
83. Reilly N et al Am J Physiol Regul Integr Comp Physiol 2007 E-pub
42
Reilly et al - 1 PROBIOTICS POTENTIATE IL-6 PRODUCTION IN IL-1-TREATED CACO-2 CELLS THROUGH A HEAT SHOCK-DEPENDENT MECHANISM Natasha Reilly 1 , Vitaliy Poylin 1 , Michael Menconi 1 , Andrew Onderdonk 2 , Stig Bengmark 3 , and Per-Olof Hasselgren 1 Department of Surgery, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA 1 ; Department of Pathology, Brigham Women’s Hospital, Harvard Medical School, Boston, MA 2 ; Liver Institute, University College, London University, United Kingdom 3 Running title: Probiotics and enterocyte IL-6 production Supported in part by NIH grant R01 DK60546 (POH) Address for correspondence: Per-Olof Hasselgren, M.D., Department of Surgery, Beth Israel Deaconess Medical Center, 330 Brookline Avenue ST919, Boston, MA 02215; Tel (617) 667-1810; Fax (617) 667-1819; e-mail [email protected] Page 1 of 42 Articles in PresS. Am J Physiol Regul Integr Comp Physiol (July 18, 2007). doi:10.1152/ajpregu.00770.2006 Copyright © 2007 by the American Physiological Society.
Transcript of 83. Reilly N et al Am J Physiol Regul Integr Comp Physiol 2007 E-pub

Reilly et al - 1
PROBIOTICS POTENTIATE IL-6 PRODUCTION IN IL-1β-TREATED CACO-2
CELLS THROUGH A HEAT SHOCK-DEPENDENT MECHANISM
Natasha Reilly1, Vitaliy Poylin1, Michael Menconi1, Andrew Onderdonk2,
Stig Bengmark3, and Per-Olof Hasselgren1
Department of Surgery, Beth Israel Deaconess Medical Center, Harvard Medical School,
Boston, MA1; Department of Pathology, Brigham Women’s Hospital, Harvard Medical
School, Boston, MA2; Liver Institute, University College, London University, United
Kingdom3
Running title: Probiotics and enterocyte IL-6 production
Supported in part by NIH grant R01 DK60546 (POH)
Address for correspondence: Per-Olof Hasselgren, M.D., Department of Surgery, Beth
Israel Deaconess Medical Center, 330 Brookline Avenue ST919, Boston, MA 02215; Tel
(617) 667-1810; Fax (617) 667-1819; e-mail [email protected]
Page 1 of 42Articles in PresS. Am J Physiol Regul Integr Comp Physiol (July 18, 2007). doi:10.1152/ajpregu.00770.2006
Copyright © 2007 by the American Physiological Society.

Reilly et al - 2
ABSTRACT
Interleukin-6 (IL-6) may exert anti-inflammatory and protective effects in intestinal
mucosa and enterocytes. The influence of probiotics on mucosal and enterocyte IL-6
production is not known. We tested the hypothesis that the probiotic bacteria
Lactobacillus paracasei and Lactobacillus plantarum regulate IL-6 production in
intestinal epithelial cells. Cultured Caco-2 cells were treated with 1 ng/ml of IL-1β in the
absence or presence of different concentrations of Lactobacillus paracasei or
Lactobacillus plantarum followed by measurement of IL-6 production. The role of heat
shock response was examined by determining the expression of hsp70 and hsp27, by
down-regulating their expression with siRNA, or by treating cells with quercetin.
Treatment of the Caco-2 cells with IL-1β resulted in increased IL-6 production,
confirming previous reports from this laboratory. Probiotics alone did not influence IL-6
production but addition of probitoics to IL-1β-treated cells resulted in a substantial
augmentation of IL-6 production. Treatment of the Caco-2 cells with live Lactobacillus
paracasei increased cellular levels of hsp70 and hsp27 and the potentiating effect on IL-6
production was inhibited by quercetin and by hsp70 or hsp27 siRNA. Results suggest that
probiotics may enhance IL-6 production in enterocytes subjected to an inflammatory
stimulus and that this effect may, at least in part, be heat shock-dependent.
Page 2 of 42

Reilly et al - 3
INTRODUCTION
The intestinal mucosa is an active participant in the inflammatory response to injury,
sepsis, and endotoxemia and becomes the site of cytokine production during these
conditions (10,21,35). In previous studies, we have been particularly interested in
mucosal production of IL-6 and have found evidence that mucosal IL-6 levels are
increased during endotoxemia and sepsis in mice (23,48). Although multiple cell types
may contribute to increased IL-6 production in the mucosa during inflammation,
experiments in IL-1β-treated cultured intestinal epithelial cells (22,30) suggest that the
enterocyte is an important source of IL-6 during inflammation.
IL-6 is a pleiotropic cytokine that can have both pro- (28,32,46) and anti-
inflammatory properties (51). Previous studies provided evidence that IL-6 may exert
protective effects in various tissues during inflammation caused by injury and sepsis
(2,40). In recent experiments in our laboratory, treatment of cultured enterocytes with IL-
6 prevented cell death caused by hyperthermia (11), further supporting the concept that
IL-6 can have beneficial and protective effects. Interestingly, in previous experiments we
found that induction of the heat shock (stress) response potentiated the effect of IL-1β on
IL-6 production in cultured enterocytes (29,34). In other studies we found that induction
of the stress response in mice resulted in potentiated IL-6 production in intestinal mucosa
during sepsis and endotoxemia (47). Because these effects of the stress response were
associated with improved intestinal integrity (45), it is possible that increased mucosal
IL-6 levels, at least in part, are responsible for the beneficial effects of the heat shock
response during sepsis and endotoxemia.
Page 3 of 42

Reilly et al - 4
Multiple studies, both in humans and experimental animals, suggest that so called
probiotics exert protective effects in intestinal mucosa during various inflammatory
conditions (3,4,8,9,13-15). In addition, recent studies suggest that probiotics may reduce
the incidence of postoperative complications, in particular infectious complications, in
patients undergoing major surgical procedures (36). It should be noticed that although
multiple experimental studies support the concept that probiotics may be beneficial in the
treatment of patients with inflammatory diseases of the gut, additional controlled studies
are needed to more definitively identify the role of probiotics in the management of these
patients. Probiotics are living bacteria with low or no pathogenicity that exert beneficial
effects on the health of the host (13,15,38). Different mechanisms are probably involved
in the beneficial effects of probiotics, including inhibited mucosal and enterocyte
production of pro-inflammatory cytokines (18,19), inhibited activation of NF-kB (26,31),
and stimulated production of the antimicrobial peptide human beta defensin-2 in
intestinal epithelial cells (49). The influence of probiotics on enterocyte IL-6 production
is not known but considering our previous observation that IL-6 exerts protective effects
in the mucosa and enterocyte (11,45,47), the present experiments were performed to test
the hypothesis that probiotics may stimulate IL-6 production in intestinal epithelial cells.
Because, in a recent study, treatment of cultured enterocytes with probiotics resulted in
heat shock response (31), an effect that may be caused by a soluble factor released from
probiotics (43), we also examined the potential role of heat shock proteins for IL-6
production in cultured enterocytes treated with probiotics. Results suggest that probiotics
can potentiate the effect of IL-1β on IL-6 production in cultured enterocytes and that this
response, at least in part, is mediated by the heat shock response.
Page 4 of 42

Reilly et al - 5
MATERIAL AND METHODS
Cell cultures
Caco-2 cells, a human colon adenocarcinoma cell line that displays enterocyte-like
features in culture (33), were obtained from American Type Culture Collection
(Rockville, MD). Cells were grown at 37°C in 5% CO2 in Dulbecco’s modified Eagel’s
medium (DMEM; Gibco-BRC, Grand Island, NY) supplemented with 10% fetal bovine
serum, nonessential amino acids, 6 mM glutamine, 10 mM HEPES, 10 µg/ml
apotransferrin, 1 mM pyruvate, 24 mM NaHCO3, 100 U/ml penicillin, and 100 µg/ml
streptomycin. Cells, between passages 5 and 25, were seeded at a density of 100,000
cells/cm2 onto 6-well tissue culture plates. Cells were grown for 72 h to approximately
80-90% confluence before use. Cells were treated with 1 ng/ml of human recombinant
IL-1β (Biosource International, Camarillo, CA) or 1 ng/ml of human recombinant TNFα
(Biosource International) for 20 h and IL-6 and IL-8 production was determined by
measuring cytokine levels in the medium as described below. Untreated Caco-2 cells
served as controls. In order to examine the influence of probiotics on cytokine
production, live or heat-inactivated Lactobacillus paracasei subsp. paracasei F19 (L.
paracasei, Belgian Coordinated Collection of Microorganisms, BCCM, deposition
number LMG P-17806) or L. plantarum 2362 (BCCM deposition number LMG P-20606)
(both strains obtained from Medipharm Inc., Kagerod, Sweden and Des Moines, Iowa,
USA) were added to the culture medium at concentrations described below. In some
experiments, quercetin (Sigma, St Louis, MO) was added to the culture medium at a
concentration of 100 µM. Although most experiments were performed in 80-90%
confluent Caco-2 cells, in a control experiment we used Caco-2 cells that had been
Page 5 of 42

