
2005 Helmer et al Durham
13
This PDF file of your paper in First Steps of Animal Domestication belongs to the publishers Oxbow Books and it is their copyright. As author you are licenced to make up to 50 offprints from it, but beyond that you may not publish it on the World Wide Web or in any other form.
Transcript of 2005 Helmer et al Durham

This PDF file of your paper in First Steps of AnimalDomestication belongs to the publishers Oxbow Booksand it is their copyright.
As author you are licenced to make up to 50 offprintsfrom it, but beyond that you may not publish it on theWorld Wide Web or in any other form.

Proceedings of the 9th Conference of the International Councilof Archaeozoology, Durham, August 2002Series Editors: Keith Dobney, Peter Rowley-Conwy and Umberto Albarella
Edited byJ.-D. Vigne, J. Peters and D. Helmer
First Steps ofAnimal Domestication
New archaeozoological approaches
© Oxbow Books 2005
ISBN 1 84217 121 6

Contents
Preface .............................................................................................................................................................................. vii Keith Dobney, Peter Rowley-Conwy and Umberto Albarella
1. New archaeological approaches to trace the first steps of animal domestication:general presentation, reflections and proposals ......................................................................................................... 1Jean-Denis Vigne, Daniel Helmer and Joris Peters
Principles and concepts2. Experimental animal domestication and its application to the study of animal exploitation in prehistory ............... 18
Benjamin S. Arbuckle
3. The domestication of the wolf – the inevitable first? ............................................................................................... 34Werner Müller
4. Breaking the mold: a consideration of variation in the evolution of animal domestication ..................................... 41Richard W. Redding
New techniques and their application5. Assessing the origin and diffusion of domestic goats using ancient DNA ............................................................ 50
Helena Fernández, Pierre Taberlet, Marjan Mashkour, Jean-Denis Vigne and Gordon Luikart
6. Kernel smoothing and mixture analyses for the determination of the sex ratios at death,at the beginning of domestication of ungulates ....................................................................................................... 55Hervé Monchot, Marjan Mashkour and Jean-Denis Vigne
7. Two novel methods for the study of dental morphological variation in Sus scrofa,in order to identify separate breeding groups within archaeological assemblages ................................................. 61Sylvia Warman
Animal domestication in West Asia8. Pig exploitation at Hagoshrim; a prehistoric site in the Southern Levant ................................................................ 80
Annat Haber, Tamar Dayan and Nimrod Getz
9. Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphratesusing sexual dimorphism ........................................................................................................................................... 86Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Peters and Maria Saña Segui
10. The upper Euphrates-Tigris basin: cradle of agro-pastoralism? ............................................................................. 96Joris Peters, Angela von den Dreisch and Daniel Helmer
11. A View from the Zagros: new perspectives on livestock domestication in the Fertile Crescent .......................... 125Melinda A. Zeder
Animal domestication in East Asia12. Wild pig? Or domesticated boar? An archaeological view on the domestication of Sus scrofa in Japan .............. 148
Akira Matsui, Naotaka Ishiguro, Hitomi Hongo and Masao Minagawa
13. Wild boar remains from the Neolithic (Jomon Period) sites on the Izu Islandsand in Hokkaido Island, Japan ................................................................................................................................. 160Kyomi Yamazaki, Osamu Takahashi, Hiroki Sugawara, Naitaka Ishiguro and Hideki Endo

86 Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Peters & Maria Saña Segui
9. Identifying early domestic cattle from Pre-PotteryNeolithic sites on the Middle Euphrates using sexualdimorphism
Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Petersand Maria Saña Segui
9th ICAZ Conference, Durham 2002The First Steps of Animal Domestication (eds J.-D. Vigne, J. Peters and D. Helmer) pp. 86–95
Introduction
Understanding the initial process of the domestication ofanimals is one of the fundamental aspects of archaeo-zoology. The beginning of animal husbandry was acritical step for the “Neolithic Revolution” (see Cauvin2000). In the Near East, where the transition from huntingto a full production economy took two millennia, thisshift in subsistence clearly went beyond a simple changein diet. We contend that the origins of domestication arenot only environmental (Cauvin et al. 1998, Helmer etal. 1998), but also the result of sociological developmentsand cultural evolution. For the Pre-Pottery Neolithiccommunities where these changes took place first,domestication implies a prerequisite knowledge of thesocial behaviour of the species in question as well as anunderstanding of herd management in terms of theeconomic needs and objectives of the communities.
In archaeozoology, fully domesticated species can bereadily separated from their wild ancestors, but during
the initial stages of domestication this distinction is farmore difficult. The fundamental question is how toidentify from faunal remains the earliest signs ofdomestication. In order to approach this question, weexamined a large number of Bos bones from the middleEuphrates, which covers a sequence from the early 10thmillennium BC (non calibrated) with wild cattle at TellMureybet to the late 9th millennium BC at Tell Halula,where domestic cattle are present (Fig. 1).
A reduction in size is commonly considered one of thebasic criteria for identifying morphological domest-ication. But when such a reduction is observed for agiven species, in a given area and through a well-datedarchaeological sequence, one a priori cannot be sure ofthe cause of this variation. In fact, it can be the result ofat least four different factors: climatic deterioration, over-hunting, selective hunting of females or, more pertinently,domestication. In order to identify domestication one mustassess the potential influence of all these factors.
The identification of domestication in ruminants is usually carried out using biometrical analysis and comparingthe results with data from wild specimens. For cattle as well as other taxa, the major difficulty when one observesa reduction in size is to establish if this reduction is due to climatic factors, a cynegetic stress (over-hunting), apreferential hunting of females, or as a result of domestication. While in the first two cases, one must rely on otherstudies or even complementary disciplines, biometrical analysis is essential for sex differentiation which isfundamental for distinguishing between selective hunting of females and domestication. The methods regularlyemployed in archaeozoology have a major drawback because it is not possible to determine the number, the meanor the standard deviation for each sex. Thus, we had used two new methods: the Kernel analysis and the mixtureanalysis.
The recent excavations (Göbekli, Dja’de, Halula) and the study of material from Mureybet provide a significantsample from a relatively small region with comparable climatic conditions, where sites are separated by less than150 km and range from PPNA to Middle PPNB.
