allowed them to support similar leaf production to plants receiving intermediate nutrient supply, and twice the phos- phorus incorporation rate of those plants. The observed differences in population dynamics across the gradient in nutrient supply demonstrate the power of comparative analyses of seagrass stands as an additional approach to examine the importance of nutrient supply in controlling seagrass growth and productivity. *Corresponding author. SSDI 0304-3770 (93) 00363-D
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Growth plasticity in Cymodocea nodosa stands: the importance of nutrient supply

Aquatic Botany, 47 (1994) 249-264 249 0304-3770/94/$07.00 © 1994 - Elsevier Science B.V. All rights reserved
Growth plasticity in Cymodocea nodosa stands: the importance of nutrient supply
M a r t a P e r e z *,a, C a r l o s M . D u a r t e b, J a v i e r R o m e r o a, K a j S a n d - J e n s e n c, T e r e s a A l c o v e r r o a
"Departament d'Ecoiogta, Universitat de Barcelona, Diagonal 645, 08028 Barcelona, Spain bCentre d'Estudis Avaneats de Blanes, Camino de Santa B,~rbara, 17300 Blanes, Girona, Spain
CFreshwater Biological Laboratory, University of Copenhagen, 51Helsingorsgade, 3400 Hillerod, Denmark
(Accepted 27 October 1993 )
Abstract
We examine here the growth plasticity (as the variability in leaf growth and production, and shoot mortality rate) and stand structure (rhizome, root and shoot biomasses and nutrient contents, and shoot age structure), of the Mediterranean seagrass Cymodocea nodosa (Ucria) Aschers. growing across an existing gradient of nutrient supply in shallow water (from 2/JM soluble reactive phospho- rus at nutrient-rich, to 0.08/JM at nutrient-poor sites). Phosphorus concentrations in plant tissues increased strongly (three-fold in leaves, and ten-fold in rhizomes and roots) from nutrient-poor to nutrient-rich sites. Leaf phosphorus concentrations at nutrient-poor sites were only half of those re- quired for balanced seagrass growth, indicating the importance of phosphorus in limiting C. nodosa growth at the nutrient-poor sites. Phosphorus-limited plants allocated a greater proportion of the lim- iting resource (32% of the P standing stock, compared with about 10% at nutrient-sufficient sites), and a greater proportion of the produced biomass (about 40e of the biomass, compared with about 15% of the biomass at nutrient-sufficient sites), to root development, thereby increasing their capac- ity to acquire phosphorus from the sediment at the expense of impaired shoot growth and size. Re- sources were relocated as nutrient shortage ameliorates towards nutrient-rich sites, yielding a reduc- tion in the relative allocation to below-ground biomass (which represented about 80% and 50% of the biomass at nutrient-poor and nutrient-rich sites, respectively). Shoot growth at the nutrient-rich site exceeded that at sites receiving intermediate and low nutrient loadings by two-fold and three-fold, respectively, yet the biomass of plants growing under high nutrient supply was almost half of that of the plants receiving intermediate nutrient loading. This was due to reduced shoot density at the nu- trient-rich site, resulting from higher shoot mortality at higher nutrient concentrations, which suggests a shift from phosphorus as the limiting resource towards limitation by light or oxygen supply to the roots. Despite their lower biomass, the fast growth of plants at the nutrient-rich site allowed them to support similar leaf production to plants receiving intermediate nutrient supply, and twice the phos- phorus incorporation rate of those plants. The observed differences in population dynamics across the gradient in nutrient supply demonstrate the power of comparative analyses of seagrass stands as an additional approach to examine the importance of nutrient supply in controlling seagrass growth and productivity.
*Corresponding author.
SSDI 0 3 0 4 - 3 7 7 0 ( 9 3 ) 0 0 3 6 3 - D

250
Introduction
M. Perez et al. /Aquatic Botany 47 (1994) 249-264
The Mediterranean seagrass Cymodocea nodosa (Ucria) Aschers. is char- acterised by an extreme size plasticity. Mature leaves range in length from a few centimetres to well over halfa metre between populations (C.M. Duarte, unpublished data, 1992). This size plasticity is often displayed across rela- tively short distances where genetic isolation is unlikely, suggesting that the observed plasticity is largely phenotypic. A phenotypic origin of seagrass size plasticity is supported by the finding that the size of the related seagrass Zos- tera marina L. is dependent on sediment fertility (Ostenfeld, 1908; Phillips, 1972). Reduced leaf size is characteristic of plants that allocate carbon to root growth to reduce nutrient shortage (e.g. Chapin, 1980; Ingestad and/~,gren, 1991 ). Thus, differences in nutrient supply may be an important source of differences in leaf size between different populations of individual seagrass species (e.g. Short, 1983). This hypothesis is supported by experimental nu- trient enrichment of seagrass beds, which generally results in increases in leaf and shoot size as well as in plant biomass and productivity (Orth, 19717; Har- lin and Thorne-Miller, 1981; Short, 1987; Powell et al., 1989; Short et al., 1990; Perez et al., 1991 ). Also, recent evidence shows that differences in leaf phosphorus content, which reflect differences in phosphorus availability for
ii .....
• I • I i I
k , ', I ~ I .
",i" ~ ~'-., ~ ~ ~ / ' " "=~" ~"" " " ::"" 9__Kin
Fig. 1. Study area, showing the location of the studied C. nodosa populations (H, high nutrient supply; M, medium nutrient supply; L, low nutrient supply) and the channels that drain nu- trient-rich water from the rice paddy fields into Alfacs Bay.

M. Perez et al. ~Aquatic Botany 47 (1994) 249-264 2 51
plant growth, account for about 40% of the differences in Thalassia testu- dinum Banks ex KSnig biomass in Florida Bay (Fourqurean et al., 1992 ).
