
1.0 introduction - University of Lagos
172
1 1.0 INTRODUCTION Nigeria is home to the largest population of sickle cell anemia patients, estimated to be in excess of 4 million patients with more than 150 000 children born annually with the disease (World Health Organisation, 2006). The condition is also incident in the African diaspora population in North America and Europe. The SCA patient‟s quality of life and productivity are significantly reduced; the condition causes episodes of pain that can last for hours or days at a time (Steinberg, 2003; Todd et al., 2006; WHO, 2006; Hankins, 2009). There are few effective therapies for SCA, a chronic blood disorder which interferes with the body‟s oxygen transfer process and can lead to the formation of small blood clots, which over time deprive organs and tissues of oxygen. Despite decades of research, only one drug, hydroxyurea, approved by the American Food and Drug Agency, is available for the treatment of sickle cell anaemia (Hankins and Aygun, 2009; Brandow et al., 2010). This chemotherapeutic agent stimulates healthy production of fetal haemoglobin to counter the sickling process (Steinberg, 2003). Although long-term clinical studies have shown the drug to improve survival, while concerns of genotoxic and carcinogenic effects are yet to be ascertained (Lal and Ames, 2011). Developing novel drugs from traditional medicinal plants can serve as a means to improve the treatment of this disease. Research on phytomedicine for the management of SCA has led to the development of a herbal based drug called Niprisan which has been patented by the National Institute for Pharmaceutical Research and Development (NIPRD), Abuja, Nigeria. This development indicates that more of such herbal based drugs could be produced consequent upon scientific investigations on medicinal plants that are used in traditional African medicine.
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of 1.0 introduction - University of Lagos

1
1.0 INTRODUCTION
Nigeria is home to the largest population of sickle cell anemia patients, estimated to be in
excess of 4 million patients with more than 150 000 children born annually with the disease
(World Health Organisation, 2006). The condition is also incident in the African diaspora
population in North America and Europe. The SCA patient‟s quality of life and productivity
are significantly reduced; the condition causes episodes of pain that can last for hours or days
at a time (Steinberg, 2003; Todd et al., 2006; WHO, 2006; Hankins, 2009). There are few
effective therapies for SCA, a chronic blood disorder which interferes with the body‟s oxygen
transfer process and can lead to the formation of small blood clots, which over time deprive
organs and tissues of oxygen.
Despite decades of research, only one drug, hydroxyurea, approved by the American Food
and Drug Agency, is available for the treatment of sickle cell anaemia (Hankins and Aygun,
2009; Brandow et al., 2010). This chemotherapeutic agent stimulates healthy production of
fetal haemoglobin to counter the sickling process (Steinberg, 2003). Although long-term
clinical studies have shown the drug to improve survival, while concerns of genotoxic and
carcinogenic effects are yet to be ascertained (Lal and Ames, 2011). Developing novel drugs
from traditional medicinal plants can serve as a means to improve the treatment of this
disease.
Research on phytomedicine for the management of SCA has led to the development of a
herbal based drug called Niprisan which has been patented by the National Institute for
Pharmaceutical Research and Development (NIPRD), Abuja, Nigeria. This development
indicates that more of such herbal based drugs could be produced consequent upon scientific
investigations on medicinal plants that are used in traditional African medicine.

2
1-1 BACKGROUND OF STUDY
Sickle cell anemia is inherited in an autosomal recessive manner. A person heterozygous for
the disease is only a carrier with sickle cell trait (AS or HbAHb
S). In some cases there are
certain, mild symptoms but nothing close to life threatening. A person homozygous for the
disease - referred to as SS or HbSHb
S - has full fledged sickle cell anemia disease and exhibits
a myriad of symptoms. An affected individual will begin to show symptoms after six months
of age, this is when the fetal haemoglobin (HbF) subsides and the mutant HbS dominates.
Patients with sickle cell anemia suffer from episodic and painful vascular occlusions known
as crises that can result in organ damage and, ultimately, premature death. Currently, in
clinical practice, clotrimazole, hydroxyurea and erythropoietin are used in SCA management,
but the side effects of these drugs limit their clinical use (Rifai et al; 1995; Mehanna, 2001;
Elliott et al., 2006). In Nigeria, herbal medications are used as alternatives to orthodox
western medicine due to the perceived minimal side-effects, affordability and easy of
acquisition (Ogunkunle and Ladejobi, 2006; Okigbo et al., 2009).
Potent bioactive compounds formed during normal metabolic processes are found in specific
part(s) of medicinal plant(s), thus making such plant(s) components ideal in preparing
pharmaceutical products for medical treatments. Scientific investigation on such medicinal
plants could be of tremendous help in developing efficacious and safer drugs for SCA
treatment.

3
1.2 STATEMENT OF PROBLEM
An affected (HbSHb
S) individual becomes symptomatic, after six months of age, when the
synthesis of fetal hemoglobin (HbF) subsides and the mutant HbS dominates. Under reduced
oxygen tension, HbS polymerises and deforms the RBC. The multifaceted clinical features of
this disease are the consequences of malformed properties of sickle cells due to a point
mutation in the gene coding for the beta globin moiety of hemoglobin. The deformed
structure of the red blood cells results in two problems that lead to a multitude of clinical
manifestations. These two problems are haemolysis, which is the premature destruction of the
red blood cells, and vaso-occlusion, which is the blockage of blood flow. Symptoms of sickle
cell anemia can be categorized as either acute, episodic such as the trademark crises or they
may be chronic. Many of the symptoms linked to haemolysis are due to the body trying to
compensate by increasing erythrocyte production, resulting in abnormality of bone shape or
size and heart dilation. The main clinical consequences of haemolysis include megaloblastic
erythropoiesis, aplastic crisis, clinical jaundice and gallstones.
Even though scientists have carried out researches on sickle cell disease (SCD) for over fifty
years they are yet to find a reliable therapy. Based on current knowledge of the disease, three
different approaches to therapy have been clinically and laboratory tested (Hankins and
Aygun, 2009). One approach is the chemical inhibition of HbS polymerization. This
approach has not yet been successful enough to warrant implementation, and to date, none of
the tested antisickling drug has an acceptable efficacy to toxicity ratio. The second approach
is through the reduction of the intracellular hemoglobin concentration. Progress has since
been made in the development of drugs that inhibit potassium and water loss from SS red
cells. The most successful approach is the induction of fetal hemoglobin (HbF). The objective
is to raise the levels of HbF which inhibits sickling. The antitumor drug hydroxyurea, which

4
is also a diuretic, succeeded in increasing HbF in primates as well as patients with SCA
(Bunn, 1993), however not all patients respond to this drug and there are concerns over its
toxicity and carcinogenic effects (Weinfeld et al., 1994; IARC, 2000; Lal and Ames, 2011;
Latagliata et al., 2012).
1.3 AIMS AND OBJECTIVES
The aim of this study is to examine the efficacy of Senna alata and Senna podocarpa as
herbal remedies for SCA.
The specific objectives are as follows:
(i) To evaluate the human HbSS erythrocyte membrane stabilising effect of the
medicinal plant(s) extract and the influence of the medicinal plant(s) extract on
polymerisation/depolymerisation of mutant haemoglobin
(ii) To determine the antioxidant activity of the medicinal plant(s) extract
(iii) To evaluate the long term toxicological effect(s) of the medicinal plant(s) extract
using albino rats
(iv) To determine the effect of the medicinal plant(s) extract on rat RBC membrane
protein profile
(v) To evaluate the cytotoxic effect of the medicinal plant(s) extract on K562 Cells
1.4 SIGNIFICANCE OF STUDY
The study will help identify a herbal preparation that is safe and efficacious in protecting the
sickle cell membrane. This will help to reduce or eliminate RBC hemolysis, thereby
inhibiting haemoglobin polymerisation thus decreasing frequency of vaso-occlusive events.
Plant(s) / formulation(s) that can prevent dehydration, maintain the HbSS erythrocyte

5
membrane integrity and have minimal or no toxicity would be of potential benefit to HbSS
patients.
1.5 HYPOTHESES
The working hypotheses are:
1. Increased RBC fragility, an hallmark of SCA may be reduced by protecting the
membrane from oxidative stress thus improving the clinical outcome / outlook of SCA
patients.
2. Senna alata and Senna podocarpa have been implicated in water homeostasis, and
since the pathophysiological root of SCA is RBC dehydration, they may mediate the
underlying process of dehydration.

6
1.6 OPERATIONAL DEFINITION OF TERMS
Electrophoresis: A technique for separating the components of a mixture of molecules by
size or shape as a result of an electric field within a support gel.
Aqueous-methanolic extract: Freeze dried product of plant material(s) soluble in 80%
methanol.
Fetal haemoglobin (HbF): A type of haemoglobin made up of two alpha-globin chains and
two gamma-globin chains found majorly in the erythrocytes of neonates.
HbSS: The specific mutated allele composition of an individual with respect to the beta
globin gene located on chromosome 11, which codes for sickle haemoglobin.
K562 Cell line: Cell line originally established from a pleural effusion of a patient with
chronic myelogenous leukemia in terminal blast crisis.
Red Blood Cell (RBC): A type of cell in the blood that contains haemoglobin, which is the
oxygen carrying pigment that gives blood its characteristic red colour.
SDS-PAGE: Sodium Deodocyl Sulphate PolyAcrylamide Gel Electrophoresis - A type of
vertical gel electrophoresis used for separating mixture of molecules according to size.
Sickle Cell Anemia (SCA): A type of genetic disorder resulting from a point mutation (A-T)
on the sixth codon of the beta-globin gene located on chromosome 11, and often leading to
anemia.
Sickle haemoglobin (HbS): A type of haemoglobin made up of two alpha-globin chains and
two attenuated beta-globin chains found in the erythrocyte.

7
2.0 LITERATURE REVIEW
Nigeria has the largest population of SCA patients, estimated to be in excess of 4 million
patients with more than 150,000 children born annually with the disease (WHO, 2006). The
homozygous state of SCA is a major health challenge in Nigeria and other developing
countries, because the condition is associated with complications and reduced life
expectancy. Carrier frequency of HbS varies significantly around the world, due to high
malaria occurrence, since carriers are somewhat protected against malaria.
The genetic basis of the disease has long been established, but it is the molecular remedy that
scientists are continually investigating. With advances such as the transgenic mice,
hydoxyurea and new herbal therapies being discovered, complications and premature death of
SCA patients may in the very near future, be a thing of the past. Hydroxyurea which is
currently the drug of choice is potentially mutagenic and carcinogenic (Latagliata, 2012).
Cancer and leukemia have been reported in hydroxyurea-treated sickle
cell disease patients,
but whether the incidence is higher than in the general population is not known (Steinberg et
al., 2003). The relative risk of leukemia in hydroxyurea-treated sickle cell anemia is much
less than the observed risk in myeloproliferative disorders. To put
this into perspective, the
risk of death from the complications of adult sickle cell disease appears to be at least 10-times
greater than the possible incidence of leukemia in hydroxyurea-treated sickle cell anemia
patients (Buchanan et al., 2004).
2.1 Historical Perspective
The acutely painful episodes that characterize sickle-cell anemia disease were described in
1872 by Africanus Horton, though the mechanism remained uncertain until, nearly thirty
years later, when James Herrick (1910) coined the term „sickle cell‟ to describe the peculiar

8
morphology of the red blood cell of his patient, a dental student who presented with
pulmonary symptoms. He was not sure then, whether the blood condition was a new disease
or a manifestation of another disease, but over the next 15 years, several similar cases were
described, supporting the idea that this was a new disease and provided the platform for a
preliminary clinical and pathological description. Hahn and Gillespie (1927) thereafter,
suggested that anoxia caused RBC sickling by demonstrating that shape changes could be
induced by saturating a cell suspension with carbon dioxide. This concept was proven by
Scriver and Waugh (1930), in vivo by inducing venous stasis in a finger using rubber band in
experiments that would undoubtedly not receive institutional review board approval today.
They showed that stasis-induced hypoxia dramatically increased the proportion of sickle-
shaped cells from approximately 15% to more than 95%. These seminal studies were noted
by Linus Pauling, who was the first to hypothesize in 1945 that the disease might originate
from an abnormality in the hemoglobin molecule. His hypothesis was validated in 1949 by
the demonstration of the differential migration of sickle versus normal hemoglobin as
assessed by starch gel electrophoresis. Both hemoglobins migrated towards the positive end
but HbS migrated lesser, thus revealing that HbA had a greater net negative charge (Klug et
al., 1974). That same year, the autosomal recessive inheritance of the disease was elucidated
by James Neel and E.A. Beet through pedigree analysis, which revealed three phenotypes and
genotypes controlled by a single pair of alleles (HbA, Hb
S). Around the same time, Watson et
al. (1948) predicted the importance of fetal hemoglobin (HbF) by suggesting that its presence
could explain the longer period necessary for sickling of newborn RBC compared with those
from mothers who had “sicklemia”. Ingram and colleagues demonstrated shortly (1954)
thereafter that the mutant sickle hemoglobin (HbS) differed from normal hemoglobin A
(HbA) by a single amino acid in the primary structure of the globin portion. This finger
printing technique involved the enzymatic digestion of protein portion of hemoglobin into

9
fragments which are then subjected to an electric field for initial migration and then placed in
a solvent where chromatographic action causes a second migration. These migrations result
in a two dimensional pattern. (Klug et al., 2010) This was followed by studies that analyzed
the structure and physical properties of HbS, which formed intracellular polymers upon
deoxygenation. These studies placed SCD at the leading edge of investigations to elucidate
the molecular basis of human diseases.
Sickle cell anemia (SCA), one of the most prevalent hereditary disorders with prominent
morbidity and mortality is an autosomal recessive disease caused by a missense point
mutation in the haemoglobin beta gene found on chromosome 11p15.4 (Harrison‟s, 1998).
This mutation results in the substitution of valine (a hydrophobic amino acid) for glutamic
acid (a hydrophilic amino acid) as the sixth amino acid residue on the beta globin chain of the
structurally abnormal haemoglobin (Hb), called sickle haemoglobin (HbS). The sickle cell
anemia pathology results from the acquisition of a novel property due to the change in amino
acid sequence, rather than from the loss of normal function (Presely et al., 2010).
Haemoglobin is an oxygen carrying protein that gives red blood cells their characteristic red
colour. Patients who are homozygous for the mutation in the gene encoding the chain of
hemoglobin (globin) that results in the substitution of valine for glutamic acid at position 6
(betaGlu6Val
) have hemoglobin S (HbS) and sickle cell anemia. Hemoglobin from these
patients polymerizes in the deoxy conformation into long fibers composed
of strands of HbS
tetramers. The betaGlu6Val
mutation in deoxy-HbS favors a hydrophobic interaction between
each strand and its neighbour. Other residues on the chain participate in the binding
of
adjacent tetramers within each strand and between strands. The interaction between the valine
at position 6 with the phenylalanine at position 85 and the leucine at position 88 on the

10
partner strand is stereochemically unavailable in oxyhemoglobin. The
fibers of polymerized
deoxy-HbS are responsible for dehydration, rigidity, and lysis of red cells (Geva et al, 2004)
In persons who are heterozygous for the betaGlu6Val
mutation, hemoglobin can polymerize if
the non-HbS allele encodes permissive mutant hemoglobin (such as HbC, HbD, or HbO
Arab). In other words, patients with sickling disorders due to two heterozygous mutations
in
globin tend to have compound heterozygosity, such as HbS/C or HbS/D.
2.2 Red Blood Cells (RBC): Erythrocytes
During erythrocyte maturation, the blast forming unit-erythrocyte (BFU-E) matures into the
colony forming unit-erythrocyte (CFU-E), which finally evolves into a mature erythrocyte
after extrusion of the nucleus, acquisition of haemoglobin and a mature cytoskeletal support
system. Mature RBCs are discoid-shape and devoid of a nucleus and mitochondria. They are
rich in the heme-containing protein, haemoglobin.
Accelerated erythrocyte formation due to anemia may result in the release of RBCs prior to
the loss of the nucleus. These immature erythrocytes known as reticulocytes can be found
circulating in the blood in increased numbers (greater than 5% of the total RBCs). Once the
red blood cell is released into circulation, it has a normal life span of 120 days. Damaged or
senescent RBCs are sequestered in the spleen and destroyed by the splenic macrophages.
Pluripotent cells of the bone marrow differentiate by feedback inhibition mechanism into
various blood components. Body cells deprived of oxygen due to reduced level of circulating
oxygen transporting red blood cells triggers differentiation of bone marrow stem cells into
erythroblast which are eventually released into the blood stream.

11
2.3 Hemoglobin
The biosynthesis of haemoglobin begins in the mitochondrion, continues in the cytoplasm
and is completed in the mitochondrion. A functional haemoglobin molecule consists of four
globin chains i.e two alpha and two non-alpha globin chains referred to as a tetramer, which
are the expression products of genes on the alpha globin and beta globin loci located on
chromosome 16 and 11 respectively. Transcriptional control elements positioned within
(Persons and Nienhuis, 2000) and upstream (Li et al., 2004) of this region coordinate the
sequential activation and silencing of these genes during three defined periods of human Hb-
Grower1 (δ2 ε2 ), Hb-Grower2 ( α2 ε2) and Hb- Portland (δ2 γ2) ( gestational weeks 4-8),
fetal globin – Hb-F (α2 γ2) (gestational week 8 through parturition) and adult globins – Hb-
A(α2 β2 ) and Hb-A2(α2 δ2) ( from birth onward) (He and Russels, 2002). The expression of
embryonic globin, like fetal haemoglobin (HbF), is normally silenced in adults despite the
fact that its encoding gene remains structurally intact (Bank, 2005). The red blood cells are
initially made in the liver and spleen, then in the long bones after which production in the
liver and spleen ceases.
Polymerisation of deoxy-HbS is greatly reduced in the presence of substantial fractions of
HbA or HbF within the RBC‟s (Steinberg et al., 2003). Erythrocytes from heterozygous AS
subjects, containing about 40% HbS do not sickle in vivo under normal oxygen tension,
except under conditions where their internal Hb concentrations is markedly increased by
osmotic shrinkage due to the hypertonic environment as of the renal medulla (Ataga and
Orringer, 2000). Individuals with the sickle cell trait are asymptomatic and have normal
haematological profiles, except under high stress of maximum exercise or low oxygen
tension (high altitude or sudden decompression in aircraft) (Bunn and Forget, 1986;
Gendreau and DeJohn, 2002; Lee et al., 2002), but individuals doubly heterozygous

12
(HbSHb
F) for sickle Hb and for hereditary persistence fetal Hb, show no clinical evidence of
sickling and have normal haematological profiles. In sickle cell (SS) patients Hb conbinations
that prevent sickling invivo also prevent the development of the haematological and clinical
manifestations associated with dehydrated RBC‟s, despite substantial proportions of HbS
within the cells.
Human RBC‟s are water permeable sacs with the highest soluble protein concentration of any
cell approximately 7.2 mmol Hb/cell water. The RBC‟s characteristic biconcave shape and
flexibility is made possible due to its two dimensional spectrin-actin based meshlike
cytoskeleton, with a highly structured link to integral membrane proteins embedded in the
lipid bilayer. (Chasis et al., 1989; Palek and Lambert, 1990). Continued osmotic equilibrium
in the plasma is achieved by the RBCs‟ high water permeability so that they can swell or
shrink only by the loss or gain of a fluid isoosmotic with surrounding plasma. Since the
plasma protein concentration is less than 1mM, there is powerful pressure driving water into
the cells. Osmotic stability over the approximate 120 day circulatory life span of the mature
RBCs is actualised due to an evolved strategy of maintaining a nearly constant volume with
minimal energy consumption. Because of their low Na+ and K
+ permeabilities which require
relatively few metabolically fuelled Na+ pumps per cell to balance their cation gradient
driven leaks, RBCs have evolved an extremely efficient CO2 ferry between tissues and lungs,
based on the high Cl- and HCO3
- exchange (Lew and Bookchin,1986) and electrodiffusional
fluxes (Knauf et. al,. 1983). This homeostatic balance is markedly altered in sickle Hb
containing RBCs whose ion content regulation, ion fluxes and hydration state become highly
distrupted in the circulation. Enucleated erythroid cells termed reticulocytes released from the
bone marrow into the circulation differentiate into mature RBCs within 2-3 days. The
maturation process involves loss of residual RNA as well as protein synthesis with

13
concomitant reduction or outright loss of many metabolic functions including a gradual
decline in monovalent cation transport activity and the reduction or inactivation of selected
transporters such as K+-Cl
- cotransporter and Na
+ pumps, via the formation of exosomes
(Johnstone, 1992).
2.4 Transport Proteins Involved In RBC Dehydration
Human RBC dehydration in vivo may arise from the single or synergistic activation of the
K+-Cl
- cotransporter (KCC), the Na
+ pump or the Gardos channel. These three transporters
expressed in the RBC plasma membrane differ considerably in their dehydrating potencies,
modalities and distribution.
2.4.1 K+-Cl
– Cotransporter (KCC)
The KCC, regulated by internal pH and cell volume, is functionally active in reticulocytes
and much less so in mature AA and SS RBCs. Most of the K+ traffic is mediated by this
transporter, which may be regulated by cell swelling, intracellular acidification, and
oxygenation state of Hb (Joiner and Franco, 2001) and to a lesser extent low intracellular
Mg2+
concentrations. Its functional state appears to be regulated by classical serine/threonine
and tyrosine residue (Merciris et al., 2003) phosphorylation and dephosphorylation (Lauf and
Adragna, 2000). Acidification in RBCs with active KCC may induce dehydration (Lauf et al.,
1992; Gillen et al, 1996), the extent of which may be limited by the inhibitory effect of cell
shrinkage, while in SS reticulocytes acid actification is abnormally exaggerated and may
therefore overcome the intensity of the volume-regulatory “brake”. Inhibitory agents may act
directly on the KCC or on its regulatory intermediates (Jennings and Schulz, 1991).

14
2.4.2 The Na+ Pump
The Na+-K
+ flux ratio through the Na
+ pump is 3:2 (Lew et al., 1991). The high anion
permeability of reticulocytes and RBCs, require that electroneutrality be maintained, majorly
by anion efflux balancing the extra Na+ efflux. Elevated internal Na+/ K
+ concentration ratios
stimulate the Na+ pump, resulting in an increased NaCl flux, which if not compensated by
passive fluxes would induce dehydration (Joiner et al., 1986). Model simulations (Glynn and
Karlish, 1975; Brugnara et al., 1989; Skou and Esmann, 1992) confirms this effect to be
rather slow, moreover, in SS cells Na+ pumps have been found to be substantially inhibited
(Clark and Rossi, 1990; Ortiz et al., 1990), despite elevated internal Na+/K
+ . This inhibition
was due to elevated Mg/ATP ratio, because Mg2+
released from 2, 3-diphosphoglycerate (2,3-
DPG) during deoxygenation, pushed the ratio further away from the optimal (Ortiz et al.,
1990). Thus Na+ pump-mediated fluxes may contribute only marginally to SS cell
dehydratioin.
2.4.3 Gardos Channel
The Gardos channel is a Ca2+
-sensitive, small-conductance, K+-selective
channel which is
activated when the physiological intracellular RBC Ca2+
concentration levels of about 20-50
nM is increased above its activation threshold of 150 nM (Tiffert and Lew, 2001). A
maximally activated Gardos channel would ensure fast dehydration of the RBC. Unlike
erythroid cells from other mammalian species, whose Gardos channel activity
is lost during
maturation (Brown et al., 1978), mature human RBCs show persistent Gardos channel
activity. The capability of Gardos channel, when fully activated, to mediate rapid RBC
dehydration far exceeds that of the other transporters. Dehydration via Gardos channels may
be reduced by direct inhibition of the channels with charybdotoxin (IC50 1.2 nM) or
clotrimazole (IC50 51 nM) (Brugnara et al., 1993), or indirectly, by inhibition of the anion

15
permeability (Bennekou et al., 2001). Mature RBCs, when compared with other cell types
lack specialised Ca2+
accumulating organelles, Ca 2+
signalling functions as well as minimal
cytoplasmic buffering capacity (Tiffert and Lew, 1997).
2.5 Dehydration of RBC
The relationship between SS cell transport and sickling was investigated in 1955 by
Tosteeson and co-workers, who showed that upon deoxygenation of fresh, heparinised whole
blood obtained from HbSS patients their RBCs had a much larger gain of Na+ and loss of K
+
than RBCs from HbAA (normal) controls. But on reoxygenation, HbSS cells gained K+ and
lost Na+, affirming that the effects of deoxygenation were reversible. Further transport
experiments with tracers of Na
+, K
+, and Cs
+ indicated that sickling induced a reversible
and
poorly selective increase in the electrodiffusional cation permeability of the RBC membrane.
A sickling induced permeability pathway was proposed by Lew et al. (1997) When HbSS
RBCs was ultracentrifuged, a cell subpopulation accumulated at the bottom of the column,
these cells had normal Hb content, and their high density reflected a dehydrated state. The
distinct morphological features of this subpopulation earned the name irreversibly sickled
cells (ISCs), due to the fact that when fully oxygenated, they still retained the sickled shape.
Isotopic labelling techniques revealed that
ISCs were a relatively young HbSS cell
subpopulation which, after release from the bone marrow as reticulocytes, was rapidly
transformed into ISCs within 4–7 days, and then disappeared from the
circulation faster than
other mature, non-ISC RBCs. ISCs were therefore a subpopulation of SS RBCs in rapid
circulatory turnover. The exclusion of most high Hb F-containing HbSS cells (whose
polymerization
was reduced or inhibited) from the hyperdense, ISC-rich cell
fraction
suggested that sickling was necessary for ISC formation. Most research in the field was
dominated by the notion of gradual dehydration, with attention focused on the three transport

16
systems potentially involved in HbSS RBC dehydration, notably, the Gardos channel, KCC
and Na+ pump.
2.6 Pathophysiology
The sickle phenotype results from the intracellular polymerization of deoxygenated α2βs2
heterotetramers (HbSS), into extended 14-strand fibres that disrupt both the shape and the
function of the mature erythrocyte (Bunn, 1997). The deformation of RBC causes changes in
their membrane structure (Ohnishi and Ohnishi, 2001), thereby enhancing the adhesion of the
cells to vascular endothelial cells (Hoover, 1979; Brittain et al., 2004). This, together with
elongated cell shapes, induces the obstruction of blood flow and causes the painful sickle cell
crisis. The red blood cell membranes of SCA patients are osmotically and mechanically more
fragile than those of normal subjects. For these reasons, sickle RBCs are easily destroyed and
removed from the circulation in the spleen, thus causing anemia. The average life span of
sickle erythrocytes is 10 - 20 days (Franco et al., 1998), as opposed the 120 days for the
normal RBC. Patients suffer from chronic anemia, frequent painful episodes (crisis) and
resultant malfunction of organs (especially the spleen) and degeneration of bone joints
(Serjeant, 1997).
The total cellular calcium content of sickle RBC is markedly abnormal (50µmol/L as against
5µmol/L for normal RBC), but oxygenated sickle RBC have moderately enhanced
permeability to external calcium and deoxygenation boosts calcium influx by about five fold
(Etzion et al.,1993). Though excess calcium is stored internally in vesicles without increase
in detectable ionized calcium, this influx activates a calcium-dependent potassium channel
called Gardos channel, through which potassium is extruded from the sickle RBC, thus
resulting in dehydration. Stimulation of sickle RBC membrane Ca2+
ATPase activity under
optimized conditions among patients have been found to be increased, deficient and normal,

17
suggesting an insignificant effect on the calcium pumping capacity under physiological
conditions. Stimulation of the Na+K
+ATPase by deoxygenation of sickle RBC somewhat
increases passive K efflux and Na influx three to five fold, this leak sites (Lew et al., 1991),
areas of the membrane affected by speculation – a sickling effect – requires cell deformation
for induction of monovalent cation leak, though oxidative perturbation of the sickle
membrane may however not be ruled out. Some studies on RBC suggest that full homeostatic
capacity of the cell (- Quabain) leads to cellular dehydratioin while the passive component of
sickling-induced leak (+ Quabain) causes balanced leak. Cell swelling or low pH (<7.4)
activates in sickle reticulocytes, a potassium-chloride cotransport (KCC) pathway.
Irreversibly sickled cell (ISC) generated by repeated deoxygenation / oxygenation of sickle
RBC, regarded as the end-stage defects of highly damaged cells, can constitute anywhere
from a few percent to about 50% of RBC population. Formation of ISC in vitro is generally
prevented by an absence of external calcium, inhibition of Gardos channel, calcium channel
blockers and calmodulin-mediated processes. The calcium antagonist, zinc, appeared to
reduce ISC counts in patients consuming it, but formation of ISC formation in vitro was not
inhibited by in the presence of zinc. The implied role of calmodulin is interesting given its
interaction with the RBC cytoskeleton and the effects of calcium on certain protein / protein
interactions (Blackman et al., 2001; Chang and Low, 2001).
2.7 Cytoskeletal Dysfunction
The flexibility and strength of the normal RBC is derived from the protein component of its
membrane, the essential features of which is an underlying cytoskeleton ( spectrin, actin,
band 4.1 and other minor components) connected by linking units (ankyrin and band 4.1) to
proteins embedded in the lipid bilayer (band 3 and glycophorin). In sickle cell anemia, this
actin / spectrin lattice „locks‟, making red blood cells much less deformable, and causing

18
them to obstruct the microcirculation. Sickle inside-out vescicles (IOV) have a diminished
capacity for ankyrin-dependent binding of spectrin (Platt et al., 1985) and
aminophospholipid, sickle IOV were found to have a slightly diminished quantity of band 4.1
(Schwartz et al., 1987) although artifacts due to protease action was not excluded. Sickle cell
patients administered vitamin E tend to have normalised band 3 anion transport activity and
high affinity ankyrin binding sites (Joiner et al., 1989). Careful analysis of sickle RBC
membranes showed a small amount of abnormal spectrin / globin adduct (Presley et al.,
2010), favoured by dehydration, reflecting an oxidative process possibly involving spectrin
(not Hb) thiols in the presence of peroxide (Rank et al., 1985)
Sickle RBC fragility has been observed to increase in patients undergoing vigorous exercise.
Invitro studies have documented notable reduction in the shear sensitivity of intact sickle
cells on rehydration and ghosts made from most dense sickle RBC are mechanically fragile.
This membrane instability reflects some protein/protein junctional association failure and
destabilization of the cytoskeleton maybe due to free heme found to be in excess in both
sickle membrane and sickle cytosol (Shalev and Hebbel, 1996).
Decreasing deformability of deoxygenating sickle RBC is highly dominated by development
of Hb polymer and cytoplasmic viscosity. It may be anticipated that abnormal microrheology
of sickle cell membrane would participate in vasoocclusion as increased static rigidity would
enhance mechanical trapping of RBC during their attempted passage through the
microvasculature, or capillaries of limiting diameter and flow velocity of cells therein
(Ferrone, 2004).