On condition that the distinction between male and female specimens is carried out, it is possible to provideevidence for cattle domestication in the region from the Early PPNB onwards.

87Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates
Climatic changes in the area for the period concerned(Dryas III and the Preboreal) appears to have been of lowamplitude (Bottema 1995, Helmer et al. 1998). Theearliest levels considered in this study coincide with theend of the Dryas III, which witnessed a climaticamelioration, but for the period which follows there is noevidence in the region for any other major climatic event.
Over-hunting, which could induce size reductionthrough stress, certainly would result in frequencieschanges of the major large species. That the frequenciesof Bos examined here do not vary greatly during thecrucial period (PPNA and Early PPNB) implies that over-hunting did not occur. In addition, evidence from othertaxa which were not domesticated but frequently hunted,such as equids and gazelles, indicate no change in size(Helmer et al. 1998; Peters et al. 1999), which supportsthe hypothesis that neither climatic change nor over-hunting affected the mammalian faunas during the periodconcerned.
Thus, while the first two factors can a priori beexcluded as responsible for the reduction in size in theBos samples studied, accurate sex determination isrequired in order to be able to distinguish betweenselective hunting of wild females on the one hand anddomestication of Bos on the other hand. This particularissue will be dealt with in the following.
Methods
For the Bos remains from each site, the sex ratio wascalculated using the second anterior phalanges, themagnum (=carpal II+III), the astragalus and the centro-quartal bone, which are relatively frequent and oftencompletely preserved, and from which standard measure-ments are easy to take. In order to compare the sizevariation in Bos from different archaeological sites,measurements based on the second anterior phalangesand the magnum were used because sexual dimorphismin the forelimbs is proportionally more developed. Thebones of the hind limb (astragalus, centroquartal)provided similar results, sexual dimorphism, however,being less pronounced. The latter bones will therefore betaken into account only for the estimation of the sexratio.
By plotting measurements of a given bone in the formof scatter diagrams, clusters can be recognized even incases where sexual dimorphism is poorly developed. Butin case there is strong overlapping between populations,it is not possible to calculate the number, the mean andthe standard deviation for each group. This precludes areliable estimate of the sex ratio.
Log-Size-Index (LSI) is a commonly applied statisticalmethod for distinguishing size variation between different
Aleppo
Damascus
Euphrates
Tig
ris
Me
dite
rra
ne
an S
ea
1
2
3
45
6
7
Figure 1. Map of the sites which are refered to in the text: 1=Neval� Çori, 2=Göbekli Tepe, 3=Dja’de, 4=Halula,5=Jerf El Ahmar, 6=Cheikh Hassan, 7=Mureybet.
88 Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Peters & Maria Saña Segui
populations of a given species (e.g., Meadow 1999).However, since various parameters are combined usingthis method, differences related to the sex of the animalswithin a given population cannot be distinguished fromthe effect of potential allometries on the overall dis-tribution.
In order to solve these problems, we applied two newstatistical methods which were modified by Monchot andLéchelle (2002) to test archaeological samples. The firstmethod or mixture analysis is based on the method ofmaximum likelihood. The non-parametric bootstrap andjack-knife techniques are used here to calculate a con-fidence interval for each estimated parameter. Theseresults can be presented in scatter diagrams together withthe absolute values. The second method or Kernel analysisis an experimental density smoothing for the estimationof the distribution (for further details see Monchot &Léchelle 2002). Both methods are complementary (seebelow and Monchot et al., this volume).
Results
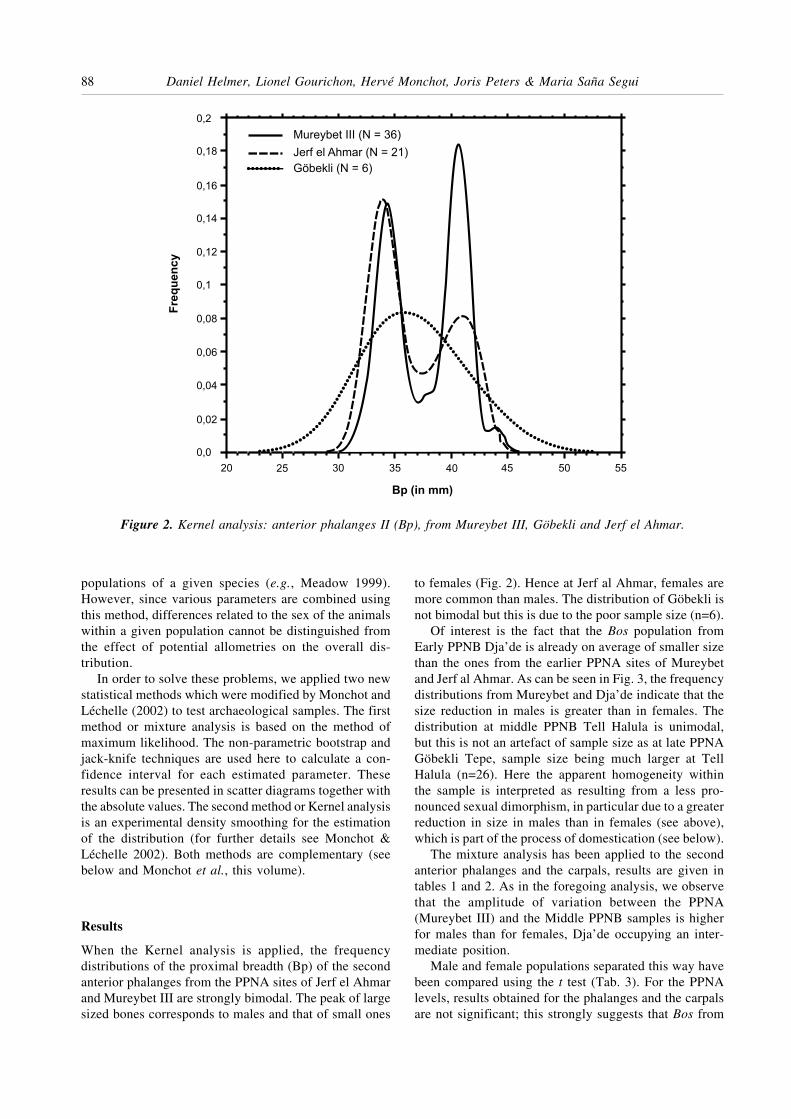
When the Kernel analysis is applied, the frequencydistributions of the proximal breadth (Bp) of the secondanterior phalanges from the PPNA sites of Jerf el Ahmarand Mureybet III are strongly bimodal. The peak of largesized bones corresponds to males and that of small ones
to females (Fig. 2). Hence at Jerf al Ahmar, females aremore common than males. The distribution of Göbekli isnot bimodal but this is due to the poor sample size (n=6).
