
2012 Ternianov et al Neurobiol Aging Oct 25
16
Overexpression of CB2 cannabinoid receptors results in neuroprotection against behavioral and neurochemical alterations induced by intracaudate administration of 6-hydroxydopamine Alexander Ternianov a , José M. Pérez-Ortiz a,b , María E. Solesio a , María S. García-Gutiérrez b , Antonio Ortega-Álvaro a , Francisco Navarrete b , Carlos Leiva c , María F. Galindo a , Jorge Manzanares a,b, * a Unidad de Neuropsicofarmacología Traslacional, Complejo Hospitalario Universitario de Albacete, Albacete, Spain b Instituto de Neurociencias, Universidad Miguel Hernández-CSIC, San Juan de Alicante, Alicante, Spain c Department of Neurology, Hospital General de Alicante, Alicante, Spain Received 5 March 2010; received in revised form 7 September 2010; accepted 14 September 2010 Abstract The role of CB2 cannabinoid receptors in the behavioral and neurochemical changes induced by intracaudate administration of 6-hydroxydopamine (6-OHDA) was evaluated. 6-OHDA (12 g/4 L) or its vehicle was injected in the caudate-putamen (CPu) of mice overexpressing the CB2 cannabinoid receptor (CB2xP) and wild type (WT) mice. Motor impairment, emotional behavior, and cognitive alterations were evaluated. Tyrosine hydroxylase (TH), glial fibrillary acidic protein (GFAP), and ionized calcium-binding adapter molecule 1 (Iba-1) were measured by immunocytochemistry in the CPu and/or substantia nigra (SN) of CB2xP mice and WT mice. Oxidative/ nitrosative and neuroinflammatory parameters were also measured in the CPu and cortex of 6-OHDA-treated and sham-treated mice. 6-OHDA-treated CB2xP mice presented significantly less motor deterioration than 6-OHDA-treated WT mice. Immunocytochemical analysis of tyrosine hydroxylase in the SN and CPu revealed significantly fewer lesions in CB2xP mice than in WT mice. GFAP and Iba-1 immunostaining revealed less astrocyte and microglia recruitment to the treated area of the CPu in CB2xP mice. Malonyldialdehyde (MDA) concentrations were lower in the striatum and cerebral cortex of sham-treated CB2xP mice than in sham-treated WT mice. The administration of 6-OHDA increased MDA levels in both WT mice and CB2xP mice; it increased the oxidized (GSSG)/reduced (GSH) glutathione ratio in the striatum in WT mice alone compared with matched sham-treated controls. The results revealed that overexpression of CB2 cannabinoid receptors decreased the extent of motor impairment and dopaminergic neuronal loss, reduced the recruitment of astrocytes and microglia to the lesion, and decreased the level of various oxidative parameters. These results suggest that CB2 receptors offer neuroprotection against dopaminergic injury. © 2010 Elsevier Inc. All rights reserved. Keywords: Parkinson’s disease; 6-OHDA; Animal model; Neuroprotection; CB2; Cannabinoids 1. Introduction Study of the endogenous cannabinoid system has re- sulted in the identification of 2 receptors (CB1 and CB2) and 2 main families of endogenous ligands (anandamide and 2-arachidonoyl-glycerol) (Mechoulam et al., 1998), with the corresponding metabolizing enzymes (fatty acid amide hydrolase, and monoacylglycerol lipase) of these ligands (Giuffrida et al., 2001), and a re-uptake system. CB1 cannabinoid receptor distribution was described in rat brain almost 20 years ago (Herkenham et al., 1990). CB1 recep- tors are localized mainly in the central nervous system and are abundant in the basal ganglia, hippocampus, cerebellum, and cortical areas (Herkenham et al., 1991). Furthermore, * Corresponding author at: Universidad Miguel Hernández, Instituto de Neurociencias, Avda. Ramón y Cajal s/n, 03550 San Juan de Alicante, Spain. Tel.: 34 96 591 9248; fax: 34 96 5919549. E-mail address: [email protected] (J. Manzanares). Neurobiology of Aging xx (2010) xxx www.elsevier.com/locate/neuaging 0197-4580/$ – see front matter © 2010 Elsevier Inc. All rights reserved. doi:10.1016/j.neurobiolaging.2010.09.012
-
Upload
hospitalesperitsant -
Category
Documents
-
view
1 -
download
0
Transcript of 2012 Ternianov et al Neurobiol Aging Oct 25

A
6oa1n6aicagoan©
K
1
sa
NS
Neurobiology of Aging xx (2010) xxx
0d
Overexpression of CB2 cannabinoid receptors results in neuroprotectionagainst behavioral and neurochemical alterations induced by
intracaudate administration of 6-hydroxydopamine
Alexander Ternianova, José M. Pérez-Ortiza,b, María E. Solesioa, María S. García-Gutiérrezb,Antonio Ortega-Álvaroa, Francisco Navarreteb, Carlos Leivac, María F. Galindoa,
Jorge Manzanaresa,b,*a Unidad de Neuropsicofarmacología Traslacional, Complejo Hospitalario Universitario de Albacete, Albacete, Spain
b Instituto de Neurociencias, Universidad Miguel Hernández-CSIC, San Juan de Alicante, Alicante, Spainc Department of Neurology, Hospital General de Alicante, Alicante, Spain
Received 5 March 2010; received in revised form 7 September 2010; accepted 14 September 2010
bstract
The role of CB2 cannabinoid receptors in the behavioral and neurochemical changes induced by intracaudate administration of-hydroxydopamine (6-OHDA) was evaluated. 6-OHDA (12 �g/4 �L) or its vehicle was injected in the caudate-putamen (CPu) of miceverexpressing the CB2 cannabinoid receptor (CB2xP) and wild type (WT) mice. Motor impairment, emotional behavior, and cognitivelterations were evaluated. Tyrosine hydroxylase (TH), glial fibrillary acidic protein (GFAP), and ionized calcium-binding adapter molecule(Iba-1) were measured by immunocytochemistry in the CPu and/or substantia nigra (SN) of CB2xP mice and WT mice. Oxidative/
itrosative and neuroinflammatory parameters were also measured in the CPu and cortex of 6-OHDA-treated and sham-treated mice.-OHDA-treated CB2xP mice presented significantly less motor deterioration than 6-OHDA-treated WT mice. Immunocytochemicalnalysis of tyrosine hydroxylase in the SN and CPu revealed significantly fewer lesions in CB2xP mice than in WT mice. GFAP and Iba-1mmunostaining revealed less astrocyte and microglia recruitment to the treated area of the CPu in CB2xP mice. Malonyldialdehyde (MDA)oncentrations were lower in the striatum and cerebral cortex of sham-treated CB2xP mice than in sham-treated WT mice. Thedministration of 6-OHDA increased MDA levels in both WT mice and CB2xP mice; it increased the oxidized (GSSG)/reduced (GSH)lutathione ratio in the striatum in WT mice alone compared with matched sham-treated controls. The results revealed that overexpressionf CB2 cannabinoid receptors decreased the extent of motor impairment and dopaminergic neuronal loss, reduced the recruitment ofstrocytes and microglia to the lesion, and decreased the level of various oxidative parameters. These results suggest that CB2 receptors offereuroprotection against dopaminergic injury.
2010 Elsevier Inc. All rights reserved.
eywords: Parkinson’s disease; 6-OHDA; Animal model; Neuroprotection; CB2; Cannabinoids
www.elsevier.com/locate/neuaging
awalcata
. Introduction
Study of the endogenous cannabinoid system has re-ulted in the identification of 2 receptors (CB1 and CB2)nd 2 main families of endogenous ligands (anandamide
* Corresponding author at: Universidad Miguel Hernández, Instituto deeurociencias, Avda. Ramón y Cajal s/n, 03550 San Juan de Alicante,pain. Tel.: �34 96 591 9248; fax: �34 96 5919549.
aE-mail address: [email protected] (J. Manzanares).
197-4580/$ – see front matter © 2010 Elsevier Inc. All rights reserved.oi:10.1016/j.neurobiolaging.2010.09.012
nd 2-arachidonoyl-glycerol) (Mechoulam et al., 1998),ith the corresponding metabolizing enzymes (fatty acid
mide hydrolase, and monoacylglycerol lipase) of theseigands (Giuffrida et al., 2001), and a re-uptake system. CB1annabinoid receptor distribution was described in rat brainlmost 20 years ago (Herkenham et al., 1990). CB1 recep-ors are localized mainly in the central nervous system andre abundant in the basal ganglia, hippocampus, cerebellum,
nd cortical areas (Herkenham et al., 1991). Furthermore,
tnTcclnps(edhoa
rpCacCdp2n2Am2socptaae
tsccSertk22T
efa
StgCCtpCbtAtam6nscota(flt
2
2
taCpmSeawtcfohmr(o
2
msnA
2 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
he presynaptic localization of CB1 receptors suggests aeuromodulatory role in synaptic neurotransmitter release.his receptor is present in other tissues and in nonneuronalell types such as astrocytes (Bouaboula et al., 1995), mi-roglia (Waksman et al., 1999), and oligodendrocytes (Mo-ina-Holgado et al., 2002). Research on the CB1 cannabi-oid receptor has shown that it is involved in severalrocesses, such as pain management, emotional behavior,tress response, addictive conditions, and motor responseLastres-Becker et al., 2002; Manzanares et al., 1999; Ortizt al., 2004). Advances in the design of drugs that act onifferent elements of the endogenous cannabinoid systemave resulted in various selective agonists and antagonistsf CB1 and CB2 receptors, as well as inhibitors of anand-mide re-uptake and fatty acid amide hydrolase blockers.
However, little information is available regarding theole of the CB2 cannabinoid receptor in brain function,articularly in the regulation of dopamine neurons. Initially,B2 receptor was found in peripheral tissues (e.g., spleen)nd the immune system (macrophages, microglia, lympho-ytes, and mastocytes) (Galiegue et al., 1995). IncreasedB2 receptor gene expression is related to physiologicalefense processes against several insults, such as tumorrogression (Joosten et al., 2002), glioma (Guzman et al.,001), oxidative stress (Yiangou et al., 2006), acute andeuropathic pain (Ibrahim et al., 2005; Racz et al., 2008a,008b), atherosclerotic plaques, and microglia activation inlzheimer’s disease (Ehrhart et al., 2005), and simian im-unodeficiency virus-induced encephalitis (Benito et al.,
005). In fact, CB1 and CB2 receptors are detected in theenile plaques of Alzheimer patients, together with markersf microglial activation (Ramirez et al., 2005). In particular,annabinoid treatment with WIN-55,212-2 (CB1 agonist)rotects from �-amyloid neurotoxic effects in vivo, andreatment with WIN-55,212-2, HU-210 (analog of �9-THC)nd JWH-133 (CB2 agonist) prevents microglial activationnd neurotoxicity in vitro (Marsicano et al., 2002; Ramirezt al., 2005).
Recent studies demonstrate the presence of CB2 recep-ors in the central nervous system (cerebellum, brainstem,pinal nucleus, hippocampus, olfactory tubercle, cerebralortex, amygdala, striatum, and thalamic nuclei) under basalonditions (Gong et al., 2006; Onaivi et al., 2006; Vanickle et al., 2005). Based on all these data, CB2 receptorsmerge as an element that is likely to be involved in neu-oprotection and could be a potential therapeutic target forhe treatment of neurodegenerative disorders such as Par-inson’s disease (Croxford et al., 2008; Fernandez-Ruiz,009; Fernandez-Ruiz et al., 2007; Garcia-Arencibia et al.,007; Howlett et al., 2002; Lastres-Becker et al., 2005;agliaferro et al., 2006).
Presently, enough data show the involvement of thendocannabinoid system in the central regulation of motorunctions and Parkinson’s disease (Brotchie, 2003; de Lago
nd Fernandez-Ruiz, 2007; Garcia-Arencibia et al., 2007). mome studies even consider that the endocannabinoid sys-em is a key modulatory element in the activity of the basalanglia (Fernandez-Ruiz, 2009). Most findings focus onB1 receptors, but the information available on the role ofB2 receptors in the brain is insufficient and contradic-
ory (Campillo and Paez, 2009; Morgan et al., 2009),articularly in the pathology of Parkinson’s disease. TheB2 receptor is devoid of addictive properties and coulde an interesting therapeutic target for further examina-ion of the treatment of neurodegenerative disorders, suchlzheimer’s disease (Campillo and Paez, 2009) or Hun-
ington’s disease (Sagredo et al., 2009). It is proposed thatn increase in CB2 receptors may protect against the dopa-inergic neuronal loss and motor deteriorations induced by
-hydroxydopamine (6-OHDA). To this end, behavioral andeurochemical alterations related to the depletion of nigro-triatal dopaminergic neurons produced by unilateral intra-audate administration of 6-OHDA were examined in miceverexpressing CB2 cannabinoid receptors (CB2xP) andheir wild type (WT) littermates. The presence of astrocytesnd microglia in the treated area of the caudate-putamenCPu) was analyzed and oxidative/nitrosative and neuroin-ammatory parameters were also estimated in 6-OHDA-
reated and sham-treated mice.
