
OPERKULUM: BAGIAN KUNCI UNTUKIDENTIFlKASI...
14
sebaliknya semakin banyak perberbedaan karakter yang dimiliki maka hubungan kekerabatannya semakin jauh. Kelas Gastropoda memiliki bagian- bagian tubuh yang dijadikan sebagai karakter untuk kunci identifikasi. Beberapa bagian tubuh bahkan merupakan karakter khusus yang banya dimiliki oleh Kelas Gastropoda, salah satunya adalah operkulum. Posisi operkulum pada Gastropoda ditunjukkan pada Gambar I. Operkulum, yang berarti tutup kecil (jamak: opercula) adalah struktur berkapur pada banyak jenis (tidak semua) Gastropoda. Hampir semua siput Pulmonata bersifat inoperculate, yaitu I) UPT Loka Konservasi Biota Laut, Pusat Penelitian Oseanografi-LIP1, Bitung ldentifikasi makhluk hidup secara konvensional awaJnya dilakukan dengan cara membandingkan aspek morfologi dan anatomi, yaitu membandingkan bagian-bagian tubub dari makhluk hidup yang diidentiflkasi. Persamaan dan perbedaan yang muneul antar jenis makhluk hidup akhimya rnenjadi dasar pengklasifikasian, membagi makhluk hidup ke dalam beberapa kelompok, filum, kelas, ordo, famili dan seterusnya sampai ke tingkat jenis. Semakin banyak karakter yang sama maka memiliki hubungan kekerabatan yang semakin dekat, dan PENDAHULUAN OPERCULUM: KEY CHARACl'ER FOR GASTROPODS IDENTIFICATION THAT IS OFfEN OVERLOOKED. Gastropodshavebodyparts thatcanbeusedaskeycharacters for identification. A special character which isfound only in gastropods, one of which is the operculum. Operculum shows a similarity to the shell, and there is a possibility of an evolutionary link between these secretory organs. The growth of operculum is following the growth of the shell, so the size of the operculum remains proportional to the size of the aperture. The type and form of operculum are used to assist the identification and classification to species level. The most important function of the operculum is to keep water in the body as well as a protection device against predators when the body is inside the shelf. Operculum of the gastropods often utilized by humans , ,. for making several kinds of materials. Evolutionary development of operculum morphology can be evidencedfrom the analysis of fossils. ABSTRACT Ucu Yanu Arbi') Oleb OPERKULUM: BAGIAN KUNCI UNTUK IDENTIFlKASI GASTROPODA YANG SERING TERABAIKAN ISSN 0216-1877 Oseana, Volume XXXVln, Nomor I, Tahun 2013: 1- 14
Transcript of OPERKULUM: BAGIAN KUNCI UNTUKIDENTIFlKASI...

sebaliknya semakin banyak perberbedaankarakter yang dimiliki maka hubungankekerabatannya semakin jauh.
Kelas Gastropoda memiliki bagianbagian tubuh yang dijadikan sebagai karakteruntuk kunci identifikasi. Beberapa bagian tubuhbahkan merupakan karakter khusus yang banyadimiliki oleh Kelas Gastropoda, salah satunyaadalah operkulum. Posisi operkulum padaGastropoda ditunjukkan pada Gambar I.Operkulum, yang berarti tutup kecil (jamak:opercula) adalah struktur berkapur pada banyakjenis (tidak semua) Gastropoda. Hampir semuasiput Pulmonata bersifat inoperculate, yaitu
I)UPT Loka Konservasi Biota Laut, Pusat Penelitian Oseanografi-LIP1, Bitung
ldentifikasi makhluk hidup secarakonvensional awaJnya dilakukan dengan caramembandingkan aspek morfologi dan anatomi,yaitu membandingkan bagian-bagian tubub darimakhluk hidup yang diidentiflkasi. Persamaandan perbedaan yang muneul antar jenis makhlukhidup akhimya rnenjadi dasar pengklasifikasian,membagi makhluk hidup ke dalam beberapakelompok, filum, kelas, ordo, famili danseterusnya sampai ke tingkat jenis. Semakinbanyak karakter yang sama maka memilikihubungan kekerabatan yang semakin dekat, dan
PENDAHULUAN
OPERCULUM: KEY CHARACl'ER FOR GASTROPODS IDENTIFICATION THAT ISOFfEN OVERLOOKED. Gastropodshave bodyparts that can be used as key charactersfor identification. A special character which is found only in gastropods, one of whichis the operculum. Operculum shows a similarity to the shell, and there is a possibilityof an evolutionary link between these secretory organs. The growth of operculum isfollowing the growth of the shell, so the size of the operculum remains proportional tothe size of the aperture. The type and form of operculum are used to assist theidentification and classification to species level. The most important function of theoperculum is to keep water in the body as well as a protection device against predatorswhen the body is inside the shelf. Operculumof the gastropods often utilized by humans, ,.for making several kinds of materials. Evolutionary development of operculummorphology can be evidenced from the analysis of fossils.
ABSTRACT
Ucu Yanu Arbi')
Oleb
OPERKULUM: BAGIAN KUNCI UNTUK IDENTIFlKASI GASTROPODAYANGSERING TERABAIKAN
ISSN 0216-1877Oseana, Volume XXXVln, Nomor I, Tahun 2013: 1- 14

2
Operkulum menunjukkan kemiripandengan eangkang, kemungkinan terdapathubungan evolusi antara kedua organsekretorik tersebut, Hashimoto et al. (2012)menemukan bahwa beberapa gen yang terlibatdalam pembentukan eangkang juga terdapat
STRUKTUR ANATOMI DAN MORFOLOGlOPERKUUJM
Gambar I. Terminologi Gastropoda seeara umum yang menunjukkan bahwa operkuJum merupakanbagian penting dari Gastropoda (Sumber: Koleksi pribadi).
dalam operkulum, seperti decapentaplegic(OPP) dan grainyhead. Bukti lain menunjukkanbahwa OPP memiliki peran kunei dalampengembangan operkulum. Berdasarkanpengamatan, bahwa OPP penting bagipengembangan cangkang dan operkulum, danOPP di bagian posterior kakimemiliki kontribusiterbadap inovasi dari operkulum. Bentuk dasaroperkulum ditunjukkan pada Gambar 2.
