
朱林山 PB12207061
11
of March 14, 2014. This information is current as Gag-Specific CD8 T Cell Responses Cells from HIV Controllers: Association with Heterogeneity in HIV Suppression by CD8 T Lambotte, Alain Venet and Gianfranco Pancino Jean-François Delfraissy, Françoise Barré-Sinoussi, Olivier Boufassa, Véronique Avettand-Fènoël, Christine Rouzioux, Urrutia, Pierre Versmisse, Christine Lacabaratz, Faroudy Asier Sáez-Cirión, Martine Sinet, So Youn Shin, Alejandra http://www.jimmunol.org/content/182/12/7828 doi: 10.4049/jimmunol.0803928 2009; 182:7828-7837; ; J Immunol References http://www.jimmunol.org/content/182/12/7828.full#ref-list-1 , 33 of which you can access for free at: cites 46 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2009 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology at NSTL China Trial on March 14, 2014 http://www.jimmunol.org/ Downloaded from at NSTL China Trial on March 14, 2014 http://www.jimmunol.org/ Downloaded from
Transcript of 朱林山 PB12207061

of March 14, 2014.This information is current as
Gag-Specific CD8 T Cell ResponsesCells from HIV Controllers: Association with Heterogeneity in HIV Suppression by CD8 T
Lambotte, Alain Venet and Gianfranco PancinoJean-François Delfraissy, Françoise Barré-Sinoussi, Olivier Boufassa, Véronique Avettand-Fènoël, Christine Rouzioux,Urrutia, Pierre Versmisse, Christine Lacabaratz, Faroudy Asier Sáez-Cirión, Martine Sinet, So Youn Shin, Alejandra
http://www.jimmunol.org/content/182/12/7828doi: 10.4049/jimmunol.0803928
2009; 182:7828-7837; ;J Immunol
Referenceshttp://www.jimmunol.org/content/182/12/7828.full#ref-list-1
, 33 of which you can access for free at: cites 46 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
at N
STL
China T
rial on March 14, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Heterogeneity in HIV Suppression by CD8 T Cells from HIVControllers: Association with Gag-Specific CD8 T CellResponses1
Asier Saez-Cirion,2* Martine Sinet,3†‡ So Youn Shin,3* Alejandra Urrutia,†‡ Pierre Versmisse,*Christine Lacabaratz,4†‡ Faroudy Boufassa,§ Veronique Avettand-Fenoel,¶�
Christine Rouzioux,¶� Jean-Francois Delfraissy,†‡# Francoise Barre-Sinoussi,*Olivier Lambotte,†‡# Alain Venet,†‡ and Gianfranco Pancino*; for the ANRS EP36 HIVControllers Study Group
“HIV controllers” (HICs) are rare individuals in whom HIV-1 plasma viral load remains undetectable without antiretroviraltreatment. This spontaneous viral control in HICs is usually associated to strong functional HIV-specific CD8� T cell responses.Accordingly, we have recently shown that CD8� T cells from HICs strongly suppress ex vivo HIV-1 infection of autologous CD4�
T cells, suggesting a crucial role of this response in vivo. Knowledge of the mechanisms underlying the CD8� T cell antiviralactivity might help to develop effective T cell-based vaccines. In the present work, we further characterized the HIV-suppressivecapacity of CD8� T cells in 19 HICs. CD8� T cells from 14 of the 19 HICs showed strong HIV-suppressive capacity ex vivo. Thiscapacity was stable over time and was partially effective even on other primate lentiviruses. HIV-suppressive capacity of CD8�
T cells correlated strongly with the frequency of HIV-specific CD8� T cells, and in particular of Gag-specific CD8� T cells. Wealso identified five HICs who had weak HIV-suppressive CD8� T cell capacities and HIV-specific CD8� T cell responses. Amongthese five HICs, at least three had highly in vitro replicative viruses, suggesting that the control of viremia in these patients is notdue to replication-defective viruses. These results, on the one hand, suggest the importance of Gag responses in the antiviralpotency of CD8� T cells from HICs and, on the other hand, propose that other host mechanisms may contribute to restrainingHIV infection in HICs. The Journal of Immunology, 2009, 182: 7828–7837.
A t least partial control of HIV can be achieved by CD8�
T cells (1–3). Some HLA class I molecules, particularlyalleles B27 and B57, have been linked to better control
of HIV infection (4–6). The presence of Gag-specific CD8� Tcells and the breadth of their specificities have also been linked tolow HIV viremia (7–9). One of the most compelling indications ofthe pressure exerted by CD8� T cell responses is the emergence ofvariants that escape recognition by these cells (10–14). However,most HIV-infected individuals have uncontrolled viremia andeventually progress to AIDS despite strong CD8� T cellresponses.
Rare individuals called “HIV controllers” (HICs)5 spontane-ously and durably control HIV infection in the absence of therapy,possibly illustrating what truly effective CD8� T cell responsescan achieve (15, 16). HICs have extremely low and stable amountsof viral DNA in their PBMC (17) and undetectable plasma viralload (18). The protective HLA alleles B27 and B57 are overrep-resented among these individuals (4, 6, 19–21). Despite very lowlevels of Ag in blood (22), most but not all HICs have high fre-quencies of HIV-specific CD8� T cells that preferentially targetthe viral Gag protein (20, 23, 24). Studies of CD8� T cell re-sponses in HICs have revealed important characteristics of func-tional HIV-specific CD8� T cells in HIV infection. Contrary tocells from viremic individuals, HIV-specific CD8� T cells fromHICs can, upon stimulation with their cognate Ag, proliferate andgenerate a multifunctional response that includes perforin expres-sion, degranulation, and chemokine/cytokine secretion (25–27).This could be related to a peculiar activation phenotype of thesecells (21) and to constitutive telomerase activity that protects themagainst senescence (28). However, how much of this is the causeand how much the consequence of viral control and low-level im-mune activation remains to be determined. We have recentlyshown that CD8� T cells from most HICs are endowed with astriking capacity to suppress HIV infection ex vivo (21), a propertythat is likely to be relevant in vivo. To further characterize thisHIV-suppressive activity we extended our analysis to a larger
*Institut Pasteur, Unite de Regulation des Infections Retrovirales, Paris, France;†INSERM Unite 802, Le Kremlin-Bicetre, France; ‡Universite Paris-Sud, Facultede Medecine Paris XI, Le Kremlin-Bicetre, France; §INSERM Unite 822, HopitalBicetre, Le Kremlin-Bicetre, France; ¶AP-HP, CHU Necker-Enfants Malades,Laboratoire de Virologie, Paris, France; �Universite Paris-Descartes, Faculte deMedecine, Paris, France; #AP-HP, Hopital Bicetre, Service de Medecine Interneet Maladies Infectieuses, Le Kremlin-Bicetre, France
Received for publication November 25, 2008. Accepted for publication April 3, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was financially supported by the Agence Nationale de Recherches sur leSIDA. S.Y.S. was supported by Korea Science and Engineering Foundation and theInstitut Pasteur Korea.2 Address correspondence and reprint requests to Dr. Asier Saez-Cirion, Unite deRegulation des Infections Retrovirales, Institut Pasteur, 25 rue du Dr Roux, 75725Paris Cedex 15, France. E-mail address: [email protected] M.S. and S.Y.S. contributed equally to this work.4 Current address: INSERM Unite 841, Faculte de Medecine Henri Mondor, CreteilFrance.
5 Abbreviations used in this paper: HIC, HIV controller; moi, multiplicity of infec-tion; PBMC, peripheral blood mononuclear cell; SFC, spot-forming cell.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0803928
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
group of 19 HICs and evaluated the relationship between this ac-tivity and HIV-specific CD8� T cell responses.
Materials and MethodsStudy subjects
Nineteen patients diagnosed with HIV-1 infection at least 10 years previ-ously who had never received antiretroviral treatment and in whom � 90%of plasma HIV RNA assays gave values�400 copies/ml were studied (Ta-ble I): 8 have been described elsewhere (21), and 11 were newly recruitedfrom the ANRS EP36 national monitoring program on HIV controllers.The subjects were serologically HLA typed by complement-mediated lym-phocytotoxicity testing (InGen One Lambda). All had very weak and stableDNA load (Table I).
All of the subjects gave their written informed consent.
HIV DNA quantification
Total DNA was extracted from whole blood with QIAamp DNA minikits(Qiagen), according to the manufacturer’s instructions. HIV-1 DNA wasthen quantified by real-time PCR (LTR amplification; Agence Nationale deRecherches sur le SIDA) (29). Four PCRs, each testing 1 �g of total DNA,were performed per extract in this ultrasensitive assay (threshold of 10copies/million leukocytes) (30).
Primary cell culture
CD4� and CD8� cells were purified (�97%) from freshly isolated PBMCby positive and negative selection, respectively, with Ab-coated magneticbeads (Miltenyi Biotec). CD4� cells were stimulated for 3 days with phy-tohemagglutinin (PHA) at 2 �g/ml in the presence of IL-2 (Chiron) at 100IU/ml. The culture medium was RPMI 1640 containing 10% FCS andpenicillin/streptomycin (100 U/ml). CD8� T cells were kept in culturewithout mitogens or cytokines.
