
1DETERMINACIÓN DEL EFECTO DEL COBRE ... - CORE
10
Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela 1 DETERMINACIÓN DEL EFECTO DEL COBRE SOBRE ALGUNAS FUNCIONES DEL SISTEMA COLINÉRGICO EN VARIAS ESTRUCTURAS DE CEREBRO DE RATA SPRAGUE- DAWLEY EN DESARROLLO * Elsa Ysmelia Gutiérrez – Reyes; * William José López Hernández; * Nelson Enrique Loureiro Dos Santos; ** Carmine Pascuzzo Lima; ** Jham Frank Papale Centofanti; ** José Moreno. PALABRAS CLAVE: Acetilcolina. Colinacetiltransferasa. Acetilcolinesterasa. RESUMEN La Acetilcolina (ACh) es un neurotransmisor sintetizado por la enzima Colinacetiltransferasa (ChAT) y degradado por la Acetilcolinesterasa (AChE). Nuestros resultados en trabajos previos, demostraron que el cobre incrementa el número de receptores colinérgicos muscarínicos en regiones específicas del cerebro de ratas acompañado también de manifestaciones conductuales de tipo colinérgicas, sin embargo, no se ha demostrado su participación en otros puntos de la vía de transmisión colinérgica. El objetivo de este estudio fue determinar el efecto del cobre sobre las actividades de las enzimas ChAT y de AChE en Corteza Cerebral Total (CT), el Cuerpo Estriado (CE) y en Hipocampo (H) de cerebro de rata en desarrollo. Se utilizaron ratas Sprague-Dawley recién destetadas tratadas con cobre a una dosis de 1 mg/kg/día durante 15, 30 y 60 días. Los resultados muestran que la concentración de cobre solo incrementó en CE al final de 15 días de tratamiento sin modificaciones a los 30 ni 60 días, análogamente, no se obtuvieron cambios significativos en las otras dos estructuras. Por otro lado, el cobre no produjo modificaciones significativas en la ChAT en ninguna de las regiones estudiadas. En cuanto a la actividad de la AChE, solo se observó un incremento en el H a los 15 días, que desapareció en las consecutivas edades. Ni en CE ni en CT, se observaron cambios en la AChE. Se plantea la necesidad de evaluar el efecto del cobre en edades extremas en estas mismas estructuras, ya que parece haber efectos de este en función a la duración del tratamiento. KEY WORDS: Acetylcholine. Choline Acetyltransferase. Acetylcholinesterase. ABSTRACT The Acetylcholine (ACh) is a neurotransmisor synthesized by the enzyme Choline acetyltransferase (ChAT) and degraded by the Acetylcholinesterase (AChE). Previous results of us, demonstrated that the copper increases the number of muscarinic acetylcholine receptors in specific regions of the rat brains, also, accompanied of behavioral manifestations of cholinergic type, nevertheless, there has not been demonstrated his participation in other points of the route of cholinergic neurotransmition. The aim of this study was determined the effect of the copper on the activities of the ChAT and of AChE enzymes in Total Cerebral Cortex (CT), the Corpus striatum (CE) and in Hippocampus (H) of developing rat brain. Sprague-Dawley rats were treated with copper to a dose of 1 mg/kg/day during 15, 30 and 60 days. The results show that copper increased in CE at 15th day of treatment without modifications neither to the 30th nor 60th days, analogous, significant changes did not obtain in other two structures. On the other hand, the copper did not produce significant modifications in ChAT in any of the studied regions. As for the AChE activity, only an increase was observed in the H to 15th day, which disappeared in the consecutive ages. Neither in CE nor in CT change was observed in the AChE activity. These results lead the need to evaluate the copper effect in extreme ages in the same structures, since there seem to be effects of this one in function of the duration of the treatment. * Unidad de Fisiología, Decanato de Medicina – UCLA ** Unidad de Bioquímica, Decanato de Medicina – UCLA Proyecto financiado por el CDCHT (012-ME-2002), UCLA. Autor de correspondencia: Nelson Enrique Loureiro Dos Santos: Fax 2591840, correo electrónico: [email protected] brought to you by CORE View metadata, citation and similar papers at core.ac.uk provided by Intellectual Repository Production UCLA
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of 1DETERMINACIÓN DEL EFECTO DEL COBRE ... - CORE

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
1DETERMINACIÓN DEL EFECTO DEL COBRE SOBRE ALGUNAS FUNCIONES DEL SISTEMA COLINÉRGICO EN VARIAS ESTRUCTURAS DE CEREBRO DE RATA SPRAGUE-
DAWLEY EN DESARROLLO * Elsa Ysmelia Gutiérrez – Reyes; * William José López Hernández; * Nelson Enrique Loureiro Dos Santos; ** Carmine Pascuzzo Lima; ** Jham Frank Papale Centofanti; ** José Moreno. PALABRAS CLAVE: Acetilcolina. Colinacetiltransferasa. Acetilcolinesterasa. RESUMEN La Acetilcolina (ACh) es un neurotransmisor sintetizado por la enzima Colinacetiltransferasa (ChAT) y degradado por la Acetilcolinesterasa (AChE). Nuestros resultados en trabajos previos, demostraron que el cobre incrementa el número de receptores colinérgicos muscarínicos en regiones específicas del cerebro de ratas acompañado también de manifestaciones conductuales de tipo colinérgicas, sin embargo, no se ha demostrado su participación en otros puntos de la vía de transmisión colinérgica. El objetivo de este estudio fue determinar el efecto del cobre sobre las actividades de las enzimas ChAT y de AChE en Corteza Cerebral Total (CT), el Cuerpo Estriado (CE) y en Hipocampo (H) de cerebro de rata en desarrollo. Se utilizaron ratas Sprague-Dawley recién destetadas tratadas con cobre a una dosis de 1 mg/kg/día durante 15, 30 y 60 días. Los resultados muestran que la concentración de cobre solo incrementó en CE al final de 15 días de tratamiento sin modificaciones a los 30 ni 60 días, análogamente, no se obtuvieron cambios significativos