
A Book is a Negotiated Order
9
JOURNAL OF ULTRASTRUCTURE AND MOLECULAR STRUCTURE RESEARCH 101, 243-251 (1988) Etude ultrastructurale du spermatozoide du coelacanthe: Latimeria chalumnae XAVIER MATTEI, YVES SIAU, ET BERNARD SERET* DCpartement de Biologie Animale, FacultC des Sciences, Universitt Cheikh Anta Diop, Dakar, SCnt?gal, et *Antenne ORSTOM, Laboratoire d’lchtyologie GLn&ale et Appliqute, MusCum National d’Histoire Naturelle, 43, rue Cuvier, 75231 Paris, France Received October 25, 1988 The study of the ultrastructure of the Latimeria chalumnae spermatozoon was carried out using the C3 coelacanth fished on September 24, 1953 and preserved in 10% formahn at the Museum National d’Histoire Naturelle in Paris. The spermatozoon has a tapered acrosome 2.5 pm long. The nucleus is 25 pm in length. Its posterior region, which is thinner for about 5 pm, is surrounded by a mitochondrial sheath. The nucleus is lengthwise crossed by a canal which contains three rods which extend to the anterior tip of the acrosome. The two centrioles are located at the base of the nucleus and arranged perpendicular to each other. The flagellum contains a 9 + 2 pattern axoneme and two longitudinal elements opposed to doublets 3 and 8. The ultrastructure of the Latimeria spermatozoon does not show a clear relationship between this fish and another group of fishes or tetrapods. 0 1988 Academic Press, Inc. L’ultrastructure du spermatozoi’de est connue dans tous les groupes de poissons actuels: Clasmobranches (Mattei, 1970), ho- locephales (Stanley, 1983), dipneustes (Boisson et al., 1967; Jespersen, 1971) bra- chiopterygiens (Mattei, 1970), chon- drosteens (Ginsburg, 1977), holosteens (Af- zelius, 1978) et teleosteens (Billard, 1970; Mattei, 1970, etc.) Q l’exception des crossopterygiens dont les spermatozoi’des n’ont CtC Ctudies qu’en microscopic photo- nique (Tuzet et Millot, 1959). Nous don- nons ici une description ultrastructurale du spermatozdide de Latimeria chalumnae. MATERIEL ET METHODES Une etude classique en microscopic Blectronique a transmission impose de nombreux imp&at&, l’animal a etudier doit en particulier &tre fraichement p&he, et a maturitC sexuelle. Devant les difIicultes de nous pro- curer un coelacanthe presentant ces conditions, nous avons utilise l’exemplaire C3 conserve dans le Labo- ratoire d’Anatomie Comparee du Museum National d’Histoire Naturelle de Paris (Millet et al., 1972). Cet animal, p&he le 24 septembre 1953 dans l’archipel des Comores, a tte tixt dans du form01 quelques heures apres sa capture. Apres plus de 34 ans de sejour dans le formol, des fragments de testicule et des fragments de rein ont etb places dans du tampon cacodylate de sodium 0,l M, CaCl, 0,2 m, saccharose 0,l m a pH 7,2 durant deux jours. Les prelevements ont ensuite ete places durant 12 heures dans du ghrtaraldehyde a 2,5% puis 1 heure dans du tetroxyde d’osmium a 1% dans le mtme tampon. Les prtlbvements ont ensuite 6tC d&hydrates par l’lthanol et l’oxyde de propylene avant d’etre inclus dans I’tpon. Les coupes ont tte reahsbes a I’ultramicrotome Porter Blum MTl, puis contrasttes par I’acbtate d’uranyle et le citrate de plomb. Les observations ont tte faites aux micro- scopes tlectronique Siemens Elmiskop 101 et JEOL 100 cx II. Pour la microscopic tlectronique a balayage, nous avons prdleve des fragments de testicule que nous avons dilaceres et places darts de l’eau distillee pen- dant 12 heures. Cette suspension a ensuite et6 soumise aux ultrasons pendant 3 minutes. Apres deshydrata- tion par l’ethanol, les prelevements ont et6 places sur une membrane millipore 0,45 urn, trait& au point cri- tique puis m&&is& et observes au microscope elec- tronique JEOL JSM 35 CF. La mise en evidence des mitochondries en micro- scopie photonique a et6 realide sur des frottis de sper- matozo’ides trait& selon la technique de Bensley (Mar- toja et Martoja-Pierson, 1%7), par une laque cuprique d’hematoxyline apres post-chromisation du materiel formold. OBSERVATIONS Dans le testicule, les spermatozoi’des sont rassembles en faisceaux ou libres dans les tubes seminiferes. 11s sont de forme al- 243 0889-1605/88 $3.00 Copyright 0 1988 by Academic Press, Inc. All rights of reproduction in any form reserved.