. Methods
.1. Animals
Adult male mice overexpressing cannabinoid CB2 recep-ors (CB2xP) and wild-type littermates (WT) were used inll experiments. Transgenic mice overexpressing the murineB2 receptor were developed in our laboratory as describedreviously (Racz et al., 2008b). Male Swiss albino ICRice (WT) were purchased from Harlan (Harlan, Barcelona,pain). The CB2xP mice and WT mice used in a givenxperiment were originated from the same breeding seriesnd were matched for age and weight (age � 2–3 months;eight � 25–30 g). Mice were maintained at a constant
emperature of 21 � 2 °C under a 12:12 hour light-darkycle (light from 8:00 to 20:00 hours), with free access toood and water. All experimental procedures were carriedut between 09:00 and 13:00 hours and complied with theighest standards of animal care, monitoring health, andinimizing pain and suffering, in accordance with the Eu-
opean Community Council Directive of 24 November 198686/609/EEC) and Spanish national law on the care and usef laboratory animals.
.2. Surgical procedures
Mice were anesthetized using a ketamine/xylazine (88g/kg and 20 mg/kg, respectively) solution and placed in a
tereotaxic frame (Stoelting Co, Wood Dale, IL, USA) withose and ear bars specially adapted for mice (Cunninghamdaptor, Stoelting). Two microliters of 6-hydroxy-dopa-
ine (6-OHDA) hydrobromide (Sigma-Aldrich, Madrid,
SafiismamipsCdSti
2
2
Ssoarra5Cmon
2
cttai5
2
danm
2
3ttwaa
pt
2
wtmpBtswMpiagt
2
iMXioLt(Xrd0wvscuUt
2
gfwceesmhg
3A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
pain), 3 �g freebase/�L in 0.9% saline and 0.02% ascorbiccid, was injected at 2 different coordinates, resulting in anal dosage of 12 �g/mouse. A unilateral lesion was made
n the right caudate-putamen (CPu) using a 10-�L Hamiltonyringe and 2 different coordinates: anterior-posterior: �1.0m; medial-lateral: �2.1 mm; Dorsal-Ventral: �2.9 mm
nd Anterior-Posterior: �0.3 mm; Medial-Lateral: �2.3m; Dorsal-Ventral: �2.9 mm; Tooth-bar at �0 mm. The
njection rate was 0.5 �L/minute and the needle was left inlace for another 5 minutes after injection before it waslowly withdrawn. To serve as controls, WT mice andB2xP mice were subjected to the same surgical proce-ures, but only vehicle was injected in the same coordinates.even weeks after the lesion, mice were killed by decapi-
ation in order to study brain histochemical and neurochem-cal parameters.
.3. Behavioral analysis
.3.1. Apomorphine-induced rotationApomorphine hydrochloride (Sigma-Aldrich, Madrid,
pain) was dissolved in saline solution and administered totudy the rotational behavior in mice injected with 6-OHDAr vehicle. Tests were performed at 3 different time pointsfter the injection and apomorphine-induced contralateralotation was measured over the 7-week experimental pe-iod. Mice received 2 mg/kg apomorphine subcutaneouslynd were placed in individual glass bowls of 25 � 25 cm forminutes to allow them to habituate to the environment.ontralateral turns to the lesion were recorded over 1inute at 5, 10, 15, and 20 minutes after the administration
f apomorphine. Results were averaged and expressed aset contralateral turns/minute.
.3.2. Light-dark box testThe light-dark box model (Crawley and Goodwin, 1980)
onsisted of 2 methacrylate boxes 20 � 20 � 15 cm, 1ransparent and 1 black and opaque, linked by an opaqueunnel (4 cm). Light from a 60-W desk lamp located 25 cmbove the light box provided room illumination. Mice werendividually placed facing the black box and tested in-minute sessions.
.3.3. Elevated plus maze testThe elevated plus maze test was carried out as previously
escribed (Lister, 1987). The apparatus was elevated 75 cmbove the floor. The time spent in the open arms (s) andumber of entries into the open and closed arms wereeasured during a 5-minute period.
.3.4. Step down inhibitory avoidance testThe step down inhibitory avoidance apparatus was a
1 � 19 � 15-cm acrylic box with a platform located nexto a grid. Mice were placed on the platform and the latencyime until they stepped down onto the grid with all 4 pawsas measured, following a modified protocol (Izquierdo et
l., 1998). During the training session, the animals received
2-second, 0.4-mA scrambled foot shock right after step- ding down onto the grid and were immediately taken out ofhe box.
.4. Immunohistochemical analysis
Mice overexpressing cannabinoid CB2 receptor and theirild-type littermates (n � 3–5 mice per group) were anes-
hetized with ketamine/xylazine (2:1 v/v, 0.2 mL perouse) and intracardially perfused with 200 mL of 4%
araformaldehyde in phosphate buffer (PB; 0.1 M, pH 7.4).rains were dissected, postfixed in the same fixative solu-
ion overnight at 4 °C, frozen, and cut into coronal 40-�mections using a vibratome. Then, floating sections wereashed 3 times with phosphate-buffered saline (PBS; 0.1, pH 7.3): sections were incubated with 1% hydrogen
eroxide in PBS for 20 minutes at room temperature tonhibit endogenous peroxidase, washed 3 times with PBS,nd then incubated for 1 hour in a solution of 10% normaloat serum (NGS) in PBS � 0.3% triton X-100 at roomemperature.
.4.1. Tyrosine hydroxylase expressionFor tyrosine hydroxylase (TH) expression, sections were
ncubated in primary TH antibody (Chemicon-Millipore,adrid, Spain) diluted to 1:5000 in PBS � 0.3% triton-100 overnight at room temperature. Sections were rinsed,
ncubated for 1 hour at room temperature in a 1:200 dilutionf biotinylated goat anti-rabbit secondary antibody (Vectorabs, Burlingame, CA, USA) in PBS � 0.3% triton X-100,
hen rinsed again and incubated in extravidin-peroxidaseSigma-Aldrich) diluted 1:2000 in PBS � 0.2% triton-100 for 1 hour at room temperature. Finally, sections were
insed and then incubated in a solution containing 0.05%iaminobenzidine-nickel (DAB-Ni) (Sigma-Aldrich) and.003% hydrogen peroxide for color deposition. Sectionsere mounted on coated slides, dehydrated, cover slipped,iewed, and photographed using a Zeiss and Leitz micro-cope (Zeiss, Thornwood, NY, USA) and a Nikon digitalamera (Nikon Corp., Tokyo, Japan). Images were editedsing Photoshop (vCS3; Adobe Systems) San Jose, CA,SA) and quantified using ImageJ software (NIH, Be-
hesda, MD, USA) after background subtraction.
.4.2. Quantification of TH immunolabeled cellsStereological cell count was performed using the Cell-
raph system (Microptic SL, Barcelona, Spain). The opticalractionator technique (adapted to 10-�m thick sections)as used to obtain estimates of total TH immunopositive
ell numbers within the CPu (Gundersen et al., 1999; Westt al., 1991). A systematically sampled series of sectionsvery 300 �m spanning the entire extent of the striatum waselected for quantification. The number of striatal TH im-unopositive cells was quantified on 6-OHDA-lesioned
emisphere of CB2xP and WT groups (n � 4–6 mice/roup). During quantification, a sampling grid of known
imensions (22,500 �m2) was positioned over each area and
cgstf
2
ciGI1XraCb1tb(ddZItb
2
twamd(b(rtWmBd(raiimmvsT
wi
2
2m
cfGsvabSdaatrsu
2
potiwtt2vcalM
2
wftPessmCwsmoS
4 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
ounting was conducted using a 40� lens. The countsenerated by the application of the dissectors represent aample from a known fraction of the entire CPu and allowo extrapolate a final estimation of the total neuron numberor each area (West et al., 1991).
.4.3. GFAP and Iba-1 expressionsFor glial fibrillary acidic protein (GFAP) and ionized
alcium-binding adapter molecule 1 (Iba-1), sections werencubated overnight at room temperature in either primaryFAP antibody (Dako, Glostrup, Denmark) or primary
ba-1 antibody (Wako Chemicals, Osaka, Japan), diluted to:5000 and 1:300, respectively, both in PBS � 0.3% triton-100. Sections then were rinsed, incubated for 1 hour at
oom temperature in a 1:200 dilution of biotinylated goatnti-rabbit secondary antibody (Vector Labs, Burlingame,A, USA) in PBS � 0.3% triton X-100, rinsed, and incu-ated in extravidin-peroxidase (Sigma-Aldrich) diluted to:2000 in PBS � 0.2% triton X-100 for 1 hour at roomemperature. Finally, sections were rinsed and then incu-ated in a solution containing 0.05% diaminobenzidineSigma-Aldrich) and 0.003% hydrogen peroxide for coloreposition. Sections were mounted on coated slides, dehy-rated, cover slipped, viewed, and photographed using aeiss and Leitz microscope and a Nikon digital camera.
mages were edited using Photoshop (vCS3; Adobe Sys-ems) and quantified using ImageJ software (NIH) afterackground subtraction.
.5. CB2 receptor gene expression by qRT-PCR
Mice were decapitated and brains were removed fromhe skull and frozen over dry ice. Brain sections of 500 �mere cut at different levels containing the regions of interest
ccording to Paxinos and Franklin (2001). Sections wereounted on slides and stored at �80 °C. Sections were
issected following the method described by Palkovits1983). Total ribonucleic acid (RNA) was obtained fromrain punches using Biozol® Total RNA extraction reagentBioflux-Inilab, Madrid, Spain). After DNAse digestion,everse transcription was carried out following the instruc-ions of the manufacturer (Epicentre Tech Corp., Madison,
I, USA). CB2 (Mm 00438286_m1) gene expression waseasured using TaqMan Gene Expression assay (Appliediosystems) as a double-stranded DNA-specific fluorescentye and performed on the AbiPrism 7700 Real Time CyclerApplied Biosystems). The reference gene used was 18SRNA, detected using TaqMan ribosomal RNA control re-gents. Briefly, the data for each target gene were normal-zed to the endogenous reference gene, and the fold changen target gene abundance was determined using the 2���Ct
ethod (Livak and Schmittgen, 2001). The quantificationethod used involves comparing the threshold cycle (Ct)
alues of the samples of interest with a control or calibrator,uch as a nontreated sample or RNA from normal tissue.
he Ct values of both the calibrator and samples of interest aere normalized to an appropriate endogenous housekeep-ng gene.
.6. Oxidative level
.6.1. Oxidized (GSSG) and reduced (GSH) glutathioneeasurementGlutathione, the major antioxidant produced by living
ells, directly participates in the neutralization of harmfulree radicals and reactive oxygen species. An increasedSSG-to-GSH ratio is considered indicative of oxidative
tress. Glutathione measurements were performed as pre-iously described (Jordan et al., 2002). Briefly, striatumnd cortex were homogenized in 5% sulfosalicylic aciduffer (1:3, w/v) at 4 °C and centrifuged at 12,000g.upernatants were transferred to Eppendorf tubes kept onry ice until assayed for glutathione content. Standardsnd tissue homogenates were assayed by calculating therithmetic means of at least triplicate samples. The reac-ion was started by adding the sample and glutathioneeductase. The levels of glutathione were measured intriatum and cortex homogenates corresponding to gen-ine GSSG and GSH.
.6.2. Determination of lipid peroxidationMalonyldialdehyde (MDA) is the final product of lipid
eroxidation. MDA concentration represents a scoreboardf oxidative stress. The process of lipid peroxidation leadso the production of peroxides and their derivatives, result-ng in a loss of membrane function. MDA concentrationsere measured under basal conditions and in 6-OHDA-
reated WT mice and CB2xP mice. MDA levels were de-ermined as previously described (Gomez-Lazaro et al.,007). Each sample was briefly homogenized in a 3-foldolume of ice-cold 10 mM Tris-HCl buffer, pH 7.4. Afterentrifugation for 6 minutes at 3000 rpm and 4 °C, themount of MDA was determined in supernatant using aipid peroxidation assay kit from Calbiochem (Calbiochem,
adrid, Spain).
.6.3. Western blot analysesTissue samples were homogenized in lysis buffer (1:10,
/v). The suspension was centrifuged at 13,000–14,000gor 5 minutes. The protein concentration from each condi-ion was quantified spectrophotometrically (Micro BCArotein Reagent Kit, Pierce, Rockford, IL, USA), and anqual amount of protein (35 �g) was loaded onto 7.5%odium dodecylsulfate-polyacrylamide for gel electrophore-is. After electrophoresis, proteins were transferred to im-obilon polyvinylidene difluoride membranes (Milliporeorp., Billerica, MA, USA). Nonspecific protein bindingas blocked with 4% w/v nonfat dried milk, 4% bovine
erum albumin and 0.1% Tween 20 in PBS for 1 hour. Theembranes were incubated with anti-iNOS (inducible nitric
xide synthase; 1:1000 dilution of anti-rabbit polyclonal,anta Cruz Biotechnology, Santa Cruz, CA, USA) and
nti-COX2 (cyclooxygenase 2; 1:1000, Cell Signaling
TwTsi0eBeUq(
2
wfipmdpcb
3
3ab
nawat
6s6
36m
dTi6tztclF2gtg
itprtmgWF0ti6
FiTrCC
5A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
echnology, Boston, MA, USA). After washing with 4%/v nonfat dried milk, 4% bovine serum albumin and 0.1%ween 20 in PBS, the membranes were incubated with aecondary antibody (1:5000, Promega, Madison, WI, USA)n 4% w/v nonfat dried milk, 4% bovine serum albumin and.1% Tween 20 in PBS. The signal was detected using annhanced chemiluminescence detection kit (GE Healthcare,uckinghamshire, UK). Immunoblots were developed byxposure to X-ray film (Eastman Kodak, Rochester, NY,SA). The density of the bands on the Western blot wasuantified by densitometric analysis using ImageJ softwareNIH). The relative levels were normalized to �-tubulin.
