
2Methoxyestradiol overcomes drug resistance in multiple myeloma cells
39
doi:10.1182/blood-2002-02-0376 Prepublished online May 24, 2002; Kenneth C Anderson Martin Sattler, Paul Richardson, Robert L Schlossman, Klaus Podar, Edie Weller, Nikhil Munshi and Dharminder Chauhan, Laurence Catley, Teru Hideshima, Guilan Li, Richard Leblanc, Deepak Gupta, cells 2-Methoxyestradiol (2ME2) overcome drug resistance in multiple myeloma (4212 articles) Neoplasia Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: digital object identifier (DOIs) and date of initial publication. the indexed by PubMed from initial publication. Citations to Advance online articles must include final publication). Advance online articles are citable and establish publication priority; they are appeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet Copyright 2011 by The American Society of Hematology; all rights reserved. 20036. the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.org From For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.org From
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of 2Methoxyestradiol overcomes drug resistance in multiple myeloma cells

doi:10.1182/blood-2002-02-0376Prepublished online May 24, 2002;
Kenneth C AndersonMartin Sattler, Paul Richardson, Robert L Schlossman, Klaus Podar, Edie Weller, Nikhil Munshi and Dharminder Chauhan, Laurence Catley, Teru Hideshima, Guilan Li, Richard Leblanc, Deepak Gupta, cells2-Methoxyestradiol (2ME2) overcome drug resistance in multiple myeloma
(4212 articles)Neoplasia �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

1
2-METHOXYESTRADIOL (2ME2) OVERCOME DRUG RESISTANCE IN MULTIPLE
MYELOMA CELLS
Dharminder Chauhan, Laurence Catley, Teru Hideshima, Guilan Li,
Richard Leblanc, Deepak Gupta, Martin Sattler, Paul Richardson,
Robert L. Schlossman, Klaus Podar, Edie Weller, Nikhil Munshi,
Kenneth C. Anderson$
Department of Adult Oncology, Dana-Farber Cancer Institute;
Harvard Medical School, Boston, MA 02115
Running title: 2ME2 overcomes drug resistance in MM
This investigation was supported by National Institutes of Health
Grants R0-1 50947 and PO-1 78378, the Multiple Myeloma Research
Foundation (MMRF) Senior Research Awards (DC, TH), International
Myeloma Foundation Fellow Award (KP), and the Doris Duke
Distinguished Clinical Research Scientist Award (KA)
$Address correspondence and reprints to:
Kenneth C. Anderson, MD
Dana-Farber Cancer Institute
44 Binney Street, Boston, MA 02215
phone: (617)-632-2144 FAX:(617)-632-2569
E-mail: [email protected]
Total text Word count: 4689 Total abstract word count: 220
Scientific Heading: Neoplasia
Copyright 2002 American Society of Hematology
Blood First Edition Paper, prepublished online May 24, 2002; DOI 10.1182/blood-2002-02-0376 For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

2
Abstract
2-Methoxyestradiol (2ME2), an estrogen-derivative, induces
growth arrest and apoptosis in leukemic cells and is also
antiangiogenic. In this study, we demonstrate that 2ME2 inhibits
growth and induces apoptosis in MM cell lines and patient cells.
Importantly, 2ME2 also inhibits growth and induces apoptosis in
MM cells resistant to conventional therapies including melphalan
(LR-5), doxorubicin (Dox-40 and Dox-6), and dexamethasone
(MM.1R). In contrast to its effects on MM cells, 2ME2 does not
reduce survival of normal peripheral blood lymphocytes.
Moreover, 2ME2 enhances Dex-induced apoptosis and its effect is
not blocked by interleukin-6 (IL-6). We next examined the effect
of 2ME2 on MM cells in the bone marrow (BM) milieu. 2ME2
decreases survival of BM stromal cells (BMSCs) and secretion of
vascular endothelial growth factor (VEGF) as well as IL-6
triggered by adhesion of MM cells to BMSCs. We show that
apoptosis induced by 2ME2 is mediated by release of mitochondrial
cytochrome-c (cyto-c) and Smac; followed by activation of
caspases-8, -9, and -3. Finally, 2ME2 inhibits MM cell growth,
prolongs survival, and decreases angiogenesis in a murine model.
These studies therefore demonstrate that 2ME2 mediates anti-MM
activity both directly on MM cells and in the BM
microenvironment. They provide the framework for the use of
2ME2, either alone or in combination with Dex, to overcome drug
resistance and improve outcome in MM.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

3
Introduction
2-Methoxyestradiol (2ME2), a natural metabolite of
estradiol, is a potent anti-tumor and anti-angiogenic agent in
leukemic cells 1-7. Although 2ME2 has potent anti-proliferative
activity in various tumorigenic and non-tumorigenic cell lines,
the mechanisms mediating 2ME2 biologic effects remain unclear.
We and others have demonstrated that multiple myeloma (MM) cells
express estrogen receptors (ERs) and that anti-estrogen compounds
such as, tamoxifen and ICI 182, 780 induce apoptosis in MM cells
8-11. Treatment of MM cells with estrogen (E2) alone did not
alter the growth of MM cells 9. Moreover, despite being a
natural derivative of estradiol (E2), 2ME2 binds poorly to the
ERs and therefore the anti-proliferative effects of 2ME2 are not
mediated by ERs 12,13. These findings, coupled with our studies
suggest that 2ME2-mediated effects on MM cells are independent of
ER.
Previous studies using xenografts and metastatic disease
models in mice have suggested that 2ME2 targets not only the
tumor cell, but also the tumor vasculature 2,14. Recent reports
of increased bone marrow (BM) angiogenesis in multiple myeloma
(MM) 15-17, coupled with the known anti-angiogenic properties of
2ME2 2,14, provide a potential rationale for evaluating 2ME2 as a
novel therapeutic in MM.
In the present study, we characterize the in vitro effects
of 2ME2 against MM cells in the BM microenvironment, as well as
its in vivo anti-MM activity in an animal model. We demonstrate
that 2ME2 induces apoptosis in MM cells resistant to conventional
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

4
therapies and delineate the signaling cascades mediating its
anti-MM activity. Importantly, we show that 2ME2 directly
decreases cytokine secretion in bone marrow stromal cells
(BMSCs), thereby modulating the tumor BM environment. These
data, along with our demonstration of its anti-MM and anti-
angiogenesis effects in vivo, provide the framework for clinical
trials of 2ME2 to improve patient outcome in MM.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