Tulisan ini merupakan sebuah tinjauanyang dirangkum dari berbagai sumber. Penulismeneoba memberikan informasi mengenaipentingnya peranan operkulum baik bagikehidupan Gastropoda dan untuk kepentinganidentifikasi jenis Gastropoda, dengan harapandapat menjadi tambahan pengetahuan tentangperanan operkulum.
tidak memiliki operkulum, keeuali pada FamiliAmphiboloidea. Namun, beberapa spesiesPulmonata terestrial mampu mensekresi suatuepiphragm, yaitu struktur sementara yang daJambeberapa kasus dapat memiliki fungsi yang sarnaseperti operkulum. Epiphragm dapat dibedakandari operkulum dalamhal homogenitas dan tandapertumbuhan (Cbeca & Jimenez-Jimenez, 1998.

3
Lapisan luar adalah struktur homolog. Keluarganeritopsine dikategorikan ke dalam kelompokcangkang empat lapis (NeritiIiidae,Neritidae, danPbenacoJepadidae) dan keJompok cangkang tigalapis (keJuarga lain). Sebaliknya, strukturoperkulum merupakan variabel nyata diNeritopsina, dan sedikit korelasi yang dapatdibentuk dalammenerangkan evolusi filogenetikatau adaptasi (Sasaki, 200 1). Salah satu strukturoperkulum yang telah diteliti adalam operkulumdari Neritopsis radula yang ditunjukkan padaGambar3.
Struktur cangkang dan operkulum dariNerita {Theliostylaj albicilla dan Cinnalepetapulchella digambarkan dan dibandingkandengan anggota yang masih ada lainnyaNeritopsina. Cangkang N. (1'.) albiclilla terdiridari empat lapisan: lapisan terluar prismatik,diikuti oleb lapisan silang-lamelar sederbana,myostracum, dan lapisan daJam silang-lameLarkompleks. Operkulum terdiri dari tiga lapisanprismatik diendapkan pada kedua sisi lapisaoorganik. C. pulchella juga memiliki cangkangempat lapis, tetapi tidak memiliki operkulum.
Gambar2. OperkuJumMarstonia comalensis sebagai contob operkulum yang memiliki bentuk anatomidasar seperti cangkang (Kaim & Sztajner, 2005).

4
OperkuIurnmelekat pada akhir dari ototcolumellar dengan punggung lempeng opercularrnengarah ke permukaan atas dari bag ianbelakang kaki. Pertumbuhao operkulummengikuti pertumbuhan ukuran cangkang,sehingga ukuran operkulum tetap sebandingdeogan ukuran aperture (bukaan cangkang).Pada banyak spesies, ketika hewan tersebut aktifdan merangkak, bagian dari bawah cangkangterletak pada permukaan luar operkulum
Gambar 3. Mikrostruktur operkulum Neritopsis radula dari Mauritius, Sarnudera Hindia. A) BagiandaJarnoperkulum yang mengalami kalsifikasi; B) Bagian dalarn yang mengalami kalsifikasidiselubungi bagian aragonitik; C) Aragonitik, bagian luar prismatik-berserat; D) Perbatasanantara aragonitik, bagian luar prisma-berserat dan bagian pertama yang mengalamikalsifikasi, bagian dalarnprismatik-komposit; E) Bagian pertama yangmengalami kalsifikasi,bagian dalam prismatik-kornposit; F) Bagian pertama yang mengalarni kalsifikasi, bagiandalarn prismatik-kornposit dan bagian dalam prismatik-berserat kedua yang dibatasi olehlapisan organik; 0) Bagian dalam prismatik-berserat yang mengalarni kalsifikasi kedua; H)Permukaan lapisan organik dengan ujung prisma (Kaim & Sztajner, 2005).
tersebut. Dalam banyak spesies dari siput tautbercangkang yang hidup pada zona subtidal;ukuran operkulum umumnya tereduksi menjadikecil, dan tidak lagi berfungsi untuk menutupaperture. Bahkan dalam sejumlah famiListrukturoperkulum telah teJah benar-benar tereduksisepenuhnya.
Menurut Kaim & Sztajner (2005) duatipe dasar operkulum daJamhal komposisi bahanpenyusunnya:

5
Gambar 4. Variasi bentuk dasar operkulum pada Gastropoda (Kaim & Sztajner, 2(05).
genus Purpura, Xenophora, danPaludomus.
• Berbentuk cakar atau unguiculate: intiapikal atau di depan, seperti pada genusTurbinella dan Fusus; dan yang berbentukcakar dan bergerigi, seperti pada Strombus.
• Spiral: twnbuh hanya pada satu sisi danberputar seperti tumbuh, selalu sinistralpada kulit dextral.
• Paucispiral atau O)igog}'TOUS:sedikit spiral,pada genus Littorina.
• Subspiral atau bampir spiral: pada genusThiara.
• Multispiral atau polygyrous: memilikistruktur spiral berjarak dekat dengan jumlahbanyak, kadang-kadang sampai dna puluh,dimana jumlab putaran pada operkulumtidak dipengaruhi oleh jumlah whorleangkang, tetapi oleh kelengkunganaperture, seperti pada genus Trochus.
• Terartikulasi: ketika memiliki proyeksi,seperti pada genus Nerita.
• Terpancar: merupakan modifikasi darioperkulum terartikulasi di mana bentukspiral tidak begitu jelas, seperti pada genusNavicella.