.7. Statistical analyses
Statistical analyses were made using the Student t testhen comparing 2 groups and 2-way analysis of variance
ollowed by the Student Newman-Keuls test when compar-ng 4 groups. Differences were considered significant if therobability of error was less than 5%. Data are presented asean � standard error of the mean (SEM). The number of
epicted characters are in accordance with their respectivevalues mentioned in the text: 1 character for p � 0.05, 2
haracters for p � 0.01, and 3 characters for p � 0.001 andeyond.
. Results
.1. Time course effects of intracaudate-putamendministration of 6-OHDA (weeks 1–7) on rotationalehavior in CB2xP mice and WT mice
The administration of apomorphine (2 mg/kg, subcuta-eously) produced a time-dependent decrease (weeks 2, 4,nd 6) in contralateral rotations in CB2xP mice comparedith WT mice (n � 12–16 mice per group). In addition,
nalysis of the rotational behavior revealed that 6-OHDA-reated CB2xP mice presented fewer rotations than
ig. 1. Apomorphine-induced rotations. Effect of a unilateral intracaudatenjections) or its saline vehicle (0.9% saline in 0.02% ascorbic acid; 2 �Lests were performed 2, 4, and 6 weeks after treatment and apomorphine-i
ecorded over 1 minute at 5, 10, 15, and 20 minutes after apomorphine admolumns represent the means and vertical lines mark the � standard erro
B2xP mice that differed significantly (Student t test, p � 0.01 and p � 0.001,-OHDA-treated WT mice at the 3 different time pointstudied (week 2, p � 0.002; week 4, p � 0.001; and week, p � 0.001) (Fig. 1).
.2. Assessment of emotional and cognitive behaviors in-OHDA-treated and sham-treated CB2xP mice and WTice
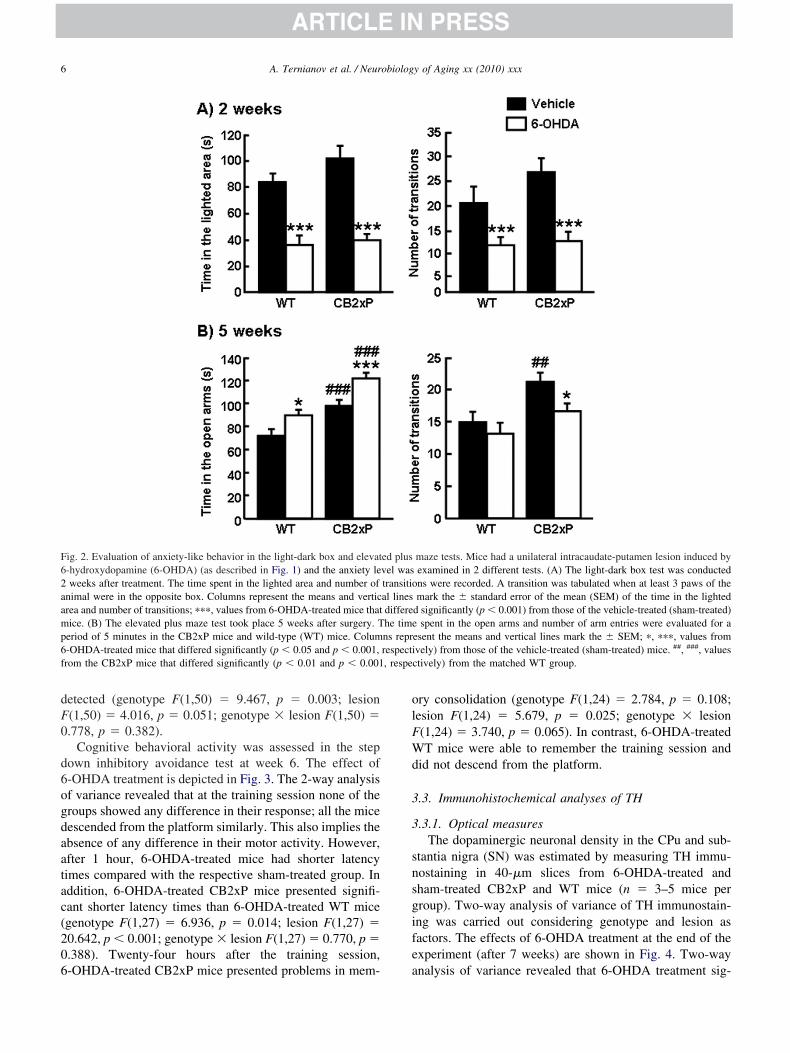
Emotional behavior activity was assessed by the light-ark box test (week 2) and elevated plus maze test (week 5).he effects of 6-OHDA 2 weeks after treatment are depicted
n Fig. 2A. Two-way analysis of variance revealed that-OHDA treatment significantly decreased the time spent inhe illuminated area and the number of transitions betweenones in both 6-OHDA-treated CB2xP mice and 6-OHDA-reated WT mice compared with the respective sham-treatedontrol groups (genotype F(1,39) � 2.080, p � 0.158;esion F(1,39) � 51.382, p � 0.001; genotype � lesion(1,39) � 0.860, p � 0.360, for time; genotype F(1,53) �.072, p � 0.156; lesion F(1,53) � 21.057, p � 0.001;enotype � lesion F(1,53) � 1.153, p � 0.288, for transi-ions). No differences were found between sham-treatedroups.
The effect of 6-OHDA treatment after 5 weeks is shownn Fig. 2B. In this case, the elevated plus maze test was usedo avoid the bias likely to result from habituation during therevious light-dark box test. Two-way analysis of varianceevealed that 6-OHDA treatment significantly increased theime spent in the open arms in both CB2xP mice and WTice compared with the respective sham-treated control
roup, and also between 6-OHDA-treated CB2xP mice andT mice (genotype F(1,35) � 37.997, p � 0.001; lesion
(1,35) � 19.004, p � 0.001; genotype � lesion F(1,35) �.383, p � 0.540). In contrast, a higher number of transi-ions between the open and closed arms was only foundn the control groups; a significant difference between-OHDA-treated and sham-treated CB2xP mice was also
n lesion, induced with 6-hydroxydopamine (6-OHDA) (12 �g/mice in 2n), on the rotational behavior of CB2xP mice and wild type (WT) mice.contralateral rotation was measured. Contralateral turns to the lesion wereon. Results were averaged and expressed as net contralateral turns/minute.mean (SEM) of the net contralateral turns/minute. ��, ���, values from
-putame/injectionducedinistratir of the
respectively) from those of WT mice.
dF0
d6ogdaatac(206
olFWd
3
3
snsgife
F62aamp6f , respe
6 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
etected (genotype F(1,50) � 9.467, p � 0.003; lesion(1,50) � 4.016, p � 0.051; genotype � lesion F(1,50) �.778, p � 0.382).
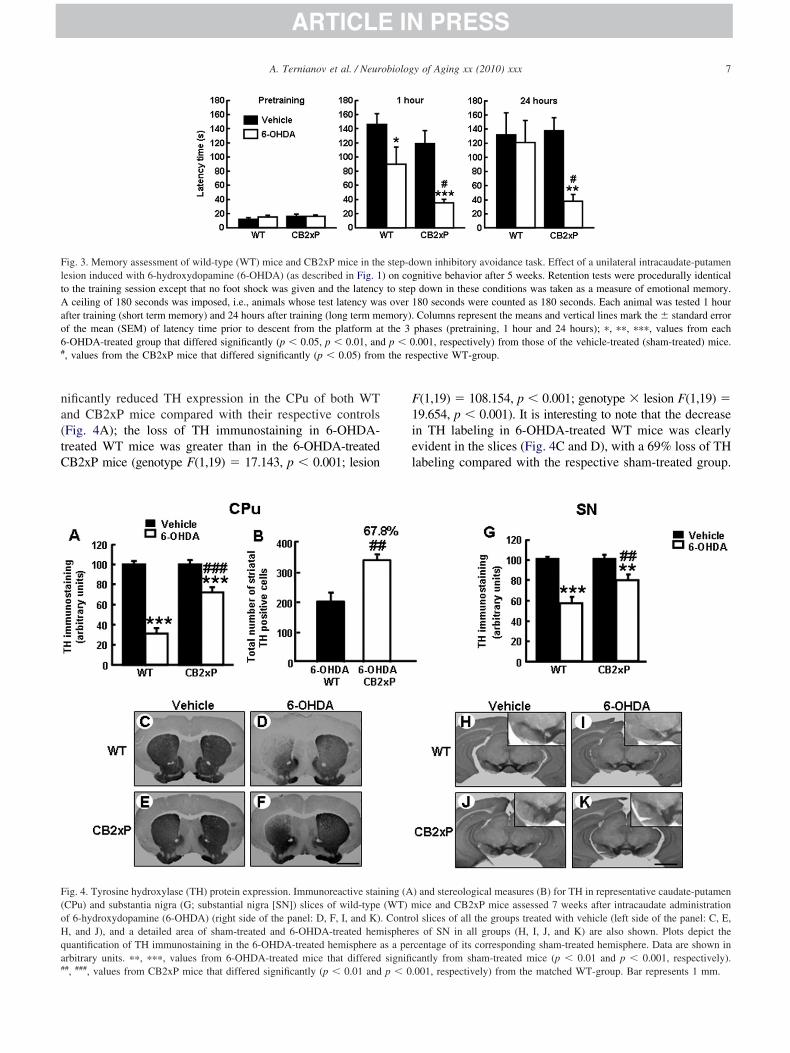
Cognitive behavioral activity was assessed in the stepown inhibitory avoidance test at week 6. The effect of-OHDA treatment is depicted in Fig. 3. The 2-way analysisf variance revealed that at the training session none of theroups showed any difference in their response; all the miceescended from the platform similarly. This also implies thebsence of any difference in their motor activity. However,fter 1 hour, 6-OHDA-treated mice had shorter latencyimes compared with the respective sham-treated group. Inddition, 6-OHDA-treated CB2xP mice presented signifi-ant shorter latency times than 6-OHDA-treated WT micegenotype F(1,27) � 6.936, p � 0.014; lesion F(1,27) �0.642, p � 0.001; genotype � lesion F(1,27) � 0.770, p �.388). Twenty-four hours after the training session,
ig. 2. Evaluation of anxiety-like behavior in the light-dark box and elevat-hydroxydopamine (6-OHDA) (as described in Fig. 1) and the anxiety leweeks after treatment. The time spent in the lighted area and number of
nimal were in the opposite box. Columns represent the means and verticrea and number of transitions; ���, values from 6-OHDA-treated mice thatice. (B) The elevated plus maze test took place 5 weeks after surgery. T
eriod of 5 minutes in the CB2xP mice and wild-type (WT) mice. Colum-OHDA-treated mice that differed significantly (p � 0.05 and p � 0.001,rom the CB2xP mice that differed significantly (p � 0.01 and p � 0.001
-OHDA-treated CB2xP mice presented problems in mem- a
ry consolidation (genotype F(1,24) � 2.784, p � 0.108;esion F(1,24) � 5.679, p � 0.025; genotype � lesion(1,24) � 3.740, p � 0.065). In contrast, 6-OHDA-treatedT mice were able to remember the training session and
id not descend from the platform.