5
Materials and Methods
Cell culture and reagents. Dex-sensitive MM.1S and Dex-
resistant MM.1R human MM cell lines were kindly provided by Dr
Steven Rosen (Northwestern University, Chicago, IL) 18. Dox-6,
Dox-40, and LR-5- drug resistant RPMI 8226 human MM cell lines
were kindly provided by Dr. William Dalton (Moffit Cancer Center,
Tampa, FL) 19. U266 MM cells were obtained from the American
Type Culture Collection (Rockville, MD). All cell lines were
grown in RPMI-1640 media supplemented with 10% heat inactivated
fetal-bovine serum, 100 units/ml penicillin, 100 µg/ml
streptomycin, and 2 mM L-glutamine. Drug resistant cell lines
were cultured with either Dox or Dex to confirm their lack of
drug sensitivity. Mononuclear cells were prepared from MM
patient BM samples by Ficoll-Hypaque density gradient
centrifugation. The MM cells were freshly isolated from patients
relapsing after multiple prior therapies including Dexamethasone,
Melphalan, and thalidomide. An informed consent was obtained from
all patients in accordance with the Helsinki protocol. Tumor
cells (CD138+ 97 + 2.0%) were isolated by CD138 positive
selection method using CD138 (Syndecan-1) Micro Beads and Auto
MACS magnetic cell sorter machine, according to manufacturer's
instructions (Miltenyi Biotec Inc., Auburn, CA). CD138+ myeloma
cells were viable (94-97%) for 2-3 weeks in vitro. Cells were
treated with 0.05 µM Dex (Sigma Chemical Co, St. Louis, MO) or
2ME2 (1-20 µM).
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

6
Proliferation assays. The BrdUrd incorporation assay was
performed using BrdUrd proliferation assay kit, according to
manufacturer's instructions (Calbiochem, San Diego, CA) with some
modifications 20. Briefly, cells were seeded onto 96-well plates
in RPMI 1640 medium containing 10% FBS. Medium was replaced 24h
later with RPMI-1640 medium containing 2% FBS, with or without
2ME2. Plates were incubated for 48h and 72h, pulsed for 2h-6h
with BrdUrd primary and secondary antibodies, and developed using
a colorimetric reaction. The absorbance at 460 nm was measured
using a Molecular Device plate reader.
MTT assays. Cell viability was assessed by 3-(4,5-
dimethylthiozol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Chemicon International Inc., Temecula, CA) assay according to
manufacturer's instructions (Roche Molecular Biochemical’s), with
some modifications. Cells were seeded in 96-well plates in RPMI-
1640 medium containing 5% FBS. 2ME2 or Dex was added 24h later
and incubated for 72h. Cell viability was evaluated as
previously described 21,22.
Enzyme linked Immunosorbent (ELISA) assays. Conditioned
media were generated from 24h cultures of MM patient-derived
BMSCs, MM.1S cells, and BMSCs + MM.1S cells, either untreated or
treated with 3 µM 2ME2; IL-6 and VEGF levels were measured using
ELISAs (R & D Systems, Minneapolis, MN), as previously described
23. BMSC viability at 24h was >80%, as determined by Trypan blue
exclusion assay. Briefly, 96 well plates were coated with either
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

7
anti-human VEGF or IL-6 antibodies overnight, washed, and then
blocked with 300 µL PBS, 1% bovine serum albumin (BSA), 5%
sucrose and 0.05% NaN3 for 2 h. After washing, 100 µL of sample
or standards diluted in Tris-Cl, 0.1% BSA, and 0.05% Tween 20 (pH
7.3) were added to the wells and incubated for 2 h at room
temperature (RT). The cells were rinsed, and either biotinylated
anti-human VEGF or IL-6 antibodies were added for 2 h at RT.
After a further washing step, the wells were incubated with
streptavidin horseradish peroxidase for 20 min at RT and rinsed.
The reaction was started by the addition of 100 µL H2O2 and
tetramethylbenzidine for 30 min at RT. After stopping the
reaction with 1 µM H2SO4, the optical density of each well was
detected by means of a microtitre plate reader at 450nm, with
correction at 540 nm.
Western blotting. Total cell lysates and cytosolic
extracts were prepared as previously described 24. Briefly,
equal amounts of proteins were resolved by 10% SDS-PAGE and
transferred onto nitrocellulose membranes. Proteins were then
transferred to nitrocellulose filters, blocked by incubation in
5% dry milk in PBST (0.05% Tween-20 in PBS), and probed with
anti-cyto-C, anti-Smac (kindly provided by Dr. Xiaodong Wang,
University of Texas Southwestern Medical Center at Dallas), anti-
Caspase-3 (Santa-Cruz, CA), anti-caspase-8 and 9 (Transduction
Labs, MI), or anti-SHP2 (Santa Cruz, CA) Abs. Blots were then
developed by enhanced chemiluminesence (ECL; Amersham, Arlington
Heights, IL). Preparation of cell lysates for PARP immunoblot
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

8
analysis was performed as described using C-2-10 anti-PARP
monoclonal antibody 25.
Quantification of apoptosis. Dual fluorescence staining
with DNA-binding fluorochrome Hoechst 33342 (HO) and propidium
iodide (PI) was used to quantitate the percentage of apoptotic
(PI-HO
+) cells using flow cytometry (The Vantage, Becton
Dickinson), as previously described 26. DNA fragmentation assays
were also performed, as in prior studies 27.
Preparation of S-100 cytosolic fractions from MM.1S cells.
MM.1S MM cells were washed twice with PBS, and the pellet was
suspended in 5 ml of ice cold buffer A (20 mM HEPES, pH 7.5, 1.5
mM MgCl2, 10 mM KCl, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 0.1 mM PMSF,
10 µg/ml leupeptin and aprotinin and pepstatin A) containing 250
nM sucrose. Cells were homogenized by douncing three times in a
Dounce homogenizer with a sandpaper-polished pestle. Cytosolic
fractions were isolated as previously described 24. The
immunoblots were scanned using an LKB produkter (Bromma, Sweden)
Ultrascan XL laser densitometer and analysed with the Gelscan
software package. Signal intensity was determined in a linear
range and normalized to that for SHP2.
Drug preparation and in vivo evaluation in murine tumor
model. For animal studies, 2ME2 was kindly provided by Entremed,
Inc. (Rockville, MD). 2ME2 (100 mg/kg) was suspended in 0.5%
carboxymethycellulose (Sigma). Six-week-old triple immune
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

9
deficient beige-nude-xid (BNX) mice were obtained from Frederick
Cancer Research and Development Center (Frederick, MD). All
animal studies were conducted according to protocols approved by
the Animal Ethics Committee of the Dana-Farber Cancer Institute.
Mice were observed daily for signs of toxicity. Terminal bleeding
was done under anesthesia using isoflourane inhalation, and
animals were sacrificed by CO2 asphyxiation.
To determine the in vivo anti-MM activity of 2ME2, mice were
inoculated subcutaneously in the flank with 3x107 RPMI 8226 MM
cells in 100 µl of RPMI-1640 media, together with 100 µl matrigel
(Becton Dickinson, Bedford, MA). Drug treatment was started
after the development of measurable tumor. The drug (100 mg/kg)
was given daily using an oro-gastric feeding tube, with
carboxymethylcellulose 0.5% as a control. Serial caliper
measurements of perpendicular diameters were done every other day
to calculate tumor volume, using the following formula: 4/24 x
(shortest diameter)2 x (longest diameter). Animals were
sacrificed if the tumor was > 2 cm or necrotic. Tumor was then
harvested for H/E staining and microvessel density (MVD)
evaluation. For tumor growth studies, 10 mice were used in each
group.
CD31 immunostaining. Tumor tissues were fixed in 10%
neutral buffered formalin and embedded in paraffin, according to
standard histological procedures. After deparaffinization,
tissue sections were pretreated with proteinase K (Roche
Molecular Biochemical’s, Indianapolis, IN) at 37°C for 30 min
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