VARIASI BEN1lJK OPERKULUMJenis dan bentulc operkulum
digunakan untuk membantu mengidentifikasidan mengkJasifikasikan grup terkait (genus),babkan sampai Icetingkat spesies. Variasi bentukstruktur operkulum dapat digambarkan sebagaiberikut:• Konsentris: intinya sebagai pusat atau
subcentral, seperti pada genusLithoglyphus dan Ampul/aria, dan intilainnya dekat margin parietal dati cangkang.
• Terimbrikasi atau pipih: tumbuh banya padasatu sisi dengan inti marjinal, seperti pada
1. Jenisyang paling umum dati ope:rkulum tipisterdiri dari bahan protein corneous agaktebal, yang berwarna kuning sampaikecoklatan dan biasanya agak tembus,lentur, tetapi mungkin menjadi rapuh ketikakering. Bentuknya bervariasi, tergantungpada kelompok siput dan bentuk bukaancangkangnya.
2 Jenis lain dari operkulum terbatas padabeberapa famHi, misal Turbinidae, yangmemiliki basis corneous berkapur yangpejal. Permukaan berkapur dalam beberapaf3mili meniliki wama atau omamenmisalnyaberupa pustula dan alur cangkang.
6
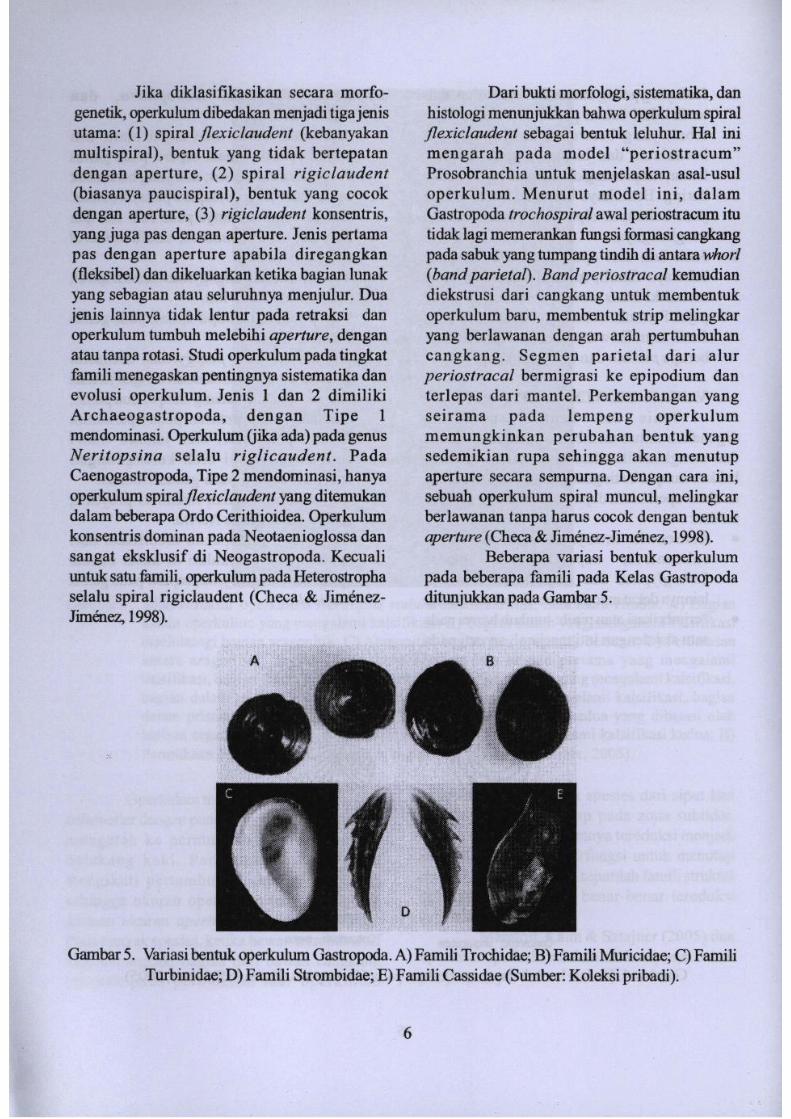
Gambar 5. Variasi bentuk operkulum Gastropoda. A) Famili Trochidae; B) Famili Muricidae; C) FamiliTurbinidae; D) Famili Strombidae; E) Famili Cassidae (Sumber: Koleksi pribadi).
Dari bukti morfologi, sistematika, danbistologi menunjukkan babwa operkulum spiralflexiclaudent sebagai bentuk leluhur, Hal inimengarah pada model "periostracum"Prosobranchia untuk menjelaskan asal-usuloperkulum. Menurut model ini, dalamGastropoda trochospiralawal periostracum itutidak lagi memerankan fimgsi forrnasi cangkangpada sabuk yang tumpang tindih di antara whorl(bandparietal). Band periostracal kemudiandiekstrusi dari cangkang untuk membentukoperkulum baru, membentuk strip melingkaryang berlawanan dengan arab pertumbuhancangkang. Segmen parietal dari alurperiostracal bermigrasi ke epipodium danterlepas dad mantel. Perkembangan yangseirama pada lempeng operkulummemungkinkan perubahan bentuk yangsedemikian rupa sehingga akan menutupaperture secara sempurna. Dengan cara ini,sebuah operkulum spiral muncul, melingkarberlawanan tanpa harus cocok dengan bentukaperture(Checa & Jimenez-Jimenez, 1998).
Beberapa variasi bentuk operkulumpada beberapa farnili pad a Kelas Gastropodaditunjukkan pada Gambar 5.