.3. Immunohistochemical analyses of TH
.3.1. Optical measuresThe dopaminergic neuronal density in the CPu and sub-
tantia nigra (SN) was estimated by measuring TH immu-ostaining in 40-�m slices from 6-OHDA-treated andham-treated CB2xP and WT mice (n � 3–5 mice perroup). Two-way analysis of variance of TH immunostain-ng was carried out considering genotype and lesion asactors. The effects of 6-OHDA treatment at the end of thexperiment (after 7 weeks) are shown in Fig. 4. Two-way
maze tests. Mice had a unilateral intracaudate-putamen lesion induced byexamined in 2 different tests. (A) The light-dark box test was conductedns were recorded. A transition was tabulated when at least 3 paws of themark the � standard error of the mean (SEM) of the time in the lighted
d significantly (p � 0.001) from those of the vehicle-treated (sham-treated)spent in the open arms and number of arm entries were evaluated for a
esent the means and vertical lines mark the � SEM; �, ���, values fromvely) from those of the vehicle-treated (sham-treated) mice. ##, ###, valuesctively) from the matched WT group.
ed plusvel wastransitioal linesdifferehe timens reprrespecti
nalysis of variance revealed that 6-OHDA treatment sig-
na(tC
F1iel
FltAao6# the re
F(oHqa#
7A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
ificantly reduced TH expression in the CPu of both WTnd CB2xP mice compared with their respective controlsFig. 4A); the loss of TH immunostaining in 6-OHDA-reated WT mice was greater than in the 6-OHDA-treatedB2xP mice (genotype F(1,19) � 17.143, p � 0.001; lesion
ig. 3. Memory assessment of wild-type (WT) mice and CB2xP mice in theesion induced with 6-hydroxydopamine (6-OHDA) (as described in Fig. 1o the training session except that no foot shock was given and the latency
ceiling of 180 seconds was imposed, i.e., animals whose test latency wafter training (short term memory) and 24 hours after training (long term mf the mean (SEM) of latency time prior to descent from the platform a-OHDA-treated group that differed significantly (p � 0.05, p � 0.01, an, values from the CB2xP mice that differed significantly (p � 0.05) from
ig. 4. Tyrosine hydroxylase (TH) protein expression. Immunoreactive staiCPu) and substantia nigra (G; substantial nigra [SN]) slices of wild-typef 6-hydroxydopamine (6-OHDA) (right side of the panel: D, F, I, and K), and J), and a detailed area of sham-treated and 6-OHDA-treated hemuantification of TH immunostaining in the 6-OHDA-treated hemisphere arbitrary units. ��, ���, values from 6-OHDA-treated mice that differed
#, ###, values from CB2xP mice that differed significantly (p � 0.01 and p � 0.(1,19) � 108.154, p � 0.001; genotype � lesion F(1,19) �9.654, p � 0.001). It is interesting to note that the decreasen TH labeling in 6-OHDA-treated WT mice was clearlyvident in the slices (Fig. 4C and D), with a 69% loss of THabeling compared with the respective sham-treated group.
own inhibitory avoidance task. Effect of a unilateral intracaudate-putamennitive behavior after 5 weeks. Retention tests were procedurally identicaldown in these conditions was taken as a measure of emotional memory.80 seconds were counted as 180 seconds. Each animal was tested 1 hourColumns represent the means and vertical lines mark the � standard error
phases (pretraining, 1 hour and 24 hours); �, ��, ���, values from each.001, respectively) from those of the vehicle-treated (sham-treated) mice.spective WT-group.
) and stereological measures (B) for TH in representative caudate-putamenice and CB2xP mice assessed 7 weeks after intracaudate administration
l slices of all the groups treated with vehicle (left side of the panel: C, E,s of SN in all groups (H, I, J, and K) are also shown. Plots depict thecentage of its corresponding sham-treated hemisphere. Data are shown inantly from sham-treated mice (p � 0.01 and p � 0.001, respectively).
step-d) on cogto step
s over 1emory).t the 3d p � 0
ning (A(WT) m
. Controispheres a persignific
001, respectively) from the matched WT-group. Bar represents 1 mm.
6i
wsvccti6(60aadiwtt
3
o4nw(oT
3
areGtpna
F(6I(*(
8 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
-OHDA-treated CB2xP mice only had 28% less TH label-ng than the sham-treated mice (Fig. 4, E and F).
In SN, the level of lesion reached 7 weeks after surgeryas less pronounced compared with CPu, although the
ame pattern of damage was found. Thus, 2-way analysis ofariance showed that 6-OHDA treatment significantly de-reased TH immunostaining in both WT and CB2xP miceompared with their respective controls (Fig. 4G), and de-ected the same trend toward more loss of TH immunostain-ng in the 6-OHDA-treated WT mice compared with the-OHDA-treated CB2xP mice as was observed in the CPugenotype F(1,17) � 8.374, p � 0.012; lesion F(1,17) �8.270, p � 0.001; genotype � lesion F(1,17) � 8.236, p �.012). In this case, the 6-OHDA-treated WT mice showed43% decrease with respect to controls (Fig. 4, pictures H
nd I), whereas the 6-OHDA-treated CB2xP mice showed aecrease of only 21% (Fig. 4, J and K). Representativemages in Fig. 4G, H, I and J show slices at the level of SN,ith detail over the treated area. These results suggest that
he CB2xP mice were less vulnerable to 6-OHDA treatmenthan the WT mice.
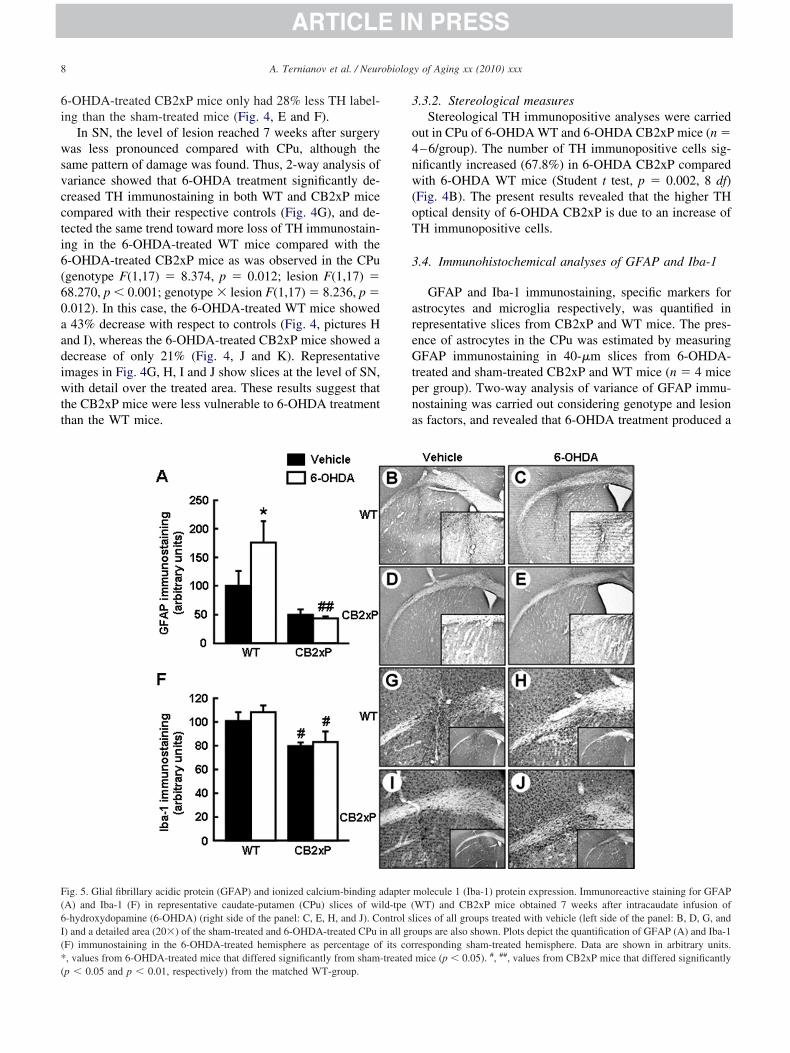
ig. 5. Glial fibrillary acidic protein (GFAP) and ionized calcium-binding aA) and Iba-1 (F) in representative caudate-putamen (CPu) slices of wi-hydroxydopamine (6-OHDA) (right side of the panel: C, E, H, and J). Co) and a detailed area (20�) of the sham-treated and 6-OHDA-treated CPu iF) immunostaining in the 6-OHDA-treated hemisphere as percentage of, values from 6-OHDA-treated mice that differed significantly from sham-
p � 0.05 and p � 0.01, respectively) from the matched WT-group..3.2. Stereological measuresStereological TH immunopositive analyses were carried
ut in CPu of 6-OHDA WT and 6-OHDA CB2xP mice (n �–6/group). The number of TH immunopositive cells sig-ificantly increased (67.8%) in 6-OHDA CB2xP comparedith 6-OHDA WT mice (Student t test, p � 0.002, 8 df)
Fig. 4B). The present results revealed that the higher THptical density of 6-OHDA CB2xP is due to an increase ofH immunopositive cells.
.4. Immunohistochemical analyses of GFAP and Iba-1
GFAP and Iba-1 immunostaining, specific markers forstrocytes and microglia respectively, was quantified inepresentative slices from CB2xP and WT mice. The pres-nce of astrocytes in the CPu was estimated by measuringFAP immunostaining in 40-�m slices from 6-OHDA-
reated and sham-treated CB2xP and WT mice (n � 4 miceer group). Two-way analysis of variance of GFAP immu-ostaining was carried out considering genotype and lesions factors, and revealed that 6-OHDA treatment produced a
molecule 1 (Iba-1) protein expression. Immunoreactive staining for GFAPWT) and CB2xP mice obtained 7 weeks after intracaudate infusion ofices of all groups treated with vehicle (left side of the panel: B, D, G, andups are also shown. Plots depict the quantification of GFAP (A) and Iba-1
responding sham-treated hemisphere. Data are shown in arbitrary units.mice (p � 0.05). #, ##, values from CB2xP mice that differed significantly
dapterld-tpe (ntrol sln all groits cor
treated
s(A(gs(20sptiCaot6Tc�C(F0aaddm
3
mv
iF06F1ts
3
baCt0Mcat�rs7a8F0FrttpFF
FatfW
9A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
ignificant increase in GFAP level only in WT mice�75%) compared with their respective controls (Fig. 5A).strocyte recruitment was lower in both CB2xP groups
ranging from �49% to �42%) compared with the WTroups. The difference in astrocyte recruitment reachedtatistical significance between the 6-OHDA-treated groupsgenotype F(1,15) � 16.109, p � 0.002; lesion F(1,15) �.253, p � 0.159; genotype � lesion F(1,15) � 3.221, p �.098). Representative images (Fig. 5B, C, D, and E) arehown with detail over the 6-OHDA-treated areas, whichresented a higher number of astrocytes. The microglia inhe CPu were estimated by measuring Iba-1 immunostain-ng in 40-�m slices from 6-OHDA-treated and sham-treatedB2xP and WT mice (n � 8–11 mice per group). Two-waynalysis of variance of Iba-1 immunostaining was carriedut considering genotype and lesion as factors, and revealedhat 6-OHDA treatment slightly increased Iba-1 level in-OHDA-treated mice compared with controls (Fig. 5F).he microglia density was lower in the CB2xP groupsompared with the WT groups (ranging from �21% to18%). Only the difference between the 6-OHDA-treatedB2xP and 6-OHDA-treated WT groups was significant
p � 0.013) (genotype F(1,37) � 10.429, p � 0.003; lesion(1,37) � 0.549, p � 0.464; genotype � lesion F(1,37) �.088, p � 0.768). Representative images (Fig. 5G, H, I,nd J) are shown, with detail over the 6-OHDA-treatedreas, which presented higher microglia expression. Theseata suggest that CB2xP mice experienced fewer lesionsue to a lower participation and/or activation of these im-unoreactive cells.
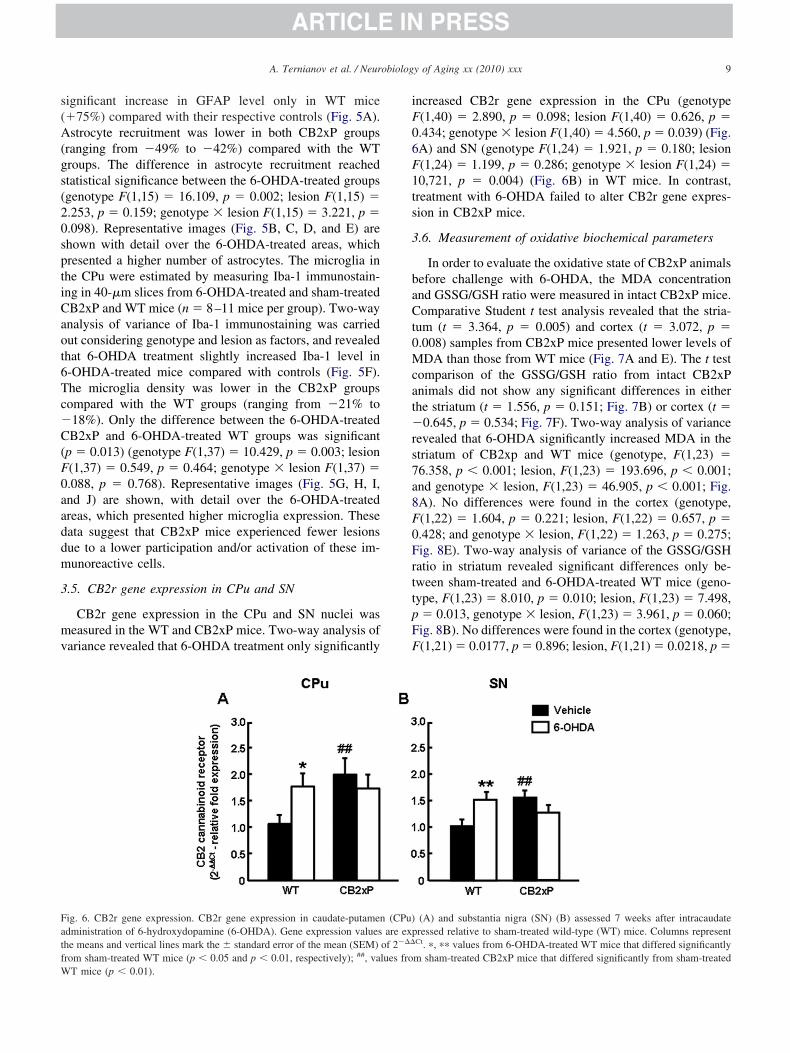
.5. CB2r gene expression in CPu and SN
CB2r gene expression in the CPu and SN nuclei waseasured in the WT and CB2xP mice. Two-way analysis of
ariance revealed that 6-OHDA treatment only significantly
ig. 6. CB2r gene expression. CB2r gene expression in caudate-putamendministration of 6-hydroxydopamine (6-OHDA). Gene expression valueshe means and vertical lines mark the � standard error of the mean (SEM)rom sham-treated WT mice (p � 0.05 and p � 0.01, respectively); ##, va
T mice (p � 0.01).
ncreased CB2r gene expression in the CPu (genotype(1,40) � 2.890, p � 0.098; lesion F(1,40) � 0.626, p �.434; genotype � lesion F(1,40) � 4.560, p � 0.039) (Fig.A) and SN (genotype F(1,24) � 1.921, p � 0.180; lesion(1,24) � 1.199, p � 0.286; genotype � lesion F(1,24) �0,721, p � 0.004) (Fig. 6B) in WT mice. In contrast,reatment with 6-OHDA failed to alter CB2r gene expres-ion in CB2xP mice.