Reilly et al - 6
cultured for 3 weeks in transwell bicameral chambers to induce full differentiation as
described in detail previously (24).
Cell transfections
In order to examine the role of hsp70 and hsp27, the expression of these genes was
silenced by transfecting cells with appropriate siRNA’s. Caco-2 cells were transfected
with pre-designed siRNA against hsp70 (sc-29352) or hsp27 (sc-29350). Other cells were
transfected with non-specific (scrambled) siRNA. All siRNA duplexes and reagents were
from Santa Cruz Biotechnology (Santa Cruz, CA) and were used according the
manufacturer’s protocol.
Preparation of bacteria
The probiotic bacteria L. paracasei and L. plantarum were cultured in De Man-
Rogosa-Sharpe (MRS) broth under anaerobic conditions at 37°C. In other experiments, L.
jensenii (Gram-positive bacteria that are part of the normal vaginal flora) were cultured
under identical conditions and were used instead of L. paracasei and L. plantarum. After
culture, bacteria were collected by centrifugation (8,000 x g for 3 min), washed in
phosphate buffered saline (PBS), pH 7.4, and resuspended in DMEM cell culture
medium. Bacteria were added to the cultured Caco-2 cells at concentrations (determined
as colony forming units, cfu) described in Results either alone or in comination with IL-
1β. In some experiments, bacteria were heat-inactivated in a water bath at 80°C for 10
min or sonicated with four 20 sec bursts before they were added to the Caco-2 cells. The
heat-inactivation was confirmed by culture to make certain the preparations were sterile.
Page 6 of 42

Reilly et al - 7
In other experiments, the medium from cultured L. paracasei was passed through a 0.22-
µm filter (Millipore, Bedford, MA) and aliquots of the filtered medium were added to the
Caco-2 cells.
When the effects of Eschericia coli (E. coli) bacteria were tested, DH5α E. coli were
first cultured in Trypticase soy broth at 37°C, washed, and grown again at 37°C under
unaerobic conditions in MRS broth. The bacteria were then washed, resuspended in
DMEM cell culture medium and added to the Caco-2 cells.
Determination of IL-6 and IL-8
IL-6 and IL-8 levels were determined in cell culture medium by commercially
available ELISA kits (Endogen, Woburn, MA). The limit of detection as described by the
manufacturer was 1 pg/ml for both assays.
Western blotting
Western blotting was performed to determine cellular levels of hsp70 and hsp27.
After incubation, cells were lysed in 300 µl of lysis buffer (0.5 M Tris HCl, pH 7.4, 1.5
M NaCl, 10% NP-40, 10 mM EDTA, and 2.5% deoxycholic acid). Cell debris were
removed by centrifugation at 3,800 x g for 7 min. The supernatant (whole cell lysate) was
stored at -80°C until further analysis.
For Western blot analysis, aliquots of the cell lysates containing 50 µg of protein, as
determined by using the Bicinchoninic Acid (BCA) Protein Assay (Pierce, Rockford, IL),
were boiled in equal volumes of loading buffer (125 mM Tris-HCl, pH 6.8, 4% sodium
dodecyl sulfate, 20% glycerol, and 10% 2-mercaptoethanol) for 7 min and then separated
Page 7 of 42

Reilly et al - 8
by electrophoresis on a 10% Tris-glycine gradient gel (Novex, San Diego, CA). A protein
ladder (See-Blue Standard, Novex) was included as a molecular weight marker. The
proteins were transferred to nitrocellulose membranes (Xcell II Blot Module, Novex).
The membranes were blocked with 5% nonfat dried milk in Tris-buffered saline, pH 7.6,
containing 1% Tween-20 for 60 min followed by incubation with a 1:2000 dilution of
mouse anti-human hsp70 (Santa Cruz Biotechnology) or a 1:2000 dilution of rabbit anti-
human hsp27 antibody as primary antibodies. A goat anti-mouse IgG (Promega,
Madison, WI) or a goat anti-rabbit IgG (Santa Cruz Biotechnology) secondary antibody
was used at a dilution of 1:5000. The blots were then incubated in enhanced
chemiluminescence reagent (ECL, Amersham Life Sciences, Buckingham, UK) and
exposed on radiographic film (X-Omot AR, Eastman-Kodak, Rochester, NY).
Determination of mRNA levels
For determination of IL-6 mRNA levels, RNA was extracted and real-time PCR was
performed as described in detail previously (50).
Statistics
Experiments were performed at least three times to ensure reproducibility of results.
The results are reported as means ± SEM. Analysis of variance followed by Tukey’s test
was used for statistical comparisons. p<0.05 was considered statistically significant.
Page 8 of 42

Reilly et al - 9
RESULTS
In the present study, we examined the effects of two intestinal Lactobacillus strains
because recent reports suggest that several species of Lactobacillus are probiotics that
have protective immunomodulating properties (1,14,27), are able to prevent adhesion of
pathogenic bacteria to the intestinal wall (44), and can preserve mucosal integritiy (17).
In initial experiments, we determined IL-6 production in cultured Caco-2 cells treated
with IL-1β, live L. paracasei or live L. plantarum, either alone or in combinations.
Treatment of the Caco-2 cells with IL-1β (1.0 ng/ml) for 20 h resulted in increased IL-6
production (Fig 1A), confirming previous reports from our laboratory (29,30,34). In
separate experiments we found that cellular levels of IL-6 remained constant during
incubation for 20 h in the absence or presence of IL-1β (data not shown), supporting the
interpretation that the increased levels of IL-6 in the culture medium of IL-1β-treated
Caco-2 cells reflected increased production of IL-6 rather than increased transport of
cellular IL-6 into the medium. This was also supported by previous experiments in our
laboratory in which treatment of cultured Caco-2 cells with IL-1β resulted in increased
IL-6 mRNA levels in addition to increased amounts of the cytokine in the incubation
medium (29,30,34). Treatment of the Caco-2 cells with L. paracasei (108 bacteria/ml) or
L. plantarum (108 bacteria/ml) alone did not influence IL-6 production. In contrast, when
the bacteria were added to IL-1β-treated cells, the IL-6 production was substantially
increased, consistent with a synergistic (potentiating) effect of the probiotics (Fig 1A).
The potentiating effect of L. paracasei was approximately 6-fold (compared with the IL-
6 production in cells treated with IL-1β alone) and of L. plantarum approximately 2-fold.
Page 9 of 42

Reilly et al - 10
Because the effect on IL-6 production was most pronounced for L. paracasei, this
Lactobacillus strain was used in subsequent experiments.
Because previous reports suggest that some (but not all) effects of probiotics can be
exerted by both live and heat-inactivated bacteria (18,49), we next compared the effects
of live and heat-inactivated L. paracasei on IL-6 production in Caco-2 cells. As shown in
Fig 1B, the potentiating effect on IL-6 production in IL-1β-treated Caco-2 cells was
similar after treatment with live and heat-inactivated L. paracasei.
In order to test whether the potentiating effect of L. paracasei on IL-6 production
was caused by a product secreted from the bacteria, we next treated Caco-2 cells with
medium from cultured L. paracasei. Result from that experiment showed that medium
from cultured L. paracasei did not exert a potentiating effect on IL-6 production in IL-
1β-treated Caco-2 cells (Fig 2A). Taken together with the results in Fig 1, these
observations suggest that a factor present in live and heat-inactivated L. paracasei
potentiates IL-6 production in IL-1β-treated Caco-2 cells. In order to determine whether
this factor requires intact bacterial cell walls to be effective, we next compared the effect
of live intact L. paracasei with that caused by sonicated L. paracasei. Similar to intact L.
paracasei, sonicated L. paracasei potentiated the effect of IL-1β and the effect of
sonicated bacteria was of the same magnitude as that of intact bacteria (Fig 2B). Notably,
in this experiment, IL-6 production in Caco-2 cells treated with L. paracasei alone (third
bar in Fig 2B) was higher than in the preceding experiments (compare with Fig 1A and
B). This result probably reflected a day-to-day variability with regards to absolute IL-6
levels and illustrates the importance of including all study groups simultaneously when
experiments are performed. Importantly, although the basal levels of IL-6 were somewhat
Page 10 of 42