Transcript of A Book is a Negotiated Order

JOURNAL OF ULTRASTRUCTURE AND MOLECULAR STRUCTURE RESEARCH 101, 243-251 (1988)
Etude ultrastructurale du spermatozoide du coelacanthe: Latimeria chalumnae
XAVIER MATTEI, YVES SIAU, ET BERNARD SERET* DCpartement de Biologie Animale, FacultC des Sciences, Universitt Cheikh Anta Diop, Dakar, SCnt?gal, et
*Antenne ORSTOM, Laboratoire d’lchtyologie GLn&ale et Appliqute, MusCum National d’Histoire Naturelle, 43, rue Cuvier, 75231 Paris, France
Received October 25, 1988
The study of the ultrastructure of the Latimeria chalumnae spermatozoon was carried out using the C3 coelacanth fished on September 24, 1953 and preserved in 10% formahn at the Museum National d’Histoire Naturelle in Paris. The spermatozoon has a tapered acrosome 2.5 pm long. The nucleus is 25 pm in length. Its posterior region, which is thinner for about 5 pm, is surrounded by a mitochondrial sheath. The nucleus is lengthwise crossed by a canal which contains three rods which extend to the anterior tip of the acrosome. The two centrioles are located at the base of the nucleus and arranged perpendicular to each other. The flagellum contains a 9 + 2 pattern axoneme and two longitudinal elements opposed to doublets 3 and 8. The ultrastructure of the Latimeria spermatozoon does not show a clear relationship between this fish and another group of fishes or tetrapods. 0 1988 Academic Press, Inc.
L’ultrastructure du spermatozoi’de est connue dans tous les groupes de poissons actuels: Clasmobranches (Mattei, 1970), ho- locephales (Stanley, 1983), dipneustes (Boisson et al., 1967; Jespersen, 1971) bra- chiopterygiens (Mattei, 1970), chon- drosteens (Ginsburg, 1977), holosteens (Af- zelius, 1978) et teleosteens (Billard, 1970; Mattei, 1970, etc.) Q l’exception des crossopterygiens dont les spermatozoi’des n’ont CtC Ctudies qu’en microscopic photo- nique (Tuzet et Millot, 1959). Nous don- nons ici une description ultrastructurale du spermatozdide de Latimeria chalumnae.
MATERIEL ET METHODES
Une etude classique en microscopic Blectronique a transmission impose de nombreux imp&at&, l’animal a etudier doit en particulier &tre fraichement p&he, et a maturitC sexuelle. Devant les difIicultes de nous pro- curer un coelacanthe presentant ces conditions, nous avons utilise l’exemplaire C3 conserve dans le Labo- ratoire d’Anatomie Comparee du Museum National d’Histoire Naturelle de Paris (Millet et al., 1972). Cet animal, p&he le 24 septembre 1953 dans l’archipel des Comores, a tte tixt dans du form01 quelques heures apres sa capture. Apres plus de 34 ans de sejour dans le formol, des fragments de testicule et des fragments de rein ont etb places dans du tampon cacodylate de
sodium 0,l M, CaCl, 0,2 m, saccharose 0,l m a pH 7,2 durant deux jours. Les prelevements ont ensuite ete places durant 12 heures dans du ghrtaraldehyde a 2,5% puis 1 heure dans du tetroxyde d’osmium a 1% dans le mtme tampon. Les prtlbvements ont ensuite 6tC d&hydrates par l’lthanol et l’oxyde de propylene avant d’etre inclus dans I’tpon. Les coupes ont tte reahsbes a I’ultramicrotome Porter Blum MTl, puis contrasttes par I’acbtate d’uranyle et le citrate de plomb. Les observations ont tte faites aux micro- scopes tlectronique Siemens Elmiskop 101 et JEOL 100 cx II.