en las otras dos estructuras. Por otro lado, el cobre no produjo modificaciones significativas en la ChAT en ninguna de las regiones estudiadas. En cuanto a la actividad de la AChE, solo se observó un incremento en el H a los 15 días, que desapareció en las consecutivas edades. Ni en CE ni en CT, se observaron cambios en la AChE. Se plantea la necesidad de evaluar el efecto del cobre en edades extremas en estas mismas estructuras, ya que parece haber efectos de este en función a la duración del tratamiento. KEY WORDS: Acetylcholine. Choline Acetyltransferase. Acetylcholinesterase. ABSTRACT The Acetylcholine (ACh) is a neurotransmisor synthesized by the enzyme Choline acetyltransferase (ChAT) and degraded by the Acetylcholinesterase (AChE). Previous results of us, demonstrated that the copper increases the number of muscarinic acetylcholine receptors in specific regions of the rat brains, also, accompanied of behavioral manifestations of cholinergic type, nevertheless, there has not been demonstrated his participation in other points of the route of cholinergic neurotransmition. The aim of this study was determined the effect of the copper on the activities of the ChAT and of AChE enzymes in Total Cerebral Cortex (CT), the Corpus striatum (CE) and in Hippocampus (H) of developing rat brain. Sprague-Dawley rats were treated with copper to a dose of 1 mg/kg/day during 15, 30 and 60 days. The results show that copper increased in CE at 15th day of treatment without modifications neither to the 30th nor 60th days, analogous, significant changes did not obtain in other two structures. On the other hand, the copper did not produce significant modifications in ChAT in any of the studied regions. As for the AChE activity, only an increase was observed in the H to 15th day, which disappeared in the consecutive ages. Neither in CE nor in CT change was observed in the AChE activity. These results lead the need to evaluate the copper effect in extreme ages in the same structures, since there seem to be effects of this one in function of the duration of the treatment.
* Unidad de Fisiología, Decanato de Medicina – UCLA ** Unidad de Bioquímica, Decanato de Medicina – UCLA Proyecto financiado por el CDCHT (012-ME-2002), UCLA. Autor de correspondencia: Nelson Enrique Loureiro Dos Santos: Fax 2591840, correo electrónico: [email protected]
brought to you by COREView metadata, citation and similar papers at core.ac.uk
provided by Intellectual Repository Production UCLA

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
INTRODUCCIÓN La Acetilcolina (Ach) es un neurotransmisor que participa en la función del Sistema Nervioso Autónomo y la Transmisión Motora Somática (Bloom, 2001; Hoffman y Taylor, 2001). Este neurotransmisor juega un importante papel en el control de diversas funciones cerebrales neuropsíquicas tales como la memoria, el aprendizaje, la actividad locomotora, la atención, la regulación cardiovascular y las respuestas emocionales de modulación del estrés (Fibiger, 1991, Oda, 1999, Kandel y Siegelbaum, 2000; Bloom, 2001). La síntesis de ACh se lleva a cabo gracias a la acción de la enzima Colinacetiltransferasa (ChAT). Se ha demostrado que la ChAT puede ser regulada por diferentes factores que actúan a corto (minutos a horas) o a largo plazo (horas a días). Los mecanismos a corto plazo incluyen modificaciones del medio iónico de la neurona a nivel presináptico, fosforilaciones mediadas por cinasas y cambios en la distribución subcelular de la enzima (Rossier et al, 1977; Hersech y Peet, 1978, Luine et al., 1984; Loureiro, 2000). La regulación a largo plazo conlleva a cambios que pueden permanecer por tiempo indefinido en las neuronas, como son los cambios en la cantidad de enzima (Gnahn et al., 1983; Nonner et al., 2000; Loureiro, 2000; Auld et al., 2001). La acción de la ACh sobre los receptores cesa con su hidrólisis que es catalizada por la acetilcolinesterasa (AChE), enzima asociada con la membrana de los terminales colinérgicos (Lefkowitz et al, 1996, Bloom, 1996). Diversos estudios enfatizan la intervención de metales de transición en la regulación del sistema colinérgico (Díaz-Arrastia et al, 1985; Moreno et al, 1995; Díaz et al, 1998; Gutiérrez-Reyes et al, 2002). El cobre es uno de ellos. Este metal está ampliamente distribuido en diversos órganos y tejidos de mamíferos, incluyendo el sistema nervioso central, donde juega un importante papel bioquímico (Kosma et al., 1981; Deibel et al, 1996; Waggoner et al, 1999). Existen pocos trabajos que demuestran los efectos neurológicos de la administración crónica de cobre en animales de experimentación. Resultados obtenidos en nuestro laboratorio demostraron por primera vez que la administración intraperitoneal de cobre (1 mg/Kg peso corporal) produjo un
incremento del número de Receptores Colinérgicos Muscarínicos (Bmax) en homogeneizados de CT, CE e H de cerebro de ratas Sprague-Dawley, acompañado de cambios conductuales relacionados con el sistema colinérgico, confirmándose que el incremento de cobre en el Sistema Nervioso Central interviene en la regulación de la función colinérgica central (Gutiérrez-Reyes et al., 1992). No se ha demostrado la participación del cobre a nivel de otros puntos de la vía de transmisión colinérgica. Los efectos a nivel de receptores, sugieren que probablemente la supersensibilidad colinérgica producida por el cobre pudiera responder a efectos presinápticos o postsinápticos, que modifiquen la síntesis, degradación o liberación de ACh. Esta hipótesis llevó a plantear como objetivo determinar el efecto de la administración crónica de sulfato de cobre sobre la actividad de las enzimas de síntesis (ChAT) y