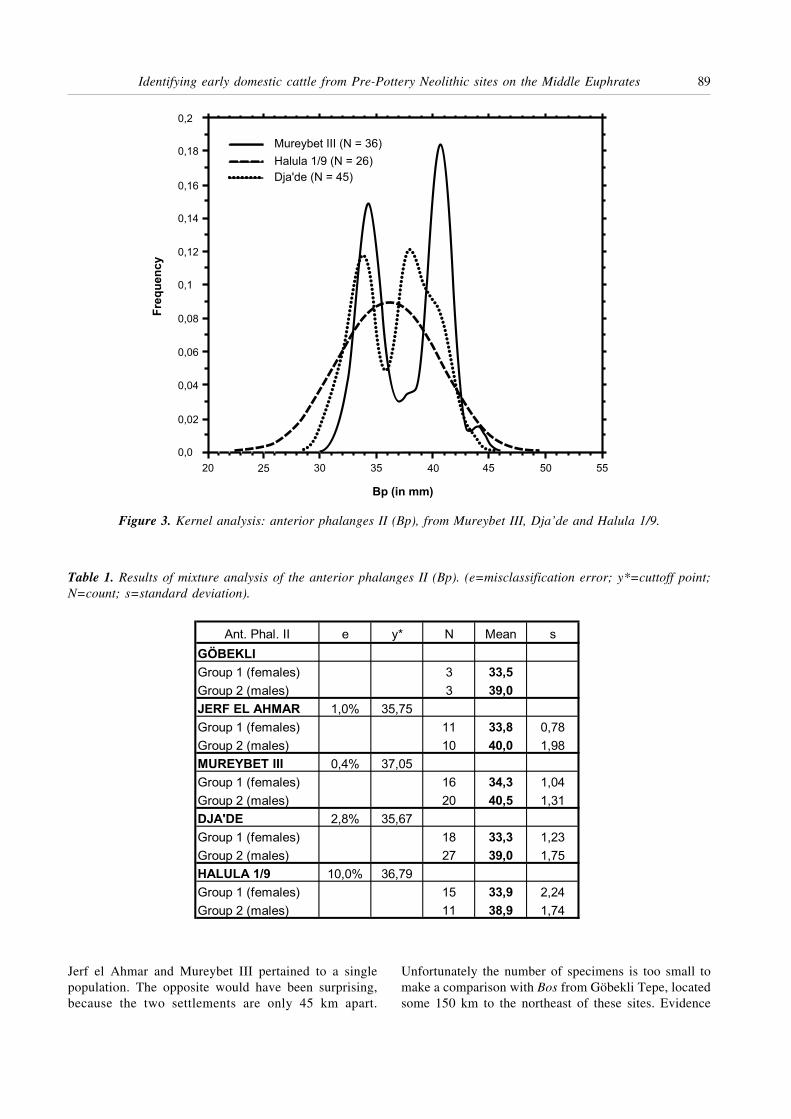
Of interest is the fact that the Bos population fromEarly PPNB Dja’de is already on average of smaller sizethan the ones from the earlier PPNA sites of Mureybetand Jerf al Ahmar. As can be seen in Fig. 3, the frequencydistributions from Mureybet and Dja’de indicate that thesize reduction in males is greater than in females. Thedistribution at middle PPNB Tell Halula is unimodal,but this is not an artefact of sample size as at late PPNAGöbekli Tepe, sample size being much larger at TellHalula (n=26). Here the apparent homogeneity withinthe sample is interpreted as resulting from a less pro-nounced sexual dimorphism, in particular due to a greaterreduction in size in males than in females (see above),which is part of the process of domestication (see below).
The mixture analysis has been applied to the secondanterior phalanges and the carpals, results are given intables 1 and 2. As in the foregoing analysis, we observethat the amplitude of variation between the PPNA(Mureybet III) and the Middle PPNB samples is higherfor males than for females, Dja’de occupying an inter-mediate position.
Male and female populations separated this way havebeen compared using the t test (Tab. 3). For the PPNAlevels, results obtained for the phalanges and the carpalsare not significant; this strongly suggests that Bos from
Figure 2. Kernel analysis: anterior phalanges II (Bp), from Mureybet III, Göbekli and Jerf el Ahmar.
0,2
0,18
0,16
0,14
0,12
0,1
0,08
0,06
0,04
0,02
0,0
20 25 30 35 40 45 50 55
Mureybet III (N = 36)
Jerf el Ahmar (N = 21)
Göbekli (N = 6)
Bp (in mm)
Fre
qu
en
cy
89Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates
Figure 3. Kernel analysis: anterior phalanges II (Bp), from Mureybet III, Dja’de and Halula 1/9.
Ant. Phal. II e y* N Mean s
GÖBEKLI
Group 1 (females) 3 33,5
Group 2 (males) 3 39,0
JERF EL AHMAR 1,0% 35,75
Group 1 (females) 11 33,8 0,78
Group 2 (males) 10 40,0 1,98
MUREYBET III 0,4% 37,05
Group 1 (females) 16 34,3 1,04
Group 2 (males) 20 40,5 1,31
DJA'DE 2,8% 35,67
Group 1 (females) 18 33,3 1,23
Group 2 (males) 27 39,0 1,75
HALULA 1/9 10,0% 36,79
Group 1 (females) 15 33,9 2,24
Group 2 (males) 11 38,9 1,74
Table 1. Results of mixture analysis of the anterior phalanges II (Bp). (e=misclassification error; y*=cuttoff point;N=count; s=standard deviation).
