
(2010) A taxonomic revision of the tarantula spider genus Lyrognathus Pocock 1895 (Araneae,...
43
Accepted by R. Raven: 14 Dec. 2009; published: 18 Feb. 2010 1 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2010 · Magnolia Press Zootaxa 2362: 1–43 (2010) www.mapress.com/ zootaxa/ Article A taxonomic revision of the tarantula spider genus Lyrognathus Pocock 1895 (Araneae, Theraphosidae), with notes on the Selenocosmiinae RICK C. WEST 1 & STEVEN C. NUNN 2 1 3436 Blue Sky Place, Victoria, BC, Canada V9C 3N5. E-mail: [email protected] 2 43 Range Road, Sarina, Qld, 4737, Australia. E-mail: [email protected] Table of contents Abstract ............................................................................................................................................................................... 1 Introduction ......................................................................................................................................................................... 2 Material and methods ......................................................................................................................................................... 2 Taxonomy ............................................................................................................................................................................ 5 Selenocosmiinae Simon 1889 ............................................................................................................................................. 5 Lyrognathus Pocock 1895 ................................................................................................................................................... 5 Identification key for adult females of Lyrognathus .......................................................................................................... 6 Lyrognathus crotalus Pocock 1895 ............................................................................................................................. 6 Lyrognathus achilles sp. nov. .................................................................................................................................... 11 Lyrognathus fuscus sp.nov. ........................................................................................................................................ 19 Lyrognathus lessunda sp. nov. ................................................................................................................................... 25 Lyrognathus robustus Smith 1988 ............................................................................................................................. 31 Lyrognathus saltator Pocock 1900 ............................................................................................................................ 36 Character discussion .......................................................................................................................................................... 38 Acknowledgements .......................................................................................................................................................... 42 References ......................................................................................................................................................................... 42 Abstract The tarantula genus Lyrognathus Pocock 1895 is revised with three new species described: L. achilles sp. nov. from Gunung Nibung, Borneo; L. fuscus sp. nov. from Simpang Kubur, Borneo and L. lessunda sp. nov. from Lombok Island, Indonesia. The male of L. robustus Smith 1988 is described for the first time, the female is redescribed. L. crotalus Pocock 1895 is considered a senior synonym of L. pugnax Pocock 1900. Several new characters are discussed, with Lyrognathus keyed to species. The diagnosis of the Selenocosmiinae is amended. Zoogeographic distribution, natural history and subfamilial relationships with proposed sister genera Selenocosmia Ausserer 1871 and Coremiocnemis Simon 1892 are discussed. Lyrognathus saltator is redescribed. Selenocosmia lyra Strand 1913 is considered a nomen dubium. Key words: Mygalomorph, new species, Lombok, Malaysia, India, Borneo, Indonesia, distribution, zoogeography, natural history
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of (2010) A taxonomic revision of the tarantula spider genus Lyrognathus Pocock 1895 (Araneae,...

ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2010 · Magnolia Press
Zootaxa 2362: 1–43 (2010) www.mapress.com/zootaxa/ Article
A taxonomic revision of the tarantula spider genus Lyrognathus Pocock 1895 (Araneae, Theraphosidae), with notes on the Selenocosmiinae
RICK C. WEST1 & STEVEN C. NUNN2 13436 Blue Sky Place, Victoria, BC, Canada V9C 3N5. E-mail: [email protected] Range Road, Sarina, Qld, 4737, Australia. E-mail: [email protected]
Table of contents
Abstract ............................................................................................................................................................................... 1Introduction ......................................................................................................................................................................... 2Material and methods ......................................................................................................................................................... 2Taxonomy ............................................................................................................................................................................ 5Selenocosmiinae Simon 1889 ............................................................................................................................................. 5Lyrognathus Pocock 1895 ................................................................................................................................................... 5Identification key for adult females of Lyrognathus .......................................................................................................... 6
Lyrognathus crotalus Pocock 1895 ............................................................................................................................. 6Lyrognathus achilles sp. nov. .................................................................................................................................... 11Lyrognathus fuscus sp.nov. ........................................................................................................................................ 19Lyrognathus lessunda sp. nov. ................................................................................................................................... 25Lyrognathus robustus Smith 1988 ............................................................................................................................. 31Lyrognathus saltator Pocock 1900 ............................................................................................................................ 36
Character discussion .......................................................................................................................................................... 38Acknowledgements .......................................................................................................................................................... 42References ......................................................................................................................................................................... 42
Abstract
The tarantula genus Lyrognathus Pocock 1895 is revised with three new species described: L. achilles sp. nov. from Gunung Nibung, Borneo; L. fuscus sp. nov. from Simpang Kubur, Borneo and L. lessunda sp. nov. from Lombok Island, Indonesia. The male of L. robustus Smith 1988 is described for the first time, the female is redescribed. L. crotalusPocock 1895 is considered a senior synonym of L. pugnax Pocock 1900. Several new characters are discussed, with Lyrognathus keyed to species. The diagnosis of the Selenocosmiinae is amended. Zoogeographic distribution, natural history and subfamilial relationships with proposed sister genera Selenocosmia Ausserer 1871 and CoremiocnemisSimon 1892 are discussed. Lyrognathus saltator is redescribed. Selenocosmia lyra Strand 1913 is considered a nomen dubium.
Key words: Mygalomorph, new species, Lombok, Malaysia, India, Borneo, Indonesia, distribution, zoogeography, natural history
Accepted by R. Raven: 14 Dec. 2009; published: 18 Feb. 2010 1

Introduction
Lyrognathus Pocock 1895 belongs to the subfamily Selenocosmiinae Simon 1889 of the mygalomorph family Theraphosidae Thorell 1869 and was formerly known from northeastern India (three species) and West Malaysia (one species). Pocock described Lyrognathus crotalus Pocock 1895 from Assam, northeastern India. This spider was unique amongst all known selenocosmiine species at that time in that leg IV was very incrassate with undivided scopula on metatarsi IV that covered the full length ventrally. This new character combination suggested to Pocock (1895) the presence of a new genus, L. crotalus being the type species by monotypy and original designation. Shortly thereafter, Pocock described two more species of Lyrognathus, L. saltator Pocock 1900 from North Khasi Hills and L. pugnax Pocock 1900 from Shillong, Meghalaya State (formerly Assam State before 1972), India. Additionally, Pocock (1900) revised the genus, keyed the group in a review of Indian theraphosid genera and keyed Lyrognathus to species level.
Simon (1903) followed Pocock’s work in a complete key to Selenocosmiinae genera. The first male of Lyrognathus was described and named Selenocosmia pugnax (Gravely 1935). Gravely (1935) suggested all Lyrognathus might best fit into Selenocosmia Ausserer 1871. Earlier, Gravely (1915b) had also identified a non-type male of L. pugnax from the Garo Hills, and a non-type male of L. crotalus from the North Khasi Hills (det. Hirst) in Meghalaya State and at that time, questioned the validity of Pocock’s three species. However, Gravely had not examined Pocock’s types, so his suggestion was largely ignored. Roewer (1942) listed Lyrognathus as did Bonnet (1957); Brignoli (1983) followed from Roewer. Raven (1985) reviewed the Theraphosidae and keyed the genus in his systematic overview of Mygalomorphae. Smith (1986, 1987) reviewed Lyrognathus in his work on Theraphosidae and later (Smith, 1988) described L. robustus Smith 1988, from the Cameron Highlands, Pahang, West Malaysia. That work enlarged our knowledge of the distribution range of the previously considered endemic Indian genus. L. liewi West 1991 was described from Pahang, Perak and Kedah States in West Malaysia. Schmidt & von Wirth (1992) synonymised L. liewi into L. robustus, based on geographic locality and similarities of morphology from original descriptions.
Among Selenocosmiinae, Lyrognathus is one of the most widespread genera, with a range similar to that of Chilobrachys Karsch 1891 and appears to be a highland dwelling group, with all species found at high elevations.
With the inclusion here of three new species and L. robustus into Lyrognathus, along with the new synonymy, herein, of L. pugnax into L. crotalus, a new key for all species of Lyrognathus is given.
Material and methods
Taxonomic description style follows Raven (2005) and West et al. (2008). All measurements are given in millimeters (mm) and were made to the nearest 0.01 mm with the aid of a squared eyepiece and digital dial callipers. Imaging was taken by mounting a Nikon CP8800 directly to the eyepiece of a WILD Heerbrugg M3Z PLAN stereozoom microscope via the Scopetronix Maxview TM system, with external cold light illumination. Each species diagnosis defines each species from all others within the genus, not just its closest relative. Leg and pedipalp measurements were taken from their dorsal aspect on the left side (unless appendages were lost or obviously regenerated) of all specimens and coxae and trochantera were measured from their ventral aspect, on the same side. Eyes in sizes of largest to smallest (including distances taken at widest points in length/width, respectively). Claws are not included in measurements of tarsi. Total body length measurements include chelicerae. Sternal sigilla measurements do not include labio-sternal sigilla. Hair types of new species are described. Spermathecae were dissected from females and cleaned in concentrated lactic acid for 24 hours. The Leg formula is given in order of size from largest to smallest in both length and width. Leg relation factor [RF~] (following von Wirth & Striffler 2005) is calculated by multiplying the length of leg I by 100, then dividing by the length of leg IV. 100 = equal sized leg I–IV, above 100 on a sliding scale represents a longer leg I, below 100 on a sliding scale represents a longer leg IV. Trichobothrial terminology follows Raven (2005). Scopula extents are given as distal extents, i.e., 2/3 means for distal two
WEST & NUNN2 · Zootaxa 2362 © 2010 Magnolia Press

thirds and refer to the ventral extent. DIVA-GIS Version 5.4.0.1 was utilized for geographic plotting (Fig. 114). All newly deposited type material is housed at the QM. Because mature females display more informative characters in Lyrognathus than do males, females were selected as holotypes. All previous types from this genus are female and some males are not known, therefore, the dichotomous key applies only to females.
Material from the following institutions were cited, examined or images of type material were supplied to the authors: IMC—Indian Museum, Calcutta; MNHN—Muséum National d’Histoire Naturelle, Paris; NHM—Natural History Museum, London; NMW—Naturhistorisches Museum, Wien; NRM—Naturhistoriska Riksmuseet, Stockholm; NUS—National University of Singapore, Singapore; QM—Queensland Museum, Brisbane; RMBR—Raffles Museum of Biodiversity Research, Singapore; SMF—Senckenberg Museum, Frankfurt–am–Main; ZMH—Zoologisches Institut und Zoologisches Museum, Universitat Hamburg, Hamburg; ZMUC—Zoological Museum University of Copenhagen, Copenhagen; ZSI—Zoological Survey of India, Calcutta.
Abbreviations: cox.—coxa, tro.—trochanter, fem.—femur, pat.—patella, tib.—tibia, met.—metatarsus, tar.—tarsus, AME—anterior median eyes, ALE—anterior lateral eyes, PME—posterior median eyes, PLE—posterior lateral eyes, OT—ocular tubercle, P—prolateral, R—retrolateral, D—dorsal, V—ventral, DV—distoventral, DL—distolateral, DD—distodorsal, DPD—distal prodorsal, DRD—distal retrodorsal, DPL—distal prolateral, DPV—distal proventral, DRV—distal retroventral. PMS—posterior median spinnerets, PLS—posterior lateral spinnerets, PSS—posterior sternal sigilla.
Material. Type material: Chilobrachys andersoni (Pocock 1895), male paratype, 1891.10.15.6, India, NHM.—images (R. Raven, pers. comm.).
Chilobrachys flavopilosus (Simon, 1884) female paratype, 79/25, Thayetmyo (19°19' N, 95°10' E), Myanmar, NHMS.—images (R. Raven, pers. comm.).
Chilobrachys sericeus (Thorell 1887), male syntype, 1895.9.21.21, Myanmar, NHM.—images (R. Raven, pers. comm.).
Coremiocnemis cunicularia (Simon 1892), holotype jar with 1 female specimen, 1 juvenile, AR4547, Penang Hill (= Bukit Bendera, 5°25' N, 100°16' E), Penang Island, Penang, West Malaysia, MNHN.—images (F. Vol, pers. comm.).
Coremiocnemis valida Pocock 1895, female holotype, 60.15, East Indies, NHM.–images (R. Raven, pers. comm.)–examined.
Orphnaecus pellitus Simon 1892, male holotype, AR4678, Calapnitan Caves (= Kulapnitan Caves, 13°45' N, 122°58' E), Camarines Sur Province, Luzon Island, Philippines, MNHN.—images (F. Vol, pers. comm.).
Phlogiellus atriceps Pocock 1897, male holotype and 1 female paratype, 1897.11.1.6-7, Buitenzorg (= Bogor, 6°36′ S, 106°48′ E), Java, NHM. —images (R. Raven, pers. comm.).—examined.
Phlogiellus baeri (Simon 1877), female holotype, AR 46711146, Manila (14°35′ N, 120°58′ E), Luzon Island, Philippines, MNHN.—images (R. Raven, pers. comm.).
Selenocosmia aruana Strand 1911, male holotype, 2685-84, Aru Island (6°10′ S, 134°30′ E), Indonesia, SMF.—images (R. Raven, pers. comm.).
Selenocosmia deliana Strand 1913, female holotype, 2688-84, Sumatra, Indonesia, SMF.—images (R. Raven, pers. comm.).
Selenocosmia hirtipes Strand 1913, female syntype, 2690-84, Moluccas (2°00′ S, 128°00′ E), Indonesia, SMF.—images (R. Raven, pers. comm.).
Selenocosmia kovariki (Schmidt & Krause 1995), female holotype, 37350-84, Tam Dao (21°27' N, 105°38' E), Vietnam, SMF.—images (R. Raven, pers. comm.).
Selenocosmia lyra Strand 1913, juvenile male holotype (new det.), 2691-84, Sumatra, Indonesia, SMF.—images (P. Jaeger, R. Raven, pers. comm.)—examined.
Selenocosmia orophila (Thorell 1897), juvenile holotype, 597, Carin–Cheba or Bia-Po (= Karen Mountains, 19°16' N, 96°35' E), Toungoo District, Karen State, Myanmar, ZMH.—images (R. Raven, pers. comm.).
Zootaxa 2362 © 2010 Magnolia Press · 3REVISION OF TARANTULA GENUS LYROGNATHUS

Selenocosmia peerboomi (Schmidt 1999), female holotype, 39185-84, Negros Island (10°0' N, 123 °0' E), Philippines, SMF.—images (R. Raven, pers. comm.).
Selenocosmia strubelli Strand 1913, syntype female syntype, 2694-84, Java, Indonesia, SMF.—images (R. Raven, pers. comm.).
Selenocosmia tahanensis Abraham 1924, male holotype, 1924.2.27.22, Gunung Tahan (4°38' N, 102°14' E), Pahang, West Malaysia NHM.—images (J. Beccaloni, pers. comm.)—examined.
Selenotholus foelschei Hogg 1902, female holotype, 1924.111.1.22, Palmerston (12°27' S, 130°57' E), Northern Territory, Australia, NHM.—illustrations (A. Smith, pers. comm.).
Selenotypus plumipes Pocock 1895, female holotype, 1892.5.15.1, Majors Creek (19°40' S, 147°4' E), Queensland, Australia, NHM.—images and illustrations (R. Raven, A. Smith, pers. comm.). Other material: Chilobrachys fumosus Pocock 1895, 2 males, S83830 and S83831, Ri-Bhoi District (25°55' N, 91°53' E), Meghalaya State, India, QM.—examined.
Chilobrachys huahini Schmidt and Huber 1996, 2 females, 2 males, S83774, S8376, S83770 and S83780, respectively, Thailand, QM.—examined.
Chilobrachys sericeus (Thorell 1895), 1 male, S74077, Northwest Thailand, QM.—examined. Coremiocnemis cunicularia (Simon 1892), 2 females, S83742 and S83744, Penang Hill, Penang Island,
Penang, West Malaysia, QM.—examined. Coremiocnemis sp. West & Nunn (in press), 2 females, 1 male, S83826, S83825 and QM S83927,
respectively, Gunung Ledang (2°22' N, 102°36' E), Johor, West Malaysia, QM.—examined.Coremiocnemis sp. West & Nunn (in press), 1 female, ZRC ARA.745, Fraser’s Hill (3°43' N, 101°45' E),
West Malaysia, RMBR.—images (D. Court, pers. comm.)—examined. Coremiocnemis sp. West & Nunn (in press), 3 females, 1 male, 2 female juveniles S74074, S88076,
S88078, S88022, S88077 and S88079, respectively, Fraser’s Hill, Selangor, West Malaysia, QM—examined; 3 females ZRC ARA.741, ZRC ARA.67 and ZRC ARA.756, respectively, Fraser’s Hill, Selangor, West Malaysia, RMBR (Gopal series).—images (D. Court, pers comm.).
Coremiocnemis sp. West & Nunn (in press), 3 males, S83823, S83821 and S83824 from Tapah (4°06' N, 101°25' E), Perak, West Malaysia, QM—examined; 1 male, ZRC ARA.743, Tapah, Perak, West Malaysia from Pahang, West Malaysia, RMBR.—images (D. Court, pers. comm.)—examined.
Coremiocnemis sp. West & Nunn (in press), 1 female, S83759, Ketambe Research Station (3º41´ N, 97º 39´ E), Gunung Leuser National Park, Aceh Teneggara, Sumatra, QM.—examined.
Coremiocnemis sp. West & Nunn (in press), 5 females, S83745, S83747, S83748, S83750 and S88749, respectively, Mile 19 (4°20' N, 101°20' E), Cameron Highlands, Perak, West Malaysia, QM—examined; 1 female, 1 juvenile, ZRC ARA.742, Renglet Camp (= Ringlet, 4°26’ N, 101°23' E), Cameron Highlands, RMBR.—images (D. Court, pers. comm.)—examined.
Haplocosmia nepalensis Schmidt and von Wirth 1996, 3 males, S83827, S83828 and S83829, Katmandu (27°42' N, 85°19' E), Nepal, QM.—examined.
Orphnaecus pellitus Simon 1892, 1 female, S83791, Sorsogon Province (12°50′ N, 123°55′ E), Luzon Island, Philippines, QM.—examined.
Phlogiellus inermis (Ausserer 1871), 1 male, 1 female, S83766 and S83767, respectively, Kent Ridge (1°17' N, 103°46' E), Singapore, QM.—examined.
Selenobrachys philippinus Schmidt 1999, 1 female, S83833, Sipalay (9°45' N, 122°26' E), Negros Island, Philippines, QM.—examined.
Selenocosmia obscura Hirst 1909, 1 male, S83832, Kota Belud (6°21' N, 116°26' E), Sabah, Borneo, QM.—examined.
Selenocosmia sp. 1 female, S74072, Vietnam, QM.—examined.Selenocosmia sp. (javanensis?) 1 female, S83754, Bogor Botanical Gardens (6°35' N, 106°47' E), Bogor,
Java, Indonesia, QM.—examined.Selenocosmia sp. 1 female, S74078, Thailand, QM.—examined.Yamia watasei Kishida 1920, 1 male, S83757, Lanyu (= Orchid) Island (22°30' N, 121°31' E), Taiwan,
QM.—examined.
WEST & NUNN4 · Zootaxa 2362 © 2010 Magnolia Press