Productive infection in vitro
CD4� T cells (105) were superinfected with HIV-1 BaL (R5) in triplicateat a multiplicity of infection (moi) of 10�3.6 in 96-well plates with a spi-noculation protocol (31). For some experiments, SIVagm.Gril, SIV-mac.251, and HIV-2.SBL and autologous primary viruses were used forinfection. For coculture, 105 CD4� T cells were mixed with 105 CD8� Tcells (CD8/CD4 ratio of 1:1) at the moment of infection. After challengethe cells were washed and cultured for 14 days. Viral replication was mon-itored every 3–4 days in supernatants by p24 or p27 ELISA (Zeptometrix).Infectivity assays were conducted in the presence of 100 IU/ml IL-2. We
have previously shown that the presence of this cytokine during the infec-tivity assays did not affect the suppressive capacity of unstimulated CD8�
T cells (21).
Intracellular p24 assay
Activated CD4� lymphocytes (5 � 104) were superinfected with HIV-1BaL (R5) as described above. Various dilutions of virus (moi of 10�1.6 to10�2.6) were used in parallel to obtain similar levels of infection in eachindividual/experiment. CD4� T cells were culture in the presence or ab-sence of unstimulated CD8� T cells (CD8/CD4 ratio of 1:1).
Seventy-two hours after infection, cells were harvested and stained withCD4-ECD (SFCI12T4D11) and CD8-PC5 (B9.11). Cells were then perme-abilized (Cytofix/Cytoperm fixation and permeablization kit; BD Biosciences)and stained with KC57-FITC (FH190-1-1) to detect intracellular HIV Ags.Abs were from Beckman Coulter. Flow cytometry was performed with a Cy-tomix FC500 and CXP acquisition software (Beckman Coulter).
Viral isolation from peripheral blood CD4� T cells
Between 2 and 5 � 106 CD4� T cells from each patient were activatedwith PHA and IL-2 as described above. Viral production in culture super-natants was monitored for 28 days by p24 ELISA. When required, CD4�
T cells were reactivated on day 10 with CD8-depleted PHA-preactivatedallogeneic PBMC, PHA, and IL-2. Virus-containing supernatants fromCD4 T cell cultures were titrated on mixed PHA-activated CD4� T cellsfrom two blood donors.
ELISPOT assay
IFN-� secretion by HIV-specific CD8� T cells was quantified ex vivo withan ELISPOT assay using appropriate stimuli (32). We used a set of 124peptides corresponding to known optimal CTL epitopes derived from theHIV-1 Env, Gag, Pol, and Nef proteins (National Institutes of Health HIVMolecular Immunology Database; www.hiv.lanl.gov/content/immunology/index.html). These peptides were synthesized by Neosystem and used at afinal concentration of 2 �g/ml. For each subject, optimal peptides weretested depending on the results of HLA typing with an average of 36 � 9peptides tested per subject. IFN-� spot-forming cells (SFCs) were countedwith a KS-ELISPOT system (Carl Zeiss Vision) and expressed as SFCs/106 PBMC after subtracting the background of control unstimulated cells.Wells were considered positive if they contained at least 50 SFCs/106
PBMC and exhibited at least twice the background level.
Depletion of HIV-specific CD8� T cells
Depletion of CD8� T cells producing IFN-� upon stimulation with HIVpeptides was performed with an IFN-� secretion assay enrichment kit
Table I. Characteristics of HIV controllers included in the study
Patienta SexAge
(years)HIV
DiagnosedMode of
Infectionb
Subtype ofInfecting
Virus HLAMedian CD4 Count(first–last) (cells/ml)
ViralFollow-up
(years)
Median RNAViral Load(copies/ml)
Viral LoadBlips (50–400
copies/ml)c
HIV DNA(log copies/106
cells)
A6 M 69 1985 Hem B A2/A74/B27/B57 1041 (775-1528) 11 �50 7/16 NAA9 M 45 1992 MMS B A2/A29/B27/B57 844 (1107-522) 15 �50 0/13 NAA12 M 46 1986 IDU NA A1/A2/B44/B57 632 (1151-632) 10 �50 4/13 1.67A21 M 45 1987 Unknown B A1/A2/B51/B57 839 (1217-785) 10 �50 9/18 1.62A7 M 39 1983 Hem B A2/B44/B57 613 (601-408) 11 �50 7/18 1.77A2 M 49 1985 Het B A3/A23/B7/B57 709 (599-493) 6 �50 1/12 0.5A1 F 49 1988 Het B A2/B27/B57 958 (1290-849) 12 �50 1/15 0.5A4 M 48 1987 IDU B A2/A32/B27/B60 597 (602-649) 10 �50 0/15 1.67B5 M 59 1989 MMS B A2/A68/B14/B57 890 (1000-937) 12 �50 0(2)/16 1.31A15 F 50 1987 Het B A3/A30/B27/B51 640 (883-330) 11 �50 2/20 1.04A3 F 42 1993 Het B A3/A30/B57/B63 1035 (960-1033) 11 �50 0/11 1.04A18 F 44 1996 Het NA A2/A31/B39/B57 896 (665-877) 11 �50 3(1)/16 1.04A17 M 46 1989 IDU B A2/B7/B57 1194 (601-1196) 11 �50 0/8 1.94A11 M 44 1989 IDU B A3/A30/B7/B57 504 (390-556) 11 �50 2/19 1.28
A16 M 50 1985 IDU B A24/A32/B27/B62 976 (1107-1142) 9 �50 0/17 2.08A13 M 47 1992 MMS B A3/A29/B35/B44 727 (653-828) 11 �100 0/21 1.64A23 F 38 1994 Unknown NA A43/A74/B57/B72 771 (902-773) 9 �50 0/11 0.5A22 F 40 1991 Het NA A11/A30/B13/B27 800 (800-717) 10 �50 0/9 NAA19 F 39 1994 Het A2 A2/A68/B60/B14 928 (790-808) 12 �50 0/10 1.57
a Patients are sorted in function of the suppressive capacity of their CD8� T cells (log p24 decrease, Fig. 1C). Horizontal rule separates strong and weak responders.b Hem, hemophilia; Het, heterosexual sex; IDU, injection drug use; MMS, male-male sex.c Number of viral load between 50 and 400 copies per total determinations with detection limit �50 copies/ml. Viral loads of �400 RNA copies/ml are indicated in
parentheses.
7829The Journal of Immunology
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
(Miltenyi Biotec) as recommended by the manufacturer. Briefly, purifiedCD8� T cells were stimulated for 6 h with appropriate pools of specificHIV peptides. Subsequently, the cells were labeled (5 min at 4°C) with anIFN-� catch reagent that attached to the cell surface of all leukocytes. Thecells were then incubated for 45 min at 37°C to allow IFN-� secretion. Thesecreted IFN-� was captured by the IFN-� catch reagent on the positive, se-creting cells. These cells were subsequently labeled with a second IFN-�-specific Ab conjugated to R-PE. The IFN-�-secreting cells were magneticallylabeled with anti-PE magnetic beads and depleted by magnetic field separa-tion. Purity of the depleted fractions was evaluated by flow cytometry.
Antibodies
The following Abs were used: CD8-ECD or -PC5 (clone B9.11), CD3-PC5(UCHT1), CD45RO-ECD (UCHL1), HLA-DR-ECD (Immu-357), andCD38-FITC (T16), all from Beckman Coulter; and CD27-FITC (M-T271)from BD Biosciences.
Pentamer staining and phenotyping
HIV-specific CD8� T cells were identified by using soluble PE- orallophycocyanin-labeled peptide-HLA class 1 multimers (Proimmune;Beckman Coulter Immunomics). The following epitopes were used: theHLA-A*0201-restricted peptide ligands SLYNTVATL (Gag 77– 85)and ILKEPVHGV (Pol 476 – 484), the A*0301-restricted peptide li-gands RLRPGGKKK (Gag 20 –28) and QVPLRPMTYK (Nef 73– 82),
the B*2705-restricted peptide ligand KRWIILGLNK (Gag 263–272),and the B*5701-restricted peptide ligands KAFSPEVIPMF (Gag 162–172), TSTLQEQIGW (Gag 240 –249), and QASQDVKNW (Gag 308 –316). PBMC were incubated with pentamers (1 �g/ml) for 30 min andthen with relevant Abs for 15 min. Cells were washed in Cell Wash (BDBiosciences) plus 1% BSA, incubated for 10 min with FACS lysingsolution (BD Biosciences). After washing, cells were fixed in 1% para-formaldehyde for flow cytometry with an Epics XL (Beckman Coulter)or a FACSCanto flow cytometer (BD Biosciences) and analyzed withRXP software (Beckman Coulter).
Proliferation assay
The proliferative capacity of HIV-specific CD8� T cells was evaluated byflow cytometry. PBMC were stained with 0.35 �M CFSE (MolecularProbes) for 10 min at 37°C, and, after washing they were stimulated for 5days with 2 �g/ml peptide or medium alone. After labeling with pentamer,anti-CD8, and anti-CD3 Abs, PBMC were fixed in 1% paraformaldehydefor flow cytometry as described above.
Statistical analyses
All values throughout the text are means � SD. Values of p were calcu-lated with the rank sum test. Correlations were identified by simple linearregression analysis and Spearman’s rank correlation test. SigmaStat 3.5software was used (Systat Software).