degradación (AChE) en la CT, el CE y el H de cerebro de ratas Sprague-Dawley, con el propósito de ayudar a discernir los mecanismos por los cuales el cobre afecta algunas funciones de la transmisión sináptica colinérgica central. MATERIALES Y MÉTODOS Animales: Se utilizaron 72 ratas Sprague-Dawley, machos, de 21 días de edad provenientes del Bioterio Central, Decanato de Ciencias Veterinarias de la Universidad Centroccidental “Lisandro Alvarado”. Las ratas recibieron agua y comida ad libitum y se mantuvieron en cajas de acero inoxidable con camas de cáscara de arroz. Este grupo etario se escogió considerando que a esta edad la barrera hematoencefálica está inmadura, suponiendo una mayor transferencia de cobre al encéfalo. Para la determinación de la actividad de la ChAT y cobre tisular, 36 ratas fueron divididas aleatoriamente en dos (2) grupos de 18 animales cada uno, grupo control y grupo experimental respectivamente, a los cuales se les administró solución fisiológica y cobre (como sulfato de cobre) a la dosis de 1 mg/kg/día. El tratamiento se administró disuelto en solución salina isotónica (NaCl, 0,9%), en un volumen de 0,5 ml por cada 100 gramos de peso corporal. El tratamiento se administró diariamente por vía intraperitoneal. Se procedió de manera similar con otras 36 ratas destinadas para la determinación de la actividad de la AchE

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
A los 15, 30 y 60 días de estar recibiendo los tratamientos se sacrificaron 6 animales de cada grupo (12 por vez). Los animales fueron decapitados y sus cerebros fueron removidos rápidamente sobre una cápsula de Petri en hielo. Se disecó la CT, el CE y el H, según lo descrito por Glowniski e Iversen (1966). Las estructuras disecadas fueron divididas en fragmentos, unos destinados para la determinación de ChAT o AChE y otros para la determinación del cobre tisular. Los tejidos fueron almacenados a –70 ºC hasta su análisis. Determinación de cobre tisular: Para la determinación de cobre se usó agua pentadestilada y el material de vidrio utilizado fue sumergido durante 24 horas en ácido nítrico al 12%. Se siguió la técnica descrita por Christian (1969) y modificada por Eller (1977) utilizando un equipo de absorción atómica de flama. Ensayos enzimáticos: Los segmentos destinados para los ensayos enzimáticos fueron homogeneizados con un homogeneizador Thomas de vidrio con pistilo de teflón, tres sesiones de tres golpes cada una separadas entre si por descansos de 1 minuto. Para la determinación de la actividad de la AChE, se procedió de manera similar usando el segundo grupo de 36 ratas divididas en 2 subgrupos (experimental y control) y sacrificadas en los mismos tiempos (15, 30 y 60 días). Cuantificación de la actividad de la enzima Colinacetiltransferasa (ChAT): Para la determinación de esta actividad se siguió la técnica descrita por Fonum (1975), con algunas modificaciones descrita por Loureiro Dos Santos et al (2000). Cuantificación de la actividad de la enzima Acetilcolinesterasa (AChE): Para la determinación de esta actividad se siguió la técnica descrita por Ellman (1961) adaptándola para la lectura en microplacas usando un lector de ELISA. Determinación de Proteínas: Las proteínas se determinaron por el método descrito por Lowry (1951). Todos los reactivos utilizados en este trabajo fueron de grado analítico. Análisis de los resultados: Los datos generados fueron analizados utilizando una regresión lineal
seguida de un análisis de varianza (ANOVA) de dos vías, considerando que se tienen como variables el tratamiento administrado y el tiempo de administración (desde 15 días hasta 60 días), para obtener por separado los efectos del tratamiento y del tiempo, así como de la posible interacción entre ambos. Se aceptó como estadísticamente significativa la dependencia del tratamiento y/o del tiempo cuando, al analizar cada regresión, si la misma resultara con un valor de p < 0,05, para todos los análisis. Todos los análisis fueron realizados utilizando el paquete GraphPad Prism (GraphPad Prism Software, San Diego, Ca), versión 3.0. RESULTADOS Como se mencionó anteriormente, todos los datos se analizaron por medio de un ANOVA de dos vías, para obtener por separado los efectos del tratamiento y del tiempo, así como de la posible interacción entre ambos. En cada uno de los gráficos se muestra además un análisis de regresión lineal que resume la tendencia global de la variación de cada parámetro estudiado. NIVELES TISULARES DE COBRE Con el objetivo de observar si el cobre administrado se acumulaba en las regiones escogidas se determino su concentración en cada una de ellas. El curso temporal de la acumulación de cobre en el CE de rata (figura 1), se apreció que la concentración de este metal, se mantuvo sensiblemente igual en las tres edades. También se pudo observar que en el grupo tratado con sulfato de cobre los niveles temporales siempre fueron mayores en relación con el grupo control. En esta estructura cerebral solo influyó el tratamiento (p=0,0061), ya que el tiempo no tuvo influencia, es decir, la diferencia entre los controles y tratados se mantuvo a lo largo de la duración del estudio. Se puede observar que en el curso temporal de la acumulación de cobre en el H (figura 2), los niveles incrementaron inicialmente (día 15) y luego disminuyeron, manteniéndose similares hasta el día 60 de tratamiento. A pesar de que en el grupo tratado con sulfato de cobre los niveles temporales siempre fueron más altos en relación con el grupo control, este aumento no fue significativo en las ratas tratadas. El tiempo de tratamiento tampoco influyó significativamente en el patrón de acumulación de cobre en el H.