Jika diklasifikasikan secara morfogenetik, operkulum dibedakan menjadi tiga jenisutama: (1) spiral jlexic/audent (kebanyakanmultispiral), bentuk yang tidak bertepatandengan aperture, (2) spiral rigiclaudent(biasanya paucispiral), bentuk yang cocokdengan aperture, (3) rigiclaudent konsentris,yang juga pas dengan aperture. Jenis pertamapas dengan aperture apabiJa diregangkan(tleksibel) dan dikeluarkan ketika bagian lunakyang sebagian atau seluruhnya menjuiur. Duajenis lainnya tidak lentur pada retraksi danoperkulum tumbuh melebihi aperture, denganatau tanpa rotasi. Studi operkulum pada tingkatfamili menegaskan pentingnya sistematika danevolusi operkulum. Jenis 1 dan 2 dimilikiArchaeogastropoda, dengan Tipe 1mendominasi. Operkulum (jika ada) pada genusNerttopsina selalu riglicaudent . Pad aCaenogastropoda, Tipe 2 mendominasi, banyaoperkulum spiraljlexiclaudenl yang ditemukandalam beberapa Ordo Cerithioidea. Operkulumkonsentris dominan pada Neotaenioglossa dansangat eksklusif di Neogastropoda. Kecualiuntuk satu famili, operkulum pada Heterostrophaselalu spiral rigiclaudent (Checa & JimenezJimenez, 1998).
7
tereduksi, dan yang tidak mampu menutupaperture. Operkulum kadang termodifikasi,misalnya pada Famili Strombidae yangberbentuk seperti eakar dan digunakan untukmendorong ke substrat pada saat bergerakmelompat. Pada Famili Conidae, operkulummemanjang dan berbentuk seperti cakar, dandigunakan untuk menggali pasir sehinggamemungkinkan siput untuk melakukan gerakanmelompat. Mekanisme pemanfaatan operkulumuntuk melompat digambarkan pada Gambar 7.
FUNGSI OPERKUWM BAGIGASTROPODA
Operkulum melekat pada permukaanalas kaki dan dalam keadaan yang palinglengkap, berfungsi sebagai semaeam "jebakan"untuk menutup lubang eangkang ketika bagianlunak hewan tersebut ditarik ke dalam eangkang.Sebuah lubang udara keeil mungkin tetaptersedia untuk membantu pemapasan. BentukoperkuJum sangat bervariasi mengikuti bentukaperture.Banyak familimemiliki operkulum yang
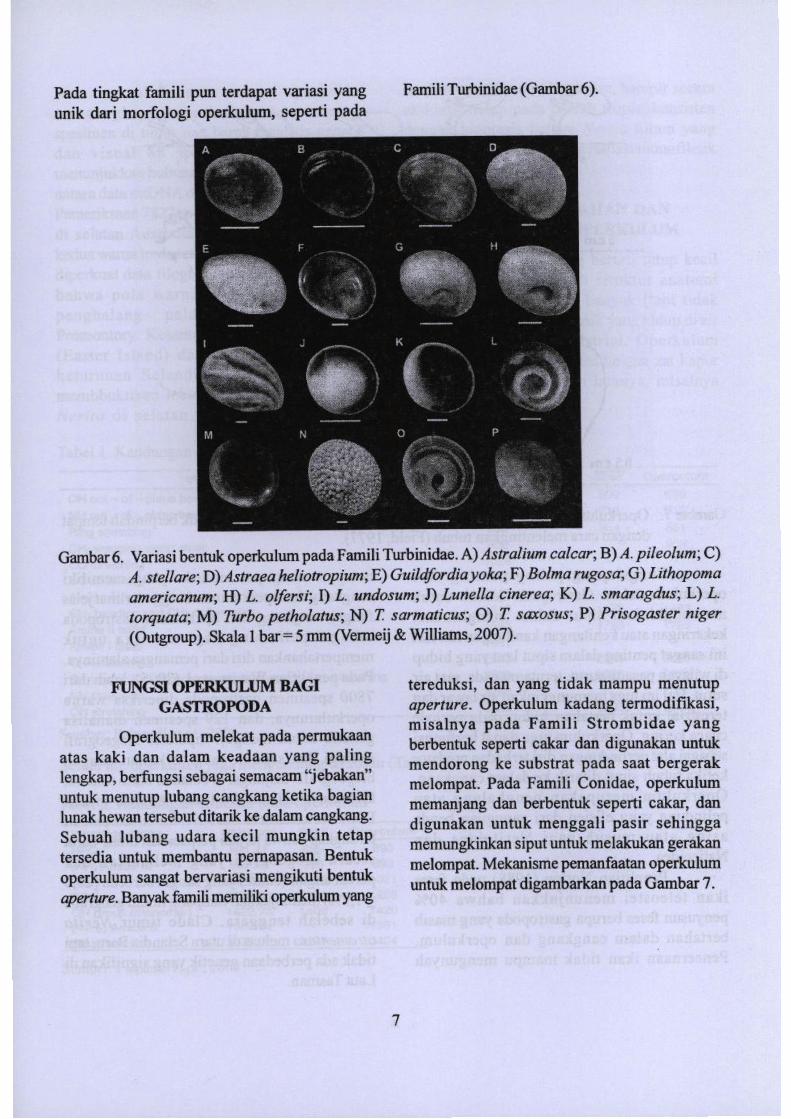
Gambar6. Variasi bentuk operkulwn pada Famili Turbinidae. A)Astralium calcar; B) A. pileolum; C)A. stellare; D)Astraea heliotropium; E) Guildfordia yoka; F)Bolma rugosa; G) Lithopomaamericanum; H) L. olfersi; J) L. undosum; J) Lunella cinerea; K) L. smaragdus; L) L.torquata; M) Turbo petholatus; N) T. sarmaticus; 0) T. saxosus; P)Prisogaster niger(Outgroup). Skala 1bar = 5 mm (Venneij&Williams, 2007).