Pour la microscopic tlectronique a balayage, nous avons prdleve des fragments de testicule que nous avons dilaceres et places darts de l’eau distillee pen- dant 12 heures. Cette suspension a ensuite et6 soumise aux ultrasons pendant 3 minutes. Apres deshydrata- tion par l’ethanol, les prelevements ont et6 places sur une membrane millipore 0,45 urn, trait& au point cri- tique puis m&&is& et observes au microscope elec- tronique JEOL JSM 35 CF.
La mise en evidence des mitochondries en micro- scopie photonique a et6 realide sur des frottis de sper- matozo’ides trait& selon la technique de Bensley (Mar- toja et Martoja-Pierson, 1%7), par une laque cuprique d’hematoxyline apres post-chromisation du materiel formold.
OBSERVATIONS
Dans le testicule, les spermatozoi’des sont rassembles en faisceaux ou libres dans les tubes seminiferes. 11s sont de forme al-
243 0889-1605/88 $3.00 Copyright 0 1988 by Academic Press, Inc. All rights of reproduction in any form reserved.

244 MATTEL SlAU. ET SERET
longee (Figs. 1 et 2). 11s possedent un ac- rosome long d’environ 3 pm, dispose late- ralement a l’extremite anterieure du noyau et le prolongeant par une pointe effilee de 0,2 pm de diametre (Fig. 3). Cet acrosome est une formation creuse. 11 renferme 3 ba- guettes qui s’etendent jusqu’a son extremite anterieure (Figs. 4-7). Le noyau mesure environ 25 p,rn de long. Le contenu chromatique est dense et compact (Figs. l- 8), alors que dans la spermatide agee des lames de chromatine sont bien visibles (Fig. 9). L’extrCmitC anterieure du noyau est ef- filee en biseau et creusee d’une cavite cen- trale a la man&e d’une aiguille hypoder- mique (Fig. 3). Le canal intranucleaire s’etend sur toute la longueur du noyau mais ne perfore pas la base de celui-ci (Figs. I2- 14). Sa section est trilobee (Figs. 6-9). 11 renferme les 3 baguettes qui se prolongent dans l’acrosome. Ces baguettes ont un di- ametre de 50 nm et sont disposees de facon tres regulibre comme le montrent les coupes transversales de noyaux (Figs. 7 et 8). Nous avons trouve parmi les tres nom- breux spermatozoldes observes, deux noyaux anormaux: I’un dont le canal intra- nucleaire Ctait de section ovoi’de et qui ne renfermait que deux baguettes (Fig. 10) et l’autre possedant 3 canaux nucleaires et 4 baguettes (Fig. 11). Le diametre du noyau n’est pas constant (Figs. 1, 2, 24 et 25). II est de 0,4 p,rn a la base de l’acrosome, 0,6 pm dans la region moyenne et 0,3 pm dans la region posterieure sur une longueur de 5 pm environ. L’extremite posterieure du noyau montre un Clargissement de meme que le canal endonucleaire a ce niveau (Figs. 12-14, 21).
La region nucleaire posterieure qui
presente sur 5 p,rn in diametre reduit de moitit par rapport a la region moyenne est entouree d’un manchon cylindrique qui le plus generalement n’est pas totalement complet (Figs. 1, 2, 12, 21, 22, 25). Ce man- chon perinucleaire apparait de nature ho- mogene et semble limit6 par une double membrane sur certaines images (Fig. 21). 11 est absent dans certains spermatozojdes (Figs. 13, 24).
Les spermatozoi’des observes en micro- scopie photonique aprbs mise en evidence des mitochondries montrent un noyau mesurant 20 km de long et une piece inter- mtdiaire mitochondriale de 5 pm de long (Fig. 23).