10
before staining with rat anti-mouse CD-31 Ab (Pharmingen, San
Diego, CA). Positive staining was detected using secondary
biotinylated rabbit anti-rat Ab, followed by incubation with
streptavidin-horseradish peroxidase (DAKO, Carpenteria, CA). 3-
Amino-9-ethylene carbazol (DAKO) was used as the chromogenic
substrate, and sections were counterstained with Gill’s
Hematoxylin (Fisher Scientific, Fair Lawn, NJ). MVD was
determined by light microscopy, as previously described 28. MVD
assessment was done without knowledge of mouse treatment history.
Areas of most intense neovascularization were counted at high
magnification (400 X), and two investigators using double-headed
light microscopy analyzed at least 5 separate fields. The mean
MVD was compared among the treatment groups and analyzed using
the student's t test.
Statistical Analysis. Non-parametric tests and mixed models
were used to analyze the data. This includes the Wilcoxon's
signed rank test to compare proliferation in untreated and
treated patient cells and the Jonchkeere-Terpstra (J-T) trend
test for assessing viability of MM cell lines resistant to
conventional therapy and of normal donors. The mixed model was
used to adjust for repeated measures in the analysis of the 2ME2
patient MM-derived BMSCs. Analysis of in vivo data included a
Wilcoxon's rank-sum test on the change in the tumor volume and a
log-rank test to compare the overall survival.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

11
Results and Discussion
2ME2 induces apoptosis in MM.1S cells. To examine whether
2ME2 induces apoptosis in the MM.1S MM cell line, we performed
both DNA-fragmentation and flow cytometric analyses. MM.1S MM
cells were cultured in the presence or absence of 2ME2 (3 µM) for
24h and 48h. DNA cleavage was analyzed by agarose gel
electrophoresis of the α32P-ATP labeled genomic DNA (Fig 1A, upper panel). Low to undetectable DNA cleavage was observed in
the untreated cells; in contrast, 2ME2 induced significant
cleavage of DNA into oligonucleosomal fragments of approximately
200 base pairs. This pattern was more pronounced following
treatment with 2ME2 at 48h. The finding that 2ME2 induces
fragmentation of DNA into multiples of nucleosome-sized fragments
confirms changes in chromatin structure observed during apoptosis
or program cell death 29. Apoptosis induced by 2ME2 was further
confirmed by flow cytometric analysis using dual PI and Hoechst
staining. Treatment of cells with 2ME2 (3 µM) significantly (P <
0.005) increases the percentage of apoptotic cells (Fig 1A, lower
panel).
Effect of 2ME2 on MM cells resistant to conventional
therapy.
The cytotoxic effects of 2ME2 on various MM cell lines, including
cell lines resistant to conventional therapy (RPMI 8226, Dox-40,
Dox-6, LR5, and MM.1R) was determined using an MTT assay, in the
presence or absence of drug at various concentrations (0.5 µM, 1
µM, 3 µM, 10 µM, and 20 µM.). 2ME2 induced a significant (P <
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

12
0.0001) decrease in cell viability in a dose dependent manner for
all cell lines (Fig 1B). Fifty-percent decrease in viable cells
was noted at 1.0-3.0 µM of 2ME2 in most of the cell lines. IC50
of 2EM2 for LR-5 cells was 0.5-1.0 µM. We next examined the
effect of 2ME2 on apoptosis in these cells. PI staining
demonstrated a dose-dependent increase in the percentage of
apoptotic (Sub-G1) cells in response to 2ME2 (P < 0.001);
however, estrogen (E2) alone did not induce apoptosis in these
cells (data not shown). This finding is consistent with our
prior study 9, showing that exposure to E2 does not affect growth
kinetics or induce apoptosis of these cells.
Effect of 2ME2 on normal lymphocytes. Normal lymphocytes
from five healthy donors were also treated with various does (0.5
µM, 1.0 µM, 3.0 µM, 10 µM, and 20 µM) of 2ME2 and analyzed for
both inhibition of proliferation and apoptosis by BrdUrd and PI
staining, respectively. In contrast to MM cells, survival of
normal lymphocytes from five healthy donors was not altered
significantly (P = 0.23 from J-T trend test), even at higher
doses (3-20 µM) of 2ME2 (Fig 1C). Moreover, no significant (P =
0.31) apoptosis of normal lymphocytes was observed in response to
2ME2 (data not shown). Taken together, these findings indicate
that 2ME2 has selective anti-MM activity.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

13
Fig. 1. Selective cytotoxicity of 2ME2 against human MM cells. (A) MM.1S cells
were treated with 2ME2 (3 µM) for the indicated times and analyzed for apoptosis by
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

14
both DNA fragmentation (upper panel is a representative of three separate experiments
with similar results) and flow cytometric analysis of PI- and HO+ apoptotic cells
(lower panel is the mean + SD from three independent experiments, P < 0.005). (B) MTT
assay were performed after incubation of MM cell lines (MM.1S --∆--; RPMI-8226 --Ο---;
LR-5 --♦--; Dox-6 --◊--; Dox-40 --�--; MM.1R --�--) with the indicated doses of 2ME2
for 72h. Results are mean + S.D from five independent experiments, P < 0.0001 for all
cell lines. (C) Effect of treatment with 2ME2 (0-20 µM) for 72h on normal lymphocytes
viability, assessed by MTT assay. Results are the mean + SD of five independent
experiments; P = 0.23 from Jonchkeere-Tepstra (J-T) test for trend. (D) MM cells
(CD138+) from five patients (#1-5) were treated with 2ME2 (9 µM) for 72h, followed by
BrdUrd assay. Values are the mean + SD of triplicate samples, P = 0.06; experiments
were repeated three times with similar results. (E) 2ME2 induces proteolytic cleavage
of PARP in patient MM cells. CD138+ cell from 2 MM patients and normal lymphocytes
from 2 healthy donors were treated with 2ME2 (10 µM) for 72h, harvested, and total
protein lysates were subjected to SDS-PAGE analysis. Immunoblot analysis of the
lysates was performed with anti-PARP Ab. 'FL' indicates 'full length' and 'CF' denotes
'cleaved fragment'.
Effect of 2ME2 on patient MM cells. We next determined the
effects of 2ME2 on MM cells freshly isolated from patients
relapsing after multiple prior therapies including Dexamethasone,
melphalan and thalidomide. Tumor cells were purified from bone
marrow aspirates by CD138 positive selection using CD138
(Syndecan-1) Micro Beads and Auto MACS magnetic cell sorting.
Treatment of MM cells from 5 patients (Patients # 1-5), with 2ME2
(9 µM) significantly (P = 0.06) decreased growth, as measured by
BrdUrd incorporation (Fig 1D). In order to further determine
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