We have recently shown (Perez et al., 1991 ) that summer growth of the seagrass C. nodosa, the dominant primary producer in Alfacs Bay (Ebro Delta, Spain), is strongly phosphorus limited, similar to seagrasses growing in other carbonate-rich sediments (Short, 1987; Short et al., 1990). Phosphorus ad- ditions to sediments supporting C. nodosa stands resulted in increased leaf turnover, shoot size and above-ground biomass and shoot density (Perez et al., 1991 ). Thus, it is likely that the considerable variability in C. nodosa stands in Alfacs Bay (e.g. a 100-fold range in biomass and cover (Perez and Camp, 1986) ) is attributable to differences in the nutrient supply to seagrass stands across the bay. In the present study, we demonstrate the large variability in growth and stand structure possible in neighbouring stands of the Mediterra- nean seagrass C. nodosa, and provide evidence that differences in nutrient supply are responsible for this variability.
Study area
Alfacs Bay is an estuarine embayment in the north-eastern coast of Spain (Fig. 1 ). The bay (56 km 2) is shallow (mean depth 3.2 m, maximum depth 6 m), has a short water residence time of about 10 days (Camp and Delgado, 1987), and is closed to the Mediterranean Sea by a long, narrow sand bar (Fig. 1 ). Nutrient inputs to the bay, annually 518 tons N and 47 tons P (J. Camp, unpublished results, 1992), are dominated by inputs from channels collecting the runoff from neighbouring rice paddy fields (Pratet al., 1988),
Table 1 Summary of environmental differences between the high-nutrient northern shore and the low-nu- trient southern shore of Aifacs Bay (data from Delsado and Camp (1987), Vidal ( 1991 ), Palacin et al. (1991)and Mallo et al. (1993))
Variable Northern shore Southern shore ( high nutrient load ) (low nutrient load)
Open water Dissolved inorganic nitrogen (/aM) 100 Soluble reactive phosphorus (#M) 2
3 0.08
Sediments (top 2 cm) Pore-water dissolved inorganic
nitrogen (#M) 400 200 Pore-water soluble reactive
phosphorus (/aM) 30 4 Redox potential (mY) - 150 + 160 Organic content (% DW) 8 1 Density (gcm -3) 0.5 1.2
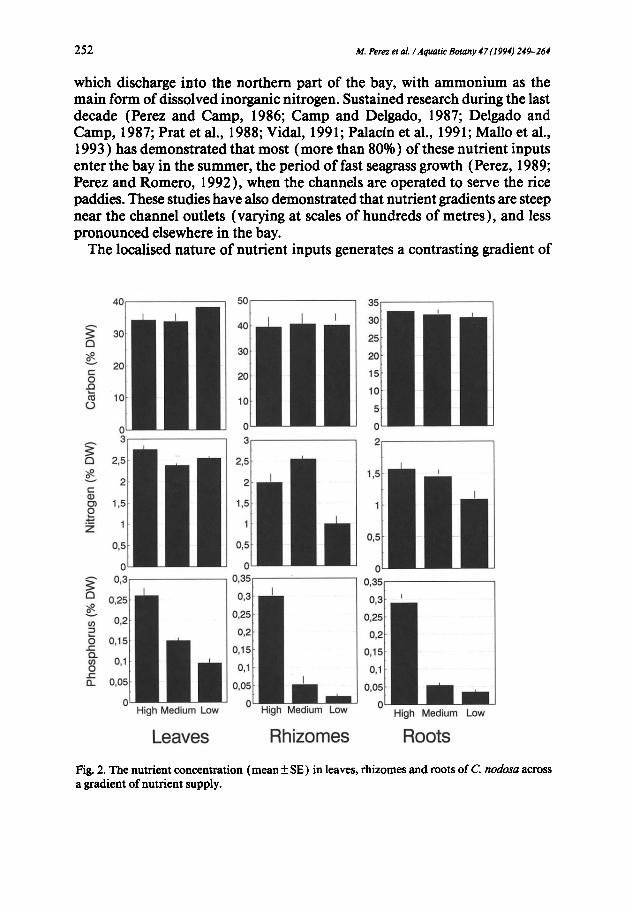
2 5 2 M. Perez et aL / Aquatic Botany 47 (1994) 249- 264
which discharge into the northern part of the bay, with ammonium as the main form of dissolved inorganic nitrogen. Sustained research during the last decade (Ferez and Camp, 1986; Camp and Delgado, 1987; Delgado and Camp, 1987; Prat et al., 1988; Vidal, 1991; Palacin et al., 1991; Mailo et al., 1993 ) has demonstrated that most (more than 80%) of these nutrient inputs enter the bay in the summer, the period of fast seagrass growth (Perez, 1989; Ferez and Romero, 1992), when the channels are operated to serve the rice paddies. These studies have also demonstrated that nutrient gradients are steep near the channel outlets (varying at scales of hundreds of metres), and less pronounced elsewhere in the bay.
The localised nature of nutrient inputs generates a contrasting gradient of
401 5 "
a~" 30
20 t - O
10 0
t-' O)
o
Z
09 2 0
t'~
0 t -
fit=
4
7=
1
3
2,5
2
1,5
1
0,5
0,35
0,3
0,25
0,2
0,15
0,1
0,05
2
1,5
1
0,5
High Medium Low High Medium Low High Medium Low
Leaves R h izo m es Roots
Fig. 2. The nutrient concentration (mean + SE) in leaves, rhizomes and roots of C nodosa across a gradient of nutrient supply.

M. Perez et al. / Aquatic Botany 47 (1994) 249-264 253
nutrient availability between the northern, nutrient-rich, and the southern, nutrient-poor, shores (Table 1 ), which is evident both in water and sedi- ments (Table 1 ). The turbid waters discharged by the channels also increase light attenuation at the northern shore compared with that at the southern shore (Perez, 1989). The strong gradient in nutrient availability within the bay is associated with major biological differences, such as reduced densities of benthic macro- and meiofauna (Palacin et al., 1991 ) and increased bacte- rial activity in the anoxic sediments (Table 1 ) of the nutrient-rich area (Mallo et al., 1993). Meadows of C. nodosa cover the shallow (less than 4 m) sedi- ments of the bay, showing order-of-magnitude differences in plant cover and biomass around the bay. Dense meadows grow along the nutrient-rich north- ern shore in contrast to the sparse patches of the southern shore (Perez and Camp, 1986; Duarte and Sand-Jensen, 1990a,b; Perez and Romero, 1992). Fertilisation experiments in the bay (Perez et al., 1991 ) have demonstrated that, similar to observations elsewhere (Fourqurean et al., 1992), nutrient, and particularly phosphorus, contents of the leaves can be used as indicators of relative nutrient availability to the plants.