Le cou renferme les deux centrioles et des formations paracentriolaires disposees entre le centriole proximal et le noyau ainsi qu’autour des deux centrioles (Figs. 12-14). Sa longueur est de l’ordre de 0,8 km. Le centriole proximal constitue de 9 triplets est dispose parallelement a la base du noyau (Fig. 14). Le centriole distal est perpendi- culaire au proximal. 11 est pourvu a son extremite distale de 9 rayons (Fig. 15). La region du cou se prolonge par un manchon membranaire dont la longueur varie de 0,6 a 1 km (Figs. 12, 13, 18, 21, 24, 25). Le man- chon membranaire est relic au centriole distal par l’intermediaire de ses rayons (Fig. 16); 9 granules denses sont visibles a ce niveau.
Le flagelle debute dans le manchon mem- branaire (Figs. 12, 17, 18). 11 est cylindrique sur une longueur de 0,l p,rn puis deux elk- ments lateraux s’associent a l’axonbme. A la sortie du manchon membranaire, le flagelle libre est constitue de l’axoneme et des 2 elements associes. Ces demiers ont
-..-I_ _l__l_.__
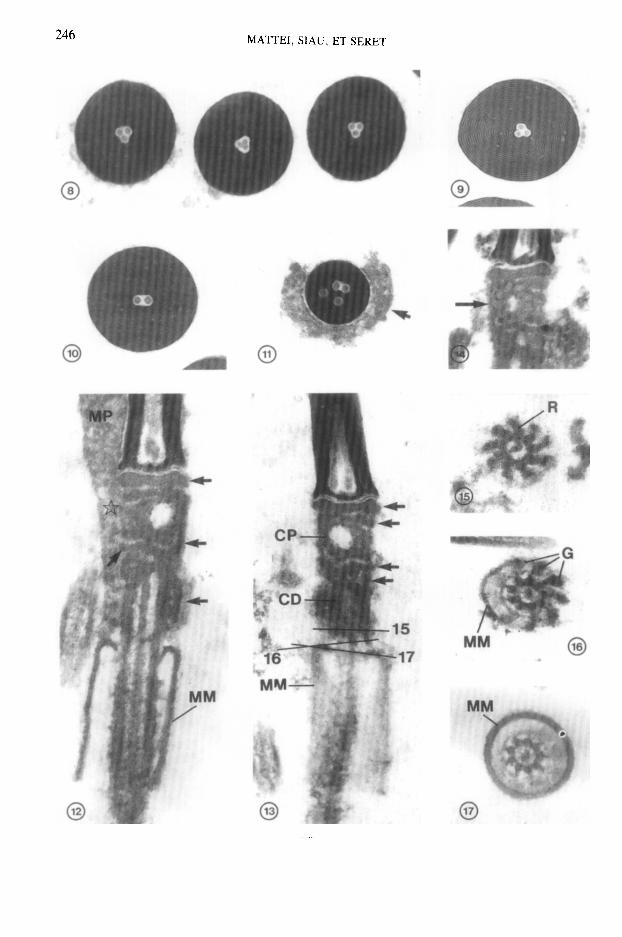
FIGS. I ET 2. Coupes longitudinales (L) et transversales (T) de spermatozoi’des. Le diametre du noyau dam se region posttrieure entourte du manchon pkinucleaire (MP) est reduit par rapport a celui de la region moyenne. A, acrosome; C, cou. La flkhe indique le manchon membranaire. Fig. 1, x 6000; Fig. 2, x 8000.
FIG. 3. Coupe longitudinale de la region nucleaire anterieure et de I’acrosome. A, acrosome; CI, canal intranucleaire; N, noyau. x 37 000.
FIGS. 4-7. Coupes transversales de la region anterieure du spermatozolde aux niveaux indiques sur la Fig. 3. CI, canal intranuclCaire contenant les trois baguettes; EA, extremitt anttrieure de I’acrosome; EN, extremitb anterieure du noyau. Fig. 4, x 43 000; Fig. 5, x 65 000: Fig. 6, x 50 000; Fig. 7. x 45 000.

LE SPERMATOZOjiDE DE Latimeria chalumnae 245
246 MATTEI, SAti, ET SEREl

LE SPERMATOZOIDE DE Latimeriu chalumnae 247
une section moins importante que celle qu’ils presentaient au niveau du manchon. 11s sont associes aux doublets no 3 et 8 de l’axonbme (Fig. 18). Nous avons observe une majorite de flagelles qui Ctaient depour- vus de ces elements lateraux (Figs. 19, 20). L’axoneme est de type classique 9 + 2. Les doublets peripheriques sont pourvus de deux bras et ils sont relies aux tubules cen- traux par des fibres rayonnantes.