FamiliTurbinidae (Gambar 6).Pada tingkat famili pun terdapat variasi yangunik dari morfologi operkulum, seperti pada

8
maupun mengupas moluska yang memilikicangkang dan operkulum sehingga terlihat jelasfungsi cangkang dan operkulum gastropodasebagai pelindung bagi moluska untukmempertahankan diri dari pemangsa aJaminya.Pada penelitian Waters et aJ.(2005), lebih dari7800 spesimen gastropoda diperiksa wamaoperkulumnya, dan 129 spesimen dianalisagenetik untuk menguji hipotesis biogeografiekologi dan sejarah yang berkaitan denganbiogeografis disjungsi dibelahan bumi selatan,khususoya selatan Australia. Analisa urutanDNA mitokondria dari Nerita atramentosaterungkap bahwa terjadi perpecahan timur-baratsecara filogeografi yang melibatkan duapercabangan (clade)yang sangat berbeda (26,0± 1,94'10) menun jukkan twnpang tindih geografisdi sebelah tenggara. Clade timnr Neritaatramentosa meluas diutara Selandia Barn, tapitidak ada perbedaan genetik yang signifikan diLaut Tasman.
Fungsi yang paling penting darioperkulum daJam Gastropoda adaJah untukmemungkinkan siput dalarn menghindarikekeringan atau kehilangan kandungan air. Halini sangat penting dalam siput laut yang hidupdi wiJayah pasang surut, terutama pada saat airsurut. Hal inijuga memunglrinkan air tawar danterestrial untuk bertahan hidup pada periodecuaca kering. Operkulumjuga dapat berfungsisebagai alat perlindungan diri terhadap predatorketika tubuh siput ditarik ke dalam cangkang.Operkulum merupakan penghalang ataupelindung yang efisien dari gangguan bendaasing atau musuh bagi Neritoidea danNaticoidea.
Penelitian Norton (1988) pada fesesikan teleostei menunjukkan bahwa 40%penyusun feses berupa gastropoda yang masihbertahan dalam cangkang dan operkuJum.Pencernaan ikan tidak mampu mengunyah
Gambar7. Operkulum pada Famili Strombidae bc:rfungsi sebagai alat bantu untuk berpindah tempatdengan caramelentingkan tubuh (Field, 1977).
\ 0.2 em",\.
c
7••Scm
,3

9
Vibration mode Chito .. n Shell 0E!!rculumHPO/ 891.41 856 893(NH) Amide III 897.41 896 893po," 1026.63 1032 1021PO,' 1259.54 1264 1255OH group (monomer) 1422.73 1408 1420(·NH2) Amide II 1587.94 1581Structural unit 3377.95 33n 3404
Sumber: Palpandi et al., 2009
Tabel 2. Kandungan chitosan pada operkulum
Sumber: Palpandi et al., 2009
Selaio itu, operkulum juga meogandung chitosan (Tabel 2).
Standard a - chitin (cm··) Shell Operculum690 699 699752 713 712896 8S4 861952 90810261073 1083 108211161418 1478 14831563 1788 178816812878 2853 28532930 2923 292132683439 3395 3401
OH out - of - plane benOlIlQNH out - of - plane betdingRing stretchingC~ wagging along chainCO stretchingCO stretchingAsymmetric In - phase ring stretching modeCH2bending and CH3defomtalionAmide II bandAmide I bandCH stretchingSymmetric CH3stretching and Blymmetrlc CH2 stretchingNH stretchingOH stretching
Vlbration modeS
Operkulurn, yang berarti tutup kecil(jamak: opercula) adalah struktur anatomiberkapur yang ada pada banyak (tapi tidaksemua) jenis Gastropoda, baik yang hidup di airlaut, tawar maupun terestrial. Operkulumtemyata selain memiliki kandungan zat kapurjuga memiliki kandungan lainnya, misalnyachitin (Tabel l).
KANDUNGAN BAHAN DANPEMANFAATAN OPERKULUM
sembilan spesies dari Neritidae, hampir secaraeksklusif hidup pada daerah tropis, konsistendengan hipotesis bahwa Nerita hitam yanghidup di selatan merupakan radiasi monofiletik(Waters et al., 2005).
Tabel 1. Kandungan chitin pada operkuJum
Analisis pola warna operkulummengungkapkan perbedaan besar antaraspesimen eli timur dan barat. AnaJisis genetikdan visual 88 spesimen dari Australiamenunjukkan hubungan yang benar-benar acakantara data mtDNA dan pola wama operkulum.Pemeriksaan 7822 spesimen lebih dari 14 lokasidi selatan Australia mengungkapkan bahwakedua warna terdapat pada semua lokasi, namundiperkuat data filogeografi yang menunjukkanbahwa pola warna berhubungan denganpenghalang palaeogeografi: WilsonsPromontory. Kesamaan genetik Nerita morio(Easter Island) dan Australia timur danketurunan Selandia Baru (l , 1 + 0,3%)membbuktikan lebih lanjut dari penyebaranNerita di selatan. Filogenetik hubungan

IO
Penempelan tubub pada fase larvaPhestilla (Nudibrancha) ke cangkang danoperkulum terjadi di lokasi penyisipan otot. Saatmetamorfosis, baik cangkang maupunoperkulum mereduksi dan akhirnya hilang.Iovestigasi mikroskopis elektron terhadapcangkang dan lampiran operkulum selamametamorfosis telab menunjukkan bahwahemidesmosom apikal kebilangan integritasdan bundel tonofilament menarik diri dariplasmalemma apikal sel-sel epiteLsebagai selyang tidak berhubungan dengan cangkang atauoperkulum (Bonar, 1978).
Ontogeni awal gastropoda (yaitu,planktotroflk versus nonplanktotrofik) mungkinmerujuk dari morfologi protoconch padaGastropoda dewasa. Protoconch terdiri dariembrio dan larva dalam spesies denganpengembangan planktotrofik, bentuk embriocangkang pada peri ode intracapsuJar dancangkang larva berikutnya secara bertahapberkembang selama periode larva. Spesies
SEJARAH EVOLUSI DANPERKEMBANGAN OPERKULUM
adaLab spesies yang operkulumnya palingbanyak digunakan, meskipun operkulum darispesies Turbo lain kadang-kadang jugadigunakan.OperkuIum inihampir selalumemilikistruktur solid, karena memiliki satu sisi datardan tidak menarik corneousyang melekat padabagian lunak hewan, dan sisi lainnya mengkilapkasar hemispherical, yang dalam kasus T.petholatus memiliki area hijau gelap yangmeoarik.