Methods
To examine the variability in structure and dynamics of seagrass stands in the bay, we selected, on the basis of previous surveys of the bay (Perez and Camp, 1986), three contrasting shallow (about 1 m) C. nodosa stands along
Table 2 Evaluation of the extent (as proportions---nested ANOVA) of variability in nutrient concentration, plant biomass and growth of Cymodocea nodosa shoots along a gradient of nutrient supply, compared with the variability within stands; also shown are the F values and probability values for the ANOVA testing for differences among stands
Variable Between (%) Within (%) F (P)
Leaf phosphorus 96.6 3.4 45.8 ( <0.001 ) Leaf nitrogen 99.1 0.9 25.2 ( < 0.001 ) I.¢af carbon 93.4 6.6 13.0 ( < 0.005) Rhizome phosphorus 97.7 2.3 62.4 ( <0.001 ) Rhizome nitrogen 95.5 4.5 21.1 ( < 0.001 ) Rhizome carbon 5.9 94.1 0.06 (0.94) Root phosphorus 99.9 0.1 170 ( <0.001 ) Root nitrogen 90.1 9.9 9.1 ( <0.01 ) Root carbon 37.1 62.9 0.6 (0.57) Shoot biomass 96.5 3.5 29.3 ( < 0.001 ) Rhizome biomass 87.2 12.8 6.9 (0.003) Root biomass 93.2 6.8 13.6 ( < 0.001 ) Leaf appearance rate 91.2 8.8 7.8 ( < 0.005) Shoot length 97.2 2.8 34.7 ( <0.001 ) Shoot elongation 96.7 2.3 33.8 ( < 0.001 )
254 M. Perez et al. / Aquatic Botany 47 (1994) 249-264
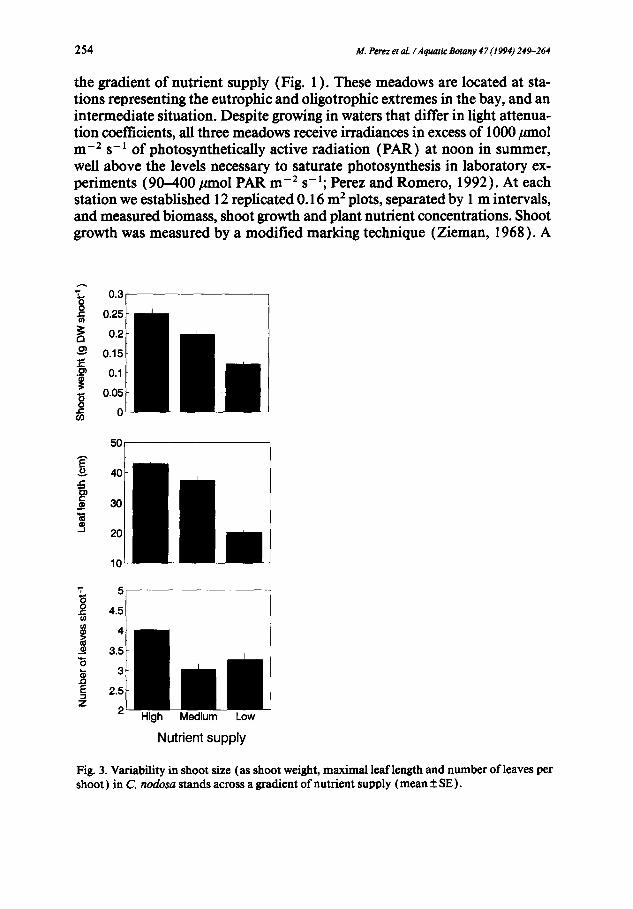
the gradient of nutrient supply (Fig. l ). These meadows are located at sta- tions representing the eutrophic and oligotrophic extremes in the bay, and an intermediate situation. Despite growing in waters that differ in light attenua- tion coefficients, all three meadows receive irradiances in excess of 1000/xmol m -2 s-1 of photosynthetically active radiation (PAR) at noon in summer, well above the levels necessary to saturate photosynthesis in laboratory ex- periments (90-400 #mol PAR m -2 s-l; Perez and Romcro, 1992). At each station we established 12 replicated 0.16 m 2 plots, separated by 1 m intervals, and measured biomass, shoot growth and plant nutrient concentrations. Shoot growth was measured by a modified marking technique (Zieman, 1968). A
0 e- ¢0
I=
e- ¢0 v
5O
o~ 40 =.
l~ 30
20
10
4 .
3. "6 == E 2. Z
High Medium Low
Nutrient supply
Fig. 3. Variability in shoot size (as shoot weight, maximal leaf length and number of leaves per shoot) in C. nodosa stands across a gradient of nutrient supply (mean_ SE).