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
Los niveles tisulares de cobre en la CT de rata se muestran en la figura 3. En esta estructura llama la atención que hasta el día 60 de tratamiento los niveles de cobre permanecieron sin cambios importantes en el grupo experimental, en relación con el grupo control. Tampoco hubo influencia del tiempo de administración. ACTIVIDAD DE LA ENZIMA ChAT En las tres siguientes figuras se puede observar la actividad de ChAT en las regiones estudiadas durante 15, 30 y 60 días de tratamiento. En la figura 4, podemos apreciar que el tratamiento con sulfato de cobre, no produjo cambios significativos en la actividad de la ChAT en CE, sin embargo con el tiempo, la actividad decayó significativamente tanto en el grupo control como en el experimental (p=0,0243). Los niveles de ChAT en H de rata se muestran en figura 5, y se observó que son mayores en las ratas tratadas, sin embargo, cuando se analizaron todos los datos bajo una ANOVA de dos vías, se pudo apreciar que este incremento no fue significativo. Por otro lado el tiempo si influyó, encontrándose un aumento sostenido desde los 15 días hasta los 60 días, en las ratas control de forma paralela al de las ratas experimentales (p<0,0001). La actividad de ChAT en CT de rata no mostraron ningún aumento significativo con el tratamiento de sulfato de cobre (figura 6), no obstante, el tiempo si influyó, puesto que los valores disminuyeron hasta el día 30, y luego incrementaron en ambos grupos hasta el día 60 (ANOVA de dos vías, p= 0,0001). ACTIVIDAD DE LA ENZIMA ACETILCOLINESTERASA (AChE) Los niveles de AChE en el CE de ratas (figura 7), analizados bajo una ANOVA de dos vías, no mostraron cambios significativos en ninguno de los períodos estudiados, así como tampoco hubo modificaciones de la actividad a lo largo de todo el estudio. Por otro lado el sulfato de cobre produjo un incremento significativo (p=0,0157) de la actividad de la AChE en el H (figura 8). El tiempo también influyó, aumentando significativamente la actividad en ambos sin embargo, mucho más pronunciado en el grupo tratado. En la CT no se obtuvieron cambios significativos respecto a la actividad enzimática (figura 9). El tiempo resultó ser un factor significativamente influyente en la
actividad de AChE, siendo el efecto similar en ambos grupos (p = 0,008). Figura 1.- Curso temporal de los niveles de cobre en el CE de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo (una determinación por animal). Los niveles de cobre aumentaron significativamente (regresión lineal, seguido de ANOVA de dos vías, p=0,0061) en el grupo tratado con sulfato de cobre. Aunque la diferencia entre los grupos tratado y control tendió a decrecer para el día 60, el tiempo no influyó significativamente en la acumulación temporal de cobre en el CE, es decir, la diferencia entre los controles y tratados se mantuvo a lo largo de la duración del estudio. Figura 2.- Curso temporal de los niveles de cobre en H de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo (una determinación por animal). Ni el tratamiento ni el tiempo influyeron significativamente en la acumulación temporal de cobre en el H.
Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
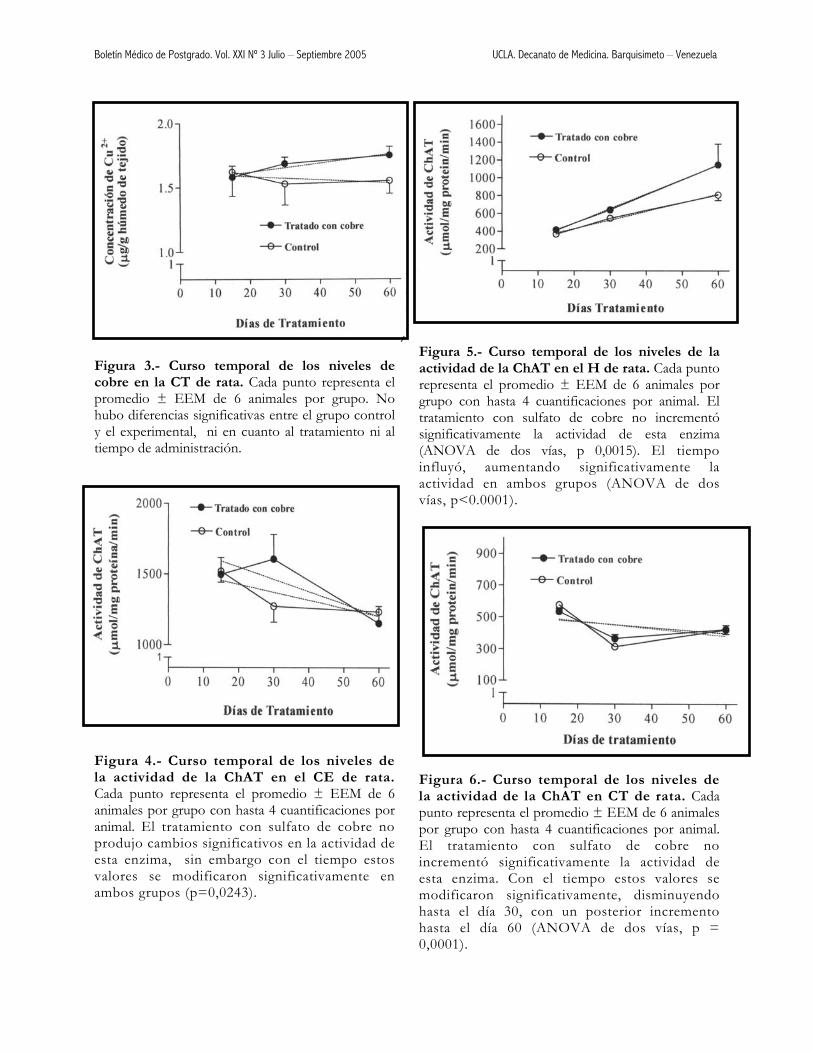
Figura 3.- Curso temporal de los niveles de cobre en la CT de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo. No hubo diferencias significativas entre el grupo control y el experimental, ni en cuanto al tratamiento ni al tiempo de administración. Figura 4.- Curso temporal de los niveles de la actividad de la ChAT en el CE de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo con hasta 4 cuantificaciones por animal. El tratamiento con sulfato de cobre no produjo cambios significativos en la actividad de esta enzima, sin embargo con el tiempo estos valores se modificaron significativamente en ambos grupos (p=0,0243).