Taxonomy
Selenocosmiinae Simon 1889
Selenocosmiae Simon 1889: 204; Simon 1892: 147; Raven 1985: 118.Poecilotherieae Simon 1889: 204; Simon 1892: 144.Phlogieae Simon 1892: 144. Synonymised by Simon 1903: 953.
Diagnosis (modified from Raven 1985, 2005): With oval patch of modified setae (often bacillate) on prolateral maxillary face and originating medially, not ventrally from the oral fringe, opposing strong spiniform setae (strikers) on ectal cheliceral surface to stridulate: maxillary lyra reduced or absent in some genera, cheliceral strikers retained, sternal sigilla largely removed from sternal margins, many labial cuspules (>300), leg spines found only distally on metatarsi and tibial apophyses on leg I of mature males absent.
Genera included:
Chilobrachys Karsch 1891Coremiocnemis Simon 1892Haplocosmia Schmidt & von Wirth 1996Lyrognathus Pocock 1895Orphnaecus Simon 1892Phlogiellus Pocock 1897Poecilotheria Simon 1885Selenobrachys Schmidt 1999Selenocosmia Ausserer 1871Selenotholus Hogg 1902Selenotypus Pocock 1895Yamia Kishida 1920
Remarks: With the transfer of the Psalmopoeus Pocock 1895 out of the Selenocosmiinae and into the Aviculariinae Simon 1874 (West et al. 2008), the need to redefine the stridulatory morphology used in the selenocosmiine subfamily diagnosis was urgent and is proposed herein. Additionally, in species with maxillary lyra reduced to absent (viz., Phlogiellus baeri, Yamia watasei), the opposing cheliceral strikers are retained. The new inclusion of diagnostic leg spination in Selenocosmiinae sheds light on the placement of the Selenogyrinae Smith 1990 which might best be placed within the Ischnocolinae (Simon 1892).
Lyrognathus Pocock 1895(Figs 1–114)
Lyrognathus Pocock 1895: 170, 175, 1900: 187, 202; Simon 1903: 954, 956; Gravely, 1915a: 415; 1915b: 284; 1935: 83, Roewer, 1942: 6; Raven 1985: 38, 118; Smith 1986, 1987: 121; Platnick, 2009.
Diagnosis: Lyrognathus differs from other Selenocosmiinae genera in having dense penicillate retrolateral setal brushes along tibiae and metatarsi IV (Figs 3, 20, 54, 71, 98), very incrassate tibiae IV (Figs 8, 21, 53, 71, 99), retrolateral surface of coxae IV entirely hirsute (except L. robustus) and undivided metatarsi IV scopula in females (Figs 19, 54, 74), except L. robustus, which shows a possibly unique form of division of the metatarsi IV scopula by extremely stout, spiniform setae (Fig. 98). Lyrognathus further differs in that the male embolus is entirely lanceolate with a singular prolateral keel extending ventrally and proximally (Figs 5, 40, 89, 90). Differs from Selenocosmia from Java and Sumatra and from Coremiocnemis from Malaysia (males and females) in the above traits, in the large size of posterior sternal sigilla and that the clavate
Zootaxa 2362 © 2010 Magnolia Press · 5REVISION OF TARANTULA GENUS LYROGNATHUS

trichobothrial field on tarsi I–IV occur in distal 1/2 (in Malaysian, Javan and Sumatran species of Selenocosmia and Coremiocnemis, clavates extend to distal 3/4 of all tarsi), and general leg morphology as diagnosed above.
Type species: Lyrognathus crotalus Pocock 1895, by original designation and monotypy.Description: A selenocosmiine theraphosid genus with incrassate leg IV combined with penicillate
retrolateral setal brushes in both sexes (except in male L. robustus), leg length formula IV, I, II, III (both sexes), leg width formula IV, III, I, II (females), circular ocular tubercle–higher posteriorly, with fovea of equal width, clypeus absent, retrolateral surface of coxae IV entirely hirsute (except in L. robustus), >2 DV, 1DPV, 1DRV, 1DD metatarsal III/IV spine arrangement, no basofemoral spines, retrolateral basomedial spine cluster on chelicerae (present/absent), keeled/terete lanceolate embolus with small basoventral projection present/absent, intercheliceral peg or spiniform setae, bilobular spermathecae with lateral lobes at 2/3 to 3/4 length of medials (most species) or unilobular spermathecal receptacles (L. crotalus), spermathecae heavily sclerotized distally, grading to clear shafts proximally, metatarsi IV scopula divided (L. robustus) or undivided, metatarsi IV scopula in distal 1/3 (L. robustus) or for full length, unarmed paired claws, third claw on leg IV present (L. robustus) or absent (all other species) and clavate trichobothria on distal 1/2 of all tarsi, very long. No dorsal carapace thorns.
Species included: Lyrognathus crotalus Pocock 1895Lyrognathus achilles sp. nov.Lyrognathus fuscus sp. nov. Lyrognathus lessunda sp. nov. Lyrognathus robustus Smith 1988 Lyrognathus saltator Pocock 1900
Distribution: Northeastern India, West Malaysia, Borneo and Lombok Island, Indonesia (Fig. 114)
Identification key for adult females of Lyrognathus
1 Scopula on metatarsi IV divided by band of strong spiniform setae (Fig. 98) ........................................... L. robustus- Scopula on metatarsi IV undivided (Fig. 19)................................................................................................................ 22 Basodorsal intercheliceral surface with strong spiniform setae (Fig. 10) .................................................................... 3- Basodorsal intercheliceral surface with peg shaped thorns (Fig. 28) ........................................................................... 43 Metatarsi IV with 6 apical spines ................................................................................................................. L. saltator- Metatarsi IV with 5 apical spines ................................................................................................................. L. crotalus4 Disto-medial lobe of spermathecae as in lateral lobe, “mushroom cap” form (Fig. 70) ............... L. lessunda sp. nov.- Disto-medial lobe of spermathecae not as in lateral lobe, not “mushroom” cap form (Figs 23, 51) .......................... 55 Retrolateral tibiae IV brush uniform along entire surface (Fig. 20) ............................................... L. achilles sp. nov.- Retrolateral tibiae IV brush lacking proximally, increasing distally (Fig. 53) ................................. L. fuscus sp. nov.
Lyrognathus crotalus Pocock 1895 (Figs 1–12, 114)
Lyrognathus crotalus Pocock 1895: 175; Platnick, 2009.Lyrognathus pugnax Pocock 1900: 203; Platnick, 2009. New synonymy.
Types: Lyrognathus crotalus Pocock 1895, female holotype, 1854.16, Assam State, northeastern India, NHM.—images and illustrations (R. Raven, A. Smith, V. von Wirth, pers. comm.).
WEST & NUNN6 · Zootaxa 2362 © 2010 Magnolia Press

FIGURES 1–2. 1. Lyrognathus crotalus Pocock, female in life. 2. L. crotalus, male in life. Photos: J-M. Verdez.
Zootaxa 2362 © 2010 Magnolia Press · 7REVISION OF TARANTULA GENUS LYROGNATHUS

FIGURES 3–10. 3. Lyrognathus species. L. crotalus Pocock, female habitus, dorsal view. 4. L. pugnax, holotype female habitus, dorsal view. 5. L. crotalus, male embolus, prolateral view. 6. L. crotalus, paratype female spermathecae, dorsal view. 7. L. crotalus, holotype maxillae, prolateral view. 8. L. crotalus, holotype right tib. IV, dorsal view. 9. L. crotalus, holotype chelicerae, ectal view showing strikers. 10. L. crotalus, holotype chelicerae, prolateral view. Scale bar = 10mm for 3, 4; 1mm for 5; 0.5mm for 6. Figs: 3–6: S. C. Nunn; Photos: 7–10, R. Raven.
Lyrognathus pugnax Pocock 1900, female holotype, 1895.1.28.1, Shillong (25°34' N, 91°53' E), Meghalaya State, northeastern India, NHM.—images, illustrations (A. Smith, V. von Wirth, pers. comm.).
Other material: Lyrognathus crotalus Pocock 1895, female topotype, 09.10.7.36, North Khasi Hills (25°34' N, 91°37' E), Meghalaya State, northeastern India, NHM.–illustrations (A. Smith, pers. comm.); male, 00000641, North Khasi Hills, Meghalaya State, northeastern India, ZMUC.—illustrations (A. Smith, pers.
WEST & NUNN8 · Zootaxa 2362 © 2010 Magnolia Press

comm.); 1 male, 1853.54-11, Shillong, Meghalaya State, northeastern India, ZSL.—illustrations (A. Smith, pers. comm.)(new det.).
Diagnosis (females): Differs from other Lyrognathus species in the intercheliceral spines, not pegs (Fig. 10, except L. saltator) and unilobular spermathecae (Fig. 6). Differs from L. saltator in having only 5 apical spines on metatarsi IV and highly incrassate tibia IV (Fig. 8). Differs from L. achilles sp. nov., L. fuscus sp. nov., L. lessunda sp. nov. and L. robustus in having unilobular spermathecae, presence of intercheliceral spiniform setae (not pegs) and absence of retrolateral basomedial cheliceral spines. Leg formula length IV, I, II = III (41, 32, 27, 27) and width IV, III, I, II. Leg RF~78.05.
FIGURES 11–12. 11. �Lyrognathus crotalus Pocock, biotope, William Nagar, Meghalaya, India. 12. Detail of burrow entrance in open soil. Photos: J-M. Verdez.
Description: Female holotype with body length: 26.4. Color (in life, Fig. 1): entirely dark grey to black.Carapace (Fig. 1): length 13.5, width 10.0. Fovea wide, procurved, deep, equal in width to OT. Eyes: AME, ALE, PLE, PME. Anterior row transverse, posterior row recurved. OT highest posteriorly
behind AME, sloping gradually anteriorly. Chelicerae (left, Figs 9, 10): ectal lyrate region a series of strikers (>65) in 7 horizontal rows (Fig. 9).
Strongest and longest strikers on lowest rows. Each striker needle-form. Teeth, 10, >50 basomesal teeth. Intercheliceral spines (>8) in tight cluster on basodorsal surface (Fig. 10). Retrolateral surface lacks basomedial spine cluster.
Maxillae (Fig. 7): prolaterally plano-convex, anterior lobe well pronounced, many cuspules (>120) on inner basoventral surface. Lyra (Fig. 7): bacilliform rods (>155) form dense, ovoid patch on prolateral surface, lowest row with >26 bacillae, longest rods medially in lowest row. Rods distally paddle-shaped with medium to long shafts, largest rods lacking distal blades. At widest point, lyrate patch 11 rows deep with smallest rods dorsally. Posterior end of patch truncate but rounded, anterior end rounded. Immediately above maxillary suture >11 elongated spines on anterior margin, rows disordered. Labium: many small cuspules (>450) along anterior 1/4 surface.
Sternum: saddle-form. Posteriorly between left and right coxae IV, border highly acuminate, lateral points slightly acuminate. Sigilla: 3 pairs (not including labio-sternal sigilla), posterior, large in size. Ovular, 1.5 of their length apart, 1.0–1.2 of their length from margin. Median pair, 1/2 size of posterior, similar in form and 0.6–0.7 of their length from margin. Anterior pair very small, somewhat obscured and border margin. Labio-sternal sigilla largest of all, 1.2 times size of PSS.
Legs (Figs. 3, 4, 8): formula (length); IV, I, II, III: (width); IV, III, I, II. Leg RF~78.05. Leg lengths: palp: 19, I: 32, II: 27, III: 27, IV: 41. Tib. IV with strong retrolateral villous setal fringe, proximally to distally uniform–entire (Fig. 8).
Zootaxa 2362 © 2010 Magnolia Press · 9REVISION OF TARANTULA GENUS LYROGNATHUS

Scopulae: met. I–IV undivided, tar. I–II. undivided, tar. III partially divided proximally (by one row of sparse setae >5, nonexistent in distal 1/2), tar. IV divided by 2–3 rows of setae. Met. I–IV, covers full length ventrally.
Coxae: some small black thorns prolatero-dorsally, no thorns retrolaterally on I–IV. Coxae easily seen dorsally. Cox. I longest, ca. 1.3 times length of II. IV widest, as long as III, basally rectangular with rounded corners. Coxae with small ventral thorns prolaterally on I–IV. I–III ventrally with numerous long thick blunt setae proximally, pallid. No short black setae. IV with mixture of long thick blunt setae entirely, pallid intermixed with shorter thin pallid setae. Ventral surface of coxae I–IV gently sloping anteriorly. Retrolateral setation: I–III with median narrow light brush, IV entirely covered in short thin pallid setae. I–IV retrolaterally lack ventral ledge.
Trichobothria: on all tarsi basal filiform field slightly wider than clavate field, merges evenly. Clavates on tar. I in distal 1/2 (very dark), long filiforms only in basal 2/3, shorter filiforms intermixed with clavates distally. Clavate extent on tar. II–IV cf. I, in distal 1/2. Shorter filiforms for length. Short epitrichobothrial field on I shorter than clavates, increasing in length proximally.
Spines: met. I with 1 DV, met. II with 1 DV, 1 DPV, 1 DRV, met. III with 2 DV, 1DPV, 1 DRV, 1 DD, 1 DPL and met. IV with 2 DV, 1DPV, 1 DRV, 1 DPD.
Claws: unarmed claws on all legs and palps. Reduced third claw absent on leg IV. Abdomen (Fig. 3): ovular, elongated, yellow brown in alcohol, black in life. Genitalia (Fig. 6): spermathecae paired but not fused, unilobular apically, each lobe with mild apical
rounded expansions, ovular in form, heavily sclerotized gradually transition to weakly sclerotized shafts proximally. In L. crotalus topotype (09.10.7.36, Fig. 6), lobes bend slightly laterally and medially, in L. pugnax holotype (1895.1.28.1), lobes lack bend.
Distribution and natural history (Figs 11, 12, 114): Known only from the type locality of Assam State and from Shillong, Meghalaya State with confirmed reports also throughout Meghalaya State to the south and west (S. Rafn, M. Siliwal and J-M. Verdez, pers. comm.). Assam State is located directly south of the Eastern Himalayas and shares borders with the countries of Bhutan and Bangladesh. To the south of Assam State is Meghalaya State. About one-third of the small state of Meghalaya is heavily forested, the Sanskrit name itself translates in English to “The Abode of Clouds”. The spiders were widely distributed but common in partially shaded laterite roadside or trail embankments in open areas of bamboo and evergreen deciduous forest of the East Garo Hills, Meghalaya State, India at elevations of 300–550 meters.
L. crotalus construct simple flare-mouthed tubular burrows, utilizing surrounding organic debris or vegetation around the mouth, on almost vertical embankments (Figs 11, 12). Adult female burrows were J-shaped and entirely lined with silk, terminating in an enlarged chamber about 30–50 cm deep. An ultimate male was found in a burrow in October, females were observed with egg sacs in February and young instars were observed in mid-April to early May (S. Rafn and J-M. Verdez, pers. comm.).
Remarks: When Pocock (1895) described L. crotalus, he had putatively examined two specimens he identified belonging to that species, sent to the NHM in 1854 (Smith 1988). He discussed one (1854.16) as the ‘type’ in 1895, but neglected to mention the other (09.10.7.36). Although Pocock had putatively examined both specimens prior to his publication, the later specimen can only be considered a "topotype", i.e., of no formal nomenclatural status.
Siliwal (pers. comm.) noted significant variation within Lyrognathus specimens collected in and around the L. crotalus type locality: Guwahati City (26°11' N, 91°45' E) and Nameri National Park (26°57' N, 92°45' E), Sonitpur District, Assam State, Northeast India. Clypeus widths seem to vary in conspecifics, as does the partial division of tarsi III scopula previously thought diagnostic of L. crotalus. In addition, the types only vary in size (L. pugnax is slightly larger), the tarsi III scopula division and slight spermathecal variation. The spermathecal variation is minimal (in the L. crotalus topotype female 09.10.7.36, the spermathecae bend slightly laterally and medially, in L. pugnax holotype female, they do not), particularly considering both specimens were dry stored at the NHM for over 30 years before being placed into wet preservative and may therefore be distorted.
WEST & NUNN10 · Zootaxa 2362 © 2010 Magnolia Press