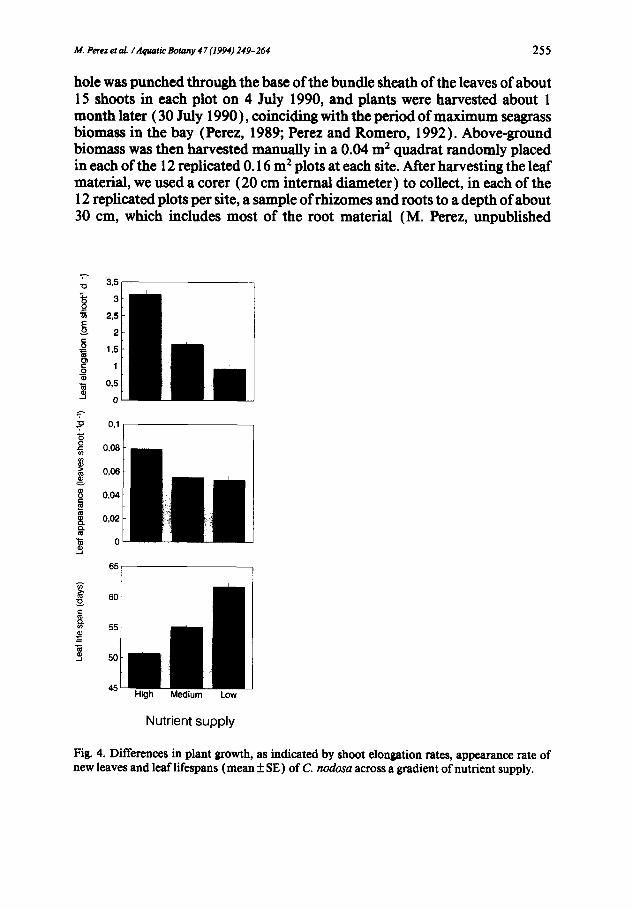
114. Perez et al. ~Aquatic Botany 47 (1994) 249-264 255
hole was punched through the base of the bundle sheath of the leaves of about 15 shoots in each plot on 4 July 1990, and plants were harvested about 1 month later (30 July 1990), coinciding with the period of maximum seagrass biomass in the bay (Perez, 1989; Perez and Romero, 1992). Above-ground biomass was then harvested manually in a 0.04 m 2 quadrat randomly placed in each of the 12 replicated 0.16 m 2 plots at each site. After harvesting the leaf material, we used a corer (20 cm internal diameter) to collect, in each of the 12 replicated plots per site, a sample of rhizomes and roots to a depth of about 30 cm, which includes most of the root material (M. Perez, unpublished
• o 3 , 5
2,s E
8 1,5
o ,~ 0,5
=' o
"o 0,1 f
0,08
,~ o,o6
0,04
~. 0,02
'~ 0 =,
65
60
~ ss .-=
~ 5 0
45 High Medium Low
Nut r ien t s u p p l y
Fig. 4. Differences in plant growth, as indicated by shoot elongation rates, appearance rate of new leaves and leaf lifespans (mean _ SE) of C. nodosa across a gradient of nutrient supply.
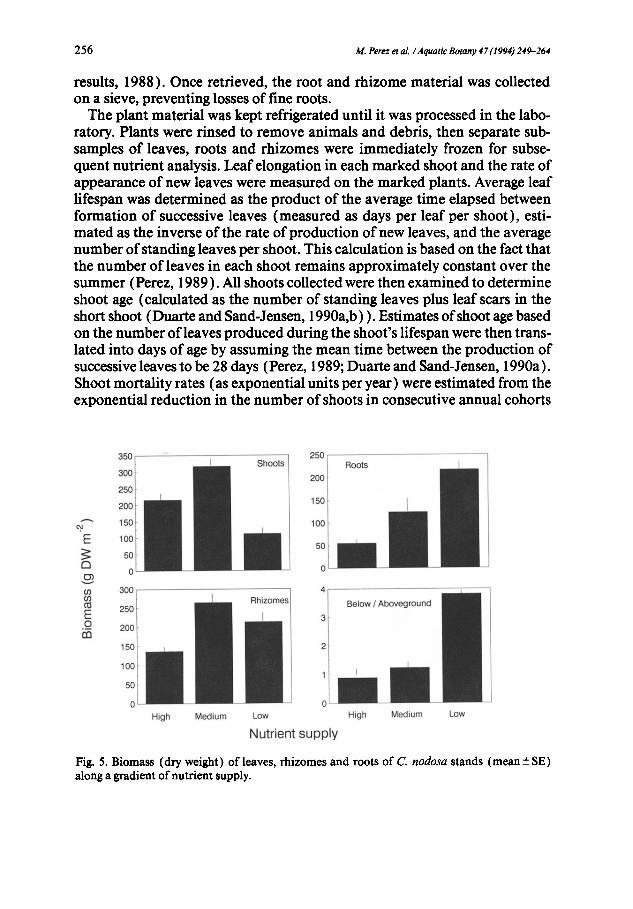
2 5 6 M. Perez et al. / Aquatic Botany 47 (1994) 249-264
results, 1988). Once retrieved, the root and rhizome material was collected on a sieve, preventing losses of fine roots.
The plant material was kept refrigerated until it was processed in the labo- ratory. Plants were rinsed to remove animals and debris, then separate sub- samples of leaves, roots and rhizomes were immediately frozen for subse- quent nutrient analysis. Leaf elongation in each marked shoot and the rate of appearance of new leaves were measured on the marked plants. Average leaf lifespan was determined as the product of the average time elapsed between formation of successive leaves (measured as days per leaf per shoot), esti- mated as the inverse of the rate of production of new leaves, and the average number of standing leaves per shoot. This calculation is based on the fact that the number of leaves in each shoot remains approximately constant over the summer (Perez, 1989 ). All shoots collected were then examined to determine shoot age (calculated as the number of standing leaves plus leaf scars in the short shoot (Duarte and Sand-Jensen, 1990a,b ) ). Estimates of shoot age based on the number of leaves produced during the shoot's lifespan were then trans- lated into days of age by assuming the mean time between the production of successive leaves to be 28 days (Perez, 1989; Duarte and Sand-Jensen, 1990a). Shoot mortality rates (as exponential units per year) were estimated from the exponential reduction in the number of shoots in consecutive annual cohorts
E
£3 E~
v
o~
E O °_
133
350
300
250
200
150
100
50
0
300 I
250
200
150
100
50
0
250
200
150
100
50
0
4 I
3
2
1
0 High Medium Low High Medium
Nutrient supply Low
Fig. 5. Biomass (dry weight) of leaves, rhizomes and roots of C. nodosa stands (mean+SE) along a gradient of nutrient supply.