19
2.8 Cell / Cell Interactions
Abnormal attachment of sickle RBC to mononuclear phagocytes or vascular endothelial cells
indicates relevance to haemolytic anemia and vasocclusive events respectively. Sickle RBC
have been found to be abnormally adherent to normal marrow, splenic or alveolar
macrophages and normal peripheral blood monocytes, after which they are phagocytosed
more readily than normal RBC. The ability of externalised phosphatidylserine (Yamaja Setty
and Betal, 2008) in promoting adherence to macrophages in vitro is enhanced by
deoxygenation, but exposure of normal RBC to peroxide or malondoaldehyde (peroxidation
by product) facilitates their phagocytosis. Abnormal adherence of sickle RBC to vascular
endothelial cells derived from bovine aorta, rat microcirculation and human umbilical vein is
enhanced by dehydration or calcium loading (Hoover et al., 1989; Kaul et al., 1993).
2.9 Pathophysiologic Role of Membrane Abnormalities
Emphases on HbS polymerization have dominated considerations of sickle cell disease
pathophysiology generally but it is now recognised widely that this disease is exceedingly
complex. Attempts to explain it based on a single feature would be artificial and the
intriguing heterogeneity amongst patients would be grossly underestimated. Severity of
anemia for sickle cell patients correlate directly with the magnitude of the very-dense cell
population.
2.10 Oxidant Levels
Increased oxidant susceptibility of sickle red blood cells (RBC) has been demonstrated to
play a major role in pathophysiology of the disease, in effect, reactive oxygen species (ROS)
produced during redox cycles appear to cause premature ageing and altered morphology of
RBCs, leading to their early removal from circulation. Another name for redox cycle

20
formation of reactive oxygen species and damage to the RBC is „oxidative stress‟. In other
cases toxicants can cause a disruption in oxidative metabolism within the cell, leading to
generation of reactive oxygen species. An important potential consequence of free-radical
formation is the occurrence of lipid peroxidation in the membranes within the cell. Lipid
peroxidation occurs when free radicals attack the unsaturated bonds of fatty acids,
particularly those in phospholipids. The free radical reacts with the fatty acid carbon chain,
abstracting one hydrogen atom. This causes a fatty acid carbon to become a radical, with
rearrangement of double bonds in the fatty acid carbon chain. This carbon radical in the fatty
acid reacts with oxygen in a series of steps to produce a lipid hydroperoxide and a lipid
radical that can then react with another fatty acid carbon. The peroxidation of the lipid
becomes a chain reaction, resulting in fragmentation and destruction of the lipid. Because of
the importance of lipids in membrane structure, the principal consequence of lipid
peroxidation for the cell is loss of membrane function. The reactive products generated by
lipid peroxidation can interact with other components of the cell as well, and this also could
contribute to cellular perturbation. Protecting the RBC membrane from oxidative stress may
thus improve the clinical manifestations of the patients (Ohnishi and Ohnishi, 2001).
2.11 Factors Contributing to Vasocclusion
HbS polymerization in vasocclusive events for most sickle RBC is never in equilibrium and
sickling is dominated by kinetics, so that clinical effects depend critically on the relationship
between microcirculatory transit time (short) and the unavoidable and relatively longer delay
time (about 15 seconds) for the onset of Hb polymerization (Bunn, 1997). It is controversial
whether or not the RBC membrane directly influences polymerization the occurrence of
painful crises actually correlates inversely with percentage dense cells and percentage ISC,
but correlates positively with adequate RBC deformability.

21
2.11.1 Environmental factors
The role of blood factors including plasma proteins (as modulators of RBC adhesivity), blood
pH (as stimulant for cation depletion, as well as Hb polymerization), osmolarity (as
determinant for dehydration) and zinc levels (as calcium antagonist) in vasocclusive crisis
cannot be underemphasized. Since cellular pathobiology is influenced by oxidative
phenomena, there is the possibility of accessibility to dietary factors like selenium, riboflavin,
vitamins E and C, exerting some beneficial influence. Presence of other globins like HbA or
HbF (levels varying enormously amongst patients), inhibit polymer formation while others
e.g. HbD and HbOArab
promote polymer formation.
2.11.2 Internal factors
Proper functioning of various RBC enzymes [(Gluthathione-S-transferase (GST),
superoxidedismutase (SOD), Catalase (CAT), Methemoglobin reductase (NADHmr),
Diaphorase (NADPHd) and Glucose-6-phosphate dehydrogenase (G-6PD)] systems play
crucial roles in sickling. The SODs are a group of metalloenzymes that catalyze the
conversion of reactive superoxide anions (O2-) to yield H2O2, which is in itself, is an
important ROS as well. Hydrogen peroxide is subsequently detoxified by two types of
enzymes; catalases and glutathione-dependent peroxidases (GPOXs). Superoxide dismutases
are considered to play a pivotal antioxidant role. Their importance is indicated by their
presence in all aerobic organisims examined (Stegeman et.al., 1992). The SOD-CAT system
provides the first defence against oxygen toxicity. Superoxide dismutase catalyses the
dismutation of the O2- to molecular oxygen and hydrogen peroxide which is detoxified by
CAT activity to water and oxygen. (see equation below)
Equation. 2O2- + SOD → H2O2 + O2 + CAT → H2O + O2

22
Fig.1 A schematic diagram showing the inter relatioship between the possible role of Hb
polymerization, membrane defects and abnormal behaviour of HbS in sickle cell anemia disease
pathophysiology
Sickle
Hemoglobin
Hb polymerization
Decreased solubility
Vaso-occlusion and
Hemolytic anemia
RBC sickling
Oxidant generation
Iron compartmentalization
Hb/membrane interaction
Abnormal microrrheology
Endothelial adhesivity
Marcrophage interaction
Phospholipid destabilization
Protein defect
Decreased stability
Cellular dehydration

23
2.12 Histopathology
Indicators of stress at several levels of biological organisation have been used to evaluate
effects of contaminants / toxicants at organizational level. These approaches vary from
measures of genetic integrity to growth and reproductive competence of the individual (Teh
et. al., 1996). As far as the individual organism is concerned, manifestations of stress at the
tissue level represent an intermediate effect between the biochemical and reproductive levels
(Hinton, 1990). Histopathological characteristics of specific organs express condition and
represent time-intergrated endogenous and exogenous impact on the organism stemming
from alterations at lower levels of biological organisation (Chavin, 1973). According to
Segner and Braunbeck (1998), histological changes occur earlier than reproductive changes
and are more sensitive than growth or reproductive parameters and, as an integrative
parameter, provide a better evaluation of organism health than a single biochemical
parameter. Many toxicants can damage specific cell types or organ systems.
2.13 Orthodox Treatment
The main drug therapy for sickle cell anemia, hydroxyurea, is thought to reduce the formation
of sickle haemoglobin and hence ameliorate the structural consequences, while also
decreasing neutrophil numbers which promote adhesion of sickled cells to blood vessel walls.
However, a recent study suggests that hydroxyurea acts directly on the plasma membrane.
The drug is known to decrease expression of adhesion molecules on the red blood cells, one
of which is phosphatidylserine, usually expressed on the outer surface of some RBCs in
sickle cell anemia. In a study by Covas et al., 2004, sickle cell patients receiving 12 months
of hydroxyurea treatment had significantly reduced phosphatidylserine levels in their blood
samples.

24
The multifaceted clinical features of this disease are the consequences of malformed
properties of sickle cells. The deformed structure of the red blood cells results in two
problems that lead to a multitude of clinical manifestations. These two problem mechanisms
are haemolysis, which is the premature destruction of the red blood cells, and vaso-occlusion,
which is the blockage of blood flow. Symptoms of sickle cell anemia can be categorized as
either acute, episodic such as the trademark crises or they may be chronic. Many of the
symptoms linked to haemolysis are the results of the body trying to compensate. The body
compensates by increasing erythrocyte production, resulting in abnormality of bone shape or
size and heart dilation due to increased heart action. The main clinical consequences of
haemolysis include megaloblastic erythropoiesis, aplastic crisis, clinical jaundice and
gallstones (Issa and Al-Salem, 2010).
The life span of erythrocyte in people with sickle cell disease is ten to twenty days as
opposed to a normal 120 day span (Franco, 1998). This increased rate of destruction demands
a significant increase in bone marrow activity. Because of the raised level of bone marrow
activity a greater amount of folic acid is required. The shortage of this acid results in
megaloblastic erythropoiesis with low reticulocyte counts, increasing mean cell volume, and
reduced haemoglobin. The increased metabolic demand of the bone marrow competes with
the demands of growth plates, usually resulting in impaired growth. Aplastic crisis, from
bone marrow depression may occur and its characteristic symptoms include lethargy, dyspnea
and possibly coma. The rapid haemolysis means increased bilirubin excretion which is
associated with clinical jaundice and the formation of gallstones (Alexander-Reindorf et
al.,1990; Issa and Al-Salem, 2010).
The second problematic mechanism, of the deformed red blood cells is the blockage of
venous microcapillary, resulting in impeded blood flow. Tangled, sickled cells obstruct

25
vessels, causing tissue anoxia and possible necrosis. Manifestation of symptoms depends on
which vessels that are being blocked. Damage to the splenic vasculature via obstruction, is an
early pathological signature of sickle cell anemia. The spleen normally acts like a filter,
removing damaged red blood cells and bacteria from the blood stream; however, in sickle cell
patients the damaged red blood cells obstruct this filter, predisposing the individual to
infection. The most common crisis, vaso-occlusive crisis, is caused by this blockage
mechanism. Vaso-occlusive crisis (painful episode) is the term used for the excruciatingly
painful period while the vessels are occluded by cells and tissue is being deprived of oxygen.
It is characterized by severe thoracic, abdominal, muscular or bone pain. This form of crisis
may last from a few hours to several weeks (Okpala and Tawil, 2002; Wright and Ahmedzai,
2010). Vessel blockage leads to extra complications in pregnant women, where mother and
foetus face increased risk due to oxygen variability and children also face the devastating risk
of stroke due to the occlusion of major cerebral vessels.
Sickle cell patients may develop chronic complications such as leg ulcers, below average
growth rate and complications from organ infarctions such as retinopathy, nephropathy and
possibly auto spleenectomy (Hassell, 1994).
Diminished solubility of HbS in its deoxygenated state results in the formation of a network
of fibrous polymers, which stiffen and distorts the red blood cell‟s shape. This polymerization
produces the classic sickle shape. The kinetic feature of this polymerization is critical. When
rapid deoxygenation occurs, polymerization doesn‟t alter the cell‟s disk-like shape. However,
when HbS red cells are slowly deoxygenated a nucleus of HbS molecules combine, grow the
fibers and transform into the sickle shape (Bunn, 1997).
The rate and extent of polymer formation in the SS red cells depends on three variables that
dictate the extent of polymerization and thus, to an uncertain degree, the severity of sickle

26
cell disease. Those variables are the degree of deoxygenation, intracellular hemoglobin
concentration, and presence of fetal hemoglobin. Deoxygenation causes polymerization
induced damage which leads to dehydration resulting in formation of dense cells which cause
sickling. This appears to be the basic domino effect. The polymerization rate of deoxygenated
HbS, is dependent on the hemoglobin concentration. This translates into the fact that dense
HbS cells are more likely to sickle. Dense cells are partly created by dehydration. There are
two main contributors to dehydration in these mutant red blood cells. First is the greatly
increased rate of potassium chloride co-transport in the erythrocytes. The second contributing
factor results from the increased calcium triggering a potassium channel, thus providing
another pathway for the loss of water (Bunn, 1997). The third variable in the severity of the
expression of the disease is the presence of fetal hemoglobin, which until approximately the
sixth month of life is the predominant type of haemoglobin (Stamatoyannopoulos, 2005;
Oneal et al., 2006). Thus affected infants are more or less symptom free. After this time
period the HbS is at normal levels of production and symptoms are revealed. In adults, the
level of HbF correlates with the parental HbF levels in a genetically dominant pattern (Milner
et al., 1984). HbF inhibits polymerization which halts the domino effect that generally results
in sickled cells.
Research on sickle cell disease has been on for over fifty years and the current knowledge of
the disease dictates three different approaches to therapy which have been clinically and
laboratory tested. One approach is the chemical inhibition of HbS polymerization. The
challenge faced is a formidable one, since the drug must be absorbed, circulate in the plasma
without binding strongly to plasma proteins, penetrate the erythrocyte membrane and bind
specifically to the HbS. To date, no antisickling drug has been tested with an acceptable
efficacy to toxicity ratio (Bunn, 1993).

27
A second plan of attack is the reduction of the intracellular hemoglobin concentration. This
approach relies on the dependency of the rate of polymerization of HbS on the HbS
concentration. Progress has seen made in the development of drugs that inhibit potassium and
water loss from SS red cells. This in turn reduces the intracellular hemoglobin concentration.
The drug Clotrimazole inhibits potassium loss at a specific channel and has proven effective
in transgenic mice and patients with sickle cell anemia (Bunn, 1993).
The other mode of cell dehydration, potassium-chloride co-transport, has not yet fully
conformed to pharmacological manipulation, despite it‟s inhibition by high intracellular mg2+
(Buchanan et al., 2004)
The most successful of the three approaches is the induction of fetal hemoglobin (HbF). The
objective is to raise the HbF levels because it inhibits sickling. The antitumor drug
hydroxyurea succeeded in increasing HbF in primates as well as patients with SS disease
(Bunn, 1993).
Jayabose and associates (1996) ran a study on treatment with hydroxyurea of children with
severe sickle cell anemia. Their aim was to determine the effect on the hemoglobin levels, to
evaluate the toxicity of the drug and to detect any change in the frequency of the vaso-
occlusive crises. Group one was made up of children with sickle cell anemia who suffered
frequent vaso-occlusive crises. The second group consisted of children with sickle cell and
severe anemia. Each group was given 20-35mg/kg/day. The frequency of vaso-occlusive
crises before and after treatment and the peak hemoglobin levels before and during treatment
were recorded. The trial resulted in a statistically significant drop in the number of crises,
hospital days and transfusions per year. The hemoglobin levels increased from 8-50% over
baseline values. Multiple factors such as increasing mean corpuscular volume and decreasing

28
adhesion of red blood cells to the endothelium, are probably responsible for the clinical
benefits of hydroxyurea.
One of the most important breakthroughs dealing with this disease did not even involve a
human patient, but rather mice were in the limelight. Two separate research teams developed
mutant mice that mimic human sickle cell anemia. The mice globins were completely
eliminated and complement human genes, including the sickling mutant form of beta globin,
were introduced. These mice make only the disease causing, mutant form of hemoglobin that
clumps when deoxygenated and sickles. The mice suffer the same array of symptoms that a
human with sickle cell disease endures. The creation of this genetically unique mouse is so
valuable because now drugs and therapies can be tested just as they would in a person with
the disease. The mice have exactly the same mutant beta globin that the majority of affected
humans possess. Until the development of these mice, experiments were conducted in test
tubes or on patients where a full range of possibilities couldn‟t be tested (Barinaga, 1997).
The effectiveness of the drug clotrimazole in reducing the intracellular hemoglobin
concentration was also tested on these transgenic mice.
Prevention is the best form of treatment to prevent the intravascular sickling. The trademark
crises of sickle cell are precipitated by fever, dehydration and cold exposure. To prevent these
is an effort to prevent the extremely dangerous crises. Sudden transition to high altitudes and
exposure to freezing temperatures should be avoided. Hydration is essential, as are
immunizations.
The most effective of the non-controversial therapies available is the blood transfusion. This
therapy includes packed red cell transfusions at three week intervals. This is generally enough
of an effort to maintain the donor cell (HbA) circulation above fifty percent. The transfusions
reduce the tendency for sickling by diluting the mutant host cells, temporarily suppressing the

29
production of erythrocytes containing HbS, thus improving blood and tissue oxygenation.
However this therapy does have its downsides. It requires a tremendous blood resource and
the risks of hepatitis and alloimmunization are potentially life threatening (Sandler, et al.,
1997).
Bone marrow transplants have the potential to regulate the hemoglobin synthesis. This
procedure causes life threatening complications in itself. Once these complications are
improved, it will most likely become an option for severe sickle cell anemia patients (Ferster
et al., 1992).
Since it is apparent that prevention of complications is the best form of treatment for this
disease, it is helpful to detect the mutation early on. When it comes to genetic screening,
sickle cell disease has had a rocky past. In the early 1970‟s efforts were made to identify
carriers of the recessive allele for the disease. The goal of this search was to advise the carrier
which would help in family planning which would hopefully lower the disease incidence.
However this plan backfired for several reasons. The counselling was not adequate and
resulted in people confusing the harmless trait for the deadly disease. People were
discriminated against at jobs and in society in general. It was even beleived that this was the
white man‟s way of medically reducing the reproduction of African Americans. Needless to
say the screenings were stopped. Several key ideas were learned from this fiasco which is
now been implemented in screenings for diseases from HIV to breast cancer. The test must
have a direct benefit to the person tested. The results must be kept in the strictest confidence
and the screening must be followed by counseling (Klug et al, 2010).
Prenatal diagnosis allows for preventative measures to begin while the child is still in the
mother‟s womb. Fetal cells may be acquired by amniocentesis. The DNA is extracted from
the cells and digested with restriction enzymes. The enzyme would cut twice within the

30
normal beta globin gene, once at each site. Thus a normal gene produces two small DNA
fragments. However, in the mutant alleles the second site has been destroyed by the mutation
resulting in the production of one long restriction fragment. When run on a gel, one long band
with two short fragments identifies a heterozygous state. This person will be a carrier. A gel
showing one long band identifies a person homozygous for the sickle cell mutation (Klug et
al, 2010).
With the aforementioned advances such as the transgenic mice and discovery of hydoxyurea,
that positive headline may be in the very near future. Due to the genotoxicity, carcinogenicity
of hydroxyurea as well as the non-responsiveness of some patients to hydroxyurea therapy,
other therapeutic options are being considered.
2.13.1 Therapeutic options based on disease pathophysiology
2.13.1.1 HbS Polymerization
Deoxygenation induced polymerisation of HbS (α2ß2S) is the
proximate cause of sickle cell
anemia and a necessary but insufficient precursor of the disease phenotype. HbS and its
polymer induce a variety of cellular and tissue injuries, but neither the fetal
hemoglobin
(HbF) tetramer (α2γ2) nor the α2ßSγ hybrid
tetramer is incorporated into the HbS polymer,
thus providing the rationale for treatments aimed at increasing HbF concentration.
2.13.1.2 Sickle Erythrocyte Damage and Dehydration
The presence of dense, dehydrated erythrocytes and abnormal reticulocytes is a distinguishing
feature of sickle cell anemia. Since polymerization of HbS is uniquely dependent on the
cellular concentration of HbS, the tendency
for HbS polymerization and cell sickling is
markedly enhanced by the increased cellular hemoglobin concentration of dehydrated sickle

31
erythrocytes. Of the four pathways that have been implicated in the dehydration of sickle
erythrocytes, and modulation of these pathways, especially the Gardos channel
and K
+-Cl
– co-
transport, is a potential means of treatment.
2.13.1.3 Endothelial damage
Cellular damage enables adhesive interactions between sickle cells, endothelial cells and
leukocytes. Vasoconstriction may be favoured as nitric oxide (NO) production is impaired in
a perturbed endothelium. Several agents directed at the endothelial receptors for sickle
erythrocytes or leukocytes may interrupt cell-cell interactions.
2.13.1.4 Inflammation, reperfusion injury, oxidant radical production
Neutrophils mediate inflammation and tissue damage. Neutrophil numbers are increased and
they may be abnormally activated and adherent. An "oxidant" environment may also be
present in the sickle erythrocyte and endothelium. Novel ways of countering
oxidant-induced
injury have been proposed, chiefly, via exogenous antioxidant intake
2.13.2 HbS Polymerization
Several classes of drug, when titrated optimally, can increase levels of HbF in most patients
with sickle cell anemia. Only one is approved by the American Food and Drug Agency for
treating sickle cell anemia.
2.13.2.1 Hydroxyurea
Hydroxyurea, the sole US Food and Drug Administration (FDA)–approved drug for treating
sickle cell anemia, could be used in all adults where indications for this treatment are present
(Steinberg, 1999). Unfortunately, for complex reasons, only a fraction of patients who might
benefit from treatment receive it. Hydroxyurea increases HbF in sickle
cell anemia because its

32
cytotoxicity causes erythroid regeneration and perhaps because its metabolism leads to NO–
related increases in soluble guanylate cyclase (sGC) with an increase
of cGMP that augments
gamma-globin gene expression (Cokic et al., 2003). A multicenter trial of hydroxyurea in
adults with sickle cell anemia, where the drug was given at sub-toxic doses, showed that
hydroxyurea reduced the incidence of pain and acute chest syndrome by nearly
half, with
little risk seen during more than 9 years of observation. Cumulative mortality was reduced
nearly 40%, and a favourable result was related to the ability of the drug to increase HbF
and
reduce painful episodes and acute chest syndrome (Steinberg et al., 2003) No relationship
between decrements in neutrophil counts and mortality was found. In infants, children and
adolescents with sickle cell anemia, their HbF response to hydroxyurea is more robust than in
adults. In a study of more than 100 children who received maximal drug
doses, HbF increased
to almost 20% and the treatment effects were sustained for 7 years without clinically
important toxicity (Zimmerman et al., 2004). HbF levels achieved during treatment were
associated with baseline HbF level, hemoglobin level, reticulocyte count, and leukocyte
count
and with compliance to treatment. According to some experts, pushing the drug dose to near
toxic levels is not necessary for a clinically beneficial result. Some have proposed using
hydroxuyrea for secondary prevention of stroke in children (Ware et al., 1999). Hydroxyurea
may also conserve resting energy expenditure by curbing the hypermetabolic state observed
in children with sickle cell disease (Fung et al., 2001)
Hydroxyurea is potentially mutagenic and carcinogenic (Latagliata, 2010). Cancer
and
leukemia have been reported in hydroxyurea-treated sickle cell disease patients, but whether
the incidence is higher than in the general population is not known (Steinberg et al., 2003).
The relative risk of leukemia in hydroxyurea-treated sickle cell anemia is much less
than the
observed risk in myeloproliferative disorders. To put this into perspective, the risk of death

33
from the complications of adult sickle cell disease appears to be at least 10-times
greater than
the possible incidence of leukemia in hydroxyurea-treated
sickle cell anemia patients
(Buchanan et al., 2004).
2.13.2.2 Decitibine
A less-toxic analog of 5-azacytidine, 5-aza-2'-deoxycytidine (decitibine), may affect HbF
levels by causing hypomethylation of the gamma-globin genes. In 8 symptomatic sickle cell
anemia patients who failed to respond to hydroxyurea, decitibine treatment (0.2
mg/kg
subcutaneously 1–3 times per week for 2 6-week cycles) led to an increase in HbF from 6.5%
to 20.4%, with an increase in hemoglobin concentration from 7.6 to 9.6 g/dL and a fall
in
reticulocytes from 231 to 163 x 109/L (Saunthararajah et al., 2003).
2.13.2.3 Short chain fatty acids
By acting as inhibitors of histone deacetylase (HDAC) and causing histone hyperacetylation
and changes in chromatin structure, short-chain fatty acids, their derivatives, and other
compounds with HDAC activity can enhance gamma-globin gene expression in erythroid
cells of patients with sickle cell anemia and ß thalassemia. Very low concentrations of one
HDAC inhibitor, an analog of trichostatin A, induced gamma-globin gene expression in an
erythroleukemia cell line transfected with a reporter construct and in erythroid
colonies of
normal adults (Cao, 2004)
In the most advanced clinical trials of HDAC inhibitors, still only Phase II studies, arginine
butyrate given by infusion once or twice a month was associated with a mean increase in HbF
from 7% to 21% in 11 of 15 patients with sickle cell anemia. In some individuals, this level
was maintained for 1–2 years (Atweh et al., 1999). No HDAC inhibitor of any class other
than butyrate and phenylbutyrate has yet been used clinically.

34
2.13.3 Sickle Erythrocyte Dehydration
2.13.3.1 Inhibition of K+-Cl
– co-transport
Oral magnesium supplementation inhibits erythrocyte K+-Cl
– co-transport in vivo. Following
studies that showed a beneficial effect on the erythrocyte membrane of transgenic sickle
mice, a 6-month clinical trial of oral Mg pidolate improved erythrocyte
hydration and was
associated with a reduction in the number of painful days (De Franceschi et al., 2000).
Studies to evaluate the effects of long-term magnesium supplementation in adult and pediatric
patients with sickle cell anemia are ongoing, and additional studies are planned
in HbSC
disease patients where activated K+-Cl
– co-transport
and cell dehydration are likely to play a
major pathophysiological role (Buchanan et al., 2004)
2.13.3.2 Inhibition of Gardos channel
Clotrimazole is an inhibitor of the human red cell Gardos channel but its use was associated
with dysuria and reversible hepatocellular toxicity. ICA 17043, a clotrimazole derivative
lacking the toxic imidazole residue, was a 10-fold more potent blocker of the
Gardos channel
than the native drug. In a recently completed Phase II trial of this agent in patients with sickle
cell disease, cell density and hemolysis were decreased while hemoglobin concentration
was
increased. To see whether clinical benefit accrues from these erythrocyte and hematological
changes will require a Phase III clinical trial (Ataga et al., 2008).
2.13.3.3 Inhibition of other channels
Movement of K+ via the Gardos channel requires the parallel
movement of Cl
– anions to
maintain electroneutrality. High-affinity blockers of Cl
– conductance can reversibly
block
human erythrocyte chloride conductance in vitro without directly affecting the Gardos
channel or the K+-Cl
– co-transport.
In sickle mice treated with a Cl
– conductance inhibitor,

35
packed cell volume (PCV) increased and mean corpuscular hemoglobin
concentration
(MCHC) decreased with an increase in cell K+. A selective loss of the densest erythrocytes,
with a shift from sickled to well-hydrated discoid erythrocytes, was seen.
Clinical trials of this
class of agent have not been done (Buchanan et al., 2004).
2.13.4 Anti-Adherence Therapy
Anti-adherence therapy for sickle cell disease targets the abnormal interactions among
erythrocytes, endothelial cells, leukocytes and platelets that are part of the pathophysiology of
the disease process. Potential anti-adherence agents have been studied in
acute painful events,
where, through poorly understood mechanisms, they restore microvascular circulation and
improve tissue ischemia. In Phase II studies of a non-ionic surfactant copolymer, poloxamer
188, this agent reduced the duration and increased the resolution of acute painful episodes, an
effect especially notable in children
less than 15 years old and in patients receiving
hydroxyurea (Orringer et al., 2001). Whether poloxamer 188 exerts its effects by modifying
interactions of sickle cells or other blood cells to endothelium is not known.
Its clinical
effectiveness as a single agent to treat or prevent vasoocclusive complications is, at best,
modest, and currently no additional trials of this agent are on-going.
Endothelium-dependent vasodilation is disturbed in sickle cell disease (Eberhardt et al., 2003)
and superoxide generation induces chronic inflammation
that may be inhibited by
apolipoprotein (apo) A-1 mimetics.
L-4F, an apoA-1 mimetic, inhibited superoxide
production and improved vasodilation in sickle mice (Ou et al., 2003) While a mechanism of
action is not yet totally clear, this agent may preserve endothelial function and endothelial
nitric oxide synthase (eNOS) activity. This type of drug may have widespread application in
vascular diseases including sickle cell anemia (Ansell et al., 2003). Most agents that might

36
disrupt the adhesive interactions and inflammation
hypothesized to presage sickle
vasoocclusion have not been studied clinically.
2.13.5 Nitric Oxide
A potent vasodilator, NO is an important regulator of vascular tone and, because of its
interaction with hemoglobin, blood vessels and blood cells, has been hypothesized to have
several advantageous effects in sickle cell disease. Generated from
L-arginine by NO
synthases, NO activates soluble guanylate cyclise (sGC), to produce the second messenger,
cGMP (Buchanan et al., 2004). Nitric oxide inhibits the adhesive interactions
among
platelets, leukocytes and sickle erythrocytes (Hankins and Aygun, 2009). It also decreases
vascular cell adhesion molecule-1 (VCAM-1) expression in endothelial cells.
While a
biologically sound rationale exists for using NO in the acute chest syndrome and pulmonary
hypertension of sickle cell disease, a controlled trial of this treatment has not been reported. It
should be remembered that this approach could be deleterious as NO can be metabolized
to
damaging oxidants like nitrite (NO2–) and peroxynitrite
(ONOO
–). In a clinical trial, inhaled
NO was associated with a small reduction in pain score and opioid use in children
with acute
painful episodes (Weiner et al., 2003).
2.14 Unorthodox Treatment
In Nigeria, herbal medications are used as alternatives to orthodox western medicine due to
the perceived minimal side-effects, affordability and easy of acquisition (Ogunkunle and
Ladejobi, 2006; Okigbo et al., 2009). Medicinal plants can be described as plants whose part
/ organ contains a substance that can be used for therapeutic purposes or as a precursor for the
synthesis of antimicrobials or other useful drugs (Sofowora, 1982). World Health
Organization (W.H.O.) as cited by Okigbo et al. (2009) defined medicinal plants as herbal

37
preparations produced from chemical, physical, or biological processes which may be used
for immediate consumption or as a basis of herbal products. Medicinal and aromatic plants
used in such herbal medications contain biologically active substances (secondary
metabolites) such as saponins, tannins, flavonoids, alkaloids and other chemical substances
which have curative properties (Okigbo et al., 2009). Most of these phytochemical
constituents formed during normal metabolic processes are potent bioactive compounds
found in specific plant parts, thus many plant components are used in preparing
pharmaceutical products for medical treatments. It has also been noted that most treatments
for diseases and infections in developing regions such as Africa largely depend on herbal
preparations (Okigbo et al., 2009). WHO estimates that 80% of the world population,
approximately 4 billion people, uses herbal medicine for primary health care (Macedo et al.,
2008). Plant materials thus play a major role in primary health care as therapeutics in many
developing countries. Herbal medicine has a long and respected history as the oldest form of
healthcare to mankind being used worldwide in various forms.
The use of phytomaterials such as Piper guinensis, Pterocarpa osun, Eugenia caryophylla
and Sorghum bicolor extracts for the treatment of SCA has been reported by Wabembe et al.
(2001). The extracts of Pterocarpus santolinoides and Aloe vera were reported to increase the
polymerisation time of Hbf and inhibit sickling in vitro (Ugbor, 2006). Sofowora and Isaac-
Sodeye (1971) reported the reversal of sickling by root extracts of Fagara zanthoxylloides.
Terminalia catappa could be effective antisickling agents that inhibit osmotically induced
haemolysis of human erythrocytes (Mgbemene and Ohiri, 1999). The aqueous extract of
Gracinia kola has been described by Elekwa et al.(2003) as having a higher membrane
stabilising effect than phenylalanine, while 2 – hydroxybenzoic acid has been identified as
the antisickling agent in aqeous extract of Zanthoxylum macrophylla roots (Elekwa et al.,

38
2005). The in vitro HbS gelling time was reportedly increased by the extract of Pterocarpus
santolinoides and Aloe vera (Mohanty et al., 2012). Antisickling activity of twelve Congolese
plants namely Alchornea cordifolia, Afromomum albo violaceum, Annona senegalensis,
Cymbopogon densiflorus, Bridelia ferruginea, Ceiba pentandra, Morinda lucida,
Hymenocardia acida, Coleus kilimandcharis, Dacryodes edulis, Caloncoba welwithsii, and
Vigna unguiculata have been reported (Mpiana et al., 2007). It has been suggested that
Cajanus cajan seeds (Onah et al., 2002), aged garlic (Ohnishi and Ohnishi, 2001; Moriguchi
et al., 2001), unripe Carica papaya fruit (Oduola et al., 2006; Thomas and Ajani, 1987),
leaves (Imaga et al., 2009), the roots of Cssus populnea L. CPK (Moody et al., 2003), Khaya
senegalensis (Fall et al., 1999 ), Scoparia dulcis (Ahmed and Jakupovic, 1990) as well as
Parquetina nigrescens (Wambabe et al., 2005) have various degree of antisickling activities.
The Nigerian Zanthoxylum has been extensively studied (Sofowora and Isaacs-Sodeye, 1971;
Sofowora, 1974; Sofowora, 1975a; 1975b; Adesanya and Sofowora, 1983). The bioactive
compounds responsible for anti-sickling properties were identified as vanillic acid, p-hydroxy
benzoic acid and p-fluoro benzoic acid. There are some other drugs prepared from medicinal
plants in the market today under various names. The most prominent and widely used of them
all is Ciklavit developed by Prof G Ekeke after eighteen years of intensive research in
collaboration with Neimeth Pharmaceuticals, Lagos, Nigeria. (Okpuzor, et al., 2008)
Research on phytomedicine for the treatment of SCD has led to the development of Nicosan
formerly called Niprisan (herbal based drug) which has been patented by the National
Institute for Pharmaceutical Research and Development (NIPRD), Abuja, Nigeria and
produced to meet increasing global demand by sufferers of SCA. This development indicates
that more of such herbal based drugs could be consequent upon scientific investigations on
plants that are used in folklore medicine.