S. Rafn (pers. comm.) reported only spiders conspecific with L. crotalus in Shillong district (type locality for L. pugnax) which show a blend between partially divided and undivided tarsi III scopula, but no distinct division between specimens collected. We consider that, as L. pugnax is based only one (type) specimen with a reduced clypeus and minimal variation of tarsi III scopula division in comparison to sympatric L. crotalus types, that L. pugnax is clearly a large L. crotalus with undivided tarsi III scopula and reduced clypeus.Prior to 1903, tarsal scopula division was considered of great taxonomic use (incorrectly considering juveniles: see Guadanucci 2005), possibly explaining Pocock actions at that time. We, therefore, consider L. crotalus Pocock 1895 a senior synonym of L. pugnax Pocock 1900.
Lyrognathus achilles sp. nov.(Figs 13–44, 114)
Types: female holotype, S88049; male paratype S88050; 2 female paratypes S88047 and S88048, Gunung Nibung (2°07' S, 112°00' E), Kotawaringin Barat, Borneo, Indonesia, col. H. Yanto, 2 Apr. 2009, QM.—examined.
Etymology: After the mythical Greek warrior of Homer’s poem, The Iliad, due to the unique tarsal scopula division: distinct proximal bald patch on ventral tarsi (Fig. 19) this species displays.
Diagnosis: Differs from both L. crotalus and L. saltator in having intercheliceral pegs (not spines), and retrolateral basomedial cheliceral spines. Differs from L. robustus in the undivided scopula for the full length of metatarsi, third claw absent on tarsi IV and swollen distal lateral lobes of the spermathecae. Differs from L. lessunda sp. nov. in the absence of “mushroom cap” medial lobes of the spermathecae. Differs from L. fuscussp. nov. in the more pronounced anterior maxillary lobe, distal cream/white leg segment bands, larger size, color and uniform retrolateral tibiae IV with a penicillate brush (grading distally in L. fuscus sp. nov.).
Description: Female holotype (Fig. 13) with body length: 50.57. Color (in life, Fig. 13): anteriorly orange/brown, grading medially to dark brown/black posteriorly and
entirely, with distinct cream/white bands on all leg segments distally. Dorsal coxae, trochantera palest of all, light grey. Ventrally dark brown/black on sternum, coxae, with legs same as seen dorsally.
Carapace (Fig. 15): length 21.45, width 13.8 (width across anterior edge 9.45). Fovea 2.71 wide, procurved, deep, smaller in width than OT. Distance from anterior carapace to fovea 13.7. Carapace with 4 discernible hair types (C1 to C4): type C1, long, needle-form pallids bordering carapace entirely (darker basally): type C2, short blunt-tipped, basally dark grading to distally pallid and cover entire carapace evenly, but reasonably sparse: type C3, short, thin wavy pallids, form basis of carapace mat: type C4 emerge from what resemble bothrial collars, long, distally spiniform, pallid and lie in uniform arrangement of (>5) along anterior edge of each radial groove, (>4) along anterior edge of each thoracic groove, (>5) along anterior edge of each sulcal groove, (>10) along anterior edge of each caput groove and line of >5 along medial caput ridge (between OT and fovea). >4 more located behind each side of OT, nearest median caput row.
Eyes (Fig. 16): ocular tubercle; length 2.25, width 3.0. OT highest posteriorly behind AME, sloping gradually anteriorly. Anterior row transverse, posterior row recurved. Eyes in lengths/widths: AME: 0.71/0.70, ALE: 0.69/0.27, PLE: 0.49/0.25, PME: 0.46/0.27. Inter-ocular distances: AME–AME: 0.29, AME–ALE: 0.21, AME–PME: 0.18, ALE–ALE: 1.85, ALE–PME: 0.38, PME–PME: 1.51, PME–PLE: 0.19, PLE–PLE: 2.35, ALE–PLE: 0.26. OT hairs: >15 (C2) directly posterior to AME’s, 4–5 between PME and center of OT (directly behind each AME). 20 (C2) along median anterior OT surface. OT entirely covered in small, weak, pallid setae (C3), directionally pointing to center of OT, posteriorly and anteriorly. Lateral small pallids very sparse with no uniform direction/random pointing.
Chelicerae (left, Figs 24–28): length 9.8, width at base of each lobe 4.25 (at bases widest point, dorsally), height: 7.1. Ectal lyrate region a series of strikers (>95) in 8 horizontal rows (Fig. 25). Strongest and longest strikers on lowest rows. Each striker very stout, blade or scimitar-shaped, with long filiform ends. Cheliceral teeth, 12, >65 basomesal teeth. Intercheliceral pegs (>9 larger, >28 smaller) in tight cluster on basodorsal surface, very stout, 1/3 as wide as long (Fig. 28). On retrolateral cheliceral surface, basomedially, with small
Zootaxa 2362 © 2010 Magnolia Press · 11REVISION OF TARANTULA GENUS LYROGNATHUS

cluster of short but strong spiniform setae (>5) and 1 spine pointing distad (Fig. 26). Surrounding these spines, notable patch of fine setae, dark, of medium length, blends into typical thicker pallid retrolateral cheliceral setae. Setae filiform, seem to emerge from hemispherical bases.
FIGURES 13–14. 13. Lyrognathus achilles sp. nov. 13. Holotype female in life. 14. Paratype male in life. Photos: R. C. West.
WEST & NUNN12 · Zootaxa 2362 © 2010 Magnolia Press
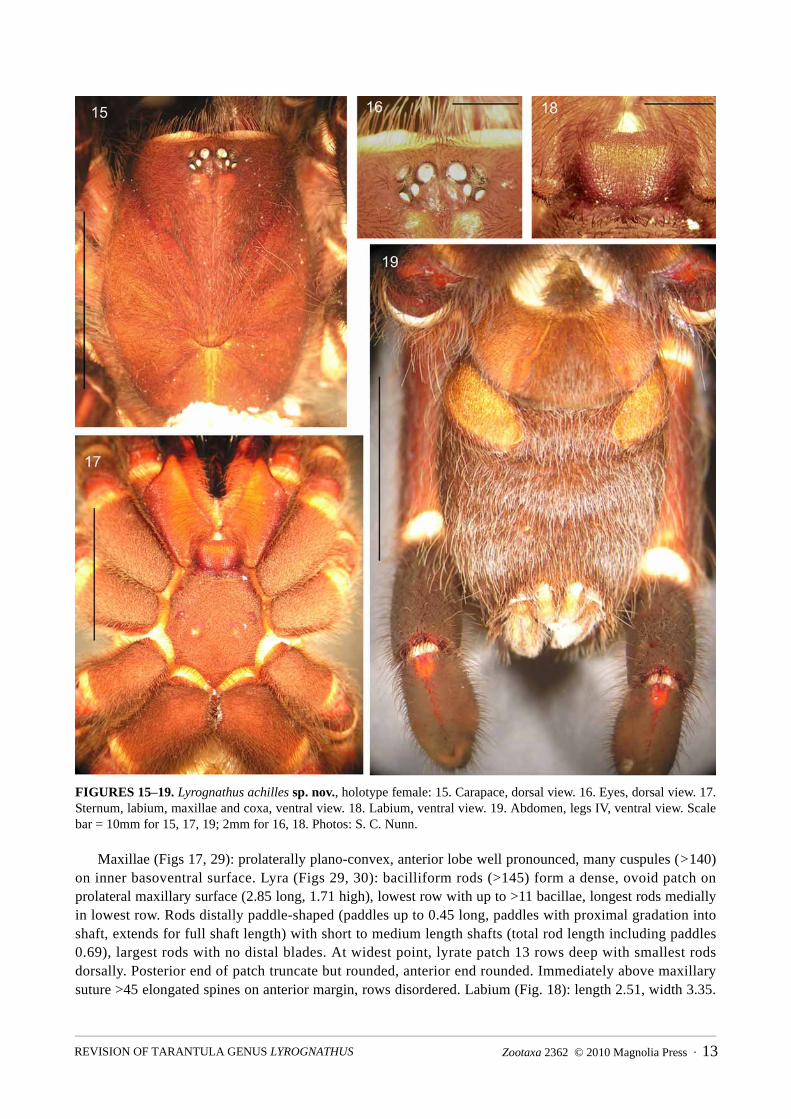
FIGURES 15–19. Lyrognathus achilles sp. nov., holotype female: 15. Carapace, dorsal view. 16. Eyes, dorsal view. 17. Sternum, labium, maxillae and coxa, ventral view. 18. Labium, ventral view. 19. Abdomen, legs IV, ventral view. Scale bar = 10mm for 15, 17, 19; 2mm for 16, 18. Photos: S. C. Nunn.
Maxillae (Figs 17, 29): prolaterally plano-convex, anterior lobe well pronounced, many cuspules (>140) on inner basoventral surface. Lyra (Figs 29, 30): bacilliform rods (>145) form a dense, ovoid patch on prolateral maxillary surface (2.85 long, 1.71 high), lowest row with up to >11 bacillae, longest rods medially in lowest row. Rods distally paddle-shaped (paddles up to 0.45 long, paddles with proximal gradation into shaft, extends for full shaft length) with short to medium length shafts (total rod length including paddles 0.69), largest rods with no distal blades. At widest point, lyrate patch 13 rows deep with smallest rods dorsally. Posterior end of patch truncate but rounded, anterior end rounded. Immediately above maxillary suture >45 elongated spines on anterior margin, rows disordered. Labium (Fig. 18): length 2.51, width 3.35.
Zootaxa 2362 © 2010 Magnolia Press · 13REVISION OF TARANTULA GENUS LYROGNATHUS

Many small cuspules (>560) along anterior 1/4 surface. Labium with 2 discernible hair types (L1, L2); type L1: long, reasonably dark spiniform, curving distally toward anterior, most concentrated along lateral borders. Setae clearly longest along anterior edge (>35); type L2: 1/2 length of L1, interspersed with L1 but darker, more needle-form.
FIGURES 20–23. Lyrognathus achilles sp. nov., holotype female: 20. Leg IV, dorsal view. 21. Leg IV, prolateral view. 22. Tar. IV, ventral view. 23. Spermathecae, dorsal view. Scale bar = 10mm for 20, 21; 5mm for 22; 2mm for 23. Photos: S. C. Nunn.
Sternum (Fig. 17): length 9.58, width 7.35, saddle-form; with 5 discernible hair types (S1 to S5): type S1, longest, thick, darker basally, becoming pallid distally, entirely over surface, sparse: type S2, elongate spiniforms, dark basally, distally pallid, on margins, most concentrated posteriorly: type S3, short spiniforms,
WEST & NUNN14 · Zootaxa 2362 © 2010 Magnolia Press

entirely, sparse: type S4, short, thin pallids, form basis of mat: type S5 dark basally, but apically pallid, short and blunt tipped, entirely cover in sparse disordered arrangement. Posteriorly between left and right coxae IV, border highly acuminate, lateral points slightly acuminate. Sigilla (Fig. 17): 3 pairs (not including labio-sternal sigilla), posterior, large, right: length 1.38, width 0.75, left with length 1.35, width 0.65. Ovular, 1.76 apart, 1.65 to 1.7 from margin. Median pair, 1/2 size of posterior, similar shape and 0.7–1.0 of their length from sternal margin. Anterior pair large, as big as medials, somewhat obscured and border margin. Labio-sternal sigilla largest, 1.6 times size of PSS.
FIGURES 24–28. Lyrognathus achilles sp. nov., paratype female QM S88047: 24. Left chelicerae, retrolateral view. 25. Cheliceral strikers, ectal view. 26. Basomedial cheliceral spine, prolateral view. 27. Left chelicerae, prolateral view. 28. Intercheliceral pegs, prolateral view. Scale bar = 5mm for 24, 25, 27; 1mm for 26, 28. Photos: S. C. Nunn.
Legs: formula (length); IV, I, II, III: (width); IV, III, I, II. Leg RF~84.51. Leg lengths (fem., pat., tib., met., tar., total): palp: 9.46, 5.98, 6.74, 0.00, 5.75, 27.93. I: 13.21, 8.87, 9.98, 9.15, 5.34, 46.55. II: 11.64, 7.96, 8.48, 7.59, 5.22, 40.89. III: 9.64, 6.79, 6.78, 8.57, 5.01, 36.79. IV: 15.55, 8.44, 11.33, 14.16, 5.6, 55.08. Leg widths (fem., pat., tib., met., tar., total.): palp: 2.15, 2.71, 2.42, 0.00, 2.54, 9.82. I: 3.38, 3.37, 2.99, 2.34, 2.14, 14.22. II: 3.37, 3.26, 2.79, 2.35, 2.23, 14.00. III: 4.07, 3.57, 3.05, 2.33, 2.18, 15.20. IV: 4.08, 3.61, 4.05, 2.83, 2.49, 17.06.Tib. IV with strong retrolateral villous setal fringe, proximally to distally uniform–entire (Fig. 20).
Scopula: met. and tar. I–III undivided, met. IV undivided, tar. IV (Fig. 22) only partially divided proximally (by one row of sparse setae >5, nonexistent in distal 1/2). Met. I–IV, for full length.
Zootaxa 2362 © 2010 Magnolia Press · 15REVISION OF TARANTULA GENUS LYROGNATHUS

Coxae: some small black thorns prolatero-dorsally, no thorns retrolaterally on I–IV. Coxae easily seen dorsally. Cox. I longest, ca. 1.2 times length of II. IV widest, as long as III, basally rectangular with rounded corners. Coxae with small ventral thorns prolaterally on I–IV. I–III ventrally with many long thick blunt setae proximally, pallid. No short black setae. IV with mixture of long thick blunt setae entirely, pallid intermixed with shorter thin pallid setae. Ventral surface of coxae I–IV gently sloping anteriorly. Retrolateral setation: I–III with median narrow light brush, IV entirely covered in short thin pallid setae. I–IV retrolaterally lack ventral ledge. Ventral measurements for coxae: palp–length 9.78, width 4.94; I–9.18, 4.16; II–7.53, 3.63; III–6.24, 4.02; IV–7.53, 4.96. Trochantera: palp–length 2.85, width 2.9; I–3.31, 4.06; II–2.9, 3.88; III–3.26, 3.6; IV–4.14, 4.11.
Trichobothria: on all tarsi basal filiform field slightly wider than clavate field, merges evenly. Clavates on tar. I in distal 1/2 (very dark), long filiforms only in basal 2/3, shorter filiforms intermixed with clavates distally. Clavate extent on tar. II–IV cf. I, in distal 1/2. Shorter filiforms for length. Short epitrichobothrial field on I shorter than clavates, increasing in length proximally. Tarsal organ evident on legs II–IV under stereo microscope (up to x64), coniform without noticeable ridges. On tar. I, entire field 4.56 long and 1.32 wide distally, to 1.85 wide proximally. On tar. IV, field 4.78 long and 0.89 wide distally, increasing to 1.95 proximally. Metatarsi: trichobothrial field not evident laterally or proximally in any form. Tibia: on tib. I prolaterally and proximally, short, possibly clavate trichobothria noted (>8 per 0.25mm squared) in L-shaped patch 0.25 long, 1.25 wide, clearly pallid: >6 filiforms in semicircular pattern proximally to rest of patch. On tib. IV prolaterally and proximally, short dark clavates (>7 per 0.25mm squared) in patch 1.66 long, 0.22 wide distinct: >7 filiforms in semicircular pattern proximally to rest of patch.
Spines: met. I with 1 DV, met. II with 1 DV, 1 DPV, 1 DRV, met. III with 2 DV, 1DPV, 1 DRV, 1 DD, 1 DPL and met. IV with 2 DV, 1DPV, 1 DRV, 1 DD, 1 DPD.
Claws: unarmed paired claws on all legs. Reduced third claw absent on leg IV. Abdomen (Fig. 19): ovular, elongated, yellow brown (in alcohol); with 4 discernible hair types (A1 to
A4): Dorsally with 3 hair types: type A1 long, dark blunt tipped, uniform: type A2 mid-length, also dark, but distally spiniform, unlike type A1: type A3 form dense mat, mid-length, uniformly dark, needle-form (some stand erect, some posteriorly, but share similar morphology). Longest hairs (type A1) more concentrated posteriorly toward spinnerets, point distad, but toward proximal region, hairs stand erect, medially curved (toward posterior). Ventral hair types almost same as dorsal, although type A2 and A3 more dense entirely. Type A4 hair in very dense patch, trapezoid, posterior of pedicel between median regions of cox. IV. Hairs of medium length, wavy and pallid. Between anterior pair of booklungs, setal mat uniform with rest of venter (Fig. 19).
Genitalia (Fig. 23): epigastric fold 6.51 wide. Spermathecae paired but not fused, separated by 0.8 (width including spermathecae 2.4), bilobular apically, each lobe with strong apical rounded appendages, lateral/outer lobes distally swollen, ovular: lateral lobe distal widths 0.45 (left)–0.41 (right): medial lobe distal widths 0.26 (left)–0.27 (right), heavily sclerotized gradually transition to weakly sclerotized shafts proximally. Lobes apically swollen, lateral lobes (including shaft length) at 4/5 length to medials. Epigastic fold extends to ca. 2/3 length of spermathecal lobes.
Spinnerets: PMS: length 2.1, width (medially) 0.95. PLS: section lengths; basal 3.45, medial 1.45, apical 2.55. PLS section widths (medially); basal 1.78, medial 1.32, apical 0.85. Spinnerets with 2 discernible hair types (SP1, SP2): SP1 very short and distally blunt pallid, covers both pairs ventrally, second form (SP2), very similar but up to three times longer, dorsally on all segments.
Male paratype (Fig. 14, as in female except) with body length: 34.09. Color (in life, Fig. 14): With brass sheen to dorsal carapace, coxae and trochantera hairs, also less distinct
in color anteriorly to posteriorly, clearly paler distal leg segments (pat., tib., met., tar.). Carapace (Fig. 31): length 13.85, and width 13.1 (width across anterior edge 7.35). Fovea 1.68 wide,
procurved, deep, narrower than OT. Distance from anterior carapace to fovea, 9.89. Carapace hair types as in female except C1, C2 and C4 are brass color, iridescent in alcohol. Eyes (Fig. 32): ocular tubercle; length 1.75, width 2.43.
WEST & NUNN16 · Zootaxa 2362 © 2010 Magnolia Press
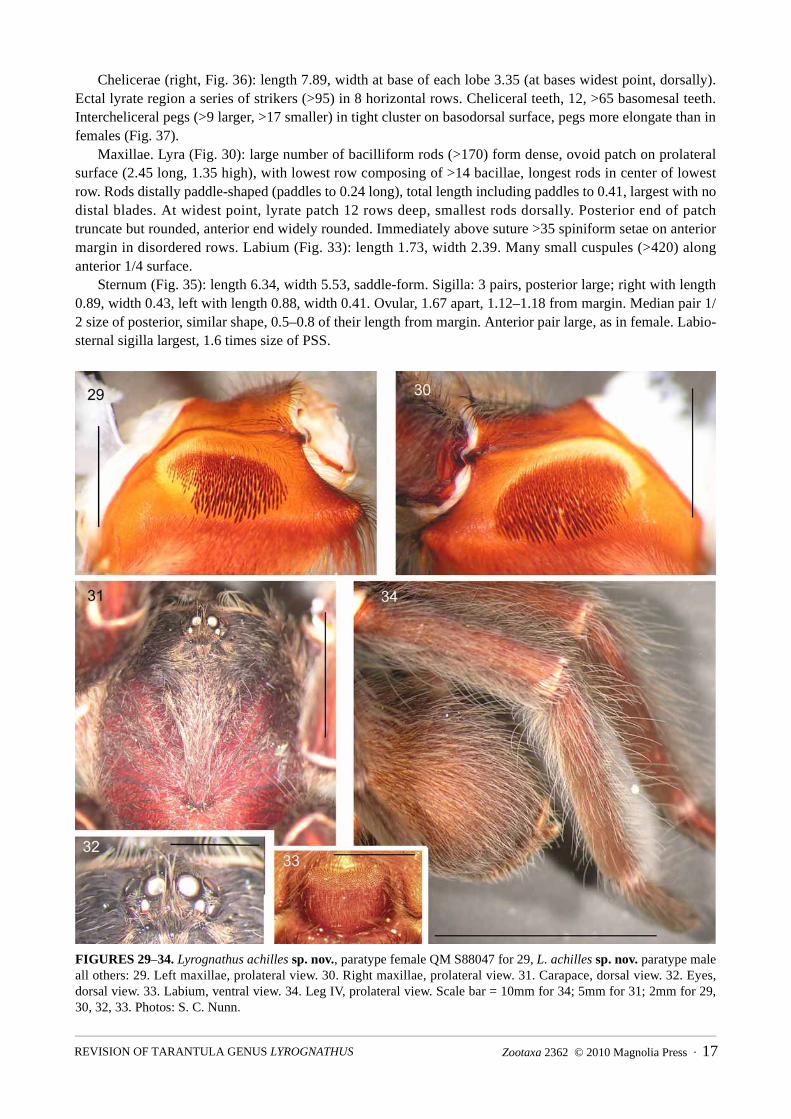
Chelicerae (right, Fig. 36): length 7.89, width at base of each lobe 3.35 (at bases widest point, dorsally). Ectal lyrate region a series of strikers (>95) in 8 horizontal rows. Cheliceral teeth, 12, >65 basomesal teeth. Intercheliceral pegs (>9 larger, >17 smaller) in tight cluster on basodorsal surface, pegs more elongate than in females (Fig. 37).
Maxillae. Lyra (Fig. 30): large number of bacilliform rods (>170) form dense, ovoid patch on prolateral surface (2.45 long, 1.35 high), with lowest row composing of >14 bacillae, longest rods in center of lowest row. Rods distally paddle-shaped (paddles to 0.24 long), total length including paddles to 0.41, largest with no distal blades. At widest point, lyrate patch 12 rows deep, smallest rods dorsally. Posterior end of patch truncate but rounded, anterior end widely rounded. Immediately above suture >35 spiniform setae on anterior margin in disordered rows. Labium (Fig. 33): length 1.73, width 2.39. Many small cuspules (>420) along anterior 1/4 surface.
Sternum (Fig. 35): length 6.34, width 5.53, saddle-form. Sigilla: 3 pairs, posterior large; right with length 0.89, width 0.43, left with length 0.88, width 0.41. Ovular, 1.67 apart, 1.12–1.18 from margin. Median pair 1/2 size of posterior, similar shape, 0.5–0.8 of their length from margin. Anterior pair large, as in female. Labio-sternal sigilla largest, 1.6 times size of PSS.
FIGURES 29–34. Lyrognathus achilles sp. nov., paratype female QM S88047 for 29, L. achilles sp. nov. paratype male all others: 29. Left maxillae, prolateral view. 30. Right maxillae, prolateral view. 31. Carapace, dorsal view. 32. Eyes, dorsal view. 33. Labium, ventral view. 34. Leg IV, prolateral view. Scale bar = 10mm for 34; 5mm for 31; 2mm for 29, 30, 32, 33. Photos: S. C. Nunn.
Zootaxa 2362 © 2010 Magnolia Press · 17REVISION OF TARANTULA GENUS LYROGNATHUS