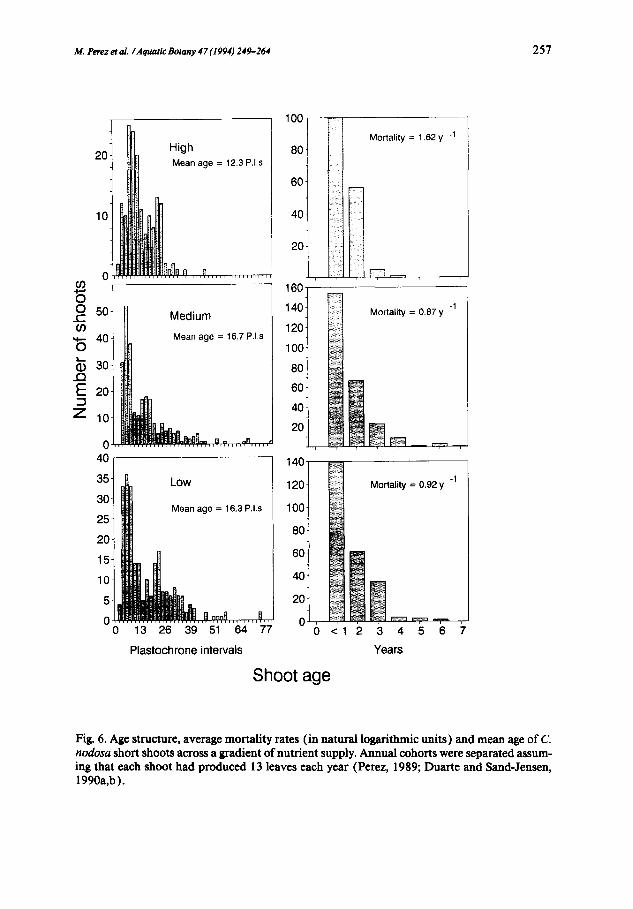
M. Perez et al. /Aquatic Botany 47 (1994) 249-264 257
O O t -
O t ,
(D .£3
E Z
20 H i g h
Mean age = 12.3 P.I.s
50 M e d i u m
4 0 - Mean age = 16.7 P.Ls
3 0 -
2 0 -
10
0 L ~ . n L . . . .
40 -r 1
35 -
3 0
2 5
2 0
1 5
1 0
5
ol 0
L o w
Mean age = 16.3 P.I.s
100 I
8ol J
60~
,° t 20
1 160~
1401
1201
1001 80~
°° t 40
20
] 40
120
100
80
60
40
20~
0
Mortality = 1.62 y -1
-1 Mortality = 0.87 y
-1 Mortal i ty = 0.92 y
< 1 2 3 4 5 6 ' :1':3' ' ' '26 ' ' ' 39 ' ' ' £)i' ' 6 4 ' ' :77 0
P l a s t o c h r o n e i n t e r va l s Years
Shoot age
Fig. 6. Age structure, average mortality rates (in natural logarithmic units) and mean age of C. nodosa short shoots across a gradient of nutrient supply. Annual cohorts were separated assum- ing that each shoot had produced 13 leaves each year (Perez, 1989; Duarte and Sand-Jensen, 1990a,b).
258 M. Perez et al. I Aquatic Botany 47 (1994) 249-264
(Duarte and Sand-Jensen, 1990b). Plant dry weights were determined after drying overnight at 65 °C.
Carbon and nitrogen concentrations in plant material (leaves, rhizomes and roots) were determined in duplicate subsamples using a Carlo-Erba (Milan, Italy) CHN analyser, and tissue P concentrations were determined colorimetdcally in duplicate subsamples after wet acid digestion (Koroleff, 1983 ). Nested analysis of variance was used to test for the importance and significance of differences between and within sites on aspects of the plant growth measured (Sokal and Rohlf, 1969).
300
E v
t- O
E v
t-- o')
Z
250
200
150
100
50
0
1
1.4
1.2 E O3 1
v
0.8 2 O 0.6 t'- t~ 0.4 O9 O t - 0.2
I:L 0
High Medium Low
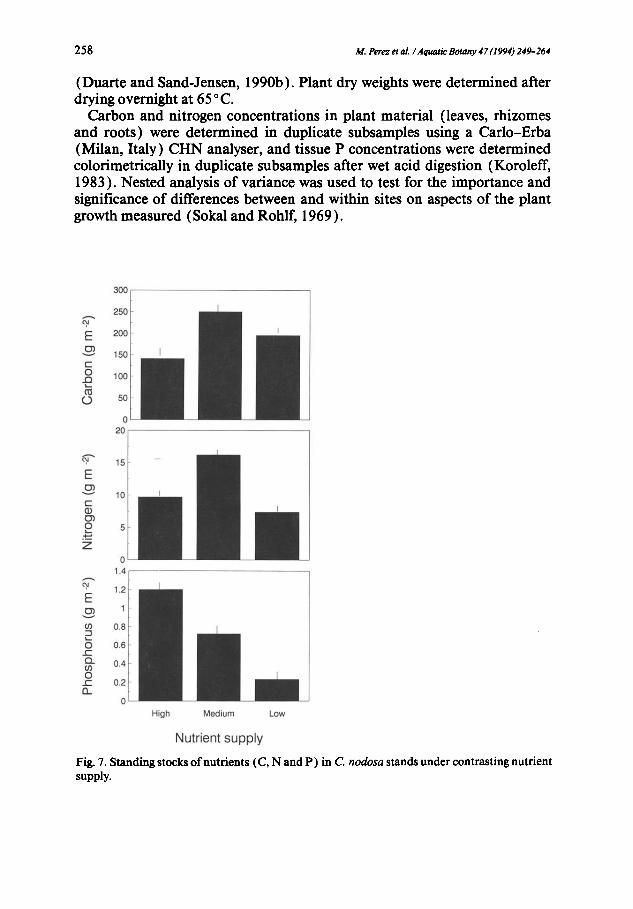
Nutrient supply Fig. 7. Standing stocks of nutrients (C, N and P ) in C. nodosa stands under contrasting nutrient supply.
M. Perez et al. / Aquatic Botany 47 (1994) 249-264
Results
259
Nutrient concentrations in plant tissues during mid-summer differed con- siderably between the three populations studied (Fig. 2 ), indicating the large differences in nutrient availability to the plants. Carbon concentrations var- ied only slightly between populations (Table 2) and between plant compo-
60
40
"5 r - 3 0 0
. 1
1:: 20 0 Q. 0 10
0
t - (9 O) 9 e-,
0 c 0
° ~
8 o_
2 o e"
r- Q.