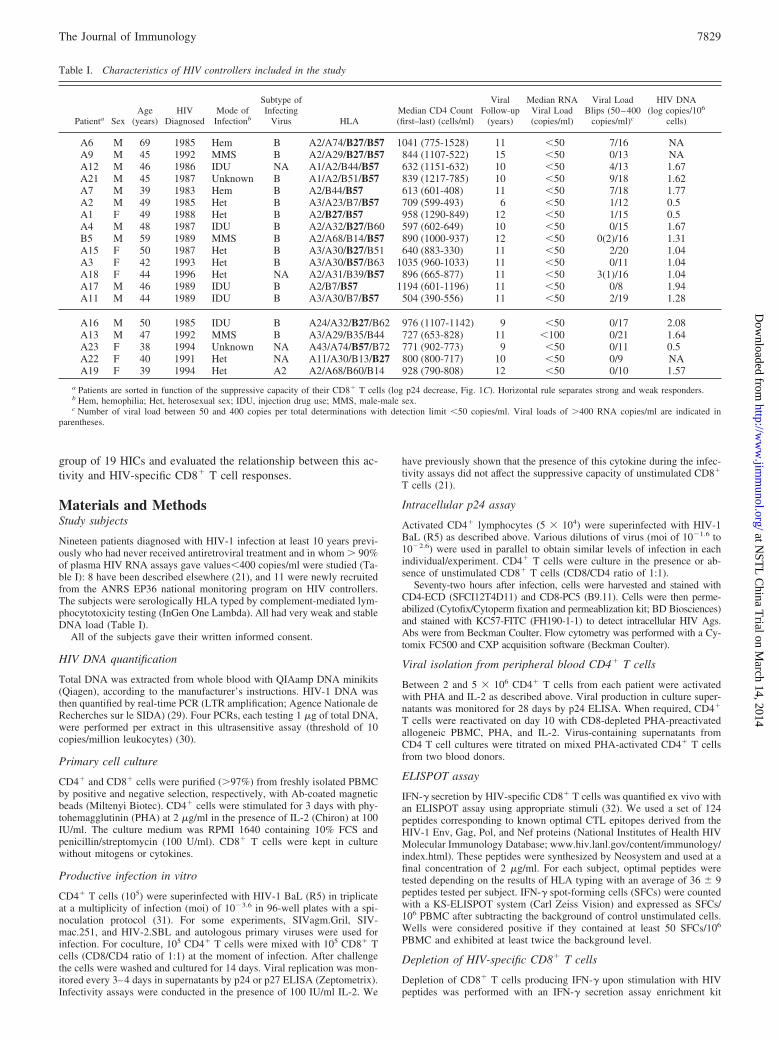
FIGURE 1. A, PHA-activated CD4� T cells were infected, in the absence (F) or presence (E) of autologous unstimulated CD8� T cells (1:1 ratio), withreplicative HIV-1 BaL. Circles represent the average (n � 3 independent infections) peak p24 values detected in culture supernatants for each individual.Horizontal lines indicate median values. B, PHA-activated CD4� T cells from HICs A6 and A13 were superinfected with HIV-1 Bal and left alone (centralpanels) or cocultured with autologous unstimulated CD8� T cells (right panels). Three days later the level of infection was determined by quantifyingintracellular p24 on CD8� cells. C, The HIV-suppressive capacity of CD8� T cells, as determined by the log fold decrease in the level of secreted p24(CD4 vs CD4/CD8 cell cultures), was compared in the 19 HICs (F), 13 chronically HIV-infected subjects with viremia �7000 copies/ml (Œ), and 8HAART-treated patients with virologic control (plasma HIV RNA �50 copies/ml) for �23 mo (f). Horizontal lines indicate median values. D, p24production in culture supernatants (mean � SD, n � 3) at the peak of viral replication after superinfection of CD4� T cells from HIV controllers withequivalent infectious doses (moi of 10�3.6) of HIV-1 Bal or filtered supernatants containing autologous HIC viruses. CD4� T cells were cultured alone(filled bars) or in the presence of non prestimulated CD8� T cells (open bars).
7830 HETEROGENEITY OF CD8 T CELL HIV SUPPRESSION IN HIV CONTROLLERS
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

ResultsUnstimulated CD8� T cells from most HICs have strongHIV-suppressive capacity
In a previous study we found that undetectable viremia in 9 out of10 HICs was associated with a remarkably strong capacity of theircirculating CD8� T cells to control in vitro HIV-1 infection ofautologous CD4� T cells (21). To extend this observation, we usedthe same viral suppression assay to assess the ex vivo anti-HIVcapacity of CD8� T cells from 19 HICs, 11 of whom were newlyrecruited for this study and 8 were retested (Table I). Viral repli-cation was readily detected in the supernatants of purified CD4� Tcells from all 19 HICs after PHA activation and challenge withHIV-1 BaL (Fig. 1A). A marked reduction in HIV-1 infection (un-detectable in eight HICs) was generally observed when autologousunstimulated CD8� T cells from HICs were added to the culture(Fig. 1A). The associated CD8� T cell-mediated decrease in thelevel of HIV proteins was due to the absence of infected CD4� Tcells in the coculture (Fig. 1B). As a whole, the HIV-suppressivecapacity of CD8� T cells from HICs (2.79 � 1.31 log p24 de-crease, CD8/CD4 vs CD4) was much stronger than that of cellsboth from viremic individuals (0.82 � 0.53 log p24 decrease,CD8/CD4 vs CD4), confirming our previous results (21), as wellas from HAART-treated individuals with undetectable viral load(0.62 � 0.63 log p24 decrease, CD8/CD4 vs CD4) (Fig. 1C). Inparticular, CD8� T cells from 14 of the 19 HICs suppressed HIVfar more strongly (log p24 decrease �2) than did cells from bothviremic and treated individuals (Fig. 1C). These subjects are re-ferred to below as strong responder HICs. Longitudinal analysis(�12 mo) of CD8� T cell antiviral activity in five strong responder
HICs included in our previous study suggested that this HIV-suppres-sive capacity is a stable characteristic (Table II). In contrast, here weidentified five “weak responder” HICs (Table I) whose CD8� T cellscould not efficiently control HIV infection of autologous CD4� Tcells (log p24 decrease �2) (Fig. 1, B and C): the HIV-suppressivecapacity of these subjects’ CD8� T cells was not stronger than that ofviremic or HAART-treated patients (Fig. 1C). We have reported thatsusceptibility of CD4� T cells from HICs to in vitro HIV infectionwas not different than that of cells from healthy blood donors (21), andno significant differences were found either between weak responderand strong responder HICs ( p � 0.331) (Fig. 1A).
To determine whether the weak HIV-suppressive activity observedin certain HICs was due to our use of a laboratory-adapted HIV strain,we analyzed the capacity of nonstimulated CD8� T cells from weakresponders A13 and A19 and from strong responder A21 to suppresssuperinfection of their own CD4� T cells by autologous viruses pre-viously obtained in primary culture of these individuals’ cells (seebelow). CD8� T cells from strong responder A21 equally controlledCD4� T cell superinfection by HIV-BaL and by autologous virus(Fig. 1D). In contrast, the weak CD8-mediated HIV suppression insubject A13 was not improved when his autologous virus was used tochallenge his CD4� T cells (0.01 vs 0.16 log p24 decrease with HIV-BaL and the autologous virus, respectively) (Fig. 1D). CD8� T cellsfrom weak responder A19 showed a stronger capacity to inhibit in-fection by autologous viruses (0.33 vs 1.76 log p24 decrease for HIVBaL and autologous virus infection, respectively) (Fig. 1D), althoughthe level of suppression did not reach that observed in strong respond-ers. Interestingly, while most HICs were infected by subtype B vi-ruses, subject A19 was infected by HIV-1 subtype A2 (Table I).Therefore, although the use of nonautologous viruses might lead to anunderestimation of the HIV-suppressive activity of CD8� T cells, itwas unlikely to explain the differences observed between weak andstrong responders.
CD8-mediated HIV-suppressive capacity in HICs correlateswith the frequency of IFN-�-producing cells
We examined whether the difference between strong and weak re-sponder HICs was associated with a difference in the magnitude ofHIV-specific CD8� T cell responses. To quantify the HIV-specificCD8� T cell response, we used the standard determination of thefrequency of IFN-�-secreting CD8� T cells upon stimulation with
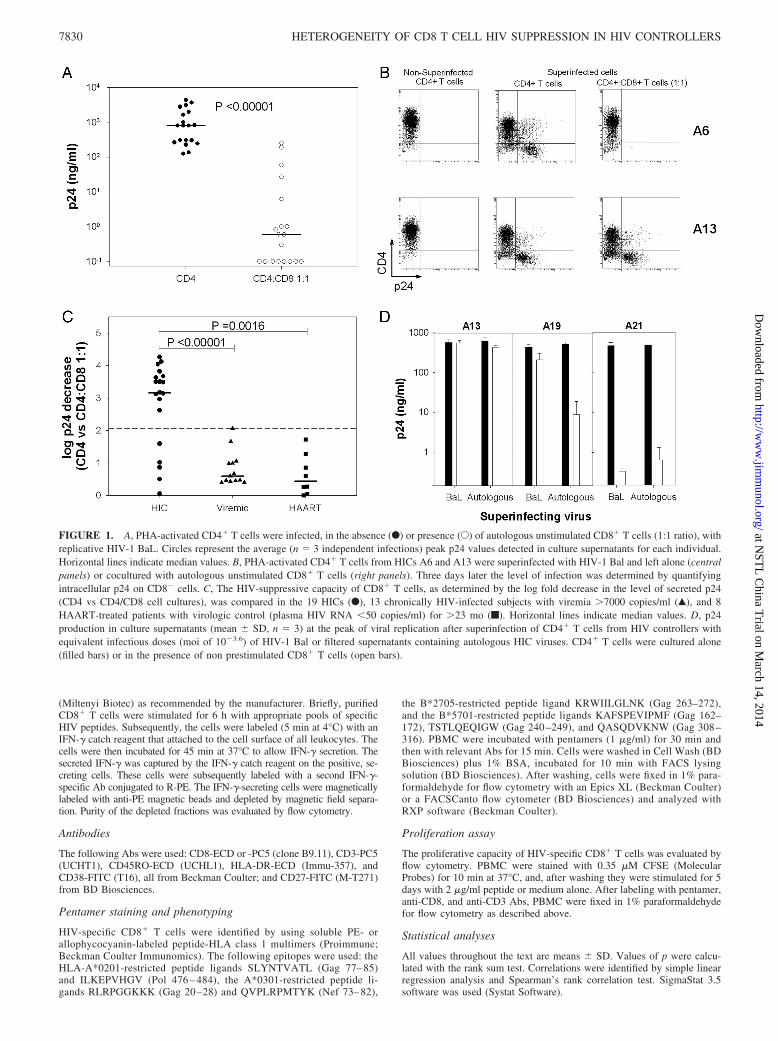
FIGURE 2. A, Frequencies of HIV-specific IFN-�-secreting CD8� T cells in strong responder HICs (log p24 decrease �2) (SR) and in weak responderHICs (log p24 decrease �2) (WR). An average of 36 � 9 peptides were tested in each subject, depending on the results of HLA typing. Each symbolcorresponds to the sum of SFCs/106 PBMC obtained with individual peptides described as being restricted by HLA Ags. Horizontal lines are median valuesfor each group. B, Correlation between the HIV-suppressive capacity of CD8� T cells from HICs (log p24 decrease as shown in Fig. 1C) and their frequencyof IFN-�-producing CD8� T cells upon HIV peptide stimulation. Each symbol represents one HIC. Vertical dashed line separates weak responder andstrong responder HICs. C, Percentage of HIV-specific cells (based on HIV multimer and CD8 expression) from strong and weak responder HICs thatexpressed ex vivo HLA-DR and CD38, coexpressed CD27 and CD45RA, or proliferated (and lost CFSE labeling) after 5 days of peptide stimulation. Eachsymbol represents one specificity for one HIC. Horizontal lines are mean values for each group.