15
whether growth inhibition correlates with apoptosis in patient MM
cells, we next determined the effects of 2ME2 (9 µM) on poly
(ADP-ribose) polymerase (PARP) cleavage, a known signature event
during apoptosis 30,31. 2ME2 (9 µM) induced PARP cleavage in
patient MM cells, consistent with apoptosis, but failed to induce
PARP cleavage in normal lymphocytes (Fig 1E). These findings
correlate with our earlier results (Fig 1C) as well as other
study 32 demonstrating lack of cytotoxicity of 2ME2 (9 µM) on
normal lymphocytes. Taken together, our results suggest that
2ME2 inhibits growth and induces apoptosis in patient MM cells.
Effect of 2ME2 on patient MM-derived Bone Marrow Stromal
Cells (BMSCs). In MM, tumor cells are predominantly localized in
the BM microenvironment due to their adherence both to
extracellular matrix proteins and to BMSCs. This interaction
between MM cells and BMSCs triggers production of cytokines
mediating autocrine and paracrine growth and survival of MM
cells, as well as protection against drug-induced apoptosis 33.
We therefore next examined the effect of 2ME2 on patient MM-
derived BMSCs from 5 MM patients. Treatment of BMSCs with 2ME2
significantly (Slope= -5.5, P = 0.002) decreases the viability
(Fig 2A, 3 Patients A-C are shown). In order to determine
whether 2ME2-induced growth inhibition correlates with apoptosis
in
patient BMSCs, we next examined the effects of 2ME2 (9 µM) on
PARP cleavage. 2ME2 (3 µM) induced PARP cleavage, consistent
with apoptosis (Fig 2B, patient A and B). Taken together, these
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

16
results suggest that 2ME2 acts not only directly on MM cells, but
also affects the BM microenvironment.
Fig. 2. Effect of 2ME2 on BMSCs and IL-6 secretion (A) Patient MM-derived BMSCs
(Patients A-C) were treated with 2ME2 (3-9 µM) for 72h, and viability was assessed by
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

17
MTT assay. Results are the mean + SD from triplicate samples, P = 0.002. (B) 2ME2
induce proteolytic cleavage of PARP in patient MM-BMSCs cells. BMSCs from 2MM
patients were treated with 2ME2 (3 µM) for 72h, and harvested; total protein lysates
were subjected to SDS-PAGE analysis. Immunoblot analysis of the lysates was performed
with anti-PARP antibody. 'FL' indicates 'full length' and 'CF' denotes 'cleaved
fragment. (C) Effect of 2ME2 on MM cell adhesion-induced IL-6 secretion in BMSCs. IL-
6 levels were measured using IL-6 specific ELISA in supernatants of 24h cultures of
BMSCs, MM.1S cells, and BMSCs + MM.1S cells, in the presence or absence of 2ME2 (3
µM).
Effect of 2ME2 on Interleukin-6 (IL-6) secretion in BMSCs.
Adhesion of MM cells to BMSCs trigger secretion of factors such
as, IL-6, from BMSCs, which regulate the growth of MM cells in
autocrine or paracrine manner 34. In order to determine whether
2ME2 also affect secretion of IL-6, we next examined the effect
of 2ME2 on IL-6 production in the BM microenvironment. As seen
in Fig 2C, 2ME2 significantly (P < 0.005) inhibits IL-6 secretion
in MM patient BMSCs triggered by MM cell adhesion (Fig 2C).
Reports that high serum levels of IL-6 contribute to clinical
chemo resistance and treatment failure 35, coupled with our
demonstration that 2ME2 decreases the MM adhesion-induced IL-6
secretion from BMSCs, suggests that 2ME2 may overcome drug
resistance in patients with advanced MM.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom
18
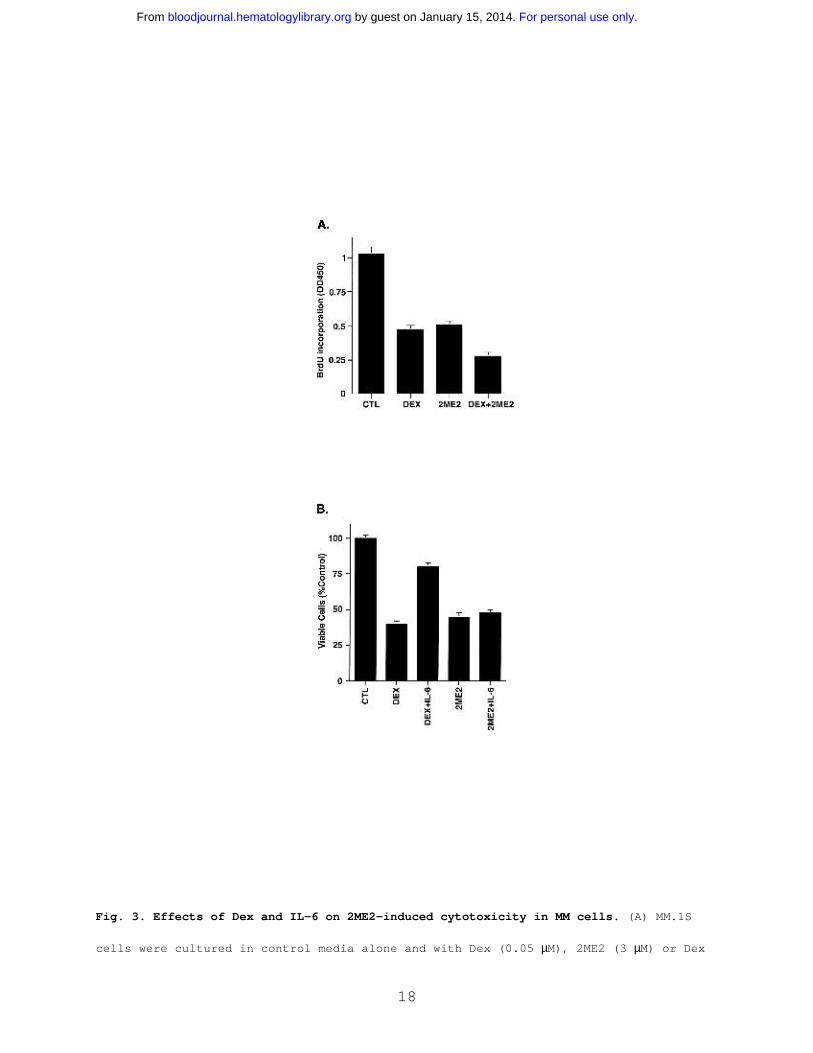
Fig. 3. Effects of Dex and IL-6 on 2ME2-induced cytotoxicity in MM cells. (A) MM.1S
cells were cultured in control media alone and with Dex (0.05 µM), 2ME2 (3 µM) or Dex
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