Reilly et al - 11
different, the potentiating effect of L. paracasei on IL-6 production was seen also in this
experiment.
In order to further characterize the influence of L. paracasei on IL-6 production in
the Caco-2 cells, we examined the effects of different concentrations of the bacteria
added to the enterocytes as well as the time course for the potentiating effects. When
different concentrations (107 – 109 bacteria/ml) of L. paracasei were added to IL-1β-
treated Caco-2 cells, the maximum effect on IL-6 production was noticed for 108
bacteria/ml (Fig 3A). None of the concentrations tested here influenced IL-6 production
when added alone to the Caco-2 cells. The potentiating effect of L. paracasei on IL-6
production in IL-1β-treated Caco-2 cells was noticed already after 3 h, at which time
point the IL-6 production was increased approximately 2-fold, and increased
progressively to be approximately 4-fold after 20 h (Fig 3B).
In order to test whether the potentiating effects of L. paracasei on IL-6 production in
IL-1β-treated Caco-2 cells noticed here are specific for probiotics, we next treated Caco-
2 cells with L. jensenii. These bacteria are Gram-positive (similar to L. paracasei and L.
plantarum) but non-probiotic bacteria that are commonly found in the normal vaginal
flora. When cultured Caco-2 cells were treated with L. jensenii (108 bacteria/ml), the IL-
1β-induced IL-6 production was potentiated to a similar degree as was noticed after
treatment of the Caco-2 cells with L. paracasei or L. plantarum. In this experiment, IL-6
production was 3.0 ± 0.8 pg/ml in IL-1β treated Caco-2 cells and 14.9 ± 1.2 pg/ml in
Caco-2 cells treated with IL-1β + L. jensenii. Treatment of the Caco-2 cells with L.
jensenii alone did not result in IL-6 production. Thus, not only probiotic Gram-positive
Page 11 of 42

Reilly et al - 12
bacteria but other Gram-positive bacteria as well may potentiate IL-6 production in IL-
1β-treated cultured enterocytes.
We next examined whether gram-negative bacteria can also potentiate the effects of
IL-1β on IL-6 production in cultured Caco-2 cells. This was done by treating cultured
Caco-2 cells with the E. coli strain DH5α. Although IL-6 production was somewhat
higher in Caco-2 cells treated with both E. coli and IL-1β than in Caco-2 cells treated
with IL-1β alone, this difference was much smaller than the approximately 6-fold
potentiating effect noticed in L. paracasei-treated cells in the same experiment (Fig 4A).
We next determined whether L. paracasei can influence IL-6 production in Caco-2 cells
stimulated with a cytokine other than IL-1β. Treatment of the Caco-2 cells with TNFα
did not result in measurable IL-6 production and this unresponsiveness to TNFα was not
influenced by L. paracasei (Fig 4B).
In order to test whether L. paracasei can potentiate the production of cytokines other
than IL-6, we examined IL-8 production in cultured Caco-2 cells. Similar to previous
reports (39), treatment of the Caco-2 cells with IL-1β resulted in a robust increase in IL-8
production (Fig 4C). L. paracasei alone or in combination with IL-1β did not influence
IL-8 production in the cultured Caco-2 cells.
Most of the experiments in the present study were performed in 80-90% confluent
Caco-2 cells, similar to previous reports from our laboratory in which the regulation of
IL-1β-induced IL-6 production was examined (29,30). In another study from our
laboratory, this model was validated by showing that IL-1β increased IL-6 production in
fully differentiated Caco-2 cells grown on filters in transwell bicameral chambers (24).
Here, we performed a control experiment using the same cell culture system as described
Page 12 of 42

Reilly et al - 13
in detail previously (24) to test whether L. paracasei can potentiate IL-1β-induced IL-6
production in fully differentiated Caco-2 cells. This also allowed us to treat the Caco-2
cells with IL-1β from the basal chamber and with L. paracasei in the apical chamber,
mimicking the situation in vivo when enterocytes are exposed to bacteria from the
intestinal lumen and to circulating cytokines from the blood stream. When this
experimental approach was used, IL-6 production (determined in the basal chamber) in
IL-1β-treated Caco-2 cells was potentiated by 57% by the addition of L. paracasei (108
bacteria/ml) to the apical chamber (IL-6 levels being non-detectable in control cells, 11.5
pg/ml after treatment with IL-1β alone for 20 h and 18.1 pg/ml after treatment with IL-1β
and L. paracasei; means from two individual experiments). In a separate experiment, L.
paracasei alone did not influence IL-6 production in fully differentiated Caco-2 cells.
Although the effect of L. paracasei on IL-6 production in IL-1β-treated fully
differentiated Caco-2 cells noticed here was less pronounced than in 80-90% confluent
Caco-2 cells, the results suggest that L. paracasei can influence the response to IL-1β
with regards to IL-6 production in fully differentiated Caco-2 cells as well.
The next set of experiments was designed to elucidate the mechanism of the
potentiating effect of L. paracasei on enterocyte IL-6 production. Because Petrof et al
(31) reported recently that treatment of cultured mouse colonocytes with probiotics
induced increased expression of the heat shock proteins hsp25 and hsp72 (corresponding
to human hsp27 and 70, respectively) and because we found previously that induction of
the stress response by hyperthermia or treatment with proteasome inhibitors potentiated
the IL-6 production in IL-1β-treated Caco-2 cells (29,34), we examined the role of the
heat shock response in the potentiated IL-6 production noticed here after treatment with
Page 13 of 42

Reilly et al - 14
L. paracasei. First, we tested whether treatment of the Caco-2 cells with L. paracasei
induced a heat shock response. As seen in Fig 5A and B, treatment of the Caco-2 cells
with live L. paracasei resulted in increased levels of both hsp70 and hsp25 with the most
pronounced effect seen for hsp70. Treatment of the cells with IL-1β did not result in
changes in the heat shock protein concentrations and the combined treatement with IL-1β
and L. paracasei did not influence the changes induced by L. paracasei alone.
If the heat shock response induced by L. paracasei participates in the regulation of
IL-6 production in IL-1β-treated Caco-2 cells, inhibition of the heat shock response
should prevent the potentiation of IL-6 production. When cells were treated with
quercetin, a substance known to inhibit the induction of the heat shock response (25), the
potentiating effect of L. paracasei on IL-6 production was abolished (Fig 5C). Although
this result lends strong support to the notion that the effect of live L. paracasei observed
in the present study is mediated by the heat shock response, one potential pitfall of the
experiment using quercetin is that the drug may have non-specific effects other than
inhibition of the heat shock response. In addition, it is not known from this experiment
which specific heat shock protein that is involved in the effect of L. paracasei.
To address these questions, we next silenced the hsp70 and hsp27 genes by
transfecting cells with siRNA’s against hsp70 and hsp25 mRNA. Treatment of Caco-2
cells with hsp70 siRNA resulted in a substantial reduction of hsp70 levels and prevented
the increase in hsp70 levels induced by live L.paracasei (Fig 6A). Measurement of IL-6
production in the same cells showed that the potentiating effect of L. paracasei was
substantially reduced after treatment with siRNA (from an approximately 3-fold increase
in IL-6 production to an approximately 50% increase over the IL-6 production caused by
Page 14 of 42

Reilly et al - 15
IL-1β alone) (Fig 6B). When the corresponding experiments were performed in Caco-2
cells treated with siRNA against hsp27 mRNA, the L. paracasei-induced increase in
hsp27 levels was blocked (Fig 7A) and the potentiating effect of L.paracasei on IL-6
production was abolished (Fig 7B). Note that in this experiment, IL-6 production in
untreated control cells was higher than in most of the other experiments in this study.
Although we do not have a definitive explanation for this finding, it is consistent with a
day-to-day variation of basal IL-6 production commonly seen in the current experimental
model. This is why it is important to include all treatment groups and control group
simultaneously in each expeiment as was done in the present study. Taken together, the
results in Fig 6 and 7 suggest that both hsp70 and hsp27 participate in the potentiating
effect of L.paracasei on IL-6 production in IL-1β-treated Caco-2 cells.
In order to further assess the role of the heat shock response in the potentiating effect
of L. paracasei on IL-6 production, two additional experiments were performed. Because
we found in initial experiments, that both heat-inactivated and sonicated L. paracasei
potentiated IL-6 production in IL-1β-treated Caco-2 cells (see Fig 1B and 2B), we tested
whether heat-inactivated and sonicated L. paracasei induced a heat shock response
(similar to live L. paracasei). Surprisingly, when cultured Caco-2 cells were treated for
20 h with heat-inactivated or sonicated L. paracasei under identical experimental
conditions as used for the experiments shown in Fig 1B and 2B, hsp27 and hsp70 levels
as determined by Western blotting followed by densitometry were not increased (data not
shown). In the same experiment, treatment of the Caco-2 cells with live L. paracasei
resulted in a 48% and 44% increase in hsp27 and hsp70 levels, respectively, confirming
the results shown in Fig 5A and 5B. These observations suggest that additional
Page 15 of 42