Table II. Log p24 decrease (CD4 vs CD4/CD8 of 1:1) duringfollow-up (�12 mo) of HICs
HIC na Median Value First Sample Last Sample
A1 7 3.5 Sept 2005 4.3 May 2008 3.5A3 3 3.1 Jan 2006 3.1 Jan 2007 3.3A4 5 3.8 Jan 2005 4.2 Apr 2008 3.5A6 7 3.8 Sept 2005 4.3 Nov 2007 3.8B5 7 3.5 Oct 2005 1.2 May 2008 3.5
a Number of blood samples analyzed.
7831The Journal of Immunology
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
appropriate HLA-defined optimal HIV-1 Env, Gag, Pol, and Nef pep-tides in an ELISPOT assay. The numbers of IFN-�-secreting cellswere heterogeneous (Fig. 2A), in agreement with recent reports (20,24). The highest frequencies of HIV-specific CD8� T cells were ob-served in strong responders (8517 � 4038 vs 1058 � 903 SFCs/106
PBMC in weak responder HICs, p � 0.0014) (Fig. 2A). The fre-quency of HIV-specific CD8� T cells in HICs was not significantlydifferent, as a whole (6843 � 4866 SFCs/106 PBMC), from that ob-served in chronically viremic patients (4616 � 4148 SFCs/106 PBMCfor 18 patients with �3 years of infection and plasma viral load�1000 RNA copies/ml, p � 0.20). The magnitude of the CD8� T cellresponse in weak responder HICs was similar to that in HAART-treated patients (865 � 1071 SFCs/106 PBMC for 11 patients with�2 years of treatment and plasma viral load �50 RNA copies/ml,p � 0.50; and Ref. 32).
Interestingly, we found a strong correlation between the fre-quency of IFN-�-producing CD8� T cells upon peptide stimula-tion and the HIV-suppressive capacity of unstimulated CD8� Tcells (Spearman 0.835, p � 0.00001) (Fig. 2B). This supports thepossibility that the ex vivo anti-HIV activity of CD8� T cells fromHICs is driven by HIV-specific cells, in keeping with an MHCclass I-mediated mechanism (21). This correlation further distin-guished strong and weak responder HICs (Fig. 2B).
We explored whether differences could also be observed be-tween strong and weak responder HICs at the phenotypical level oftheir HIV-specific CD8� T cells. Due to the low frequency ofthese cells in weak responders, we could perform these analysesonly in three of them. HIV-specific CD8� T cells from strongresponder HICs possessed a discordant activation phenotype withhigh expression of the activation marker HLA-DR associated witha low CD38 expression (Fig. 2C), in keeping with our previousstudy (21). In contrast, the expression of both activation markerswas low in the cells from weak responders (Fig. 2C), a phenotypethat is found in HAART subjects (21). HIV-specific CD8� T cellsfrom both strong and weak responders had high proliferative po-tential (Fig. 2C), which is a hallmark of a high-quality HIV-spe-cific CD8� T cell response in HICs (26). Interestingly, we foundin weak responders an increase of a subpopulation of HIV-specificCD8� T cells characterized by the coexpression of CD27 andCD45RA (Fig. 2C). We have recently reported that this subpopu-lation is characteristically abundant in HIV patients treated duringacute primary HIV infection and may represent a stable quiescentlong-term memory pool (33).
CD8-mediated HIV-suppressive capacity in HICs correlatesstrongly with the magnitude of Gag-specific CD8� T cellresponses
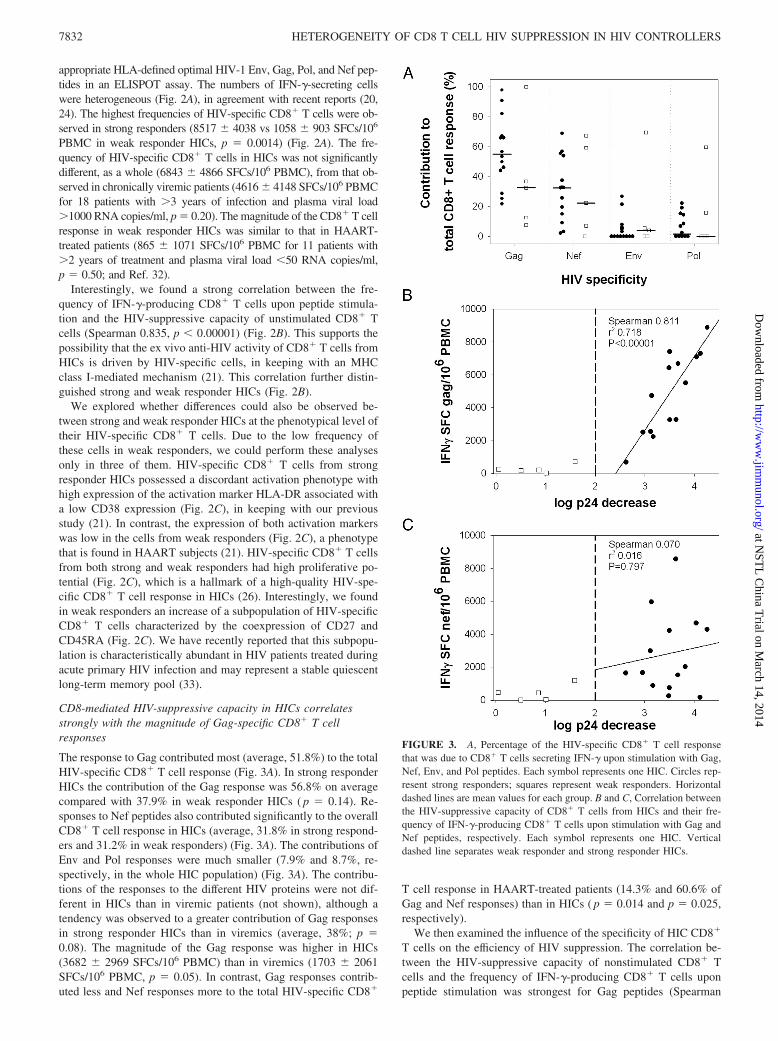
The response to Gag contributed most (average, 51.8%) to the totalHIV-specific CD8� T cell response (Fig. 3A). In strong responderHICs the contribution of the Gag response was 56.8% on averagecompared with 37.9% in weak responder HICs ( p � 0.14). Re-sponses to Nef peptides also contributed significantly to the overallCD8� T cell response in HICs (average, 31.8% in strong respond-ers and 31.2% in weak responders) (Fig. 3A). The contributions ofEnv and Pol responses were much smaller (7.9% and 8.7%, re-spectively, in the whole HIC population) (Fig. 3A). The contribu-tions of the responses to the different HIV proteins were not dif-ferent in HICs than in viremic patients (not shown), although atendency was observed to a greater contribution of Gag responsesin strong responder HICs than in viremics (average, 38%; p �0.08). The magnitude of the Gag response was higher in HICs(3682 � 2969 SFCs/106 PBMC) than in viremics (1703 � 2061SFCs/106 PBMC, p � 0.05). In contrast, Gag responses contrib-uted less and Nef responses more to the total HIV-specific CD8�
T cell response in HAART-treated patients (14.3% and 60.6% ofGag and Nef responses) than in HICs ( p � 0.014 and p � 0.025,respectively).