39
On the understanding that herbal remedies and medicinal plants products from indigenous
flora have long been used in folk medicine in the management of SCA, it appears that proper
and in-depth scientific investigation on such medicinal plants could be of tremendous help in
developing efficacious and safer drugs for SCA treatment.
The species Senna alata (Fabaceae) and Senna podocarpa (Fabaceae) are examples of plants
commonly used in popular Nigerian folk medicine. Both S. alata and S. podocarpa bear a
common Yoruba name „asunwon‟ while S. alata is referred to as „asunwon oyinbo‟, the
suffix meaning exotic or introduced species and S. podocarpa is known as „asunwon gidi‟
the suffix meaning indigenous or native species (Odugbemi, 2006; Ogunkule and Ladejobi,
2006).
2.14.1 Physical Description of Senna alata
Senna alata (Fabaceae) is a plant growing 6-12 feet as a shrub with erect waxy yellow spikes
resembling fat candles before individual blossoms open. The leaves are lanceolate in shape,
bilaterally symmetrical opposed with smooth margins, folding together at night. The leaflets
are 8-20 in number arranged in four pairs. The buds of the plant are rounded with five
overlapping sepals, five free or less equal petals narrowed at the base. Fruits are winged pods
with seeds small and square. It is found in the tropics (West Africa, Islands of Central and
South Pacific, South America and Australia) in secondary vegetations or along riverbanks. It
is commonly referred to as Ringworm Cassia, Candle Bush, Candelabra Bush, Empress
Candle Plant, Ringworm Tree or “Candletree”. Its conservation status is secure. In Southwest
Nigeria it is referred to as “Asunwon oyingbo” in the Yoruba language for its exotic origin;
Crawcraw or Ringworm plant for its use in skin diseases (Odugbemi, 2006; Ogunkunle and
Ladejobi, 2006).

40
2.14.1.1 Medicinal Attributes of Senna alata
Senna alata is a widely recognized medicinal plant amongst herbal traditional practitioners
(Ogunkunle and Ladejobi, 2006). It is used in form of poultice, decoctions with plant parts
such as leaves, stem, bark and pods used in preparations and are widely believed to be
applicable in skin diseases, dysentery, abortifacient, antihelminthic, itch, ringworm, measles,
eczema, bronchitis, black menstruation, venereal disease and stomach ache (Odugbemi,
2006). Other members of the Caesalpinioideae subfamily are recognized as well for their
medicinal properties-Senna podocarpa, Cassia fistula, S. tora, C. obtustifolia, C. nigricans,
C.acutifolia, C. sieberiana, C. occidentalis, C. angustifolia, C. acutifolia, C. spectabilis (Ayo
and Amupitan, 2007). For example, S. tora has been noted for its anti-microbial and anti-
oxidant abilities (Uddin et al., 2008). Out of the thirty-three species found in Nigeria, Senna
alata has been noted for its use in the treatment of skin ailments and its use as a laxative or
purgative with anti-microbial action against dermatophyte bacterial and fungal infections
(Owoyale et al., 2005; Ayo and Amupitan, 2007). It has been stated that all the plant parts
can be used as an oral laxative due to its anthraquinone content (Owoyale et al., 2005).
The leaves of Senna alata are used as infusions to treat constipation or poultice to treat
oedema and skin disease (Okigbo et al., 2009). The leaf sap has been used to treat ringworm,
scabies, ulcers, swellings, inflammation and skin parasites (Owoyale et al., 2005; Pieme et
al., 2006). The decoction of the leaf, flower bark and wood has been used in treating eczema
and allergy and the decoction of the leaf flower only has been used as an expectorant in
bronchitis, dyspnoea and as an astringent and mouthwash (Pieme et al., 2006; Idu et al.,
2006). The methanolic extract has been shown to have a strong growth inhibition against
bacteria and fungi- Mucor sp., Rhizopus sp., Aspergillus niger, Escherichia coli, Bacillus

41
subtilis, Salmonella typhii, Pseudomonas aeruginosa and Staphylococcus aureus (Owoyale et
al., 2005).
Further research has shown that the methanolic extract of S. alata also inhibited the growth of
Microsporum canis, Trichophyton simii, Fusarium solani, Staphylococcus pyogenes,
Corynebacterium diptheriae, Dermatophilus congolensis, Actinomyces bovis, Blastomyces
dermatitidis, Trichophyton mentagrophytes, Candida albicans, Aspergillus flavus (Makinde
et al., 2007). Plants possess primary metabolites and secondary metabolites; some secondary
metabolites formed during normal metabolic processes may be responsible for their
therapeutic ability. Phytochemical analysis of S. alata leaves shows the presence of phenolic
compounds, eugenol, glycosides, saponin, flavonoids, alkaloids, tannins and anthroquinone.
Anti-microbial activity of the methanolic extract of the leaves on the isolates of
Staphylococcus aureus, Escherichia coli, Proteus vulgaris, Pseudomonas aeruginosa,
Bacillus subtilis, Microsporum gypseum and Trichophyton rubrum has been established
(Phongpaichit et al., 2004; Ogunkunle and Ladejobi, 2006; Idu et al., 2006). S. alata has been
found to contain azulene, guiane, 1, 5, 7-trihydroxy-3-methylanthraquinone, rhein (1, 8-
dihydroxyanthraquinone), chrysophanic acid, chrysophenol and aloe-emodin (Hauptmann
and Nazario, 1950; Phongpaichit et al., 2004; Ayo and Amupitan, 2007). Kaempferol is a
flavonoid compound isolated from the methanolic extract of leaves of S. alata plant found to
be responsible for the antioxidant effect of the extract (Panichayupakaranat and Kaewsuwan,
2004). Ethanolic extract of Cassia alata was found to exhibit protective antioxidant activities
in rats exposed to carbon tetrachloride (Wegwu, 2005). The methanolic extract of S. alata
leaves was found to be more effective than the chloroform, petroleum ether or water/aqueous
extracts in anti-microbial activity (Owoyale et al., 2005; Idu et al., 2006; Makinde et al.,

42
2007). Pieme et al. (2006) concluded in their in vivo experiments on mice and rats that the
aqueous ethanolic extract of the leaves of S. alata had low toxicity and was safe to use.
2.14.2 Physical Description of Senna podocarpa
Senna podocarpa (Fabaceae) formally known as Cassia podocarpa is an herbal shrub that
grows up to 5m high with dense racemes of yellow flowers and thin flat pods. It has a
perennial life span and is widely distributed in Africa, and in some part of Asia. S. podocarpa
is found in the savanna forest area of West Africa (Akanmu et al., 2005). The plant is found
locally, on old farmlands in both western and northern parts of Nigeria (Dalziel and
Hutchinson, 1958).
2.14.2.1 Medicinal Attributes of Senna podocarpa
This plant has been used extensively in the folklore medicines for the treatment of a variety
of skin diseases such as scabies, ringworm and eczema (Sofowora, 1986) especially amongst
the Igbo and Yoruba speaking tribes of Nigeria where it is locally known as Ageloo-ogala
and Asunwon gidi, respectively (Adefemi et al., 1998). The leaves have been extensively
used based on their antigonorrehoeal and purgative properties as well as sore-healing remedy
(Elujoba et al., 1994; Akanmu, 1999; WHO, 1999). The virucidal property against Herpes
simplex virus and African swine fever virus and their infections was established by Silva et
al. (1997) Decoction of the leaves is given as a mild laxative and in large doses, it acts as a
purgative (Trease and Evans, 1996). In Liberia folk medicine, amongst the Kpelle people, the
effects of stem bark extracts in vitro and in vivo on microfilaria were examined and found to
possess curative effect (Kilian et al., 1990). According to Gomes et al. (1997) S. podocarpa
is widely used by Fulani traditional healers living in Guinea Bissau to treat veneral diseases.

43
Ehtnobotanical and phytochemical studies of the leaves and pods of S. podocarpa have
identified some of the plant constituents, which include rhein, emodin, chrsophanol, free or
combined anthraquinones (Rai and Obayemi, 1973; Rai and Abdulahi, 1978; Elujoba et al.,
1994). Anthraquinones derivatives (Akinyemi et al., 2000) obtained from the pods and leaves
have been held responsible for the plants‟ laxative as well as purgative activites. Other
phytochemical groups in the leaf extracts of the plants include flavonoids, tannins,
phlobatannins and saponins (Ogunkunle and Ladejobi, 2006)
2.15 Cytotoxicity testing
Sickle cell anemia (SCA) can potentially be cured by gene therapy (Perumbeti and Malik,
2010) and hematopoietic stem cell transplantation (Bernaudin et al., 2007) but neither is
currently applicable to most patients with this disease due to technical reasons, cost and lack
of highly sophisticated medical care necessary to provide these therapies in Nigeria. Over 50
agents have been described that increase γ-globin expression and/or HbF in primary human
erythroid cell cultures or when administered to humans. But most inducing agents, including
histone deacetylase inhibitors, DNA methyltransferase inhibitors and cancer chemotheraphy
drugs may be cytotoxic, and may alter epigenic marks on a genome-wide basis, damage DNA
or suppress erythropoiesis (Mabaera et al., 2008). Hydroxyurea is the only approved drug for
use in humans, though it has provided a major advance in the care of people with SCA, it is
only effective in one – half of SCA patients (Steinberg et al. 1997a) and it produces
significant suppression of blood counts. These and other issues have led to the low rate of
compliance in Africa (Aliyu and Babdok, 2007) and United States (Hampton, 2008).
In view of the shortcomings of current inducing agents, there are no drugs that are effective,
safe and have the ease of use that would make them applicable to most patients with SCA.
There is therefore an urgent need to screen for natural inducers of HbF and identify new types

44
of agents that can induce HbF production with greater efficiency and lower toxicity (Lal and
Vichinsky, 2004). In developing countries, so many constituents of plants capable of diseases
treatment have been investigated by scientists with a view to contributing to the search for
substances that would be effective in the management of the diseases. Crude extracts from
plants have been used in treating many diseases since ancient times, although, the biological
concentration and toxicity of such plant extracts are seldom known. The safety of any
therapeutic agent is of paramount importance and appropriate toxicity assays to evaluate
biological safety of biomaterials are essential. In vitro cytotoxicity assays with cultured cells
are very common because they are rapid, inexpensive and do not have ethical implications
(Ishiyama et al., 1996; Harbell et al., 1997; Fernandes et al., 2005). Several methods for
determining cellular viability and/or growth following in vitro exposure to compounds have
been reported. These methods used well know dyes that develop a colour, allowing a
colorimetric measurement of cell viability.
Cytotoxicity is a technique for studying the effectiveness of substances on the survival and
proliferation of cells before testing them in patients (Carney and Winkler, 1985). The assay
measures the toxicity of a treatment on cells at a wide range of doses and it is frequently used
in cancer research laboratories to determine the effect of drugs on proliferating tumor cells
(Hoffman and Robert, 1991). In vitro cytotoxicity tests are based on the idea of „basal‟
cytotoxicity – that toxic substances affect basic functions of cells which are common to all
cells, and that the toxicity can be measured by assessing cellular damage. The development of
in vitro cytotoxicity assays has been driven by the need to rapidly evaluate the potential
toxicity of large numbers of compounds, to limit animal experimentation whenever possible,
and to carry out tests with small quantities of compound (Barile et al., 1994). Evidence for
the use of in vitro cytotoxicity tests has led many pharmaceutical companies to screen

45
compound libraries to remove potentially toxic compounds early in the drug discovery
process (Davila et al., 1990; Barile et al., 1994 and Todd et al., 1999). Early identification of
toxic effects can help project between chemical series and identify toxicity relationships that
may exist between the chemicals and the cells (Todd et al., 1999).
A reduction in cellular ATP levels or mitochondrial activity is an early indication of cellular
damage. It has been shown that changes in metabolic activity are better indicators of early
cell injury, and that effects on membrane integrity are indicative of more serious injury,
leading to cell death (Davila et al., 1990). The key features of cytotoxicity assay include low
compound consumption and multiple assays. Mitochondrial enzyme, succinate-tetrazolium
reductase, present in viable cells reduces tetrazolium salts to soluble or insoluble coloured
formazan compounds that can be quantified spectrophotometrically. The most commonly
used tetrazolim salt is MTT [3-(4, 5-Dimethylazol-2-yl)-2,5-diphenyltetrazolium bromide],
however, the disadvantage is that the formazan dye produced from MTT is extremely water
insoluble, so an additional extraction step is needed for spectrophotometric quantification,
which can be avoided with the use of WST-1 [2,(4-Iodophenyl)-3-(4-nitrophenyl)-5-(2,4-
disulfophenyl)-2H-tetrazolium]. Since the bio reduction of tetrazolium salt, is a sensitive
chromogenic indicator for NADH produced by metabolically active cells in culture, the
amount of formazan dye formed directly correlates to the number of metabolically active
cells in the culture (Riss and Moravec, 2004).
One parameter for cell death is the integrity of the cell membrane, which can be measured by
the cytoplasmic enzyme activity released by damaged cells (Riss and Moravec, 2004).
Lactate dehydrogenase (LDH) is a stable cytoplasmic enzyme present in all cells. It is rapidly
released into the cell culture supernatant upon damage of the plasma membrane
(Korzeniewski and Callewaert, 1983).The LDH activity is determined in an enzymatic test.

46
The first step is the reduction of NAD+ to NADH/H+ by the LDH catalyzed conversion of
lactate to pyruvate. In a second step, the catalyst (diaphorase) transfers H/H+ from
NADH/H+ to the tetrazolium salt 2-(4-iodophenyl)-3-(4-nitrophenyl)-5-phenyltetrazolium
chloride (INT), which is reduced to a red formazan (Nachlas et al., 1960; Decker and
Lohmann-Matthes, 1988; Lappalainen et al., 1994). Neutral red (3-amino-m-dimethylamino-
2-methylphenazine hydrochloride) has been used previously for the identification of vital
cells in cultures (DeRenzis and Schechtman, 1973). This assay quantifies the number of
viable, uninjured cells after their exposure to toxicants; it is based on the uptake and
subsequent lysosomal accumulation of the supravital dye, neutral red. Quantification of the
dye extracted from the cells has been shown to be linear with cell numbers, both by direct cell
counts and by protein determinations of cell populations (Borenfreund and Puerner, 1985,
1986).
Cell lines are often used for in-vitro cytotoxicity testing, and an experimental model system
employing the human erythroid-like K562 cell line, isolated and characterized by Lozzio and
Lozzio (1975) from a patient with chronic myelogenous leukemia in blast crisis, has been
extensively employed as a very useful in vitro model to study the molecular mechanism(s)
regulating the expression of embryonic and fetal human globin genes (Bianchi et al., 2000),
as well as to determine the therapeutic potential of new differentiation-inducing compounds
(Osit et al., 1997;Bianchi et al., 2001). K562 cells grow in culture as single, undifferentiated,
cells in suspension, with low production of hemoglobins (Bianchi et al., 2007) but when
stimulated by various agents, they respond within few days with a significant increase in the
production of hemoglobins and -globin mRNA.

47
3.0 MATERIALS AND METHODS
3.1.1 Plant Materials
The Cajanus cajan (beans and leaves), Smilax kraussiana (roots), Alchornea cordifolia
(leaves), Senna alata (leaves), Senna podocarpa (leaves) used in this study were sourced,
during the rainy season (July – September) from 2006 - 2007, and authenticated by herbarium
curators at the Forestry Research Institute of Nigeria (FRIN), Ibadan. The voucher specimens
of Senna alata leaves (No.FHI.106929) and Senna podocarpa leaves (No.FHI.106995) were
deposited at FRIN herbarium. “Jana”, an herbal formulation, was graciously provided by
Prof. (Mrs.) Okpuzor of the Department of Cell Biology and Genetics, University of Lagos
3.1.2 Participants
The informed consent of the volunteers (SCA patients) was obtained after counseling in
accordance with the approved University protocols (Apendix 1). Blood was collected from
HbSS patients in steady states who had not been on any herbal therapy for at least six months,
but were on sustainance therapy (folic acid and paludrin). Blood (3ml) from donors was
taken into heparin or EDTA tubes and used within two hours post collection.
3.1.3 Experimental Animals
Male albino rats (94 -110 g) were purchased from the University of Ibadan animal house. The
rats were allowed to acclimatize for 2 weeks at the University of Lagos, Department of Cell
Biology and Genetics animal house experimental facility, provided standard feed for
laboratory rodent, regular tap water ad libitum. All animals had free access to water and
standard rat pellets (Pfizer). Rats were fasted (12-15hrs) prior to the administration of extract.

48
3.2 Preparation of Extracts
The plant materials (200g) were extracted with either 80% ethanol or 80% methanol using a
Soxhlet apparatus, concentrated under reduced pressure to a viscous liquid, freeze dried,
weighed and stored at 4 oC until use. For water extract, 10 g of powdered leaf material was
mixed with boiling water (500 ml) for 15 min. The extract was filtered and evaporated under
vacuum below 70 °C on a rotary evaporator according to Duh and Yen, (1997).
3.3 Preliminary Screening
The hydroethanolic extract of Cajanus cajan (beans and leaves), Smilax kraussiana (roots),
Alchornea cordifolia (leaves), Senna alata (leaves), Senna podocarpa (leaves) as well as Jana
were evaluated for their HBSS erythrocyte membrane stabilizing activity using the osmotic
fragility test (Dacie and Lewis, 1991)
3.4 Phytochemical Screening
The phytochemical compositions of the extracts were evaluated according to the methods of
Trease and Evans (2002) and Harborne (1991).
3.5 Proximate Analysis
3.5.1 Preparation of the plant materials for chemical analyses:
Senna alata leaves and Senna podocarpa leaves were washed with distilled water dried at
room temperature to remove residual moisture, and oven-dried at 55 ºC for 24 hours (Aletor
and Adeogun, 1995; Abuye et al., 2003). The dried leaves were ground into powder using a
blender (MARLEX; CM/L 7371373), and sieved through 20-mesh sieve. The leaf powder
was used for the chemical composition analyses.

49
The methods recommended by the Association of Official Analytical Chemists were used to
determine ash (#942.05), crude lipid (#920.39), crude fibre (#962.09) and nitrogen content
(#984.13). All the proximate values were reported in % (AOAC, 1990).
3.5.2 Determination of Moisture Content
Moisture content of the leaf of the two Senna species were determined by drying leaf samples
at 105oC in a hot air oven to constant weight (3 hrs) (AOAC, 1990).
3.5.3 Determination of Ash Content
Two gram aliquot of the sample was placed in a pre-weighed crucible; this was heated on a
hot plate until no smoke was visible. The residue containing crucible was transferred into a
muffle furnace set at 550oC for 3 hours after which the ash containing crucible was cooled in
a dessicator and weighed. The weight of the ash containing crucible was subtracted from that
of the pre-weighed crucible to obtain the ash content. (AOAC, 1990)
3.5.4 Determination of crude lipid and crude fibre content:
Two grams of dried leaves were weighed in a porous thimble of a Soxhlet apparatus. The
thimble was placed in an extraction chamber, which was suspended above a pre-weighed
receiving flask containing petroleum ether (b.p.40-60ºC). The flask was heated on a heating
mantle for eight hours to extract the crude lipid. After the extraction, the thimble was
removed from the Soxhlet apparatus and the solvent distilled off. The flask containing the
crude lipid was heated in the oven at 100ºC for 30 min to evaporate the solvent, then cooled
in a desiccator, and reweighed. The difference in weight was expressed as percentage crude
lipid content. Crude fibre was estimated by acid-base digestion with 1.25 % H2SO4 (prepared
by diluting 7.2 ml of 94 % conc. acid of specific gravity 1.835 g ml-1
per 1000 ml distilled
water) and 1.25 % NaOH (12.5 g per 1000 ml distilled water) solutions. The residue after

50
crude lipid extraction was placed in a 600 ml beaker and 200 ml of boiling 1.25 % H2SO4
added. The contents were boiled for 30 minutes, cooled, filtered through a filter paper and the
residue washed three times with 50 ml aliquots of boiling water. The washed residue was
returned to the original beaker and further digested by boiling in 200 ml of 1.25 % NaOH for
30 minutes. The digest was filtered to obtain the residue. This was washed three times with
50 ml aliquots of boiling water and finally with 25 ml ethanol. The washed residue was dried
in an oven at 130ºC to constant weight and cooled in a dessicator. The residue was scraped
into a pre–weighed porcelain crucible, weighed, ashed at 550ºC for two hours, cooled in a
dessicator and reweighed. Crude fibre content was expressed as percentage loss in weight on
ignition, (AOAC, 1990; Nesamvuni et al., 2001).
3.5.5 Determination of nitrogen content and estimation of crude protein:
Macro-Kjeldahl method was used to determine the nitrogen content, which is the major
component of amino acid, a precursor for protein. Two grams of dried leaves were digested
in a 100 ml Kjeldahl digestion flask by boiling with 10 ml of concentrated tetraoxosulphate
(VI) acid (H2SO4) and a Kjeldahl digestion tablet (a catalyst) until the mixture was clear. The
digest was filtered into a 100 ml volumetric flask and the solution made up to 100 ml with
distilled water. Ammonia in the digest was steam distilled from 10ml of the digest to which
had been added 20 ml of 45 % sodium hydroxide solution. The ammonia liberated was
collected in 50 ml of 20 % boric acid solution containing a mixed indicator. Ammonia was
estimated by titrating with standard 0.01 M HCl solution. Blank determination was carried
out in a similar manner. Crude protein was estimated using the following formula:
% Crude protein = % Nitrogen content X 6.25 (AOAC, 1990).

51
3.5.6 Determination of carbohydrate and energy values:
Available carbohydrate was estimated, by subtracting the total sum of percent crude protein,
crude lipid, crude fibre and ash from 100 % dry weight of the plant. The plant calorific value
(kJ) was estimated by multiplying the percentages of crude protein, crude lipid and
carbohydrate by the factors 16.7, 37.7 and 16.7 respectively and adding up the products.
(AOAC, 1990).
3.6 Invitro Studies
3.6.1 Effect of Extract on HbSS Erythrocytes
3.6.1.1 Determination of Osmotic Fragility
Osmotic fragility of red cells was determined according to the modified method of Dacie and
Lewis (1991). A 10% buffered saline solution was prepared by dissolving 45g NaCl, 6.828g
Na2HPO4 and 0.935g NaH2PO4 in 500 ml of distilled water. A 1 % buffered saline solution
was then prepared and its pH adjusted to 7.4 using a pH meter. Graded concentrations of
buffered saline solution (0.00 - 0.85 %) were prepared and 5.0 ml of each concentration was
dispensed into prelabelled tubes (control), while 4.0 ml of each concentration was dispensed
into another set of eight tubes and 1.0 ml of buffered extract was added to each of the tubes
(experimental). A drop (0.02 ml) of heparinised whole blood was then added to each tube and
mixed thoroughly by five gently inversions, then incubated at room temperature for 30 mins
after which the contents of the tubes was again mixed and centrifuged for five minutes at
2500 rpm. The supernatant was decanted into cuvettes and the optical density read at a
wavelength of 540 nm using the supernatant from the 0.85 % tube as blank for each set of
tubes. Lysis was recorded as a percentage (optical density of the solution of tube without
NaCl was taken as 100 % hemolysis), and plotted against percentage sodium chloride to

52
obtain a fragility curve. The mean corpuscular fragility (MCF) index, which is the
concentration of saline solution (NaCl g/L) that caused 50% lysis of erythrocytes, was
interpolated from the plot and the % membrane stabilisation was calculated using the
following formula : % Membrane Stability = (MCFcontrol – MCFtest / MCFcontrol)*100.
(Okpuzor and Adebesin, 2006; Chikezie and Uwakwe, 2011)
3.6.1.2 Activity of the Extracts on Haemoglobin Polymerisation
The venous blood was washed with incubation media and collected by centrifugation at 2500
rpm for five minutes, until the supernatant was clear. A 10 % or 35% hematocrit was
prepared for antisickling or desickling experiment respectively. All prepared blood samples
were stored at 4oC until use and allowed to attain room temperature before use.
3.6.1.2.1 Antisickling Activity of the Extracts
A 2000 µl aliquot of washed red blood cells (10 % hematocrit) was mixed with 2000 µl of
buffered plant extract (5 mg ml-1
) or control. The mixture was gently shaken and at 30 mins
interval, a 400 µl aliquot was withdrawn and mixed with 600 µl of freshly prepared 2 %
sodium metabisulphite solution in a fresh tube. The resulting mixture was layered with 600µl
of liquid paraffin and incubated at 37oC. A 100 µl aliquot was withdrawn every 35 mins into
a fresh Eppendorf tube, mixed thoroughly with 200 µl of 5 % buffered formalin and then
incubated at room temperature until ready to prepare slides. For the positive controls, namely
NICOSAN (an herbal formulation currently used by SCA patients) (5 mg ml-1
),
parahydroxybenzoic acid (synthetic drug) (5 mg ml-1
) and negative control (sodium chloride)
a 1000 µl aliquot of the washed RBC and 1000 µl of control (NICOSAN /
parahydroxybenzoic acid / sodium chloride) were thoroughly mixed at 37oC and immediately
(zero time) a 200 µl aliquot was withdrawn and at intervals of 30mins thereafter, into fresh

53
Eppendorf tubes, mixed with 300 µl of freshly prepared 2 % sodium metabisulphite solution
and the resulting mixture was further diluted with 500 µl of buffered formalin.
3.6.1.2.2 Desickling Activity of the Extracts
A 2000 µl aliquot of washed red blood cells (35 % hematocrit) was layered with liquid
paraffin and 2000 µl of freshly prepared 2 % sodium metabisulphite solution was added with
the aid of a needle and syringe. The mixture was gently rolled between the palms and
incubated for 2 ½
hours at 37oC with occasional shaking. After incubation a 500 µl aliquot of
the mixture was withdrawn and mixed with 500 µl of Senna alata (5 mg mlˉ¹), Senna
podocarpa (5 mg mlˉ¹), parahydroxybenzoic acid (5 mg mlˉ¹) or physiological saline
solution in a fresh tubes layered with liquid paraffin and preincubated at 37oC. A 100 µl
aliquot was withdrawn every 30 mins into a fresh Eppendorf tube, mixed thoroughly with
200 µl of 5 % buffered formalin and then incubated at room temperature until ready to
prepare slides.
3.6.1.2.3 Preparation of Microscopic Slides
The mixture containing buffered formalin and control was centrifuged at 2000 rpm for 5
minutes and 250 µL of the supernatant was removed. With the aid of a capillary tube, a drop
was taken and placed on a microscope slide. A smear was made and fixed with 95 %
methanol. The slide was stained with Giemsa stain and excess stain was washed off. The
slide was air dried and examined under light microscope. About 100 RBCs were counted
from five different fields.

54
3.6.2 Antioxidant Potential of Extracts
3.6.2.1 Total Antioxidant Capacity
The total antioxidant capacity of the plant extract(s) was determined using the method of
Prieto et al. (1999) with slight modifications. The assay is based on the formation of a green
phosphate- molybdenum (V) complex by the reduction of molybdenum (VI) at acidic pH.
Aliquots of 0.1 ml of each extract or the standard - ascorbic acid (100 µg ml-1
) was mixed
with 3 ml of working reagent (0.6 M sulphuric acid, 28 mM sodium phosphate and 4 mM
ammonium molybdate). The tubes were incubated at 95oC for 90 minutes, cooled to room
temperature and the absorbance of each solution was measured at 695 nm against a blank
(working reagent plus solvent-methanol/water) using a spectrophotometer. The experiment
was performed in triplicates. The total antioxidant capacity was expressed as milligram
equivalent of ascorbic acid.
3.6.2.2 Total Reducing Power
The reducing power of the plant extract(s), gallic acid and ascorbic acid was determined by
assessing the ability to reduce ferric chloride solution (Athukorala et al., 2006). Aliquots of
1ml graded concentration (25-100 µg ml-1
) of the crude extract or standard (gallic acid or
ascorbic acid) was mixed with equal volume of 1% ferriccyanide and 200 mM phosphate
buffer, pH. 6.6. the mixture was incubated at 50oC for 20 minutes. An equal volume of 10 %
trichloroacetic acid was then added and the resulting solution centrifuged at 6000 rpm for 10
minutes. The resulting supernatant, distilled water and 0.1 % ferric chloride were mixed in
the ratio 1:1:2 and the absorbance measured at 700nm. A higher absorbance indicated higher
reducing power

55
3.6.2.3 Total Phenolic Content
The total phenolic content of the plant extract(s) was determined according to the method of
Singleton and colleagues (1999), using the folin-ciocalteau reagent (FCR) which is reduced
by phenols to a mixture of blue oxides with maximal absorbance at around 760 nm. Equal
volumes (100 µl) of plant extract or gallic acid (0-100 µg) were mixed with 500 µl of FCR
and 1.5 ml of 20 % sodium carbonate. The resulting mixture was thoroughly shaken, made up
to 10 ml with distilled water and incubated at room temperature for 2 hours. The absorbance
at 760 nm was determined against a reagent blank. The experiment was performed in
triplicate and the total phenolic content expressed as gallic acid equivalents (GAE)
3.6.2.4 2, 2-Diphenyl-1-picryl-hydrazyl (DPPH) Radical Scavenging Activity
The DPPH radical scavenging activity (DRSA) of the plant extract(s) or standard (ascorbic
acid) was assayed with slight modification of the method of Uddin et al. (2008) which
depends on the production of yellow coloured diphenylhydrazine via reduction of purple
DPPH (maximum absorbance at 517 nm). Graded concentrations (25-100 µl ml-1
) of the
extract or vitamin C were prepared. A 2 ml aliquot of each test solution was then added to 0.5
ml of 1 mM DPPH solution in methanol and the mixture incubated in the dark for 15 minutes
at 37oC, after which the absorbance was measured at 517 nm against a blank containing
methanol and DPPH. This was performed in triplicate. The DPPH radical scavenging activity
was calculated using the equation:
% DRSA = 1- (As /Ab) x100
Where As and Ab are absorbance of the sample and blank respectively. A decrease in
absorbance indicates an increase in DPPH radical scavenging activity.

56
3.6.2.5 DNA Protection Assay
Antioxidant / pro-oxidant activity of crude plant extracts assessed in aqueous system via the
method of hydroxyl radical-induced DNA damage in plasmid pBR322 as described
previously by Russo et al. (2001) with slight modification. To test for DNA damage induced
by hydroxyl radicals, the reaction was conducted in an Eppendorf tube at a total volume of 12
µl containing 0.5 µg pBR322 DNA in 3 µl of 50 mM phosphate buffer (pH 7.4), 3 µl of 2
mM FeSO4 and 2 µl of the hydromethanolic leaf extract of S.alata or S.podocarpa at
concentrations of 10, 100 and 1000 μg ml-1
followed by the addition 4 μl of 30% H2O2. The
resulting mixture was incubated for 60 min at 37 °C. The DNA (super-coiled, linear and open
circular) was analyzed on 1% agarose gels and visualized using ethidium bromide staining.
3.7 In Vivo Studies
3.7.1 Toxicological Studies
3.7.1.1 Organ Toxicity
3.7.1.1.1Treatment of Animals
One hundred and twelve young albino rats (6 – 8 weeks old) were randomly distributed into
seven groups of 16 rats each. Group A which served as control did not receive any dose of
the extract but was administered 1 ml of distilled water daily. Groups B, C, D, received 1 ml
daily dose of 200, 600 and 1000 mg kg-1
body weight extract of Senna alata while groups E,
F, G, received 1 ml daily dose of 200, 600 and 1000 mg kg-1
body weight extract of Senna
podocarpa respectively, for 15, 35 or 63 days. The animals were deprived of food but not
water (8-14 hours) prior to administration of extract. During the period of administration, the
animals were weighed, food and water intake were monitored daily. After 15, 35 or 63 days,
all surviving animals were fasted overnight. Animals were sacrificed after anesthesia

57
(chloroform) by decapitation and blood from each animal was collected separately in labelled
heparinised / EDTA tubes. Selected organs namely the brain, heart, kidney, liver, lungs,
spleen, testes and cauda epididymes were harvested and washed instantly in normal saline.
The washed organs were blotted dry with filter paper, weighed and labelled separately. About
0.2 g of the organs were cut with surgical scissors, kept separately in labelled containers and
frozen (-200C) for further analysis of biochemical parameters. The left over organs were
immediately preserved in 1 % formal saline for histopathological studies.
3.7.1.2 Haematological Parameters
Blood collected in anticoagulant (EDTA) tubes were used to determine populations of white
blood cells (WBC) and red blood cells (RBC) according to Dacie and Lewis, 1991. Packed
cell volume (PCV) was also evaluated.
3.7.1.2.1 Determination of packed cell volume (PCV)/ Haematocrit
PCV was determined by the microhaematocrit method. Haematocirt is the volume of red
blood cells expressed as a percentage of the volume of whole blood in the sample. Lithium
heparinised capillary tube coated with anticoagulant were filled with blood obtained from
EDTA bottles. One end of the capillary tube was sealed with plastercine, and centrifuged for
5 mins at 13000 rpm in a high speed haematocrit centrifuge. The packed cell height of the
sample expressed in percentage (%) was determined using a PCV chart.
3.7.1.2.2 Determination of white blood cell count (WBC)
A white blood cell count estimates the total number of white blood cells in 1 mm3 of blood. A
20 µl aliquot of blood sample was mixed with 380 µl of Turk‟s fluid and the mixture was put
in a Neubauer counting chamber which was subsequently viewed under low power objective

58
(X10) of a light microscope. The four larger corner squares containing uniformly distributed
WBCs were observed and the cells therein counted.
3.7.1.2.3 Determination of differential white blood cell count
A differential white blood cell count provides information on the different types of white
blood cells present in the circulating blood which include neutrophils, lymphocytes,
monocytes, basophils (rarely seen) and eosinophils. Blood from EDTA tube was used to
prepare a thin blood smear on a clean grease free labelled microscope glass slide. The smear
was air dried, fixed with absolute methanol and stained with Leishman stain. The stained
slide was allowed to dry and then examined under the light microscope using the oil
immersion objective.
3.7.1.3 Biochemical Assay
Blood collected into anticoagulant (lithium heparin) tubes were centrifuged at 3000 rpm for
10 minutes at room temperature. The plasma samples were obtained and stored at -40C in
labeled sample bottles until required. The resulting plasma and 10 % liver homogenate was
analyzed to determine the activity of some marker enzymes. Alanine aminotransferase
(ALT), Aspartate aminotransferase (AST) and alkaline phosphatase (ALP) were analyzed by
the method of Obidike et al., 2011 and total protein using the Biuret method. The catalase
and thiobarbituric acid reactives substances (TBARS) levels, (a measure of antioxidant
status) of plasma as well as organ homogenates were also determined according to the
method of Niehaus and Sammuelson, (1968). The 10 % homogenate of selected organs were
also prepared by macerating 1 g of each organ with 9 ml of phosphate buffered saline. The
mixture was centrifuged at 5000 rpm for 5 minutes and the supernatant transferred into
prelabeled Eppendorf tubes.