Reilly et al - 16
mechanisms (in addition to induction of the heat shock response) may be involved in the
potentiating effects of L. paracasei on IL-6 production in IL-1β-treated Caco-2 cells, at
least in the potentiating effects caused by heat-inactivated and sonicated L. paracasei.
In a second experiment designed to further assess the role of the heat shock response
in the potentiating effect of L. paracasei on IL-6 production, we compared the early time
courses for L. paracasei-induced heat shock response and L. paracasei-induced
potentiation of IL-6 expression in IL-1β-treated Caco-2 cells. In this experiment, IL-6
mRNA levels were determined by real-time PCR because we hypothesized that IL-6
mRNA levels would be influenced by IL-1β and L. paracasei at early time points.
Treatment of the Caco-2 cells with IL-1β alone resulted in an early increase in IL-6
mRNA levels noticed already after 1 h and present up to 3 h after addition of IL-1β (Fig
8). In additional experiments, IL-6 mRNA levels had returned to basal levels after 20 h
(data not shown), a time point at which IL-6 protein levels were increased. These
observations are in line with previous reports from our laboratory showing an early
upregulation (within 1 h) of IL-6 mRNA levels in IL-1β-treated Caco-2 cells (29,30) and
support the concept that IL-1β-induced IL-6 production is regulated at the transcriptional
level. Importantly, in the present experiments, addition of live L. paracasei to the Caco-2
cells resulted in a potentiation of the IL-1β-induced IL-6 mRNA levels with an almost 5-
fold potentiation noticed after 1 h and an approximately 10-fold potentiation noticed after
3 h (Fig 8). Western blotting followed by densitometry showed a 23%, 51%, and 20%
increase in hsp27 levels in L. paracasei-treated Caco-2 cells at 1 h, 2 h, and 3 h,
respectively (means from 2 experiments). The corresponding figures for hsp70 levels
were 20%, 57%, and 33%. Thus, treatment of Caco-2 cells with live L. paracasei resulted
Page 16 of 42

Reilly et al - 17
in an early heat shock response that was present when an early potentiation of IL-6
mRNA expression was noticed. Although not conclusive, the results are supportive of a
role of the heat shock response in the potentiating effects of live L. paracsei on IL-6
production in IL-1β-treated Caco-2 cells.
It should be noted that hsp27 and hsp70 were upregulated earlier (1-3 h) in the
current study than in the study by Petrof et al (31) where hsp25 and hsp72 were increased
12 h after treatment of cultured young adult mouse colon (YAMC) cells with the
probiotic formulation VSL#3. The reasons for the different time course of heat shock
protein induction between our study and that of Petrof et al (31) are not known but may
be differences in cell types being used or different probiotics used for treatment of the
cells.
DISCUSSION
In the present study, the probiotic bacterium L. paracasei potentiated the effects of
IL-1β on IL-6 production in cultured Caco-2 cells. In addition, treatment of the Caco-2
cells with live L. paracasei increased the expression of hsp70 and hsp27 and silencing of
these genes with siRNA or treatment of the cells with quercetin prevented the effects of
live L. paracasei suggesting that the potentiating effect of L. paracasei on IL-6
production was, at least in part, regulated by the heat shock response. Because heat-
inactivated and sonicated L. paracasei also potentiated the IL-6 production in IL-1β-
treated Caco-2 cells, without inducing a heat shock response, multiple mechanisms in
addition to the heat shock response are probably involved in the potentiating effect of
probiotics on enterocyte IL-6 production.
Page 17 of 42

Reilly et al - 18
It should be noted that most experiments in the present study were performed in 80-
90% confluent Caco-2 cells rather than in fully differentiated enterocytes. This
experimental design was chosen to make comparisons possible with multiple previous
studies in which we examined IL-6 production in approximately 80-90% confluent Caco-
2 cells, including studies in which the potentiating effects of the heat shock response on
IL-6 production were determined (29,30,34). Importantly, we and others found recently
that although the absolute amounts of IL-6 produced from the apical and basolateral
membranes in fully differentiated cultured intestinal epithelial cells may be different,
treatment of the cells with IL-1β and other cytokines induced qualitatively similar
responses at the apical and basolateral membranes (24,41). In a control experiment
performed in the current study, L. paracasei potentiated the IL-1β-induced IL-6
production in fully differentiated Caco-2 cells cultured in a transwell bicameral system
(although to a lesser degree than noticed in 80-90% Caco-2 cells), further validating the
use of 80-90% confluent Caco-2 cells for the study of mechanisms involved in the
regulation of IL-6 production by probiotics.
Although beneficial effects of probitotics have been reported in several recent
clinical studies in the treatment of various inflammatory bowel diseases (3,8,9,13-15) and
as preoperative treatment of patients undergoing major surgical procedures (4,36),
additional well-controlled clinical studies are needed to further define the role of
probiotics in the management of inflammatory bowel disease. Probiotics probably exert
beneficial effects through multiple mechanisms. In general, probiotics may provide
benefits by bacterial interference with intestinal pathogens (5,37) and by direct
interaction with cells in the intestinal mucosa (18,19,26,31,49). In previous studies,
Page 18 of 42

Reilly et al - 19
treatment of cultured enterocytes with probiotics resulted in inhibited activity of the
transcription factor NF-kB (26,31), upregulated expression of the antimicrobial peptide
human beta defensin-2 (49), improved epithelial barrier function (19), prevention of
cytokine-induced apoptosis (52) and reduced production of the pro-inflammatory
cytokine IL-8 (39). The apparently contradictory results in the study by Ma et al (18), i.e.,
reduced IL-8 production in stimulated enterocytes treated with probiotics, and the results
in the present report of unchanged IL-8 production in L. paracasei-treated Caco-2 cells,
may reflect different stimuli used to induce IL-8 production [TNFα in the study by Ma et
al (18) and IL-1β in the present study], different types of probiotics [L. reuteri in the
study by Ma et al (18) versus L. paracasei in the present study] and different enterocyte
cell lines [T84 and HT-29 cells in the study by Ma et al (18) and Caco-2 cells in the
present report].
An additional potential mechanism by which probiotics may provide mucosal
protection, perhaps secondary to induction of the heat shock response, is increased
production of the anti-inflammatory cytokine IL-10 (45). Interestingly, recent studies
suggest that local delivery in the gut of IL-10 by genetically manipulated Lactococcus
lactis may be beneficial in the treatment of patients with Crohn’s disease (6,42).
In the present study, we found a novel mechanism by which probiotics may exert
beneficial effects, i.e., potentiated IL-6 production by enterocytes subjected to an
inflammatory stimulus. This observation is important because there are multiple lines of
evidence suggetsing that IL-6 has mainly anti-inflammatory and protective effects in the
intestinal mucosa. For example, in previous studies we found that heat shock-induced
potentiation of mucosal IL-6 production in endotoxemic mice prevented mucosal injury
Page 19 of 42

Reilly et al - 20
(45,47). In other studies, IL-6 exerted cell protective effects in cultured enterocytes by
inducing thermotolerance (11). Studies from other laboratories as well support the
concept that IL-6 may act as an anti-inflammatory cytokine controlling both local and
systemic inflammatory responses (2,40,51).
Interestingly, in the present study, L. paracasei and L. plantarum did not influence
IL-6 production in the cultured Caco-2 cells under basal conditions but only in cells that
were treated with IL-1β. This observation supports clinical observations that probiotics
provide beneficial effects in the setting of intestinal inflammation (3,4,8,9,13-15). A lack
of effect of probiotics on basal cytokine production in enterocytes has been reported by
others as well (18,19).
In the present study, both live and heat-inactivated L. paracasei enhanced IL-6
production in IL-1β-stimulated enterocytes. In contrast, treatment of the cells with culture
medium from the probiotics did not influence IL-6 production. These observations
suggest that the effects of L. paracasei noticed in the present study were caused by a
factor present in the bacteria, possibly in the bacterial wall, rather than by a secreted
product. In previous reports, the effects of probiotics in cultured intestinal epithelial cells
were induced by live (18) or inactivated bacteria (49) or by secreted product(s) present in
the culture medium of the probiotics (31,43). These apparently conflicting results may
reflect differences in probiotic bacteria as well as different intestinal epithelial cell lines
being used in previous studies. It is also possible that different effects of probiotics are
caused by different mechanisms. Regardless, the observations suggest that probiotics can
exert their biological effects by various cell-associated and secreted factors.
Page 20 of 42

Reilly et al - 21
Induction of the heat shock response by probiotics, as observed in the present study,
is similar to a recent report by Petrof et al (31) in which treatment of cultured mouse
colonic epithelial cells with the probiotics VSL #3 resulted in increased expression of
hsp72 and 25. In that study (31), evidence was found that inhibition of the chymotrypsin-
like activity of the proteasome may be a mechanism by which probiotics induce the heat
shock response. Although we did not examine the effect of L. paracasei on Caco-2 cell
proteasome activity in the current report, in recent experiments we found that treatment
of cultured Caco-2 cells with proteasome inhibitors resulted in a heat shock response (34)
similar to the effects of L. paracasei noticed here. Induction of the heat shock response
by proteasome inhibition was reported in a number of other cell types as well
(7,16,20,53).
Our results from experiments in which treatment of the Caco-2 cells with quercetin
or hsp70 or hsp27 siRNAs blocked the L. paracasei-induced potentiation of IL-6
production strongly suggest that the heat shock response was involved in the regulation
of IL-6 production. The observation that the heat shock response regulates IL-6
production in stimulated enterocytes is in line with previous reports from this laboratory
in which the heat shock response was induced by other mechanisms. Thus, in previous
experiments we found that when the heat shock response was induced in cultured Caco-2
cells by hyperthermia, the IL-1β-induced IL-6 production as well as the expression of IL-
6 mRNA were upregulated (29). Further support for a role of the heat shock response in
enterocyte IL-6 production was found in subsequent experiments in which treatment of
cultured Caco-2 cells with proteasome inhibitors induced a heat shock response and
potentiated IL-6 production (34). In additional experiments, induction of the heat shock
Page 21 of 42