We then examined the influence of the specificity of HIC CD8�
T cells on the efficiency of HIV suppression. The correlation be-tween the HIV-suppressive capacity of nonstimulated CD8� Tcells and the frequency of IFN-�-producing CD8� T cells uponpeptide stimulation was strongest for Gag peptides (Spearman
FIGURE 3. A, Percentage of the HIV-specific CD8� T cell responsethat was due to CD8� T cells secreting IFN-� upon stimulation with Gag,Nef, Env, and Pol peptides. Each symbol represents one HIC. Circles rep-resent strong responders; squares represent weak responders. Horizontaldashed lines are mean values for each group. B and C, Correlation betweenthe HIV-suppressive capacity of CD8� T cells from HICs and their fre-quency of IFN-�-producing CD8� T cells upon stimulation with Gag andNef peptides, respectively. Each symbol represents one HIC. Verticaldashed line separates weak responder and strong responder HICs.
7832 HETEROGENEITY OF CD8 T CELL HIV SUPPRESSION IN HIV CONTROLLERS
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

0.907, p � 0.00001) (Fig. 3B). This correlation was unlikely to bedue to a bias for HLA-B57-restricted Gag responses since in the 13individuals carrying this HLA allele, HIV-specific CD8� T cellresponses targeting HLA-B57 restricted Gag epitopes represented,on average, 26 � 12% of their total response. Other responseswere either restricted by HLA-B57 but not directed at Gag (19 �18%) or restricted by other alleles and directed at Gag (28 � 28%of the response) or at other proteins (27 � 20%). As mentioned,Nef was also a main target of the HIV-specific CD8� response;however, only a weak correlation was found with the magnitude ofNef responses (Spearman 0.473, p � 0.040) (Fig. 3C). Further-more, this correlation with Nef responses was completely lostwhen weak responder HICs were excluded from the analyses(Spearman 0.070, p � 0.797). Most interestingly, in the group ofstrong responder HICs, the CD8� T cell anti-HIV capacity stillcorrelated more tightly with the magnitude of Gag responses (Spear-man 0.812, p � 0.00001) than with the total frequency of IFN-�-producing CD8� T cells (Spearman 0.634, p � 0.007). Overall theseresults suggest that the numbers of CD8� T cells responding to Gagepitopes influence the capacity of CD8� T cells from HICs to sup-press HIV infection of autologous CD4� T cells.
To evaluate more directly the impact of Gag responses in theHIV-suppressive activity of CD8� T cells from strong responderHICs, we first tried to compare the HIV-suppressive capacity ofFACSAria-sorted pentamer-positive cell fractions. Unfortunately,and despite a fairly good viability, functionality of these cells wascompromised. Hence, we compared the relative weight of Gag andNef responses by assessing the HIV-suppressive capacity of CD8�
T cell fractions depleted of either one response or the other. Theseexperiments were performed with cells from three HICs (A3, A6,and A11) with similar numbers of HIV-specific cells (11,270,12,473, and 12,612 SFC/106 PBMC, respectively) and a contribu-tion of the Gag response to the total HIV-CD8� T cell responseclose to 50% (49.8%, 63.0%, and 55.3%, respectively). CD8� Tcells isolated from HICs were stimulated with 1) a pool of all theoptimal HIV-1 peptides that were recognized in individualELISPOT assays (not shown); (2) a pool of Gag peptides only; (3) a
pool of Nef peptides only. As shown in Fig. 4A, the suppression ofHIV infection observed when autologous unstimulated CD8� Tcells from strong responder HICs were added to CD4� T cellcultures was lost when the CD8� T cells that produced IFN-� uponstimulation with the complete pool of recognized HIV peptideswere removed. In the case of A3, both the cell fractions depletedof Gag-specific or Nef-specific CD8� T cells retained strong HIV-suppressive capacity (Fig. 4B). For A11, depletion of Gag-specificcells caused the nearly complete loss of HIV-suppressive capacity,whereas depletion of Nef-specific cells had no effect (Fig. 4B). ForA6, the depletion of Gag-specific cells also caused a strong loss ofHIV-suppressive capacity (Fig. 4B). Removal of Nef-specific cellsoccasioned a more modest loss of HIV-suppressive capacity. Insummary, although the respective contributions of Gag and Nefresponses were difficult to quantify precisely, Gag-specific CD8�
T cells seemed to strongly contribute to the HIV-suppressive ca-pacity of CD8� T cells in all three strong responder HICs evalu-ated, in agreement with the correlations described above. In con-trast, the contribution of Nef responses was more variable.
CD8-mediated HIV-suppressive capacity in strong responderHICs is broad
We have already reported a broad capacity of CD8� T cells fromstrong responder HICs to effectively control superinfection by dif-ferent HIV-1 subtypes (21). Interestingly, CD8� T cells fromstrong responder HICs also partially suppressed infection of CD4�
T cells by other human lentiviruses such as HIV-2, SIVmac, andSIVagm (Fig. 4C). At least some of the HIV-1 epitopes recognizedby HIV-specific CD8� T cells from strong responders HICs wereconserved in the other lentiviruses used in our experiments (notshown). In accordance with a MHC-mediated mechanism, the ca-pacity to suppress SIV infection was totally lost when CD8� Tcells were separated from autologous CD4� T cells by semiper-meable membranes (not shown), as was shown in the case ofHIV-1 (21).
FIGURE 4. A, p24 production in culture supernatants (mean � SD, n � 3) at the peak of viral replication after superinfection of CD4� T cells fromA6 with HIV-1 Bal. CD4� T cells were cultured alone or in the presence (CD4/CD8 of 1:1) of non-prestimulated CD8� T cells or CD8� T cells depletedof HIV-specific CD8� T cells. These results are representative of experiments with 5 HICs. B, HIV-suppressive capacity of CD8� T cells (mean � SD,n � 3), as determined by the log fold decrease in the level of secreted p24 (CD4 vs CD4/CD8, 1:1 cell cultures), after depletion of HIV-specific cells (blackbars), Gag-specific (open bars), or Nef specific (gray bars) cell fractions. Pie charts at the bottom represent the relative contribution of Gag and Nefresponses to the total HIV-specific CD8� T cell response. C, CD4� T cells from HIV controllers were infected with replicative HIV-1.BaL (black bars),SIVagm.Gril (open bars), SIVmac.251 (gray bars), or HIV-2.SBL (patterned bars) and cultured alone or with autologous unstimulated CD8� T cells. Viralreplication was monitored by p24 or p27 ELISA on culture supernatants. Bars indicate the level of suppression at the peak of viral replication when CD8�
T cells were present in the culture (mean � SD, n � 3). n.d., Experiment not done.
7833The Journal of Immunology
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
Weak responder HICs carry infectious replicative viruses
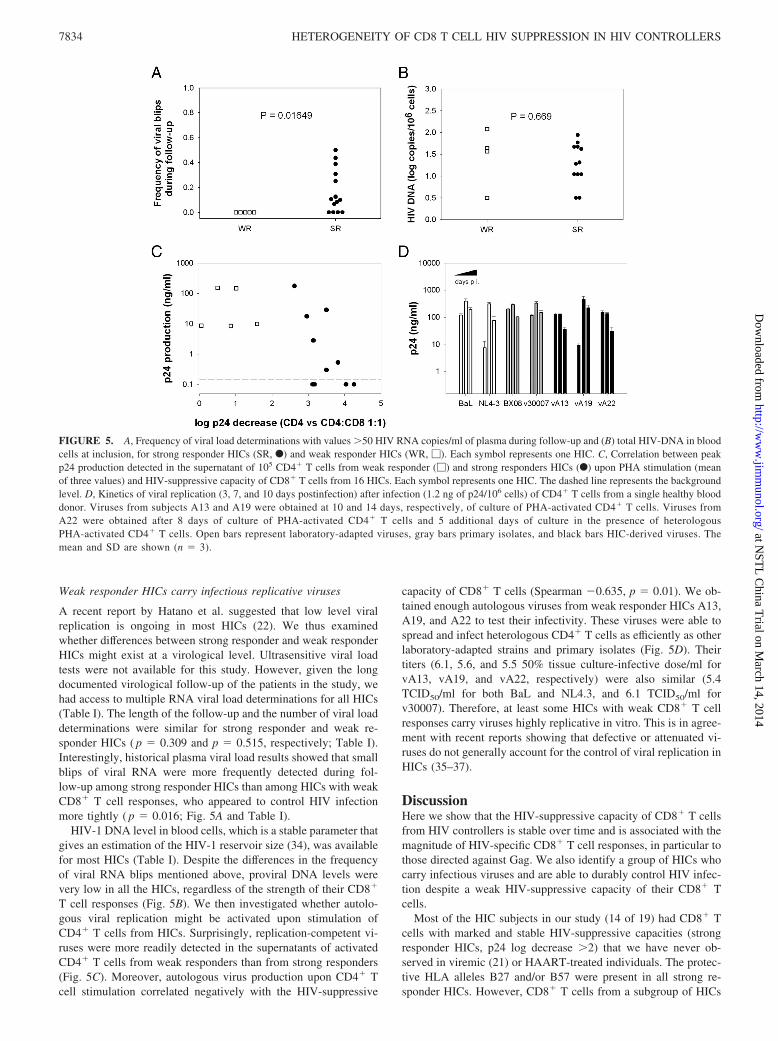
A recent report by Hatano et al. suggested that low level viralreplication is ongoing in most HICs (22). We thus examinedwhether differences between strong responder and weak responderHICs might exist at a virological level. Ultrasensitive viral loadtests were not available for this study. However, given the longdocumented virological follow-up of the patients in the study, wehad access to multiple RNA viral load determinations for all HICs(Table I). The length of the follow-up and the number of viral loaddeterminations were similar for strong responder and weak re-sponder HICs ( p � 0.309 and p � 0.515, respectively; Table I).Interestingly, historical plasma viral load results showed that smallblips of viral RNA were more frequently detected during fol-low-up among strong responder HICs than among HICs with weakCD8� T cell responses, who appeared to control HIV infectionmore tightly ( p � 0.016; Fig. 5A and Table I).