FIGURES 35–40. Lyrognathus achilles sp. nov., paratype male: 35. Sternum, labium, maxillae and coxa, ventral view. 36. Right maxillae, retrotrolateral view showing strikers. 37. Right chelicerae, intercheliceral pegs, prolateral view. 38. Right distal palp showing bulb, prolateral view. 39. Right distal palp showing bulb, retrolateral view. 40. Right bulb and embolus, ventro-anterior view showing keeled embolus. Scale bar = 10mm for 35; 2mm for 38, 39; 1mm for 36, 37, 40. Photos: S. C. Nunn.
Legs: formula (length); IV, I, II, III: (width); III, IV, I, II. Leg RF~93.14. Leg lengths (fem., pat., tib., met., tar., total): palp: 8.83, 5.0, 6.77, 0.00, 2.13, 22.73. I: 12.25, 6.83, 10.93, 9.78, 5.26, 45.05. II: 10.66, 6.38, 8.54, 8.17, 4.88, 38.63. III: 8.51, 5.26, 6.72, 7.8, 4.27, 32.56. IV: 13.28, 6.84, 10.38, 12.68, 5.19, 48.37. Leg widths (fem., pat., tib., met., tar., total.): palp: 2.22, 1.92, 1.90, 0.00, 1.97, 8.01. I: 2.9, 2.53, 2.13, 1.52, 1.6, 10.68. II: 2.8, 2.56, 1.91, 1.35, 1.43, 10.05. III: 3.73, 2.7, 2.36, 1.55, 1.56, 11.9. IV: 2.75, 2.58, 2.39, 1.72, 1.77, 11.21. Tib. IV with retrolateral villous setal fringe, proximally to distally uniform, not as elongate or dense as in females. Ventral measurements for coxae: palp–length 5.41, width 2.57; I–5.89, 2.67; II–5.45, 2.67; III–3.89, 2.78; IV–5.1, 3.78. Trochantera: palp–length 2.57, width 2.1; I–3.34, 3.1; II–2.78, 2.54; III–2.43, 2.45; IV–3.35, 2.93.
Genitalia: Palp (Figs 38–40): tibia mildly incrassate, tapering distally. Cymbium bipartite, tegulum large. Lanceolate embolus slightly tapers distally, very little flaring, proximally emerges anterior to tegulum. Relative length of palpal tibia about three times length of embolus. Longitudinal keel present, distinct (embolus not terete), along entire length of embolus, twisted through 90° prolaterally distally (Fig. 40).
Spinnerets: PMS: length 1.23, width (medially) 0.51. PLS: section lengths; basal 1.95, medial 1.25, apical 1.85. PLS section widths (medially); basal 1.12, medial 0.75, apical 0.55.
WEST & NUNN18 · Zootaxa 2362 © 2010 Magnolia Press

Distribution and natural history (Figs 41–44, 114): Known only from the type locality, Gunung Nibung, Kotawaringin Barat, Borneo, Indonesia. Little is known; however, L. achilles is a fossorial spider that lives in higher shaded tropical forests away from the riparian flood plains. Burrows were found only on sloped embankments amongst leaf litter and low vegetation and were formed in clay-like soil. A mature male was found in early April (H. Yanto, pers. comm.).
FIGURES 41–44. 41. Lyrognathus achilles sp. nov. Upper slopes of Gunung Nibung, Borneo, Indonesia. 42. Biotope, Gunung Nibung. 43. Biotope with arrow pointing to burrow entrance. 44. Detail of burrow entrance among foliage. Photos: 41–44: H. Yanto.
Lyrognathus fuscus sp.nov.(Figs 45–62, 114)
Types: female holotype, 3 female paratypes S88054, S88051, S88052 and S88053, respectively, Simpang Kubur (2°29' S, 111°25' E), Katawaringin, Kotawaringin Barat, Borneo, Indonesia, col. H. Yanto, 2 Apr. 2009, QM.—examined.
Etymology: From Latin meaning dark brown/grey. The genus name is masculine, thus fuscus is the masculine form of the adjective.
Common name: Hariyanto’s tarantula, after the collector of this species.Diagnosis: Differs from both L. crotalus and L. saltator in having intercheliceral pegs (not spines), and
retrolateral basomedial cheliceral spines. Differs from L. robustus in having undivided scopula for full length of metatarsi IV, swollen distal lateral lobes of spermathecae but no third claw on tarsi IV. Differs from L. lessunda sp. nov. in the absence of “mushroom cap” medial lobes of the spermathecae. Differs from L. achilles sp. nov. in less pronounced maxillary lobe, lack of distal cream/white leg segment bands, smaller
Zootaxa 2362 © 2010 Magnolia Press · 19REVISION OF TARANTULA GENUS LYROGNATHUS

size, color and proximal-distal gradation of retrolateral tibiae IV penicillate brush (uniform for length in L. achilles sp. nov.).
Description: Female holotype (Fig. 45) with body length: 45.87.
FIGURES 45–46. Lyrognathus fuscus sp. nov. 45. Holotype female in life. 46. Male in life. Photos: 45, R. C. West; 46, H. Yanto.
WEST & NUNN20 · Zootaxa 2362 © 2010 Magnolia Press

FIGURES 47–52. Lyrognathus fuscus sp. nov. holotype female: 47. Carapace, dorsal view. 48. Eyes, dorsal view. 49. Sternum, labium, maxillae and coxa, ventral view. 50. Labium, ventral view. 51. Spermathecae, dorsal view. 52. Abdomen, ventral view. Scale bar = 10mm for 47, 49, 52; 2mm for 48, 50; 1mm for 51. Photos: S. C. Nunn.
Color (in life, Fig. 45): Entirely dark brown/grey, no distal leg segment bands.Carapace (Fig. 47): length 16.96, width 12.77 (width across anterior edge 8.85). Fovea 2.20 wide,
procurved, deep, smaller in width than OT. Distance from anterior carapace to fovea, 12.51; with 4 discernible hair types (C1 to C4): type C1, long needle-form pallids border carapace entirely; type C2, short black, blunt tipped, entirely over carapace; type C3, short thin, wavy pallids, form basis of mat; type C4, emerge from bothrial-like collars, long, distally spiniform, pallid, in uniform arrangement: (>5) along anterior edge of each radial groove, (>4) along anterior edge of each thoracic groove, (>5) along anterior edge of sulcal groove,
Zootaxa 2362 © 2010 Magnolia Press · 21REVISION OF TARANTULA GENUS LYROGNATHUS

(>6) along anterior edge of each caput groove, ( >4) along medial caput ridge (between OT/fovea), (>3) located behind each side of OT, nearest median caput row.
Eyes (Fig. 48): ocular tubercle; length 1.95, width 2.91. OT highest posteriorly behind AME, sloping gradually anteriorly. Anterior row slightly procurved. Posterior row recurved. Eyes in lengths/widths: AME: 0.67/0.51, ALE: 0.71/0.27, PLE: 0.41/0.25, PME: 0.40/0.28. Inter-ocular distances: AME–AME: 0.28, AME–ALE: 0.24, AME–PME: 0.15, ALE–ALE: 1.79, ALE–PME: 0.41, PME–PME: 1.43, PME–PLE: 0.19, PLE–PLE: 2.0, ALE–PLE: 0.38. OT hairs: >15 (C2) directly posterior to AME’s, two between PME and center of OT (behind AME). 16 (C2) along median anterior OT surface. OT entirely covered in small, weak, pallid setae (C3), directionally point to center of OT, posteriorly–anteriorly. Lateral small pallids very sparse with no uniform direction/random arrangement.
Chelicerae (right, Figs 58–62): length 8.90, width at base of lobe 4.15 (bases widest point, dorsally). Ectal lyrate region a series of strikers (>75), 8 horizontal rows (Fig. 59). Strongest and longest strikers on lowest rows. Each striker very stout, blade or scimitar-shaped with long filiform ends. Teeth, 13, >40 basomesal teeth. Intercheliceral pegs (>11 larger, >18 smaller) in tight cluster on basodorsal surface, extremely stout, 1/2 as wide as long (Fig. 62). Retrolaterally and basomedially, small cluster of short strong spiniform setae (>7) point distad (Fig. 60). Surrounding spines a patch of fine setae, dark, of medium length, blends into typical thicker pallid cheliceral setae. Setae filiform and seem to emerge from hemispherical bases.
FIGURES 53–57. Lyrognathus fuscus sp. nov. holotype female (except paratype QM S88051; 56–57): 53. Left leg IV, dorsal view. 54. Left leg IV, retrolateral view. 55. Left met, tar. IV, prolateral view. 56. Left maxillae, prolateral view. 57. Maxillary lyra, prolateral view. Scale bar = 10mm for 53–55; 2mm for 56; 1mm for 57. Photos: S. C. Nunn.
WEST & NUNN22 · Zootaxa 2362 © 2010 Magnolia Press

FIGURES 58–62. Lyrognathus fuscus sp. nov. paratype female QM S88051: 58. Left chelicerae, retrolateral view. 59. Left cheliceral strikers, ectal view. 60. Basomedial cheliceral spines, retrolateral view. 61. Left chelicerae, prolateral view. 62. Left intercheliceral pegs, prolateral view. Scale bar = 5mm for 58, 61; 2mm for 59; 1mm for 60, 62. Photos: S. C. Nunn.
Maxillae (Figs 49, 56): prolaterally plano-convex, anterior lobe well pronounced, many cuspules (>105) on inner basoventral surface. Lyra (Figs 56, 57): large number of bacilliform rods (>180) form dense, ovoid patch on prolateral maxillary surface (3.23 long, 1.72 high), lowest row with up to >18 bacillae, longest rods in center of lowest rows. Rods distally paddle-shaped (paddles up to 0.19 long) with short to medium length shafts (total length 0.69), largest rods with no distal blades. At widest point, lyrate patch 10 rows deep, smallest rods dorsally. Posterior end of patch truncate, anterior end not pointed distad/widely rounded. Immediately above maxillary suture >45 elongated spines on anterior margin in disordered rows. Labium (Fig. 50): length 2.35, width 3.0. Many small cuspules (>540) along anterior 1/4 surface. Labium with 2 discernible hair types (L1, L2); type L1: long, reasonably dark spiniform, curve distally toward anterior, most concentrated along lateral borders. Setae longest along anterior edge (>35); type L2: 1/2 length of L1, interspersed with L1 but darker, more needle-form.
Sternum (Fig. 49): length 7.65, width 6.68, saddle-form; with 5 discernible hair types (S1 to S5): type S1, longest, thick blunt tipped, darker basally, become pallid distally, entire but sparse; type S2, elongate spiniforms, entirely dark, on borders, most concentrated posteriorly; type S3, similar morphology to S1, although only 1/4–1/2 length in comparison, over entire surface, sparse; type S4, short, thin pallids, form basis of mat; type S5, short, black blunt tipped, entirely cover sternum in sparse disordered arrangement. Posteriorly between left and right coxa IV, border highly acuminate, lateral points slightly acuminate. Sigilla: 3 pairs, posterior, large; right with length 1.31, width 0.65, left length 1.0, width 0.6. Ovular, 1.65 apart, 1.35–1.38 from margin. Median pair, 1/2 size of posterior, similar shape, 1.0–0.6 of their length from margin.
Zootaxa 2362 © 2010 Magnolia Press · 23REVISION OF TARANTULA GENUS LYROGNATHUS