Les spermatozoi’des que nous avons ob- serves dans les tubes renaux presentent la meme structure que ceux que nous avons d&its dans le testicle.
DISCUSSION
Dans une etude precedente (Mattei et Mattei, 1976), nous avions Ctudie en micro- scopie electronique a transmission les flagelles spermatiques d’un Poisson te- leosteen qui avait sejourne 4 ans dans du formol. I1 nous avait CtC possible de de- terminer que I’axonbme Ctait de type 9 + 0. L’animal que nous Ctudions ici a sejourne plus de 34 ans dans le form01 mais il a CtC fixe dans d’excellentes conditions. Les deux premiers coelacanthes pCchCs en 1938 et 1952 avaient CtC tres ma1 conserves et ne permettaient aucune etude histologique. Les scientifiques de l’epoque Ctaient impa- tients d’obtenir un animal en bon Ctat, c’est la raison pour laquelle ce troisieme coela- canthe (C3) a CtC particulierement bien fix& 11 est “le premier en bon &at et susceptible des plus fructueuses etudes” (Millot, 1954).
C’est a partir de cet animal que l’etude du spermatozdide en microscopic photonique a CtC realisee (Tuzet et Millot, 1959). Les images que nous donne la microscopic Clec- tronique, sans Ctre d’excellente qualite, sont suffisantes pour permettre une etude anatomique de ce spermatozoide. La mem- brane plasmique du gamete n’est pas con- servee a l’exception du manchon mem- branaire qui fait suite au cou. La persis- tance de la membrane plasmique dans cette region est peut Ctre due au fait qu’elle est d’une part reliee au centriole distal et d’autre part repliee sur elle meme pour con- stituer une structure plus resistante. L’absence de manchon perinucleaire dans certains spermatozdides est peutdtre due a sa fragilite.
Recemment, Leung (1988), Ctudie en mi- croscopie Clectronique des spermatozdides d’un Poisson teleosteen rare, fixes preala- blement au form01 avant d’etre postfix& dans du tetroxyde d’osmium a 1% dans du tampon cacodylate. Les resultats qu’il ob- tient sont de meilleure qualite que ceux que nous presentons ici, mais l’auteur ne precise pas si l’animal a sejourne longue- ment dans le formol.
L’etude precedente realisee en micro- scopie photonique (Tuzet et Millet, 1959) donnait comme dimensions du spermatozo- ‘ide dans le testicule les valeurs suivantes: acrosome, 2 pm; noyau, 20 pm; et piece intermediaire, 5 pm. Nous avons obtenu les memes valeurs pour les spermatozo’ides
FIG. 8. Coupe transversale de noyaux spermatiques. X 30 000. FIG. 9. Coupe transversale du noyau d’une spermatide trts &gee. La chromatine presente un aspect lamel-
laire. X 45 000. FIGS. 10 ET 11. Coupe transversale du noyau de deux spermatozo’ides anormaux. Le premier renferme deux
baguettes dam un canal intranucltaire reduit. Le second contient trois canaux intranucldaires et quatre ba- guettes. La fleche indique le manchon perinucleaire. X 56 000.
FIGS. 12 ET 13. Coupes longitudinales du spermatozoi’de passant au niveau du cou et du manchon mem- branaire (MM). Le noyau et le canal intranuckaire sont Clargis dans la region nucleaire posterieure. Le manchon perinucleaire (MP) se prolonge au niveau du cou (etoile) (Fig. 12), II est absent dans la Fig. 13. Des formations paracentriolaires (fleches) sont associees aux centrioles proximal (CP) et distal (CD). Fig. 12, x 57 000: Fig. 13, x 5oooo.
FIG. 14. Le centriole proximal (fltche) est constitue de neuf triplets. X 50 000. FIGS. 15-17. Coupes transversales de la base du centriole distal et du flagelle aux niveaux indiques sur la Fig.