19
+ 2ME2. At 48 h, cells were harvested and analyzed by BrdUrd assay. Results are mean
+ SD of three independent experiments (P < 0.005). (B) MM.1S cells were treated with
2ME2 (3 µM) or Dex (0.05 µM), in the presence or absence of IL-6 (10 ng/ml). At 48 h,
cells were harvested and viability analyzed by MTT assay. The median viability was
41% for Dex and 78% for Dex + IL-6 (P = 0.05, as determined by one-sided Wilcoxon
rank-sum test), whereas for 2ME2 the median viability was 45% with 2ME2 alone and 48%
with 2ME2 + IL-6 (P = 0.20, Wilcoxon test, as above). Results are mean + SD of three
independent experiments.
Effect of Dex and IL-6 on response of MM cells to 2ME2. To
determine whether the effects of 2ME2 are additive with
conventional therapies, we next examined whether Dex (0.0005- 0.5
µM) enhanced 2ME2 (3 µM) effects on proliferation of Dex-
sensitive (MM.1S) MM cells. As seen in Fig 3A, the combination
of 2ME2 and Dex induces more growth inhibition as compared to the
treatment of MM.1S cells with 2ME2 alone. Dex (0.05 µM) and 2ME2
(3 µM) increased growth inhibition by 30%, relative to cultures
with 2ME2 alone (P = 0.05, one-sided Wilcoxon rank-sum test).
PI staining after Dex (0.05 µM) and 2ME2 (3 µM) revealed 93 +
6.1% apoptosis versus 66 + 4.3% cell death induced by Dex alone.
We and others have shown that IL-6 protects against Dex-
induced apoptosis 36-39, and we therefore next examined the
effect of IL-6 on 2ME2-induced apoptosis. MM.1S cells were
treated with 2ME2 (3 µM) or Dex (0.05 µM ), in the presence and
absence of IL-6 (10 ng/ml). As seen in Fig 3B, the median
viability was 41% for Dex and 78% for Dex + IL-6 (P = 0.05, as
determined by one-sided Wilcoxon rank-sum test), whereas for 2ME2
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

20
the median viability was 45% with 2ME2 alone and 48% with 2ME2 +
IL-6 (P = 0.20, Wilcoxon test, as above). These findings suggest
that IL-6 fail to abrogate effects of 2ME2 on MM.1S cells. In
contrast and as in our prior studies 38, IL-6 blocks Dex-induced
decreased in MM.1S cell viability. Taken together, our data
suggest that 2ME2 overcomes the growth and protective effects of
IL-6 on MM cells and also indicate distinct mechanisms of action
for 2ME2 and Dex against MM cells.
2ME2 induces mitochondrial release of cytochrome-c and Smac.
Having shown the biologic sequelae of 2ME2 in MM cells, we next
examined the signaling mechanisms mediating apoptosis triggered
by 2ME2. Our prior studies have characterized Dex-induced
apoptotic mechanisms in MM.1S MM cells, and we therefore studied
2ME2 effects using these cells in order to compare differential
signaling cascades triggered by Dex versus 2ME2. Previous
studies have shown that 2ME2-induced apoptosis involves
mitochondrial damage 32, we first therefore determined whether
treatment of MM.1S cells is associated with the release of
mitochondrial apoptogenic proteins, including cytochrome-c (cyto-
c) and second mitochondrial activator of caspases (Smac). Our
recent studies have demonstrated that Smac, but not cyto-c, is
released from mitochondria into the cytosol during Dex-induced
apoptosis in MM.1S cells 24,40. To determine whether 2ME2-
induced apoptosis in MM.1S cells is associated with the
mitochondrial release of cyto-c and/or Smac, we treated MM.1S
cells with 2ME2 for various times and analyzed cytosolic and
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

21
mitochondrial extracts for levels of cyto-c and Smac. As seen in
Fig. 4A and 4B (upper panels), 2ME2 increase cyto-c and Smac
levels in cytosol at 48 h. Densitometric analysis of the
immunoblot demonstrated that 2ME2 induces a 4-5 fold increase in
the cytosolic cyto-c and Smac levels, as compared to untreated
cells (Fig 4A and 4B). The increase in cyto-c and Smac levels
was associated with a concomitant decrease in mitochondrial Smac
or cyto-c levels (data not shown). Moreover, the 2ME2-induced
increases in cytosolic cyto-c or Smac levels were specific, since
no changes were observed in levels of cytosolic SHP2 (Fig 4A and
4B, lower panels). These results suggest that 2ME2-induced
apoptosis is accompanied by accumulation of both Smac and cyto-c
in the cytosol. In contrast, and as in our prior studies 24,
Dex-induced apoptosis was associated with the mitochondrial
release of Smac, but not of cyto-c (Fig 4A and 4B).
To determine whether 2ME2 also induces cyto-c or Smac
release in other MM cell lines, we treated Dox-40, MM.1R and LR-5
MM cells with 2ME2 (3 µM) for 48h, followed by analysis of
cytosolic extracts for levels of cyto-c and Smac. As seen in
Fig. 4C and 4D (upper panels), 2ME2 also triggers accumulation of
cyto-c and Smac levels in the cytosol of Dox-40 and LR-5 MM
cells. Similar results were obtained using MM.1R MM cell line
(data not shown). 2ME2-induced increases in cytosolic cyto-c or
Smac levels were specific, since no changes were observed in
levels of cytosolic SHP2 (Fig 4C and 4D, lower panels). Taken
together, these results suggest that 2ME2-induced apoptosis is
accompanied by accumulation of both Smac and cyto-c in the
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

22
cytosol.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

23
Fig. 4. 2ME2 induces mitochondrial release of cytochrome-c (A) and Smac (B).
MM.1S cells were treated with 2ME2 (3 µM) or Dex (0.05••M) and harvested at 48h.
Cytosolic proteins were separated by 12.5% SDS-PAGE and analyzed by immunoblotting
with anti-cyto-c (A, upper panel) or anti-Smac (B, upper panel) Abs. As a control for
equal loading of proteins, filters were also reprobed with anti-SHP2 Ab (A, and B,
lower panels). Blots are representative of three independent experiments. (C) and
(D) Dox-40 and LR-5 MM cells were treated with 2ME2 (3 µM) and harvested at 48h.
Cytosolic proteins were separated by 12.5% SDS-PAGE and analyzed by immunoblotting
with anti-cyto-c (C, upper panels) or anti-Smac (D, upper panel) Abs. As a control
for equal loading of proteins, filters were reprobed with anti-SHP2 Ab (C, and D,
lower panels). Blots are representative of three independent experiments.
Densitometric analysis of the immunoblot demonstrated that 2ME2 induces a 4-5 fold
increase in the cytosolic cyto-c and Smac levels, as compared to untreated cells.
2ME2-induced apoptosis is mediated by activation of caspase
cascade. Smac promotes caspase activation via the cyto-c/Apaf-
1/caspase-9 pathway 41, and our prior study in MM cells
demonstrated that Dex triggers release of Smac from mitochondria
into cytosol, with sequential activation of caspase-9 and
caspase-3 40. To examine whether 2ME2-induced Smac release and
apoptosis in MM cells is associated with activation of caspase-9,
cytosolic extracts from 2ME2-treated MM.1S cells were subjected
to immunoblot analysis using anti-caspase-9 antibody. As seen in
Fig. 5A, treatment of MM.1S cells with 2ME2 (3 µM) induces
proteolytic cleavage of procaspase-9 into 37 and 35 KDa
fragments. We also assayed for catalytic activity of caspase-9,
using LEHD-pNA conjugated substrate in a colorimetric protease
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