Operkulum sering digunakan dalamseni asli Northwest Coast, misalnya untukasesoris pada masker, panel dan dayung. Obyekinidigunakan untukmewakili gigi dalam masker,dan digunakan sebagai elemen dekoratif dalambanyak benda yang merupakan bagian pentingdalam seni dan budaya masyarakat NorthwestCoast, termasuk piring untuk makanan danlainnya.
·Chitin dan cbitosan selain terkandung dalamoperkulum juga terkandung daLamcangkang danbagian tubuh yang memiliki struktur kerangkakapur lainnya, seperti halnya yang terkandungdalam rangka luar atau kulit udang, kepiting danberbagai jenis Crustacea lainnya (palpandi etal.,2009).
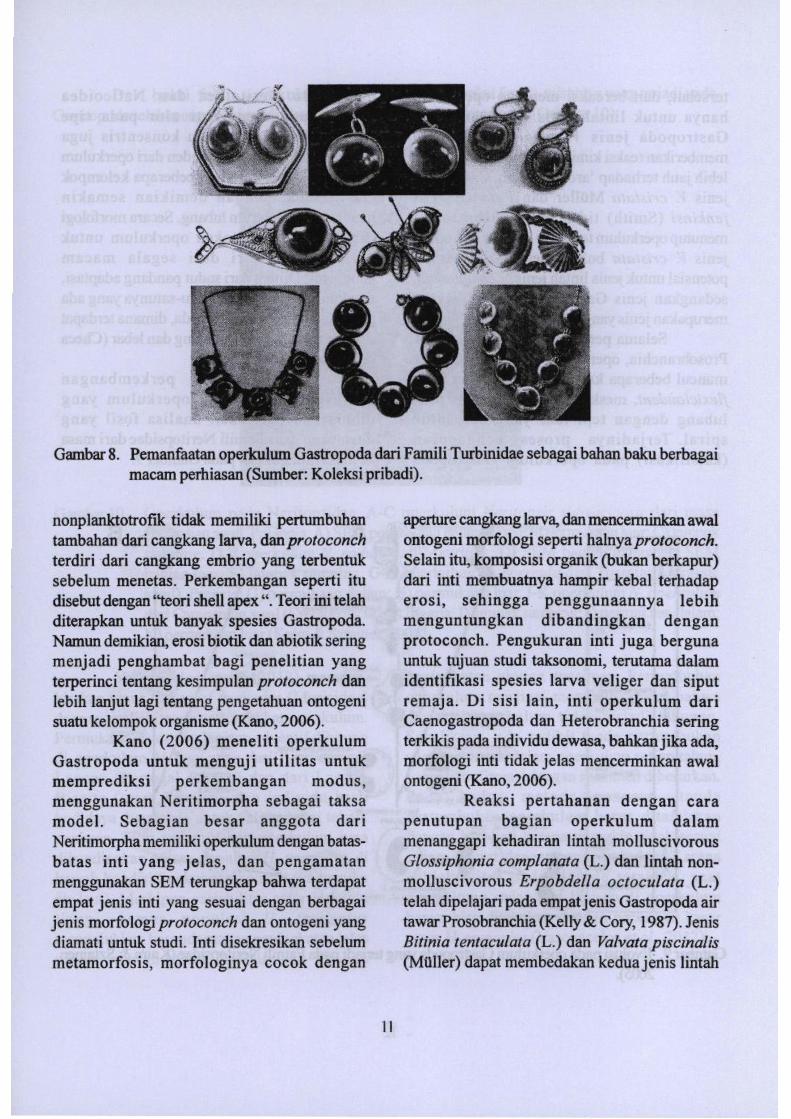
Operkulum selaio memiliki fungsipenting bagi siput ternyata juga rnemilikibeberapa macam bahan yang terkandung didalamoya yang sering dimanfaatkao olehmanusia. Operkulum dari Gastropoda terteotu,terutama varietas dari Laut Merah, telah lamadimanfaatkan sebagai bahan dupa dalam tradisiYahudi kuno serta budayaArab. Operkulum dariStrombus tricornis dan Lambis Iruncataseringdigunakan di daerah dekat Timur Tengah untukkeperluan serupa. Bubuk operkulum jugamerupakan bahan penting dalam pembuatandupa di China dan Jepang yang disebut "BElxiang" (Chinese: .lll .fi: , IiIseashell! aroma) atau"kai kou" (Jepang: It' =if , lit shell! aroma bajubesi.) Produsen di negara-negara tersebutmenggunakan operkulum dari berbagai jewssiput, termasuk yang mudah ditemukan di AsiaTenggara, Arnerika Selatan, dan Afrika Timur.Bau amis operkulum dibilangkan secaratradisional dengan cuka, alkobol dan air.Operkulum yang telah bersih kemudian digilingmenjadi bubuk dan digunakan sebagai fiksatifaroma, daLam teknik yang sama dengan yangdigunakan daLam parfum dari resin tanamantertentu. Ketika dibakar, operkulum berkualitastinggi berbau kastor, sedangkan yangberkualitas rendah aromanya seperti rambutyang dibakar.
Operkulwn spesies tertentu (misalnyadari Famili Turbinidae) kadang-kadangdigunakan sebagai "batu pennata" organik yangsangat murah pada cincin, gelang, jimat danlainnya. Operkulum ini umumnya dikenaJ dilingkungan oelayan dan pedagang sebagai"mata kucing" (atau lebib bam-bam ini disebutsebagai "mata Siwa"). Siput Turbopetholatus
II
aperture cangkang larva, danmencenni.nkan awalootogeni morfologi seperti halnya protoconch.Selain itu, komposisi organik (bukan berkapur)dari inti membuatnya hampir kebal terhadaperosi, sehingga peogguoaannya lebihmenguntungkan dibandiogkan denganprotoconch. Pengukuran inti juga bergunauntuk tujuan studi taksonomi, terutama dalamidentifikasi spesies larva veliger dan siputremaja. Di sisi lain, inti operkulum dariCaenogastropoda dan Heterobranchia seringterkikis pada individu dewasa, bahkanjika ada,morfologi inti tidak jelas mencerminkan awalontogeni (Kano, 2006).