13. Des granules denses (Ci) sont associes aux rayons (R) du centriole distal et au manchon membranaire (MM). x 5oooo.

248 MATTEI. SIAU. ET SERET
0 18 . . ;
FIG. 18. Coupes transversales du spermatozoide passant au niveau du noyau (N), du manchon membranaire (MM) et de la base du flagelle libre (F). Les flkches indiquent les dkments latelraux associds aux doublets no 3 et 8 de I’axontme. x 22 000.
FIGS. 19 ET 20. Coupes transversales de flagelles. Fig. 19, x 40 000; Fig. 20, x 60 000.
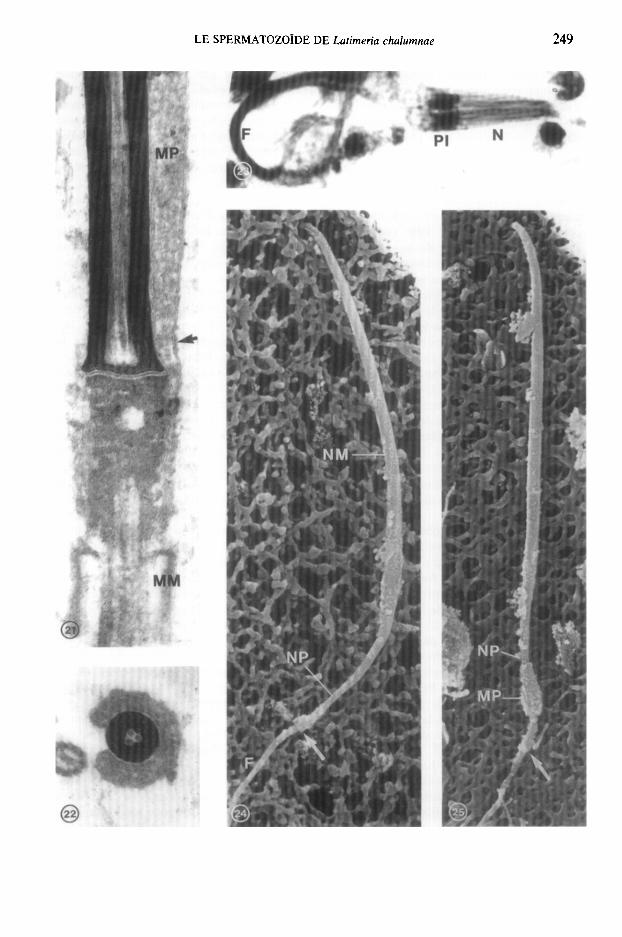
FIG. 21. Coupe longitudinale du spermatozoide au niveau du manchon p&inucltaire (MP) et du cou. Le manchon p&inuclCaire est limit6 par une double membrane (flkhe). MM, manchon membranaire. x 60 000.
FIG. 22. Coupe transversale du noyau et du manchon pCrinuclCaire. x 21 000. FIG. 23. Frottis de spermatozoides observks en microscopic photonique apr6s coloration des mitochondries.
F, flagelles; N, noyaux; PI, pike intermkdiaire. x 1250. FIG. 24. Spermatozoide observe en microscopic B balayage. Le rkgion nuclCaire posttrieure (NP) a un
diamktre rkduit par rapport & la rtgion nuclkaire moyenne (NM). Le manchon perinuclkaire est absent. La fleche indique le manchon membranaire. F, flagelle. x 8000.
FIG. 25. Spermatozoide observe en microscopic A balayage. Le manchon ptkinuclkaire (MY) entoure la region nucltaire posttrieure (NP). La flkche indique le manchon membranaire. F, Flagelle. x 8000.
LE SPERMATOZOPDE DE Latimeria chalumnae 249

250 MATTEI. SIAU. ET SERET
preleves dans le testicule et observes en mi- croscopie photonique apres coloration des mitochondries. Une difference apparait en- tre la longueur du noyau du spermatozoi’de observe en microscopic photonique (20 pm) et celle observee en microscopic elec- tronique (25 pm), pour des gametes preleves dans les testicules. Cette diffe- rence de 5 pm correspond a la longueur de la piece intermediaire mise en evidence en mi- croscopie photonique. Elle correspond Cgalement a la longueur du noyau dans sa partie posterieure Ctroite entouree du man- chon pkinucleaire tel que nous le montre la microscopic electronique. Nous en de- duisons que les mitochondries des spermato- zoi’des preleves dans les testicules consti- tuent le manchon perinucleaire et s’etendent jusqu’au niveau du cou. La piece intermediaire observee en micro- scopie photonique correspond en realite a la region posterieure du noyau entouree des mitochondries. Une telle disposition est ex- ceptionnelle et n’a CtC d&rite que chez un nemerte (Afzelius, 1971). Les spermatozo- ides que nous avons preleves dans les tubes renaux presentent la m&me structure que ceux qui proviennent des tubes semi- niferes.