59
3.7.1.3.1 Determination of biochemical Parameters
3.7.1.3.1.1 Determination of albumin
A 100 µl aliquot of water (blank), plasma or standard (albumin) (100 mg dl-1
) was mixed
with 2500 µl of buffered bromocresol green and incubated for 10 mins at room temperature
after which the absorbance was read against a blank.
Albumin concentration = (OD sample / OD standard) x 100 mg dl-1
3.7.1.3.1.2 Determination of total protein (TP)
Proteins react with cupric ions in alkaline solution to form a purple complex. A 20µl aliquot
of plasma sample, water (blank) or standard (80 g l-1
protein and 0.095 % sodium azide) was
mixed with 1000 µl aliquot of working solution [sodium hydroxide (200 µl), potassium
sodium tartarate (32 mmol l-1
), cupper sulphate and potassium iodide (30 mmol l-1
)],
incubated for 10 mins at room temperature then transferred into cuvettes and the absorbance
of the samples and standard read against the reagent blank within 30mins at 546 nm.
The total protein concentration (g l-1
) = 80 x (OD sample / OD stamdard)
3.7.1.3.2 Determination of enzyme activities
3.7.1.3.2.1 Determination of alanine aminotransferase (ALT)
Alanine aminotransferase catalyse the production of pyruvate from α-ketoglutarate and L-
alanine. Pyruvate, a measure of ALT activity, in the presence of 2,4-dinitrophenylhydrazone
forms pyruvate-hydrazone complex.
A 100 µl aliquot of water/plasma samples was mixed with 500 µl aliquot of working solution
[100 mmol l-1
phosphate buffer (pH 7.4), L-alanine (200 mmol l-1
and α-ketoglutarate (2.0

60
mmol l-1
)] and incubated for exactly 30 mins at 37 oC, then 500 µl of 2,4-
dinitrophenylhydrazine (2 mmol l-1
) was 3added. The resulting solution was further incubated
for 20 mins at room temperature, after which 5000 µl of sodium hydroxide (0.4 mmol l-1
) was
added and the absorbance read against the blank at 546 nm in a spectrophotometer.
3.7.1.3.2.2 Determination of aspartate aminotransferase (AST)
Aspartate aminotransferase catalyse the production of oxaloacetate from α-oxoglutarate and
L-aspartate. oxaloacetate a measure of AST activity, in the presence of 2,4-
dinitrophenylhydrazone forms pyruvate-hydrazone complex.
A 100 µl aliquot of water (blank) or plasma sample was mixed with 500 µl aliquot of
working solution [100 mmol l-1
phosphate buffer (pH 7.4), L-aspartate (200 mmol l-1
) and α-
oxoglutarate (2.0 mmol l-1
)] and incubated for exactly 30 mins at 37 oC, then 500 µl of 2,4-
dinitrophenylhydrazine (2 mmol l-1
) was added. The resulting solution was further incubated
for 20 mins at room temperature, after which 5000 µl of sodium hydroxide (0.4 mmol l-1
) was
added and the absorbance read against a the blank at 546 nm in a spectrophotometer.
3.7.1.3.2.3 Determination of alkaline phosphatase (ALP)
A 500 µl aliquot of plasma sample was mixed rapidly with 3000 µl aliquot of working
reagent (p-nitrophenylphosphate (10 mmol l-1
), diethanolamine buffer (1 mol l-1
, pH. 9.8) and
0.5 mm l-1
magnesium chloride) in a cuvette and the absorbance read at 1 min interval for 3
mins at 405 nm, in a spectrophotometer.

61
3.7.2 Antioxidant studies
3.7.2.1 Determination of Catalase (CAT) activity
Catalase activity was assayed colorimetrically at 620 nm and expressed a µmoles of hydrogen
peroxide consumed per min or hydrogen peroxide consumed per min per mg of protein as
described by Sinha, 1972. The reaction mixture (1.5 ml) contained 1ml 0.01 M phosphate
buffer (pH 7.0), 100 µl of sample and 400 µl of H2O2.
Catalase enzyme activity = (ΔOD/min/E) x (V/v) x 103
Where : ΔOD/min = change in absorbance/min
E = Molar concentration co-efficient (40)
V = Total volume of reacting sample
v = Volume of sample
3.7.2.2 Determination of lipid peroxidation
Lipid peroxidation resulting in the formation of thiobarbituric acid reactive substances
(TBARS) was measured by the method of Rael et al. (2004). A 100 µl aliquot of plasma was
mixed with 2000 µl working reagent (equal volumes of thiobarbituric acid (0.37 %),
trichloroacetic acid (15 %) and 0.25 M HCl). The mixture was boiled for 15 mins in a water
bath, cooled, centrifuged at 1000 rpm for 10 mins at room temperature and the absorbance of
the clear supernatant was measured against a blank at 535 nm in a spectrophotometer.
The TBARS level expressed as malonyldialdehyde (MDA) level (nmol ml-1
)
= (OD/E) x (V/v) x 106
Where : OD = change in absorbance/min
E = Molar concentration co-efficient (40)
V = Total volume of reacting sample

62
v = Volume of sample
3.7.3 Electrophoresis of Red Blood Cell (RBC) Membrane Proteins
Albino rat erythrocyte plasma membranes (ghosts) were prepared using a modification of the
procedure of Johanning and O'dell (1989). Blood obtained from rats dosed daily for 63 days
with the plants extract was collected into EDTA bottles and centrifuged at 2500 rpm for 15
minutes at 4oC. The plasma and buffy coat were then carefully removed using a Pasteur
pipette. Erythrocytes were then washed 3 times in cold 5 mM sodium phosphate / 0.15 M
sodium chloride and centrifuged as before. Careful aspiration was performed after each wash,
sacrificing packed cells at the interphase to ensure removal of the buffy coat. Packed cells (1
mL) were then lysed in 20 mL of cold 0.01% saponin in 5 mM sodium phosphate, pH 8.0.
The mixture was incubated at 4oC for 10 minutes and centrifuged at 16,000 rpm for 20
minutes at 4oC. The resultant deep red supernatant lysates (enriched cytostolic content) were
discarded, leaving a red pellet of packed ghost over a creamy white layer of leukocyte debris.
The ghost was carefully separated from this protease rich layer, by tilting and rotating the
tube to free the ghost and then aspirating it into a new tube containing ice cold 5 mM sodium
phosphate, pH 8.0 and washed severally by centrifuge until an opaque pellet was obtained.
The protein content of the resulting pellet was assayed using the Biurets method and final
pellet was solubilised with 2 volumes of SDS-PAGE sample loading buffer and boiled for 5
minutes. The membrane protein profile was analysed by SDS-polyacrylamide (10 %) gel
electrophoresis under denaturating conditions, using the discontinuous buffer system of
Laemmli (1970). Equal concentrations of protein were loaded per track on each gel.

63
3.8.1 Genotoxicity Studies
3.8.1.1 Genotoxicological Investigation of Extract Using an Animal (Albino Rat) Model
3.8.1.1.1 Sperm-Head Morphology Assay
Male albino rats (35) were divided into seven experimental groups comprising of 5 animals
per group and one group served as control. The control group received by gavage, 1 ml of
water daily for 63 days, while the experimental groups received also by gavage, 1 ml of sub
chronic doses of 200, 600 and 1000 mg kg-1
body weight, corresponding to low, medium and
high doses of each plant extract. After 63 days from the first exposure, the animals were
sacrificed under anaesthesia, and the cauda epididymis removed and 1mg macerated in
prelabelled tubes containing 5 mls of normal saline. A fraction of each suspension was mixed
with 1% eosin Y solution and a smear made on a clean glass slide. The air dried smear slides
were coded (to avoid bias) and scored for the different sperm head morphologies.
3.8.2 CELL LINE STUDIES
The cytotoxic effect of the extracts on the human erythroid-like K562 cell line was
determined using WST-1 [2-(4-Iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-
tetrazolium] cell viability and proliferation assay kit (ScienCell), neutral red uptake assay
(Creppy et al., 2004) and LDH leakage assay (Upur et al., 2008).
3.8.2.1 Cell Culture
Human chronic myelogenous leukemia K562 cells purchased from Cell Line Services
(Germany), were cultured and maintained in a Roswell Park Memorial Institute (RPMI) 1640
medium supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin (Sigma
Aldrich). Cells were grown at 37o C in a humidified atmosphere of 5% CO2/air.

64
3.8.2.2 Cell viability Assay (WST-1)
The hydromethanolic leaf extract of Senna alata and Senna podocarpa were tested for in
vitro cytotoxicity, using K562 cells. Cytotoxicity was measured by the reduction of
tetrazolim salt - WST-1 [2-(4-Iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-
tetrazolium] - to colored formazan compounds by succinate-tetrazolim reductase which exists
in viable cells. Briefly, 50 μl of media (RMPI 1640) was added to wells in the 96-well plates
from row A to row F (triplicate). Then, 100 μl of diluted plant extract (1mg/ml) was added to
the wells on the first column of rows A - F. Starting from the first column the 100 μl of
solution (50 μl + 50 μl media) was mixed and 50 μl transferred to wells on the second column
by using multichannel micro-pipettor and a serial dilution was done up to the tenth column.
Finally, excessive 50 μl from the tenth column were discarded. The final volume for each
well was 50 μl. The cultured K562 cells were resuspended in RPMI 1640 at 1.39 x 104 cells /
ml after verifying cell viability by a trypan blue dye exclusion assay. Then, 50µl aliquot of
the suspension was pipetted into each well, except the 12th
column wells, in the micro-titer
plate containing medium or extract (final volume 100 μl). Finally, 50 μl of medium was
added to the 12th
column wells as medium control, while column eleven wells served as cell
control. Each sample was replicated 3 times and the cells incubated at 37°C in a humidified
5% CO2 incubator for 24 h. After the incubation period, WST-1 reagent (10 μl of 3.26
mg/ml) was added into each well, the plates carefully shaken and the cells incubated for 3h.
The absorbance values of each well at OD450 – OD630 were read using an ELISA Reader
(Molecular Device, Menlo Park, USA). The IC50 values were determined by plotting the drug
concentration versus the survival ratio of the treated cells. Assays were performed at least
three times, and data shown are representative of those assays.
% Cytotoxicity = 1- [(Absorbance of test / Absorbance of control) x 100]

65
A dose-response curve was plotted to enable the calculation of the concentration that killed
50% (CC50) of the K562 cells.
3.8.2.3 Cell Viability (Neutral Red Uptake Assay)
Cells were seeded in 24-well microplates (x104 cells
ml
-1), routinely cultured in a humidified
incubator for 24 h. Cells were maintained in culture and exposed to Senna alata or Senna
podocarpa aqueous methanolic leaf extract as well as nicosan – a herbal drug, over a range
of concentrations (0.0 – 100.0 µg ml–1
). After 24 h exposure to the test sustance, neutral red
(NR) uptake test was performed according to the procedure described by Upur et al. (2008).
Briefly, at the end of the treatment, the medium with or without test substance was discarded
and 200 µl of freshly prepared neutral red solution (50 µg ml
–1) was added to every well
and
re-incubated for an additional 4 h at 37°C. Thereafter, the cells were carefully washed twice
with 200 µl of PBS to eliminate extracellular neutral red. The incorporated dye was eluted
from the cells by adding 200 µl elution medium (50% ethanol supplemented with 1% acetic
acid, v/v) into each well followed by gentle shaking of microplate for 15 min. The plates were
then read at 540 nm using a microplate reader (Molecular Device, Menlo Park, USA). The
percentage of viable cells was calculated and the IC50 was determined and expressed as
microgram per milliliter.
3.8.2.4 Cell Death (Lactate Dehydrogenase Leakage Assay)
Lactate dehydrogenase (LDH) is a soluble cytosolic enzyme, present in most eukaryotic cells,
which is released into the culture medium upon cell death due to plasma membrane damage.
K562 cells (5 x 104 cells ml
–1well
–1) were pre-incubated
in 24-well plates for 24 hours at 5%
CO2–95% air 37°C. Cell viability was assessed by lactate dehydrogenase (LDH) leakage
through the membrane into the medium. Aliquots (150µl) of the cell culture supernatants

66
from control and serial concentration of extract treated cultures were tested after 6 or 7 days
incubation for the presence of LDH using a LDH assay kit (ScienCell).
In this test, three wells
were used for each concentration of
extract, and three independent experiments were
performed.
3.9 DATA ANALYSIS
Statistical Analysis: Data was analyzed using one-way analysis of variance (ANOVA)
followed by Tukey Post Hoc Test, using SPSS 16.0 computer software package (SPSS Inc;
Chicago, U.S.A). Differences at P<0.05 were considered significant.

67
4.0 RESULTS
4.1 PRELIMINARY SCREENING
Five plants namely Cajanus cajan (beans and leaves), Smilax kraussiana (leaves), Alchornea
cordifolia (leaves), Senna alata (leaves), Senna podocarpa (leaves) and Jana, a herbal
formulation were screened for their membrane stabilizing activity, using the osmotic fragility
test.
4.1.1 MEMBRANE STABILISING EFFECT OF AQUEOUS ETHANOLIC
EXTRACT
Membrane stabilization was conferred on HbSS RBC by the hydroethanolic extract of five
medicinal plants namely Cajanus cajan seed (38.34 – 40.18%)(Fig.2) and leaf extract of
(Fig.3) Smilax kraussiana (18.97 – 45.52%), (Fig.4) Alchornea cordifolia ( -67.63 –
62.85%), (Fig.5) Senna alata (34.63 – 44.17%), (Fig.6) Senna podocarpa (34.25 – 67.12%),
(Fig.7) Cajanus cajan (31.85 - 57.53%), as well as (Fig.8) Jana-herbal formula ( 64.34 -
77.26%). Optimal membrane stabilizing concentration of Senna podocarpa and JANA was
1000 µg ml-1
, while 600 µg ml-1
was recorded for the other extracts. Two plants Senna alata
and Senna podocarpa were thus selected for further investigation, since there is no
documented report on these plants in relation to SCA management.

68

69

70
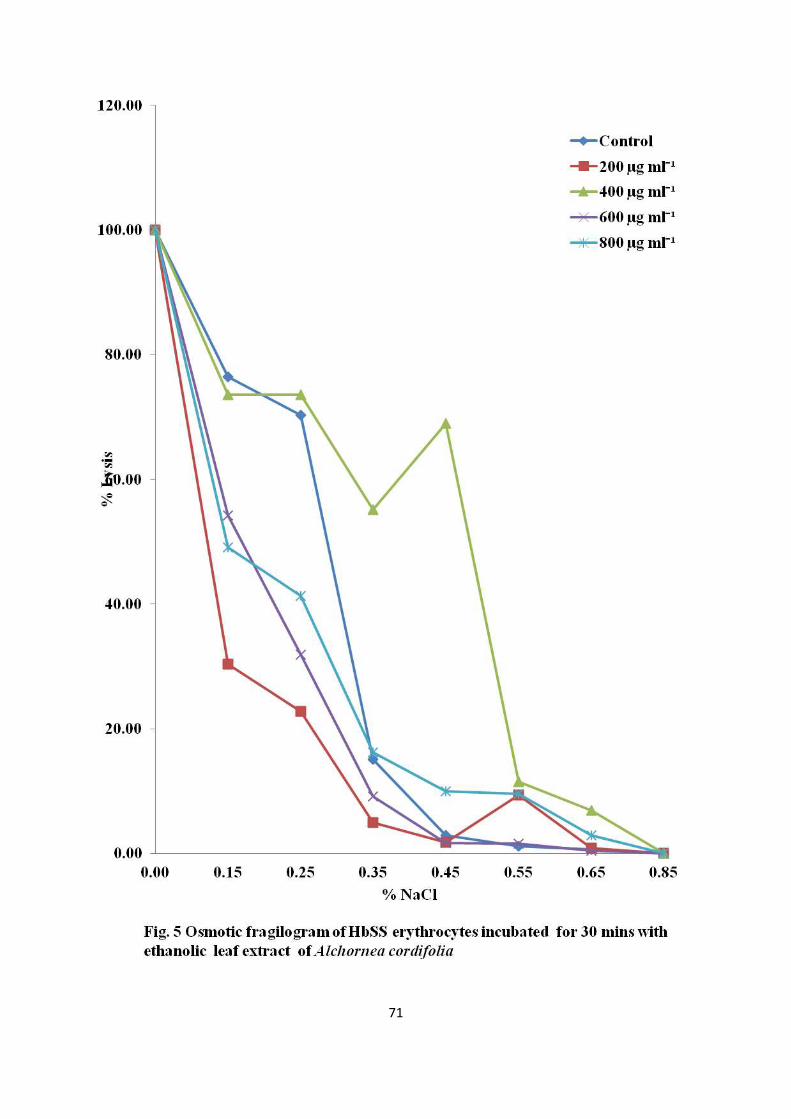
71

72

73

74

75
4.1.2 EXTRACTION YIELD AND PHYTOCHEMICAL COMPOSITION
For S. alata, aqueous methanolic extract gave the highest yield, followed by water extract and
lastly aqueous ethanolic extract (Table 1). On the other hand, S. podocarpa water extract
gave the highest yield, followed by aqueous methanol and then aqueous ethanol.
Senna alata contained keto-sugar, while alkaloid was detected in S.podocarpa and both S.
alata as well as S. podocarpa contained deoxy-sugar (Table 2a). Presence of tanins, saponins,
flavonoids, steroids, anthraquinones and cardiac glycosides were confirmed (Table 2b) in S.
alata and S. podocarpa, while phlobatanin was found only in S. podocarpa

76
Table 1. Percentage yield of plant material(s) after extraction with different solvents
% Yield
Ethanol (80%) Methanol (80%) Water
Alchornea cordifolia
(Leaves) 26.51 ND ND
Cajanus cajan (seeds) 20.32 ND ND
Cajanus cajan (leaves) 22.01 ND ND
Senna alata (leaves) 23.17 27.67 24.03
Senna podocarpa (leaves) 5.5 9.81 13.95
Smilax kraussiana (leaves) 11.9 ND ND

77
Table 2a Phytochemical screening of ethanolic (80%) extracts
TEST AC CCB CCL SA SK SP
Fehlings soln + - + + + -
Fehlings +H2SO4 + Heat + _ + + + -
Dragendoff - - - - - +
Meyers - - - - - -
Barfoed’s (Monosacchs) + - + - - -
Resorcinol (Keto-sugar) ++ + ++ + + -
Phlorogucinol (Pentose) - - - - - -
Keller-Killanni (Deoxy-sugar) + +++ + + + +
Key: - Negative; + positive; ++ more positive; +++ most positive
AC – Alchornea cordifolia leaves;CCB – Cajanus cajan beans;CCL – Cajanus
cajan leaves; SA – Senna alata leaves; SK – Smilax kraussiana leaves; SP – Senna
podocarpa leaves
Table 2b Phytochemical screening of methanolic (80%) extracts
TEST S. alata S. podocarpa
TANIN + +
SAPONIN + +
FLAVONOID + +
PHLOBATANIN - +
STEROIDS + +
TERPENOID - -
ANTHRAQUINONE + +
CARDIAC GLYCOSIDE + +
Key: - Negative; + positive

78
4.1.3 PROXIMATE COMPOSITION
The chemical compositions (Table 3) of the two plants showed no significant difference in
their ash or moisture content. S. alata had significantly higher lipid, protein and crude fibre
contents while the carbohydrate and energy contents of S. podocarpa were significantly
higher.

79
Table 3 Proximate composition of aqueous-methanolic leaf extract of Senna alata and
Senna podocarpa
Parameter S.alata S.podocarpa
%Moisture 5.67±0.14 6.20±0.15
%Lipids 11.2±0.18ᵃ 3.35±0.10
%Protein 5.28±0.09ᵃ 1.97±0.06
%Crude fibre 33.33±0.18ᵃ 20.72±0.59
%Ash 6.61±0.20 6.78±0.16
%Carbohydrate 43.58±0.81 67.18±0.13ᵃ
Energy (Kcal) 295.94± 0.62 306.19±0.97ᵃ
Values (g/100gwt) expressed as percentage mean ± SEM; n=3; superscript (a) shows
significant difference (P<0.05) between means on the same row.

80
4.2 IN VITRO STUDIES
4.2.1 MEMBRANE STABILISING EFFECT OF AQUEOUS METHANOLIC
EXTRACT
The mean corpuscular fragility values of S.alata and S.podocarpa hydromethanolic leaf
extract treated HbSS RBC were significantly (P < 0.05)lower than the untreated HbSS RBC.
The hydromethanolic leaf extract of S.alata (Fig.9) exhibited a membrane stabilizing effect
(44.44 – 66.01%) with a 200 µg ml-1
optimal concentration, whereas hydromethanolic leaf
extract of S.podocarpa (Fig.10) showed a much lower level of membrane stabilization (5.72 –
17.71%) with 800 µg ml-1
supplementation conferring optimal stabilizing effect.

81

82

83
4.2.2 EFFECT OF EXTRACT ON HEMOGLOBIN POLYMERIZATION
The test substances, S.podocarpa, S.alata, parahydroxybenzoic acid and Nicosan, had
significantly higher antisickling activity when compared to the untreated control at all
sampling times (Fig.11). After 120 minutes of incubation, Nicosan exhibited a significantly
(P < 0.05) higher antisickling activity than S.podocarpa as well as parahydroxybenzoic acid,
while after 150 minutes incubation time the antisickling activity of S.podocarpa was
significantly lower than that of S.alata, parahydroxybenzoic acid and Nicosan in that order
(Fig.12).
Pre incubation of HBSS RBCs with S.alata for up to 90 minutes did not significantly alter the
antisickling profile after 175 minutes. Optimal pre incubation time of 30 minutes prior to
deoxygenation conferred an antisickling effect that lasted for 210 minutes (Fig.13). Pre
incubation of HBSS RBCs for 150 minutes with S.podocarpa prior to deoxygenation,
conferred an antisickling effect that lasted for 210 minutes, while no significant (P > 0.05)
changes in antisickling profile was observed for 30, 60, 120 and 180 minutes pre incubation
time (Fig.14).

84
s.podoca
rpa
S.alata
Nicosa
n
PABA
Buffer
ed saline
-300
-200
-100
0
100
Fig.11 Antisickling efficiency of Senna alata, Senna podocarpa, nicosan,
parahydroxybenzoic acid and and buffered saline. Suspension (10% hematocrit) of HbSS
erythrocytes were incubated with test solution (2.5 mg ml-1
) for three hours at 37oC,
aliquots were then withdrawn, deoxygenated with 2% sodium metabisulphite and the
percentage of sickle cells determined. (PABA- Parahydroxybenzoic acid)
TREATMENT GROUP
An
tisic
kli
ng
acti
vit
y (
%)

85
30 60 90 120
150
180
-300
-200
-100
0
100
s.podocarpa
S.alata
PABA
Nicosan
Buffered saline
Fig.12 Antisickling potencies of Senna alata, Senna podocarpa, nicosan,
parahydroxybenzoic acid and and buffered saline. Suspension (10% hematocrit) of HbSS
erythrocytes were incubated with test solution (2.5 mg ml-1
) at 37oC, then aliquots were
withdrawn at various times, deoxygenated with 2% sodium metabisulphite and the
percentage of sickle cells determined. (PABA- Parahydroxybenzoic acid)
An
tisic
kli
ng
Acti
vit
y(%
)

86
-150
-100
-50
0
50
0mins
30mins
60mins
90mins
150mins
180mins
120mins
Time (mins)
Fig.13 Antisickling potency of Senna alata. Suspension (10% hematocrit) of HbSS
erythrocytes was incubated with S.alata (2.5 mg ml-1
) at 37oC, then aliquots were
withdrawn every thirty minutes, deoxygenated with 2% sodium metabisulphite at 37oC, and
the percentage of sickle cells determined at thirty-five (35) minutes interval .
An
tisic
kli
ng
Axti
vit
y(%
)
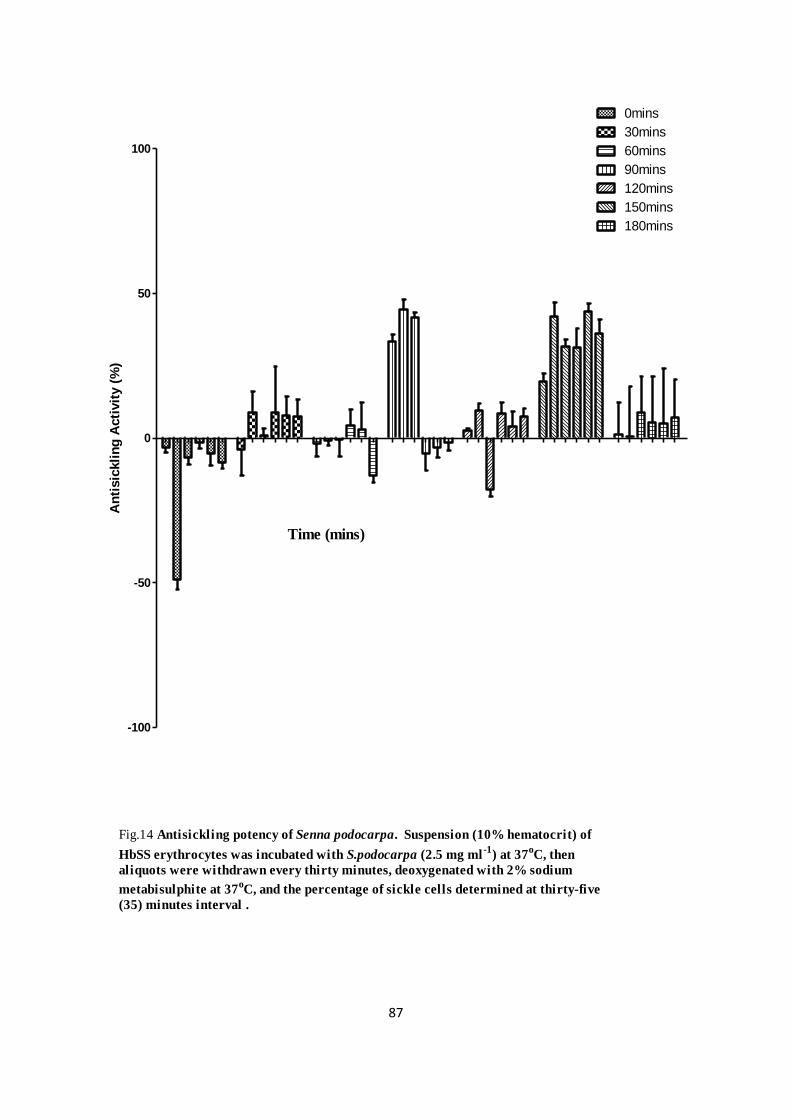
87
-100
-50
0
50
100
0mins
30mins
60mins
90mins
150mins
180mins
120mins
Fig.14 Antisickling potency of Senna podocarpa. Suspension (10% hematocrit) of
HbSS erythrocytes was incubated with S.podocarpa (2.5 mg ml-1
) at 37oC, then
aliquots were withdrawn every thirty minutes, deoxygenated with 2% sodium
metabisulphite at 37oC, and the percentage of sickle cells determined at thirty-five
(35) minutes interval .
Time (mins)
An
tisic
kli
ng
Acti
vit
y (
%)

88
4.2.3 EFFECT OF EXTRACT ON HEMOGLOBIN DEPOLYMERIZATION
A contact time dependent significant (P < 0.01) increase in reversal activity by Senna
podocarpa (2.5 mg ml-1
) was observed (Fig. 15) after 60 mins, though the level of reversal
activity was extremely significantly (P < 0.001) lower when compared with the positive
control parahydroxybenzoic acid (PABA) (2.5 mg ml-1
) and Senna alata (2.5 mg ml-1
). S.
alata showed a highly significantly (P < 0.001) increased contact time dependent reversal
activity when compared to with normal saline, S. podocarpa and PABA

89
30
60
90
120
-50
0
50
100
Senna alata
PABA
NS
senna podocarpa
Fig. 15 Effect of Senna alata (SA), Senna podocarpa (SP), parahydroxybenzoic acid (PABA) and and buffered saline (NS)
on presickled RBC. Suspension (35% hematocrit) of HbSS erythrocytes was layered with liquid parrafin and
incubated with 2% sodium metabisulphite for 21/2
hours at 37oC. After incubation, the test solution was carefully
introduced into the solution under oil, aliquots were then withdrawn at 30 minute intervals, and the percentage of
sickle cells determined. Data expressed as mean sem; ANOVA and Tukey Post Hoc test; *** significantly different
from control (buffered saline) P 0.001
***
***
***
***
***
***
***
***
***
***
***
Time (mins)
Reversal A
ctivity(%
)

90
4.2.4 ANTIOXIDANT POTENTIAL OF EXTRACTS
Table.4 shows that the total phenolic content of the extracts expressed as mg gallic acid
equivalent per gram dry weight of extract was highest in the aqueous-methanolic extract of
S.podocarpa (154±0.34) followed by aqueous-methanolic extract of S.alata (102.21±0.11),
then aqueous extract of S.podocarpa (79.28±0.06) and lastly aqueous extract of S.alata
(68.81±0.03). There were significant differences (p<0.05) amongst the four extracts in total
phenolic compounds content. Both S.podocarpa and S.alata aqueous-methanolic extract had
higher total phenolic content than their corresponding aqueous extracts.
The aqueous extracts (Fig.16) showed poor reducing power, but gallic acid, ascorbic acid,
aqueous-methanolic extracts of S.podocarpa and S.alata, exhibited EC50 (effective
concentration) values of 28.13, 38.02, 49.47 and 61.45 µg ml-1
respectively.
The DPPH radical scavenging activity of the extracts showed a dose-dependent response. The
aqueous-methanolic extract (Fig.17) of S. podocarpa and S.alata had similar IC50 (inhibitory
concentration) values of 64.56 µg ml-1
and 65.16 µg ml-1
respectively which were about five
times less potent than ascorbic acid (13.59 µg ml-1
).