HIV-1 DNA level in blood cells, which is a stable parameter thatgives an estimation of the HIV-1 reservoir size (34), was availablefor most HICs (Table I). Despite the differences in the frequencyof viral RNA blips mentioned above, proviral DNA levels werevery low in all the HICs, regardless of the strength of their CD8�
T cell responses (Fig. 5B). We then investigated whether autolo-gous viral replication might be activated upon stimulation ofCD4� T cells from HICs. Surprisingly, replication-competent vi-ruses were more readily detected in the supernatants of activatedCD4� T cells from weak responders than from strong responders(Fig. 5C). Moreover, autologous virus production upon CD4� Tcell stimulation correlated negatively with the HIV-suppressive
capacity of CD8� T cells (Spearman �0.635, p � 0.01). We ob-tained enough autologous viruses from weak responder HICs A13,A19, and A22 to test their infectivity. These viruses were able tospread and infect heterologous CD4� T cells as efficiently as otherlaboratory-adapted strains and primary isolates (Fig. 5D). Theirtiters (6.1, 5.6, and 5.5 50% tissue culture-infective dose/ml forvA13, vA19, and vA22, respectively) were also similar (5.4TCID50/ml for both BaL and NL4.3, and 6.1 TCID50/ml forv30007). Therefore, at least some HICs with weak CD8� T cellresponses carry viruses highly replicative in vitro. This is in agree-ment with recent reports showing that defective or attenuated vi-ruses do not generally account for the control of viral replication inHICs (35–37).
DiscussionHere we show that the HIV-suppressive capacity of CD8� T cellsfrom HIV controllers is stable over time and is associated with themagnitude of HIV-specific CD8� T cell responses, in particular tothose directed against Gag. We also identify a group of HICs whocarry infectious viruses and are able to durably control HIV infec-tion despite a weak HIV-suppressive capacity of their CD8� Tcells.
Most of the HIC subjects in our study (14 of 19) had CD8� Tcells with marked and stable HIV-suppressive capacities (strongresponder HICs, p24 log decrease �2) that we have never ob-served in viremic (21) or HAART-treated individuals. The protec-tive HLA alleles B27 and/or B57 were present in all strong re-sponder HICs. However, CD8� T cells from a subgroup of HICs
FIGURE 5. A, Frequency of viral load determinations with values �50 HIV RNA copies/ml of plasma during follow-up and (B) total HIV-DNA in bloodcells at inclusion, for strong responder HICs (SR, F) and weak responder HICs (WR, �). Each symbol represents one HIC. C, Correlation between peakp24 production detected in the supernatant of 105 CD4� T cells from weak responder (�) and strong responders HICs (F) upon PHA stimulation (meanof three values) and HIV-suppressive capacity of CD8� T cells from 16 HICs. Each symbol represents one HIC. The dashed line represents the backgroundlevel. D, Kinetics of viral replication (3, 7, and 10 days postinfection) after infection (1.2 ng of p24/106 cells) of CD4� T cells from a single healthy blooddonor. Viruses from subjects A13 and A19 were obtained at 10 and 14 days, respectively, of culture of PHA-activated CD4� T cells. Viruses fromA22 were obtained after 8 days of culture of PHA-activated CD4� T cells and 5 additional days of culture in the presence of heterologousPHA-activated CD4� T cells. Open bars represent laboratory-adapted viruses, gray bars primary isolates, and black bars HIC-derived viruses. Themean and SD are shown (n � 3).
7834 HETEROGENEITY OF CD8 T CELL HIV SUPPRESSION IN HIV CONTROLLERS
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

had only weak HIV-suppressive capacity. In agreement with recentreports (20, 24), the HICs we studied had heterogeneous levels ofHIV-specific CD8� T cells, as estimated by the frequency of IFN-�-producing CD8� T cells. The magnitude of the HIV-specificCD8� T cell response correlated strongly with the capacity ofCD8� T cells from HICs to control HIV infection of autologousCD4� T cells in vitro. Accordingly, the lowest frequencies of IFN-�-producing CD8� T cells were found in weak responder HICs.
Some underestimation of the CD8� T cell response in HICs maycome for the use of peptides derived from consensus sequences forELISPOT determinations, or of a laboratory-adapted HIV strainfor HIV-suppression analyses. However, CD8� T cells from twoweak responder HICs had limited suppressive capacity even whenautologous viruses were used, which further supported a trulyweak CD8� T cell response in these individuals. We cannot ex-clude that control of viremia in weak responders may be due torobust HIV-specific CD8� T cell responses in lymphoid tissues,and actually Ferre and collegues have recently shown that HICshave polyfunctional HIV-specific T cell responses in rectal mucosathat were frequently stronger than in blood (38). However, thepresence in this study of a few HICs with very weak responsesboth in the blood and in the rectal mucosa is interesting. Althougha weak high quality CD8� T cell response might be sufficient tocontrol viremia in vivo, it seems unlikely to be the case in weakresponder HICs. The absence of viral blips during the follow-up ofweak responders and our finding that at least some of these HICscarry viruses that are highly infectious in vitro and readily detect-able upon in vitro activation endorse the idea of a very tight andactive host-restraint of HIV-1 infection. Our phenotypical analysesof the HIV-specific CD8� T cells in weak responders showed anincreased proportion of a CD27�CD45RA� subset of cells, pre-viously observed in patients treated during primary HIV infection,and that might represent a quiescent and stable memory pool ableto proliferate and acquire effector capacities upon Ag stimulation(33). Although these cells may provide an effective response in theeventuality of viral replication, their increased proportion in weakresponder HICs together with the low expression of HLA-DR sug-gest a long period without antigenic stimulation of the CD8� T cellresponse.
Therefore, an alternative mechanism is probably responsible forcontrolling HIV-1 in these HICs. The lower antiviral activity ofCD8� T cells in weak responder HICs did not seem to be com-pensated for by other cell populations within PBMC (e.g., NK cellsor �� T cells), as illustrated by HIV-suppressive experimentswhere nonstimulated PBMC (depleted of CD4� cells), used in-stead of CD8� T cells, were also unable to control HIV superin-fection of autologous CD4� T cells (not shown). Interestingly,persistent lack of low-level detectable viremia in one HIC has beenrecently associated to low levels of HIV Abs and remarkably lowlevels of T cell activation (22). Further virologic studies (such asviral sequencing or determination of tissular viral replication) andthe analysis of innate responses and regulatory T cells (39) mighthelp to identify new mechanisms of control in HICs.
Unlike the cells from weak responder HICs, CD8� T cells fromstrong responder HICs had a broad capacity to suppress superin-fection of their own CD4� T cells by a wide range of HIV-1 strains(21) and, at least partially, by other lentiviruses. This could berelated to the presence of HIV-specific CD8� T cells targetingepitopes located within highly conserved regions of the virus. Re-sponses against Gag and Nef epitopes together accounted for thebulk of total CD8� T cell responses in strong responder HICs, andno phenotypic differences were observed between Gag-specificand Nef-specific CD8� T cell responses in these individuals (21).Interestingly, we observed a strong correlation between the HIV-
suppressive capacity of CD8� T cells in strong responder HICsand the number of Gag-specific CD8� T cell responses. Moreover,the analysis of the relative HIV-suppressive capacity of the Gagresponse in three strong responder HICs showed that, for all threeHICs, Gag-specific CD8� T cells possess the strongest antiviralcapacities. Thus, Gag responses appear to be strongly involved inthe antiviral potency of CD8� T cells. This is in agreement with areport showing evidence of CD8� T cell selective pressure on gagin HICs (40). Increasing evidence suggests that Gag-specificCD8� and CD4� T cell responses are associated with better con-trol of HIV viremia (7–9, 20, 41). Gag epitopes are presented onthe surface of infected CD4� T cells early after viral entry, beforeDNA integration and viral protein synthesis (42), and this mightallow Gag-specific CD8� T cells to recognize and eliminate in-fected cells before the infection is properly established and beforeNef-mediated down-regulation of MHC class I molecules occurs(43). Other factors such as functional avidity (41, 44) or lytic gran-ule loading (45) might contribute to an enhanced HIV-suppressivecapacity of Gag-specific CD8� T cells.
No correlation was found between HIV-suppressive capacity ofCD8� T cells in strong responder HICs with Nef-specific CD8� Tcell responses. However, this observation does not preclude a con-tribution of responses targeting Nef (or other viral proteins) to theHIV-suppressive capacity of CD8� T cells. Actually, our experi-ments of selective depletion of HIV-specific cell fractions showedvariable capacities (from strong to none) of Nef-specific CD8� Tcells from HICs to suppress HIV infection, perhaps depending onthe frequency of the Nef responses that were targeting epitopesrestricted by HLA-B57. Along these lines, escaping mutations arealso found in Nef epitopes in HICs, although less frequently thanin Gag epitopes (46).
Escaping mutations in epitopes located within structurally im-portant regions of the virus could limit the capacity of the virus tomutate to escape immune pressure, as variations in these regionshave a fitness cost (13, 47). Although we did not directly addressthis issue, the difficulties to detect HIV-1 replication in the culturesupernatants of activated CD4� T cells from strong responderHICs might reflect the impact of the pressure exerted by CD8� Tcell responses on viral fitness. Nevertheless, we cannot excludethat, given the extraordinary antiviral potency of CD8� T cellsfrom strong responder HICs, the few remaining CD8� T cells inthe �97% pure CD4� T cell fractions used in these experimentswere enough to efficiently suppress autologous virus replication.