24
assay 40. Incubation of cytosolic extracts from 2ME2-treated
MM.1S cells with LEHD-pNA was associated with cleavage of LEHD-
pNA (data not shown). Taken together, these findings demonstrate
that treatment of MM.1S cells with 2ME2 is associated with
activation of caspase-9.
Multiple studies have shown that upstream events mediating
mitochondrial cyto-c release induce sequential activation of
caspase-8, cleavage of Bid, translocation of cleaved Bid from the
cytoplasm into mitochondria, and release of cyto-c 42,43. Since
2ME2 induces release of cyto-c, we next assayed for activation of
caspase-8 by 2ME2. As seen in Fig. 5B, 2ME2 (3 µM) also triggers
proteolytic cleavage of caspase-8. In contrast, and as in our
prior study 40, Dex-induced apoptosis is not associated with
caspase-8 activation (Fig 5B).
Previous studies have demonstrated that both caspase-8 and -
9 activate downstream apoptotic caspase-3 (CPP32) 44.
Furthermore, in a cell free system, the addition of purified
cyto-C to cyto-c-depleted extracts activates caspase-3 and DNA-
fragmentation 45. Since treatment of MM.1S cells with 2ME2
activates both caspase-8 and -9, we next determined whether
caspase-3 is cleaved and activated in response to 2ME2. MM.1S
cells were treated with 2ME2 (3 µM), and cell lysates were
subjected to immunoblotting with either caspase-3 or its known
substrate PKC δ 24. 2ME2 induces caspase-3 cleavage and
activation, as evidenced by cleavage of caspase-3 (Fig 5C and
data not shown). Dex treatment served as a positive control.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

25
Fig 5. Delineation of 2ME2 and Dex-induced caspase cascade. 2ME2 induces
activation of caspase-9 (A) and -8 (B). MM.1S cells were treated with 2ME2 (3 µM) and
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

26
Dex (0.05•µM) and harvested at 48h. Cytosolic proteins were separated by 12.5% SDS-
PAGE and analyzed by immunoblotting with anti-caspase-8 (cas-8) and -9 (Cas-9) Abs.
Blots are representative of three independent experiments. (C) Cleavage of caspase-3
induced by 2ME2. MM.1S cells were treated with 2ME2 (3 µM) and Dex (0.05•µM), and
harvested at 48h. Total cell lysates were analyzed by immunoblotting with anti-
caspase-3 Ab. 'FL' indicates 'full length' and 'CF' denotes 'cleaved fragment'. (D)
2ME2 and Dex-induced differential signaling cascades in MM.
Collectively, a comparative analyses of 2ME2 and Dex-induced
apoptotic signaling cascades therefore suggests that 2ME2, in
contrast to Dex, triggers multiple signaling pathways (Fig. 5D).
For example, 2ME2 induces release of both cyto-c and Smac,
whereas Dex triggers the release of only Smac; 2ME2 activates
both caspase-8 and -9, whereas Dex triggers activation of only
caspase-9; and finally, IL-6 protects against Dex-, but not 2ME2-
induced apoptosis. Since 2ME2 triggers apoptosis in Dex-
resistant MM cells, elucidating the 2ME2-induced apoptotic
signaling pathways may not only identify additional apoptotic
targets, but also delineate abnormalities in signaling pathways
which confer Dex-resistance.
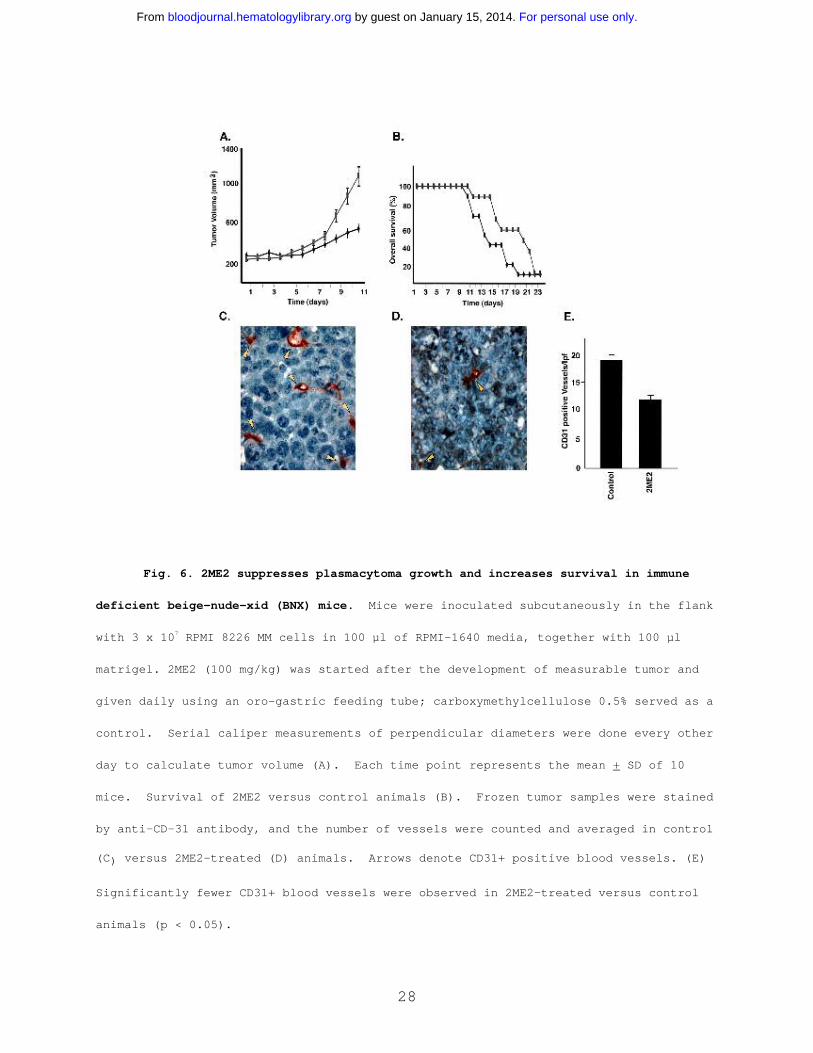
2ME2 suppresses in vivo tumor growth in Scid-mouse model.
Having shown the signaling mechanisms mediating the anti-MM
effects of 2ME2 in vitro, we next determined whether these in
vitro effects correlate with in vivo activity of 2ME2, using our
Scid-mouse model. Immune deficient beige-nude-xid (BNX) mice
were inoculated subcutaneously in the flank with 3x107 RPMI 8226
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