91
Table 4. Total amount of phenolic compounds, free amino acids and total anti oxidant
capacity of aqueous and methanolic (80%) plant extract
Plant Total phenolic
(mgGAE/gplant extract) Free amino acids (mg/100g)
Total antioxidant capacity
(mgAAE/gplant extract)
Methanol
(80%) Aqueous
Methanol
(80%) Aqueous
Methanol
(80%) Aqueous
S.alata 102.21±0.11 68.81±0.03 192±0.12 542.92±0.63 855.58±0.44 298.78±0.38
S.podocarpa 154.85±0.34a 79.28±0.06a 418.31±0.99a 547.74±0.45a 958.19±0.59a 559.74±0.07a
a- significant difference (p<0.05) exist within column

92

93

94
4.2.5 DNA PROTECTION ASSAY
Aqueous-methanolic leaf extract of S.podocarpa exhibited a dose dependent DNA protection
activity, while aqueous-methanolic leaf extract of S.alata at 10 µg ml-1
and 1000 µg ml-1
exhibited pro-oxidant activity, but 100 µg ml-1
showed DNA protection activity (Fig. 18).

95
Fig.18 Effect of aqueous-methanolic leaf extract of S.alata / S.podocarpa on hydroxyl radical
induced DNA damage.
Lanes 1-pBR322; 2-plasmid + FeSO4; 3-plasmid + H2O2; 4-plasmid + FeSO4 + H2O2; 5-7 plasmid +
FeSO4 + H2O2+ S.alata (1000 µg ml-1, 100 µg ml-1, 10 µg ml-1 respectively); 8-10 plasmid + FeSO4 +
H2O2+ S.podocarpa (1000 µg ml-1, 100 µg ml-1, 10 µg ml-1 respectively)
Open Circular
linear
SuperCoil
ed
Direction
Of
Run

96
4.3 IN VIVO STUDIES
4.3.1 Sub acute toxicological effects of S.alata and S.podocarpa on biochemical
parameters of albino rats
Following the sub acute administration of S.alata for 15 days (Fig.19), the AST activity was
significantly (p < 0.05) reduced in the plasma at doses of 200- and 600 mg ml-1
, but at 1000
mg ml-1
dose, it showed a very significant (p < 0.01) reduction in AST activity. A
concomitant, though non significant (p > 0.05) reduction in ALT activity was also revealed.
A significant (p < 0.05) and very significant (p < 0.01) reduction in plasma urea level by 200-
and 600 mg ml-1
of S.alata respectively was observed with no significant (p > 0.05) changes
in the albumin level (Fig.20). S.podocarpa (Fig.21) on the other hand, did not significantly (p
> 0.05) alter the plasma AST and ALT activity at all doses tested, but reduced the urea level
very significantly (p < 0.01) at 1000 mg ml-1
dose over the control while the albumin level
was not significantly (p > 0.05) altered (Fig.22).

97
AST
ALT
0
100
200
300
400C0NTROL
SAM
SAH
SAL
Fig.19 Effect of hydromethanolic extract of Senna alata leaves on activity of selected enzymes in the
plasma of albino rat administered 1ml daily gavage dose of water (CONTROL), 200mg/kg.bwt(SAL),
600mg/kg.bwt(SAM) or 1000mg/kg.bwt(SAH) for 15 days. Data are mean SEM; n=5; ANOVA (P 0.05);
significantly different from control at *(P 0.05) **(P 0.01)
*
*
**
ENZYME
AC
TIV
IT
Y (U
/L
)

98
ALB
UREA
0
20
40
60
80
CONTROL
SAM
SAH
SAL
Fig.20 Effect of hydromethanolic extract of Senna alata leaves on the level of selected analytes in the
plasma of albino rat administered 1ml daily gavage dose of water (CONTROL), 200mg/kg.bwt(SAL),
600mg/kg.bwt(SAM) or 1000mg/kg.bwt(SAH) for 15 days. Data are mean SEM; n=5; ANOVA (P 0.05);
significantly different from control at *(P 0.05) or **(P 0.01)
**
*
ANALYTE
An
alyte level (g
/l);(m
g/d
l)

99
AST
ALT
0
100
200
300
400CONTROL
SPM
SPH
SPL
Fig.21 Effect of hydromethanolic extract of Senna podocarpaa leaves on activity of selected enzymes in
the plasma of albino rat administered 1ml daily gavage dose of water (CONTROL), 200mg/kg.bwt(SAL),
600mg/kg.bwt(SAM) or 1000mg/kg.bwt(SAH) for 15 days. Data are mean SEM; n=5; ANOVA (P 0.05).
ENZYME
AC
TIV
IT
Y (U
/L
)
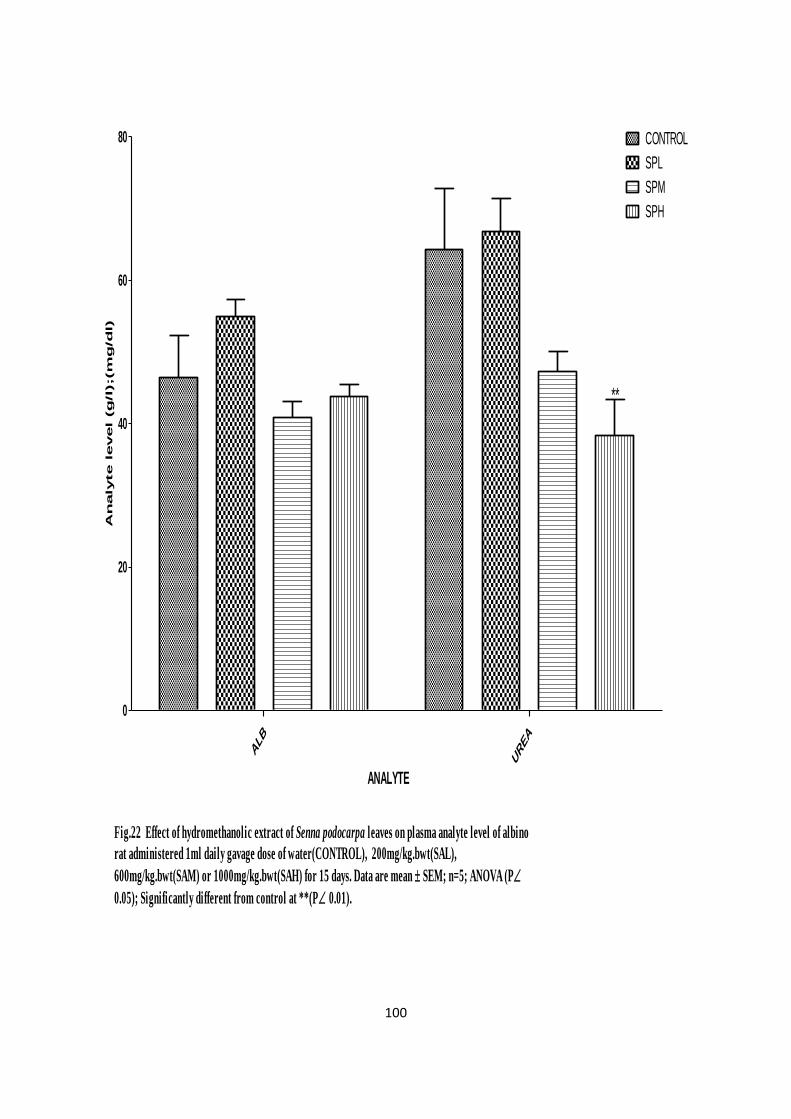
100
ALB
UREA
0
20
40
60
80 CONTROL
SPM
SPH
SPL
Fig.22 Effect of hydromethanolic extract of Senna podocarpa leaves on plasma analyte level of albino
rat administered 1ml daily gavage dose of water(CONTROL), 200mg/kg.bwt(SAL),
600mg/kg.bwt(SAM) or 1000mg/kg.bwt(SAH) for 15 days. Data are mean SEM; n=5; ANOVA (P
0.05); Significantly different from control at **(P 0.01).
**
ANALYTE
An
alyte level (g
/l);(m
g/d
l)

101
4.3.2 Sub chronic toxicological effect S.alata and S.podocarpa on body and
organ weights of albino rats
S.alata at 200 mg kg.bwt-1
significantly (p < 0.05) increased weight of the animals (Fig.23).
The weight of liver relative to the body weight (Fig.24) was significantly increased by 600
mg kg.bwt-1
of S.alata, while no significant changes in the relative weights of other organs
was noticed when compared to the control. The high dose (1000 mg kg.bwt-1
) of S.alata
induced a very significant (p < 0.01) increase in the liver weight relative to brain weight
(Fig.25) when compared to the control.
Senna podocarpa (Fig.26) at 200 mg kg.bwt-1
dose significantly (p < 0.05) increased liver
weight and concomitantly decreased the kidney weight, with a highly significant (p < 0.01)
increase in the weight of the lungs and testes when compared with the control. The 600 mg
kg.bwt-1
dose however, induced a high significant (p < 0.01) increase and decrease in lungs
and testicular weights respectively. The liver and lungs weight was significantly (p < 0.05)
increased and decreased respectively by 1000 mg kg.bwt-1
dose of S.podocarpa when
compared with the control. The liver weight expressed as fraction of the brain weight (Fig.27)
was very highly significantly (p < 0.01) increased over the control in the 1000 mg kg.bwt-1
dose group.

102
CONTROL
SAL
SAM
SAH
SPL
SPM
SPH
0
10
20
30
40
50
*
Fig.23 Effect of Senna alata (SA)/Senna podocarpa (SP) hydro-methanolic leaf extract on
live weight of Albino rats administered 1ml daily gavage dose of 200mg/kg.bwt(L),
600mg/kg.bwt(M) or 1000mg/kg.bwt (H) for 35 days
Dose Group
% B
OD
Y W
EIG
HT
CH
AN
GE

103
BRAIN
HEART
KID
NEY
LIVER
LUNGS
SPLE
EN
TESTE
S
0
1
2
3
4
5CONTROL
SAL
SAM
Fig.24 Effect of Senna alata (SA-)hydro-methanolic leaf extract on live weight of Albino
rats administered 1ml daily gavage dose of 200mg/kg.bwt(L), 600mg/kg.bwt(M) or
1000mg/kg.bwt (H) for 35 days
*SAH
ORGAN
%B
OD
Y W
EIG
HT

104
HEART
KID
NEY
LIVER
LUNGS
SPLE
EN
TESTE
S
0
100
200
300
400
CONTROL
SAL
SAM**
SAH
Fig.25 Effect of Senna alata (SA-) hydro-methanolic leaf extract on live weight of Albino
rats administered 1ml daily gavage dose of 200mg/kg.bwt(L), 600mg/kg.bwt(M) or
1000mg/kg.bwt (H) for 35 days
ORGAN
% B
RA
IN W
EIG
HT
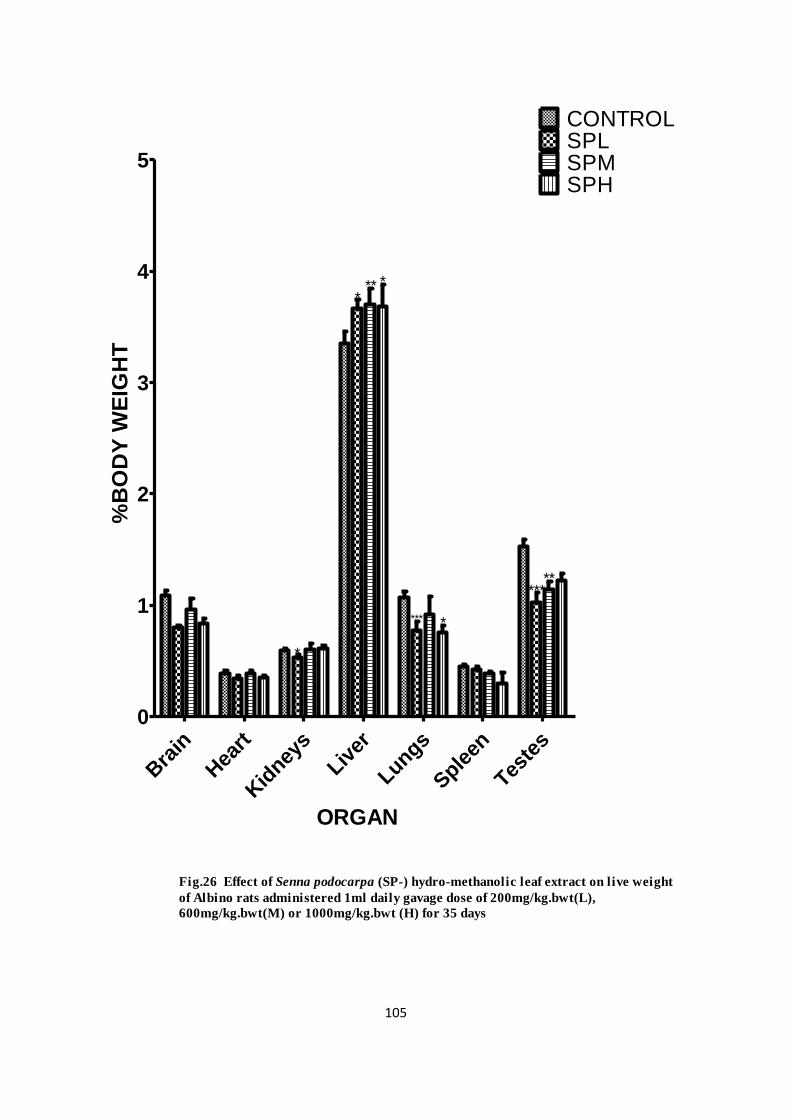
105
Bra
in
Hea
rt
Kid
neys
Liver
Lungs
Sple
en
Teste
s
0
1
2
3
4
5
CONTROLSPLSPMSPH
*
*** *
***
*
*****
Fig.26 Effect of Senna podocarpa (SP-) hydro-methanolic leaf extract on live weight
of Albino rats administered 1ml daily gavage dose of 200mg/kg.bwt(L),
600mg/kg.bwt(M) or 1000mg/kg.bwt (H) for 35 days
ORGAN
%B
OD
Y W
EIG
HT

106
Hea
rt
Kid
neys
Liver
Lungs
Sple
en
Teste
s0
100
200
300
400
500CONTROL
SPL
SPM***
****** SPH
Fig.27 Effect of Senna podocarpa (SP) hydro-methanolic leaf extract on live weight of
Albino rats administered 1ml daily gavage dose of 200mg/kg.bwt(L), 600mg/kg.bwt(M)
or 1000mg/kg.bwt (H) for 35 days
ORGAN
% B
RA
IN W
EIG
HT

107
At nine weeks, daily dosage of S.alata or S.podocarpa (Fig.28) increased with very high
significance (p < 0.001) the weight of the rats at all doses tested, except the 1000 mg kg.bwt-1
dose of S.alata when compared to the control. The liver weight compared to the control was
significantly (p < 0.05) reduced by 600- and 1000 mg kg.bwt-1
dose of S. alata, (Fig.29)
while the liver weight expressed as a fraction of the brain weight (Fig.30) was however
increased very significantly (p < 0.01) and very highly significantly (p < 0.001) at 200- and
1000 mg kg.bwt-1
dose respectively. S. podocarpa (Fig.31) did not significantly (p < 0.05)
induce weight change in the organs at all the tested doses when compared to the control. The
liver weight as a fraction of brain weight (Fig.32) was very significantly (p < 0.01) elevated
over the control by 200- and 600 mg kg.bwt-1
dose of S.podocarpa, while no significant
weight change was noticed in other organs.

108
0
20
40
60
80
100
CONTROLSALSAMSAH
SPMSPH
SPL
Fig.28 Effect of 63 day daily administration 1ml gavage dose of water(Control),
200mg/kg.bwt(-L), 600mg/kg.bwt(M) or 1000mg/kg.bwt(H) of S.alata(SA-) or S.podocarpa(SP-)
hydromethanolic leaf extract on weight change. Values represent mean sem; n=5; ANOVA.
***Significantly different from the control (p0.001)
***
***
***
***
***
Dose Group
% W
eig
ht
Ch
an
ge
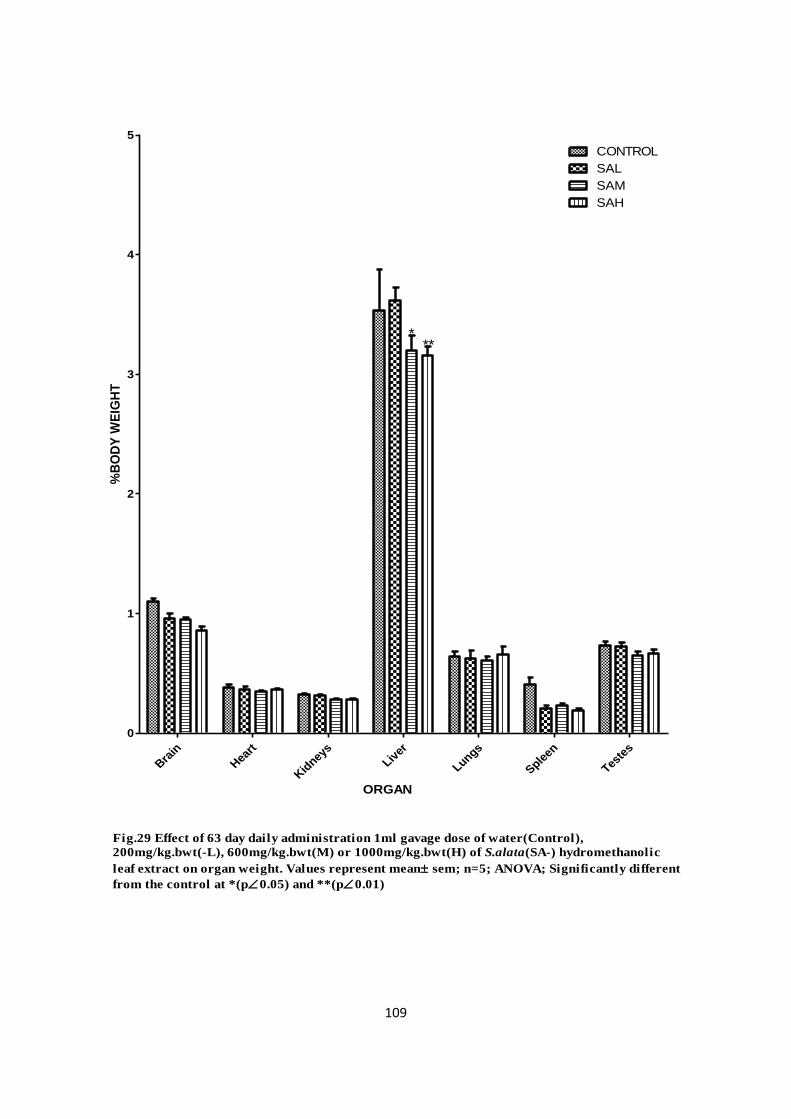
109
Bra
in
Hea
rt
Kid
neys
Liver
Lungs
Spleen
Teste
s
0
1
2
3
4
5
CONTROL
SAL
SAM
SAH
***
Fig.29 Effect of 63 day daily administration 1ml gavage dose of water(Control),
200mg/kg.bwt(-L), 600mg/kg.bwt(M) or 1000mg/kg.bwt(H) of S.alata(SA-) hydromethanolic
leaf extract on organ weight. Values represent mean sem; n=5; ANOVA; Significantly different
from the control at *(p0.05) and **(p0.01)
ORGAN
%B
OD
Y W
EIG
HT

110
Hea
rt
Kid
neys
Liver
Lungs
Sple
en
Teste
s
0
100
200
300
400
500CONTROL
SAL
SAM
***
**
Fig.30 Effect of 63 day daily administration 1ml gavage dose of water(Control),
200mg/kg.bwt(-L), 600mg/kg.bwt(M) or 1000mg/kg.bwt(H) of S.alata(SA-) hydromethanolic
leaf extract on organ weight. Values represent mean sem; n=5; ANOVA; Significantly different
from the control at **(p0.01) and ***(p0.001)
SAH
ORGAN
% B
RA
IN W
EIG
HT

111
Bra
in
Hea
rt
Kid
neys
Liver
Lungs
Sple
en
Teste
s
0
1
2
3
4
5
CONTROL
SPL
SPM
SPH
Fig.31 Effect of 63 day daily administration 1ml gavage dose of water(Control),
200mg/kg.bwt(-L), 600mg/kg.bwt(M) or 1000mg/kg.bwt(H) of S.podocarpa(SP-)
hydromethanolic leaf extract on organ weight. Values represent mean sem; n=5; ANOVA (p
0.05)
ORGAN
%B
OD
Y W
EIG
HT

112
Hea
rt
Kid
neys
Liver
Lungs
Spleen
Teste
s0
100
200
300
400
500CONTROL
SPL
SPM
Fig.32 Effect of 63 day daily administration 1ml gavage dose of water(Control),200mg/kg.bwt(-L), 600mg/kg.bwt(-M) or 1000mg/kg.bwt(-H) of S.podocarpa(SP-)hydromethanolic leaf extract on organ weight. Values represent mean sem; n=5; ANOVA
(p 0.05)
***
****** SPH
ORGAN
% B
RA
IN W
EIG
HT

113
4.3.3 Sub chronic toxicological effect S.alata and S.podocarpa on hematologic
parameters of albino rats
There were no significant (p > 0.05) changes in WBC (total and differential) and platelets
populations (Table 5) between the control and the experimental (Senna alata or Senna
podocarpa) groups, and between the experimental groups. The PCV was however
significantly (p < 0.05) increased in the experimental groups when compared with the
control.

114
Table 5. Effect of nine weeks administration of hydromethanolic leaf extract of Senna
alata and Senna podocarpa on hematologic parameters of albino rats.
HEMATOLOGIC PARAMETER
CONTROL (Water)
200 mg kg.bwt-1 600 mg kg.bwt-1 1000 mg kg.bwt-1
S.alata S.podocarpa S.alata S.podocarpa S.alata S.podocarpa
Hematocrit (%)
31.6±0.68 41.00±1.03* 39.40±2.04* 42.17±0.95* 40.00±1.52* 40.40±1.69* 38.40±0.70*
WBC (x10³) (x103/mm3)
6.04±3.31 7.33±6.23 6.64±7.54 6.32±5.18 6.24±5.87 4.44±1.20 5.70±3.42
Platelets (x103/mm3)
40.20±3.10 44.67±1.41 41.40±1.66 42.17±0.91 34.80±3.37 44.00±1.38 38.20±3.41
Leucocytes (x103/mm3)
58.80±3.81 51.00±1.91 56.80±2.13 54.33±0.99 64.80±3.23 51.00±1.18 61.40±3.70
Monophils (x103/mm3)
0.40±0.4 2.00±0.00* 0.80±0.37 2.00±0.0* 0.00±0.00 1.80±0.37* 0.40±0.40
Eosophils (x103/mm3)
0.60±0.6 2.67±0.61 1.00±0.45 1.50±0.22 0.00±0.00 3.20±0.97* 0.00±0.00
Values are expressed mean ± SEM (n=5); P<0.05 level of significant using ANOVA
followed by tukey multiple comparism test. Asterisk (*) shows that the mean is significantly
different (P<0.05) when compared with the control on the same column.

115
4.3.4 Biochemical effects of S.alata and S.podocarpa on ALT, AST and ALP activity in
albino rats
Table 6 Shows that nine weeks orally administered daily gavage dose of aqueous methanolic
leaf extract of S.alata or S.podocarpa had no significant (P > 0.05) effect on the levels of
ALT and AST activities in the liver at all doses tested compared to the control group, but
200- and 600 mg kg.bwt-1
dose of S.alata caused a significant (P < 0.05) and very highly
significant (P < 0.001) reduction in total protein level respectively. Neither extract
significantly (P > 0.05) altered the activity of plasma ALT, but 1000 mg kg.bwt-1
dose of
S.alata decreased significantly (P < 0.05) the AST activity while ALP activity was
significantly (p < 0.05) increased when rats were treated with 600 mg kg.bwt-1
dose of
S.podocarpa versus the control. Also albumin level was increased, very significantly (P <
0.05) when 1000 mg kg.bwt-1
dose of S.podocarpa was given, while all other treatments were
not significantly (P > 0.05) different from the control.
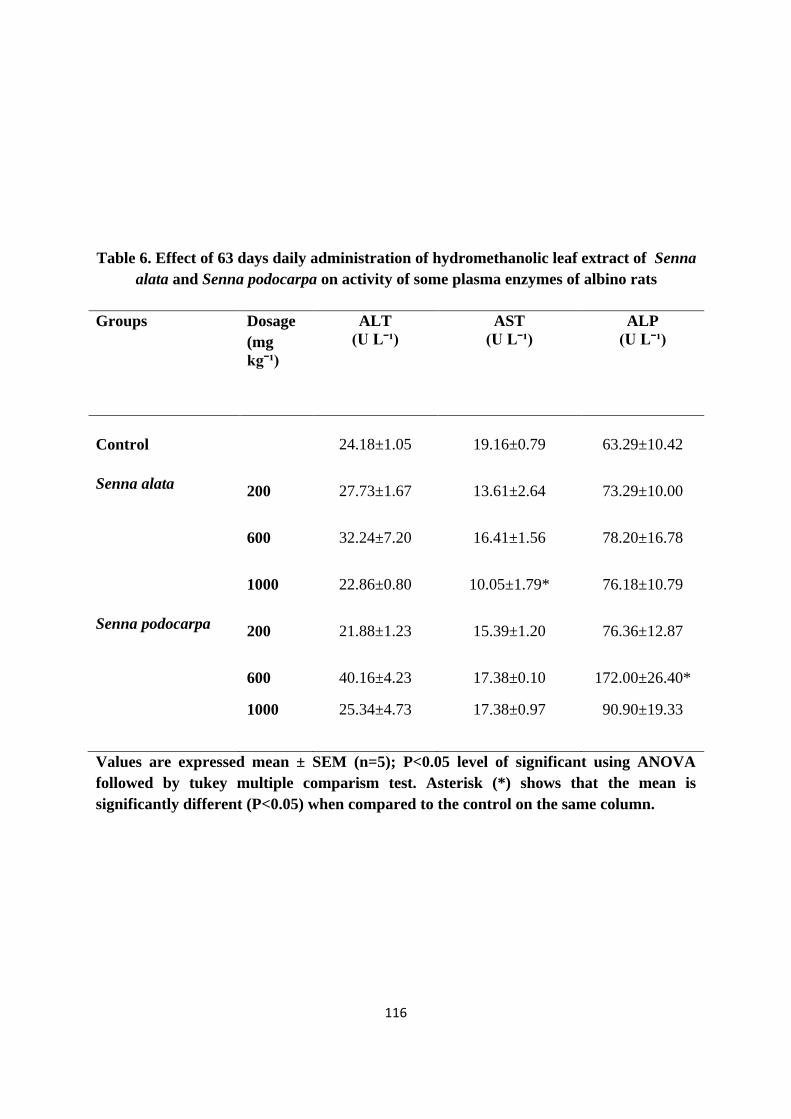
116
Table 6. Effect of 63 days daily administration of hydromethanolic leaf extract of Senna
alata and Senna podocarpa on activity of some plasma enzymes of albino rats
Groups Dosage
(mg
kgˉ¹)
ALT
(U Lˉ¹)
AST
(U Lˉ¹)
ALP
(U Lˉ¹)
Control
24.18±1.05 19.16±0.79 63.29±10.42
Senna alata 200 27.73±1.67 13.61±2.64 73.29±10.00
600 32.24±7.20 16.41±1.56 78.20±16.78
1000 22.86±0.80 10.05±1.79* 76.18±10.79
Senna podocarpa 200 21.88±1.23 15.39±1.20 76.36±12.87
600 40.16±4.23 17.38±0.10 172.00±26.40*
1000 25.34±4.73 17.38±0.97 90.90±19.33
Values are expressed mean ± SEM (n=5); P<0.05 level of significant using ANOVA
followed by tukey multiple comparism test. Asterisk (*) shows that the mean is
significantly different (P<0.05) when compared to the control on the same column.

117
4.3.5 Antioxidant activity of S.alata and S.podocarpa in albino rats
S.alata and S.podocarpa (Table. 7) did not significantly alter plasma catalase activity at all
doses tested when compared to the control, but 200 mg kg.bwt-1
dose of S.alata reduced
significantly (P<0.05) the liver catalase activity. The level of liver lipid peroxidation was
reduced in a very highly significant (P<0.001) manner over the control, by all the tested doses
of S.alata, while S.podocarpa on the other hand had no significant (P < 0.05) effect on the
liver lipid peroxidation level. The level of lipid peroxidation in the plasma was significantly
(P<0.05) reduced over the control by 200 and 1000mg/kg.bwt of dose of S.alata, but
S.podocarpa at 200 and 600mg/kg.bwt caused a very high significant (P<0.001) reduction in
the level of lipid peroxidation in the plasma.

118
Table 7. Antioxidant effect of 63 days daily administration of hydromethanolic leaf
extract of Senna alata and Senna podocarpa on albino rats.
Groups Dosage
(mg kgˉ¹)
CATALASE TBARS
LIVER PLASMA LIVER PLASMA
CONTROL 0.34±0.04 0.084±0.05 11.35±6.26 5.48±4.68
Senna alata 200 0.07±0.10* 0.297±0.10 5.90±3.24* 2.03±1.08*
600 0.08±0.09 0.037±0.40 7.28±3.55* 3.51±1.24
1000 0.05±0.08 0.042±0.25 6.77±7.57* 2.06±1.04*
Senna
podocarpa
200 0.33±0.05 0.024±0.02 10.12±1.31 2.71±4.20*
600 0.40±0.04 0.012±0.00 10.15±0.86 2.25±1.04*
1000 0.17±0.04 0.098±0.04 10.55±1.04 4.52±1.98
Values are expressed mean ± SEM (n=5); P<0.05 level of significant using ANOVA
followed by tukey multiple comparism test. Asterisk (*) shows that the mean is
significantly different (P<0.05) when compared to the control on the same column.

119
4.3.6 Effect of plant extract on albino rat erythrocyte membrane protein profile
The protein profile (Fig. 33) of albino rat RBC of the control and S. alata experimental
groups did not show distinct differences in their banding pattern. No dose related difference
in the albino rat RBC protein profile (Fig. 34) existed between the control and S. podocarpa
experimental groups

120

121

122
4.3.7 Histopathology
Control liver (Plate 1) exhibited dilated central veins, moderate lymphocytes in portal tracts,
while the S. podocarpa group showed (Plate 2) congestion of sinus and hepatic veins with
moderate lymphocytic inflammation. Moderate lymphocytes in the portal tracts with
destruction of limiting plate were noticed (Plate 3) in S.alata group.

123
Plate 1. Control liver with normal parenchyma and dilated central vein (arrowed)
harbouring moderate lymphocytes. Section stained with haematoxylin - eosin (HE)
solution for light microscopy
X 400

124
Plate 2. Senna podocarpa liver Sinusoidal congestion of parenchyma with dialated
central vein (arrowed) and lymphocytic inflammation. Sextion stained with haematoxylin - eosin (HE) solution for light microscopy
X 400

125
Plate 3. senna alata liver moderately congested parenchyma with dialated
central vein (arrowed) habouring lymphocytes. Section stained with
haematoxylin - eosin (HE) solution for light microscopy
X 400

126
4.3.8 Genotoxic potential of plant extract on albino rats
There was no significant (p > 0.05) increase in the average weight of the testis and cauda
epididymis in the hydromethanolic extract fed albino rat groups (Table 8). In the 1000 mg
kg.bwt-1
S.podocarpa extract treated group, increase in testes weight was significant when
expressed as relative weight (organ weight/brain weight).
Figure 35 shows the frequency of occurrence of the different types of spermheads observed in
albino rats exposed to extracts of S.alata and S.podocarpa The mean percentage of normal
sperm ranged from 51% to 97% in all treatment groups, and not different significantly (P <
0.05) among S. alata treated group or between S. alata treated groups compared with control.
However, treatment with S. podocarpa significantly (P < 0.001) reduced the occurrence
(50.8% to 53.33%) of normal sperms in rats, while various morphological (Plate 4) sperm
abnormalities were recorded in the control and the S. alata / S. podocarpa treatment groups.
Significant increase (P < 0.001) in frequency of spermhead abnormalities including pinhead,
wide acrosome and amorphous head were observed in the S. podocarpa treated group, while
the short hook rat spermhead morphology has the highest occurrence (5.21%) in the 1000 mg
kg.bwt-1
S. podocarpa treated group.