Jerf el Ahmar and Mureybet III pertained to a singlepopulation. The opposite would have been surprising,because the two settlements are only 45 km apart.
Unfortunately the number of specimens is too small tomake a comparison with Bos from Göbekli Tepe, locatedsome 150 km to the northeast of these sites. Evidence
0,2
0,18
0,16
0,14
0,12
0,1
0,08
0,06
0,04
0,02
0,0
20 25 30 35 40 45 50 55
Mureybet III (N = 36)
Halula 1/9 (N = 26)
Dja'de (N = 45)
Bp (in mm)
Fre
qu
en
cy
90 Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Peters & Maria Saña Segui
Table 2. Results of mixture analysis of the magnum (GB). (e=misclassification error; y*=cuttoff point; N=count;s=standard deviation).
Magnum e y* N Mean s
GÖBEKLI
Group 1 (females) 0
Group 2 (males) 0
JERF EL AHMAR
Group 1 (females) 5 39.7 1.61
Group 2 (males) 0
MUREYBET III 0.3% 46.52
Group 1 (females) 10 40.7 2.27
Group 2 (males) 10 49.5 1.07
DJA'DE 4.0% 43.62
Group 1 (females) 14 40.4 1.76
Group 2 (males) 5 46.3 2.08
HALULA 1/9 1.4% 43.48
Group 1 (females) 8 39.3 1.96
Group 2 (males) 7 45.3 0.77
Table 3. Means comparison: A: results of t test for the PPNA sites (9800 to 8800 BC cal.); B: results of t test forMureybet III (9800 to 8800 BC cal.), Dja’de (8800 to 8300 BC cal.) and Halula 1/9 (8300 to 7500 BC cal.).
t test Females Males
A Sites Jerf Göb Jerf Göb
Phal. II ant. (BP) Mb III 1.5 0.9
Jerf
Magnum (GB) Mb III 0.9
Jerf
B Sites Dja'de Hal 1/9 Dja'de Hal 1/9
Phal. II ant. (BP) Mb III 2.5 0.6 3.2 2.8
Dja'de 1.1 0.1
Magnum (GB) Mb III 0.4 1.6 4.0 4.6
Dja'de 1.6 0.3
that the animals from Mureybet III, Jerf el Ahmar andGöbekli indeed represent aurochs comes from thesimilarities in shape and size of the horn cores and fromthe fact that these animals were of the same staturecompared to their Early to Middle Holocene wild relatives
in Europe (cf. Grigson 1989). In contrast, the individualsfrom Dja’de and Halula are clearly smaller.
The sex ratio obtained from the combined samples ofastragalus, second anterior phalanges, centroquartal andmagnum (Tab. 4) is similar at Göbekli Tepe, Mureybet
91Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates
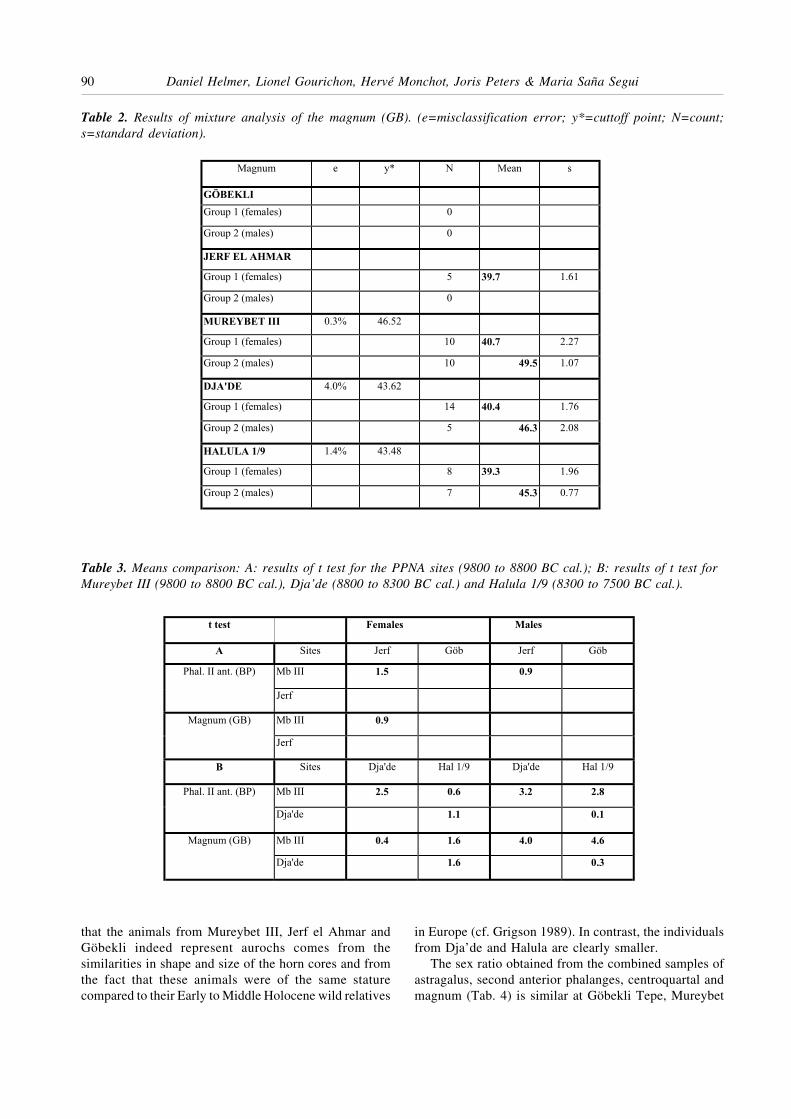
III, Dja’de and Tell Halula, with almost one female forevery male. The two later sites (Dja’de and Tell Halula),however, have a more variable sex ratio between thedifferent skeletal parts than the earlier sites (GöbekliTepe and Mureybet III), which are much more homo-genous. These variations could be interpreted as resultingfrom the initial steps of domestication; the number ofsites considered in this study, however, is too limited totest this assumption. Only at Jerf al Ahmar the sex ratiois in favour of female individuals (one bull for threecows). This high frequency probably indicates selectivehunting of females. This hypothesis is reinforced by thekill-off pattern deduced on the basis of tooth eruptionand abrasion (Fig. 4), where a predominance of calves ofless than two years and of adults surpassing four yearsconceivably correspond to the hunting of a herdcomprising essentially females with their young (familygroups). On the contrary, the kill-off pattern fromMureybet III is interpreted as a hunting pattern in-dependent of the animal’s sex.