1 2
3 4 5 6 7
Figura 5.- Curso temporal de los niveles de la actividad de la ChAT en el H de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo con hasta 4 cuantificaciones por animal. El tratamiento con sulfato de cobre no incrementó significativamente la actividad de esta enzima (ANOVA de dos vías, p 0,0015). El tiempo influyó, aumentando significativamente la actividad en ambos grupos (ANOVA de dos vías, p<0.0001). Figura 6.- Curso temporal de los niveles de la actividad de la ChAT en CT de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo con hasta 4 cuantificaciones por animal. El tratamiento con sulfato de cobre no incrementó significativamente la actividad de esta enzima. Con el tiempo estos valores se modificaron significativamente, disminuyendo hasta el día 30, con un posterior incremento hasta el día 60 (ANOVA de dos vías, p = 0,0001).
Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
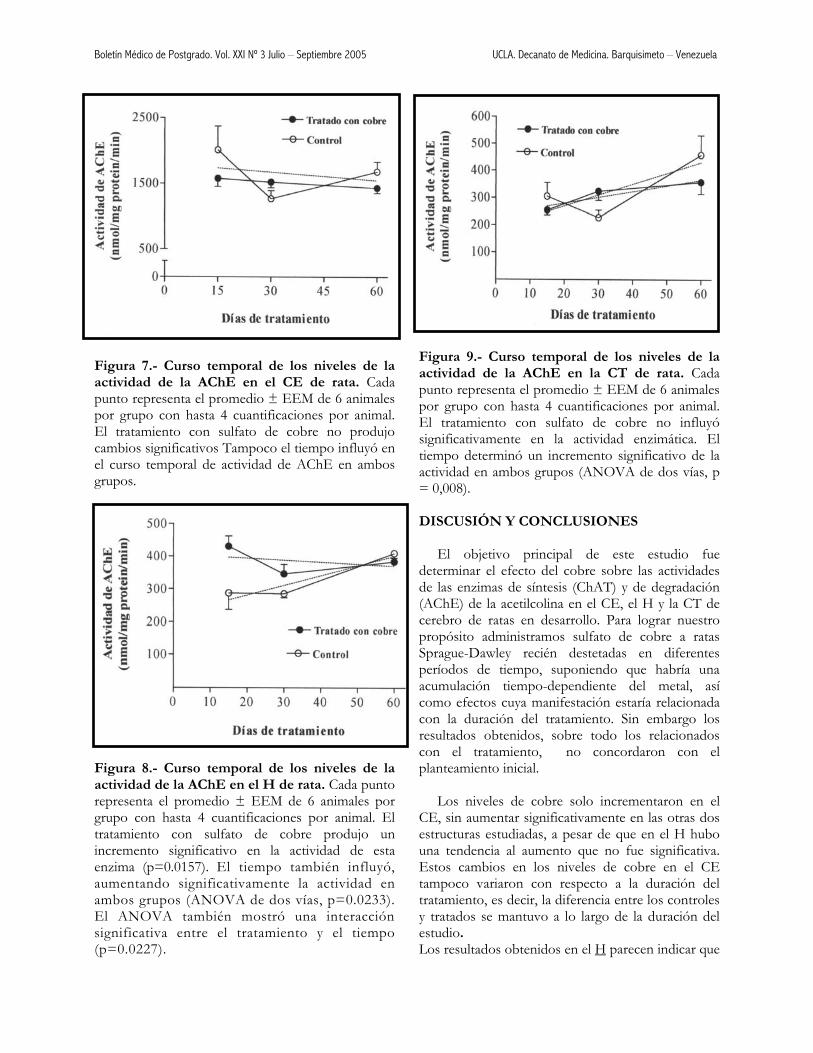
Figura 7.- Curso temporal de los niveles de la actividad de la AChE en el CE de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo con hasta 4 cuantificaciones por animal. El tratamiento con sulfato de cobre no produjo cambios significativos Tampoco el tiempo influyó en el curso temporal de actividad de AChE en ambos grupos.
Figura 8.- Curso temporal de los niveles de la actividad de la AChE en el H de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo con hasta 4 cuantificaciones por animal. El tratamiento con sulfato de cobre produjo un incremento significativo en la actividad de esta enzima (p=0.0157). El tiempo también influyó, aumentando significativamente la actividad en ambos grupos (ANOVA de dos vías, p=0.0233). El ANOVA también mostró una interacción significativa entre el tratamiento y el tiempo (p=0.0227).