Reilly et al - 22
response in vivo by hyperthermia or treatment with sodium arsenite augmented mucosal
IL-6 production in endotoxemic mice (47). Similar to the results in the present study, heat
shock by itself did not influence enterocyte or mucosal IL-6 production in our previous
studies but augmented IL-6 production that was already increased by endotoxemia in
mice (47) or treatment of Caco-2 cells with IL-1β (29,34). Although the mechanisms by
which the heat shock response potentiates IL-6 production in stimulated enterocytes are
not fully known at present, we recently found evidence that upregulated expression and
activity of the transcription factor C/EBPβ may play an important role (12).
In conclusion, the present study provides the first evidence that the probiotic
bacterium L. paracasei may enhance IL-6 production in enterocytes subjected to an
inflammatory stimulus and that the effect of live L. paracasei is, at least in part, heat
shock-dependent. Because other studies have shown that IL-6 has anti-inflammatory and
protective effects in the intestinal mucosa, the present results offer a novel mechanism by
which probiotics may exert some of their beneficial effects although additional
experiments will be needed to define the role of IL-6 in cell protective effects provided
by probiotics.
Page 22 of 42

Reilly et al - 23
REFERENCES
1. Annuk H, Shchepetova J, Kullisaar T, Songisepp E, Zilmer M, Mikelsaar M.
Characterization of intestinal lactobacilli as putative probiotic candidates. J Appl
Microbiol 94:403-412,2003.
2. Barton BE, Jackson JV. Protective role of interleukin-6 in the
lipopolysaccharide-galactosamine septic shock model. Infect Immun 61:1496-
1499,1993.
3. Bengmark S. Synbiotics and the mucosal barrier in critically ill patients. Curr
Opin Gastroenterol 21:712-716,2005.
4. Bengmark S, Martindale R. Probiotics and synbiotics in clinical medicine. Nutr
Clin Pract 20:244-261,2005.
5. Bibel DJ. Bacterial interference, bacteriotherapy, and bacterioprophylaxis. In Aly
R, Shinefield M (eds), Bacterial Interference, 1982, pp 1-12.
6. Braat H, Rottiers P, Hommes DW, Huyghebaert N, Remant E, Remon JP, van
Deventer SJ, Neirynck S, Peppelenbosch MP, Steidler L. A phase I trial with
transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin
Gastroenterol Hepatol 4:754-759,2006.
7. Bush KT, Goldberg AL, Nigam SK. Proteasome inhibition leads to a heat-shock
response, induction of endoplasmic reticulum chaperones, and thermotolerance. J
Biol Chem 14:9086-9092,1997.
8. Gionchetti P, Rizzello F, Helwig U, Venturi A, Lammers KM, Brigidi P, Vitali
B, Poggioli G, Miglioli M, Campieri M. Prophylaxis of pouchitis onset with
Page 23 of 42

Reilly et al - 24
probiotic therapy: a doube-blind, placebo-controlled trial. Gastroenterology
124:1202-1209,2003.
9. Gionchetti P, Rizzello F, Venturi A, Brigidi P, Matteuzzi D, Bazzochi G,
Poggioloi G, Miglioli M, Campieri M. Oral bacteriotherapy as maintenance
treatment in patients with chronic pouchitis: a double-blind, placebo-controlled
trial. Gastroenterology 119:305-309,2000.
10. Hasselgren PO. The significance of intestinal cytokines. In: Cytokines and the
Abdominal Surgeon. Schein M and Wise L (eds). Landes, Austin, TX, 1998, pp
197-213.
11. Hershko DD, Robb BW, Luo GJ, Paxton JH, Hasselgren PO. Interleukin-6
induces thermotolerance in cultured Caco-2 cells independent of the heat shock
response. Cytokine 21:1-9,2003.
12. Hungness ES, Robb BW, Luo GJ, Hershko DD, Hasselgren PO. Hyperthermia-
induced heat shock activates the transcription factor C/EBPβ and augments IL-6
production in human intestinal epithelial cells. J Am Coll Surg 195:619-
626,2002.
13. Isolauri E. Probiotics in human disease. Am J Clin Nutr 73:1142S-1146S,2001.
14. Isolauri E, Juntunen M, Rautanen T, Sillanaukee P, Koivula T. A human
Lactobacillus strain (Lactobacillus casei sp strain GG) promotes recovery from
acute diarrhea in children. Pediatrics 88:90-97,1991.
15. Isolauri E, Kirjavainen PV, Salminen S. Probiotics: a role in the treatment of
intestinal infection and inflammation. Gut 50 (Suppl 3):III54-III59,2002.
Page 24 of 42

Reilly et al - 25
16. Kawazoe Y, Nakai A, Tanabe M, Nagata K. Proteasome inhibition leads to the
activation of all members of the heat shock factor family. Eur J Biochem
255:356-362,1998.
17. Luyer MD, Buurman WA, Hadfoune M, Speelmans G, Knol J, Jacobs JA,
Dejong CHC, Vriesema AJM, Greve JWM. Strain-specific effects of probiotics
on gut barrier integrity following hemorrhagic shock. Infect Immun 73:3686-
3692,2005.
18. Ma D, Forsythe P, Bienenstock J. Live Lactobacillus reuteri is essential for the
inhibitory effect on tumor necrosis factor alpha-induced interleukin-8 expression.
Infect Immun 72:5308-5314,2004.
19. Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, Yachimec C, Doyle J,
Jewell L, DeSimone C. Probiotic bacteria enhance murine and human intestinal
epithelial barrier function. Gastroenterology 121:580-591,2001.
20. Mathew A, Mathur SK, Morimoto RI. Heat shock response and protein
degradation: regulation of HSF 2 by the ubiquitin-proteasome pathway. Mol Cell
Biol 18:5091-5098,1998.
21. Mester M, Tompkins RG, Gelfand JA, Dinarello CA, Burke JF, Clarke BD.
Intestinal production of IL-1α during endotoxemia in the mouse. J Surg Res
54:584-591,1993.
22. Meyer TA, Noguchi Y, Ogle CK, Tiao G, Wang JJ, Fischer JE, Hasselgren PO.
Endotoxin stimulates interleukin-6 production in intestinal epithelial cells. A
synergistic effect with prostaglandin E2. Arch Surg 129:1290-1295,1994.
Page 25 of 42

Reilly et al - 26
23. Meyer TA, Wang J, Tiao G, Ogle CK, Fischer JE, Hasselgren PO. Sepsis and
endotoxemia stimulate intestinal interleukin-6 production. Surgery 118:336-
342,1995.
24. Moon MR, Parikh AA, Pritts TA, Kane C, Fischer JE, Salzman AL, Hasselgren
PO. Interleukin-1β induces complement component C3 and IL-6 production at
the basolateral and apical membranes in a human intestinal epithelial cell line.
Shock 13:374-378,2000.
25. Nagai N, Nakai A, Nagata K. Quercetin suppresses heat shock response by
downregulation of HSF 1. Biochem Biophys Res Commun 208:1099-1105,1995.
26. Neish AS, Gewirtz AT, Zeng H, Young AN, Hobert ME, Karmali V, Rao AS,
Madara JL. Prokaryotic regulation of epithelial responses by inhibition of IkB-α
ubiquitination. Science 289:1560-1563,2000.
27. Niedzielin K, Kordecki H, Birkenfeld B. A controlled, double-blind, randomized
study on the efficacy of Lactobacillus plantarum 299V in patients with irritable
bowel syndrome. Eur J Gastroenterol Hepatol 13:1143-1147,2001.
28. Papanicolaou DA, Wilder RL, Monolapas SC, Chrousos GP. The
pathophysiologic roles of interleukin-6 in human disease. NIH conference. Ann
Intern Med 128:127-137,1998.
29. Parikh AA, Moon MR, Kane CD, Salzman AL, Fischer JE, Hasselgren PO.
Interleukin-6 production in human intestinal epithelial cells increases in
association with the heat shock response. J Surg Res 77:40-44,1998.
Page 26 of 42