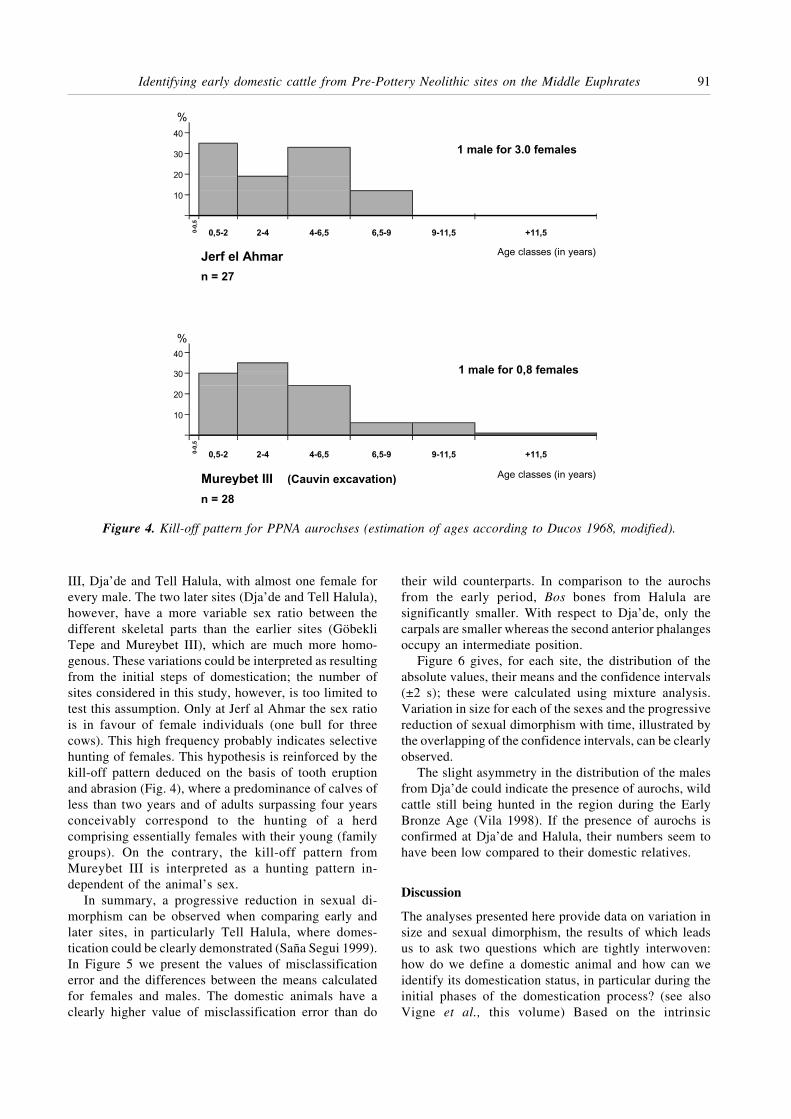
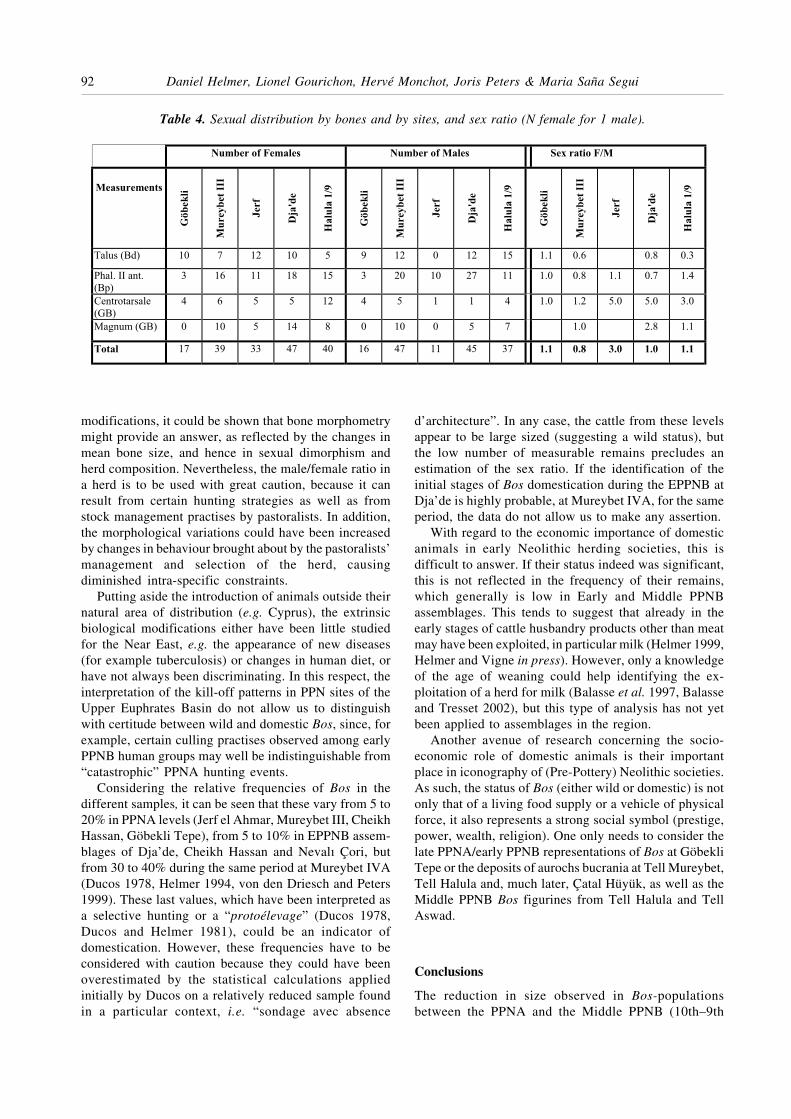
In summary, a progressive reduction in sexual di-morphism can be observed when comparing early andlater sites, in particularly Tell Halula, where domes-tication could be clearly demonstrated (Saña Segui 1999).In Figure 5 we present the values of misclassificationerror and the differences between the means calculatedfor females and males. The domestic animals have aclearly higher value of misclassification error than do
their wild counterparts. In comparison to the aurochsfrom the early period, Bos bones from Halula aresignificantly smaller. With respect to Dja’de, only thecarpals are smaller whereas the second anterior phalangesoccupy an intermediate position.