.6. Measurement of oxidative biochemical parameters
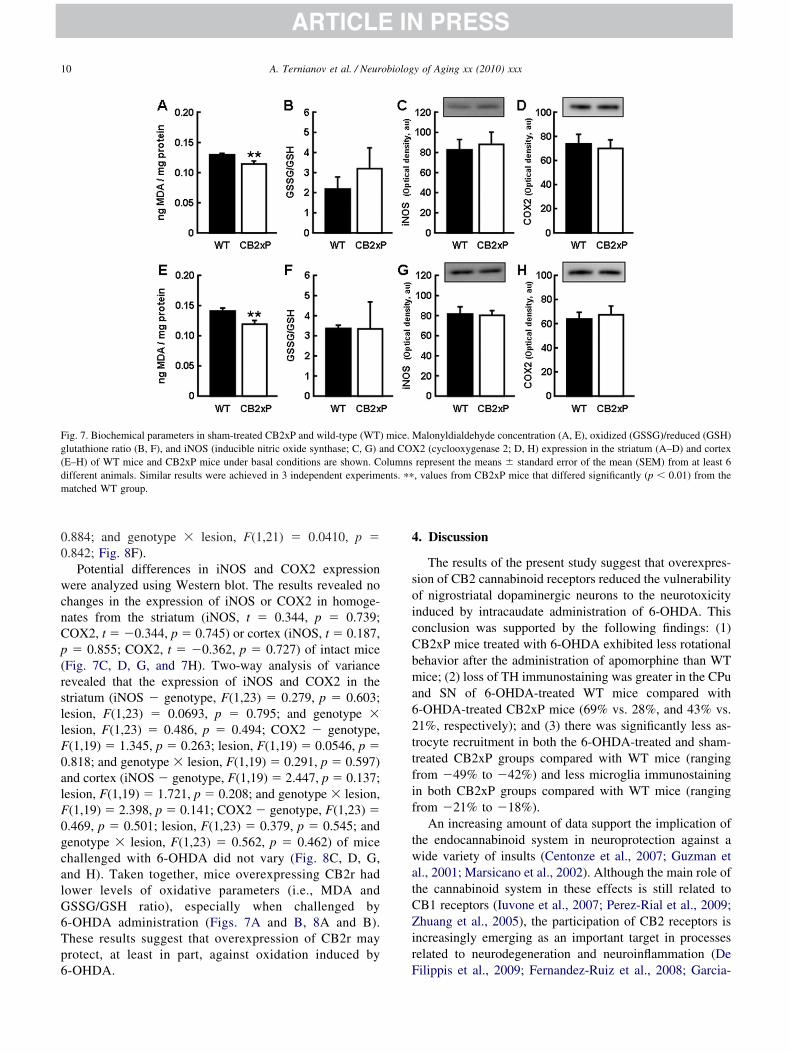
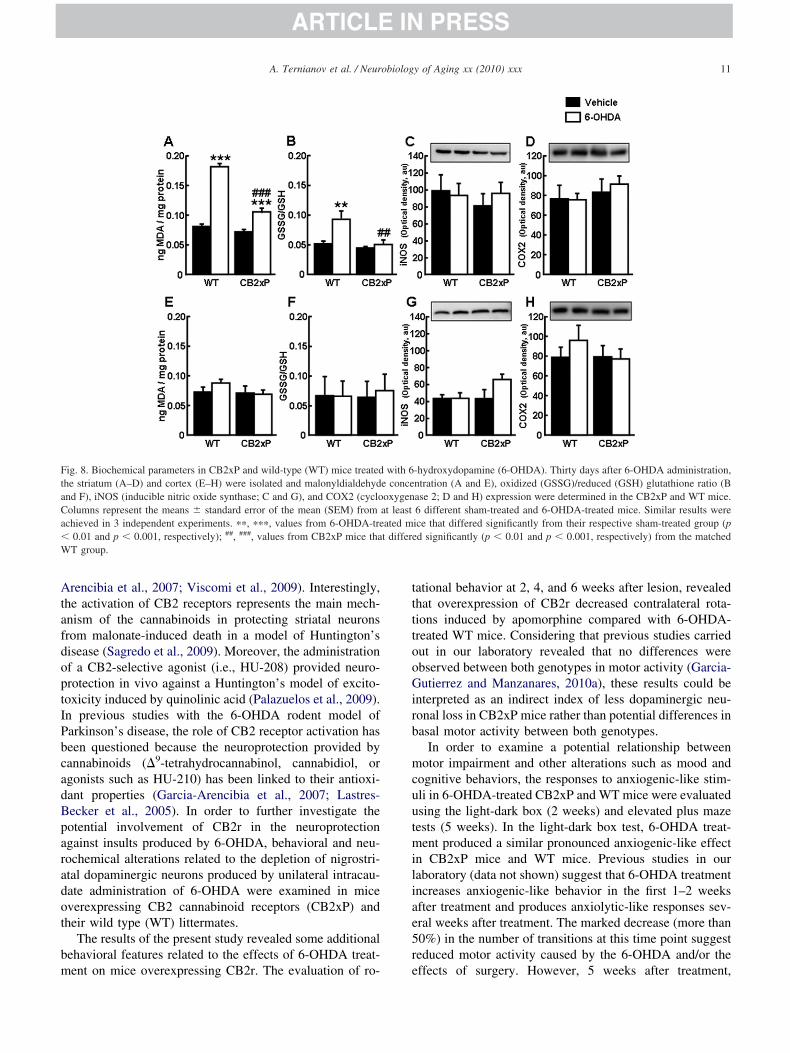
In order to evaluate the oxidative state of CB2xP animalsefore challenge with 6-OHDA, the MDA concentrationnd GSSG/GSH ratio were measured in intact CB2xP mice.omparative Student t test analysis revealed that the stria-
um (t � 3.364, p � 0.005) and cortex (t � 3.072, p �.008) samples from CB2xP mice presented lower levels ofDA than those from WT mice (Fig. 7A and E). The t test
omparison of the GSSG/GSH ratio from intact CB2xPnimals did not show any significant differences in eitherhe striatum (t � 1.556, p � 0.151; Fig. 7B) or cortex (t �
0.645, p � 0.534; Fig. 7F). Two-way analysis of varianceevealed that 6-OHDA significantly increased MDA in thetriatum of CB2xp and WT mice (genotype, F(1,23) �6.358, p � 0.001; lesion, F(1,23) � 193.696, p � 0.001;nd genotype � lesion, F(1,23) � 46.905, p � 0.001; Fig.A). No differences were found in the cortex (genotype,(1,22) � 1.604, p � 0.221; lesion, F(1,22) � 0.657, p �.428; and genotype � lesion, F(1,22) � 1.263, p � 0.275;ig. 8E). Two-way analysis of variance of the GSSG/GSHatio in striatum revealed significant differences only be-ween sham-treated and 6-OHDA-treated WT mice (geno-ype, F(1,23) � 8.010, p � 0.010; lesion, F(1,23) � 7.498,� 0.013, genotype � lesion, F(1,23) � 3.961, p � 0.060;ig. 8B). No differences were found in the cortex (genotype,(1,21) � 0.0177, p � 0.896; lesion, F(1,21) � 0.0218, p �
(A) and substantia nigra (SN) (B) assessed 7 weeks after intracaudateressed relative to sham-treated wild-type (WT) mice. Columns representCt. �, �� values from 6-OHDA-treated WT mice that differed significantlym sham-treated CB2xP mice that differed significantly from sham-treated
(CPu)are exp
of 2���
lues fro
00
wcnCp(rsllF0alF0gcalG6Tp6
4
soicCbma62ttfif
twatCZir
Fg(dm
10 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
.884; and genotype � lesion, F(1,21) � 0.0410, p �
.842; Fig. 8F).Potential differences in iNOS and COX2 expression
ere analyzed using Western blot. The results revealed nohanges in the expression of iNOS or COX2 in homoge-ates from the striatum (iNOS, t � 0.344, p � 0.739;OX2, t � �0.344, p � 0.745) or cortex (iNOS, t � 0.187,� 0.855; COX2, t � �0.362, p � 0.727) of intact mice
Fig. 7C, D, G, and 7H). Two-way analysis of varianceevealed that the expression of iNOS and COX2 in thetriatum (iNOS � genotype, F(1,23) � 0.279, p � 0.603;esion, F(1,23) � 0.0693, p � 0.795; and genotype �esion, F(1,23) � 0.486, p � 0.494; COX2 � genotype,(1,19) � 1.345, p � 0.263; lesion, F(1,19) � 0.0546, p �.818; and genotype � lesion, F(1,19) � 0.291, p � 0.597)nd cortex (iNOS � genotype, F(1,19) � 2.447, p � 0.137;esion, F(1,19) � 1.721, p � 0.208; and genotype � lesion,(1,19) � 2.398, p � 0.141; COX2 � genotype, F(1,23) �.469, p � 0.501; lesion, F(1,23) � 0.379, p � 0.545; andenotype � lesion, F(1,23) � 0.562, p � 0.462) of micehallenged with 6-OHDA did not vary (Fig. 8C, D, G,nd H). Taken together, mice overexpressing CB2r hadower levels of oxidative parameters (i.e., MDA andSSG/GSH ratio), especially when challenged by-OHDA administration (Figs. 7A and B, 8A and B).hese results suggest that overexpression of CB2r mayrotect, at least in part, against oxidation induced by
ig. 7. Biochemical parameters in sham-treated CB2xP and wild-type (WT)lutathione ratio (B, F), and iNOS (inducible nitric oxide synthase; C, G) aE–H) of WT mice and CB2xP mice under basal conditions are shown. Cifferent animals. Similar results were achieved in 3 independent experimatched WT group.
-OHDA. F
. Discussion
The results of the present study suggest that overexpres-ion of CB2 cannabinoid receptors reduced the vulnerabilityf nigrostriatal dopaminergic neurons to the neurotoxicitynduced by intracaudate administration of 6-OHDA. Thisonclusion was supported by the following findings: (1)B2xP mice treated with 6-OHDA exhibited less rotationalehavior after the administration of apomorphine than WTice; (2) loss of TH immunostaining was greater in the CPu
nd SN of 6-OHDA-treated WT mice compared with-OHDA-treated CB2xP mice (69% vs. 28%, and 43% vs.1%, respectively); and (3) there was significantly less as-rocyte recruitment in both the 6-OHDA-treated and sham-reated CB2xP groups compared with WT mice (rangingrom �49% to �42%) and less microglia immunostainingn both CB2xP groups compared with WT mice (rangingrom �21% to �18%).
An increasing amount of data support the implication ofhe endocannabinoid system in neuroprotection against aide variety of insults (Centonze et al., 2007; Guzman et
l., 2001; Marsicano et al., 2002). Although the main role ofhe cannabinoid system in these effects is still related toB1 receptors (Iuvone et al., 2007; Perez-Rial et al., 2009;huang et al., 2005), the participation of CB2 receptors is
ncreasingly emerging as an important target in processeselated to neurodegeneration and neuroinflammation (De
alonyldialdehyde concentration (A, E), oxidized (GSSG)/reduced (GSH)2 (cyclooxygenase 2; D, H) expression in the striatum (A–D) and cortex
represent the means � standard error of the mean (SEM) from at least 6, values from CB2xP mice that differed significantly (p � 0.01) from the
mice. Mnd COXolumnsents. ��
ilippis et al., 2009; Fernandez-Ruiz et al., 2008; Garcia-
AtafdoptIPbcadBparadot
bm
ttttooGirb
mcuutmiliae5r
FtaCa�W
11A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
rencibia et al., 2007; Viscomi et al., 2009). Interestingly,he activation of CB2 receptors represents the main mech-nism of the cannabinoids in protecting striatal neuronsrom malonate-induced death in a model of Huntington’sisease (Sagredo et al., 2009). Moreover, the administrationf a CB2-selective agonist (i.e., HU-208) provided neuro-rotection in vivo against a Huntington’s model of excito-oxicity induced by quinolinic acid (Palazuelos et al., 2009).n previous studies with the 6-OHDA rodent model ofarkinson’s disease, the role of CB2 receptor activation haseen questioned because the neuroprotection provided byannabinoids (�9-tetrahydrocannabinol, cannabidiol, orgonists such as HU-210) has been linked to their antioxi-ant properties (Garcia-Arencibia et al., 2007; Lastres-ecker et al., 2005). In order to further investigate theotential involvement of CB2r in the neuroprotectiongainst insults produced by 6-OHDA, behavioral and neu-ochemical alterations related to the depletion of nigrostri-tal dopaminergic neurons produced by unilateral intracau-ate administration of 6-OHDA were examined in miceverexpressing CB2 cannabinoid receptors (CB2xP) andheir wild type (WT) littermates.