Reilly et al - 27
30. Parikh AA, Salzman AL, Fischer JE, Szabo C, Hasselgren PO. Interleukin-1β
and interferon-γ regulate interleukin-6 expression in cultured human intestinal
epithelial cells. Shock 8:249-255,1997.
31. Petrof EA, Kojima K, Ropeleski MJ, Musch MW, Tao Y, DeSimone C, Chang
EB. Probiotics inhibit nuclear factor-kB and induce heat shock proteins in
colonic epithelial cells through proteasome inhibition. Gastroenterology
127:1474-1487,2004.
32. Pinsky MR, Vincent JL, Deviere J, Alegre M, Kahn RJ, Dupont E. Serum
cytokines in human septic shock: relation to multiple organ failure and mortality.
Chest 103:565-575,1993.
33. Pinto M, Robine-Leon S, Appay MD, Kedinger M, Triadon N, Dussaulx E, et al.
Enterocyte-like differentiation and polarization of the human colon carcinoma
cell line Caco-2 in culture. Biol Cell 47:323-330,1983.
34. Pritts TA, Hungness ES, Hershko DD, Robb BW, Sun X, Luo GJ, Fischer JE,
Wong HR, Hasselgren PO. Proteasome inhibitors induce heat shock response
and increase IL-6 expression in human intestinal epithelial cells. Am J Physiol
282:R1016-R1026,2002.
35. Pritts TA, Hungness E, Wang Q, Robb B, Hershko D, Hasselgren PO. Mucosal
and enterocyte IL-6 production during sepsis and endotoxemia. Role of
transcription factors and regulation by the stress response. Am J Surg 183:372-
383,2002.
36. Rayers N, Seehofer D, Theruvath T, Langrehr JM, Muller AR, Bengmark S,
Neuhaus P. Combined perioperative enteral supply of bioactive pre- and
Page 27 of 42

Reilly et al - 28
probiotics abolishes postoperative bacterial infections in human liver
transplantation – a randomized, double-blind clinical trial. Am J Transplant
5:125-130,2005.
37. Reid G, Howard J, Gan BS. Can bacterial interference prevent infection? Trends
Microbiol 9:424-428,2001.
38. Salminen S, Ouwehand A, Benno Y, Lee YK. Probiotics: how should they be
defined? Trends Food Sci Technol 10:107-110,1999.
39. Savidge TC, Newman PG, Pan WH, Weng MQ, Shi HN, McCormick BA,
Quaroni A, Walker WA. Lipopolysaccharide-induced human enterocyte
tolerance to cytokine-mediated interleukin-8 production may occur
independently of TLR-4/MD-2 signaling. Pediatr Res 59:89-95,2006.
40. Shanley TP, Foreback JL, Pemick DG, Ulich TR, Kunkel SL, Ward PA.
Regulatory effects of interleukin-6 in immunoglobulin G immune-complex-
induced lung injury. Am J Pathol 151:193-203,1997.
41. Sitaraman SV, Merlin D, Wang L, Wong M, Gewirtz AT, Si-Tahar M, Madara
JL. Neutrophil-epithelial crosstalk at the intestinal lumenal surface mediated by
reciprocal secretion of adenosine and IL-6. J Clin Invest 107:861-869,2001.
42. Steidler L, Hans W, Schotte L, Neirynck S, Obermeier F, Falk W, Fiers W,
Remaut E. Treatment of murine colitis by Lactococcus lactis secreting
interleukin-10. Science 289:1352-1355,2000.
43. Tao Y, Drabik KA, Waypa TS, Musch MW, Alverdy JC, Schneewind O, Chang
EB, Petrof EO. Soluble factors from Lactobacillus GG activate MAPKs and
Page 28 of 42

Reilly et al - 29
induce cytoprotective heat shock proteins in intestinal epithelial cells. Am J
Physiol 290:C1018-1030,2006.
44. Todoriki K, Mukai T, Sato S, Toba T. Inhibition of adhesion of food-borne
pathogens to Caco-2 cells by Lactobacillus strains. J Appl Microbiol 91:154-
159,2001.
45. Wang Q, Hasselgren PO. Heat shock response reduces intestinal permeability in
septic mice: potential role of interleukin-10. Am J Physiol 282:R669-R676,2002.
46. Wang W, Smail N, Wang P, Chaudry IH. Increased gut permeability after
hemorrhage is associated with upregualtion of local and systemic IL-6. J Surg
Res 79:39-46,1998.
47. Wang Q, Sun X, Pritts TA, Wong HR, Hasselgren PO. Induction of the stress
response increases interleukin-6 production in the intestinal mucosa of
endotoxemic mice. Clin Sci 99:489-496,2000.
48. Wang Q, Wang JJ, Boyce S, Fischer JE, Hasselgren PO. Endotoxemia and IL-1β
stimulate mucosal IL-6 production in different parts of the gastrointestinal tract. J
Surg Res 76:27-31,1998.
49. Wehkamp J, Harder J, Wehkamp K, Wehkamp-von Meissner B, Schlee M,
Enders C, Sonnenborn U, Nuding S, Bengmark S, Fellermann K, Schroder JM,
Stange EF. NF-kB- and AP-1-mediated induction of human beta defensin-2 in
intestinal epithelial cells by Escherichia coli Nissle 1917: a novel effect of a
probiotic bacterium. Infect Immun 72:5750-5758,2004.
Page 29 of 42

Reilly et al - 30
50. Wei W, Yang H, Cao P, Menconi M, Chamberlain C, Petkova V, Hasselgren PO.
Degradation of C/EBP beta in cultured myotubes is calpain-dependent. J Cell
Physiol 208:386-398,2006.
51. Xing Z, Gauldie J, Cox G, Baumann H, Jordana M, Lei XF, Achong MK. IL-6 is
an anti-inflammatory cytokine required for controlling local or systemic acute
inflammatory responses. J Clin Invest 101:311-320,1998.
52. Yan F, Polk DB. Probiotic bacterium prevents cytokine-induced apoptosis in
intestinal epithelial cells. J Biol Chem 277:50959-50965,2002.
53. Zhou M, Wu X, Ginsberg HN. Evidence that a rapidly turning over protein,
normally degraded by proteasomes, regulates hsp72 gene transcription in HepG2
cells. J Biol Chem 271:24769-24775,1996.
Page 30 of 42

Reilly et al - 31
LEGENDS TO FIGURES
Fig 1 The effects of IL-1β, L. paracasei and L. plantarum on IL-6 production in cultured
Caco-2 cells. (A) Cultured Caco-2 cells were treated for 20 h with 1 ng/ml of IL-1β,
108/ml of L. paracasei or L. plantarum alone or in combinations as indicated in the figure
followed by measurement of IL-6 levels in the culture medium. (B) Cultured Caco-2 cells
were treated for 20 h with 1 ng/ml of IL-1β, live or heat-inactivated L. paracasei (108
bacteria/ml) alone or in combinations as indicated in the figure followed by measurement
of IL-6 levels in the culture medium. Results are means ± SEM with n=3 in each group.
*p<0.05 vs control (untreated cells); +p<0.05 vs cells treated with IL-1β alone by
ANOVA. Identical experiments were performed three times with almost identical results.
Fig 2 The effects of IL-1β, medium from cultured L. paracasei, and sonicated L.
paracasei on IL-6 production in cultured Caco-2 cells. (A) Cultured Caco-2 cells were
treated for 20 h with 1 ng/ml of IL-1β, 108/ml of L. paracasei or medium from cultured
L. paracasei followed by measurement of IL-6 levels in the culture medium. (B) Cultured
Caco-2 cells were treated for 20 h with IL-1β, intact or sonicated L. paracasei alone or in
combinations as indicated in the figure and at concentrations identical to those used in
Fig 2A, followed by measurement of IL-6 in the culture medium. Results are means ±
SEM with n=3 in each group. *p<0.05 vs control; +p<0.05 vs IL-1β alone by ANOVA.
Identical experiments were performed three times with almost identical results.
Fig 3 (A) The effect of different concentrations of L. paracasei on IL-6 production in
Caco-2 cells cultured in the absence of presence of IL-1β. Cultured Caco-2 cells were
Page 31 of 42