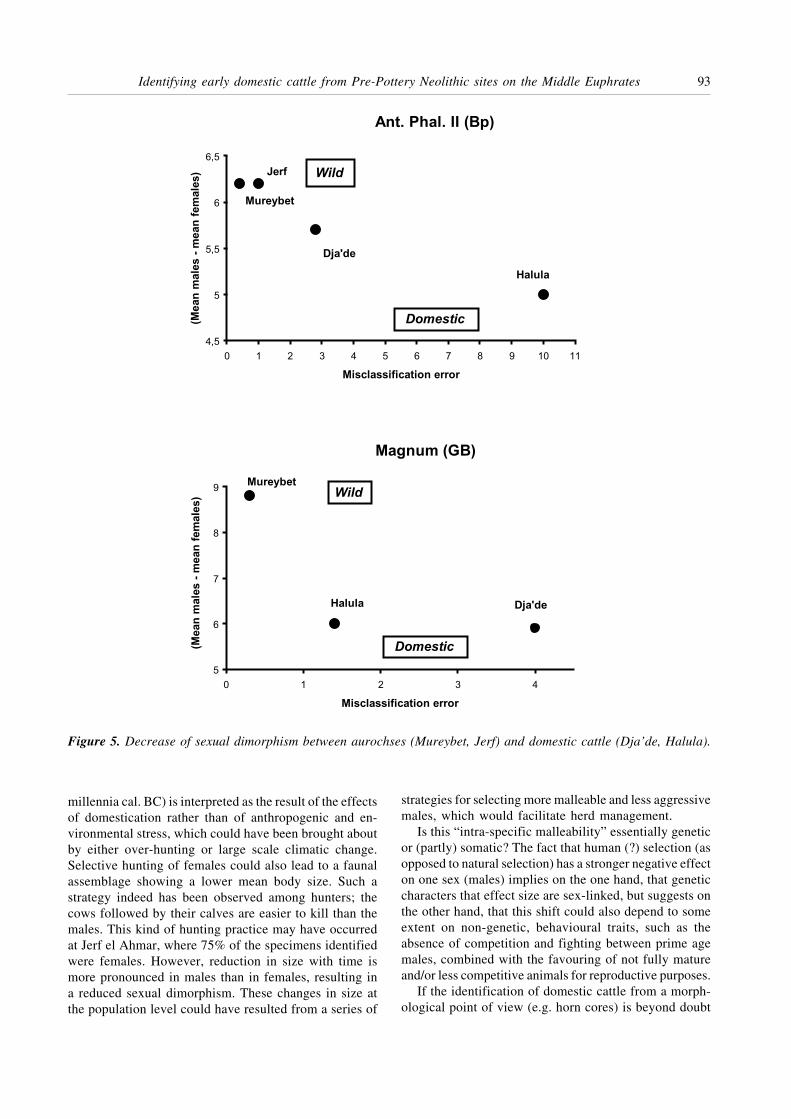
Figure 6 gives, for each site, the distribution of theabsolute values, their means and the confidence intervals(±2 s); these were calculated using mixture analysis.Variation in size for each of the sexes and the progressivereduction of sexual dimorphism with time, illustrated bythe overlapping of the confidence intervals, can be clearlyobserved.
The slight asymmetry in the distribution of the malesfrom Dja’de could indicate the presence of aurochs, wildcattle still being hunted in the region during the EarlyBronze Age (Vila 1998). If the presence of aurochs isconfirmed at Dja’de and Halula, their numbers seem tohave been low compared to their domestic relatives.
Discussion
The analyses presented here provide data on variation insize and sexual dimorphism, the results of which leadsus to ask two questions which are tightly interwoven:how do we define a domestic animal and how can weidentify its domestication status, in particular during theinitial phases of the domestication process? (see alsoVigne et al., this volume) Based on the intrinsic
Figure 4. Kill-off pattern for PPNA aurochses (estimation of ages according to Ducos 1968, modified).
0-0
,5
0,5-2 2-4 4-6,5 6,5-9 9-11,5 +11,5
Jerf el Ahmar
n = 27
0-0
,5
0,5-2 2-4 4-6,5 6,5-9 9-11,5 +11,5
Mureybet III (Cauvin excavation)
n = 28
20
10
40
30
40
Age classes (in years)
Age classes (in years)
%
30
10
%
20
1 male for 0,8 females
1 male for 3.0 females
92 Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Peters & Maria Saña Segui
modifications, it could be shown that bone morphometrymight provide an answer, as reflected by the changes inmean bone size, and hence in sexual dimorphism andherd composition. Nevertheless, the male/female ratio ina herd is to be used with great caution, because it canresult from certain hunting strategies as well as fromstock management practises by pastoralists. In addition,the morphological variations could have been increasedby changes in behaviour brought about by the pastoralists’management and selection of the herd, causingdiminished intra-specific constraints.
Putting aside the introduction of animals outside theirnatural area of distribution (e.g. Cyprus), the extrinsicbiological modifications either have been little studiedfor the Near East, e.g. the appearance of new diseases(for example tuberculosis) or changes in human diet, orhave not always been discriminating. In this respect, theinterpretation of the kill-off patterns in PPN sites of theUpper Euphrates Basin do not allow us to distinguishwith certitude between wild and domestic Bos, since, forexample, certain culling practises observed among earlyPPNB human groups may well be indistinguishable from“catastrophic” PPNA hunting events.