Anterior pair 1/3 size of posterior, somewhat obscured, border sternal margin. Labio-sternal sigilla large, only slightly smaller than PSS.
Legs: formula (length); IV, I, II, III: (width); IV, III, I, II. Leg RF~86.72. Leg lengths (fem., pat., tib., met., tar., total): palp: 9.32, 5.73, 6.13, 0.00, 5.10, 26.28. I: 12.48, 7.92, 9.26, 7.68, 4.99, 42.33. II: 10.78, 6.93, 7.21, 7.31, 4.71, 36.94. III: 9.32, 5.93, 5.5, 7.22, 4.23, 32.2. IV: 13.73, 7.39, 10.43, 11.85, 5.41, 48.81. Leg widths (fem., pat., tib., met., tar., total.): palp: 2.36, 2.61, 2.17, 0.00, 2.26, 9.4. I: 3.08, 3.11, 2.99, 2.13, 2.05, 13.36. II: 3.06, 3.12, 2.57, 1.95, 1.89, 12.59. III: 3.8, 3.32, 2.91, 2.10, 1.96, 14.09. IV: 3.41, 3.26, 3.44, 2.35, 2.01, 14.47. Tib. IV with strong retrolateral penicillate setal fringe, proximally weak, gradually increases distally (Fig. 53).
Scopula: Met., tar. I–III undivided, met. IV undivided, tar. IV partially divided proximally (by one row of sparse setae, nonexistent in distal 1/2). Met. I–IV for full length.
Coxae: some small black thorns prolatero-dorsally, no thorns retrolaterally on I–IV. Coxae easily seen dorsally. Cox. I longest, about 1.3 times longer than II. IV widest, as long as III, basally rectangular with rounded corners. Coxae with small ventral thorns prolaterally on I–IV. I–III ventrally with many long thick blunt setae proximally, pallid. No short black setae. IV with mixture of long thick blunt setae entirely, pallid intermixed with shorter thin pallid setae. Ventral surface of coxae I–IV gently sloping anteriorly. Retrolateral setation: I–III with median narrow light brush, IV entirely covered in short thin pallid setae. I–IV retrolaterally lack ventral ledge. Ventral measurements for coxae: palp–length 6.75, width 4.12; I–7.23, 3.3; II–5.68, 2.83; III–4.44, 3.43; IV–6.3, 3.95. Trochantera: palp–length 2.65, width 1.85; I–3.05, 3.41; II–2.65, 2.78; III–2.45, 2.8; IV–3.1, 3.8.
Trichobothria: All tarsi with basal filiform field slightly wider than clavate field, merges evenly. Clavates on tar. I in distal 1/2 (very dark), long filiforms only in basal 2/3, shorter filiforms intermixed with clavates distally. Clavate extent on II–IV cf. I, in distal 1/2. Shorter filiforms for length. Short epitrichobothrial field on I shorter than clavates, increase in length proximally. Tarsal organ evident on legs I–III under stereo microscope (up to x64), organ on tar. I 1/2 size cf. II and III, low, wide, domed without noticeable ridges. On tar. I, entire field 3.3 long, 0.98 wide distally, to 1.3 wide proximally. On tar. IV, field 3.89 long, 0.85 wide distally, increase to 1.35 proximally. Metatarsi: trichobothrial field not evident laterally or proximally. Tibia: on I prolaterally and proximally, short, possibly clavate trichobothria noted (>8 per 0.25mm squared) in L-shaped patch 0.25 long, 1.25 wide, clearly pallid: >6 filiforms in semicircular pattern proximally to rest of patch; on IV prolaterally and proximally, short dark clavates (>7 per 0.25mm squared) in patch 1.66 long, 0.22 wide distinct: >7 filiforms in semicircular pattern proximally to rest of patch.
Spines: met. I with 1 DV, met. II with 1 DV, met. III with 2 DV, 1DPV, 1 DRV, 1 DD, met. IV with 2 DV, 1DPV, 1 DRV, 1 DD.
Claws: unarmed claws on all legs and palps. Third claw absent on leg IV. Abdomen (Fig. 52): ovular, elongated, yellow brown (in alcohol); with 4 discernible hair types (A1 to
A4); dorsally with 3 hair types; type A1, long dark blunt tipped, uniform in color; type A2, mid-length, dark, but distally spiniform, unlike type 1; type A3, form dense mat of abdomen, mid-length, uniformly dark colored, needle-form (some stand erect, some run posteriorly along abdomen, both share similar morphology). Longest hairs (type A1) more concentrated posteriorly toward spinnerets, pointing distad, but proximally, hairs stand erect and medially curved (toward posterior). Ventral hair types almost same as dorsal, with type A2 and A3 denser entirely. Type 4 hair (A4) in very dense patch, trapezoid in shape, just posterior to pedicel, between median regions of cox. IV. Hairs of medium length, but wavy, pallid. Between anterior pair of booklungs, setal mat jet black, much darker than rest with more concentrated setal counts (Fig. 52).
Genitalia (Fig. 51): epigastric fold 6.2 wide. Spermathecae paired, not fused, separated by 0.8 (width including spermathecae 4.1), bilobular apically, each lobe with strong apical rounded appendages, lateral/outer lobes more defined distally–ovular; lateral lobe distal, widths 0.62 (left)–0.51 (right): medial lobe distal widths 0.32 (left)–0.31 (right), heavily sclerotized gradual transition to weakly sclerotized shafts proximally. Lobes apically swollen, lateral lobes (including shaft length) 4/5 length to medials (1.12 to 1.2, respectively). Epigastric fold extends to about 2/3 length of spermathecal lobes.
Spinnerets: PMS: length 0.71, width (medially) 0.56. PLS: section lengths; basal 2.75, medial 1.46, apical 2.21. PLS section widths (medially); basal 1.35, medial 1.18, apical 0.75. Spinnerets with 2 discernible hair
WEST & NUNN24 · Zootaxa 2362 © 2010 Magnolia Press

types (SP1, SP2): SP1 very short and distally blunt pallid, covers both pairs ventrally; second form (SP2), very similar but up to three times longer, dorsally on all segments.
Male unknown.Distribution and natural history (Fig. 114): Known only from the type locality, Simpang Kubur,
Katawaringin, Kotawaringin Barat, Borneo, Indonesia. Little is known; however, L. fuscus is a fossorial spider that lives in higher shaded tropical forests away from the riparian flood plains. Burrows were only found on sloped embankments amongst leaf litter and low vegetation and were formed in clay-like soil (H. Yanto, pers. comm.).
Lyrognathus lessunda sp. nov.(Figs 63–83, 114)
Types: female holotype, 1 female, 3 male paratypes, S74067, S74066, S74070, S74068 and S74069, respectively, Lombok Island (8°08' S, 117°24' E), Lesser Sunda Islands, Indonesia, col. B. Sutrino, 15 Aug. 2002, QM.—examined.
Etymology: The specific name is a contraction of Lesser Sunda Islands in the Malay Archipelago of Indonesia and, therefore, a noun in apposition.
FIGURE 63. Lyrognathus lessunda sp. nov., holotype, female in life. Photo: R. C. West.
Diagnosis: Differs from all other Lyrognathus species in the apically swollen “mushroom cap” that form bilobular spermathecae, present on both lateral and medial lobes in females and undivided scopula on tar. IV. Differs from L. crotalus in intercheliceral pegs (not spines), bilobular spermathecae and retrolateral basomedial cheliceral spines. Lyrognathus lessunda sp. nov. resembles L. achilles sp. nov., L. fuscus sp. nov.and L. robustus in having intercheliceral pegs (females and males) in combination with bilobular spermathecae and retrolateral basomedial cheliceral spines. Differs from L. achilles sp. nov. and L. fuscus sp.
Zootaxa 2362 © 2010 Magnolia Press · 25REVISION OF TARANTULA GENUS LYROGNATHUS

nov. in the apical “mushroom cap” form medial lobes of spermathecae. Differs from L. robustus in the undivided metatarsi IV scopula for the full length ventrally and the absence of a third claw on tarsi IV. Differs from L. saltator in the undivided tarsi IV scopula, intercheliceral pegs and retrolateral basomedial cheliceral spines.
FIGURES 64–70. Lyrognathus lessunda sp. nov. holotype female: 64. Carapace, dorsal view. 65. Eyes, dorsal view. 66. Sternum, labium, maxillae and coxa, ventral view. 67. Labium, ventral view. 68. Left palp, legs I, II, ventral view. 69. Abdomen, legs IV, ventral view. 70. Spermathecae, dorsal view. Scale bar = 10mm for 64, 66, 69; 5mm for 68; 2mm for 65, 67, 70. Photos: S. C. Nunn.
Description: Female holotype (in life, Fig. 63) with body length: 32.91. Color (in life): intermolt and premolt entirely reddish brown/rust, postmolt dark brown posteriorly,
shading to rust orange anteriorly. Carapace (Fig. 64): length 13.3, width 11.7 (width of anterior edge 8.7). Fovea 2.15, procurved, deep,
slightly wider than OT (Fig. 21). Carapace with 4 discernible hair types: 4 distinct types noted (C1 to C4): type C1, long, needle-form pallids border carapace entirely (darker basally): type C2, short blunt tipped,
WEST & NUNN26 · Zootaxa 2362 © 2010 Magnolia Press

basally dark gray to distally pallid, cover entire carapace evenly, but reasonably sparse: type C3, short, thin wavy pallids, form basis of carapace mat: type C4, emerge from bothria-like collars, long, distally spiniform, pallid, lie in uniform arrangement of (>4) along anterior edge of each radial groove, (>4) along anterior edge of each thoracic groove, (>6) along anterior edge of each sulcal groove, (>8) along anterior edge of each caput groove and line of >4 along medial caput ridge (between OT and fovea). >4 more located behind each side of OT, nearest median caput row.
FIGURES 71–76. Lyrognathus lessunda sp. nov., holotype female (except: paratype female QM S74066; 75, 76): 71. Abdomen and legs IV, dorsal view. 72. Left leg IV, retrolateral view. 73. Left leg IV, prolateral view. 74. Left met. and tar. IV, ventral view. 75. Left maxillae, prolateral view. 76. Left maxillary lyra, prolateral view. Scale bar = 10mm for 71–73; 5mm for 74; 2mm for 75; 1mm for 76. Photos: S. C. Nunn.
Zootaxa 2362 © 2010 Magnolia Press · 27REVISION OF TARANTULA GENUS LYROGNATHUS
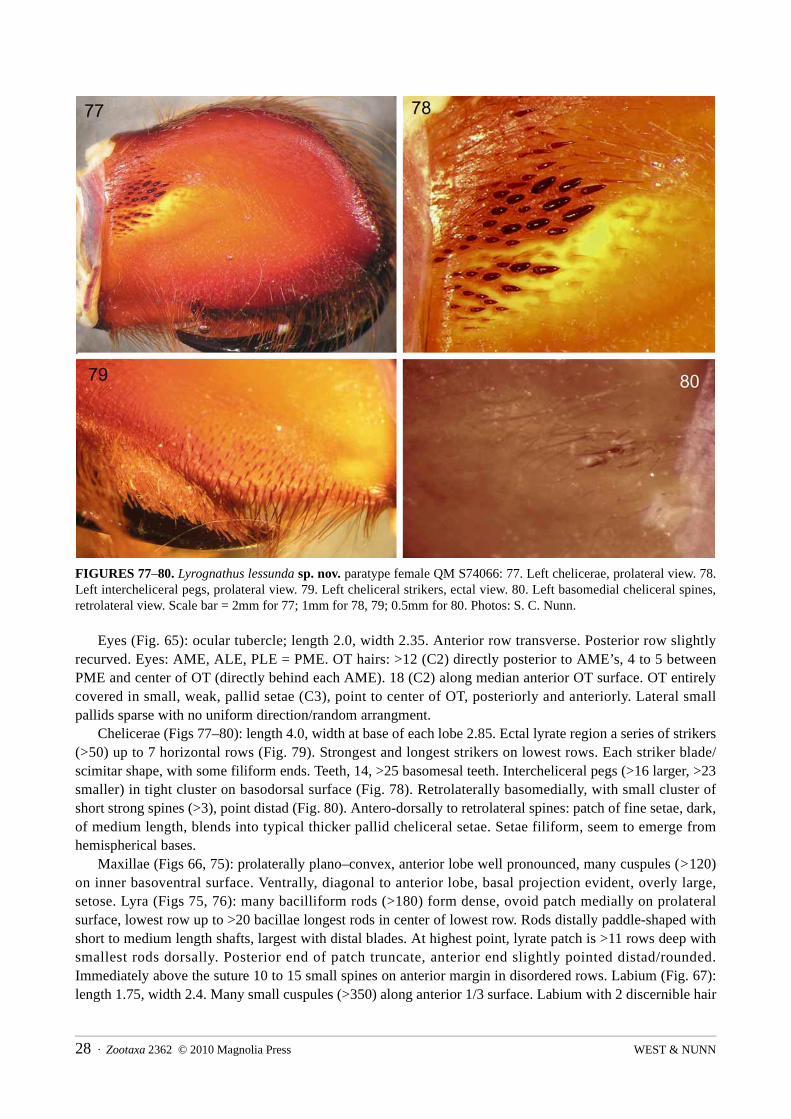
FIGURES 77–80. Lyrognathus lessunda sp. nov. paratype female QM S74066: 77. Left chelicerae, prolateral view. 78. Left intercheliceral pegs, prolateral view. 79. Left cheliceral strikers, ectal view. 80. Left basomedial cheliceral spines, retrolateral view. Scale bar = 2mm for 77; 1mm for 78, 79; 0.5mm for 80. Photos: S. C. Nunn.
Eyes (Fig. 65): ocular tubercle; length 2.0, width 2.35. Anterior row transverse. Posterior row slightly recurved. Eyes: AME, ALE, PLE = PME. OT hairs: >12 (C2) directly posterior to AME’s, 4 to 5 between PME and center of OT (directly behind each AME). 18 (C2) along median anterior OT surface. OT entirely covered in small, weak, pallid setae (C3), point to center of OT, posteriorly and anteriorly. Lateral small pallids sparse with no uniform direction/random arrangment.
Chelicerae (Figs 77–80): length 4.0, width at base of each lobe 2.85. Ectal lyrate region a series of strikers (>50) up to 7 horizontal rows (Fig. 79). Strongest and longest strikers on lowest rows. Each striker blade/scimitar shape, with some filiform ends. Teeth, 14, >25 basomesal teeth. Intercheliceral pegs (>16 larger, >23 smaller) in tight cluster on basodorsal surface (Fig. 78). Retrolaterally basomedially, with small cluster of short strong spines (>3), point distad (Fig. 80). Antero-dorsally to retrolateral spines: patch of fine setae, dark, of medium length, blends into typical thicker pallid cheliceral setae. Setae filiform, seem to emerge from hemispherical bases.
Maxillae (Figs 66, 75): prolaterally plano–convex, anterior lobe well pronounced, many cuspules (>120) on inner basoventral surface. Ventrally, diagonal to anterior lobe, basal projection evident, overly large, setose. Lyra (Figs 75, 76): many bacilliform rods (>180) form dense, ovoid patch medially on prolateral surface, lowest row up to >20 bacillae longest rods in center of lowest row. Rods distally paddle-shaped with short to medium length shafts, largest with distal blades. At highest point, lyrate patch is >11 rows deep with smallest rods dorsally. Posterior end of patch truncate, anterior end slightly pointed distad/rounded. Immediately above the suture 10 to 15 small spines on anterior margin in disordered rows. Labium (Fig. 67): length 1.75, width 2.4. Many small cuspules (>350) along anterior 1/3 surface. Labium with 2 discernible hair
WEST & NUNN28 · Zootaxa 2362 © 2010 Magnolia Press

types (L1, L2); type L1: long, reasonably dark spiniform, curving distally toward anterior, most concentrated along lateral borders. Setae longest along anterior edge (>30); type L2, 1/2 length of L1, interspersed with L1 but more needle-form.
Sternum (Fig. 66): length 7.25, width 6.2. Slightly wider posteriorly than anteriorly with 5 discernible hair types (S1 to S5): type S1, longest, thick blunt-tipped, darker basally, become pallid distally, entire cover, but sparse: type S2, elongate spiniforms, dark basally, distally pallid, on borders, most concentrated posteriorly: type S3, short spiniforms, overall sparse: type S4 short, thin pallids, form basis of mat: type S5 dark basally, but apically pallid, short blunt tipped, entirecover in sparse unorganised arrangement. Posteriorly between left/right cox. IV, border greatly acuminate, lateral sternal points also acuminate. Sigilla: 3 pairs, posterior, very large, ovoid, 1.7 lengths apart, 0.6 to 0.95 of their length from margin. Median pair 1/2 size of posterior, similar, 0.2–0.1 of their length from margin. Anterior pair somewhat obscured, on margin. Labio-sternal sigilla small, thin, but wide.
Legs. Setation: retrolateral surfaces of pat., tib., met. and tar. IV with distinct thick brushes of hair-like setae (Fig. 71), penicillate. Legs: formula; (length) IV, I, II, III, (width) IV, III, I, II. Leg RF~ 87.92. Leg lengths (fem., pat., tib., met., tar., total): holotype S74067: palp: 7.2, 4.3, 4.65, 0.00, 5.15, 21.3. I: 10.4, 6.15, 7.25, 6.3, 4.1, 34.2. II: 8.1, 5.2, 5.2, 5.25, 4.15, 27.9. III: 8.9, 4.8, 4.8, 5.6, 4.05, 28.15. IV: 10.65, 5.85, 8.25, 9.3, 4.85, 38.9. Paratype S74070: palp: 7.3, 5.15, 5.2, 0.00, 5.25, 22.9. I: 10.25, 6.7, 8.0, 7.3, 4.65, 36.9. II: 8.55, 5.9, 6.2, 6.3, 4.4, 31.35. III: 7.75, 5.25, 6.7, 6.45, 4.15, 30.3. IV: 10.3, 5.95, 9.05, 9.80, 5.35, 40.35. Leg mid-widths (fem., pat., tib., met., tar., total.): (holotype only): palp: 1.85, 2.13, 1.71, 0.00, 2.05, 7.74. I: 2.45, 2.51, 2.63, 2.1, 2.15, 11.84. II: 2.35, 2.35, 2.12, 1.95, 1.75, 10.52. III: 3.15, 2.55, 2.35, 1.95, 1.85, 11.85. IV: 2.45, 2.83, 3.02, 2.35, 2.15, 12.8.
Scopula: undivided, for full length of met./tar. on all legs.Coxae: some small black thorns prolatero-dorsally, no thorns retrolaterally on I–IV; easily seen dorsally; I
longest, about 1.6 times length of II. IV widest, as long as III, basally rectangular with rounded corners; with ventral thorns prolaterally on I–IV. I–III ventrally with many long thick blunt setae proximally, pallid. No short black setae. IV with mixture of long thick blunt setae proximally, pallid intermixed with shorter thin pallid setae. Ventral surface of I–IV gently sloping anteriorly. Retrolateral setation: I–III with median narrow light brush, IV completely cover of light setae. I–IV retrolaterally lack ventral ledge. Ventral measurements for coxae (holotype only): I–length 6.8, width 3.1; II–5.8, 2.9; III–5.1, 2.8; IV–6.1, 3.4. Trochantera: I–length 2.6, width 2.7; II–2.6, 2.1; III–2.6, 2.8; IV–3.3, 3.1.
Trichobothria: on all tar., basal filiform field slightly wider than clavate field, merging evenly. Clavates on tar. I only in distal 1/2, long filiforms only in basal 1/2, shorter filiforms intermixed with clavates distally. Clavate extent on II–IV cf. I, only in distal 1/2. Shorter filiforms for length. Short epitrichobothrial field (Raven 2005) on I shorter than clavates, uniform height for length. Tarsal organ not evident on legs I–IV.
Spines: met. I with 1 DV, met. II with 1 DV, 1 DPV, 1 DRV, met. III with 2 DV, 1 DPV, 1 DRV, 1 DD and met. IV with 2 DV, 1 DPV, 1 DRV, 1 DD, 1 DPL.
Claws: unarmed paired claws on all legs. No reduced third claw on leg IV.Abdomen (Figs 69, 71): ovular, elongate, reddish brown; with 4 discernible hair types (A1 to A4):
dorsally with 3 hair types: type A1, long, dark blunt-tipped, uniform color: type A2, mid-length, also dark, but distally spiniform, unlike type A1: type A3, form dense mat, mid-length, uniformly dark, needle-form (some stand erect, some run posteriorly but with similar morphology). Longest hairs (type A1) more concentrated posteriorly toward spinnerets, point distad, but proximally, hairs stand erect, medially curved (toward posterior). Ventral hair types almost same as dorsal, but type A2 and A3 more dense entirely. Type A4 hair, in very dense patch, trapezoidal, posterior of pedicel between median regions of cox. IV. Hairs of medium length, straight, pallid. Between anterior pair of booklungs, setal mat similar to rest of venter.
Genitalia (Fig. 70): epigastric fold 4.95. Spermathecae bilobular. Lobes apically swollen with lateral lobes 2/3 length of medials. Medial lobes very long. Epigastric fold extends ca. 2/3 length of medial spermathecal lobes.
Spinnerets: PMS: length 1.45, width (medially) 0.65. PLS: section lengths; basal 2.85, medial 1.30, apical 2.10. PLS section widths (medially); basal 1.32, medial 0.98, apical 0.67; with 2 discernible hair types (SP1,
Zootaxa 2362 © 2010 Magnolia Press · 29REVISION OF TARANTULA GENUS LYROGNATHUS