Tuzet et Millot (1959) observent des sper- matozoi’des preleves dans les deux vessies oti ils se concentrent chez les males a ma- turite sexuelle (Millot et Anthony, 1973). 11s donnent les valeurs suivantes pour les longueurs du noyau et de la piece interme- diaire, respectivement 26 et 5 pm. Nous n’avons pas pu obtenir de spermatozoides provenant des vessies du coelacanthe C3, celles-ci ayant CtC ouvertes et videes lors des etudes anterieures. Peut-Ctre les sper- matozdides subissent-ils une transforma- tion dans les vessies et les mitochondries prennent-elles alors une disposition retro- nucltaire plus classique?
Le spermatozdide de Latimeria chalum- nae est de tyye Cvolue selon le definition de Franzen (1970). Le presence d’un noyau et d’une region mitochondriale bien deve- loppts est en accord avec le mode de fe-
condation de cet animal ovovivipare (Lavett-Smith et al., 1975).
Le canal intranucleaire a CtC d&-it dans les spermatozoi’des de plusieurs Vertebres: le petromizonide Lampetra planeri (Folle- nius, 1965), le brachiopterygien Polypterus senegalus (Mattei, 1970), le dipneuste Neo- ceratodus forsteri (Jespersen, 1971), le chondrosteen Acipenser stellatus (Gins- burg, 1977) et I’amphibien Pleurodeles waltlii (Picheral, 1967). Le second genre de dipneuste Ctudit, Protopterus, possede un spermatozolde depourvu de canal endonu- cleaire (Boisson et al., 1967). Le nombre de baguettes qu’il renferme est de un (Lampe- tru, Polypterus et Pleurodeles), deux (Neo- ceratodus) oti trois (Acipenser). Ces ba- guettes sont situdes uniquement dans le canal intranucleaire chez Lampetra, Polypterus et Acipenser alors qu’elles at- teignent l’extremite anterieure de l’acrosome chez Latimeria, Neoceratodus et Pleurodeles. La reaction acrosomienne est suivie d’une expulsion de ces baguettes a l’exterieur du noyau chez Lampetra (Jaana et Yamamoto, 1981) et Acipenser (Ginsburg, 1977) tandis que dans la cas de Pleurodeles (Picheral, 1977) elle conserve sa position intranucleaire. Leur devenir n’est pas connu au tours de la fecondation chez Latimeria pas plus d’ailleurs que chez Polypterus et Neoceratodus.
La presence de rayons associes au cen- triole distal et reliant ces derniers a la mem- brane plasmique est un caractere commun dans les spermatozoides d’invertebres (Af- zelius, 1979), mais peu frequent dans les gametes de vertebres. Elle a CtC signalee chez les dipneustes (Boisson et al., 1967; Jespersen, 1971).
Le flagelle spermatique de Latimeria rap- pelle celui des tlasmobranches (Boisson et al., 1%8; Stanley, 1971) et du dipneuste Neoceratodus (Jespersen, 1971). Dans ces differents cas deux elements lateraux sont associes a l’axoneme au niveau des dou- blets no 3 et 8. Nous n’avons pu determiner la longueur de ces elements, mais comme la majorite des coupes transversales de

LE SPERMATOZOfDE DE Latimeria chalumnae 251
flagelles en sont depourvues, ils sont prob- ablement peu developpes.
Plusieurs etudes ont demontre que I’anatomie du spermatozoi’de avait une valeur phylogenetique (Wingstrand, 1972; Mattei et Mattei, 1975; Afzelius, 1983; Bac- cetti, 1986; Mattei et Marchand, 1987). L’ultrastructure du gamete de Latimeria chulumnae ne demontre pas de parente e- vidente entre ce Poisson et un autre groupes de poissons ou de tetrapodes. La diversite spermatique des poissons est grande (Mattei, 1988) et l’ultrastructure du spermatozo’ide du coelacanthe augmente encore cette diversite.