The results of the present study revealed some additionalehavioral features related to the effects of 6-OHDA treat-
ig. 8. Biochemical parameters in CB2xP and wild-type (WT) mice treatedhe striatum (A–D) and cortex (E–H) were isolated and malonyldialdehydnd F), iNOS (inducible nitric oxide synthase; C and G), and COX2 (cycloolumns represent the means � standard error of the mean (SEM) fromchieved in 3 independent experiments. ��, ���, values from 6-OHDA-tre
0.01 and p � 0.001, respectively); ##, ###, values from CB2xP mice thaT group.
ent on mice overexpressing CB2r. The evaluation of ro- e
ational behavior at 2, 4, and 6 weeks after lesion, revealedhat overexpression of CB2r decreased contralateral rota-ions induced by apomorphine compared with 6-OHDA-reated WT mice. Considering that previous studies carriedut in our laboratory revealed that no differences werebserved between both genotypes in motor activity (Garcia-utierrez and Manzanares, 2010a), these results could be
nterpreted as an indirect index of less dopaminergic neu-onal loss in CB2xP mice rather than potential differences inasal motor activity between both genotypes.
In order to examine a potential relationship betweenotor impairment and other alterations such as mood and
ognitive behaviors, the responses to anxiogenic-like stim-li in 6-OHDA-treated CB2xP and WT mice were evaluatedsing the light-dark box (2 weeks) and elevated plus mazeests (5 weeks). In the light-dark box test, 6-OHDA treat-ent produced a similar pronounced anxiogenic-like effect
n CB2xP mice and WT mice. Previous studies in ouraboratory (data not shown) suggest that 6-OHDA treatmentncreases anxiogenic-like behavior in the first 1–2 weeksfter treatment and produces anxiolytic-like responses sev-ral weeks after treatment. The marked decrease (more than0%) in the number of transitions at this time point suggesteduced motor activity caused by the 6-OHDA and/or the
-hydroxydopamine (6-OHDA). Thirty days after 6-OHDA administration,ntration (A and E), oxidized (GSSG)/reduced (GSH) glutathione ratio (Base 2; D and H) expression were determined in the CB2xP and WT mice.6 different sham-treated and 6-OHDA-treated mice. Similar results wereice that differed significantly from their respective sham-treated group (pd significantly (p � 0.01 and p � 0.001, respectively) from the matched
with 6e conceoxygenat leastated mt differe
ffects of surgery. However, 5 weeks after treatment,

6acmrodllActoasmb
atL2itt6P(tltiamsrg2abtirt
rtmr6itsop
tm6lir2dmtataptCsmrssrniab
r6ah
saltowcTattsn
ibcGsmRt
12 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
-OHDA-treated CB2xP and WT mice presented significantnxiolytic-like behaviors compared with their respectiveontrols. These results suggest 2 ideas: (1) CB2xP-vehicleice present less anxiety than WT-vehicle mice. These
esults are in agreement with previous studies performed inur laboratory in mice not submitted to any surgical proce-ure (Garcia-Gutierrez and Manzanares, 2010a), and (2) theesion increased the time that both type of mice spent in theight area. This could be interpreted as decreased anxiety.lternatively, another possibility is that mice may have
hanged their perception of the anxiogenic stimulus due tohe lesion and spent longer time in this area because thepen arms of the elevated plus maze no longer result annxiogenic stimulus. In this test, the number of transitions islightly higher in CB2xP compared with WT mice. Thisay be due to different impact of the lesion on motor
ehavior between both genotypes.Considering that connections between the basal ganglia
nd limbic system are involved in the regulation of emo-ional behavior (Ahn et al., 2007; Hikosaka et al., 2008;evine, 2007; Nieoullon and Coquerel, 2003; Xiao et al.,009), it can be speculated that the dopaminergic depletionn the CPu observed in 6-OHDA-treated mice might alterhe basal-limbic connections that mediate, at least in part,he anxiogenic-like responses observed 2 weeks after-OHDA-induced lesion. Similarly, a large proportion ofarkinson’s disease patients suffer from anxiety disordersAarsland et al., 2009). On the other hand, it is possible thathe anxiolytic-like responses observed in week 5 were re-ated to a lack of perception of the anxiogenic stimuli oncehe lesion becomes more pronounced and stable. Interest-ngly, sham-treated CB2xP mice displayed a higher level ofnxiolytic-like behavior compared with sham-treated WTice at 5 weeks. Previous studies in our laboratory demon-
trated that CB2xP mice have a behavioral endophenotypeesistant to acute and chronic anxiogenic and depresso-enic-like stimuli (Garcia-Gutierrez and Manzanares,010a; Garcia-Gutierrez et al., 2010b). In addition, Onaivind colleagues (Onaivi et al., 2008) indicate that intracere-roventricular administration of an antisense oligonucleo-ide sequence directed against the CB2r gene reduces anx-ety-like behaviors in mice. These data suggest that CB2eceptors play an important role in the regulation of emo-ional behavior.
The evaluation of cognitive behavior, based on aversion-elated conditioning, revealed no differences in latency be-ween 6-OHDA-treated and sham-treated CB2xP and WTice. However, evaluation of the latency time at 1 hour
evealed a potential alteration of short-term memory in-OHDA-treated CB2xP and WT mice that was character-zed by a shorter latency. These results are consistent withhe cognitive alterations observed in patients with Parkin-on’s disease (Aarsland et al., 2009; Kulisevsky and Pag-nabarraga, 2009; Leverenz et al., 2009). Although the
recise neurochemical mechanisms underlying this cogni- 2ive deficit are still unknown, it can be hypothesized that theolecular alterations observed in the CPu and SN after
-OHDA treatment mediated this cognitive impairment, ateast in part. It is well established that the basal ganglia arentimately connected with the brain regions involved in theegulation of cognitive processes (Nieoullon and Coquerel,003; Tisch et al., 2004). It is possible that the dopaminergicepletion observed in the CPu region of 6-OHDA-treatedice may produce alterations in key elements involved in
he consolidation of memory, such as gamma-Aminobutyriccid (GABA) neurotransmission. Recent studies revealedhat CB2xP mice present significant increase of GABAA�2
nd GABAA�2 gene expressions in amygdala and hip-ocampus (Garcia-Gutierrez and Manzanares, 2010a). In-erestingly, 24 hours after shock stimuli, 6-OHDA-treatedB2xP mice had a significantly increased latency in de-
cending from the platform compared with sham-CB2xPice. These results suggest the involvement of CB2r in the
egulation of cognitive behavior. Considering that proteinynthesis is implicated in long term memory (Bekin-chtein et al., 2008; Costa-Mattioli et al., 2009), theseesults suggest that CB2r, through an unknown mecha-ism, might be responsible for the pronounced alterationsn the consolidation of memory processes. Further studiesre needed to explore the mechanisms underlying theseehavioral alterations in CB2xP mice.
Taken together, the evaluation of anxiety and cognitiveesults point out that differences in the vulnerability of-OHDA lesion on motor behavior is not necessarily equiv-lent to alterations in anxiety-and/or cognitive-related be-aviors.
The immunohistochemical analyses of TH clearlyhowed that the dopaminergic neuronal density in the CPund SN of 6-OHDA-treated CB2xP mice was markedlyower than in the 6-OHDA-treated WT controls at the end ofhe experiment. In addition, the lower reduction of THptical density in the CPu of 6-OHDA-treated CB2xP miceas associated with a higher number of TH immunopositive
ells (67.8%) compared with 6-OHDA-treated WT mice.hese findings were also reflected in the lower number ofpomorphine-induced rotations observed in 6-OHDA-reated CB2xP mice in comparison with their 6-OHDA-reated WT littermates and indicate that CB2r overexpres-ion is related to a lower vulnerability of dopaminergiceurons to 6-OHDA toxicity.
Nigrostriatal lesion following striatal 6-OHDA infusions associated with an inflammatory response characterizedy the proliferation and activation of microglia and astro-ytes in a time-dependent manner (Cicchetti et al., 2002;omide et al., 2005; Rodrigues et al., 2004). Nevertheless,
ome neuroprotective effects have been linked to actionsediated by glial cells, particularly microglia (Fernandez-uiz et al., 2007). CB2r seems to play an important role in
he regulation of microglial cells (Fernandez-Ruiz et al.,
007; Walter and Stella, 2004; Walter et al., 2003). A role
faMteii6hnnoewToft2
mgtg(aheCiaciaspie
fabWcTrsMCtaIMoa
etrida(2ltilfksmtn
cn6moltPseu
D
chAo
spmEl
A
d“tJdc
13A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
or CB2r in the astrocytic response to noxious stimuli haslso been hypothesized (Fernandez-Ruiz et al., 2008).oreover, a recent study showed that CB2r participates in
he inhibition of proinflammatory astrocyte function (Shengt al., 2009). In this study, a striking reduction of GFAPmmunostaining and less microglial presence were detectedn the CPu of CB2xP mice 7 weeks after the insult in both-OHDA-treated and sham-treated CB2xP. This findingighlights the important role of glial cells in the response tooxious stimuli and suggests an anti-inflammatory mecha-ism that may contribute to the neuroprotective effectsbserved with CB2 overexpression. No noteworthy differ-nces were found in microglial activation after treatmentith vehicle or 6-OHDA in either WT mice or CB2xP mice.his might be because the histochemical study was carriedut 7 weeks after the lesion, when enough time had elapsedor the inflammatory microglial response that was detectedo have receded (Cicchetti et al., 2002; Rodrigues et al.,001).
CB2r gene expression was increased in 6-OHDA WTice compared with their respective controls in the 2 re-
ions analyzed (CPu and SN). These results coincide withhe findings of a previous study revealing increased CB2rene expression after exposure to different noxious stimuliPrice et al., 2009). In addition, treatment with the CB2r-gonist JWH-015 reduced 1-methyl-4-phenyl-1,2,3,6-tetra-ydropyridine (MPTP)-induced microglial activation (Pricet al., 2009). WT mice under basal conditions present lowB2r gene expression in the CPu and the lesion markedly
ncreased (approximately 30%) the expression of this genes a secondary “defensive response” against the aggressionaused by the neurotoxin. These results suggest that thencrease of CB2r gene expression is secondary to the dam-ge. In contrast, the higher basal level of CB2r gene expres-ion (approximately 50%) in the CPu of CB2xP mice ap-ears to protect against 6-OHDA and may explain, at leastn part, the fact that the 6-OHDA treatment did not alter thexpression of this gene in 6-OHDA-treated CB2xP mice.
According to the above-mentioned studies and due to theact that both Parkinson’s disease (Koutsilieri et al., 2002)nd 6-OHDA-induced lesions (Galindo et al., 2003; Grun-latt et al., 2000; Jenner and Olanow, 1996; Monteiro andinterbourn, 1989) are closely connected to oxidative pro-
esses, a number of biochemical parameters were evaluated.he results revealed that CB2r overexpression protects neu-
ons from oxidation, at least in part. This assumption wasupported by the pronounced differences in the increase ofDA and the GSSG/GSH ratio in the striatum betweenB2xP mice and WT mice treated with 6-OHDA. In con-
rast, no differences were found in the expression of iNOSnd COX2 in any of the experimental groups examined.nterestingly, intact CB2xP mice presented lower levels of
DA in both the striatum and cortex, suggesting that CB2rverexpression increased antioxidant capacity. Glutathione,
ubiquitous tripeptide thiol, is an endogenous intra- and rxtracellular protective antioxidant that plays a key role inhe control of apoptotic death. In line with the presentesults, it has been shown that 6-OHDA-induced cell deaths associated with decreased intracellular GSH levels (Jor-an et al., 2000) and treatments with drugs that enhancentioxidant defenses have been regarded as neuroprotectiveGalindo et al., 2003; Jordan et al., 2004; Tarozzi et al.,009). Indeed, CB2xP mice treated with 6-OHDA had aower GSSG/GSH ratio compared with WT mice. In addi-ion, it has been proposed that early depletion of total GSHn the SN is responsible for the subsequent oxidative stress,ipid peroxidation (Dexter et al., 1989), mitochondrial dys-unction, and dopaminergic cell loss in patients with Par-inson’s disease (Zeevalk et al., 2008). Thus, overexpres-ion of CB2r may reduce the accumulation of oxidativeediators in the brain, making the neurons less vulnerable
o the free radicals produced by 6-OHDA and improving theeurological course.