Figura 9.- Curso temporal de los niveles de la actividad de la AChE en la CT de rata. Cada punto representa el promedio ± EEM de 6 animales por grupo con hasta 4 cuantificaciones por animal. El tratamiento con sulfato de cobre no influyó significativamente en la actividad enzimática. El tiempo determinó un incremento significativo de la actividad en ambos grupos (ANOVA de dos vías, p = 0,008). DISCUSIÓN Y CONCLUSIONES El objetivo principal de este estudio fue determinar el efecto del cobre sobre las actividades de las enzimas de síntesis (ChAT) y de degradación (AChE) de la acetilcolina en el CE, el H y la CT de cerebro de ratas en desarrollo. Para lograr nuestro propósito administramos sulfato de cobre a ratas Sprague-Dawley recién destetadas en diferentes períodos de tiempo, suponiendo que habría una acumulación tiempo-dependiente del metal, así como efectos cuya manifestación estaría relacionada con la duración del tratamiento. Sin embargo los resultados obtenidos, sobre todo los relacionados con el tratamiento, no concordaron con el planteamiento inicial. Los niveles de cobre solo incrementaron en el CE, sin aumentar significativamente en las otras dos estructuras estudiadas, a pesar de que en el H hubo una tendencia al aumento que no fue significativa. Estos cambios en los niveles de cobre en el CE tampoco variaron con respecto a la duración del tratamiento, es decir, la diferencia entre los controles y tratados se mantuvo a lo largo de la duración del estudio. Los resultados obtenidos en el H parecen indicar que

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
pudo haber un fenómeno de tamponamiento de dichos niveles, puesto que luego de un incremento inicial, la concentración de cobre en el grupo tratado tendió a hacerse semejante a la del grupo control a partir del día 30. En la CT este tamponamiento quizás se establece desde una edad postnatal temprana. Estos resultados parecen indicar que tanto en el H como CT existe un control más estricto que en CE, en cuanto al ingreso y/o acumulación de cobre, lo que permitiría una correcta homeostasis de este metal desde una edad temprana. Nuestros resultados sugieren una regulación de los niveles de cobre que parece depender de cada estructura involucrada, puesto que la concentración de cobre aumentó precozmente en el CE y en el H (que tuvo un patrón bastante similar al CE, aunque no fuera significativo este aumento), mientras que en la CT los niveles se mantuvieron sin cambios a los largo del tratamiento. Estos resultados sugieren que la homeostasis del cobre se inicia a temprana edad, a diferencia del zinc, otro metal esencial, cuya homeostasis se establece en una fase postnatal tardía (Gutiérrez – Reyes, 2004, trabajo de ascenso, resultados a publicar), lo que podría depender de la acción de una proteína particular, que es la metalotioneína, proteína citosólica que participa en la detoxificación de metales pesados y radicales libres y cuya síntesis es inducida tanto por el cobre como por el zinc, asegurando que la concentración libre y potencialmente tóxica de este oligoelemento sea mantenida baja similar a lo que ocurre con el zinc. La inducción de esta proteína en estructuras encefálicas sirve para quelar al cobre, no solo el presente de manera endógena, sino también al que ingrese exógenamente al encéfalo (Hamer, 1986). Probablemente los mecanismos de inducción de esta proteína por el cobre se establecen más tardíamente en el CE, en comparación con el H y la CT, que bien podría asociarse a mecanismos regulatorios de ingreso de oligoelementos diferenciados según, la estructura involucrada. Vale la pena resaltar que las dos estructuras que parecen regular mejor el ingreso de cobre, son precisamente las que presentan un menor poder de regulación de los niveles de zinc con respecto al CE (Gutiérrez – Reyes, 2004, trabajo de ascenso, resultados a publicar).
Por otro lado, con excepción del H, el tratamiento con cobre no se asoció a cambios notables de la actividad colinérgica, sin embargo, la duración del tratamiento expresada como la variable tiempo si influyó en el curso temporal en las tres estructuras estudiadas, lo que parece indicar que la edad de la rata puede influir en el curso temporal de la actividad de la ChAT. En CE, la administración de cobre no se asoció a cambios importantes en la actividad de la ChAT, observándose apenas un ligero incremento a los 30 días, que tendió a semejarse al grupo control al final del tratamiento, a pesar de que fue la única estructura donde el metal se acumuló; sin embargo, con el tiempo se observó una disminución en la actividad de esta enzima tanto en grupos tratados como en el control. En el H, la tendencia con el tiempo fue a incrementar la actividad, mientras que en la CT no hubo un comportamiento linear sino más bien oscilante. Es interesante destacar que estos resultados coincidieron con los niveles tisulares de cobre, excepto en el caso del CE, que a pesar de que acumuló el metal no se modificó la actividad de ChAT, en cambio, ni en el H ni en la CT hubo acumulación de cobre. Estos resultados fueron inesperados, considerando el conocimiento de que la transmisión colinérgica puede ser modificada por el cobre a nivel de receptor (Moreno et al, 1995; Díaz et al, 1998; Gutiérrez – Reyes et al, 2002). A diferencia del cobre, el zinc si incrementa la actividad de la ChAT (Gutiérrez – Reyes, 2004, trabajo de ascenso, resultados a publicar). Probablemente se deba a diferencias en la afinidad de ambos metales por residuos de aminoácidos importantes para su actividad. En resumen, el tratamiento con sulfato de cobre no produjo cambios significativos de la ChAT en ninguna de las estructuras estudiadas, sin embargo y a pesar de que el patrón de cambios fue diferente desde el punto de vista cuantitativo en cada estructura, el tiempo si influyó de manera significativa, lo que parece indicar que la edad de la rata es determinante en estos resultados. Cabe señalar que los niveles de ChAT encontrados en esta investigación se encuentran en el mismo orden de magnitud que los reportados por otros investigadores, tanto para corteza total como para H (Issa et al., 1996; Luine et al, 1984).