Reaksi pertahanan dengan carapenutupan bagian operkulum dalammeoaoggapi kehadiran lintah moUuscivorousGlossiphonia complanata (L.) dan tintah nonmoUuscivorous Erpobdella octoculata (L.)telah dipelajari pada empatjenis Gastropoda airtawar Prosobranchia (Kelly& Cory, 1987). JenisBitinia tentaculata (L.) dan Valvatapiscinalis(MUlier) dapat membedakan kedua jenis lintah
oonplanktotrofik tidak memiliki pertumbuhantambahan dari cangkang larva, dan protoconchterdiri dari cangkang embrio yang terbentuksebelum menetas. Perkembangan seperti itudisebut deogan "teori shell apex". Teori ini telahditerapkan untuk banyak spesies Gastropoda.Namun demikian, erosi biotik dan abiotik seringmenjadi penghambat bagi penelitian yangterperinci tentang kesimpulan protoconch danlebih lanjut lagi tentang peogetahuan ontogenisuatu kelompok organisme (Kano, 2006).
Kano (2006) meneliti operkulumGastropoda untuk meoguji utilitas uotukmemprediksi perkembangan modus,menggunakan Neritimorpha sebagai taksamodel. Sebagian besar anggota dariNeritimorpha memiliki operkulum dengan batasbatas inti yang jelas, dan pengamatanmenggunakan SEM terungkap bahwa terdapatempat jenis inti yang sesuai dengan berbagaijenis morfologi protoconch dan ontogeni yangdiamati untuk studio Inti disekresikan sebelummetamorfosis, morfologinya cocok dengan
Gambar 8. Pemanfaatan operkulum Gastropoda dari Famili Turbinidae sebagai bahan baku berbagaimacam perhiasan (Sumber: Koleksi pribadi).
12
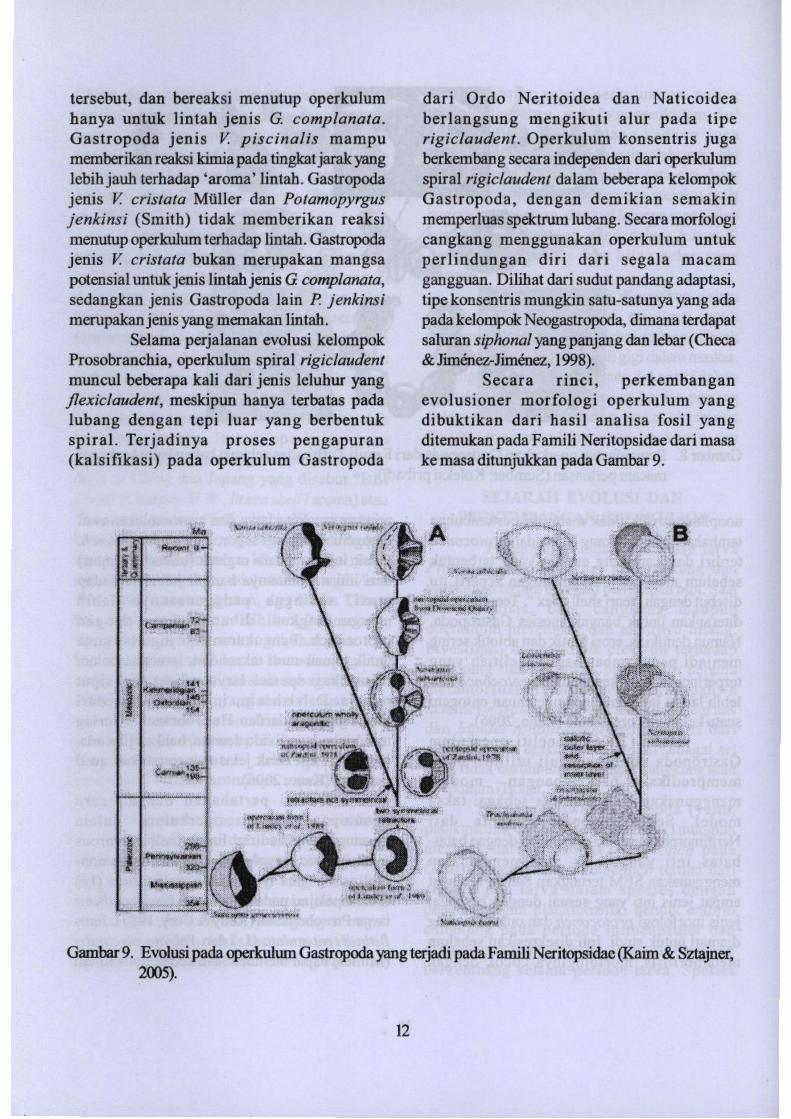
Gambar 9. Evolusipada operkulurnGastropoda yang te:rjadi pada Famili Neritopsjdae (Kaim & Sztajner,2005).
dari Ordo Neritoidea dan Natieoideaber langsung mengikuti alur pada tiperigiclaudent . Operkulum konsentris jugaberkembang secara independen dari operkulumspiral rigiclaudent dalam beberapa kelompokGastropoda, dengan demikian semakinmemperluas spektrum lubang. Secara morfologicangkang menggunakan operkulum untukperlinduogan diri dari segala macamgangguan. Dilihat dari sudut pandang adaptasi,tipe konsentris mungkin satu-sarunya yang adapada kelompok Neogastropoda, dimana terdapatsaluran siphonal yang panjang dan lebar (Checa&Jimenez-Jimenez, 1998).