Reilly et al - 32
treated for 20 h with different concentrations of L. paracasei alone or together with 1
ng/ml of IL-1β followed by measurement of IL-6 in the culture medium. (B) Time-course
for the potentiating effect of L. paracasei on IL-6 production in IL-1β-treated Caco-2
cells. Cells were treated with 1 ng/ml of IL-1β or IL-1β + 108/ml of L. paracasei for
various periods of time up to 20 h followed by measurement of IL-6 levels in the culture
medium. Results are means ± SEM with n=3 in each group. Identical experiments were
performed three times with almost identical results.
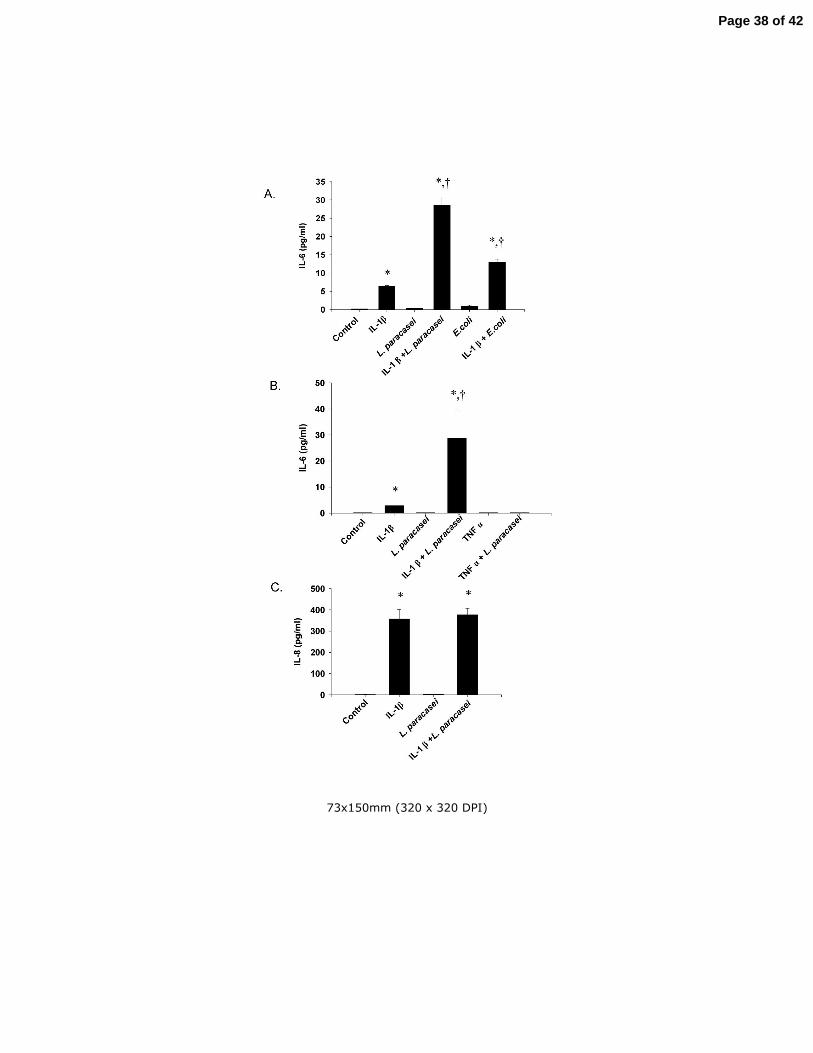
Fig 4 (A) The effects of L. paracasei and E. coli on IL-6 production in IL-1β-treated
Caco-2 cells. Cells were treated for 20 h with 1 ng/ml of IL-1β, 108/ml of L. paracasei, or
108/ml of DH5α E. coli, either alone or in combinations as indicated in the figure,
followed by measurement of IL-6 in the culture medium. (B) The effects of L. paracasei
on IL-6 production in IL-1β- and TNFα-treated Caco-2 cells. Cultured Caco-2 cells were
treated for 20 h with 1 ng/ml of IL-1β or 1 ng/ml of TNFα alone or in combination with
108/ml of L. paracasei. (C) The effect of IL-1β and L. paracasei on IL-8 production in
cultured Caco-2 cells. Cells were treated for 20 h with 1 ng/ml of IL-1β or 108/ml of L.
paracasei alone or in combination as indicated in the figure followed by measurement of
IL-8 levels in the culture medium. Results are means ± SEM with n=3 in each group.
*p<0.05 vs control; +p<0.05 vs IL-1β alone by ANOVA. Identical experiments were
performed three times with almost identical results.
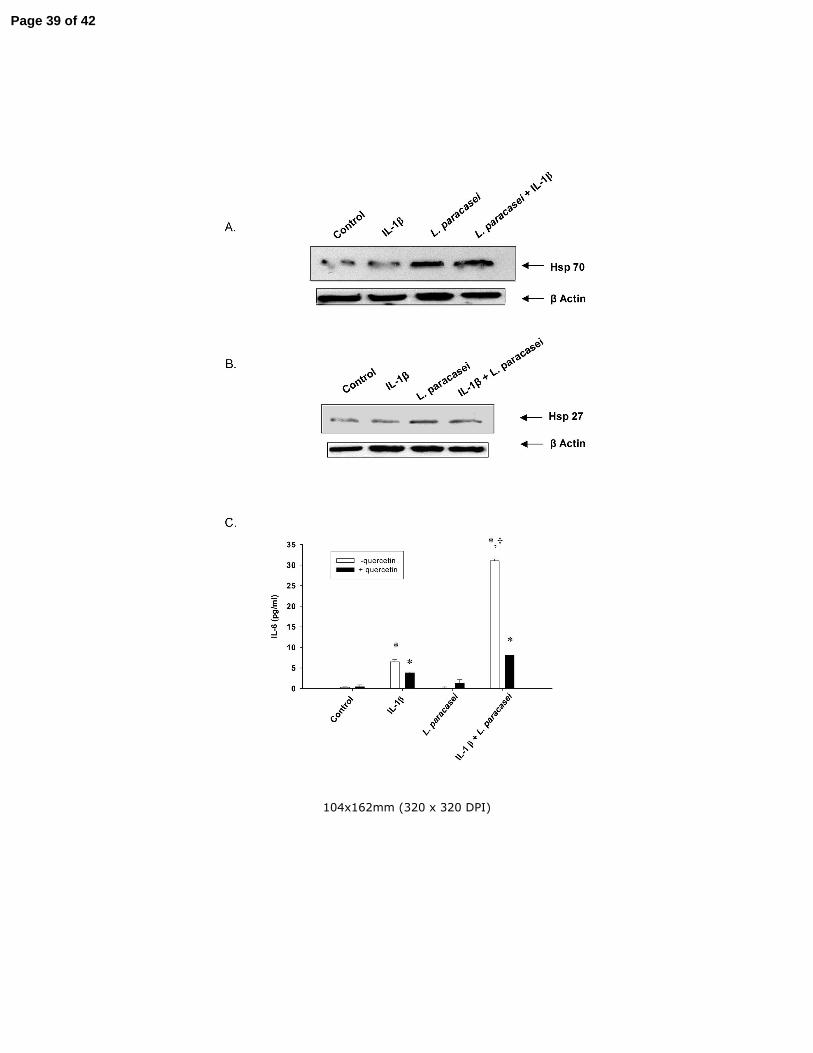
Fig 5 The influence of IL-1β and L. paracasei on cellular levels of (A) hsp70 and (B)
hsp27. Cultured Caco-2 cells were treated for 20 h with 1 ng/ml of IL-1β or 108/ml of L.
Page 32 of 42