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
En cuanto a los niveles de la actividad de la AChE, se pudo observar que a diferencia del H, el CE y la CT fueron poco sensibles a la administración del cobre. Llama la atención que solo en el H la actividad de esta enzima incrementó significativamente, a pesar de que en esta estructura la acumulación de cobre no fue significativa. Sin embargo, es de hacer notar que esta fue la única estructura en la cual también hubo una interacción significativa entre el tratamiento y el tiempo, lo que indica que la magnitud del efecto del cobre varía con el tiempo. Es posible que pequeños incrementos en la acumulación del metal, aun no significativos, sumados a la influencia del tiempo, pudo determinar un sinergismo en la actividad de esta enzima. Por otro lado, en la CT, que a pesar de que el tiempo favoreció la actividad de la enzima, la acumulación de cobre tuvo un patrón bastante similar al control, es decir, no hubo ni siquiera una tendencia al aumento en los niveles del metal. También llama la atención que en el CE, ni el tratamiento ni el tiempo influyeron en la actividad de AChE. Similar a lo que ocurrió con ChAT, estos resultados también fueron sorprendentes, considerando que la transmisión colinérgica puede ser modificada por el cobre a nivel de receptor (Moreno et al, 1995; Díaz et al, 1998; Gutiérrez – Reyes et al, 2002). Pattison y Bernhard (1978) en estudios realizados en AChE proveniente de la placa terminal de Torpedo postularon que la enzima pudiera tener dos estados conformacionales únicos, uno reactivo y otro no reactivo. Según estos investigadores, algunos cationes como el calcio y el magnesio se unen y estabilizan el estado conformacional activo. Quizás en el H, este estado conformacional activo se estabiliza de manera sustancial en presencia de pequeñas cantidades de cobre y con el tiempo, hipótesis que pudiera explicar el incremento de la actividad de la AChE demostrada en este trabajo en el H. Cabe resaltar que a diferencia de la enzima ChAT, la AChE no cuenta con blancos aparentes para la acción del cobre, es decir, residuos de aminoácidos que pudieran interaccionar con el cobre. En relación a la enzima AChE, el tratamiento con sulfato de cobre solo produjo un incremento significativo de la actividad de la AChE en el H y hubo, una interacción positiva con el tiempo, lo que parece indicar que la edad de la rata es determinante en estos resultados. Ni en el CE ni en la Corteza, el
tratamiento con Sulfato de Cobre produjo cambios en la actividad de AChE, pero similar al HH, el tiempo influyó en el curso temporal de la actividad de esta enzima. Lo importante de los resultados obtenidos en este trabajo es que se demuestra por primera vez que el cobre, a la dosis de 1 mg/kg/día, en nuestras condiciones experimentales, tiende a incrementar solo la actividad de la AChE en el H, estructura no precisamente caracterizada por tener una alta inervación colinérgica y donde tampoco hubo acumulación del metal. La edad de la rata expresada en el tiempo o duración del tratamiento, si fue importante en el perfil de la actividad colinérgica en todas las estructuras estudiadas. En conclusión, la administración crónica de cobre solo se asocia a cambios en la actividad de AChE en el H y los cambios más relevantes en la función colinérgica, dependiendo particularmente de la estructura involucrada, se ven influenciados dependiendo del grado de madurez del animal, por lo que deberán caracterizarse estos efectos en animales considerados recién nacidos y en edad senil. REFERENCIAS BIBLIOGRÁFICAS 1. BLOOM FE. 2001. Neurotransmission and the
Central Nervous System. In Goodman and Gilman’s. The Pharmacological Basis of Therapeutics. 10th Edition. Hardman JG, Limbird LE Editors. International Edition. Mc Graw Hill Companies, Inc, New York. Chapter 12: 293 – 320.
2. HOFFMAN BB AND TAYLOR P. 2001.
Neurotransmission. In Goodman and Gilman’s. The Pharmacological Basis of Therapeutics. 10th Edition. Hardman JG, Limbird LE Editors. International Edition. Mc Graw Hill Companies, Inc. New York. Chapter 6:. 115 – 53.
3. FIBIGER, H.C., 1991. Cholinergic mechanism
in learning, memory and dementia: a rewiew of recent evidence. Trends Neurosci. 14: 220 – 223.
4. ODA YOSHIO. 1999. Cholineacetystransferase:
The structure, distribution and pathologicac changes in the central nervous system. Pathol Int. 49: 921 – 937.

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
5. KANDEL ER AND SIEGELBAUM SA. 2000. Overview of synaptic transmission. In Principles of Neural Science. 4th Edition. Kandel ER, Schwartz JH, Jessell TM Editors. Mc Graw Hill, New York. Chapter 10: 175 – 86.
6. BLOMM F. E. 1996. Neurotransmission and
the Central Nervous System. in Goodman and Gilman`s. The Pharmacological Basis of Therapeutics. Ninth Ed. Hardman and Limbird Editors. International Edition. Mc Graw Hill Companies, Inc. , New York. Chapter 12: 267 – 293.
7. ROSSIER J. et al. 1977. The effect of Cl- on
cholineacetyltransferase kinetic parameters and proposed role of Cl- in the regulation of acetylcholine synthesis. J Neorochem. 29: 1007 – 1012.