In summary, the results of this study revealed that in-reased CB2r gene expression greatly attenuated the vul-erability of dopaminergic neurons to treatment with-OHDA. This may be due, at least in part, to less recruit-ent of astrocytes and microglia in the CPu and diminished
xidative parameters (i.e., MDA and the GSSG/GSH ratioevel) in the striatum. Despite the general limitations ofranslating the findings of this animal model to patients witharkinson’s disease, the results of the present study stronglyupport the CB2r as a key target that deserves furtherxploration to determine possible therapeutic strategies reg-lating the functional activity of this receptor.
isclosure statement
All authors declare that they have no actual or potentialonflicts of interest. All authors have verified that the dataave not been published or are being submitted elsewhere.ll authors have approved the procedures and the final formf the manuscript.
All experimental procedures complied with the highesttandards of animal care, monitoring health, and minimizingain and suffering, in accordance with the European Com-unity Council Directive of 24 November 1986 (86/609/EC) and Spanish national law on the care and use of
aboratory animals.
cknowledgements
This research was supported by grants from “Conselleriae Sanitat de la Generalitat Valenciana” (AP-099/09) andFundación de Neurociencias y Envejecimiento” to JM, andhe “Cátedra Distec” for research in Parkinson’s disease toM and CL. This work was also supported by the “Fun-ación para la Investigación Sanitaria en Castilla-La Man-ha (FISCAM)” project PI-2008/21 to MFG, “Incorpo-
ación de Grupos Emergentes” FIS-Carlos III and project
PJMMI((t
R
A
A
B
B
B
B
C
C
C
C
C
C
D
d
D
E
F
F
F
G
G
G
G
G
G
G
G
G
G
G
G
H
H
H
H
14 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
I-080693 from ISCII-Carlos III to MFG and MES. AT,MPO, and AOA are postdoctoral fellows from FISCAM.SGG and FN are predoctoral fellows from the Spanishinistry of Health and the Spanish Ministry of Science and
nnovation, respectively. We thank Patricia RodríguezFISCAM), Raquel Poveda (FISCAM), Carlos A. GarridoFIS-Carlos III), and Analía Rico (RETICS) for excellentechnical assistance.
eferences
arsland, D., Marsh, L., Schrag, A., 2009. Neuropsychiatric symptoms inParkinson’s disease. Mov. Disord. 24, 2175–2186.
hn, M.S., Breeze, J.L., Makris, N., Kennedy, D.N., Hodge, S.M., Herbert,M.R., Seidman, L.J., Biederman, J., Caviness, V.S., Frazier, J.A., 2007.Anatomic brain magnetic resonance imaging of the basal ganglia inpediatric bipolar disorder. J. Affect. Disord. 104, 147–154.
ekinschtein, P., Cammarota, M., Izquierdo, I., Medina, J.H., 2008. BDNFand memory formation and storage. Neuroscientist 14, 147–156.
enito, C., Kim, W.K., Chavarria, I., Hillard, C.J., Mackie, K., Tolon,R.M., Williams, K., Romero, J., 2005. A glial endogenous cannabinoidsystem is upregulated in the brains of macaques with simian immuno-deficiency virus-induced encephalitis. J. Neurosci. 25, 2530–2536.
ouaboula, M., Poinot-Chazel, C., Bourrie, B., Canat, X., Calandra, B.,Rinaldi-Carmona, M., Le Fur, G., Casellas, P., 1995. Activation ofmitogen-activated protein kinases by stimulation of the central canna-binoid receptor CB1. Biochem. J. 312 (Pt 2), 637–641.
rotchie, J.M., 2003. CB1 cannabinoid receptor signalling in Parkinson’sdisease. Curr. Opin. Pharmacol. 3, 54–61.
ampillo, N.E., Paez, J.A., 2009. Cannabinoid system in neurodegenera-tion: new perspectives in Alzheimer’s disease. Mini Rev. Med. Chem.9, 539–559.
entonze, D., Finazzi-Agro, A., Bernardi, G., Maccarrone, M., 2007. Theendocannabinoid system in targeting inflammatory neurodegenerativediseases. Trends Pharmacol. Sci. 28, 180–187.
icchetti, F., Brownell, A.L., Williams, K., Chen, Y.I., Livni, E., Isacson,O., 2002. Neuroinflammation of the nigrostriatal pathway during pro-gressive 6-OHDA dopamine degeneration in rats monitored by immu-nohistochemistry and PET imaging. Eur. J. Neurosci. 15, 991–998.
osta-Mattioli, M., Sossin, W.S., Klann, E., Sonenberg, N., 2009. Trans-lational control of long-lasting synaptic plasticity and memory. Neuron61, 10–26.
rawley, J., Goodwin, F.K., 1980. Preliminary report of a simple animalbehavior model for the anxiolytic effects of benzodiazepines. Pharma-col. Biochem. Behav. 13, 167–170.
roxford, J.L., Pryce, G., Jackson, S.J., Ledent, C., Giovannoni, G., Per-twee, R.G., Yamamura, T., Baker, D., 2008. Cannabinoid-mediatedneuroprotection, not immunosuppression, may be more relevant tomultiple sclerosis. J. Neuroimmunol. 193, 120–129.
e Filippis, D., Steardo, A., D’Amico, A., Scuderi, C., Cipriano, M.,Esposito, G., Iuvone, T., 2009. Differential cannabinoid receptor ex-pression during reactive gliosis: a possible implication for a nonpsy-chotropic neuroprotection. ScientificWorldJournal 9, 229–235.
e Lago, E., Fernandez-Ruiz, J., 2007. Cannabinoids and neuroprotectionin motor-related disorders. CNS Neurol. Disord. Drug Targets 6, 377–387.
exter, D.T., Carter, C.J., Wells, F.R., Javoy-Agid, F., Agid, Y., Lees, A.,Jenner, P., Marsden, C.D., 1989. Basal lipid peroxidation in substantianigra is increased in Parkinson’s disease. J. Neurochem. 52, 381–389.
hrhart, J., Obregon, D., Mori, T., Hou, H., Sun, N., Bai, Y., Klein, T.,Fernandez, F., Tan, J., Shytle, R.D., 2005. Stimulation of cannabinoidreceptor 2 (CB2) suppresses microglial activation. J. Neuroinflamma-
tion 2, 29.ernandez-Ruiz, J., 2009. The endocannabinoid system as a target for thetreatment of motor dysfunction. Br. J. Pharmacol. 156, 1029–1040.
ernandez-Ruiz, J., Pazos, M.R., Garcia-Arencibia, M., Sagredo, O.,Ramos, J.A., 2008. Role of CB2 receptors in neuroprotective effects ofcannabinoids. Mol. Cell. Endocrinol. 286 suppl 1, S91–S96.
ernandez-Ruiz, J., Romero, J., Velasco, G., Tolon, R.M., Ramos, J.A.,Guzman, M., 2007. Cannabinoid CB2 receptor: a new target for con-trolling neural cell survival? Trends Pharmacol. Sci. 28, 39–45.
aliegue, S., Mary, S., Marchand, J., Dussossoy, D., Carriere, D., Carayon,P., Bouaboula, M., Shire, D., Le Fur, G., Casellas, P., 1995. Expressionof central and peripheral cannabinoid receptors in human immunetissues and leukocyte subpopulations. Eur. J. Biochem. 232, 54–61.
alindo, M.F., Jordan, J., Gonzalez-Garcia, C., Cena, V., 2003. Chromaf-fin cell death induced by 6-hydroxydopamine is independent of mito-chondrial swelling and caspase activation. J. Neurochem. 84, 1066–1073.
arcia-Arencibia, M., Gonzalez, S., de Lago, E., Ramos, J.A., Mechoulam,R., Fernandez-Ruiz, J., 2007. Evaluation of the neuroprotective effectof cannabinoids in a rat model of Parkinson’s disease: importance ofantioxidant and cannabinoid receptor-independent properties. BrainRes. 1134, 162–170.
arcia-Gutierrez, M.S., Manzanares, J., 2010a. Overexpression of CB2cannabinoid receptor gene expression results in decreased vulnerabilityto anxiety and impaired action of alprazolam in mice. J. Psychophar-macol., September 13, 2010; doi: 10.1177/0269881110379507.
arcia-Gutierrez, M.S., Perez-Ortiz, J.M., Gutierrez-Adan, A., Man-zanares, J., 2010b. Depression-resistant endophenotype in mice over-expressing cannabinoid CB2 receptors. Br. J. Pharmacol. 160, 1773–1784.
iuffrida, A., Beltramo, M., Piomelli, D., 2001. Mechanisms of endocan-nabinoid inactivation: biochemistry and pharmacology. J. Pharmacol.Exp. Ther. 298, 7–14.
omez-Lazaro, M., Galindo, M.F., Melero-Fernandez de Mera, R.M.,Fernandez-Gomez, F.J., Concannon, C.G., Segura, M.F., Comella,J.X., Prehn, J.H., Jordan, J., 2007. Reactive oxygen species and p38mitogen-activated protein kinase activate Bax to induce mitochondrialcytochrome c release and apoptosis in response to malonate. Mol.Pharmacol. 71, 736–743.
omide, V.C., Silveira, G.A., Chadi, G., 2005. Transient and widespreadastroglial activation in the brain after a striatal 6-OHDA-induced partiallesion of the nigrostriatal system. Int. J. Neurosci. 115, 99–117.
ong, J.P., Onaivi, E.S., Ishiguro, H., Liu, Q.R., Tagliaferro, P.A., Brusco,A., Uhl, G.R., 2006. Cannabinoid CB2 receptors: immunohistochemi-cal localization in rat brain. Brain Res. 1071, 10–23.
runblatt, E., Mandel, S., Youdim, M.B., 2000. MPTP and 6-hydroxydo-pamine-induced neurodegeneration as models for Parkinson’s disease:neuroprotective strategies. J. Neurol. 247 Suppl 2, II95–II102.
undersen, H.J., Jensen, E.B., Kieu, K., Nielsen, J., 1999. The efficiencyof systematic sampling in stereology � reconsidered. J. Microsc. 193,199–211.
uzman, M., Sanchez, C., Galve-Roperh, I., 2001. Control of the cellsurvival/death decision by cannabinoids. J. Mol. Med. 78, 613–625.
erkenham, M., Lynn, A.B., Johnson, M.R., Melvin, L.S., de Costa, B.R.,Rice, K.C., 1991. Characterization and localization of cannabinoidreceptors in rat brain: a quantitative in vitro autoradiographic study.J. Neurosci. 11, 563–583.
erkenham, M., Lynn, A.B., Little, M.D., Johnson, M.R., Melvin, L.S., deCosta, B.R., Rice, K.C., 1990. Cannabinoid receptor localization inbrain. Proc. Natl. Acad. Sci. U. S. A. 87, 1932–1936.
ikosaka, O., Sesack, S.R., Lecourtier, L., Shepard, P.D., 2008. Habenula:Crossroad between the Basal Ganglia and the Limbic System. J. Neu-rosci. 28, 11825–11829.
owlett, A.C., Barth, F., Bonner, T.I., Cabral, G., Casellas, P., Devane,W.A., Felder, C.C., Herkenham, M., Mackie, K., Martin, B.R.,
Mechoulam, R., Pertwee, R.G., 2002. International Union of Pharma-
I
I
I
J
J
J
J
J
K
K
L
L
L
L
L
L
M
M
M
M
M
M
N
O
O
O
P
P
P
P
P
R
R
R
15A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
cology. XXVII. Classification of cannabinoid receptors. Pharmacol.Rev. 54, 161–202.
brahim, M.M., Porreca, F., Lai, J., Albrecht, P.J., Rice, F.L., Khodorova,A., Davar, G., Makriyannis, A., Vanderah, T.W., Mata, H.P., Malan,T.P., Jr, 2005. CB2 cannabinoid receptor activation produces antino-ciception by stimulating peripheral release of endogenous opioids.Proc. Natl. Acad. Sci. U. S. A. 102, 3093–3098.
uvone, T., Esposito, G., De Filippis, D., Bisogno, T., Petrosino, S.,Scuderi, C., Di Marzo, V., Steardo, L., 2007. Cannabinoid CB(1)receptor stimulation affords neuroprotection in MPTP-induced neuro-toxicity by attenuating S100B up-regulation in vitro. J. Mol. Med. 85,1379–1392.
zquierdo, I., Izquierdo, L.A., Barros, D.M., Mello e Souza, T., de Souza,M.M., Quevedo, J., Rodrigues, C., Sant’Anna, M.K., Madruga, M.,Medina, J.H., 1998. Differential involvement of cortical receptor mech-anisms in working, short-term and long-term memory. Behav. Phar-macol. 9, 421–427.
enner, P., Olanow, C.W., 1996. Oxidative stress and the pathogenesis ofParkinson’s disease. Neurology 47 suppl 3, S161–S170.
oosten, M., Valk, P.J., Jorda, M.A., Vankan-Berkhoudt, Y., Verbakel, S.,van den Broek, M., Beijen, A., Lowenberg, B., Delwel, R., 2002.Leukemic predisposition of pSca-1/Cb2 transgenic mice. Exp. Hematol.30, 142–149.
ordan, J., Galindo, M.F., Calvo, S., Gonzalez-Garcia, C., Cena, V., 2000.Veratridine induces apoptotic death in bovine chromaffin cells throughsuperoxide production. Br. J. Pharmacol. 130, 1496–1504.