127
Table 8 Effects of methanolic extract of Senna alata and Senna podocarpa on testis and
cauda epididymides of albino rats after 9 weeks exposure
TREATMENT
TESTIS CAUDA EPIDIDYMIS A
bso
lute
Weig
ht
%B
od
y w
eigh
t
% B
rain
wei
gh
t
Ab
solu
te W
eig
ht
% B
od
y w
eigh
t
% B
rain
wei
gh
t
CONTROL 0.99±0.07 0.73±0.04 66.78±4.37 0.16±0.02 0.12±0.02 11.09±1.40
Sen
na
ala
ta
200 mg
kg.bwtˉ¹ 1.10±0.04 0.73±0.04 75.91±3.10 0.21±0.02 0.14±0.02 14.35±1.38
600 mg
kg.bwtˉ¹ 1.02±0.02 0.65±0.04 68.89±3.40 0.15±0.01 0.10±0.01 10.43±0.65
1000 mg
kg.bwtˉ¹ 1.15±0.03 0.67±0.04 77.77±2..26 0.18±0.01 0.10±0.01 12.02±0.87
Sen
na
po
do
carp
a
200 mg
kg.bwtˉ¹ 1.07±0.02 0.72±0,02 77.47±5.05 0.17±0.01 0.12±0.01 12.51±1.28
600 mg
kg.bwtˉ¹ 1.02±0.05 0.60±0.03 73.99±2.80 0.20±0.02 0.12±0.00 14.29±1.21
1000 mg
kg.bwtˉ¹ 1.29±0.07 0.67±0.01 79.76
a ±2.24 0.18±0.02 0.09±0.01 11.11±1.12
aSignificantly different from the control (P<0.05)

128
Nor
mal
Sper
m H
ead
Short H
ook
Lon
g H
ook
Pinhea
d
Wid
e ac
roso
me
Am
orphou
s hea
d
0
50
100
150control
sal
sam
sah
spl
spm
sph
Fig.35 Frequency of occurrence of the different types of spermheads induced by 63 days exposure of
albino rats to Senna alata -200/kg.bwt (SAL), -600mg/kg.bwt (SAM), -1000mg/kg.bwt (SAH), Senna
podocarpa – 200mg/kg.bwt (SPL), 600mg/kg.bwt (SPM) or – 1000mg/kg.bwt (SPH). Values represent
mean SEM;ANOVA (P0.05)
SPERMHEAD MORPHOLOGY
FR
EQ
UE
NC
Y (
%)

129

130
4.4 Cytotoxic potential of plant extract on K562 cell line
Figure 36 shows the viability of K562 cells based on WST-1 assay following exposure to
varying concentrations of different extracts. At concentrations below 75 µg ml -1
all the
extracts sustained cell viability. Senna podocarpa, Nicosan and Senna alata had cell
cytotoxicity (CC50) values of 104.5 µg ml -1
, 115.3 µg ml -1
and 131.8 µg ml -1
respectively.
The neutral red uptake assay revealed that the cytotoxic effect of Senna podocarpa on K562
cells (Fig. 37) was not significantly different (p > 0.05) from that of Nicosan at all
concentrations tested, while the cytotoxicity of Senna alata was significantly (P < 0.05) lower
than that of Senna podocarpa and Nicosan at concentrations greater than 64 µg ml -1
. While
the lactate dehydrgenase leakage assay showed (Fig. 38) that, of all the tested concentrations
of the different extracts, only 8.125 µg ml -1
of Senna alata showed significant (P < 0.05)
cytotoxicity.

131
100 200 300 400 500 600
-50
0
50
100Senna podocarpa
Nicosan
Senna alata
FIG. 36 Cytotoxic effect of aqueous methanolic leaf extract of Senna alata, Senna podocarpa and an herbal drug,
Nicosan, on K562 cells. K562 cells grown on 96-well microplates were incubated for 48 hours in RPMI 1640 mediumsupplemented with 1% fetal bovine serum to which was added varying concentrations of extract. Succinatedehydrogenase activity was determined using WST-1 assay kit. Data expressed as mean ± sem.
Concentration (µg ml-1
)
% C
ell C
yto
to
xicity

132
50 100 150
-400
-200
0
200Senna podocarpa
Nicosan
Senna alata
FIG. 37 Cytotoxic effect of aqueous methanolic leaf extract of Senna alata (SA), Sennapodocarpa (SP) and an herbal drug, Nicosan (NIC), on K562 cells. K562 cells grown on 24-wellmicroplates were incubated for 48 hours in RPMI 1640 medium supplemented with 1% fetalbovine serum to which was added varying concentrations of extract. The cells were washed andthen exposed to neutral red for 4 hrs and the quantity of lysosmally accumulated dye wasdetermined. Data expressed as mean ± sem; * significant difference exist between SA and SP ( p< 0.05).
Concentration (µg ml-1
)
Cell
Cyto
to
xic
ity (
%)
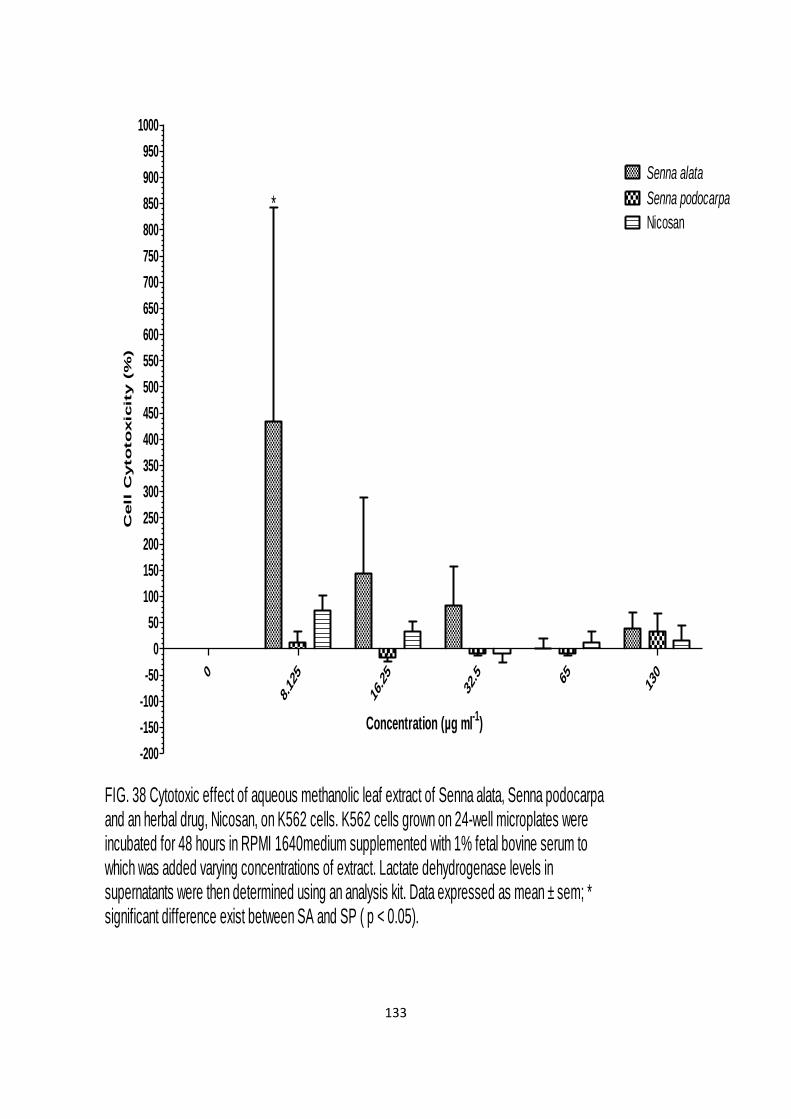
133
0
8.125
16.25
32.5 6
5130
-200
-150
-100
-50
0
50
100
150
200
250
300
350
400
450
500
550
600
650
700
750
800
850
900
950
1000
Senna podocarpa
Nicosan
Senna alata
FIG. 38 Cytotoxic effect of aqueous methanolic leaf extract of Senna alata, Senna podocarpaand an herbal drug, Nicosan, on K562 cells. K562 cells grown on 24-well microplates wereincubated for 48 hours in RPMI 1640medium supplemented with 1% fetal bovine serum towhich was added varying concentrations of extract. Lactate dehydrogenase levels insupernatants were then determined using an analysis kit. Data expressed as mean ± sem; *significant difference exist between SA and SP ( p < 0.05).
*
Concentration (µg ml-1
)
Cell C
yto
to
xicity (%
)

134
5.0 DISCUSSION
The low value of the ash content could be indicative of low mineral value especially the
macro minerals in the leaf of S. alata and S. podocarpa. The crude fat value for S.alata was
moderate, while that for S. podocarpa was high as compared to those of T. triangulare,
Amaranthus hybridus, Calchorus africanum (Akinhunsi and Salawu, 2005).
The increased left shift in osmotic fragilogram of erythrocytes supplemented with the
extracts, suggests conferment of membrane stabilization, with Jana-herbal formula having the
highest, followed by the hydroethanolic leaf extract of Senna podocarpa, Cajanus cajan,
Senna alata, Smilax kraussiana, Alchornea cordifolia, and Cajanus cajan seed in descending
order respectively. Optimal membrane stabilization was achieved by Senna podocarpa and
Jana at highest concentration tested. The fragilogram generated by Alchornea cordifolia was
erratic and showed membrane destabilizing action, while that of S.krussiana at the highest
concentration tested crenated the RBCs. The hydromethanolic leaf extract of S.alata appeared
to be a better membrane stabilizing agent than S.podocarpa, which may be indicative of
higher activity of its extractable secondary compounds composition. Plants are known to
contain compounds which facilitate the stabilization of red blood cell membranes (Smith et
al., 1983) hence the membrane stabilizing effects of these extracts may be due to the presence
of one or more naturally occurring compounds. The attenuating and potentiating effects of the
extracts on membrane stability needs further investigation with a view to understanding the
mechanism involved.
Both plant extracts inhibited the red blood cell sickling process. S.alata appears to be an early
inhibitor while S.podocarpa is a late inhibitor as indicated by the morphological
normalization of HbSS RBCs following treatment with the hydromethanolic extract. This
suggests some influence on the propensity of HbSS RBC to sickle. Although the mechanism

135
of action of has not been elucidated, HbSS erythrocyte membrane stabilization by the plants
extract may play a role in enhancing RBC rehydration rate which would increase the
solubility of deoxy HbS or the extracts may possibly possess protein binding affinity, which
could disrupt formation of tactoid or fibrilary structures due to deoxy HbS polymerization
inside the RBC, thus increasing the concentration of unpolymerised deoxy HbS in solution.
There were significant (P < 0.05) differences amongst the four extracts in total phenolic
compounds content. Both S.podocarpa and S.alata hydromethanolic extract had higher total
phenolic content than their corresponding aqueous extracts. Plant polyphenolic compounds
play an important role in stabilizing lipid oxidation, including radical scavenging properties
and may thus act as primary antioxidant free radical terminators in sickle cell anemia which
can also be viewed as having oxidant etiology. Several studies have described the antioxidant
properties of medicinal plants, foods and beverages which are rich in phenolic compounds
(Krings and Berger, 2001; Wang et. al., 2008).
The aqueous extracts of S.podocarpa and S.alata had a significantly (P < 0.05) higher content
of free amino acids than their hydromethanolic extracts. These plants may serve as a reservoir
of available amino acid when consumed as teas.
The hydromethanolic extracts of S.podocarpa and S.alata showed significantly (P < 0.05)
higher antioxidant capacities than their respective aqueous extracts. The hydromethanolic
extract of S.podocarpa showed highest overall total antioxidant capacity, which may be
attributed to its higher phenolic content. The DPPH radical scavenging activity showed a
dose-dependent response. The hydromethanolic extract of S. podocarpa and S.alata had
similar IC50 values, which were about five times less potent than ascorbic acid. This may be
attributed to its redox properties which allow them act as a reducing agent, hydrogen donors,

136
singlet oxygen quenchers, metal chelating properties. On the other hand aqueous extract of
both plants showed very weak antioxidant activity.
The aqueous leaf extract of S.podocarpa and S.alata showed poor reducing power. Gallic
acid exhibited an EC50 (effective concentration) value < ascorbic acid < hydromethanolic leaf
extract of S.podocarpa. While the hydromethanolic leaf extract of S.alata had the highest
EC50 value. This might be due to the reducing power since a positive correlation existed
between total phenolic content (TPC) and total antioxidant capacity (TAC), which is in
accordance with previous studies (Zheng and Wang, 2001; Unver et al., 2009)
The antioxidative activity of the phenolic compounds may be attributed to their redox
properties which allow them act as reducing agents, hydrogen donors, singlet oxygen
quenchers, metal chelating agents (Huang et al., 2004) or inhibit various oxidizing enzymes
and regenerate endogenous alpha tocopherol in the lipid bilayer (Letelier et al., 2009).
According to Osman et al., (2004) reducing power has a direct positive correlation with
antioxidant properties. Little quantities of different active principles are present in these
extracts which could provoke multiple and synergestic antioxidant effects (Letelier et al.,
2009)
Different values of each antioxidant assay from the same sample are found in various
publications (Surrasmo et al., 2005; Okoro et al., 2010) which could be caused by variation
in geographical location, environmental conditions or the mechanism of action of the
compounds in the natural sample.
The increased DRSA and TAC values of hydromethanolic over aqueous extract could be due
to the solvent of extraction which may have selectively solubilised more phenolic compounds
as shown by the increase in phenolic content. The hydromethanolic extracts contain more

137
apolar compounds than the water extract since increase in the polarity of a compound causes
an increase of solubility of the compound in the apolar phases in which peroxidations occur.
The free radical scavenging property may be one of the mechanisms by which these plants
are effective in traditional medicine. Phenolic compounds may be responsible for antioxidant
properties of many plants (Uddin et al., 2008). Therefore hydromethanolic extract of
S.podocarpa and S. alata can be more effective antioxidants than their water extract.
The results of the DNA protection assay indicate that hydromethanolic leaf extract of
S.podocarpa (at all doses tested) and S.alata (at 100 µg ml-1
) protects DNA against hydroxyl
mediated damage generated by Fenton chemistry. The ability of the extract to protect DNA
can be attributed to its ability to scavenge hydroxyl radicals before
the radicals oxidize DNA,
thus suppressing the formation of linear DNA and induced a partial recovery of super coiled
DNA
Senna alata and Senna podocarpa are medicinal plants widely used in south west Nigeria to
treat various diseases. The application of these herbs in the management of sickle cell anemia
is being sought; hence the toxicity profile of these herbals was investigated. A subchronic (35
day) study of the hydromethanolic leaf extract of these plants at the tested doses did not
appear to retard growth or affect food consumption, indicating that feed intake and utilisation
of protein and other nutrients were not affected by the intake of the herbals.
Analysis of organ weight in toxicology studies is paramount for identification of potentially
harmful effects of chemicals (Sellers, 2007). Anthropometric results revealed a higher body
weights in the group treated for 63 day treatment of rats with S.alata (26.28-68.15% above
controls) and S.podocarpa (59.75-64.62% above controls). Liver weight relative to body
weight was statistically decreased in rats dosed with 600- and 1000 mg kg.bwt-1
of S.alata,
whereas liver weight relative to brain weight was statistically increased in rats dosed with

138
200- and 1000 mg kg.bwt-1
of S.alata. The liver weight effects may be considered test
substance related since according to Bailey et al., (2004) analysis of organ-to-body weight
ratios is predictive for evaluating liver toxicity. Other organ weights relative to body/brain
weight from the different treatment groups did not reveal any statistical differences except the
increase in liver weight relative to brain weight of the 200- and 600 mg kg.bwt-1
S.podocarpa
treated groups
The rats treated with S. alata and S. podocarpa showed no changes in their daily physical and
behavioural activities. The intake of the extract did not affect the functions of the bone
marrow as reflected by the values of WBC (total and differential) and platelets. They were
neither quantitatively nor qualitatively affected. However, the increase in the PCV values
may be taken to be a reflection of an increased RBC volume, indicative of an impairment of
the cellular dehydration pathway, mediated by K+-Cl
- cotransporter (KCC) or Ca
2+- activated
K+ channel (Gardos pathway) (Muzyamba and Gibson, 2003).
It is established that the liver plays a significant role in various metabolic processes, and its
xenobiotic function is vital. Therefore, emphasis was laid on the effect these extracts might
have on the function of the organ, 63 days consumption of either S.alata or S.podocarpa did
not alter its total protein level.
S.alata at 200- and 1000 mg kg.bwt-1
significantly and very significantly reduced plasma
AST activity respectively while 600 mg kg.bwt-1
of S.podocarpa increased plasma ALT
activity. The changes in aspartate (AST) and alanine (ALT) aminotransferase enzyme levels
found in the rats correlate with the relative organ weight changes, though the values were not
double that of control, which is one indicator of biological significance (Rhomberg et al.,
2007). Both enzymes are not liver specific but are also found in skeletal and cardiac muscle
(Thrall, 2004). These findings are therefore not considered toxicologically significant. There

139
were no significant changes in the WBC (total and differential) and platelets counts between
the control and the experimental groups. A significant increase in the RBC counts of the
experimental groups over the control suggests that the hydromethanolic leaf extracts of
S.alata and S.podocarpa may not be toxic as they do not affect leucopoiesis but affected the
circulating erythrocytes and possibly hematopoiesis as reflected by changes in packed cell
volumes (PCV) and eosinophils. The increased PCV may also be a reflection of the hydration
state of the RBCs.
Assessment of TBARS was used to measure plasma and tissue concentrations of
malondialdehyde (MDA), a decomposition product of oxidized lipids, thus serving as an
index of plasma and tissue lipid peroxidation. The increased lipid peroxidation level of the
brain of rats administered 35 day daily dosage of S.alata leaf extract, may be due to either
artifactual ex vivo oxidation (Janero, 1990) or that the extract or its product of metabolism
crossed the blood brain barrier, whereas the reduction in hepatic, renal as well as plasma
TBARS supports the former
Thiobarbituric acid reactive substances (TBARS), as a marker of lipid peroxidation, and
catalase a cellular antioxidant were also estimated in the plasma and liver, of rats after 63 day
administration of plant extract. There was a significant reduction in hepatic (S.alata dose
group) and plasma (both plant extracts) TBARS. Overall result of the TBARS indicated
potential of crude extracts to inhibit oxidation in lipid system. The findings of this study
revealed that the leaf extract of S.alata and S.podocarpa increased the antioxidant capacity of
blood and had an inhibitory effect on the basal level of liver lipid peroxidation, a major
consequence of free radical cell damage, which may alter intrinsic membrane properties.
Natural antioxidants, e.g. plants extract, could also protect erythrocyte membrane from lipid
peroxidation induced by oxo-heme oxidants, by terminating the chain reaction or reacting

140
with free radicals, particularly peroxy radicals, which are major propagators of the auto-
oxidation chain of fat. This lends scientific support to the therapeutic use of the plant leaves
in traditional medicine.
Senna alata and Senna podocarpa leaves are employed traditionally in the treatment of a
variety of disease (either singly or in conjunction with other plants) in southwestern Nigeria.
The sperm head morphological evaluation is considered to be essential for the assessment of
male germ cell toxicity by several regulatory bodies like the Organization for Economic
Cooperation and Development (Trivedi, 2010).
The mechanism by which Senna podocarpa leaf extract caused increased a significant
increase in sperm head abnormalities is not quite clear, but generally damage to the sperm
cell may have occurred via physiological, cytotoxic or genetic means.
Testicular germ cells carrying minor gene mutations are not eliminated but are manifested as
morphologically deformed sperm. It is also documented that certain substances are germ cell
mutagens, affecting gene loci in spermatogonial cells thereby increasing the percentage of
sperm abnormality (Letz, 1990). In mice, a rodent, sperm cell morphology is genetically
controlled by numerous autosomal and sex-linked genes (Krazanowska, 1986), thus, the
abnormal sperm population observed in the Senna podocarpa treated group in this study
maybe due to mutagenic effects of some constituents of these plants extract on specific gene
loci of germ cell chromosomes involved in the maintenance of normal sperm structure. And
since no significant testicular weight loss was recorded in this study, testicular germ cells
may not have been destroyed either due to membrane damage or macromolecular
degradation, although the increased incidence of abnormal sperms is of great concern.

141
Toxicological disturbance of male reproductive function can occur at many sites producing a
range of effects, some primary, and some secondary to the initial insult. Target cells include
testis, epididymis, mature sperm and hormonal regulatory system (Creasy, 2001). Senna
podocarpa exposure could have produced pituitary-hypothalamic or sex hormonal effects
which may have affected spermatogenesis (Bruce and Heddle, 1979; Letz, 1990).
Spermatogonia located within the basal compartment of the testis, is outside the blood-tubule
barrier, hence it is exposed to any toxicant entering the interstitial fluid. Genotoxicants that
gain access to the lumenal compartment (containing spermatocytes and spermatids) may
result in irreversible DNA damage which may manifest as head abnormalities.
The sensitivity of the assays used in determining cell cytotoxicity depends on their endpoints.
The results from the WST-1 assay, indicate that both Senna alata and Senna podocarpa
possessed some active principles which increased cell viability at concentrations lower than
100 µg ml¹, these constituents might have increased the activity of mitochondrial
dehydrogenase, induced the expression of growth stimulation or excerted mitogenic effect.
But since some plant extracts have been found to reduce MTT in the absence of cells
(Shoemaker et al., 2004), which could also have been the case with WST-1. The dose-
dependent cytotoxicity observed with higher concentrations of both plant extracts maybe due
to biologically active alkaloids (Ganapaty et al., 2010) present in Senna podocarpa or
antioxidants which can be oxidised in culture medium with deleterious effect on cells in vitro
(Long et al., 2000; Morita et al., 2003; Halliwell, 2008). The lysosome is regarded as the
garbage disposal unit of the cell and its functionality connotes cell survival. Lysosomal
activity was most severely affected by Senna podocarpa (8.13 µg mlˉ¹) followed by Senna
alata (16.25 µg mlˉ¹) and nicosan (65 µg mlˉ¹). The results from the neutral red uptake assay
revealed that Senna alata at concentrations greater than (16.25 µg mlˉ¹) increased lysosomal

142
activity, while the converse was the case for Senna podocarpa as concentrations greater than
(8.13 µg mlˉ¹) impaired lysosomal activity. K562 cell membrane perturbation leads to
increased extracellular lactate dehydrogenase activity which implies cell cytotoxicity. Senna
alata appeared to cause greater membrane perturbation at lower concentrations than senna
podocarpa probably due to the presence of higher saponin content. The observation in this
study maybe of therapeutic significance, since these plants are used in complementary and
alternative medicine (CAM).
CONCLUSION
The hydromethanolic leaf extracts of Senna alata and Senna podocarpa exhibited appreciable
anti-sickling as well as de-sickling effect. Both plants provide exogenous sources of
antioxidants (possibly in form of phenolic phytochemicals) with remarkable activity, which
may be beneficial if incorporated into supplements formulated for SCA patients, to lower
DNA, membrane and possibly protein oxidative damage. The use of these extracts may help
reduce anemia, with indications that the prolonged use at high concentrations may also
induce infertility due to reduced spermatozoa. Thus the extensive prolonged utilization of
these plants orally in traditional medicine is cautioned when fertility is an issue. The results
of the present research have paved the way to envisaging in vivo studies of these extracts as a
therapeutic agent in patients with SCD and may provide a rational explanation for their use in
managing SCD by Nigerian traditional practitioners.

143
SUMMARY OF FINDINGS
OBJECTIVES
FINDINGS
To evaluate the HbSS RBC membrane stabilising effect of S. alata and S. podocarpa and their infleunce on poly merisation/depolymerisation of mutant HbS
The result indicated a significant reduction in lysis of HbSS RBC under osmotic stress and inhibition/reversal of the sickling process by S. alata and S. podocarpa extracts
To determine the antioxi dant activity of S. alata and S. podocarpa
Hydromethanolic leaf extract of S. alata and S. podocarpa significantly increased the blood antioxidant capacity and had an inhibitory effect on the basal liver lipid peroxidation level
To evaluate the long term toxicological effect(s) of S. alata and S. podocarpa on albino rats
Long term oral administratiion of hydromethanolic leaf extract of S. alata and S. podocarpa appeared toxic
To determine the effect of S. alata and S. podocarpa on RBC membrane protein Profile of albino rats
Both extract did not alter albino rat erythrocyte membrane profile
To evaluate the cytotoxic effect of S. alata and S. Podocarpa on K562 cells
Both extract reduced K562 cell viabilty

144
CONTRIBUTION(S) TO KNOWLEDGE
1. This study has established the membrane stabilizing potential of hydromethanolic leaf
extract of Senna alata and senna podocarpa, via reduction in the median corpuscular fragility
2. Hydromethanolic leaf extract of Senna alata possess greater antisickling activity than that
of Senna podocarpa
3. Hydromethanolic leaf extract of S.alata and S.podocarpa increases the antioxidant capacity
of blood and also suppress the basal level of lipid peroxidation of liver.
4. Hydromethanolic leaf extract of Senna alata and Senna podocarpa functions as an
antianemic agent by increasing the hematocrit level.

145
REFERENCES
Abuye, C., Urga, K., Knapp, H., Selmar, D., Omwega, A. M., Imungi, J. K and Winterhalter,
P. (2003). A compositional study of Moringa stenopetala leaves. East African Medical
Journal. 80:247-252.
Acquaye, C. T. A., Young, J. D., Ellory, J. C., Gorecki, M and Wilcher, M. (1982). Mode of
transport and possible mechanism of action of LPhenylalamine benzylester as an antisickling
agent. Biochimica Biophysica Acta. 693: 407-416.
Adefemi, O.A., Elujoba, A.A. and Odesanmi, W.O. (1988). Evaluation of the toxicity
potential of Cassia podocarpa with reference to official senna. West African
Journal of Pharmacology and Drug Research. 8: 41-48
Ahmed, M and Jakupovic, J. (1990). Ditrepenoids from Scoparia dulcis L. Phytochemistry.
29(9): 3035- 3037
Airy, H. K. (1973). A Dictionary of the flowering Plants and Ferns. 8th
ed. Cambridge,
University Press. London. 3-5.
Ajayi, I. A., Jonathan, S. G., Adewuyi, A. and Oderinde, R. A. (2008). Antimicrobial
Screening of the Essential Oil of Some Herbal Plants from Western Nigeria. World Applied
Science Journal. 3(1): 79-81
Akanmu, M. A. (1999). Evaluation of the toxicity potential of Cassia podocarpa and Cassia
fistula fruits. M.Sc. Thesis Obafemi Awolowo University, Ile-Ife, Nigeria. p19-20.
Akanmu, M. A., Iwalewa, E. O., Elujoba A. A. and Adelusola, K. A. (2005). Toxicity
Potentials of Senna podocarpa (Guillet Perr.) Lock Pods In Rodents. African Journal
Traditional Complementary and Alternative Medicine. 2(3): 274-281.
Akanmu, M. A., Iwalewa, E. O., Elujoba A. A and Adelusola, K. A. (2004). Toxicity
Potentials of Cassia fistula Fruits as Laxative with Reference to Senna. African Journal of
Biomedical Research. 7(1): 23-26.
Akindahunsi, A. A and Salawu, S. O. (2005). Phytochemical Screeninig and nutrient – anti-
nutrient composition of selected tropical green leafy vegetables. African Journal of
Biochemistry. 4: 97-501.
Akinyemi, K. O., Coker, A. O., Bayagbon, C., Oyefolu, A. O. B., Akinside, K. A and
Omonigbehin, E. O. (2000). Antibacterial screening of five Nigerian medicinal plants
against S. typhi and S. paratyphi. J. Nig. Infec. Con. Assoc., 3: 1-4.
Aletor, V. and Adeogun, O. (1995). Chemical analysis of the fruit of Vitex doniana
(Verbenaceae). Food Chemistry. 53: 375-379.
Alexander-Reindorf, C., Nwaneri, R. U., Worrell, R. G., Ogbonna, A. and C. Uzoma, C.
(1990). The significance of gallstones in children with sickle cell anemia. Journal of the
National Medical Association. 82(9): 645–650.

146
Aliyu, Z. Y and Babadok, A. (2007). Hydroxyurea utilization in Nigeria, a lesson in public
health [abstract]. Blood. 2007;110 Abstract 80.
Anderson, K., Crable, S. C., Gallagher, P. G and Joiner, C. H. (2002). Expression of multiple
KCl cotransporter isoforms in sickle and normal erythroid cells.[abstract].Blood
2002;100:666a.
Ansell, B. J., Navab, M., Hama, S., Kamranpour, N., Fonarow, G., Hough, G., Rahmani, S.,
Mottahedeh, R., Dave, R., Reddy, S. T and Fogelman, A. M. (2003).
Inflammatory/antiinflammatory properties of high-density lipoprotein distinguish patients
from control subjects better than high-density lipoprotein cholesterol levels and are favorably
affected by simvastatin treatment. Circulation.108:2751–2756.
AOAC. (1990). Official Methods of Analysis (15th Edition). Helrich, K. Ed; Association of
Official Analytical Chemists, Washington D.C. 43:068-069.
Araba, A. B. (1976). A survey of hematological variables in 600 healthy Nigerians. Nigerian
Medical Journal. 5: 49-53.
Ataga, K. I., Orringer, E. P., Styles, L., Vichinsky, E. P., Swerdlow, P., Davis, G. A.,
Desimone, P. A and Stocker, J. W. (2006). Dose-escalation study of ICA-17043 in patients
with sickle cell disease. Pharmacotherapy. 26:1557–1564.
Ataga, K. I and Orringer, E. U. (2000). Renal abnormalities in sickle cell disease. American
Journal of Hematology. 63: 205-211
Ataga, K. I., Smith, W. R., De Castro, L. M., Swerdlow, P., Saunthararajah, Y., Castro, O.,
Vichinsky, E., Kutlar, A., Orringer, E. P., Rigdon, G. C and Stocker, J. W. (2008). Efficacy
and safety of the Gardos channel blocker, senicapoc (ICA-17043), in patients with sickle cell
anemia. Blood. 111:3991–3997.
Athukorala, Y., Kim, K. N and Jeon, Y. J. (2006). Antiproliferative and antioxidant
properties of an enzymatic hydrolysate from brown alga. Ecklonia cava. Food and Chemical
Toxicology. 44: 1065-1074.
Atweh, G. F, Sutton, M., Nassif, I., Boosalis, V., Dover, G. J., Wright,, E., McMahon, L.,
Stamatoyannopoulos, G., Faller, D. V and Perrine, S. P. (1999). Sustained induction of fetal
hemoglobin by pulse butyrate therapy in sickle cell disease. Blood. 93:1790–1797.
Atweh, G. F and Schechter, A. N. (2003). Pharmacologic induction of fetal hemoglobin:
raising the therapeutic bar in sickle cell disease. Curr Opin Hematol, 8: 123–130.
Atweh, G. F., DeSimone, J., Saunthararajah, Y., Fathallah, H., Weinberg, R. S., Nagel, R. L.,
Fabry, M. E and Adams, R. J. (2003). Hemoglobinopathies. Hematology. 14–39.
Ayo, A. G and Amupitan, J. O. (2007). Phytochemical and Cytotoxic Screening of Leaves of
Cassia nigricans Vahl. Research Journal of Biological Science 2(1):69-71.