Several important questions await answers; that is, mainlywhether the potent CD8� T cell response observed in most HICsprecedes or follows initial viremic control, and how such a potentCD8� T cell response is maintained. The association presentedhere between blips in plasma viral RNA and stronger CD8� T cellresponses in HICs must be considered with care because of thelimited number of weak responder HICs, but it is tempting to spec-ulate that CD8� T cell control of HIV might involve a feedbackmechanism whereby occasional blips (or ongoing low-level viralreplication) are needed to boost the antiviral response. The in-creased telomerase activity in these cells would further ensure theirpersistence (28). Two scenarios can be envisaged: 1) if viremia iscontrolled by a common mechanism in weak and strong responderHICs, the presence of the protective HLA B27 and B57 alleles mayhelp to sustain control over time, in the eventuality of viral escape,through the establishment of a robust CD8� T cell response; 2)different mechanisms are responsible for initial control of HIVinfection in weak and strong responder HICs. Detailed longitudinalstudies of HICs will be necessary to answer these questions.
7835The Journal of Immunology
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

AcknowledgmentsWe thank all the members of the French National Agency for Research onAIDS and Viral Hepatitis (ANRS) EP36 HIV Controllers study group forhelpful discussions. We thank Chiraz Hamimi for technical help. We alsothank Dr. Laurence Meyer, Dr. Daniel Sereni, Dr. Caroline Lascoux, Dr.Olivier Taulera, Jeannine Delgado, Dr. Francois Bricaire, Dr. Michele Ben-tata, Dr. Pascale Kousignian, Michele Pauchard, Dr. Alain Krivitzky, Pa-tricia Honore, Marie-Therese Rannou, Dr. Jean-Paul Viard, Dr. David Zuc-man, Nadege Velazquez, and all the other physicians and nurses who caredfor the patients. We especially thank the subjects who participated in thisstudy for their cooperation.
DisclosuresThe authors have no financial conflicts of interest.
References1. Borrow, P., H. Lewicki, B. H. Hahn, G. M. Shaw, and M. B. Oldstone. 1994.
Virus-specific CD8� cytotoxic T-lymphocyte activity associated with control ofviremia in primary human immunodeficiency virus type 1 infection. J. Virol. 68:6103–6110.
2. Friedrich, T. C., L. E. Valentine, L. J. Yant, E. G. Rakasz, S. M. Piaskowski,J. R. Furlott, K. L. Weisgrau, B. Burwitz, G. E. May, E. J. Leon, et al. 2007.Subdominant CD8� T-cell responses are involved in durable control of AIDSvirus replication. J. Virol. 81: 3465–3476.
3. Jin, X., D. E. Bauer, S. E. Tuttleton, S. Lewin, A. Gettie, J. Blanchard,C. E. Irwin, J. T. Safrit, J. Mittler, L. Weinberger, et al. 1999. Dramatic rise inplasma viremia after CD8� T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 189: 991–998.
4. Flores-Villanueva, P. O., E. J. Yunis, J. C. Delgado, E. Vittinghoff,S. Buchbinder, J. Y. Leung, A. M. Uglialoro, O. P. Clavijo, E. S. Rosenberg,S. A. Kalams, et al. 2001. Control of HIV-1 viremia and protection from AIDSare associated with HLA-Bw4 homozygosity. Proc. Natl. Acad. Sci. USA 98:5140–5145.
5. Kiepiela, P., A. J. Leslie, I. Honeyborne, D. Ramduth, C. Thobakgale, S. Chetty,P. Rathnavalu, C. Moore, K. J. Pfafferott, L. Hilton, et al. 2004. Dominant in-fluence of HLA-B in mediating the potential co-evolution of HIV and HLA.Nature 432: 769–775.
6. Migueles, S. A., M. S. Sabbaghian, W. L. Shupert, M. P. Bettinotti,F. M. Marincola, L. Martino, C. W. Hallahan, S. M. Selig, D. Schwartz,J. Sullivan, and M. Connors. 2000. HLA B*5701 is highly associated with re-striction of virus replication in a subgroup of HIV-infected long term nonpro-gressors. Proc. Natl. Acad. Sci. USA 97: 2709–2714.
7. Edwards, B. H., A. Bansal, S. Sabbaj, J. Bakari, M. J. Mulligan, andP. A. Goepfert. 2002. Magnitude of functional CD8� T-cell responses to the Gagprotein of human immunodeficiency virus type 1 correlates inversely with viralload in plasma. J. Virol. 76: 2298–2305.
8. Kiepiela, P., K. Ngumbela, C. Thobakgale, D. Ramduth, I. Honeyborne,E. Moodley, S. Reddy, C. de Pierres, Z. Mncube, N. Mkhwanazi, et al. 2007.CD8� T-cell responses to different HIV proteins have discordant associationswith viral load. Nat. Med. 13: 46–53.
9. Riviere, Y., M. B. McChesney, F. Porrot, F. Tanneau-Salvadori, P. Sansonetti,O. Lopez, G. Pialoux, V. Feuillie, M. Mollereau, S. Chamaret, et al. 1995. Gag-specific cytotoxic responses to HIV type 1 are associated with a decreased risk ofprogression to AIDS-related complex or AIDS. AIDS Res. Hum. Retroviruses 11:903–907.
10. Chopera, D. R., Z. Woodman, K. Mlisana, M. Mlotshwa, D. P. Martin,C. Seoighe, F. Treurnicht, D. A. de Rosa, W. Hide, S. A. Karim, et al. 2008.Transmission of HIV-1 CTL escape variants provides HLA-mismatched recipi-ents with a survival advantage. PLoS Pathog. 4: e1000033.
11. Martinez-Picado, J., J. G. Prado, E. E. Fry, K. Pfafferott, A. Leslie, S. Chetty,C. Thobakgale, I. Honeyborne, H. Crawford, P. Matthews, et al. 2006. Fitnesscost of escape mutations in p24 Gag in association with control of human im-munodeficiency virus type 1. J. Virol. 80: 3617–3623.
12. Rousseau, C. M., M. G. Daniels, J. M. Carlson, C. Kadie, H. Crawford,A. Prendergast, P. Matthews, R. Payne, M. Rolland, D. N. Raugi, et al. 2008.HLA class I-driven evolution of human immunodeficiency virus type 1 subtypec proteome: immune escape and viral load. J. Virol. 82: 6434–6446.
13. Schneidewind, A., M. A. Brockman, R. Yang, R. I. Adam, B. Li, S. Le Gall,C. R. Rinaldo, S. L. Craggs, R. L. Allgaier, K. A. Power, et al. 2007. Escape fromthe dominant HLA-B27 restricted CTL response in Gag is associated with adramatic reduction in HIV-1 replication. J. Virol. 81: 12382–12393.
14. Goepfert, P. A., W. Lumm, P. Farmer, P. Matthews, A. Prendergast,J. M. Carlson, C. A. Derdeyn, J. Tang, R. A. Kaslow, A. Bansal, et al. 2008.Transmission of HIV-1 Gag immune escape mutations is associated with reducedviral load in linked recipients. J. Exp. Med. 205: 1009–1017.
15. Deeks, S. G., and B. D. Walker. 2007. Human immunodeficiency virus control-lers: mechanisms of durable virus control in the absence of antiretroviral therapy.Immunity 27: 406–416.
16. Saez-Cirion, A., G. Pancino, M. Sinet, A. Venet, and O. Lambotte. 2007. HIVcontrollers: how do they tame the virus? Trends Immunol. 28: 532–540.
17. Lambotte, O., F. Boufassa, Y. Madec, A. Nguyen, C. Goujard, L. Meyer,C. Rouzioux, A. Venet, and J. F. Delfraissy. 2005. HIV controllers: a homoge-
neous group of HIV-1-infected patients with spontaneous control of viral repli-cation. Clin. Infect. Dis. 41: 1053–1056.
18. Dinoso, J. B., S. Y. Kim, R. F. Siliciano, and J. N. Blankson. 2008. A comparisonof viral loads between HIV-1-infected elite suppressors and individuals who re-ceive suppressive highly active antiretroviral therapy. Clin. Infect. Dis. 47:102–104.
19. Bailey, J. R., K. G. Lassen, H. C. Yang, T. C. Quinn, S. C. Ray, J. N. Blankson,and R. F. Siliciano. 2006. Neutralizing antibodies do not mediate suppression ofhuman immunodeficiency virus type 1 in elite suppressors or selection of plasmavirus variants in patients on highly active antiretroviral therapy. J. Virol. 80:4758–4770.
20. Emu, B., E. Sinclair, H. Hatano, A. Ferre, B. Shacklett, J. N. Martin,J. M. McCune, and S. G. Deeks. 2008. HLA class I-restricted T-cell responsesmay contribute to the control of human immunodeficiency virus infection, butsuch responses are not always necessary for long-term virus control. J. Virol. 82:5398–5407.
21. Saez-Cirion, A., C. Lacabaratz, O. Lambotte, P. Versmisse, A. Urrutia,F. Boufassa, F. Barre-Sinoussi, J. F. Delfraissy, M. Sinet, G. Pancino, andA. Venet. 2007. HIV controllers exhibit potent CD8 T cell capacity to suppressHIV infection ex vivo and peculiar CTL activation phenotype. Proc. Natl. Acad.Sci. USA 104: 6776–6781.