SP2): SP1 very short and distally blunt pallid, covers both pairs ventrally; second form (SP2), very similar but up to twice as long, dorsally on all segments.
Male paratype (Fig. 81, as in female except) with body length: 30.28. Color (in life): reddish brown with slight purple sheen in natural light.Carapace (Fig. 84): length 11.9, width 9.35 (width across anterior edge 6.9). Fovea 1.75 wide, deep,
procurved.
FIGURES 81–90. Lyrognathus lessunda sp. nov. paratype male QM S74069: 81. Habitus, dorsal view. 82. Carapace, dorsal view. 83. Eyes, dorsal view. 84. Sternum, labium, maxillae and coxa, ventral view. 85. Right intercheliceral pegs, prolateral view. 86. Right cheliceral strikers, ectal view. 87. Right maxillae showing lyra, prolateral view. 88. Left bulb, retrolateral view. 89. Left bulb, prolateral view. 90. Left bulb, anterior view. Scale bar = 10mm for 81; 5mm for 82, 84; 2mm for 83, 86, 87; 1mm for 85, 88–90. Photos: S. C. Nunn.
Eyes (Fig. 83): ocular tubercle; length 1.86, width 2.17. Anterior row transverse. Posterior row slightly recurved. Eyes, largest to smallest: AME, ALE, PLE, PME.
Chelicerae (Fig. 86): length 3.9, width at base of each lobe 2.5. Teeth,12, >15 smaller basomesals. Intercheliceral pegs (>9 larger, >15 smaller) in tight cluster on basodorsal surface (Fig. 85).
WEST & NUNN30 · Zootaxa 2362 © 2010 Magnolia Press

Maxillae (Fig. 87): anterior lobe well pronounced with many cuspules (>140) on inner basoventral surface. Lyra (Fig. 87): immediately above suture 10–15 elongated spines medially to anterior margin. Bacillate rods relatively slightly shorter than in female. Labium (Fig. 84): length 1.95, width 2.65. Many small cuspules (>300) along anterior 1/4 surface.
Sternum (Fig. 84): length 5.85, width 5.05. Wider posteriorly than anteriorly, with thin but even cover of short setae, interspersed with many longer darker setae. No spines around edges. Lateral points acuminate, but not posterior angle.
Legs: formula; (length) IV, I, II, III; (width) III, IV, I, II. Leg RF~97.07 (S74069). Lengths (fem., pat., tib., met., tar., total): paratype S74069: palp: 7.25, 4.7, 6.9, 0.00, 2.4, 21.25. I: 11.4, 6.7, 9.4, 8.45, 5.45, 41.4. II: 10.4, 5.5, 8.15, 7.45, 4.6, 36.1. III: 8.55, 5.3, 5.5, 6.75, 4.15, 30.25. IV: 11.85, 5.8, 9.35, 10.85, 4.8, 42.65. paratype S74068: palp: 6.9, 4.75, 6.35, 0.00, 2.35, 20.35. I: 11.1, 6.15, 9.45, 8.1, 5.15, 39.95. II: 9.05, 5.35, 7.2, 7.25, 5.05, 33.9. III: 8.0, 4.85, 5.35, 6.85, 3.95, 29.0. IV: 11.25, 5.55, 9.25, 11.45, 4.75, 42.25. paratype S74066: palp: 8.6, 5.0, 7.1, 0.00, 2.45, 23.15. I: 12.15, 6.75, 10.6, 9.15, 5.9, 44.55. II: 10.0, 5.75, 8.05, 8.15, 5.25, 37.2. III: 8.45, 5.3, 5.65, 7.85, 4.25, 31.5. IV: 12.25, 5.65, 10.25, 12.65, 5.15, 49.15. Leg mid-widths (fem., pat., tib., met., tar., total.): paratype S74069: palp: 1.85,1.95, 1.9, 0.00, 2.03, 7.73. I: 2.26, 2.45, 1.98, 1.47, 1.35, 9.51. II: 2.25, 2.17, 1.95, 1.37, 1.27, 9.01. III: 3.15, 2.3, 2.1, 1.37, 1.51, 10.43. IV: 2.35, 2.37, 2.27, 1.52, 1.36, 9.87. Leg IV with reduced retrolateral setal brush, most concentrated on retrolateral metatarsal surface, proximally, but on tib. (Fig. 81). Coxae ventral measurements (S74069): palp–length 5.97, width 2.66; I–5.52, 2.87; II–4.69, 2.67; III–4.2, 2.73; IV–5.44, 3.2. Trochantera: palp–length 2.31, width 1.87; I–3.36, 2.78; II–3.04, 2.69; III–2.34, 2.7; IV–2.96, 2.86.
Claws: as in female, except on apical ventral region of paired claws of IV, 2–3 small teeth.Genitalia. Palp (Figs 88–90): tib. mildly incrassate, tapers distally. Cymbium bipartite, tegulum large.
Lanceolate embolus slightly tapers distally with very little flaring, proximally emerges anterior to tegulum. Relative length of palpal tibia ca. twice length of embolus. Longitudinal keel distinct (embolus not terete), runs entire length of embolus, twisted through 90° prolaterally at tip (Fig. 90).
Distribution and natural history (Fig. 114): The natural history of this spider remains unknown. It is claimed to live in ground burrows on the higher slopes of Gunung Rinjani, Lombok Island, Lesser Sunda Islands, Indonesia (G. Albanjari and I. Wayan Suana, pers. comm.).
Remarks: This is the first confirmed record of any theraphosid species on Lombok Island. Roewer (1942, p. 265) listed Phlogiellus inermis (Ausserer 1871) from Lombok Island, however, the listing is probably erroneous as no description or record of P. inermis, previous or since, cites this spider from Lombok and the original type locality for the specimens Ausserer examined were from Java and Sumatra only (Ausserer 1871, p.189).
Lyrognathus robustus Smith 1988(Figs 91–112, 114)
Lyrognathus robustus Smith 1988: 15–19, figs1–9; Platnick, 2009.Lyrognathus liewi West 1991: 615–619, figs 1–11. Schmidt & von Wirth 1992: 10.
Types: Lyrognathus robustus Smith 1988, female holotype, 1988.2.6, Cameron Highlands, Malaysia, (unknown collector and date), NHM.–images (R. Raven, pers. comm.)–examined; Lyrognathus liewi West 1991, female holotype, 2 female paratypes, S15460, S15461 and S15462, respectively, 24 mile road marker, Cameron Highlands (4°41' N, 101°52' E), Pahang, West Malaysia, col. K. C. Liew, Oct.–Nov. 1986, QM.–images (R. Raven, pers. comm.)–examined.
Other material: 3 females, 1 male, S74339, S74341, S74342 and S74340, respectively, Cameron Highlands, Pahang, West Malaysia, QM–examined; 1 male, S88073, Maxwell Hill (= Bukit Larut, 4°52′ N, 100°48′ E), Perak, West Malaysia, QM–examined ; 2 females, ZRC ARA.747, Taiping (4°51' N, 100°48' E), Perak, West Malaysia, col. H. C. Abraham, 1 Nov. 1925, RMBR.–images (D. Court, pers. comm.)–examined.
Zootaxa 2362 © 2010 Magnolia Press · 31REVISION OF TARANTULA GENUS LYROGNATHUS

Diagnosis: Differs from other Lyrognathus species in the basally incrassate metatarsi IV that tapers distally (Fig. 99). Also differs from all other species in having uniquely divided scopula on metatarsi IV, a third claw on tarsi IV (Fig. 98 and see: Character discussion) and bilobular spermathecae without apical swelling (Fig. 101).
FIGURE 91. Lyrognathus robustus, adult female habitus, dorsolateral view. Photo: R. C. West.
Description: female QM S15641 (unless otherwise stated) with body length: 38.14. Color (in alcohol): reddish brown with darker femora on all legs. Carapace: length 16.7, width 12.8 (width across anterior edge 9.6). Fovea 2.3, procurved, deep, very wide.Eyes (Fig. 93): ocular tubercle; length 1.85, width 2.65. Anterior row transverse. Posterior row slightly
recurved. Sizes: AME, ALE, PLE, PME. Chelicerae (Figs 95, 96): length 6.05, width at base of each lobe 4.25. Ectal lyrate region a series of
strikers (>80), up to 7 disordered horizontal rows (Fig. 96). Strongest and longest strikers on lowest rows. Each striker needle-form with some filiform ends. Teeth: 14 (with >2 small teeth proximally) and >60 basomesal teeth. Intercheliceral pegs (>7 larger, >20 smaller) in tight cluster on basodorsal surface (Fig. 95). On retrolateral surface basomedially, with small cluster of short strong spines (>3) point distad (Fig. 96).
Maxillae (Figs 94, 97): prolaterally plano-convex. Anterior lobe well pronounced. Many cuspules (>320) on inner basoventral surface. Diagonally of anterior lobe, a basal projection with long stout setae. Lyra (Fig. 97): many bacilliform rods (>280) form dense, ovoid patch on medial prolateral surface, lowest row with up to >40 bacillae, longest rods finish center of lowest row. Rods distally paddle-shaped with long shafts, some with distal blades. Truncate posteriorly. At widest point, lyrate patch up to 10 rows deep with smallest rods dorsally. Immediately above suture, 30 small spines on anterior margin. Labium: length 2.55, width 3.67. Many small cuspules (>600) along anterior 1/3 surface.
Sternum: length 7.65, width 6.9. Slightly wider posteriorly than anteriorly, with even cover of short setae, interspersed with longer darker setae. Lateral and posterior points acuminate with series of short stout setae cover posterior points. Sigilla: 3 pairs; posterior pair, large, 3.2 lengths apart, 1.85 of their length from margin. Median pair 1/2 size of posterior, similar shape, border margin. Anterior pair smallest, somewhat obscured,
WEST & NUNN32 · Zootaxa 2362 © 2010 Magnolia Press

border margin. Labio-sternal sigilla largest, separated by their own diameter. Some strong spiniform setae around posterior edges, border cox. III/IV.
FIGURES 92–101. Lyrognathus liewi (jun. syn. L. robustus), paratype female QM S19651.15461. 92. Habitus, dorsal view. 93. Eyes, dorsal view. 94. Sternum, labium and right maxillae, ventral view. 95. Intercheliceral pegs, prolateral view. 96. Right chelicerae, lower retrolateral view showing strikers and arrow pointing to retrolateral basomedial cheliceral spines. 97. Right maxillae showing lyra, prolateral view. 98. Right leg IV, ventral view. 99. Right leg IV, retrolateral view. 100. Right tar. IV, prolateral view showing transverse suture and reduced third claw. 101. Spermathecae, dorsal view. Scale bar = 10mm for 92, 98, 99; 5mm for 100; 2mm for 93–97; 1mm for 101. Illustrations: S. C. Nunn.
Leg setation: retrolateral surfaces of pat., tib. and met. IV with distinct thick short brushes of penicillate setae. Legs: formula; (length only) IV, I, II, III. Leg RF~82.67. Leg lengths (fem., pat., tib., met., tar., total): paratype S15460: palp: 9.08, 5.76, 6.49, 0.00, 5.94, 27.18. I: 14.02, 9.02, 9.9, 8.42, 4.95, 46.31. II: 11.72, 7.87, 7.98, 7.7, 4.57, 39.84. III: 10.78, 6.99, 6.77, 8.91, 4.4, 37.85. IV: 14.69, 8.8, 12.21, 15.4, 5.5, 56.6. S15460: palp: 9.15, 5.65, 5.85, 0.00, 5.9, 26.55. I: 11.95, 7.8, 9.3, 7.35, 5.35, 41.75. II: 10.2, 6.85, 6.45, 6.7, 5.0, 35.2.
Zootaxa 2362 © 2010 Magnolia Press · 33REVISION OF TARANTULA GENUS LYROGNATHUS

III: 8.95, 5.85, 5.6, 7.65, 5.2, 33.25. IV: 13.25, 7.1, 11.25, 12.9, 6.0, 50.5. Tib. IV incrassate. Met. IV basally incrassate, tapers distally. Tar. IV with transverse suture at distal 1/3 of tarsus, most obvious ventrally and laterally (Fig. 100).
Scopula: on met./tar. I–IV. Undivided on met./tar. I–III. Met. and tar. IV divided by band of extremely strong spiniform setae with scopula only on apical 1/3 of met. IV (Fig. 98).
FIGURES 102–112. Lyrognathus robustus male QM S74340. 102, Habitus, dorsal view. 103. Eyes, dorsal view. 104. Sternum, labium and right maxillae, ventral view. 105. Right intercheliceral pegs, prolateral view. 106. Right lower retrolateral cheliceral surface showing strikers and retrolaeral basomedial spines. 107. Right maxillae showing lyra, prolateral view. 108. Right leg IV, ventral view. 109. Right palpal tibia, prolateral view. 110. Right bulb, retrolateral view. 111. Right bulb, prolateral view. 112. Left embolus, transverse cross section. Scale bar = 20mm for 102; 10mm for 108; 5mm for 109; 2mm for 104, 107, 110, 111; 1mm for 103, 105, 106; 0.5mm for 112. Photo & illustrations: S. C. Nunn.
WEST & NUNN34 · Zootaxa 2362 © 2010 Magnolia Press

Coxae: many short black thorns prolatero-dorsally, no thorns retrolaterally I–IV; easily seen dorsally. I longest, ca. 1.8 times length of II. IV widest, 1.3 times longer than III, basally rectangular with rounded corners; with ventral thorns prolaterally on I–IV. I–III ventrally with many long thick blunt setae, pallid, some short black setae intermixed. IV with mixture of long thick blunt setae, pallid intermixed with shorter thin pallid setae. Ventral surface of I–IV gently sloping anteriorly. Retrolateral setation: I–III with median narrow light brush, IV completely covered in light setae. I–IV retrolaterally lack ventral ledge, ventrally with uniform setation. Ventral measurements: I length 7.2, width 3.1; II 6.1, 3.1; III 5.8, 2.9; IV 6.7, 4.4. Trochantera (ventral): I length 4.0, width 3.8; II 3.4, 3.1; III 2.8, 2.7; IV 4.0, 3.9.
Trichobothria: tarsi: on all, basal filiform field slightly wider than clavate field, merges evenly. Clavates on tar. I only in distal 1/2, long filiforms only in basal 1/2, shorter filiforms intermixed with clavates distally. Clavate extent on II–IV cf. I, only in distal 1/2. Shorter filiforms for length. Short epitrichobothrial field on I shorter than clavates, increasing in height proximally. Tarsal organ visible (coniform, high domed), positioned immediately behind prolateral paired tarsal claw, but obscured by border setae, on legs I–IV.
Spines: met. I with 1 DV, met. II with 1 DV, 1 DPV, 1 DRV, met. III with 2 DV, 1 DPV, 1 DRV, 1 DD and met. IV with 2 DV, 1 DPV, 1 DRV, 1 DD, 1 DL. Disordered cluster of small, short spines cover entire ventral region of met. IV outside scopulate area (Fig. 98).
Claws: unarmed paired claws on all legs. Reduced third claw on leg IV (Fig. 100). Genitalia (Fig. 101): spermathecae typically bilobular, dividing basally. Lobes not apically swollen, with
lateral lobes 2/3 length of medials. Medials very long. Heavily sclerotized distally, proximally reduces to weak sclerotization at bases of all lobes. N.B.: West (1991) noted extreme variation in types held at the QM. In one example (type: S15641), the spermathecae were trilobular, in another (type: S15460), a bud was present on one of the medial lobes.
Male QM S74340 (Fig. 102, as in female except) with body length: 28.54. Color (in alcohol): dark brown in natural lighting. Carapace (Fig. 102): length 11.2, width 10.1 (width across anterior edge 6.3). Fovea 1.8, distinctly
procurved, deep. Eyes (Fig. 103): ocular tubercle; length 1.2, width 2.1. Anterior row transverse. Posterior row slightly
recurved. Sizes: AME, ALE, PLE, PME. Chelicerae: length 6.7, width at base of each lobe 4.8. Teeth: 12 with >40 small basomesal teeth.
Intercheliceral pegs (>2 larger, >13 smaller) in tight cluster on basodorsal surface (Fig. 105).Maxillae (Figs 104, 107): anterior lobe well pronounced, many cuspules (>280) on inner basoventral
surface. Lyra (Fig. 107): immediately above suture no elongated spines (only typical hair-like setae) medially to anterior margin. Labium (Fig. 104): length 1.4, width 2.3. Many small cuspules (>500) along anterior 1/4 surface.
Sternum (Fig. 104): length 5.4, width 4.2. Clearly wider posteriorly than anteriorly, with even cover of short setae, interspersed with many longer darker setae. Differs from females in that no spiniform setae around edges.
Legs: formula; (length) IV, I, II, III; (width) III, IV, I, II. Leg RF~93.45. Leg lengths (fem., pat., tib., met., tar., total): palp: 10.1, 5.15, 9.05, 0.00, 2.9, 27.2. I: 12.5, 6.75, 10.6, 9.55, 6.43, 45.83. II: 11.2, 5.9, 8.94, 8.95, 5.6, 40.59. III: 9.05, 4.65, 6.7, 8.85, 5.53, 34.78. IV: 12.6, 6.15, 10.4, 14.55, 5.34, 49.09. Leg mid-widths (fem., pat., tib., met., tar., total.): palp: 2.15, 2.03, 2.02, 0.00, 2.4, 8.6. I: 2.45, 2.35, 2.2, 1.95, 1.63, 10.58. II: 2.44, 2.35, 2.15, 1.87, 1.5, 10.31. III: 3.9, 2.9, 2.05, 1.83, 1.75, 12.43. IV: 2.43, 2.85, 2.3, 1.82, 1.72, 11.12. Coxae ventral measurements: palp–length 4.99, width 2.16; I–5.21, 2.66; II–4.94, 2.66; III–4.04, 2.67; IV–4.52, 2.70. Trochantera ventral: palp–length 1.79, width 1.97; I–2.16, 2.29; II–2.05, 2.17; III–2.13, 2.41; IV–2.16, 2.45.
Genitalia. Palp (Figs 109–112): tib. with dorsal arrangement of distinct long pallid sensory setae 2–1–2 (proximally, medially and distally), distal pair with medial bend dorsally, as in typical metatarsal distal paired sensory setae (Fig. 109); embolus tip tapers distally with no flaring (Fig. 110). Twisted through 90° at tip. Relative length of tib. about twice length of embolus. No longitudinal keel (Fig. 112).
Zootaxa 2362 © 2010 Magnolia Press · 35REVISION OF TARANTULA GENUS LYROGNATHUS