Nous remercions Monsieur Daniel Robineau, La- boratoire d’Anatomie Comparee, Museum National d’Histoire Naturelle, Paris, qui nous a procure les fragments de testicule et de rein du coelacanthe C3 et qui nous a conseilles dans la realisation de ce travail.
REFERENCES
AFZELIUS, B. (1971). J. Submicrosc. Cytol. 3, 181- 192.
AFZELIUS, B. A. (1978). J. Ultrastruct. Res. 64, 309 314.
AFZELIUS, B. A. (1979) in FAWCETT, D. W. ET BED- FORD, J. M. (Eds.), The Spermatozoon, pp. 243- 25 1, Urban & Schwarzenberg, Baltimore.
AFZELIUS, B. A. (1983) in ANDRE, J. (Ed.), The Sperm Cell, pp. 385-394, Nijhoff, The Hague.
BACCETTI, B. (1986) Comp. Biochem. Physiol. 85,29- 36.
BILLARD, B. (1970) in BACCETTI, B. (Ed.), Compara- tive Spermatology, pp. 71-79, Academic Press, New York.
BOISSON, C., MATTEI, C., ET MATTEI, X. (1967) Bull. Inst. Fondam. Afr. Noire 29, 1097-1121.
BOISSON, C., MATTEI, X., ET MATTEI, C. (1968) Bull. Inst. Fondam. Afr. Noire 30, 659-673.
FOLLENIUS, E. (1965). J. Ultrastruct. Res. 13, 45% 468.
FRANZEN, A. (1970) In BACCETTI, B. (Ed.), Compar- ative Spermatology, pp. 29-46, Academic Press, New York.
GINSBURG, A. S. (1977) in BEWAEV, D. K. (Ed.), Problemy eksperimentalnoj biologii, pp. 246-256, Nauka, Moscow.
JAANA, H., ET YAMAMOTO, T. S. (1981) Japan. J. Ich- thyol. 28, 135-147.
JESPERSEN, A. (1971) J. Ultrastruct. Res. 37, 178-185. LAVETT-SMITH, C., RAND, C. S., ATZ, J. W., ET
SCHAEFFER, B. (1975) Science, 12, 1105-1106. LEUNG, L. K. P. (1988) Gamete Res. 19, 41-49. MARTOJA, R., ET MARTOJA-PIERSON, M. (1967) Initi-
ation aux techniques de l’histologie animale, Mas- son, Paris.
MATTEI, C., ET MATTEI, X. (1975) in AFZELIUS, B. A, (Ed.), Functional Anatomy of the Spermatozoon, pp. 211-221, Pergamon, Oxford.
MATTEI, X. (1970) in BACCETTI, B. (Ed.), Compara- tive Spermatology, pp. 57-70, Academic Press, New York.
MATTEI, X. (1988) Riot’. Cell 63, 151-158. MATTEI, X., ET MARCHAND, B. (1987) C. R. Acad.
Sci. Paris 305, 525-529. MATTEI, X., ET MATTEI, C. (1976) J. Microsc. Biol.
Cell. 2.5, 187-188. MILLOT, J. (1954) Le Naturaliste Malgache, ler supl.,
l-26. MILLOT, J., ET ANTHONY, J. (1973) C. R. Acad. Sci.
Paris 216, 2447-2448. MILLOT, J., ANTHONY, J., ET ROBINEAU, D. (1972)
Bull. Mus. Natl. Hist. Nat. Zool. 39, 533-548. PICHERAL, B. (1967) Arch. Biol. 78, 193-221. PICHERAL, B. (1977) J. Ultrastruct. Res. 60, 106-120. STANLEY, H. P. (1971) J. Ultrastruct. Res. 36, 103-
118. STANLEY, H. P. (1983) J. Ultrastruct. Res. 83, 184
194. TUZET, O., ET MILLOT, J. (1959) Ann. Sci. Nat. Zool.
1, 61-69. WINGSTRAND, K. G. (1972) Dan. Vidensk Selsk. 19,
l-72.




