22. Hatano, H., E. L. Delwart, P. J. Norris, T. H. Lee, J. Dunn-Williams, P. W. Hunt,R. Hoh, S. L. Stramer, J. M. Linnen, J. M. McCune, et al. 2009. Evidence forpersistent low-level viremia in individuals who control HIV in the absence ofantiretroviral therapy. J. Virol. 83: 329–335.
23. Gea-Banacloche, J. C., S. A. Migueles, L. Martino, W. L. Shupert, A. C. McNeil,M. S. Sabbaghian, L. Ehler, C. Prussin, R. Stevens, L. Lambert, et al. 2000.Maintenance of large numbers of virus-specific CD8� T cells in HIV-infectedprogressors and long-term nonprogressors. J. Immunol. 165: 1082–1092.
24. Pereyra, F., M. M. Addo, D. E. Kaufmann, Y. Liu, T. Miura, A. Rathod, B. Baker,A. Trocha, R. Rosenberg, E. Mackey, et al. 2008. Genetic and immunologicheterogeneity among persons who control HIV infection in the absence of ther-apy. J. Infect. Dis. 197: 563–571.
25. Betts, M. R., M. C. Nason, S. M. West, S. C. De Rosa, S. A. Migueles,J. Abraham, M. M. Lederman, J. M. Benito, P. A. Goepfert, M. Connors, et al.2006. HIV nonprogressors preferentially maintain highly functional HIV-specificCD8� T cells. Blood 107: 4781–4789.
26. Migueles, S. A., A. C. Laborico, W. L. Shupert, M. S. Sabbaghian, R. Rabin,C. W. Hallahan, D. Van Baarle, S. Kostense, F. Miedema, M. McLaughlin, et al.2002. HIV-specific CD8� T cell proliferation is coupled to perforin expressionand is maintained in nonprogressors. Nat. Immunol. 3: 1061–1068.
27. Zimmerli, S. C., A. Harari, C. Cellerai, F. Vallelian, P. A. Bart, and G. Pantaleo.2005. HIV-1-specific IFN-�/IL-2-secreting CD8 T cells support CD4-indepen-dent proliferation of HIV-1-specific CD8 T cells. Proc. Natl. Acad. Sci. USA 102:7239–7244.
28. Lichterfeld, M., D. Mou, T. D. Cung, K. L. Williams, M. T. Waring, J. Huang,F. Pereyra, A. Trocha, G. J. Freeman, E. S. Rosenberg, et al. 2008. Telomeraseactivity of HIV-1-specific CD8� T cells: constitutive upregulation in controllersand selective increase by blockade of PD ligand 1 in progressors. Blood 112:3679–3687.
29. Schvachsa, N., G. Turk, M. Burgard, D. Dilernia, M. Carobene, M. Pippo,M. Gomez-Carrillo, C. Rouzioux, and H. Salomon. 2007. Examination of real-time PCR for HIV-1 RNA and DNA quantitation in patients infected with HIV-1BF intersubtype recombinant variants. J. Virol. Methods 140: 222–227.
30. Avettand-Fenoel, V., N. Mahlaoui, M. L. Chaix, C. Milliancourt, M. Burgard,M. Cavazzana-Calvo, C. Rouzioux, and S. Blanche. 2007. Failure of bone mar-row transplantation to eradicate HIV reservoir despite efficient HAART. AIDS21: 776–777.
31. O’Doherty, U., W. J. Swiggard, and M. H. Malim. 2000. Human immunodefi-ciency virus type 1 spinoculation enhances infection through virus binding. J. Vi-rol. 74: 10074–10080.
32. Lacabaratz-Porret, C., A. Urrutia, J. M. Doisne, C. Goujard, C. Deveau,M. Dalod, L. Meyer, C. Rouzioux, J. F. Delfraissy, A. Venet, and M. Sinet. 2003.Impact of antiretroviral therapy and changes in virus load on human immunode-ficiency virus (HIV)-specific T cell responses in primary HIV infection. J. Infect.Dis. 187: 748–757.
33. Lecuroux, C., I. Girault, A. Urrutia, J. M. Doisne, C. Deveau, C. Goujard,L. Meyer, M. Sinet, and A. Venet. 2009. Identification of a particular HIV-specific CD8� T cell subset with a CD27� CD45RO�/RA� phenotype and mem-ory characteristics after initiation of HAART during acute primary HIV infection.Blood 113: 3209–3217.
34. Avettand-Fenoel, V., T. Prazuck, L. Hocqueloux, A. Melard, C. Michau,R. Kerdraon, E. Agoute, and C. Rouzioux. 2008. HIV-DNA in rectal cells is wellcorrelated with HIV-DNA in blood in different groups of patients, includinglong-term non-progressors. AIDS 22: 1880–1882.
35. Blankson, J. N., J. R. Bailey, S. Thayil, H.-C. Yang, K. Lassen, J. Lai,S. K. Gandhi, J. D. Siliciano, T. M. Williams, and R. F. Siliciano. 2007. Isolationand characterization of replication-competent HIV-1 from a subset of elite sup-pressors. J. Virol. 81: 2508–2518.
36. Lamine, A., A. Caumont-Sarcos, M. L. Chaix, A. Saez-Cirion, C. Rouzioux,J. F. Delfraissy, G. Pancino, and O. Lambotte. 2007. Replication-competent HIVstrains infect HIV controllers despite undetectable viremia (ANRS EP36 study).AIDS 21: 1043–1045.
37. Miura, T., M. A. Brockman, C. J. Brumme, Z. L. Brumme, J. M. Carlson,F. Pereyra, A. Trocha, M. M. Addo, B. L. Block, A. C. Rothchild, et al. 2008.
7836 HETEROGENEITY OF CD8 T CELL HIV SUPPRESSION IN HIV CONTROLLERS
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

Genetic characterization of human immunodeficiency virus type 1 in elite con-trollers: lack of gross genetic defects or common amino acid changes. J. Virol.82: 8422–8430.
38. Ferre, A. L., P. W. Hunt, J. W. Critchfield, D. H. Young, M. M. Morris,J. C. Garcia, R. B. Pollard, H. F. Yee, Jr., J. N. Martin, S. G. Deeks, andB. L. Shacklett. 2008. Mucosal immune responses to HIV-1 in elite controllers:a potential correlate of immune control. Blood Epub ahead of print on Dec. 23,2008: doi: 10.1182/blood-2008-10-182709.
39. Chase, A. J., H. C. Yang, H. Zhang, J. N. Blankson, and R. F. Siliciano. 2008.Preservation of FoxP3� regulatory T cells in the peripheral blood of humanimmunodeficiency virus type 1-infected elite suppressors correlates with lowCD4� T-cell activation. J. Virol. 82: 8307–8315.
40. Bailey, J. R., T. M. Williams, R. F. Siliciano, and J. N. Blankson. 2006. Main-tenance of viral suppression in HIV-1-infected HLA-B*57� elite suppressorsdespite CTL escape mutations. J. Exp. Med. 203: 1357–1369.
41. Chen, H., A. Piechocka-Trocha, T. Miura, M. A. Brockman, B. D. Julg,B. M. Baker, A. C. Rothchild, B. L. Block, A. Schneidewind, T. Koibuchi, et al.2009. Differential neutralization of human immunodeficiency virus (HIV) repli-cation in autologous CD4 T cells by HIV-specific cytotoxic T lymphocytes. J. Vi-rol. 83: 3138–3149.
42. Sacha, J. B., C. Chung, E. G. Rakasz, S. P. Spencer, A. K. Jonas, A. T. Bean,W. Lee, B. J. Burwitz, J. J. Stephany, J. T. Loffredo, et al. 2007. Gag-specific
CD8� T lymphocytes recognize infected cells before AIDS-virus integration andviral protein expression. J. Immunol. 178: 2746–2754.
43. Collins, K. L., B. K. Chen, S. A. Kalams, B. D. Walker, and D. Baltimore. 1998.HIV-1 Nef protein protects infected primary cells against killing by cytotoxic Tlymphocytes. Nature 391: 397–401.
44. Almeida, J. R., D. A. Price, L. Papagno, Z. A. Arkoub, D. Sauce, E. Bornstein,T. E. Asher, A. Samri, A. Schnuriger, I. Theodorou, et al. 2007. Superior controlof HIV-1 replication by CD8� T cells is reflected by their avidity, polyfunction-ality, and clonal turnover. J. Exp. Med. 204: 2473–2485.
45. Migueles, S. A., C. M. Osborne, C. Royce, A. A. Compton, R. P. Joshi,K. A. Weeks, J. E. Rood, A. M. Berkley, J. B. Sacha, N. A. Cogliano-Shutta, et al.2008. Lytic granule loading of CD8� T cells is required for HIV-infected cellelimination associated with immune control. Immunity 29: 1009–1021.
46. Bailey, J. R., T. P. Brennan, K. A. O’Connell, R. F. Siliciano, and J. N. Blankson.2009. Evidence of CD8� T cell mediated selective pressure on HIV-1 nef inHLA-B*57� elite suppressors. J. Virol. 83: 88–97.
47. Streeck, H., M. Lichterfeld, G. Alter, A. Meier, N. Teigen, B. Yassine-Diab,H. K. Sidhu, S. Little, A. Kelleher, J. P. Routy, et al. 2007. Recognition of adefined region within p24 Gag by CD8� T cells during primary human immu-nodeficiency virus type 1 infection in individuals expressing protective HLAclass I alleles. J. Virol. 81: 7725–7731.
7837The Journal of Immunology
at NST
L C
hina Trial on M
arch 14, 2014http://w
ww
.jimm
unol.org/D
ownloaded from




