Considering the relative frequencies of Bos in thedifferent samples, it can be seen that these vary from 5 to20% in PPNA levels (Jerf el Ahmar, Mureybet III, CheikhHassan, Göbekli Tepe), from 5 to 10% in EPPNB assem-blages of Dja’de, Cheikh Hassan and Neval� Çori, butfrom 30 to 40% during the same period at Mureybet IVA(Ducos 1978, Helmer 1994, von den Driesch and Peters1999). These last values, which have been interpreted asa selective hunting or a “protoélevage” (Ducos 1978,Ducos and Helmer 1981), could be an indicator ofdomestication. However, these frequencies have to beconsidered with caution because they could have beenoverestimated by the statistical calculations appliedinitially by Ducos on a relatively reduced sample foundin a particular context, i.e. “sondage avec absence
d’architecture”. In any case, the cattle from these levelsappear to be large sized (suggesting a wild status), butthe low number of measurable remains precludes anestimation of the sex ratio. If the identification of theinitial stages of Bos domestication during the EPPNB atDja’de is highly probable, at Mureybet IVA, for the sameperiod, the data do not allow us to make any assertion.
With regard to the economic importance of domesticanimals in early Neolithic herding societies, this isdifficult to answer. If their status indeed was significant,this is not reflected in the frequency of their remains,which generally is low in Early and Middle PPNBassemblages. This tends to suggest that already in theearly stages of cattle husbandry products other than meatmay have been exploited, in particular milk (Helmer 1999,Helmer and Vigne in press). However, only a knowledgeof the age of weaning could help identifying the ex-ploitation of a herd for milk (Balasse et al. 1997, Balasseand Tresset 2002), but this type of analysis has not yetbeen applied to assemblages in the region.
Another avenue of research concerning the socio-economic role of domestic animals is their importantplace in iconography of (Pre-Pottery) Neolithic societies.As such, the status of Bos (either wild or domestic) is notonly that of a living food supply or a vehicle of physicalforce, it also represents a strong social symbol (prestige,power, wealth, religion). One only needs to consider thelate PPNA/early PPNB representations of Bos at GöbekliTepe or the deposits of aurochs bucrania at Tell Mureybet,Tell Halula and, much later, Çatal Hüyük, as well as theMiddle PPNB Bos figurines from Tell Halula and TellAswad.
Conclusions
The reduction in size observed in Bos-populationsbetween the PPNA and the Middle PPNB (10th–9th
Table 4. Sexual distribution by bones and by sites, and sex ratio (N female for 1 male).
Number of Females Number of Males Sex ratio F/M
Measurements G
öb
ek
li
Mu
rey
bet
III
Jer
f
Dja
'de
Ha
lula
1/9
Gö
bek
li
Mu
rey
bet
III
Jer
f
Dja
'de
Ha
lula
1/9
Gö
bek
li
Mu
rey
bet
III
Jer
f
Dja
'de
Ha
lula
1/9
Talus (Bd) 10 7 12 10 5 9 12 0 12 15 1.1 0.6 0.8 0.3
Phal. II ant.
(Bp)
3 16 11 18 15 3 20 10 27 11 1.0 0.8 1.1 0.7 1.4
Centrotarsale
(GB)
4 6 5 5 12 4 5 1 1 4 1.0 1.2 5.0 5.0 3.0
Magnum (GB) 0 10 5 14 8 0 10 0 5 7 1.0 2.8 1.1
Total 17 39 33 47 40 16 47 11 45 37 1.1 0.8 3.0 1.0 1.1
93Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates
millennia cal. BC) is interpreted as the result of the effectsof domestication rather than of anthropogenic and en-vironmental stress, which could have been brought aboutby either over-hunting or large scale climatic change.Selective hunting of females could also lead to a faunalassemblage showing a lower mean body size. Such astrategy indeed has been observed among hunters; thecows followed by their calves are easier to kill than themales. This kind of hunting practice may have occurredat Jerf el Ahmar, where 75% of the specimens identifiedwere females. However, reduction in size with time ismore pronounced in males than in females, resulting ina reduced sexual dimorphism. These changes in size atthe population level could have resulted from a series of
strategies for selecting more malleable and less aggressivemales, which would facilitate herd management.
Is this “intra-specific malleability” essentially geneticor (partly) somatic? The fact that human (?) selection (asopposed to natural selection) has a stronger negative effecton one sex (males) implies on the one hand, that geneticcharacters that effect size are sex-linked, but suggests onthe other hand, that this shift could also depend to someextent on non-genetic, behavioural traits, such as theabsence of competition and fighting between prime agemales, combined with the favouring of not fully matureand/or less competitive animals for reproductive purposes.
If the identification of domestic cattle from a morph-ological point of view (e.g. horn cores) is beyond doubt
Figure 5. Decrease of sexual dimorphism between aurochses (Mureybet, Jerf) and domestic cattle (Dja’de, Halula).
Ant. Phal. II (Bp)
4,5
5
5,5
6
6,5
0 1 2 3 4 5 6 7 8 9 10 11
Misclassification error
(Mean
male
s -
mean
fem
ale
s)
Dja'de
Halula
Jerf
Mureybet
Wild
Domestic
Magnum (GB)
5
6
7
8
9
0 1 2 3 4
Misclassification error
(Mean
male
s -
mean
fem
ale
s)
Mureybet
Halula Dja'de
Wild
Domestic
94 Daniel Helmer, Lionel Gourichon, Hervé Monchot, Joris Peters & Maria Saña Segui
at Middle PPNB Tell Halula (Saña Segui 1999), then,according to the results presented here, a domestic statusof Bos is also highly probable at Early PPNB Dja’de.Although this may appear early for cattle domestication– Dja’de being dated from 8800 to 8300 cal. BC –, it isless surprising when we know that, for the same period,sheep, pig and probably goat are already fullydomesticated at Early PPNB Neval� Çori, only onehundred kilometres north-west of Dja’de (Peters et al.1999; Peters et al., this vol.), and that cattle has beentransferred to Cyprus through sea no later than 8400–8200 cal. BC (Vigne et al. 2000).
AcknowledgementsWe wish to thank G. Willcox for his constructive remarksand for his help with the English. The research presentedin this article is supported in the framework of theEuropean Science Foundation EUROCORES programme“The Origin of Man, Language and Languages”.
Bibliography
Bottema, S. (1995) The Younger Dryas in the Eastern Mediterranean.Quaternary Science Reviews 14, 883–891.