C 0 ~ 2o 8. 9 o. 0
Shoots E e =
High Medium Low
Rhizomes Roots
3 e
3
2
2
1
1
High Medium Low High Medium Low
Nutrient supply
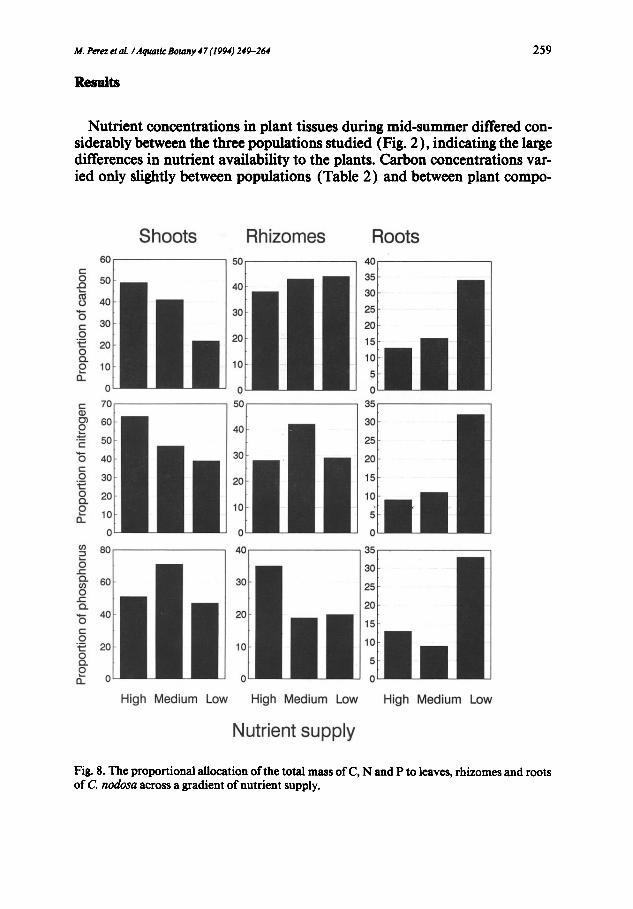
Fig. 8. T h e proportional allocation of the total mass o f C, N a n d P to leaves, rhizomes and roots o f C nodosa across a gradient of nutrient supply.

260 M. Perez et al. /Aquatic Botany 47 (1994) 249-264
nents (Fig. 2 ), but nitrogen concentrations were significantly higher in leaves than in roots (Fig. 2), and significantly (Table 2) lower in the below-ground material of the plants growing at the nutrient-poor site (Fig. 2 ). The concen- tration of phosphorus in all seagrass tissues (leaves, rhizomes and roots) de- creased (Fig. 2) significantly (Table 2) with nutrient supply, to reach values well below those believed to be indicative of phosphorus-limited seagrass growth (about 0.2% P; Duarte, 1990), thereby providing supporting evi- dence of the importance of phosphorus as the primary nutrient limiting sum- mer growth ofC. nodosa in Alfacs Bay (Perez et al., 1991 ).
The large differences in plant nutrient concentrations across the gradient of nutrient supply correlated with similarly broad differences in shoot char- acteristics and growth in the populations. Shoot size decreased as nutrient availability decreased (Fig. 3 ), and shoots growing under low nutrient supply also had fewer leaves than those at high nutrient supply (Fig. 3 ). Leaf growth also decreased, as reflected in significant (Table 2) reductions in leaf elon- gation rates and in the rate of appearance of new leaves, with decreasing nu- trient availability (Fig. 4 ). The lifespan of leaves, however, showed a signifi- cant increase (Fig. 4) as nutrient limitation increased.
The increased growth and size of C. nodosa shoots as nutrient availability increased (Figs. 3-4) were, however, not conducive to increased plant bio- mass, which was significantly (Table 2) smaller (Fig. 5 ) in the stand receiv- ing high nutrient supply, compared with that exposed to intermediate nu- trient levels. The stand receiving the lowest nutrient supply developed the lowest shoot biomass, intermediate rhizome biomass and the highest root biomass of all three populations (Fig. 5). This pattern resulted in a three-fold increase in the ratio of below-ground to above-ground biomass as nutrient limitation became more severe (Fig. 5). The low shoot biomass developed by nutrient-sufficient plants (Fig. 5) reflects their reduced shoot density (849 shoots m - 2 ) compared with that in the stand receiving intermediate nutrient supply (1594 shoots m -2), resulting from the greater mortality rate experi- enced by the plants receiving high nutrient supply (Fig. 6 ). These logarithmic mortality rate estimates (Fig. 6) allowed calculation of the half-life of the shoot population, which was found to be only half a year (0.43 years) in the population receiving the highest nutrient supply, whereas population half-lives
Table 3 Calculated C. nodosa leaf production, and nutrient incorporation per unit area at the three sites studied
Nutrient loading Leaf production N incorporation P incorporation (g C m-2 day -I ) (rag N m-2 day - I ) (rag P m-2 day - l )
High 1.18 125 12.8 Medium 1.37 107 7.7 Low 0.39 26.9 1.1

M. Perez et al. / Aquatic Botan~, 47 (1994) 249.-264 261
were twice as long in the other stands (0.80 years and 0.75 years at the inter- mediate and nutrient-poor sites, respectively). The much shorter population half-life at the nutrient-rich site implies that it should maintain a shoot (and rhizome) turnover rate twice as fast as that of the other populations to sup- port a stable population (Gallegos et al., 1993).
Comparison of the standing stocks of carbon, nitrogen and phosphorus contained in the three C. nodosa stands showed that the plants at the nutrient- rich site had the highest mass of the limiting nutrient (i.e. phosphorus) per unit area (Fig. 7). The standing stocks of nitrogen and carbon in the plants were, however, highest with the intermediate nutrient supply (Fig. 7). The proportion of nutrients allocated to roots was highest in nutrient-stressed plants, whereas shoots of plants sufficiently supplied with nutrients contained a greater proportion of total nitrogen and carbon than those of nutrient- stressed plants (Fig. 8 ).
The data on leaf growth, plant density and nutrient concentration allowed calculation of leaf production and nutrient incorporation per unit area. These calculations indicated that, despite their lower biomass, the fast growth of the plants at the nutrient-rich site allowed them to support primary production rates similar to those at the site receiving intermediate nutrient loadings, which were three-fold higher than that calculated for the nutrient-poor site (Table 3). In addition, nutrient incorporation rates were highest at the nutrient-rich site, particularly for phosphorus incorporation, which was about twice and 12-fold greater there than at the sites receiving intermediate and low nutrient loading, respectively (Table 3).