Reilly et al - 33
paracasei alone or in combination as indicated in the figure. Hsp70 and hsp27 levels
were determined by Western blotting. β-Actin levels were determined as loading
controls. The experiment was repeated three times with almost identical results. (B) The
effect of quercetin on L. paracasei-induced potentiation of IL-6 production in IL-1β-
treated Caco-2 cells. Cells were treated for 20 h with 1 ng/ml of IL-1β or 108/ml L.
paracasei in the absence or presence of 100 µM quercetin as indicated in the figure.
Results are means ± SEM with n=3 in each group. *p<0.05 vs corresponding control
group; +p<0.05 vs corresponding IL-1β group by ANOVA. Identical experiments were
performed three times with almost identical results.
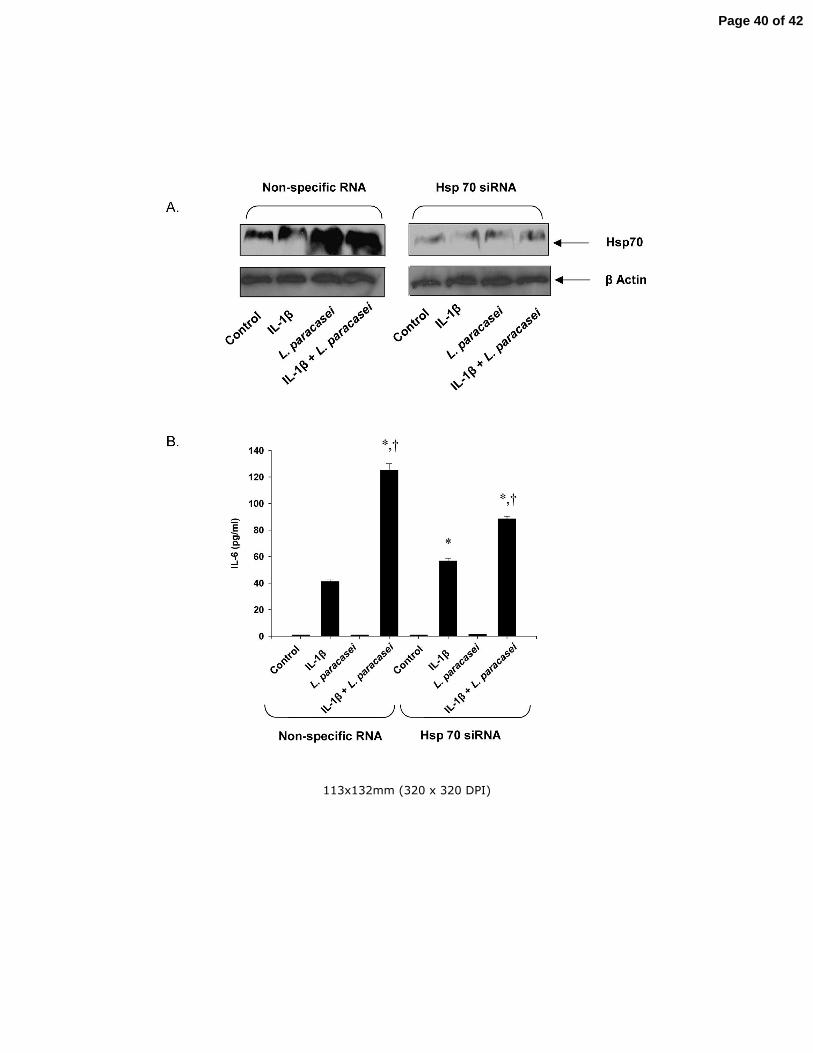
Fig 6 The influence of reduced hsp70 levels on the L. paracasei-induced potentiation of
IL-6 production in IL-1β-treated Caco-2 cells. (A) Hsp70 levels in cultured Caco-2 cells
transfected with hsp70 siRNA or control (non-specific) RNA as described in Materials
and Methods and subsequently treated with IL-1β or L. paracasei alone or in
combination as indicated in the figure. (B) The effects of IL-1β and L. paracasei on IL-6
production in Caco-2 cells transfected with hsp70 siRNA or non-specific RNA. Results
are means ± SEM with n=3 in each group. *p<0.05 vs control; +p<0.05 vs corresponding
IL-1β group by ANOVA. Identical experiments were performed 3 times with almost
identical results.
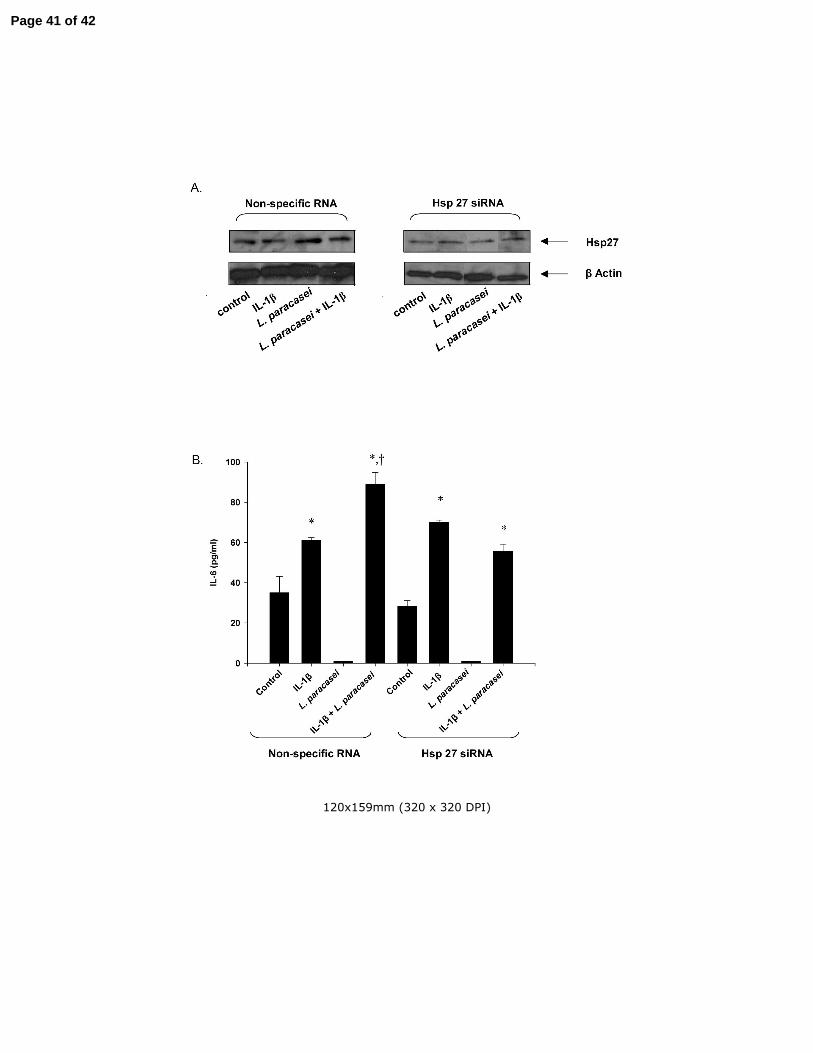
Fig 7 The influence of reduced hsp27 levels on the L. paracasei-induced potentiation of
IL-6 production in IL-1β-treated Caco-2 cells. (A) Hsp27 levels in cultured Caco-2 cells
transfected with hsp27 siRNA or control (scrambled) RNA as described in Materials and
Page 33 of 42

Reilly et al - 34
Methods and subsequently treated with IL-1β or L. paracasei alone or in combination as
indicated in the figure. (B) The effects of IL-1β and L. paracasei on IL-6 production in
Caco-2 cells transfected with hsp27 siRNA or control RNA. Results are means ± SEM
with n=3 in each group. *p<0.05 vs control; +p<0.05 vs corresponding IL-1β group by
ANOVA. Identical experiments were performed three times with almost identical results.
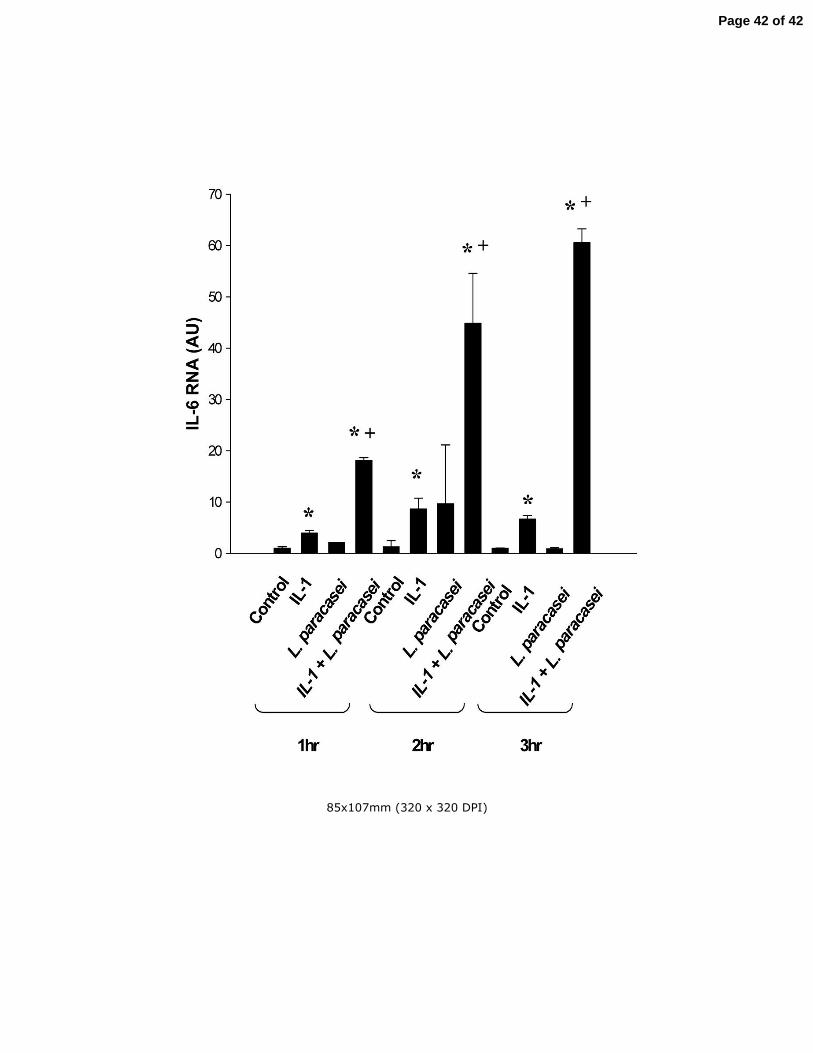
Fig 8 The effects of IL-1β and live L. paracasei on IL-6 mRNA levels in cultured Caco-2
Cells. Cells were treated for 1, 2, or 3 h with IL-1β, L. paracsei either alone or in
combination as indicated in the figure. Untreated cells served as control. IL-6 mRNA
levels were determined by real-time PCR. Results are means ± SEM with n=6 in each
group. *p<0.05 vs control at the corresponding time point; +p<0.05 vs IL-1β alone at the
corresponding time point by ANOVA.
Page 34 of 42

76x126mm (320 x 320 DPI)
Page 35 of 42

74x119mm (320 x 320 DPI)
Page 36 of 42

78x131mm (320 x 320 DPI)
Page 37 of 42
73x150mm (320 x 320 DPI)
Page 38 of 42
104x162mm (320 x 320 DPI)
Page 39 of 42
113x132mm (320 x 320 DPI)
Page 40 of 42
120x159mm (320 x 320 DPI)
Page 41 of 42
85x107mm (320 x 320 DPI)
Page 42 of 42




