Balasse, M., Bocherens, H, Tresset, A., Mariotti, A. and Vigne, J.-D (1997) Émergence de la production laitière au Néolithique ?
Contribution de l’analyse isotopique d’ossements de bovinsarchéologiques. Compte Rendus de l’Académie des Sciences,Paris, Sciences de la terre et des planètes 325, 1005–1010.
Balasse, M. and Tresset, A. (2002) Early weaning of Neolithicdomestic cattle (Bercy, France) revealed by intra-tooth variationin nitrogen isotope ratios. Journal of Archaeological Science29, 8, 853–860.
Cauvin, J. (2000) The Birth of the Gods and the Origins ofAgriculture. Cambridge, Cambridge University Press.
Cauvin, J., Cauvin, M.-C., Helmer, D. and Willcox, G. (1998)L’homme et son environnement au Levant Nord entre 30000 et7500 BP. Paléorient 23, 2, 51–69.
Driesch, A. von den, Peters, J. (1999) Vorläufiger Bericht über diearchäozoologischen Untersuchungen am Göbekli Tepe und amGürcütepe bei Urfa, Türkei. Istambuler Mitteilungen 49, 23–29.
Ducos, P. (1968) L’origine des animaux domestiques en Palestine.Bordeaux, Institut du Quaternaire.
Ducos, P. (1978) Tell-Mureybet (Syrie, IXe-VIIe millénaires). Etudearcheozoologique et précis d’écologie humaine. Lyon, CNRSEditions.
Ducos, P. and Helmer, D. (1981) Le point actuel sur l’apparition dela domestication dans le Levant. In Préhistoire du Levant(Colloques Internationaux du CNRS 598), 523–528. Paris,CNRS Editions.
Grigson, C. (1989) Size and sex: evidence for the domestication ofcattle in the Near East. In A. Milles, D. Williams and N. Gardner(eds) The Beginnings of Agriculture. British ArchaeologicalReports, International Series 496, 77–109.
Helmer, D. (1994) La domestication des animaux d’embouche dans
Figure 6. Distribution of measurements (Bp) of anterior phalanges II. Black line=confidence interval (±2 s), arrow=meanfor each sex.
Anterior phalanges II
25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45
MalesFemales
Göbekli
Halula 1/9
Dja'de
Mureybet III
Jerf el Ahmar
Bp (in mm)

95Identifying early domestic cattle from Pre-Pottery Neolithic sites on the Middle Euphrates
le Levant Nord (Syrie du Nord et Sinjar), du milieu du IXe
millénaire BP à la fin du VIIIe millénaire BP. Nouvelles donnéesd’après les fouilles récentes. Anthropozoologica 20, 41–54.
Helmer, D., Roitel, V., Saña, M. and Willcox, G. (1998)Interprétations climatiques des données archéozoologiques etarchéobotaniques en Syrie du nord de 16000 BP à 7000 BP, etles débuts de la domestication des plantes et des animaux. In M.Fortin and O. Aurenche (eds) Espace naturel, espace habité enSyrie du nord (10e-2e millénaires av. J.-C.). Québec, CanadianSociety for Mesopotamian Studies 33. Lyon, Travaux de laMaison de l’Orient 28, 9–33.
Helmer, D. (1999) Origine e sviluppo dell’allevamento nel VicinoOriente. In M. Arioti & B. Casciari (eds) Società pastorlaid’Africa e d’Asia. Brescia, Grafo edizioni, La Ricerca Folklorica40, 25–30.
Helmer, D. and Vigne, J.-D. (in press) Gestion des cheptels decaprinés au Néolithique dans le Midi de la France. In Actes du25e congrés de la Société préhistorique de France (Nanterre,Novembre 2000).
Meadow, R. (1999) The use of size index scaling for research onarchaeozoological collections from the Middle East. In C. Becker,H. Manhart, J. Peters and J. Schibler (eds) Historia animaliumex ossibus. Festschrift für Angela von den Driesch, 285–300.Rahden/Westf., Leidorf.
Monchot, H. and Léchelle, J. (2002) Statistical nonparametricmethods for the study of fossil populations. Paleobiology 28, 1,55–69.
Peters, J., Helmer, D., Driesch , A. von den and Saña Segui, M.(1999) Early animal husbandry in the Northern Levant. Palé-orient 25, 2, 27–47.
Saña Segui, M. (1999) Arqueología de la domesticacíon animal.La gestión de los recursos animales en Tell Halula (Valle delÉufrates – Siria) de 8.800 al 7.000 BP. Barcelone, Treballsd’Arqueologia del Pròxim Orient 1.
Vigne, J.-D., Carrère, I., Saliège, J.-F., Person, A., Bocherens, H.,Guilaine, J. and Briois, F. (2000) Predomestic cattle, sheep, goatand pig during the late 9th and the 8th millennium cal. BC onCyprus: preliminary results of Shillourokambos (Perkklisha,Limassol). In M. Mashkour, A.M. Choyke, H. Buitenhuis and F.Poplin (eds.) Archaeozoology of the Near East IV, Proc. 4thinternational Symposium Archaeozoology of Southwestern Asiaand adjacent areas (ASWA, Paris, 1998), 52–75. Groningen,Archaeological Research and Consultancy (Publicaties 32).
Vila, E. (1998) L’exploitation des animaux en Mésopotamie auxIVe et IIIe millénaires avant J.-C. (Monographie du CRA 21).Paris, CNRS Editions.
Daniel Helmer and Lionel GourichonMaison de l’Orient Méditerranéen–Jean PouillouxGREMMO7 rue RaullinF-69007 LyonFranceE-mail: [email protected]
Hervé MonchotLaboratoire d’AnthropologieUMR 6569Faculté de médecine NordBoulevard Pierre DramardF-13916 MarseilleFrance
Joris PetersInstitut für Palaeoanatomieund Geschichte der TiermedizinLudwig-Maximilians-Universität MünchenKaulbachstrasse 37D-80539 MünchenGermany
Maria Saña Segui,
Departament d’Anthropologia Social i prehistòriaUniversitat Autònoma de BarcelonaE-08193 Bellaterra (Barcelona)Spain




