27
MM cells in 100 µl of RPMI-1640 media, together with 100 µl
matrigel. Daily oral administration of 2ME2 (100mg/kg) starting
after the development of measurable tumor significantly reduced
MM tumor growth (35-40%, P < 0.05)(Fig 6A) and increased survival
(Fig 6B), compared with the control group treated with the
vehicle (Matrigel) only. No significant toxicity, as evidenced
by lack of weight changes, was observed in any treatment groups.
These findings suggest that 2ME2 at molar concentrations is
effective in controlling growth of human MM in mice. Other in
vivo preclinical studies have shown that 2ME2 is also effective
in reducing tumor burden in Meth-A sarcoma or B16 melanoma; in
non estrogen-dependent human breast carcinoma (MDA-MB-435); and
in human pancreatic (MIA PaCa-2) xenograft mouse models 2,14.
Since 2ME2 is a known anti-angiogenic agent and recent
reports show increased BM angiogenesis in MM 15-17, we next
examined the effect of 2ME2 on intratumoral microvessel density
(MVD), evidenced by CD-31 immunostaining of human MM tumors in
mice. A significant decrease in MVD was noted in 2ME2-treated
animals (Fig 6D) versus the cohort treated with control vehicle
alone (Fig 6C). The number of CD-31+ blood vessels per low power
(400X) field was 10.30 + 1.0 in 2ME2-treated versus 19.10 + 1.9
in control groups (P < 0.05) (Fig 6E). Taken together, these
studies demonstrate that 2ME2 is effective in inhibiting both
tumor cell growth and angiogenesis in MM.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom
28
Fig. 6. 2ME2 suppresses plasmacytoma growth and increases survival in immune
deficient beige-nude-xid (BNX) mice. Mice were inoculated subcutaneously in the flank
with 3 x 107 RPMI 8226 MM cells in 100 µl of RPMI-1640 media, together with 100 µl
matrigel. 2ME2 (100 mg/kg) was started after the development of measurable tumor and
given daily using an oro-gastric feeding tube; carboxymethylcellulose 0.5% served as a
control. Serial caliper measurements of perpendicular diameters were done every other
day to calculate tumor volume (A). Each time point represents the mean + SD of 10
mice. Survival of 2ME2 versus control animals (B). Frozen tumor samples were stained
by anti-CD-31 antibody, and the number of vessels were counted and averaged in control
(C) versus 2ME2-treated (D) animals. Arrows denote CD31+ positive blood vessels. (E)
Significantly fewer CD31+ blood vessels were observed in 2ME2-treated versus control
animals (p < 0.05).
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

29
Having shown in vivo effects of 2ME2 on angiogenesis, we
next determined whether 2ME2 altered VEGF secretion in the BM
microenvironment. 2ME2 induces a significant (P < 0.05) decrease
in VEGF secretion in MM.1S cells (34 + 2.1% decrease, n=3) and in
BMSCs (41 + 3.2% decrease, n=3). Our results are consistent with
other study suggesting that VEGF expression, as evaluated by
immunohistochemical analysis, is down regulated concomitant with
tumor angiogenic suppression. 46. Since we and others have
recently demonstrated that VEGF is not only an angiogenic factor,
but also triggers growth and migration in MM cells 16,17, our
present findings suggest an additional mechanism whereby 2ME2
mediates anti-MM activity.
Our study therefore demonstrates that 2ME2 can both directly
induce apoptosis in MM cells and act in the BM microenvironment
to decrease cytokines mediating tumor cell growth and survival,
as well as angiogenesis. These studies provide the framework for
clinical trials of 2ME2 to overcome drug resistance and improve
patient outcome in MM.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

30
Acknowledgements
We thank Dr. Xiaodong Wang for providing Smac related
reagents. The authors gratefully acknowledge the technical
assistance of Kamal Chauhan.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

31
References
1. Cushman M, He H-M, Katzenellenbogen J-A, Lin C-M, Hamel E.
Synthesis, antitubulin and antimitotic activity, and cytotoxicity
of analogs of 2-methoxyestradiol, an endogenous mammalian
metabolite of estradiol that inhibits tubulin polymerization by
binding to the colchicine binding site. J Med Chem. 1995;38:2041-
2049.
2. Klauber N, Parangi S, Flynn E, Hamel E, D'Amato R-J.
Inhibition of angiogenesis and breast cancer in mice by the
microtubule inhibitors 2-methoxyestradiol and taxol. Cancer Res.
1997;57:81-86.
3. Lottering M-L, Haag M, Seegers J-C. Effects of 17 beta-
estradiol metabolites on cell cycle events in MCF-7 cells. Cancer
Res. 1992;52:5926-5932.
4. D'Amato R-J, Lin C-M, Flynn E, Folkman J, Hamel E. 2-
Methoxyestradiol, an endogenous mammalian metabolite, inhibits
tubulin polymerization by interacting at the colchicine site.
Proc Natl Acad Sci U S A. 1994;91:3964-3968.
5. Mukhopadhyay T, Roth JA. Induction of apoptosis in human lung
cancer cells after wild-type p53 activation by methoxyestradiol.
Oncogene. 1997;14:379-384.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

32
6. Schumacher G, Neuhaus P. The physiological estrogen metabolite
2-methoxyestradiol reduces tumor growth and induces apoptosis in
human solid tumors. J Cancer Res Clin Oncol. 2001;127:405-410.
7. Kumar A-P, Garcia G-E, Slaga T-J. 2-methoxyestradiol blocks
cell-cycle progression at G(2)/M phase and inhibits growth of
human prostate cancer cells. Mol Carcinog. 2001;31:111-124.
8. Danel L, Vincent C, Rousset F, et al. Estrogen and
progesterone receptors in some human myeloma cell lines and
murine hybridomas. J Steroid Biochem. 1988;30:363-367
9. Treon SP, Teoh G, Urashima M, et al. Anti-estrogens induce
apoptosis of multiple myeloma cells. Blood. 1998;92:1749-1757.
10. Otsuki T, Yamada O, Kurebayashi J, et al. Estrogen receptors
in human myeloma cells. Cancer Res. 2000;60:1434-1441.
11. Wang L-H, Yang X-Y, Mihalic K, Xiao W, Li D, Farrar W-L.
Activation of estrogen receptor blocks interleukin-6-inducible
cell growth of human multiple myeloma involving molecular cross-
talk between estrogen receptor and STAT3 mediated by co-regulator
PIAS3. J Biol Chem. 2001;276:31839-31844.
12. Maran A, Zhang M, Kennedy AM, et al. -2-methoxyestradiol
induces interferon gene expression and apoptosis in osteosarcoma
cells. Bone. 2002;30:393-398.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