8. HERSH L.B. AND PEET M. 1978. Effects of
salts on the physical and kinetics propierties of human placental choline acetyltransferase. J Neurochem. 30: 1087 – 1093.
9. LUINE V.N. et al. 1984. Activation of choline
acetyltransferase by vasoactive intestinal peptide. J. Neurochem. 42: 1131 – 1134.
10. LOUREIRO DOS SANTOS, NELSON E.
2000. Regulaçao da expressao e da atividade funcional de marcadores colinérgicos no sistema nervoso central. Possiveis alvos de açao de peptidos amiloidogénicos. Tesis de Doutor Em Ciéncias. Universidade Federal do Rio de Janeiro. Instituto de Biofísica Carlos Chagas Filho.
11. GNAHN H. et al. 1983. H. NGF-mediated
increase in choline-acetyltransferase (ChAT) in the neonatal forebrain: evidence for a physiological role of NGF in the brain? Dev. Brain Res. 9:45 – 52.
12. NONNER D. et al. 2000. Brief exposure to
neurotrophins produces a calcium-dependent increase in choline acetyltransferase activity in cultured rat septal neurons. J Neurochem. 74 (3): 988 – 999.
13. AULD, D.S. et al. 2001. Neurotrophins
differentially enhance acetylcholine release.
Acetylcholine content and choline acetyltransferase activity in basal forebrain neurons. J Neurochem. 77: 253 – 262.
14. LEFKOWITZ R.J. et al. 1996.
Neurotransmission. In Goodman and Gilman`s. The Pharmacological Basis of Therapeutics. Ninth Ed. Hardman and Limbird Editors. International Edition. Mc Graw Hill Companies, Inc. New York. Chapter 6:., 105 – 139.
15. DÍAZ-ARRASTIA R. et al. 1985. Endogenous
inhibitor of ligand binding to the muscarinics acetylcholine receptor. J. Neurochem. 44 (2): 622 – 628.
16. DÍAZ LE. Et al. 1998. Comparación del Efecto
de Cobre y Zinc sobre la Unión de [3H] - QNB al Receptor Muscarínico Humano. Boletín Médico de Postgrado 14 (4): 207 – 215.
17. MORENO-YANES, JA. Et al.. 1995. Efecto
regulador de cobre sobre la función colinérgica muscarínica del cerebro humano. Arch Venez Farmacol Ter. 14 (1): 5 – 17.
18. GUTIERREZ - REYES E. et al. 2002.
Supersensitivity of the cholinergic muscarinic system in the rat’s brain is induced by high concentrations of Cu2+. Investigación Clínica 43 (2): 107 – 117.
19. KOSMA M. et al. 1981. Histochemical detection
of zinc and cooper in various neurons of the central nervous system. Acta Histochem. 69: 12 – 7.
20. DEIBEL MA et al. 1996. Cooper, iron and zinc
imbalances in severely degenerated brain regios in Alzheimer’disease: possible relation to oxidative stress. J Neurol Sci. 143: 137 – 42
21. WAGGONER DJ. Et al. 1999. The role of
cooper in neurodegenerative disease. Neurobiol Dis. 6: 221 – 230.
22. GLOWINSKI J. AND IVERSEN L. 1966.
Regional studies of catecholamines in the rat brain. Disposition of Norepynephryne, Dopamine and Dopa in various regions of the brain. J. Neuchem. 13: 665 – 669.

Boletín Médico de Postgrado. Vol. XXI Nº 3 Julio – Septiembre 2005 UCLA. Decanato de Medicina. Barquisimeto – Venezuela
23. CHRISTIAN G.D., 1969. Medical Trace metals and absortion spectroscopy. Anal Chem: 41: 24 – 40A..
24. ELLER P. AND HAARTZ J. 1977. An study of
methods for the determinatios of lead and cadmium. Am Ind Hyg. Assoc. 38 (3): 116 – 124.
25. LOWRY O. H. et al. 1951. Protein
measurement with the folin pheno reagent. J. Biol. Chem. 193: 265 – 275
26. FONNUM, F. 1975. A rapid radiochemical
method for the determination of choline acetyltransferase. J Neurochem. 24: 407 – 409.
27. ELLMAN G.L. et al. 1961. A new and rapid
colorimetric determination of cholinesterase activity. bichem pharmacol. 7: 88 – 95.
28. GUTIÉRREZ - REYES E, 2004. Efecto del
zinc sobre algunas funciones del sistema nervioso colinérgico en CE, H y CT de rata en desarrollo. Trabajo de ascenso a la categoría Titular, UCLA.
29. HAMER, D. H. 1986. Metallothionein. Ann. Rev. Biochem. 55: 913 – 951.
30. ISSA A.M. et al. 1996. Effects of phosphatase
inhibitors calyculin A and Okadaic acid on acetylcholine synthesis and content of rat hippocampal formation. J Neurochem. 66: 1924 – 1932.
31. PATTISON S AND BERNHARD S. 1978.
Functional consequences of ligand – dependent conformational changes in trypsin-solubilized and membrane particle constrined – acetylcholinesterase. Proc Natl Acad Sci 75 (8): 3613 – 3617.
Agradecimientos Al CDCHT de la Universidad Centroccidental Lisandro Alvarado por haber financiado este proyecto bajo el código 012-ME-2002. Al Sr. Elis Mosquera, personal del laboratorio de Fisiología Celular de la Unidad de Fisiología del Decanato de Medicina, por el apoyo técnico brindado.




