Distribution and natural history (Fig. 114): L. robustus is found among the wet montane forested regions of Cameron Highlands, Pahang State, in the main Central range and from Baling, Kedah State and Bukit Larut and Gerik, Perak State in the Eastern range of West Malaysia. L. robustus construct simple tubular burrows in clay-like soil with either a plain opening or a slight flare-mouth that utilizes surrounding organic debris or vegetation around the mouth on sloped or nearly vertical embankments. Adult female burrows were J-shaped and terminatined in an enlarged chamber about 18–30 cm deep. An ultimate male was found in a burrow in January (S. Hogg, pers. comm.). A “nervous” spider, L. robustus will readily bite if provoked (West, pers. obs.). The local native Orang Asli people know these tarantulas as “Tei Foo” (translates as “Earth Tiger”) and still believe the bite to be fatal (West 1991, p. 616).
Remarks: Von Wirth (2000, p.17) discussed some issues between L. robustus and Coremiocnemis species within Malaysia, compared to Lyrognathus of India. He assumed it might be best to transfer L. robustus into Coremiocnemis based on plesiomorphic metatarsal scopula division on leg IV that L. robustus shares with Coremiocnemis, as well as presence of intercheliceral pegs and bilobular spermathceae, not known in Lyrognathus prior to Smith’s L. robustus description. He also considered zoogeographic ranges for both groups. However, with the inclusion of three new Lyrognathus species that display traits diagnostic of Lyrognathus (undivided scopula on metatarsi IV), but share with L. robustus the presence of intercheliceral pegs and bilobular spermathecae and further, are found within Indonesia, the previously considered dubious placement of this species into Lyrognathus is no longer questionable. Further, clavate trichobothria on tarsi of L. robustus are like that in all other Lyrognathus and not those of any Malaysian Coremiocnemis species. Male embolus morphology does not conform to Coremiocnemis either, lacking a distal “kiss curl” (West & Nunn, in press). There is at most likely a close relationship shared between Lyrognathus and Coremiocnemis by way of several shared derived characters (see: retrolateral cheliceral spines and spermathecae in Character discussion), however, the presence of penicillate retrolateral setal brushes on tibiae IV in combination with incrassate tibiae IV in females and ventral projection of the male embolus is seen only in Lyrognathus. L. robustus shows some traits that differ from other members of the genus and, in that regard, might be considered the most extreme representative of Lyrognathus, but cannot be considered separate to the group as it clearly does not fit Coremiocnemis or any other selenocosmiine genus as suggested by von Wirth (2000). The inclusion of L. achilles sp. nov., L. fuscus sp. nov. and L. lessunda sp. nov., herein, shows that traits such as intercheliceral pegs, bilobular spermathecae, retrolateral basomedial cheliceral spines and zoogeographic ranges outside of India clearly fit parsimoniously within Lyrognathus.
Even though L. liewi was placed as a junior synonym of L. robustus, a subsequent search through the senior author’s collection yielded a male, now deposited in the QM. A second L. robustus male was located near the summit of Bukit Larut, Perak, West Malaysia by Mr Stephen Hogg, and then donated to the QM. So far no other males from L. robustus have ever been located, in these specimens, informative characters exist that are not present in females, hence the need to define the male here.
Lyrognathus saltator Pocock 1900(Figs 113, 114)
Lyrognathus saltator Pocock 1900: 203.
Types: Lyrognathus saltator Pocock 1900, female holotype, 1897.6.24.24, North Khasi Hills, Meghalaya State, Northeast India, NHM.—images, illustrations (A. Smith, von Wirth, pers. comm.).
Diagnosis: Differs from L. crotalus in the 6 apical spines on metatarsi lV and only mildly incrassate tibiae IV. Differs from L. achilles sp. nov., L. fuscus sp. nov., L. lessunda sp. nov. and L. robustus in the intercheliceral spiniform setae (not pegs) and absence of retrolateral basomedial cheliceral spines. Legs: Leg formula length, IV, I, II = III (49, 37, 32, 32) and width, IV, III, I, II. Leg RF~75.51. Male unknown.
Description: Female holotype with body length: 32. Color (in life, Fig. 113): dark grey to black except pat., tib., met. and tar. of legs I and II, ventrally dark
grey to black, except pat., tib., met. and tar. of legs I and II.
WEST & NUNN36 · Zootaxa 2362 © 2010 Magnolia Press

Carapace (Fig. 1): length 15, width 12.5. Fovea wide, procurved, deep, equal in width to OT. Eyes: AME, ALE, PLE, PME. Anterior row transverse, posterior row recurved. OT highest posteriorly
behind AME, sloping gradually anteriorly. Chelicerae: ectal lyrate region a series of strikers (>70) in 7 horizontal rows. Strongest and longest strikers
on lowest rows. Each striker needle-form. Teeth, 10, >50 basomesal teeth. Intercheliceral spines (>7) in tight cluster on basodorsal surface. Retrolateral surface lacks basomedial spine cluster.
FIGURE 113. Lyrognathus species (possibly L.saltator) from North Bangladesh. Photo: S. Molur.
Maxillae: prolaterally plano-convex, anterior lobe well pronounced, numerous cuspules (>125) on inner basoventral surface. Lyra: bacilliform rods (>160) form a dense, ovoid patch on prolateral maxillary surface, lowest row with >25 bacillae, longest rods medially in lowest row. Rods distally paddle-shaped with medium to long shafts, largest rods lacking distal blades. At widest point, lyrate patch 11 rows deep with smallest rods dorsally. Posterior end of patch truncate but rounded, anterior end rounded. Immediately above maxillary suture >2 small spines on anterior margin, disordered arrangment. Labium: numerous small cuspules (>450) along anterior 1/4 surface.
Sternum (damaged): saddle-form. Posteriorly between left and right coxae IV, border highly acuminate, lateral sternal points slightly acuminate. Sigilla: 3 pairs (not including labio-sternal sigilla), posterior, large in size. Ovular, 1.5 of their length apart, 0.8 (left sigilla only) of length from sternal margin. Median pair, 1/2 size of posterior, similar in form and 0.6 (left sigilla only) of length from sternal margin. Anterior pair very small, somewhat obscured and border l margin. Labio-sternal sigilla damaged.
Zootaxa 2362 © 2010 Magnolia Press · 37REVISION OF TARANTULA GENUS LYROGNATHUS

Legs (Fig. 113): formula (length); IV, I, II, III: (width); IV, III, I, II. Leg RF~75.51. Leg lengths: palp: 22.5, I: 37, II: 32, III: 32, IV: 49. Tib. IV shows strong retrolateral villous setal fringe, proximally to distally uniform–entire (Fig. 113).
Scopula: met. I–IV undivided, tar. I-III. undivided, tar. IV divided by 2-3 rows of setae. Met. I–IV, cover full length ventrally.
Trichobothria: on all tarsi basal filiform field slightly wider than clavate field, merges evenly. Clavates on tar. I in distal 1/2 (very dark), long filiforms only in basal 2/3, shorter filiforms intermixed with clavates distally. Clavate extent on tar. II–IV cf. I, in distal 1/2. Shorter filiforms for length. Short epitrichobothrial field on I shorter than clavates, increasing in length proximally.
Spines: met. I with 1 DV, met. II with 1 DV, 1 DPV, 1 DRV, met. III with 2 DV, 1DPV, 1 DRV, 1 DD, 1 DPL and met. IV with 2 DV, 1DPV, 1 DRV, 1 DPD, 1 DD.
Claws: unarmed claws on all legs and palps. Reduced third claw absent on leg IV. Abdomen (Fig. 113): ovular, elongated, yellow brown in alcohol, black in life. Genitalia: unknown.Distribution and natural history (Fig. 114): Known only from the type locality North Khasi Hills,
Meghalaya State, Northeast India. Found in sympatry with L. crotalus. The natural history of this species is unknown.
Remarks: The holotype female of L. saltator, 1897.6.24.24, NHM, was originally a dry specimen with abdomen stuffed with cotton wool that is now preserved in alcohol. It is too brittle for dissection and examination of spermathecae.
Von Wirth (2007, pers. comm.) noted the 6 apical spines on metatarsi IV in Pocock’s L. saltator holotype. This is the only confirmed example from this species known (to the authors), whether or not this is just an example of L. crotalus could not be determined by the authors through external examination of the type alone, spermathecae have never been extracted due to brittle condition of the specimen. Variation of leg segment widths (following Pocock 1900) is dubious, M. Siliwal and S. Rafn (2008, pers. comm.) have noted similar variation in L. crotalus specimens gathered from type locations. No other variation from L. crotalus is noted. Due to limited material representing L. saltator, and fragile condition of the type, no further attempt to recognise or transfer this species has been undertaken here (other than noting the significance of the 6 apical metatarsi IV spines).
Character discussion
Use of characters in systematics of Selenocosmiinae:Leg IV morphology. Legs of the Theraphosidae have been analyzed since the group was diagnosed:
several characters have been used: tarsal scopula division and function (Pocock 1897, Gerschman de Pikelin & Schiapelli 1973, Perez-Miles 1994, Guadanucci 2005), claw morphology (Pocock 1900), setal counts and trichobothrial analysis (Raven 1985, 2005). Raven (1985, see: Anterior–posterior Gradations, p. 12) discussed the importance of leg morphology in all Mygalomorphae in relation to systematics of the group.
In most Lyroganthus, Coremiocnemis and Selenocosmia species examined, a metatarsal IV spine arrangement of 2 DV, 1 DPV, 1 DRV and 1 DD are found in both sexes. In all other selenocosmiines examined (except Chilobrachys sericeus, Haplocosmia nepalensis and Selenocosmia obscura), an arrangement of 1DV, 1 DPV, 1 DRV, 1DL and 1 DPL metatarsal IV spines are found.
While all Lyrognathus species possess an incrassate leg IV (Figs 1, 13, 45, 63, 91), so do other non-congeners, such as Selenotypus plumipes Pocock (1895, p. 176, Pl. X, fig. 2a: NHM type) and Selenocosmia peerboomi (Schmidt 1999, SMF), although in those species leg IV is not quite as thick, the retrolateral penicillate tibiae IV brush is absent, the scopula on metatarsi IV remain divided by a line of setae and intercheliceral pegs/spines are absent. Within selenocosmiine relatives outside the Lyrognathus, greatly incrassate posterior legs (with the exception in some selenocosmiine mature males with incrassate femora III) are unknown; therefore this trait is considererd informative.
WEST & NUNN38 · Zootaxa 2362 © 2010 Magnolia Press

FIGURE 114. Records of Lyrognathus species in Southeast Asia. C. Senescall
Metatarsal scopula: L. robustus has uniquely divided scopula on IV (Fig. 98). The division is not typical among other Selenocosmiinae in that it is possibly a secondary reduction to accommodate highly incrassate proximal metatarsi IV (a possible shared derived state also seen in L. crotalus, although not as extreme). In all other Lyrognathus species, the metatarsi IV scopula covers the full length and is undivided, traits not known in any other selenocosmiine species.
Retrolateral surface of coxae IV: In all Lyrognathus species (except L. robustus), retrolateral surfaces of coxae IV are entirely hirsute, in all other selenocosmiine species examined, retrolateral surfaces of coxae IV were either entirely pallid or with medial brushes of fine setae, as in coxae I–III of all selenocosmiines examined.
Tarsal trichobothria: Trichobothria on tarsi I–IV were consistent between all Lyrognathus species examined herein. Clavate trichobothria occur in the distal halves of all tarsi dorsally, in a relatively wide and noticeable field (>30) separated by the short epitrichobothrial field. Clavates are noticeably longer than the surrounding trichobothria. Both proximally and distally, the epitrichobothria disperse (lengthening 1/5 on proximal tarsus), the distal end merges evenly with the clavates and also merges with a field of short filiforms (>8). Raven (2005, p. 25. Figs 31–33) showed morphology of epitrichobothrial setae in detail. A long filiform field occurs (>10) proximally and is slightly wider than the clavate field. In spiders presumed to be Selenocosmia javanensis, one Selenocosmia species, two further undescribed species of Selenocosmia (West & Nunn, in prep.) from Vietnam and North Thailand and all Malaysian Coremiocnemis species, the clavate
Zootaxa 2362 © 2010 Magnolia Press · 39REVISION OF TARANTULA GENUS LYROGNATHUS

field on tarsi I are very wide, found in distal 3/4 and are not so distinct from surrounding epitrichobothria as in Lyrognathus. Also, in the abovementioned Selenocosmia and Coremiocnemis, a tuft of long hair-like setae is intermixed with the proximal filiform field on tarsi IV. In Coremiocnemis, hairs in the tuft are strongly recurved and long filiform trichobothria follow this recurvature. In the above Selenocosmia, the tuft is not as obvious with long filiforms lacking recurvature. In Chilobrachys sericeus, clavate trichobothria occur in the distal halves of tarsi I and are distinct from surrounding epitrichobothria, however, the long filiform field in that species has more trichobothria than all other species examined.
Leg IV retrolateral setal brush: In all Lyrognathus species (and unique to it), a dense penicillate brush of long setae occurs on retrolateral surfaces of leg IV, along the tibiae and metatarsi (Figs 3, 20, 53, 71, 92). Selenotypus Pocock 1895 (p. 176 and Pl. X, fig. 2a), Selenocosmia peerboomi (Schmidt 1999) and Coremiocnemis; see: Pocock 1895 p. 175 and Pl. X, fig. 5, show long setae on leg IV; however, leg IV setae are evenly spread across all views, no penicillate retrolateral tibiae IV setal brush in either genus has been noted.
Tarsal organs in the Selenocosmiinae. In Mygalomorphae, tarsal organs have been extensively reviewed (Gertsch and Platnick 1979, 1980; Forster 1980; Raven 1981, 1985, 1994) and noted in all families; however, identification and examination of tarsal organs in Theraphosidae to date is lacking to completely absent. Raven (1985, p. 22) noted the tarsal organs in the Ischnocolinae genus Heterothele (low domed with shallow concentric ridges); however, R. C. Gallon (2008, pers. comm.) reported not to have found tarsal organs in any theraphosid.
During this study, tarsal organs were observed in L. achilles sp. nov., L. fuscus sp. nov., L. robustus (smooth and high domed, coniform), spiders presumed to be Selenocosmia javanensis (mid-domed), 4 undescribed Coremiocnemis species (low domed, slight raised ridges, West & Nunn, in press), 1 undescribed Orphnaecus species (high domed, not coniform, West & Nunn, in prep.), also in 3 Phlogiellus species (high domed, Nunn and West, in prep.). Prior identification of the organ in Selenocosmiinae is nonexistent. A maximum magnification of 40X was needed to view tarsal organs and, in most instances, setae needed to be either shaved or bent to one side for visibility. Morphology seems to greatly vary intragenerically, with examples easily seen in some species yet unidentifiable under stereomicroscopes in congenerics. In genera where multiple species displayed an obvious organ, morphology was very similar, only differing in size, not shape. We strongly suggest that future examination of any theraphosid include a search for and description of tarsal organs, for they occur in Theraphosidae and should be considered relatively easy to record among many Selenocosmiinae.
Retrolateral basomedial cheliceral spines. In southern Lyrognathus species (L. achilles sp. nov., L. fuscussp. nov., L. lessunda sp. nov. and L. robustus), a small patch of short, black spines (1–6) occurs on the retrolateral surfaces of chelicerae basomedially (Figs 20, 60, 80 and 96, respectively). These spines are also found in Selenocosmia and all Malaysian Coremiocnemis species, but not known in any other selenocosmiines, suggesting a relationship between the three groups. Further examination of this character trait needs to be done to see how common it is within Selenocosmia. The spines were not found in Indian Lyrognathus species.
Male bulb: Raven (1985, p. 9, 10) discussed systematic importance of the male bulb in Mygalomorphae, identified and discussed haemotodochae (1985, p. 8) and identified a reduced number of sclerites within the bulb as a unique character combination of the group. Morphology of the male bulb in Theraphosidae is considered by most authors to be useful to characterize taxa (Bertani 2000, p. 29), hence the need to look into morphology of selenocosmiine bulbs in further detail. In 2000, Bertani identified homologous characters (namely embolic keels) of male bulbs in Theraphosinae and further, developed new terminology to identify characters and their states.
The selenocosmiine bulb is attached to a bipartite cymbium via haemotodochae to the subtegulum, displaying a large and rounded tegulum, in some genera highly globose (between which lies the second haemotodochae), with a thin to robust embolus emerging anterior to the tegulum, except some species within the Australo-Papuan region in which the emboli emerge prolaterally. In some genera, the embolus is terete and displays strong distal flaring (some Australo-Papuan fauna, see Raven 2005, p. 23, figs 24, 25; and p. 27) or
WEST & NUNN40 · Zootaxa 2362 © 2010 Magnolia Press