33
13. Merriam G-R, MacLusky N-J, Picard M-K, Naftolin F.
Comparative properties of the catechol estrogens, I: methylation
by catechol-O-methyltransferase and binding to cytosol estrogen
receptors. Steroids. 1980;36:1-11.
14. Fotsis T, Zhang Y, Pepper MS, et al. The endogenous oestrogen
metabolite 2-methoxyoestradiol inhibits angiogenesis and
suppresses tumour growth. Nature. 1994;368:237-239.
15. Ribatti D, Vacca A, Nico B, et al. Bone marrow angiogenesis
and mast cell density increase simultaneously with progression of
human multiple myeloma. Br J Cancer. 1999;79:451-455.
16. Podar K, Tai YT, Davies FE, et al. Vascular endothelial
growth factor triggers signaling cascades mediating multiple
myeloma cell growth and migration. Blood. 2001;98:428-435.
17. Vacca A, Ribatti D, Presta M, et al. Bone marrow
neovascularization, plasma cell angiogenic potential, and matrix
metalloproteinase-2 secretion parallel progression of human
multiple myeloma. Blood. 1999;93:3064-3073.
18. Moalli P-A, Pillay S, Weiner D, Leikin R, Rosen S-T. A
mechanism of resistance to glucocorticoids in multiple myeloma:
transient expression of a truncated glucocorticoid receptor mRNA.
Blood. 1992;79:213-222
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

34
19. Damiano J-S, Cress A-E, Hazlehurst L-A, Shtil A-A, Dalton W-
S. Cell adhesion mediated drug resistance (CAM-DR): Role of
integrins and resistance to apoptosis in human myeloma cell
lines. Blood. 1999;93:1658-1667
20. Maeshima Y, Manfredi M, Reimer C, et al. Identification of
the anti-angiogenic site within vascular basement membrane-
derived tumstatin. J Biol Chem. 2001;276:15240-15248.
21. Chauhan D, Uchiyama H, Akbarali Y, et al. Multiple myeloma
cell adhesion-induced interleukin-6 expression in bone marrow
stromal cells involves activation of NF-kappa B. Blood.
1996;87:1104-1112.
22. Hideshima T, Chauhan D, Shima Y, et al. Thalidomide and its
analogues overcome drug resistance of human multiple myeloma
cells to conventional therapy. Blood. 2000;96:2943-2950
23. Gupta D, Treon SP, Shima Y, et al. Adherence of multiple
myeloma cells to bone marrow stromal cells upregulates vascular
endothelial growth factor secretion: therapeutic applications.
Leukemia. 2001;15:1950-1961.
24. Chauhan D, Pandey P, Ogata A, et al. Cytochrome-c dependent
and independent induction of apoptosis in multiple myeloma cells.
J Biol Chem. 1997;272:29995-29997
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

35
25. Desnoyers S, Shah G-M, Brochu G, Poirier G-G. Erasable blot
of poly(ADP-ribose) polymerase. Analytical Biochemistry.
1994;218:470-473
26. Chauhan D, Hideshima T, Pandey P, et al. RAFTK/PYK2-dependent
and -independent apoptosis in multiple myeloma cells. Oncogene.
1999;18:6733-6740.
27. Chauhan D, Pandey P, Hideshima T, et al. SHP2 mediates the
protective effect of interleukin-6 against dexamethasone-induced
apoptosis in multiple myeloma cells. J Biol Chem. 2000;275:27845-
27850.
28. Weidner N, Semple JP, Welch WR, Folkman J. tumor angiogenesis
and metastasis - correlation in invasive breast carcinoma. N Engl
J Med. 1991;324:1-8
29. Wyllie A-H. Chromatin cleavage in apoptosis: association
with condensed chromatin morphology and dependence on
macromolecular synthesis. J. Pathol. 1984;142:66-77
30. Lazebnik Y-A, Kaufmann S-H, Desnoyers S, Poirier G-G,
Earnshaw W-C. Cleavage of poly(ADP-ribose) polymerase by a
proteinase with properties like ICE. Nature. 1994;371:346-347.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

36
31. Aoufouchi S, Yelamos J, Milstein C. Inhibition of apoptosis
of a PARP(-)/(-)cell line transfected with PARP DNA-binding
domain mutants. J Mol Biol. 1999;290:943-949.
32. Huang P, Feng L, Oldham E-A, Keating M-J, Plunkett W.
Superoxide dismutase as a target for the selective killing of
cancer cells. Nature. 2000;407:390-395.
33. Anderson K-C. Targeted therapy for multiple myeloma. Semin
Hematol. 2001;38:286-294.
34. Anderson K-C. Novel biologically based therapies for myeloma.
Cancer J. 2001;7 Suppl 1:S19-23.
35. Kyrstsonis M-C, Dedoussis G, Baxevanis C, Stamatelou M,
Maniatis A. Serum interleukin-6 (IL-6) and interleukin-4 (IL-4)
in patients with multiple myeloma. Br J Haematol. 1996;92:420-422
36. Hardin J, MacLeod S, Grigorieva I, -et al. Interleukin-6
prevents dexamethasone-induced myeloma cell death. Blood.
1994;84:3063-3070.
37. Rowley M, Liu P, Van Ness B. Heterogeneity in therapeutic
response of genetically altered myeloma cell lines to interleukin
6, dexamethasone, doxorubicin, and melphalan. Blood.
2000;96:3175-3180.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

37
38. Chauhan D, Kharbanda S, Ogata A, et al. Interleukin-6
inhibits Fas-induced apoptosis and stress-activated protein
kinase activation in multiple myeloma cells. Blood. 1997;89:227-
234
39. Chen Y-H, Desai P, Shiao R-T, Lavelle D, Haleem A, Chen J.
Inhibition of myeloma cell growth by dexamethasone and all-trans
retinoic acid: synergy through modulation of interleukin-6
autocrine loop at multiple sites. Blood. 1996;87:314-323.
40. Chauhan D, Hideshima T, Rosen S, Reed J-C, Kharbanda S,
Anderson K-C. Apaf-1/cytochrome c-independent and Smac-dependent
induction of apoptosis in multiple myeloma (MM) cells. J Biol
Chem. 2001;276:24453-24456.
41. Du C, Fang M, Li Y, Li L, Wang X. Smac, a mitochondrial
protein that promotes cytochrome c-dependent caspase activation
by eliminating IAP inhibition. Cell. 2000;102:33-42.
42. Li H, Zhu H, Xu C-J, Yuan J. Cleavage of BID by caspase 8
mediates the mitochondrial damage in the Fas pathway of
apoptosis. Cell. 1998;94:491-501.
43. Kuwana T, Smith J-J, Muzio M, Dixit V, Newmeyer D-D,
Kornbluth S. Apoptosis induction by caspase-8 is amplified
through the mitochondrial release of cytochrome c. J Biol Chem.
1998;273:16589-16594.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom

38
44. Chinnaiyan A-M, Dixit V-M. The cell-death machine. Curr.
Biol. 1996;6:555-562
45. Liu X, Naekyung Kim C, Yang J, Jemmerson R, Wang X. Induction
of apoptotic program in cell-free extracts: requirement for dATP
and cytochrome c. Cell. 1996;86:147-157
46. Banerjee S-K, Zoubine M-N, Sarkar D-K, Weston A-P, Shah J-H,
Campbell D-R. 2-Methoxyestradiol blocks estrogen-induced rat
pituitary tumor growth and tumor angiogenesis: possible role of
vascular endothelial growth factor. Anticancer Res. 2000;20:2641-
2645.
For personal use only. by guest on January 15, 2014. bloodjournal.hematologylibrary.orgFrom




