147
Bailey, S. A., Zidell, R. H and Perry, R.W. (2004). Relationship between organ weight and
brain weight in the rats: What is the best analytical endpoint. Toxicology Pathology. 32: 448-
466.
Bakanay, S. M., Dainer, E., Clair, B., Adekile, A., Daitch, L., Wells, L., Holley, L., Smith, D
and Kutlar, A. (2005). Mortality in sickle cell patients on hydroxyurea therapy. Blood, 105:
545-547.
Balkaran, B., Char, G., Morris, J. S., Thomas, P. W., Serjeant, B. E and Serjeant G.R.
(1992). Stroke in a cohort of patients with homozygous sickle cell disease. Journal of
Pediatrics. 120:360-366.
Bank, A. (2005). Understanding globin regulation in β-thalassemia: it‟s as simple as α, β, γ,
and δ. Journal of Clinical Invesigation. 115:1470-1473.
Bank, A. (2006). Regulation of human fetal hemoglobin: new players, new complexities.
Blood. 107:435-443.
Barile, F. A., Dierickx, P. J and Kristen, U. (1994). In vitro cytotoxicity testing for prediction
of acute human toxicity. Cell Biology and Toxicology. 10: 155-162.
Barinaga, M. (1997). Mutant Mice Mimic Human Sickle Cell Anemia. Science. 278:803-804
Beet, E. A. (1949). The genetics of the sickle-cell trait in a Bantu tribe. Annals of eugenics
(London) 14:279.
Belcher, J. D., Mahaseth, H., Welch, T. E., Otterbein, L. E., Hebbel, R. P and Vercellotti, G.
M. (2006). Heme oxygenase-1 is a modulator of inflammation and vaso-occlusion in
transgenic sickle mice. Journal of Clinical Investigation. 116:808-816.
Bennekou, P., De Franceschi, L., Pedersen, O., Lian, L., Asakura, T., Evans, G., Brugnara, C
and Christophersen, P. (2001). Treatment with NS3623, a novel Cl-conductance blocker,
ameliorates erythrocyte dehydration in transgenic SAD mice: a possible new therapeutic
approach for sickle cell disease. Blood. 97(5):1451–1457
Bennekou, P., Pedersen, O., Moller, A and Cristophersen, P. (2000). Volume control in sickle
cells is facilitated by the novel anion conductance inhibitor Ns1652. Blood.95:1842-1848
Bessey, O. A., Lowry, O. H. and Brock, M. J. (1946). A Method for the rapid determination
of Acid and Alkaline phosphatase in Serum. Journal of Biological Chemistry. 164: 321-329.
Bewaji, C. O., Olorunsogo, O. O and Bababumi, E. A. (1985). Sickle cell membrane-bound
(Ca2+
and Mg2+
)- ATPase: Activation by 3,4-dihydro-2, 2-dimethyl-2H-1-1-benzopyran-6-
butyric acid. A novel antisickling agent. Cell Calcium. 6. 237-244.
Bianchi, N., Ongaro, F., Chiarabelli, C., Gualandi, L., Mischiati, C and Bergamini, P. (2000).
Induction of erythroid differentiation of human K562 cells by cisplatin analogs. Biochemical
Pharmacology. 60:31–40.
Bianchi, N., Chiarabelli, C., Borgatti, M., Mischiati, C., Fibach, E and Gambari, R. (2001).
Accumulation of gamma-globin mRNA and induction of erythroid differentiation after

148
treatment of human leukaemic K562 cells with tallimustine. British Journal of
Haematology.113:951–61.
Bianchi, N., Zuccato, C., Lampronti, I., Borgatti, M., and Gambari, R. (2007). Fetal
Hemoglobin Inducers from the Natural World: A Novel Approach for Identification of Drugs
for the Treatment of β-Thalassemia and Sickle-Cell Anemia.
eCAM,doi:10.1093/ecam/nem/39
Bize, I., Taher, S and Brugnara, C. (2003). Regulation of K-Cl cotransport during reticulocyte
maturation and erythrocyte aging in normal and sickle erythrocytes. American Journal of
Physiology: Cell Physiology. 285: C31–C38
Blackman, S. M., Hustedt, E. J., Cobb., c. E and Beth., A. H. (2001). Flexibility of
cytoplasmic domaiiin of the anion exchange protein, band 3, in human erythrocytes.
Biophysical Journal. 81:3363-3376.
Bookchin, R. M and Lew, V. L. (1980). Progressive inhibition of the Ca pump and Ca: Ca
exchange in sickle red cells. Nature. 284:561-563.
Brandow, M. A., Jirovec, D. L and Panepinto, J. A. (2010). Hydroxyurea in Children with
Sickle Cell Disease: Practice Patterns and Barriers to Utilization. American Journal of
Hematology. 85(8): 611–613.
Brittain, J. E., Han, J., Kenneth I. Ataga, K. I., Eugene P. Orringer, E. P and Parise, L. V.
(2004). Mechanism of CD47-induced α4β1 Integrin Activation and Adhesion in Sickle
Reticulocytes. Journal of Biological Chemistry. 279(41):42393–42402.
Brown, A. M., Ellory, J. C., Young, J. D and Lew, V. L. (1978). A calcium activated
potassium channel present in foetal red cells of the sheep but absent from reticulocytes and
mature red cells. Biochim Biophys Acta. 511: 163–175.
Bruce, W. R and Heddle, J. A. (1979). The mutagenic activity of 61 agents as determined by
the micronucleus, Salmonella, and sperm abnormality assays. Canadian Journal of Genetics
and Cytology. 21:319–334.
Brugnara, C. Bunn, H. F and Tosteson, D. C. (1989). Ion content and transport and the
regulation of volume in sickle cells. Annuals of New York Academy of Science. 565: 96–103
Brugnara, C., DeFranceschi, L and Alper, S. L. (1993). Ca2+
-dependent K+ transport and cell
dehydration in sickle erythrocytes by clotrimazole and other imidazole derivatives. Journal of
Clinical Investigations, 92:520-526
BrugnaraC, De Franceschi L, Bennekou P, Alper SL, and Christophersen P. (2001). Novel
therapies for prevention of erythrocyte dehydration in sickle cell anemia. Drug News
Perspect 14: 208–220
Buchanan,G. R., DeBaun, M. R., Quinn, C. T and Steinberg, M. H. (2004). Sickle Cell
Disease. Hematology.1: 35-37.
Bunn, H. F. (1993). Reversing ontogeny. New England Journal of Medicine. 328:129-131.

149
Bunn, H. F. (1997). Mechanisms of disease–pathogenesis and treatment of sickle cell disease.
New England Journal of Medicine.337:762–769.
Bunn, H. F and Forget, B. G. (1986). Hemoglobin: Molecular, Genetic and Clinical Aspects.
Philadelphia, PA: Saunders. p690
Cao, H. (2004). Pharmacological induction of fetal hemoglobin synthesis using histone
deacetylase inhibitors.Hematology. 9:223–233
Carney, D. N and Winkler, C. F. (1985). "In vitro assays of chemotherapeutic
sensitivity".Important advances in oncology: 78–103
Chaine, B., Neonato, M. G., Girot, R and Aractingi, S. (2001). Cutaneous adverse reactions
to hydroxyurea in patients with sickle cell disease. Archives of Dermatology.137(4):467–470.
Chan, K. (2003). Some Aspect of Toxic Contaminants in Herbal Remedies. A Review
Chemosphere. 52: 1361-1371.
Chandel, M., Kaur, S and Kumar, S. (2011). Studies on the genoprotective/antioxidant
potential of methanol extract of Anthocephalus cadamba (Roxb.) Miq. Journal of Medicinal
Plants Research. 5(19):4764-4770
Chang, S. H and Loow, P. S. (2001). Reguulation of the glycoprotein C-protein 4.1
membrane toskeleton bridge and evaluation of its contribution to erythrocyte membrane
stability. Journal of Biological chemistry. 276: 22223-22230
Chasis, J. A., Prenant, M., Leung, A and Mohandas, N (1989). Membrane assembly and
remodeling during reticulocyte maturation. Blood. 74: 1112–1120.
Cheesbrough, M. (1991). Medical Laboratory Manual for Tropical Countries. Vol. 1 2nd
ed.
Pp 526
Chiang, E.Y and Frenette, P. S. (2005). Sickle cell vaso-occlusion. Hematology/Oncology
Clinics of North America. 19:771-784.
Chikezie, P. C and Uwakwe, A. A (2011). Membrane stability of sickle erythrocytes
incubated in extracts of three medicinal plants: Anacardium occidentale, Psidium guajava,
and Terminalia catappa. Pharmacognosy Magazine. 7(26): 121–125
Chiu, D., Lubin, B., Roelofsen, B and van Deenen, L. L (1981). Sickled erythrocytes
accelerate clotting in vitro: an effect of abnormal membrane lipid asymmetry. Blood. 58:398-
401.
Clark, M. R and Rossi, M. E. (1990). Permeability characteristics of deoxygenated sickle
cells. Blood. 76: 2139–2145
Claussen, W. J and Franco, R. S. (2000). Dehydration of mature and immature sickle red
blood cells during fast oxygenation/deoxygenation cycles: role of KCl cotransport and
extracellular calcium. Blood. 95:2164-2168

150
Cokic, V. P., Smith, R. D., Beleslin-Cokic, B. B., Njoroge, J. M., Miller, J. L., Gladwin, M. T
and Schechter, A. N. (2003). Hydroxyl urea induces fetal haemoglobin by the nitric oxide-
dependent activation of soluble gyuanylyl cyclase. Journal Clinical Investigation. 111: 231-
239.
Conley, C. L. (1980). Sickle-cell anemia. The first molecular disease, in Blood, Pure and
Eloquent. McGraw-Hill, New York. p 319.
Covas, D. T., de Lucena Angulo, I., Vianna Bonini Palma, P and Zago, M. A. (2004).
Effects of hydroxyurea on the membrane of erythrocytes and platelets in sickle cell anemia.
Haematologica. 89(3): 273-80.
Creasy, D. M. (2001). Pathogenesis of male reproductive toxicity. Toxicology Pathology.
29:64–76.
Creppy, E. E., Chiarappa, P., Baudrimont, I., Borracci, P., Moukha, S and Carratu, M. R.
(2004). Synergistic effects of fumonisin B1 and ochratoxin A: are in vitro cytotoxicity data
predictive of in vivo acute toxicity? Toxicology. 201: 115–123
Dacie, J.V and Lewis, S. (1991). Practical hematology 7th
edition Churchill Livingstone, New
York. Pp668
Dalziel, J. M and Hutchinson, J. D. (1958). Flora of West Tropical Africa, London, Vol. 1
part 2, p450- 455.
Davila, J.C; Reddy, C.G; Davis, P.J and Acosta, D. (1990). Toxicity assessment of
papaverine hydrochloride and papaverine –derived metabolitesin primary cultures of rat
hepatocytes. In vitro Cell Developmental Biology. 26:515-524.
De Franceschi, L., Bachir, D., Galacteros, F., Tchernia, G., Cynober, T., Neuberg, D.,
Beuzard, Y and Brugnara, C. (2000). Oral magnesium pidolate: effects of long-term
administration in patients with sickle cell disease. British Journal of Haematology. 108:
284–289
De Jong, K., Larkin, S. K., Styles, L. A., Bookchin, R. M and Kuypers, F. A. (2001).
Characterization of the phosphatidylserine-exposing subpopulation of sickle cells. Blood.
98:860-867.
DeFranceschj, L and Corrocher, R. (2004). Established and experimental treatments for
sickle cell disease. Heamatologica. 89(3):348-356
Driscoll, C. M. (2007). Sickle cell disease. Pediatrics in Review. 28 (7): 259 -268
Duh, P. D and Yen, G. C. (1997). Antioxidative activity of three herbal water extracts. Food
Chemistry. 6: 639-645.
Dzandu, J. K and Johnson, R. M. (1980). Membrane protein phosphorylation in intact normal
and sickle cell erythrocytes. Journal of Biological Chemistry. 255:6382, 1980.
Eaton, J. W., Jacob, H. S and White, J. G. (1979). Membrane abnormalities of irreversibly
sickled cells. Seminars in Hematology. 16:52

151
Eaton, W. A. (2003). Linus Pauling and sickle cell disease. Biophysical Chemistry. 100:109–
116.
Eaton, W. A., Hofrichter, J and Ross, P. D. (1976). Delay time of gelation: A possible
determinant of clinical severity in sickle cell disease. Blood 47:621.
Eberhardt, R. T., McMahon, L., Duffy, S. J., Steinberg, M. H., Perrine, S. P., Loscalzo, J.,
Coffman, J. D and Vita, J. A. (2003). Sickle cell anemia is associated with reduced nitric
oxide bioactivity in peripheral conduit and resistance vessels. American Journal of
Hematology.74:104–111.
Edeoga, H. O., Okwu, D. E. and Mbaebie, B. O. (2005). Phytochemical Constitutents of
some Nigerian Medicinal plants. African Journal of Biotechnology. 4(7): 685-688.
Elekwa, I., Monanu, M. O and Anosike, E. O. (2003). Studies on the effect of aqueous
extracts of Garcinia kola seed on the human erythrocytes adenosine triphosphatase of HbAA,
HbAS, and HbSS genotypes. Global Journal of Medical Sciences. 2(2). 107-114.
Elekwa, I., Monanu, M. O., Anosike, E. O. (2005). In vitro effects of aqueous extracts of
Zanthxylum macrophyla roots on adenosine triphosphatases from human erythrocytes of
different genotypes. Biochemistry. 17(1).19-25.
Elliott, G., Liu X-H., Cusick, J. L., Menze, M. A., Vincent, J., Witt, T., Hand, S. C and
Toner, M. (2006). Trehalose uptake through P2X7 purinergic channels provides dehydration
protection. Cryobiology. 52:114–127
Elujoba, A. A., Ogunti, E. O., Soremekun, R. D and Iranloye, T. A. (1994). The
Pharmacognosy and Dosage formulation of Cassia podocarpa leaf with reference to Senna.
Journal of Pharmaceutical Sciences & Pharmaceutical Practice. 2: 14-18.
Etzion, Z., Tiffert, T., Bookchin, R. M and Lew, V. L. (1993).Effects of deoxygenation on
active and passive Ca2+ transport and on the cytoplasmic Ca2+ levels of sickle cell anemia
red cells. Journal of Clinical Investigation. 92(5): 2489–2498
Fall, A. B., vanhaelen-Fastre, R., Vanhaelen, M., Lo, I., Toppet, M., Ferster, A., and Fondu,
P. (1999). In vitro antisickling activity of a rearranged limonoid isolated from Khaya
senegalensis. Planta Medica. 65(3):209-212
Fernandes, M. J. B., Limas.C., Rossi, M. H., Gonçales, E and Simoni, I. C. (2005).
Cytotoxicity of subfractions and compounds from Polymnia sonchifolia. Brazilian Journal of
Microbiology, 36(4):
Ferrone, F. A and Rotter, M. A. (2004). Crowding and the polymerization of sickle
hemoglobin. Journal of Molecular Recognition 17: 497–504.
Ferrone, F. A. (2004). Polymerization and sickle cell disease: a molecular view.
Microcirculation. 11:115-128.

152
Franco, R. S., Lohmann, J., Silberstein, E. B., Mayfield-Pratt, G., Palascak, M., Nemeth, T.
A., Joiner, C. H., Weiner, M and Rucknagel, D. L. (1998). Time-dependent changes in the
density and hemoglobin F content of biotin-labeled sickle cells. Journal of Clinical
Investigation. 101(12): 2730–2740.
Ferster, A., De Valck, C., Azzi, N., Fondu, P., Toppet, M and Sariban, E. (1992). Bone
marrow transplantation for severe sickle cell anaemia. British Journal of Haematology.
80:102-105
Frenette, P. S. (2004). Sickle cell vasoocclusion: heterotypic, multicellular aggregations
driven by leukocyte adhesion. Microcirculation. 11:167-177.
Frenette, P. S and Atweh, G. F. (2007). Sickle cell disease: old discoveries, new concepts,
and future promise. Journal of Clinical Investigation. 117(4):850–858.
Fu, P. P., Xia, Q., Chou, M. W. and Lin, G. (2007). Detection, Hepatotoxicity and
Tumorigenicity of Pyrrolizidine Alkaloids in Chinese Herbal Plants and Herbal Dietary
Supplements. Journal of Food and Drug Analysis. 15(4): 400-415.
Fung, E. B., Barden, E. M., Kawchak, D. A., Zemel, B. S., OheneFrempong, K and Stallings,
V. A. (2001). Effect of hydroxyurea therapy on resting energy expenditure in children with
sickle cell disease. Journal of Pediatrics, Hematology and Oncology. 23:604–608.
Ganapaty, S., Ch. Bharath and Laatsch, H. (2010). Des-N-Methylnoracronycine from the
roots of Carissa conjesta. Wight. International Journal of Green Pharmacy. 4:186 – 188
Gee, B. E and Platt, O. S. (1995). Sickle reticulocytes adhere to VCAM-1. Blood. 85:268-
274.
Gendreau, M. A and DeJohn, C. (2002). Responding to medical events during commercial
airline flights. New England Journal of Medicine. 346:1067-1073
Geva, A., Clark, A. J., Zhang, Y., popowicz, A., Manning, M. J and Neufeld, E. J. (2004).
Hemoglobin Jamican Plain- a Sickling haemoglobin with Reduced Oxygen Affinity. New
England Journal of Medicine. 351:1532-1538
Gillen, C. M., Brill, S., Payne, J. A and Forbush, B III.(1996). Molecular cloning and
functional expression of the K-Cl cotransporter from rabbit, rat, and human. A new member
of the cation-chloride cotransporter family. Journal of Biological Chemistry. 271: 16237–
16244.
Glynn, I. M and Karlish, S. J. (1975). The sodium pump. Annual Review of Physiology. 37:
13–55.
Hahn, E. V and Gillespie, E. B. (1927). Report of a case greatly improved by splenectomy;
experimental study of sickle cell formation. Archives of Internal Medicine 39:233.
Halliwell, B. (2000). The antioxidant paradox. Lancet. 355:1179 – 1180

153
Halliwell, B. (2008). Are polyphenols antioxidants or pro-oxidants? What do we learn from
cell culture and in vivo studies? Archives of Biochemistry and Biophysics. 476 (2): 107–112
Halsey C and Roberts, I. A. (2003) The role of hydroxyurea in sickle cell disease. British
Journal of Haematology. 120:177-186.
Hampton, T. (2008). Strides made in red blood cell disorders, but substantial barriers to care
remain. The Journal of the American Medical Association 299(4):395–396.
Hankins, J and Aygun, B. (2009). Pharmacotherapy in sickle cell disease – state of the art
and future prospects. British Journal of Haematology. 145(3): 296–308.
Hankins, J. A. (2009). Pharmacotherapy in sickle cell disease – state of the art and future
prospects. British Journal of Haematology. 145(3):296-308.
Hankins, J. S., Ware, R. E., Rogers, Z. R., Wynn, L. W., Lane, P. A., Scott, J. P and Wang,
W. C. (2005). Long-term hydroxyurea therapy for infants with sickle cell anemia: the
HUSOFT extension study. Blood.106: 2269-2275
Harbell, J. W., Knootz, S.W., Lewis, R. W and Lovell, D. (1997). IRAG working group: Cell
cytotoxic assays. Food and Chemical Toxicology., 35: 79-126
Harborne, J. B. (1991). A list of recommended TLC systems for all major classes of plant
chemical. In Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis. 2nd
Ed. Chapman and Hall: London, p.277 - 279.
Hassell, K. L., Eckman, J. R and Lane, P. A. (1994). Acute multiorgan failure syndrome: a
potentially catastrophic complication of severe sickle cell pain episodes. American Journal of
Medicine.96:155–162.
Hattangadi, S. M and Lodish, H. F. (2007). Regulation of erythrocyte lifespan: do reactive
oxygen species set the clock? Journal of Clincal Investigation.117(8):2075–2077.
He, Z and Russe, J. E. (2002). A Human embryonic hemoglobin inhibits Hb polymerization
in vitro and restores a normal phenotype tpo mouse models of sickle cell disease.
Proceedings of National Academic of Science. 99 (16): 10635-10640
Hebbel, R. P. (2003). Sickle hemoglobin instability: a mechanism for malarial protection.
Redox report : communications in free radical research. 8: 238–240
Hebbel, R. P., Yamada, O., Moldow, C. F., Jacob, H. S., White, J. G., Eaton, J. W. (1980).
Abnormal adherence of sickle erythrocytes to cultured vascular endothelium: possible
mechanism for microvascular occlusion in sickle cell disease. Journal of Clincal
Investigation. 65:154-160.
Herrick, J. B. (1910). Peculiar elongated and sickle-shaped red corpuscles in a case of severe
anemia. Archives of Internal Medicine 6:517.
Hoffman and Robert, M. (1991). "In vitro sensitivity assays in cancer: A review, analysis and
prognosis". Journal of Clinical Laboratory Analysis 5 (2): 133-43.

154
Hoover, R., Rubin, R., Wise, G., Warren, R. (1979). Adhesion of normal and sickle
erythrocytes to endothelial monolayer cultures. Blood. 54:872-876.
Huang, D., Lin, C., Chen, H and Lin, Y. (2004). Antioxidant and antiproliferative activitie of
sweeet potato (Ipomea batatas[L] Lam „tainong‟) constituents. Botanical Bulletting of
Academy of Science. 45: 179-186
Idu, M., Orosanya, F. E., Igeleke, C. L, Omonigho, Omogbeme, O. E and Ayinde, B. A.
(2006). Preliminary investigation on the phytochemistry and antimicrobial activity of Senna
alata Leaves. Journal of Applied Sciences. 6: 14 - 22
Imaga, N. O. A., Gbenle, G. O., Okochi, V. I., Adenekan, S. O., Edeoghon, S. O., Kehinde,
M. O., Bamiro, S. B., Ajiboye, A., Obinna, A. (2010). Antisickling property of Carica
papaya leaf extract. African Journal of Biochemical Research. 3(4): 102 - 106
Imaga, N. O. A., Gbenle, G. O., Okochi, V. I., Akanbi, S. O., Edeoghon, S. O., Oigbochie,
V., Kehinde, M. O amd Bamiro, S. B. (2009). Antisickling property of Carica papaya leaf
extract. African Journal of Biochemical Research. 3(4): 102-106.
Ingram, V. M. ((1957). Gene mutations in human haemoglobin: The chemical difference
between normal and sickle cell haemoglobin. Nature 180:326.
Ingram, V. M. (1949). Abnormal human haemoglobins: III. The chemical difference between
normal and sickle cell haemoglobins. Biochimica et Biophysica Acta. 36:402.
Ingram, V. M. (1958). Abnormal human haemoglobins. I. The comparison of normal human
and sickle-cell haemoglobins by fingerprinting. Biochimica et Biophysica Acta.. 28:539-545.
International Agency for Research on Cancer (IARC) (2000). IARC Monographs on the
evaluation of the carcinogenic risk of chemicals to man. Hydroxyurea. World Health
Organisation. 76: 347
Ishiyama, M., Tominaga, H., Shiga, M and Sasamoto, K. (1996). A combined assay of cell
viability and in vitro cytotoxicity with a highly water-soluble tetrazolium salt, neutral red and
crystal violet. Biol. Pharm. Bull., 19(11):1518-1520
Issaa, H and Al-Salemb, A. H. (2010). Hepatobiliary Manifestations of Sickle Cell Anemia.
Gastroenterology Research. 3(1):1-8
Iyamu, E. W., Turner, E. A and Asakura, T. (2003). Niprisan (Nix – 0699) improves the
survival rates of transgenic sickle cell mice under acute severe hypoxic conditions. British
Journal of Hematology. 122: 1001-1008.
Jaja, S. I, Kehinde, M. O., Gbenebitse, S., Mojiminyi, F. B. O and Ogungbemi, A. I. (2000).
Effect of vitamin C on arterial blood pressure, irreversible sickled cells and osmotic fragility
in sickle cell anemia subjects. Nigerian Journal of Physiological Sciences. 16(1): 14-18.

155
Jaja, S. I., Aigbe, P. E., Gbenebitse, S., Temiye, E. O. (2005). Changes in erythrocytes
following supplementation with alpha-tocopherol in children suffering from sickle cell
anemia. Nigerian postgraduate medical journal. 12(2):110–4.
Jaja, S. I., Ikotun, A. R., Gbenebitse, S and Temiye, E. O. (2002). Blood pressure,
hematologic and erythrocyte fragility changes in children suffering from sickle cell anemia
following ascorbic acid supplementation. Journal of Tropical Pediatrics.48(6); 366 – 370.
Janero, D. R. (1990). Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices
of lipid peroxidation and peroxidative tissue injury. Free Radical Biology Medicine. 9:515-
540
Jayabose, S., Tugal, O., Sandoval, C., Patel, P., Puder, D., Lin, T and Visintainer, P. (1996).
Clinical and hematologic effects of hydroxyurea in children with sickle cell anemia. Journal
of Pediatrics. 129: 559–565.
Jennings, M. L and Schulz, R. K. (1991). Okadaic acid inhibition of KCl cotransport.
Evidence that protein dephosphorylation is necessary for activation of transport by either cell
swelling or N-ethylmaleimide. Journal of General Physiology 97: 799–817.
Jennings, M. L and Adame M. F. (2001). Direct estimate of 1:1 stoichiometry of K(+)-Cl(–)
cotransport in rabbit erythrocytes. American Journal of Physiology, Cell Physiology 281:
C825–C832
Johanning, L. G and O'dell, B. L. (1989). Effect of zinc deficiency and food restriction in rats
on erythrocyte membrane zinc, phospholipid and protein content. Journal of Nutrition. 119:
1654-1660.
Belcher, J. D., Bryant, J. C., Nguyen, J., Bowlin, P. R., Kielbik, M. C., Bischof, J. C., Hebbel,
R. P and Vercellotti, G. M. (2003). Transgenic sickle mice have vascular inflammation.
Blood. 101:3953-3959.
Johnstone, R. M. (1992). The Jeane Manery-Fishery Memorial Lecture 1991. Maturation of
reticulocytes: formation of exosomes as a mechanism for shedding membrane proteins.
Biochemistry of Cell Biology. 70: 179-190.
Joiner, C. H and Franco, R. S. (2001). The activation of KCl cotransport by deoxygenation
and its role in sickle cell dehydration. Blood Cells Molecules & Diseases 27: 158–164.
Joiner, C. H., Platt, O. S and Lux, S. E. (1986). Cation depletion by the sodium pump in red
cells with pathologic cation leaks. Sickle cells and xerocytes. Journal of Clinical
Investigation. 78: 1487–1496.
Joneckis, C.C., Ackley, R.L., Orringer, E.P., Wayner, E.A., Parise, L.V. (1993). Integrin
alpha 4 beta 1 and glycoprotein IV (CD36) are expressed on circulating reticulocytes in sickle
cell anemia. Blood. 82:3548-3555.
Kanodia, K. V., Vanikar, A. V., Goplani, K. R., Gupta, S. B and Trivedi, H. L. (2008). Sickle
cell nephropathy with diffuse proliferative lupus nephritis: a case report. Diagnostic
Pathology. 3:9

156
Kaul, D. K and Hebbel, R. P. (2000). Hypoxia/reoxygenation causes inflammatory response
in transgenic sickle mice but not in normal mice. Journal of Clinical Investigation. 106:411-
420.
Kaul, D. K., Tsai, H. M., Liu, X. D., Nakada, M. T., Nagel, R. L and Coller. B. S. (2000).
Monoclonal antibodies to alphaVbeta3 (7E3 and LM609) inhibit sickle red blood cell-
endothelium interactions induced by platelet-activating factor. Blood. 95:368-374.
Kaul, D.K., Nagel, R.L., Chen, D., Tsai, H. M. (1993). Sickle erythrocyte-endothelial
interactions in microcirculation: the role of von Willebrand factor and implications for
vasoocclusion. Blood. 81(9):2429-38
Kavitha, N., Noordin, R., Chan, K and Sasidharan, S. (2010). Cytotoxicity activity of root
extract/fractions of Eurycoma longifolia Jack root against Vero and Hs27cells. Journal of
Medicinal Plants Research. 4(22): 2383-2387
Khanna, R., Chang, S. H., Andrabi, S., Azam, M., King, A., Divera, A., Brugnara, C., Low,
P. S., Liu, s. C and Chishti, A. H. (2002). Headpiece domain of dematin is required for the
stability of the erythrocyte membrane. Proceedings of the academy of Science, U.S.A. 99:
6637-6642
Klug, P. P., Lessin, L. S and Peter Radice, B. S. (1974). Rheological aspects of sickle cell
disease. Archives of Internal Medicine. 133(4):577-590.
Klug, W. S., Cummings, M. R., Spencer, C. A and Palladino, M. A. (2010). Concepts of
genetics (10th
ed.). Pearsons. Pp. 895
Knauf, P. A., Law, F. Y and Marchant, P. J. (1983). Relationship of net chloride flow across
the human erythrocyte membrane to the anion exchange mechanism. Journal of General
Physiology 81: 95–126.
Krazanowska, H. (1986). Inheritance of sperm head abnormality types in mice and the role of
the Y chromosomes. Genetics Research. 28:189-198
Krings, U and Berger, R. G. (2001). Antioxidant activity of some roasted foods. Food
Chemistry. 72 :223–29.
Kutlar A. (2007). Sickle cell disease: a multigenic perspective of a single gene disorder.
Hemoglobin, 31:209–24.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of head of
bacteriophage T4. Nature. 227: 680-685.
Lal, A and Ames, B. N. (2011). Association of chromosome damage detected as micronuclei
with hematological diseases and micronutrient status. Mutagenesis. 26(1): 57–62.
Lal, A and Vichinsky, E. (2004). The role of fetal hemoglobin-enhancing agents in
thalassemia. Seminars in Haematology. 41:17–22.
Latagliata, R., Spadea, A., Cedrone, M., Di Giandomenico, J., De Muro, M., Villivà, N.,
Breccia, M., Anaclerico, B., Porrini, R., Spirito, F., Rago, A., Avvisati, G., Alimena, G.,
Montanaro, M., Andriani, A. and Gruppo Laziale SMPC Ph1 neg. (2012). Symptomatic

157
mucocutaneous toxicity of hydroxyurea in Philadelphia chromosome-negative
myeloproliferative neoplasms. Cancer. 118: 404–409.
Lauf, P. K and Adragna, N. C. (2000). K-Cl cotransport: properties and molecular
mechanism. Cellular Physiology and Biochemistry 10:341–354.
Lauf, P. K., Bauer, J., Adragna, N. C., Fujise, H., Zade-Oppen, A. M. M., Ryu, K. H and
Delpire, E. (1992). Erythrocyte K-Cl cotransport: properties and regulation. American
Journal of Physiology, Cell Physiology 263: C917–C932.
Lee, A. P., Yamamoto, L. G and Relles, N. L. (2002). Commercial airline travel decreases
oxygen saturation in children. Paediatric Emergency Care, 18:78-80.
Letelier, M.A., Cortes, J.F., Lepe, A.M., Jara, J.A., Molina-Berrios, A., Rodriquez, C., Iturra-
Montecinos, P. and Faundez, M. (2009). Evaluation of the antioxidant properties and effects
on the biotransformation of commercial herbal preparations using rat liver endoplasmic
reticulum. Boletin Latinamericano y del Caribe de Plantas Medicinales y Aromaticas. 8(2):
110-120
Letz, G. (1990). Male reproductive toxicology, In: J. Ladou (Ed.). Occupational Medicine.
Appleton and Lange. Connecticut, pp. 288-296.
Lew, V. L and Bookchin, R. M. (1986). Volume, pH and-ion content regulation in human red
cells: analysis of transient behavior with an integrated model. Journal of Membrane Biology.
92: 57–74.
Lew, V. L., Freeman, C. J., Ortiz, O. E and Bookchin, R. M. (1991). A mathematical model
on the volume, pH and ion content regulation of reticulocytes. Application to the
pathophysiology of sickle cell dehydration. Journal of Clinical Investigation.87: 100–112.
Lew,V. L, Ortiz-Carranza, O. E and Bookchin, R. M. (1997). Stochastic nature and red cell
population distribution of the sickling-induced Ca2+
permeability. Journal of Clinical
Investigation. 99: 2727–2735.
Lew,V. L., Etzion, Z and Bookchin, R. M. (2002). Dehydration response of sickle cells to
sickling-induced Ca(++) permeabilization. Blood. 99: 2578–2585.
Lew,V. L., Tsien, R. Y., Miner, C and Bookchin, R. M. (1982). Physiological [Ca2+
]i level
and pump-leak turnover in intact red cells measured using an incorporated Ca chelator.
Nature. 298: 478–481.
Li, Q., Fang, X., Han, H and Stamatoyannopoulos, G. (2004). The minimal promoter plays a
major role in silencing of the galago γ-globin gene in adult erythropoiesis. Proceedings of the
National Academy of Science, U S A.101:8096–8101.
Li, H., Ko, C. H., Tsang, S. K., Leung, P. C., Fung, M. C and Fung, K. P. (2011). The
Ethanol Extract of Fructus trichosanthis Promotes Fetal Hemoglobin Production via p38
MAPK Activation and ERK Inactivation in K562 Cells. Evidence -based Complement and
Alternative Medicine. 2011: 657056.