no flaring at all. In Poecilotheria Simon 1885, the embolus is piriform and spiraliform (Pocock 1900, p. 108) and in some Chilobrachys Karsh 1891, the embolus is blade-shaped (Pocock 1900, p. 193) with multiple apical keels.
The embolus is entirely but singularly keeled within Lyrognathus, Phlogiellus and Orphnaecus. In all 3 genera, the keel begins dorsally and then turns 90° prolaterally, continuing to turn ventrallyand proximally. In Phlogiellus and Orphnaecus, the keel ends with a large basal projection ventrally. While such a projection is also present in Lyrognathus, (see Smith 1986, p. 120, fig. 19h: S. pugnax; herein: L. crotalus (Fig. 5), L. achilles sp. nov. (Fig. 40) and L. lessunda sp. nov. (Fig. 98)) it is smaller, completely absent in L. robustus (Fig. 111). The basal embolus is lanceolate in Lyrognathus, whereas in Phlogiellus and Orphnaecus the basal embolus is rather stout/incrassate–obconic (West & Nunn, in prep.). In Lyrognathus, the embolus is long, just under one-third length of palpal tibia. In both Phlogiellus and Orphnaecus, the embolus is short, just under one-quarter length of palpal tibia. In all three selenocosmiine genera, the keel turns 90° towards the tip.
Remarks: Spermathecae: There is considerable variation of spermathecae between Lyrognathus species, L. crotalus has unilobular spermathecae (Fig. 6), while L. achilles sp. nov., L. fuscus sp. nov., L. lessunda sp. nov. and L. robustus have bilobular spermathecae (Figs 23, 51, 70 and 101, respectively). Spermathecae of L. saltator are unknown (see species notes). In species with bilobular spermathecae, L. lessunda sp. nov. has apically swollen “mushroom cap” shaped lobes (medial and lateral), while in L. robustus, the lobes are not apically swollen. Both L. achilles sp. nov. and L. fuscus sp. nov. have spermathecae with medial lobes differing in morphology to laterals. While describing Lyrognathus liewi (=L. robustus), West (1991) noted extreme variation of spermathecal receptacles in types. In one specimen, spermathecae were bilobular, another bilobular with an apical bud present, yet another trilobed. Because no two species share similar spermathecal morphology, Lyrognathus could become problematic in a cladistic analysis unless all species from the group are considered for careful terminal unit (TU) selection (see: Mishler 2005). Similar to L. achilles sp. nov., L. fuscus sp. nov., L. lessunda sp. nov. and L. robustus, West Malaysian Coremiocnemis and Javan/Sumatran Selenocosmia possess bilobular spermathecae. However, in both those groups, the lobes are of equal length. In all Lyrognathus with bilobular morphology, lateral lobes are shorter than medials, which remain very long. Among all Lyrognathus species, one constant trait was noted: the distal spermathecal lobes are heaviest sclerotized, reducing to clear shafts.
Intra-generic character transition within Lyrognathus: Four distinct characters show geographic variation. The first character, intercheliceral setation, presents the first state–"strong spiniform setae/intercheliceral spines", present in L. crotalus and L. saltator (NE India) and the second state–"peg-shaped setae/intercheliceral pegs", present in all other members of the genus (West Malaysia, Borneo and Indonesia). The second intercheliceral state shows strong variation, from extremely elongated pegs (e.g., male, L. robustus), to extremely stout, being half as wide–equal width/length (L. fuscus sp. nov.). Following this shift in morphology, there is no stable state to define any members of Lyrognathus from any other selenocosmiine genus (known to the authors), namely those closest related. That said, the shift in morphology is interesting, because it is not previously noted as varying within selenocosmiine genera; previously genera could be defined as having peg-shaped setae present/absent. The second character, spermathecae, presents multiple states within Lyrognathus as mentioned above. However, there is a shift in character states that is congruous with the above shift in intercheliceral states, being first state–"unilobular spermathecae" (NE India, L. crotalus, L. saltator–unknown), with second state–"bilobular spermathecae" (all other species–West Malaysia, Borneo and Indonesia). The third character, retrolateral basomedial cheliceral spines, displays two clear states, first state–absent (all in the NE Indian material), second state–present (all West Malaysian and Indonesian members of Lyrognathus). The fourth character, cheliceral strikers, displays two clear states,–“needle-form”, found in all NE Indian Lyrognathus and in L. robustus from W. Malaysia and–“blade/scimitar-form”, found in all Indonesian species.
All Lyrognathus species known have tibiae IV penicillate retrolateral setal brushes (females and some males), leg formula of length: IV, I, II, III and width: IV, III, I, II (females and some males), incrassate tibiae and metatarsi IV (females and some males), retrolaterally hirsute coxae IV (except L. robustus), intercheliceral spines/pegs, well developed ovoid lyra on prolateral maxillary surfaces (both sexes), with
Zootaxa 2362 © 2010 Magnolia Press · 41REVISION OF TARANTULA GENUS LYROGNATHUS

corresponding spiniform or scimitar shaped strikers on ectal chelicerate surfaces (both sexes). This character combination is not known in any other selenocosmiine species. L. robustus can be considered an example from Lyrognathus that has secondarily lost 1–2 leg IV traits (undivided metatarsi IV scopula that covers the full length and retrolateral coxae IV with a hirsute surface). However, that species still retains leg IV villous retrolateral setal brushes, clearly diagnostic to Lyrognathus, as well as incrassate tibiae IV and rounded ocular tubercle morphology.
Although this genus has such a large geographic range, the presence of penicillate retrolateral setal brushes on tibiae IV, highly incrassate tibiae IV, undivided scopula on metatarsi IV, retrolateral surface of coxae IV entirely hirsute and singular, entirely keeled but lanceolate emboli are found on most Lyrognathusspecies and only with that selenocosmiine genus, suggesting five synapomorphies for the group and that all Lyrognathus discussed herein (regardless of intra–generic character transition) are clearly a monophyletic unit. This hypothesis will be tested in a cladistic analysis including all Lyrognathus, Coremiocnemis (West & Nunn, in prep) and Indo-Malayasian Selenocosmia (Nunn, West and Court, in prep).
The holotype of Selenocosmia lyra Strand (1913) from Sumatra, originally described as a female, was examined on our behalf by Dr P. Jaeger and found to be an immature male. The prolateral maxillary stridulating organ of this spider does not conform to known Selenocosmia species from Sumatra, West Malaysia or Java, in that the largest bacillae have long shafts with distal paddles, a trait seen in all Lyrognathus and Coremiocnemis species from the region, to which that species might best be placed. Therefore, Selenocosmia lyra Strand 1913 is considered nomen dubium.
Acknowledgements
Thanks to the Department of Wildlife and National Parks Peninsular Malaysia for permit numbers WL (AP/PP) 0227/00 and WL (AP/PP) 0244/01 to collect and export arachnids from the country. Thanks to Dr Robert J. Raven (QM) for assistance with technical issues and comments on the manuscript. Thanks to Mr Mark Newton for early comments on the manuscript. Ms Janet Beccaloni (NHM) is thanked for loaning type material to Australia. Thanks to Mr Stephen Hogg, Mr Sanjay Molur and Mr Jean–Michel Verdez for assistance with images. Mr G. Albanjari, Mr Stephen Hogg, Mr Soren Rafn, Dr Manju Siliwal, Dr I. Wayan Suana, Mr Jean-Michel Verdez and Mr Hari Yanto also kindly provided information on Lyrognathus species within Bangladesh, Borneo, India, Lombok Island and West Malaysia. Thanks to Mr David Court (RMBR) who assisted to identify, examine and photograph Lyrognathus material held in the RMBR. Images and illustrations of type material were kindly supplied by Drs Robert Raven and Peter Jaeger, Mr Andrew Smith, Mr Fabian Vol and Mr Volker von Wirth. Dr Owen Seeman (QM) is thanked for his valuable and tireless effort transporting and preparing type material. Ms Lua Hui Kheng (RMBR) is kindly thanked for organising registered numbers for all examined RMBR material. Dr Hieronymus Dastych (ZMH) is thanked for checking data of type specimens housed in the ZMH. Dr G. B. Edwards and Dr H. Don Cameron kindly checked Latin names and GIS mapping was performed by Mr Charles Senescall.
References
Ausserer, A. (1871) Beiträge zur Kenntniss der Arachniden–Familie der Territelariae Thorell (Mygalidae Autor). Verhandlungen der k.k. zoologisch–botanischen Gesellschaft in Wien, 21, 17–224.
Bertani, R. (2000) Male palpal bulbs and homologous features in Theraphosinae (Araneae, Theraphosidae). Journal of Arachnology, 28, 29–42.
Bonnet, P. (1957) Bibliographia araneorum. Toulouse 2 (3), 1927–3026.Brignoli, P.M. (1983) A catalogue of the Araneae described between 1940 and 1981. Manchester University Press, Manchester, 755
pp.Forster, R.R. (1980) Evolution of the tarsal organ, the respiratory system and the female genitalia in spiders. Verhandlungen
Intermationaler Arachnologen–Kongress, 8, 269–284.Gerschman de Pikelin, B.S. & Schiapelli, R.D. (1973) La subfamilia ‘‘Ischnocolinae’’ (Araneae: Theraphosidae). Revista del Museo
Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Entomologia, 4, 43–77.
WEST & NUNN42 · Zootaxa 2362 © 2010 Magnolia Press

Gertsch, W.J. & Platnick, N.I. (1979) A revision of the spider family Mecicobothriidae (Araneae, Mygalomorphae). American Museum Novitates, 2687, 1–32.
Gertsch, W.J. & Platnick, N.I. (1980) A revision of the American spiders of the family Atypidae (Araneae, Mygalomorphae). American Museum Novitates, 2704, 1–39.
Gravely, F.H. (1915a) The evolution and distribution on Indian spiders belonging to the sub-family Aviculariinae. Journal and Proceedings of the Asiatic Society of Bengal. Calcutta, 10, 411–420.
Gravely, F.H. (1915b) Notes on Indian mygalomorph spiders. I. Records of the Indian Museum, Calcutta, 11, 257–287.Gravely, F.H. (1935) Notes on Indian mygalomorph spiders. II. Records of the Indian Museum, Calcutta, 37, 69–84.Guadanucci, J.P.L. (2005) Tarsal scopula significance in Ischnocolinae phylogenetics (Araneae, Mygalomorphae, Theraphosidae).
Journal of Arachnology, 33, 456–467.Karsch, F. (1891) Arachniden von Ceylon und von Minikoy gesammelt von den Herren Doctoren P. und F. Sarasin. Berliner
entomologische Zeitschrift, 36, 267–310.Mishler, B.D. (2005) The logic of the data matrix in phylogenetic analysis. In V.A. Albert (ed.), Parsimony, Phylogeny and
Genomics. Oxford University Press. pp. 57–70.Perez-Miles, F. (1994) Tarsal scopula division in Theraphosinae (Araneae, Theraphosidae): its systematic significance. Journal of
Arachnology, 22, 46–53.Platnick, N.I. (2009) The world spider catalog, version 9.5. American Museum of Natural History, available from http://
research.amnh.org/entomology/spiders/catalog/index.html (accessed 30 June 2009)Pocock, R.I. (1895) On a new and natural grouping of some of the Oriental genera of Mygalomorphae, with descriptions of new
genera and species. The Annals and Magazine of Natural History, 15(6), 165–184.Pocock, R.I. (1897) On the spiders of the suborder Mygalomorphae from the Ethiopian Region, contained in the collection of the
British Museum. Proceedings of the Zoological Society of London, 1897, 724–774.Pocock, R.I. (1900) The fauna of British India, including Ceylon and Burma. Arachnida, London, 279 pp. Raven, R.J. (1981) A review of the Australian genera of the mygalomorph spider subfamily Diplurinae (Dipluridae: Chelicerata).
Australian Journal Zoology, 29, 321–363.Raven, R.J. (1985) The spider infraorder Mygalomorphae (Araneae): Cladistics and systematics. Bulletin of the American Museum of
Natural History, New York, 182, 1–180.Raven, R.J. (1994) Mygalomorph spiders of the Barychelidae in Australia and the western Pacific. Memoirs of the Queensland
Museum, 35(2), 291–706.Raven, R.J. (2000) Taxonomica Araneae I: Barychelidae, Theraphosidae, Nemesiidae and Dipluridae (Araneae). Memoirs of the
Queensland Museum, 45, 569–575.Raven, R.J. (2005) A new tarantula species from northern Australia (Araneae, Theraphosidae). Zootaxa, 1004, 15–28.Roewer, C.F. (1942) Katalog der Araneae von 1758 bis 1940. Bremen 1, 1040 pp.Schmidt, G. (1999) Chilocosmia peerboomi spec. n., a new theraphosid spider from the island of Negros (Philippines) (Araneae:
Theraphosidae: Selenocosmiinae). Entomologische Zeitschrift, Frankfurt–am–Main 109, 280–286 (corrections, 391–392).Schmidt, G. & von Wirth, V. (1992) Neue veröffentlichungen über vogelspinnen. Arachnologischer Anzeiger, 3 (4), 10.Simon, E. (1874) Listes d'arachnides d'Algérie. Annales Société entomologique de France, Paris 4 (5), 66, 106–107, 155.Simon, E. (1877) Etudes arachnologiques. 5e Mémoire. IX. Arachnides recueillis aux îles Phillipines par MM. G. A. Baer et Laglaise.
Annales Société entomologique de France, Paris 7 (5), 53–96.Simon, E. (1885) Matériaux pour servir à la faune arachnologiques de l'Asie méridionale. I. Arachnides recuellis à Wagra–Karoor près
Gundacul, district de Bellary par M. M. Chaper. II. Arachnides recuellis à Ramnad, district de Madura par M. l'abbé Fabre. Bulletin de la Société zoologique de France, Paris 10, 1–39.
Simon, E. (1889) Arachnides. In Voyage de M. E. Simon au Venezuela (décembre 1887-avril 1888). 4e Mémoire. Annales Société entomologique de France. Paris 9 (6), 169–220.
Simon, E. (1892) Histoire naturelle des araignées. Paris 1, 1–256.Simon, E. (1903) Histoire naturelle des araignées. Paris 2, 669–1080.Smith, A.M. (1986) The tarantula: classification and identification guide. Fitzgerald Publishing, London, 176 pp.Smith, A.M. (1987) The tarantula: classification and identification guide (second ed.). Fitzgerald Publishing, London, 178 pp.Smith, A.M. (1988) Lyrognathus robustus, a new species of theraphosid spider from Malaysia. British Tarantula Society Journal, 4
(2), 15–19.Strand, E. (1913) Neue indoaustralische und polynesische spinnen des Senckenbergischen Museums. Archiv für Naturgeschichte.
Berlin 79 (A6), 113–123.Thorell, T. (1869) On European spiders. Part I. Review of the European genera of spiders, preceded by some observations on
zoological nomenclature. Nova acta regiae Societatis scientiarum Upsaliensis. Stockholm, 7 (3), 108 pp.Thorell, T. (1895) Descriptive catalogue of the spiders of Burma. British Museum, London, 406 pp.von Wirth, V. (2000) Über die tatsächliche Identität der im Vorartikel als Lyrognathus robustus Smith, 1988 bezeichneten
Vogelspinnenart (Araneae, Theraphosidae, Selenocosmiinae). Arthropoda, 3 (8), 17–19.von Wirth, V. & Striffler, B.F. (2005) Neue Erkenntnisse zur Vogelspinnen–Unterfamilie Ornithoctoninae, mit Beschreibung von
Ornithoctonus aureotibialis sp. n. und Haplopelma longipes sp. n. (Araneae, Theraphosidae). Arthropoda, 13(2), 2–27.Walckenaer, C.A. (1837) Histoire naturelle des insectes. Aptères. Paris 1, 682 pp.West, R.C. (1991) A new theraphosid spider from the Cameron Highlands, Malaysia. Memoirs of the Queensland Museum, 30, 615–
619.West, R.C., Marshall, S.D., Fukushima, C.S. & Bertani, R. (2008) Review and cladistic analysis of the neotropical tarantula genus
Ephebopus Simon 1892 (Araneae: Theraphosidae) with notes on the Aviculariinae. Zootaxa, 1849, 35–58.
Zootaxa 2362 © 2010 Magnolia Press · 43REVISION OF TARANTULA GENUS LYROGNATHUS













![(1895-1919, 1920-1990[A], & 1991-PRESENT ... - Ngin](https://static.fdokumen.com/doc/165x107/63288d2e21c8de9666002ad3/1895-1919-1920-1990a-1991-present-ngin.jpg)