Discussion
The data presented demonstrate the remarkable plasticity possible, in growth and size, of neighbouring C. nodosa stands located across a gradient of nutrient supply. These data also support our previous finding that phos- phorus is the primary nutrient limiting summer growth of C. nodosa in Alfaes Bay (Perez et al., 1991 ). The large variability in phosphorus concentrations in plant tissues observed along the nutrient supply gradient (more than ten- fold variation in the roots), is similar to the range for P tissue concentrations reported for various seagrasses (Duarte, 1990). This stresses the ability of C. nodosa to grow across a wide range of nutrient concentrations in sediments, which apparently results, in part, from its capacity to change the allocation of carbon and limiting nutrients to different compartments. Phosphorus-limited plants allocated a greater proportion of the limiting resource, as well as a greater proportion of the produced biomass, to root development, thereby increasing the capacity of roots to acquire phosphorus from the sediment. This, however, is achieved at the expense of impaired shoot growth and size. As nutrient supply ameliorates, resources are relocated to promote leaf growth

262 M. Perez et al. / Aquatic Botany 47 (1994) 249-264
and increase shoot size, yielding a shift from a dominance of below-ground to that of above-ground biomass. This capability has also been observed for ter- restrial plants (e.g. Chapin, 1980; Ingestad and ~gren, 1991 ) and seagrasses elsewhere (Short, 1983; Powell et al., 1989).
Our results demonstrate that increased shoot growth as nutrient supply in- creases may not result in a similar increase in biomass, because of parallel reductions in shoot density resulting from a decreased shoot lifespan at higher nutrient concentrations. Why shoots may show reduced lifespans at higher nutrient concentrations is unknown, but similar reductions in plant survival with increasing nutrient concentration have also been observed for terrestrial plants (e.g. Parrish and Bazzaz, 1982 ). Moreover, eutrophication often leads to massive declines in biomass of seagrasses (Orth and Moore, 1983; Cam- bridge and McComb, 1984; Larkum and West, 1990; Giesen et al., 1990), suggesting a general coupling between excess nutrient enrichment and sea- grass mortality. Maintenance of a stable meadow under the high mortality rate observed at the nutrient-rich site must involve increased shoot recruit- ment, which requires, in turn, increased rhizome growth (Duarte and Sand- Jensen, 1990b). This is consistent with the greater P concentrations and pro- portional P allocation to rhizomes of plants growing in nutrient-rich sedi- ments relative to those observed for nutrient-limited plants.
As shoots become larger with high nutrient supply, growth regulation should shift from nutrient towards light limitation, as a result of self-shading (Perez and Romero, 1992) and shading by fast-growing macroalgae (e.g. Ulva sp. in Alfacs Bay; Martinez-Arroyo, 1990), which are abundant in nutrient-rich areas. In addition, anoxic sediments in the eutrophic shore (Vidal, 1991; Mallo et al., 1993) may affect root respiration and nutrient uptake (Pregnall et al., 1984) and limit seagrass growth. Hence, seagrass responses to eutrophication compound both direct effects of increased nutrient supply and indirect ef- fects. Indirect effects involve, for instance, reduced underwater light penetra- tion and increased shading by epiphytes and fast-growing macroalgae (Orth and Moore, 1983; Cambridge and McComb, 1984; Larkum and West, 1990; Giesen et al., 1990), and altered root metabolism in anoxic sediments (Preg- nallet al., 1984). Thus, it is likely that both direct nutrient effects and indi- rect effects, derived from shading by fast-growing macroalgae (Martinez-Ar- royo, 1990) and sediment anoxia (Vidal, 1991; Mallo et al., 1993), are responsible for the changes in C. nodosa growth described here.
In summary, our results demonstrate the great plasticity in the allocation of nutrients and biomass, shoot size and growth, and population dynamics of C. nodosa in response to sustained differences in nutrient availability. The data provide evidence of population-level responses to enhanced nutrient supply (e.g. reduced shoot longevity and modified age structure) which are difficult to detect in short-term (less than 1 year) nutrient enrichment exper- iments. However, our correlational approach is, unlike experimental nutrient

M. Perez et al. /Aquatic Botany 47 (1994) 249-264 263
additions, unable to separate the direct effects of increased nutrient supply from the indirect effects, involving reduced fight availability and oxygen sup- ply to roots, of increased nutrient supply. These results, along with the exper- imental evidence reported earlier (Perez et al., 1991 ), add to recent compar- ative analyses of seagrass stands along gradients of nutrient supply (Fourqurean et al., 1992), to demonstrate the power of combining correla- tional and experimental approaches to elucidate the importance of nutrient supply for seagrass growth and productivity.
Acknowledgements
This work was funded by CICYT grant (CE-89-0017) to J. Romero, K. Sand-Jensen was supported by a grant from the Danish National Science Research Council. We thank J. Camp, M. CanyeUas, N. Marb6 and M.A. Ma- teo for assistance in the field, and Q. Garrabou for assistance in the labora- tory.
References
Cambridge, M.L. and McComb, A.J., 1984. The loss of seagrasses in Cockburn Sound, Western Australia. I. The time course and magnitude of seagrass decline in relation to industrial de- velopment. Aquat. Bot., 20: 229-243.
Camp, J. and Delsado, M., 1987. Hidrografla de las bahias del Delta (]el Ebro. Invest. Pesq., 51: 351-369.
Chapin III, F.S., 1980. The mineral nutrition of wild plants. Annn. Rev. Ecol. Syst., 11: 233- 260.
Delgado, M. and Camp, J., 1987. Abundancia y distribuci6n de nutrientes inorgimicos disueltos en las bahias del Delta del Ebro. Invest. Pesq., 51:517-548.