158
Long, L. H., Clement, M. V., and Halliwell, B. (2000). Artifacts in cell culture: rapid
generation of hydrogen peroxide on addition of (-)-epigallocatechin, (-)- epigallocatechin
gallate, (+)-catechin and quercetin to commonly used cell culture media. Biochemical and
Biophysical Research Communications. 273:50–53.
Lowry, O. H., Rosebrough, N. J., Farr, A. L and Randall, R. J. (1951). Protein measurement
with the folin phenol reagent. Journal of Biological Chemistry. 193: 265-275.
Lozzio, C. B and Lozzio, B. B. (1975). "Human chronic myelogenous leukemia cell-line with
positive Philadelphia chromosome", Blood 45 (3): 321–34
Macedo, M. F. S,. Sisenado, J. D. F., Queiroz, A. C. C., Argolo, A. C. R., Dantas Saturino, L.
C., Coelho, B. B and Batistuzzo de Medeiros, B. B. (2008). Determining the genotoxicity of
an aqueous infusion of Bauhinia monandra leaves. Brazilian Journal of Pharmacology.
18(4): 509-516.
Makinde, A. A., Igoli, J. O., TA‟Ama, L., Shuaibu, S. J and Garba, A. (2007). Antimicrobial
activity of Senna alata. African Journal of Biotechnology. 6: (13): 1509 – 1510.
McGoron, A. J., Joiner, C. H., Palascak, M. B., Claussen, W. J and Franco, R. S. (2000).
Dehydration of mature and immature sickle red blood cells during fast
oxygenation/deoxygenation cycles: role of KCl cotransport and extracellular calcium. Blood
95: 2164–2168
Mehanna, A. S. (2001). Sickle Cell Anemia and Antisickling Agents Then and Now. Current
Medicinal Chemistry. 8: 79-88
Merciris, P., Claussen, W. J, Joiner, C. H and Giraud, F. (2003). Regulation of K-Cl
cotransport by Syk and Src protein tyrosine kinases in deoxygenated sickle cells. Pflugers
Archives 446: 232–238.
Messmann, R., Gannon, S., Sarnaik, S and Johnson, R. M. (1990). Mechanical properties of
sickle cell membranes. Blood. 75: 1711-1717
Mgbemene, C. N and Ohiri, F. C. (1999). Anti-sickling potential of Terminalia catappa leaf
extract. Pharmaceutical Biology. 37(2):152-154.
Milner, P. F., Leibfarth, J. D., Ford, J., Barton, B. P., Grenett, H. E and FA Garver, F. A.
(1984). Increased HbF in sickle cell anemia is determined by a factor linked to the beta S
gene from one parent. Blood. 63: 64-72
Mohandas, N and Evans, E. (1989). Rheological and adherence properties of sickle cells:
potential contribution to hematologic manifestations of the disease. Annals of the New York
Academy of Sciences. 565:327-337
Mohanty, S., Sharma, M., Dubey, K., Amit Alexander, A., Khan, J., Khan, S and Tripathi, D.
K. (2012). Significance of Herbal Plants in the Treatment of Sickle Cell Anaemia. Research
Journal of Pharmacy and Technology. 5 (3):297-301

159
Moody, J. O., Robert, V. A., Connolly, J. D and Houghton, P. J. (2003). Anitinflammatory
activities of methanol extracts and an isolated furanoditerpene constituent of Sphenocentrum
jollyanum Pierre (Menispermaceae). Journal of Ethnopharmacology. 104(1-2): 67-91.
Moriguchi, T., Takasugi, N and Itakura, Y.(2001). The effects of AGED garlic extract on
lipid peroxidation and deformability of erythrocytes. American society for Nutritional
sciences. 131(3s):1016S-1019S.
Morita, K., Arimochi, H and Ohnishi, Y. ( 2003). In Vitro Cytotoxicity of 4-Methylcatechol
in Murine Tumor Cells: Induction of Apoptotic Cell Death by Extracellular Pro-Oxidant
Action. Journal of Pharmacology and Experimental Therapeutics. 306(1): 317-323.
Mpiana, P. T., Tshibangu, D. S. T., Shehonde, O. M and Ngbolua, K. N. (2007). In vitro
antidrepanocytary activity (antisickle cell anemia) of some Congolese plants. Phytomedicine.
14(2-3):192-195
Muzyamba, M. C. and Gibson, J. S. (2003). Effect of 1-chloro-2,4-dinitrobenzene on K+
transport in normal and sickle human red blood cells. Journal of Physiology. 547(3):903-911
Neel, J. V. (1949). The inheritance of sickle cell anemia. Science 110:64-66.
Nesamvuni, C., Steyn, N and Potgieter, M. (2001). Nutrients analysis of selected western
African foods. South African Journal of Science. 97: 51-54.
Niehaus, Jr. W.G. and Samuelson, B. (1968). Formation of malonaldehyde from phospholipid
arachidonate during microsomal lipid peroxidation. European Journal of Biochemistry. 6:
126-130.
Noguchi, C.T. and Schechter, A.N. (1978). The inhibition of sickle cell haemoglobin gelation
by amino acids and related compounds. Biochemistry, 17: 5455-59.
Obidike, I. C., Shehu Idris-Usman, I., John-Africa, L. B and Salawu, O. A. (2011). An
Evaluation of Acute and Sub Chronic Toxicological Effects of Hymenocardia acida Leaf
Extract in Adult Wistar Rats. Journal of Pharmacology and Toxicology. 6: 400-408.
Odugbemi, T. (2006). Outlines of pictures of medicinal plants from Nigeria. University of
Lagos Press. pp283.
Oduola, T, Adeniyi, F. A. A, Ogunyemi, E. O., Bello, I. S and Idowu, T. O. (2006).
Antisickling agent in an extract of unripe pawpaw (Carica papaya): Is it real? African
Journal of Biotechnology. 5(20): 1947-1949.
Ogunkunle, A.T.J. and Ladejobi,T. A. (2006). Ethnobotanical and Phytochemical studies on
some species of Senna alata. African Journal of Biotechnology. 5(21): 2020-2023.
Ohnishi, S. T and Ohnishi. T.(2001). In vitro effects of aged garlic extract and other
nutritional supplements on sickle erythrocytes. Journal of Nutrition.131:1085S-1092S.
Okigbo, R. N., Anuagasi, C. L and Amadi, J. E. (2009). Advances in selected medicinal and
aromatic plants indigenous to Africa. Journal of Medicinal Plants Research. 3(2): 86-95

160
Okoro, I. S., Osagie, A and Asibor, E. O. (2010). Antioxidant and antimicrobial activities of
polyphenols from ethnomedicinal plants of Nigeria. African Journal of Biotechnology. 9 (20):
2989-2993.
Okpala, I and Tawil, A. (2002). Management of pain in sickle-cell disease. Journal of the
Royal Society of Medicine. 95(9): 456–458.
Okpuzor, J and Adebesin, O. (2006). Membrane stabilizing effect and antisickling activity of
Senna podocarpa and Senna alata. 31st
congress for European biochemical societies,
Instanbul, Turkey, July 25-29, 2006.
Okpuzor, J., Adebesin, O., Ogbunugafor, H and Amadi, I. (2008). The potential of medicinal
in sickle cell disease control: A review. International Journal of Biomedical and Health
Sciences. 4(2): 47 – 55.
Onah, J. O., Akubue, P. I and Okide, J. F. (2002). The kinetics of reversal of presickled
erythrocytes by the aqueous extract of Cajanus cajan. Phytotherapy Research. 16(8): 748-
750
Oneal, P. A., Gantt, N. M., Schwartz, J. D., Bhanu, N. V., Terry Lee, Y., Moroney, J. W.,
Reed, C. H., Schechter, A. N., Luban, N. L. C and Miller, J. L. (2006). Fetal hemoglobin
silencing in humans. Blood. 108(6): 2081–2086.
Orringer, E. P., Casella, J. F., Ataga, K. I., Koshy, M., Adams-Graves, P., Luchtman-Jones,
L., Wun, T., Watanabe, M., Shafer, F., Kutlar, A., Abboud, M., Steinberg, M., Adler, B.,
Swerdlow, P., Terregino, C., Saccente, S., Files, B., Ballas, S., Brown, R., Wojtowicz-Praga,
S and Grindel, J. M. (2001). Purified poloxamer 188 for treatment of acute vaso-occlusive
crisis of sickle cell disease: A randomized controlled trial. Journal of the American Medical
Association. 286:2099–2106.
Ortiz, O. E., Lew, V. L and Bookchin, R. M. (1990). Deoxygenation permeabilizes sickle cell
anemia red cells to magnesium and reverses its gradient in the dense cells. Journal of
Physiology. 427: 211–226.
Osman, H., Nasarudin, R and Lee, S. L. (2004). Extract of cocoa ( Theobroma cacao L.)
leaves and their antioxidation potential. Food Chemistry. 86: 41-46
Ou, J., Ou, Z., Jones, D. W., Holzhauer, S., Hatoum, O. A., Ackerman, A. W., Weihrauch, D.
W., Gutterman, D. D., Guice, K., Oldham, K. T., Hillery, C. A., Pritchard, K. A. Jr. (2003).
L-4F, an apolipoprotein A-1 mimetic, dramatically improves vasodilation in
hypercholesterolemia and sickle cell disease. Circulation. 107:2337–2341.
Owoyale, J. A.., Olatunji, G. A and Oguntoye, S. O. (2005). Antifungal and antibacterial
activities of an alcoholic extract of Senna alataleaves. Journal of Applied Science and.
Environmental Management. 9: 105-107
Palek, J and Lambert, S. (1990). Genetics of the red cell membrane skeleton. Seminars on
Hematology 27: 290–332.

161
Patel, S., Gheewala, N., Suthar, A and Shah, A. (2009). In-Vitro Cytotoxicity Activity Of
Solanum nigrum Extract Against Hela Cell Line And Vero Cell Line. International Journal
of Pharmacy and Pharmaceutical Sciences, 1(1): 38 -46
Pauling, L. (1964). Molecular disease and evolution. Bulletin of the New York Academy of
Medicine. 40:334-342.
Pauling, L., Itano, H.A., Singer, S.J., Wells, I. C. (1949). Sickle cell anemia, a molecular
disease. Science. 110:543-548.
Persons, D. A and Nienhuis, A. W. (2000). Gene therapy for the hemoglobin disorders: Past,
present, and future. Proceedings of the National Academy of science. 97(10):5022-5024.
Perumbeti, A and Malik, P. (2010). Therapy for beta-globinopathies: a brief review and
determinants for successful and safe correction. Annals of the New York Academy of Science.
1202:36–44.
Pieme, C. A., Penlap, V. N. Nkegoum, B., Taziebou, C. L., Tekwu, E. M., Etoa, F. X and
Ngongang, J. (2006). Evaluation of acute and subacute toxicities of aqueous ethanolic extract
of leaves of Senna alata (L.) Roxb (Ceasalpiniaceae). African Journal of Biotechnology.
5(3): 283-289.
Pieme, C. A., Penlap, V. N., Nkegoum, B. and Ngogang, J. (2008). In vivo antioxidant and
potential antitumor activity of aqueous ethanol extract of leaves of Senna alata (L.) Roxb.
(Ceasalpiniaceae) on Carcinomatous Cells. International Journal of Pharmacology. 4(4):
245-251.
Platt, O. S. (2008). Hydroxyurea for the Treatment of Sickle Cell Anemia. New England
Journal of Medicine.358:1362-1369.
Platt, O. S., Falcone, J. F and Lux, S. E. (1985). Molecular Defects in the Sickle Erythrocyte
Skeleton - Abnormal Spectrin Binding to Sickle Inside-Out Vesicles. Journal of Clinical
Investigation. 75(1):266–271.
Platt, O. S., Orkin, S. H., Dover, G., Beardsley, G. P., Miller, B and Nathan, D. G. (1984).
Hydroxyurea enhances fetal hemoglobin production in sickle cell anemia. Journal of Clinical
Investigation. 74:652-656
Presley, T. D., Perlegas, A. S., Bain, L. E., Ballas, S. K., Nichols, J. S., Sabio, H., Gladwin,
M. T., Kato, G. J and Kim-Shapiro, D. B. (2010). Effects of a single sickling event on the
mechanical fragility of sickle trait erythrocytes. Hemoglobin. 34(1):24-36
Prieto, P., Pineda, M and Aguilar, M. (1999). Spectrophotometric quantitation of antioxidant
capacity through the formation of a phosphomolybdenum complex: Specific application to
the determination of vitamin E. Analytical Biochemistry.269: 337–41.
Rael, L. T., Thomas, G.W., Craun, M. L., Curtis, C. G., Bar-Or, R and Bar-Or, D. (2004).
Lipid Peroxidation and the Thiobarbituric Acid Assay: Standardization of the Fatty Acids.
Journal of Biochemistry and Molecular Biology. 37: 749-752

162
Rai, P. P and Abdulahi, N. I. (1978). Occurrences of anthraquinones in the leaves of Cassia
species. Nigerian Journal of Pharmacy. 9:160-165.
Rai, P. P and Obayemi, O. M. (1973). Anthraquinones from the leaves of Cassia podocarpa.
Current Science. 47:457-460.
Rajadurai, M. Vidhya, V.G., Ramya, M. and Bhaskar, A. (2009). Ethno-medicinal plants
used by traditional Healers of Pachamalai Hills, Tamilandu, India. Ethno-medicine. 3(1): 39-
41.
Randi, M. L, Ruzzon, E., Tezza, F., Luzzatto, G., Fabris, F. (2005). Toxicity and side effects
of hydroxyurea used for primary thrombocythemia. Platelets. 16(3-4):181–184.
Rank, B. H., Carlsson, J and Hebbel, R. P. (1985). Abnormal Redox Status of Membrane-
Protein Thiols in Sickle Erythrocytes. Journal of Clinical Investigation.75(5):1531–1537.
Rates, S. M. K. (2001). Plants as source of drugs Toxicon, 39:603-613
Reitman, S and Frankel, S. (1957). A colorimetric method for the determination of serum
glutamic oxalacetic and glutamic pyruvic transaminases. American Journal of Clinical.
Pathology. 28: 56-63
Rhoda, M. D., Apovo, M. Beuzard, Y and Giraud, F. (1990). Ca2+
permeability in
deoxygenated sickle cells. Blood. 2453-2458
Rhomberg, L. R., Baetcke, K., Blancato, J., Bus, J., Cohen, S., Conolly, R., Dixit, R., Doe,
J., Ekelman, K and Fenner-Crisp, P. (2007). Issues in the design and interpretation of chronic
toxicity and carcinogenicity studies in rodents: approaches to dose selection. Critical Reviews
in Toxicology. 37:729-837.
Rifai, Z., Welle, S., Kamp, C and Thornton, C.A. (1995). Ragged red fibers in normal aging
and inflammatory myopathy. Annals of Neurology. 37: 24–29.
Riss, T and Moravec, R. A. (2004). Use of multiple assay endpoints to investigate effects of
incubation time, dose of toxin and plating density in cell-based cytotoxicity assays. Assay
Drug Dev. Technol. 2:51–62.
Romero,,J.R., Suzuka, S.M., Nagel, R.L and Fabry, M. E. (2002). Arginine supplementation
of sickle transgenic mice reduces red cell density and Gardos channel activity. Blood,
99:1103-1108
Rotter, M. A., Kwong, S., Briehl, R. W and Ferrone, F. A. (2005). Heterogeneous nucleation
in sickle hemoglobin: experimental validation of a structural mechanism. Biophysical
Journal. 89: 2677-2684.
Russo, A., Izzo, A. A., Cardile, V and Borrelli, F. (2001). Vanella A. Indian medicinal plants
as antiradicals and DNA cleavage protector. Phytomedicine. 8: 125–32
Saad, B., Dakwar S., Said, O., Abu-Hijleh G., Al Battah, F., Kmeel, A. and Hassan, A.
(2006). Evalution of Medicinal Plant Hepatotoxicity in Co-cultures of Hepatocytes and
Monocytes. Evidence-based Complementary and Alternative Medicine. 3(1): 93-98.

163
Sakano, K., Oikawa, S., Hasegawa, K and Kawanishi, S. (2001). Hydroxyurea induces site-
specific DNA damage via formation of hydrogen peroxide and nitric oxide. Japanese journal
of cancer research: Gann. 92(11):1166–1174.
Sandler, S. G., Mallory, D. A., Wolfe, J. S., Byrne, P. C. and Lucas, D. M. (1997). Screening
with monoclonal anti-Fy3 to provide blood for phenotype- matched transfusions for patients
with sickle cell disease. Transfusion. 37: 393–397.
Santos, J. L., Bosquesi, P. L., Almeida, A. E., Chung Man Chin and Varanda, E. A. (2011).
Mutagenic and genotoxic effect of hydroxyurea. International Journal of Biomedical
Science. 7(4): 263-267.
Saunthararajah, Y., Hillery, C.A., Lavelle, D., Molokie, R., Dorn, L., Bressler, L., Gavazora,
S., Chen, Y. H., Hoffman, R and DeSimone, J. (2003). Effects of 5-aza-2′-deoxycytidine on
fetal haemoglobin levels, red cell adhesion, and haematopoietic differentiation in patients
with sickle cell disease. Blood. 102: 3865–3870.
Schwartz, R. S., Rybicki, A.C., Heath, R. H and Lubin, B. H. (1987). Protein 4.1 in Sickle
Erythrocytes - Evidence for Oxidative Damage. Journal of Biological Chemistry.
262(32):15666–15672.
Scriver, J. R and Waugh, T. R. (1930). Studies on a case of sickle cell anemia. Canadian
Medical Association Journal. 23:375-380.
Sellers, S. R., Morton, D., Michael, B., Roome, N., Johnson, J. K., Yano, B. L., Perry, R and
Schaffer, K. (2007). Society of Toxicologic Pathology Position Paper: OrganWeight
Recommendations for Toxicology Studies. Toxicologic Pathology. 35:751–755
Serjeant, G. R (1997). Sickle cell disease. Lancet. 350: 725-730
Serjeant, G.R., Ashcroft, M.T., Serjeant, B.E., Milner, P. F. (1973). The clinical features of
sickle-cell-thalassaemia in Jamaica. British Journal of Haematology. 24:19-30.
Serjeant, G.R., Sommereux, A.M., Stevenson, M., Mason, K., Serjeant, B. E. (1979).
Comparison of sickle cell-beta0 thalassaemia with homozygous sickle cell disease. British
Journal of Haematology. 41:83-93.
Shalev, O and Hebbel, R. P. (1996). Catalysis of soluble hemoglobin oxidation by free iron
on sickle red cell membranes. Blood. 87(9):3948–3952.
Shoemaker, M., Cohen, I and Campbell, M. (2004). Reduction of MTT by aqueous herbal
extracts in the absence of cells. Journal of Ethnopharmacology. 93(2-3): 381-384
Singleton, V. L., Orthofer, R and Lamuela-Raventos, R. M. (1999). Analysis of total
phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent.
Methods in Enzymology. 299: 152–78.
Sinha, K.A. (1972). Colorimetric Assay of Catalase. Annals of Biochemistry. 47:389-394

164
Skickel, F., Patsenke, E. and Schuppan, D. (2005). Herbal Hepatotoxicity. Journal of
Hepatology. 43: 901-910.
Skou, J. C and Esmann, M. (1992). The Na,K-ATPase. Journal of Bioenergetics and
Biomembranes. 24: 249–261.
Smith, E. L., Hill, R. L., Bughaith, M. F. and El-Gindy, A. R. (1983). Principles of
Biochemistry. 7th Ed., McGraw-Hill, USA. pp. 886
Sofowora, E. A., Isaacs-Sodeye, W. A., Ogunkoya, L. O. (1979). Isolation and
Characterization of an antisickling agent from the root of Fagara zanthoxyloides. African
Medicinal Plants. Proceedings of a Conference. Ed. Sofowora EA, pp. 89-97.
Sofowora, E.A. (1974). Recent Developments in Research into the Antisickling Properties of
Fagara zanthoxyloides (Orin Ata). The J. Pharm. (Nigeria) 5: 8-14.
Sofowora, E. A. (1975). Isolation and characterization of an anti-sickling agent from Fagara
zanthoxyloides. Lloydia 38:169-171.
Sofowora, A. (1986). The State of Medicinal Plants Research in Nigeria. University Press,
Ibadan, Nigeria. p86.
Sofowora, E. A. (1982). Medicinal Plants and Traditional Medicine in Africa. John Wiley
and Sons, London. p249.
Sofowora, E.A, Isaac-Sodeye, W .A and Ogunkoya, L .O. (1975). African Medicinal Plants:
Isolation and Characterization of an anti-sicking agents from Fagara zanthoxyloides
root.Lloydia 34:169-174
Sofowora, E. A and Issacs-Sodeye, W. A. (1971). Reversal of sickling and crenation in
erthrocytes by the root of Fagara xanthoxyloides. LIoydia. 34: 383.
Stamatoyannopoulos, G. (2005). Control of globin gene expression during development and
erythroid differentiation. Experimental Hematology. 33:259-271.
Stegeman, J. J., Miller, M. R and Hinton, D. E. (1989). Cytochrome P450IA1 induction and
localization in endothelium of vertebrate (teleost) heart. Molecular Pharmacology 36: 723–
729.
Steinberg, M. H., Lu, Z. H., Barton, F. B., Terrin, M. L., Charache, S and Dover, G. J
(1997a). Fetal haemoglobin in sickle cell anaemia: determinants of response to hydroxylurea.
Multicenter Study of Hydroxylurea. Blood. 89(3): 1078 - 1088
Steinberg, M. H., Nagel, R. L, Brugnara, C. (1997b). Cellular effects of hydroxyurea in Hb
SC disease. British Journal of Haematology.98:838–844.
Steinberg, M. H. (1999). Management of sickle cell disease. New England Journal of
Medicine. 340:1021-1030
Steinberg, M. H. (2003). Therapies to increase fetal hemoglobin in sickle cell disease.
Current Hematology Report 2: 95–101

165
Steinberg, M. H and Brugnara, C. (2003). Pathophysiological-based approaches to treatment
of sickle cell disease. Annual Review of Medicine. 54: 89–112
Steinberg, M. H., Barton, F., Castro, O., Pegelow, C.H., Ballas, S.K., Kutlar, A., Orringer,
E., Bellevue, R., Olivieri, N., Eckman, J., Varma, M., Ramirez, G., Adler, B., Smith, W.,
Carlos, T., Ataga, K., DeCastro, L., Bigelow, C., Saunthararajah, Y., Telfer, M., Vichinsky,
E., Claster, S., Shurin, S., Bridges, K., Waclawiw, M., Bonds, D and Terrin, M. (2003).
Effect of hydroxyurea on mortality and morbidity in adult sickle cell anemia: risks and
benefits up to 9 years of treatment. Journal of the American Medical Association. 289: 1645–
1651
Stocker, J. W., De Franceschi, L., McNaughton-Smith, G. A., Corrocher, R., Beuzard, Y and
Brugnara, C. (2003). ICA-17043, a novel Gardos channel blocker, prevents sickled red blood
cell dehydration in vitro and in vivo in SAD mice. Blood 101: 2412–2418
Surassmo, S., Nukoolkarn, V., Gritsanapan, W and Chomnawang, M. T. (2005). Effects of
Thai Medicinal Plants on Inflammation caused by Acne-Inducing Bacteria 31st Congress on
Science and Technology of Thailand at Suranaree University of Technology
Thevenin, B. J. M., Crandall, I., Ballas, S. K., Sherman, I. W and Shohet, S. B. (1997). Band
3 peptides block the adherence of sickle cells to endothelial cells in vitro. Blood. 90:4172-
4179.
Thomas, K. D and Ajani, B. (1987). Antisickling agent in an extract of unripe pawpaw fruit
(Carica papaya) 1987; Transactions of the Royal Society of Tropical Medicine & Hygiene.
81: 510-511.
Thrall, M. A. (2004). Clinical Chemistry of Mammals: Laboratory Animals and
Miscellaneous Species. In Veterinary Hematology and Clinical Chemistry. Edited by Troy
DB. Baltimore, MD: Lippincott Williams and Wilkens, 4:464-468.
Tiffert, T and Lew, V. L. (1997). Cytoplasmic Ca2_ buffers in intact human red cells.
Journal of Physiology. 500: 139–154.
Tiffert, T and Lew, V. L. (2001). Kinetics of Inhibition of the plasma membrane calcium
pump by vanadate in intact human red blood cells. Cell calcium. 30(5): 337-342
Tiffert, T., Daw, N., Perdomo, D and Lew, V. L. (2001). A fast and simple screening test to
search for specific inhibitors of the plasma membrane calcium pump. The Journal of
laboratory and clinical medicine. 137: 199–207
Todd, M. D., Lin, X., Stankowski, L. F., Desai, M and Wolfgang, G. H. I. (1999). Drug
discovery. Journal of Biomolecular Screening. 4:259-268.
Todd, K. H., Green, C., Bonham, V. L Jr., Haywood, C Jr and Ivy, E. (2006). Sickle cell
disease related pain: crisis and conflict. Journal of Pain, 7(7):453-8.
Tosteson, D. C., Carlsen, E., Dunham, E. T. (1955). The effects of sickling on ion transport:
I. Effect of sickling on potassium transport. Journal of General Physiology 39:31.
Trease, G. E and Evans, W. C. (2002). Pharmacognosy 15th
ed. W.B Saunders, Edinburgh;
585pp

166
Trivedi, P. P., Kushwaha, S., Tripathi, D. N and Jena, G. B. (2010). Evaluation of male germ
cell toxicity in rats: correlation between sperm head morphology and sperm comet assay.
Mutation Research. 703(2): 115-21
Turhan, A., Weiss, L. A., Mohandas, N., Coller, B. S and Frenette, P. S. (2002). Primary role
for adherent leukocytes in sickle cell vascular occlusion: a new paradigm. Proceedings of the
National Academy of Science. U. S. A. 99:3047-3051.
Uddin, S. N., Ali, M. E and Yesmin, M. N. (2008). Antioxidant and antibacterial activities of
Senna tora roxb. American Journal of Plant Physiology. 3:96 – 100.
Ugbor, C. (2006). The effect of vegetable extracts on the antisickling potential of Aloe vera.
www.biochemistry.org/meetings/abstracts/BS2006/BS20060567.pdf.
Unver, A., Arslam, D., Ozcan, M. M and Akbulut, M. (2009). Phenolic content and
antioxidant activity of some spices. World Applied Science Journal. 6(3): 373-377.
Upur, H., Yusup, A., Baudrimont, I., Umar, A., Berke, B., Yimit, D., Lapham, J. C., Creppy,
E. E and Moore, N. (2008). Inhibition of cell growth and cellular protein, DNA and RNA
synthesis in human hepatoma (HepG2) cells by ethanol extract of abnormal Savda Munziq of
traditional Uighur medicine. eCAM, doi:10.1093/ecam/nen062
Wambebe, C., Khamofu, H., Momoh, J.A., Ekpeyong, M., Audu, B.S., Njoku, S.O.,
Nasipuri, N.R., Kunle, O.O., Okogun, J.I., Enwerem, N.M., Gamaniel, S.K., Obodozie, O.O.,
Samuel, B., Fojule, G., Ogunyale, P.O. (2001). Double-blind, placebo-controlled, randomized
cross-over clinical trial of NIPRISAN in patients with sickle cell disorder. Phytomedicine.
8(4):252-261
Wang, R. H., Phillips, G. Jr, Medof, M. E and Mold, C. (1993). Activation of the alternative
complement pathway by exposure of phosphatidylethanolamine and phosphatidylserine on
erythrocytes from sickle cell disease patients. Journal of Clinical Investigation. 92:1326.
Wang. H., Gao, X. T., Guo, C. Z., Cai, L and Yao, W. B. (2008). In vitro and in vivo
antioxidant activity of aqueous extract from Choerospondias axillaries fruit. Food Chemistry.
106:888–95.
Ware, R. (2007). Principles and Indications of Transfusion therapy for children with Sickle
Cell Disease. Clinical Advances in Haematology and Oncology. 5: 686 - 688
Ware, R. E (2010). How I use hydroxyurea to treat young patients with sickle cell anemia.
Blood. 115(26): 5300–5311.
Ware, R. E., Zimmerman, S. A and Schultz, W. H. (1999). Hydroxyurea as an alternative to
blood transfusions for the prevention of recurrent stroke in children with sickle cell disease.
Blood.94:3022–3026.
Watson, J., Stahman, A. W and Bilello, F. P. (1948). The significance of the paucity of sickle
cells in newborn Negro infants. The American journal of the medical sciences. 215:419-423.

167
Wegwu, M. O., Ayalogu, E. O. and Sule, O. J. (2005). Anti-Oxidant Protective Effects of
Cassina alata in Rats Exposed to Carbon Tetrachloride. Journal of Applied Science and
Environmental Management. 9(3): 77-80.
Weiner, D. L., Hibberd, P. L., Betit, P., Cooper, A. B., Botelho, C. A., Brugnara, C. (2003).
Preliminary assessment of inhaled nitric oxide for acute vaso-occlusive crisis in pediatric
patients with sickle cell disease. The Journal of the American Medical Association.
289:1136–1142.
Weiner, D.L and Brugnara, C. (2003). Hydroxurea and sickle cell disease. A chance for every
patient. The Journal of the American Medical Association. 289:1692-1694
Weinfeld, A., Swolin, B. and Westin, J. (1994). Acute leukaemia after hydroxyurea therapy
in polycythaemia vera and allied disorders: prospective study of efficacy and
leukaemogenicity with therapeutic implications. European Journal of Haematology. 52: 134-
139
WHO. (1980). Expert committee on diabetes mellitus. 2nd
Report. Technical report series
646. World Health Organization. Geneva. :12
Wong, W. Y., Elliott-Mills, D and Powars, D. (1996). Renal failure in sickle cell anemia.
Hematol Oncol Clin North Am 10:1321, 1996.
World Health Organisation (2001). Legal status of Traditional medicine and
Complementary/Alternative Medicine: A World wide review. WHO Publishing 1.
World Health Organisation (2006). Fifty-ninth World Health Assembly. Sickle-cell anaemia
Report by the Secretariat, Geneva. A59/9
World Health Organization (1977). Resolution-Promotion and development of Training and
Research in Traditional Medicine. WHO document No.:30-49
World Health Organization (1999). WHO monographs on selected medicinal plants.Vol 1,
WHO Geneva. p1-8.
World Health Organization (1999). World Health Organization Monograph on selected
Medicinal Plants. WHO Graphics, Malta. 1: 1-295.
World Health Organization (2002). World Health Organization Monograph on selected
Medicinal Plants. WHO Graphics, Malta. 2: 1-358.
World Health Organization (2006). Sickle Cell Anemia Report by Secretariat, Geneva.
Wright, J and Ahmedzai, S. H. (2010). The management of painful crisis in sickle cell
disease. Current Opinion in Supportive and Palliative Care. 4(2):97-106.
Yamaja Setty, B. N and Betal, S. G. (2008). Microvascular endothelial cells express a
phosphatidylserine receptor: a functionally active receptor for phosphatidylserine-positive
erythrocytes. Blood. 111(2): 905–914.

168
Yasin Z, Witting S, Palascak, M. B., Joiner, C. H., Rucknagel, D. L and Franco, R. S. (2003).
Phosphatidylserine externalization in sickle red blood cells: associations with cell age,
density, and hemoglobin F. Blood. 102:365-370.
Yumrutas, O and Saygideger, S. D. (2012). Determination of antioxidant and antimutagenic
activities of Phlomis armeniaca and Mentha pulegium. Journal of Applied Pharmaceutical
Science 2 (01): 36-40.
Zheng, W and Wang, S.Y. (2001). Antioxidant Activity and Phenolic Compounds in Selected
Herbs. Journal Agricultural Food Chemistry. 49: 5165-5170
Zimmerman, S.A., Schultz, W. H., Davis, J. S., Pickens, C. V., Mortier, N. A., Howard, T. A
and Ware, R. E. (2004). Sustained long-term hematologic efficacy of hydroxyurea at
maximum tolerated dose in children with sickle cell disease. Blood.103: 2039-2045.

169

170

171
Photograph of Senna alata

172
Photograph of Senna podocarpa




