ordan, J., Galindo, M.F., Tornero, D., Benavides, A., Gonzalez, C.,Agapito, M.T., Gonzalez-Garcia, C., Cena, V., 2002. Superoxide an-ions mediate veratridine-induced cytochrome c release and caspaseactivity in bovine chromaffin cells. Br. J. Pharmacol. 137, 993–1000.
ordan, J., Galindo, M.F., Tornero, D., Gonzalez-Garcia, C., Cena, V.,2004. Bcl-x L blocks mitochondrial multiple conductance channelactivation and inhibits 6-OHDA-induced death in SH-SY5Y cells.J. Neurochem. 89, 124–133.
outsilieri, E., Scheller, C., Grunblatt, E., Nara, K., Li, J., Riederer, P.,2002. Free radicals in Parkinson’s disease. J. Neurol. 249 Suppl 2,II1–II5.
ulisevsky, J., Pagonabarraga, J., 2009. Cognitive impairment in Parkin-son’s disease: tools for diagnosis and assessment. Mov. Disord. 24,1103–1110.
astres-Becker, I., Hansen, H.H., Berrendero, F., De Miguel, R., Perez-Rosado, A., Manzanares, J., Ramos, J.A., Fernandez-Ruiz, J., 2002.Alleviation of motor hyperactivity and neurochemical deficits by en-docannabinoid uptake inhibition in a rat model of Huntington’s disease.Synapse 44, 23–35.
astres-Becker, I., Molina-Holgado, F., Ramos, J.A., Mechoulam, R.,Fernandez-Ruiz, J., 2005. Cannabinoids provide neuroprotectionagainst 6-hydroxydopamine toxicity in vivo and in vitro: relevance toParkinson’s disease. Neurobiol. Dis. 19, 96–107.
everenz, J.B., Quinn, J.F., Zabetian, C., Zhang, J., Montine, K.S., Mon-tine, T.J., 2009. Cognitive impairment and dementia in patients withParkinson disease. Curr. Top. Med. Chem. 9, 903–912.
evine, D.S., 2007. Neural network modeling of emotion. Phys. Life Rev.4, 37–63.
ister, R.G., 1987. The use of a plus-maze to measure anxiety in themouse. Psychopharmacology 92, 180–185.
ivak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T))Method. Methods 25, 402–408.
anzanares, J., Corchero, J., Romero, J., Fernandez-Ruiz, J.J., Ramos,J.A., Fuentes, J.A., 1999. Pharmacological and biochemical interac-tions between opioids and cannabinoids. Trends Pharmacol. Sci. 20,287–294.
arsicano, G., Moosmann, B., Hermann, H., Lutz, B., Behl, C., 2002.Neuroprotective properties of cannabinoids against oxidative stress:
role of the cannabinoid receptor CB1. J. Neurochem. 80, 448–456.echoulam, R., Fride, E., Di Marzo, V., 1998. Endocannabinoids. Eur.J. Pharmacol. 359, 1–18.
olina-Holgado, E., Vela, J.M., Arevalo-Martin, A., Almazan, G., Molina-Holgado, F., Borrell, J., Guaza, C., 2002. Cannabinoids promote oli-godendrocyte progenitor survival: involvement of cannabinoid recep-tors and phosphatidylinositol-3 kinase/Akt signaling. J. Neurosci. 22,9742–9753.
onteiro, H.P., Winterbourn, C.C., 1989. 6-Hydroxydopamine releasesiron from ferritin and promotes ferritin-dependent lipid peroxidation.Biochem. Pharmacol. 38, 4177–4182.
organ, N.H., Stanford, I.M., Woodhall, G.L., 2009. Functional CB2 typecannabinoid receptors at CNS synapses. Neuropharmacology 57, 356–368.
ieoullon, A., Coquerel, A., 2003. Dopamine: a key regulator to adaptaction, emotion, motivation and cognition. Curr. Opin. Neurol. 16,S3–S9.
naivi, E.S., Ishiguro, H., Gong, J.P., Patel, S., Meozzi, P.A., Myers, L.,Perchuk, A., Mora, Z., Tagliaferro, P.A., Gardner, E., Brusco, A.,Akinshola, B.E., Hope, B., Lujilde, J., Inada, T., Iwasaki, S., Macharia,D., Teasenfitz, L., Arinami, T., Uhl, G.R., 2008. Brain neuronal CB2cannabinoid receptors in drug abuse and depression: from mice tohuman subjects. PLoS One 3, e1640.
naivi, E.S., Ishiguro, H., Gong, J.P., Patel, S., Perchuk, A., Meozzi, P.A.,Myers, L., Mora, Z., Tagliaferro, P., Gardner, E., Brusco, A., Akin-shola, B.E., Liu, Q.R., Hope, B., Iwasaki, S., Arinami, T., Teasenfitz,L., Uhl, G.R., 2006. Discovery of the presence and functional expres-sion of cannabinoid CB2 receptors in brain. Ann. N. Y. Acad. Sci.1074, 514–536.
rtiz, S., Oliva, J.M., Perez-Rial, S., Palomo, T., Manzanares, J., 2004.Chronic ethanol consumption regulates cannabinoid CB1 receptor geneexpression in selected regions of rat brain. Alcohol Alcohol. 39, 88–92.
alazuelos, J., Aguado, T., Pazos, M.R., Julien, B., Carrasco, C., Resel, E.,Sagredo, O., Benito, C., Romero, J., Azcoitia, I., Fernandez-Ruiz, J.,Guzman, M., Galve-Roperh, I., 2009. Microglial CB2 cannabinoidreceptors are neuroprotective in Huntington’s disease excitotoxicity.Brain 132, 3152–3164.
alkovits, M., 1983. Punch sampling biopsy technique. Methods Enzymol.103, 368–376.
axinos, G., Franklin, K.B.J., 2001. The Mouse Brain in StereotaxicCoordinates. Academic Press, Harcourt Science and Technology Com-pany, New York.
erez-Rial, S., Garcia-Gutierrez, M.S., Molina, J.A., Perez-Nievas, B.G.,Ledent, C., Leiva, C., Leza, J.C., Manzanares, J., 2009. Increasedvulnerability to 6-hydroxydopamine lesion and reduced developmentof dyskinesias in mice lacking CB1 cannabinoid receptors. Neurobiol.Aging, doi:10.1016/j.neurobiolaging.2009.03.017.
rice, D.A., Martinez, A.A., Seillier, A., Koek, W., Acosta, Y., Fernandez,E., Strong, R., Lutz, B., Marsicano, G., Roberts, J.L., Giuffrida, A.,2009. WIN55,212–2, a cannabinoid receptor agonist, protects againstnigrostriatal cell loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-dine mouse model of Parkinson’s disease. Eur. J. Neurosci. 29, 2177–2186.
acz, I., Nadal, X., Alferink, J., Banos, J.E., Rehnelt, J., Martin, M.,Pintado, B., Gutierrez-Adan, A., Sanguino, E., Bellora, N., Man-zanares, J., Zimmer, A., Maldonado, R., 2008a. Interferon-gamma is acritical modulator of CB(2) cannabinoid receptor signaling duringneuropathic pain. J. Neurosci. 28, 12136–12145.
acz, I., Nadal, X., Alferink, J., Banos, J.E., Rehnelt, J., Martin, M.,Pintado, B., Gutierrez-Adan, A., Sanguino, E., Manzanares, J., Zim-mer, A., Maldonado, R., 2008b. Crucial role of CB(2) cannabinoidreceptor in the regulation of central immune responses during neuro-pathic pain. J. Neurosci. 28, 12125–12135.
amirez, B.G., Blazquez, C., Gomez del Pulgar, T., Guzman, M., deCeballos, M.L., 2005. Prevention of Alzheimer’s disease pathology bycannabinoids: neuroprotection mediated by blockade of microglial ac-
tivation. J. Neurosci. 25, 1904–1913.
R
R
S
S
T
T
T
V
V
W
W
W
W
X
Y
Z
Z
16 A. Ternianov et al. / Neurobiology of Aging xx (2010) xxx
odrigues, R.W.P., Gomide, V.C., Chadi, G., 2001. Astroglial and micro-glial reaction after a partial nigrostriatal degeneration induced by thestriatal injection of different doses of 6-hydroxydopamine. Int. J. Neu-rosci. 109, 91–126.
odrigues, R.W.P., Gomide, V.C., Chadi, G., 2004. Astroglial and micro-glial activation in the Wistar rat ventral tegmental area after a singlestriatal injection of 6-hydroxydopamine. Int. J. Neurosci. 114, 197–216.
agredo, O., Gonzalez, S., Aroyo, I., Pazos, M.R., Benito, C., Lastres-Becker, I., Romero, J.P., Tolon, R.M., Mechoulam, R., Brouillet, E.,Romero, J., Fernandez-Ruiz, J., 2009. Cannabinoid CB2 receptor ago-nists protect the striatum against malonate toxicity: relevance for Hun-tington’s disease. Glia 57, 1154–1167.
heng, W.S., Hu, S., Ni, H.T., Rock, R.B., Peterson, P.K, 2009.WIN55,212-2 inhibits production of CX3CL1 by human astrocytes:involvement of p38 MAP kinase. J Neuroimmune Pharmacol. 4, 244–248.
agliaferro, P., Javier Ramos, A., Onaivi, E.S., Evrard, S.G., Lujilde, J.,Brusco, A., 2006. Neuronal cytoskeleton and synaptic densities arealtered after a chronic treatment with the cannabinoid receptor agonistWIN 55,212-2. Brain Res. 1085, 163–176.
arozzi, A., Morroni, F., Merlicco, A., Hrelia, S., Angeloni, C., Cantelli-Forti, G., Hrelia, P., 2009. Sulforaphane as an inducer of glutathioneprevents oxidative stress-induced cell death in a dopaminergic-likeneuroblastoma cell line. J. Neurochem. 111, 1161–1171.
isch, S., Silberstein, P., Limousin-Dowsey, P., Jahanshahi, M., 2004. Thebasal ganglia: anatomy, physiology, and pharmacology. Psychiatr.Clin. North Am. 27, 757–799.
an Sickle, M.D., Duncan, M., Kingsley, P.J., Mouihate, A., Urbani, P.,Mackie, K., Stella, N., Makriyannis, A., Piomelli, D., Davison, J.S.,
Marnett, L.J., Di Marzo, V., Pittman, Q.J., Patel, K.D., Sharkey, K.A.,2005. Identification and functional characterization of brainstemcannabinoid CB2 receptors. Science 310, 329–332.
iscomi, M.T., Oddi, S., Latini, L., Pasquariello, N., Florenzano, F., Bernardi,G., Molinari, M., Maccarrone, M., 2009. Selective CB2 receptor agonismprotects central neurons from remote axotomy-induced apoptosis throughthe PI3K/Akt pathway. J. Neurosci. 29, 4564–4570.
aksman, Y., Olson, J.M., Carlisle, S.J., Cabral, G.A., 1999. The centralcannabinoid receptor (CB1) mediates inhibition of nitric oxide produc-tion by rat microglial cells. J. Pharmacol. Exp. Ther. 288, 1357–1366.
alter, L., Franklin, A., Witting, A., Wade, C., Xie, Y., Kunos, G.,Mackie, K., Stella, N., 2003. Nonpsychotropic cannabinoid receptorsregulate microglial cell migration. J. Neurosci. 23, 1398–1405.
alter, L., Stella, N., 2004. Cannabinoids and neuroinflammation. Br. J.Pharmacol. 141, 775–785.
est, M.J., Slomianka, L., Gundersen, H.J., 1991. Unbiased stereologicalestimation of the total number of neurons in thesubdivisions of the rathippocampus using the optical fractionator. Anat. Rec. 231, 482–497.
iao, D., Zikopoulos, B., Barbas, H., 2009. Laminar and molecular orga-nization of prefrontal projections to multiple thalamic nuclei. Neuro-science 161, 1067–1081.
iangou, Y., Facer, P., Durrenberger, P., Chessell, I.P., Naylor, A., Boun-tra, C., Banati, R.R., Anand, P., 2006. COX-2, CB2 and P2X7-immu-noreactivities are increased in activated microglial cells/macrophagesof multiple sclerosis and amyotrophic lateral sclerosis spinal cord.BMC Neurol. 6, 12.
eevalk, G.D., Razmpour, R., Bernard, L.P., 2008. Glutathione and Par-kinson’s disease: is this the elephant in the room? Biomed. Pharmaco-ther. 62, 236–249.
huang, S.Y., Bridges, D., Grigorenko, E., McCloud, S., Boon, A., Hamp-son, R.E., Deadwyler, S.A., 2005. Cannabinoids produce neuroprotec-tion by reducing intracellular calcium release from ryanodine-sensitive
stores. Neuropharmacology 48, 1086–1096.


![arXiv:1404.3757v2 [cs.SI] 25 Oct 2014](https://static.fdokumen.com/doc/165x107/63216b5fb71aaa142a043bed/arxiv14043757v2-cssi-25-oct-2014.jpg)













![arXiv:1710.09223v1 [cond-mat.mes-hall] 25 Oct 2017](https://static.fdokumen.com/doc/165x107/631923570255356abc08230e/arxiv171009223v1-cond-matmes-hall-25-oct-2017.jpg)