Duarte, C.M., 1990. Seagrass nutrient content. Mar. Ecol. Prog. Ser., 67: 201-207. Duarte, C.M. and Sand-Jensen, K., 1990a. Seagrass colonization: patch formation and patch
growth in C. nodosa patches. Mar. Ecol. Prog. Ser., 65: 193-200. Duarte, C.M. and Sand-Jensen, K., 1990b. Seagrass colonization: biomass development and
shoot demogi'aphy in C. nodosa patches. Mar. Ecol. Pro8~ Ser., 67: 97-103. Fourqurean, J.W., Zieman, J.C. and Powel, G.V.N., 1992. Phosphorus limitation of primary
production in Florida Bay: evidence from C:N:P ratios of the dominant seagrass Thalassia testudinum. Limnol. Oceanogr., 37: 162-171.
Gallegos, M.E., Merino, M., Marb~t, N. and Duarte, C.M., 1993. Biomass and dynamics of Thal- assia testudinum in the Mexican Caribbean: elucidating rhizome growth. Mar. Ecol. Prog. Ser., 95: 185-192.
Giesen, W.B.J.T., van Katwijk, M.M. and den Hartog, C., 1990. Eelgrass condition and turbid- ity in the Dutch Wadden Sea. Aquat. Bot., 37:71-85.
Harlin, M.M. and Thorne-Miller, B., 1981. Nutrient enrichment of seagrass beds in a Rhode Island coastal lagoon. Mar. Biol., 65: 221-229.
Ingestad, T. and ]~gren, G.I., 1991. The influence of plant nutrition on biomass allocation. Ecol. Appl., 1: 168-174.

264 M. Perez et al. /Aquatic Botany 47 (1994) 249-264
Koroleff, F. 1983. Determination of phosphorus. In: K. Grasshoff, M. Ehrardt and K. Kremling (Editors), Methods of Seawater Analysis, 2nd edn. Verlag-Chemie, Niirnberg, pp. 125-139.
Larkum, A.W.D. and West, R.J., 1990. Long-term changes of seagrass meadows in Botany Bay, Australia. Aquat. Bot., 37: 55-70.
Mallo, S., Vallespin6s, F., Ferrer, S. and Vaqu~, D., 1993. Microbial activities in estuarine sed- iments (Ebro Delta, Spain) influenced by organic matter influx. Sci. Mar., 57:31-40.
Martinez-Arroyo, A., 1990. Estudio ecol6gico de las algas efemeroflceas: papel en los flujos de materia y energta en un sistema estuMico (Bahia de los Alfaques, Delta del Ebro, Espafia). Ph.D. Thesis, University of Barcelona, 166 pp.
Orth, R.J., 1977. Effects of nutrient enrichment on growth of the eelgrass Zostera marina in the Chesapeake Bay, Virginia, USA. Mar. Biol., 44: 187-194.
Orth, R.J. and Moore, K.A., 1983. Chesapeake Bay: an unprecedented decline in submerged aquatic vegetation. Science, 222:51-53.
Ostenfeld, C.H., 1908. On the ecology and distribution of the grass wrack (Zostera marina) in Danish waters. Rep. Dan. Biol. Stn., 16: 1-62.
Palacin, C., Martin, D. and Gili, J.M., 199 I. Features of spatial distribution of benthic infauna in a Mediterranean shallow-water bay. Mar. Biol., 110: 315-321.
Parrish, J.A.D. and Bazzaz, B.A., 1982. Responses of plants from three successional communi- ties to a nutrient gradient. J. Ecol., 70: 233-248.
Pcrez, M., 1989. Fandrogamas marinas en sistemas estuaricos: produccion, factores limitantes y algunos aspectos del ciclo de nutrientes. Ph.D. Thesis, University of Barcelona, Barcelona.
Perez, M. and Camp, J., 1986. Distribuci6n espacial y biomasa de las faner6gamas marinas de las bahias del Delta del Ebro. Invest. Pesq., 50:519-530.
Pcrez, M. and Romero, J., 1992. Photosynthetic response to light and temperature of the sea- grass Cymodocea nodosa and the prediction of its seasonality. Aquat. Bot., 43:51-62.
Perez, M., Romero, J., Duarte, C.M. and Sand-Jensen, K., 1991. Phosphorus limitation of Cy- modocea nodosa growth. Mar. Biol., 109:129-133.
Phillips, R.C., 1972. Ecological life history of Zostera marina L. (eelgrass) in Puget Sound, Washington. Ph.D. Thesis, University of Washington, Seattle, WA, 154 pp.
Powell, G.V.N., Kenworthy, W.J. and Fourqurean, J.W., 1989. Experimental evidence for nu- trient limitation of seagrass growth in a tropical estuary with restricted circulation. Bull. Mar. Sci., 44: 324-340.
Prat, N., Mufioz, I., Camp, J., Comin, F.A., Lucena, J.R., Romero, J. and Vidal, M., 1988. Seasonal changes in particulate organic carbon and nitrogen in the river and drainage chan- nels of the Ebro Delta (N.E. Spain). Verh. Int. Verein. Limnol., 23:1344-1349.
Pregnall, A.M., Smith, R.D. and Alberto, R.S., 1984. Metabolic adaptation of Zostera marina (eelgrass) to diurnal periods of root anoxia. Mar. Biol., 83: 141-147.
Short, F.T., 1983. The seagrass, Zostera marina L.: plant morphology and bed structure in re- lation to sediment amonium in Izembek Lagoon, Alaska. Aquat. Bot., 16: 149-161.
Short, F.T., 1987. Effects of sediment nutrients on seagrasses: literature review and mesocosm experiment. Aquat. Bot., 27: 41-57.
Short, F.T., Dennison, W.C. and Capone, D.G., 1990. Phosphorus-limited growth of the tropi- cal seagrass Syringodium filiforme in carbonate sediments. Mar. Ecol. Prog. Ser., 62:169- 174.
Sokal, R.R. and Rohlf, F.J., 1969. Biometria. H. Blume, Madrid, 830 pp. Vidal, M., 1991. Estudios sobre la din~lrnica de nutrientes en sistemas estu~ricos: flujos sedi-
mento--agua de nutrientes y oxigeno. Ph.D. Dissertation, University of Barcelona, 305 pp. Zieman, J.C., 1968. A study of the growth and decomposition of the sea grass, Thalassia testu-
dinum. M.Sc. Thesis, University of Miami.




