Secara rinci, perkembanganevolusioner morfologi operkulum yangdibuktikan dari hasil analisa fosil yangditemukan pada Famili Neritopsidae dari masake masa ditunjukkan pada Gambar 9.
tersebut, dan bereaksi menutup operkulumhanya untuk lintah jenis G. comp/anata.Gastropoda jenis V. pisoinalis mampumemberikan reaksi kimia pada tingkat jarak yanglebih jauh terhadap 'aroma' lintah. Gastropodajenis V. cristata Muller dan Potamopyrgusjenkinsi (Smith) tidak memberikan reaksimenutup operkulum terhadap lintah. Gastropodajenis V. cristata bukan merupakan mangsapotensial untuk jenis lintah jenis G complanata,sedangkan jenis Gastropoda lain P. jenkins;merupakan jenis yang memakan lintah.
Selama perjalanan evolusi kelompokProsobranchia, operkulum spiral rigiclaudentmuncul beberapa kali dari jenis lelubur yangflexiclaudent, meskipun hanya terbatas padalubang dengan tepi luar yang berbentukspiral. Terjadinya proses pengapuran(kalsifikasi) pada operkulum Gastropoda

13
peningkatan ukuran spesimen. Pertumbuhanoperkulum marj inal meogungkapkan bahwadeposisi operkuJum tidak terjadi tahunan.Sebaliknya, ada sedikit tanda pertumbuhanyang terbentuk dalam operkuJum setiap taboo,yang dikonfumasi dengan spesirnen dibesarkan.Pada kedua metode penuaan, tandapertumbuhan operkulum harus ditafsirkandengan hati-hati kareoa mungkin tidak sesuaisecara akurat dengan umur individu, Karenasubjektivitas pada metode tersebut, saatmenghitung striae dan Japisan di operkulumhanya bisa dilakukan dengan perkiraan kasarumur H. trunculus (Vasconcelos et al., 2012).
Vasconcelos et al. (2012) memperkirakan umur Hexaplex trunculus (Muricidae)dengan analisis tanda pertumbuhan operkulum.Pennukaan striae dan lapisan adventif dihitungdi operkulum H. trunculus dari Ria FormosaLagoon (Portugal selatan) dan dari LagoonBizerte (utara Tunisia), dan individu basilbudidaya digunakan untuk validasi dari teknikpenuaan. Fitur pertumbuhan operkuJum jugadibandingkan antara populasi H. trunculus dibawah kondisi lingkungan kontras (yaitu suhuair laut). Meskipun variabiJitas antar-individutioggi, semua populasi H. trunculusmenunjukkan kecenderungan peningkatan jelasdalam jumlah striae dan lapisan dengan
Gambar 10. Operkulum pada Neritopsidae. A-C operkutum Neritopsis subvaricosa dari masaOxfordian awal, Polandia:A) SEM permukaan dalam; B) SEM permukaan luar;C) tampaksamping. D-F operkutum N. radula dari Mauritius: D) SEM bagian dalam; E) SEMbagian luar; F) tampak samping. G-H operkulum N. opercula dari Toarcian (Jurassicawal), Prancis: G) pennukaan dalam; H) permukaan luar. I-K operkulum N. operculumdart Campanian awal (Cretaceous akhir) Jnggris: I) tampak samping; 1)permukaan dalam;K) pennukaan luar (Kairn& Sztajner, 2005).
sampai tingkat famili, misalnya yang terjadi padaFamiliNeritopsidae (Gambar 10).
Dari bukti fosil, operkulum padaGastropoda telah mengalami modifikasi bentuk

14
Kano, Y. 2006. Usefulness of the opercularnucleus for inferring earlydevelopment in neritimorphgastropods. J. Morphol.267(9): 1120-1136.
Kelly, P.M. and J.S. Cory. 1987. Operculumclosing as a defence against predatoryleeches in four British freshwaterprosobranch snails. Hydrobiologia144(2): 121-124.
Norton, S.F. 1988. Role of the gastropod sheUand operculum in inhibiting predationbyfisbes. Science241 (4861): 92-94.
Palpandi,C., V. Shanmugam andA. Shanmugam.2009. Extraction ofchitin and chitosanfrom shell and operculum ofmangrovegastropod Nerita (Dostia)crepidularia Lamarck. InternationalJournal of Medicine and MedicalSciences 1(5): 198-205.
Sasaki, T. 200 1.Macro- and microstructure ofshell and operculum in two recentgastropod species, Nerita(Theliostyla) albici/la andCinnalepeta pulchella (Neritopsina:Neritoidea), PaleontologicalRes.5 (1):21-31.
Vasconcelos, P., I.H. Gharsallah, P.Moura, N.Zamouri-Langar, A. Gaamour, H.Missaoui, O. Jarboui andM.G Gaspar.2012. Appraisal of the usefulness ofoperculum growth marks for ageingHexaplex trunculus (Gastropoda:Muricidae): Comparison betweensurface striae and adventitious layers.Marine Biology Research 8 (2): 141-15l
Waters, J.M., T.M. King,P.M. O'Loughlin andfiG Spencer. 2005. Phylogeographicaldisjunction in abundant high-dispersallittoral gastropods. Mol.Ecol. 14 (9):2789-2802.
Bonar, D.B. 1978. Fine structure of muscleinsertions on the larval shell andoperculum of the nudibranch Pheslillasibogae (Mollusca: Gastropoda)before and during metamorphosis.1issueCell. 10(1): 143-152.
Checa, A.G and A.P. Jimenez-Jimenez. 1998.Cmstructionalmorphology origin, andevolution of gastropod operculum.Paleobiology24 (1): 109-132.
Field, L.H. 1977. An experimental analysis ofthe escape response of the gastropodStrombus maculatus. Pacific Science31(1): 1-11.
Hashimoto, N., Y. Kurita and H.Wada. 2012.Developmental role of DPP in thegastropod shell plate and co-option ofthe DPP signaling pathway in theevolution of the operculum. Dev. Bioi.IS; 366 (2): 367-373.
Kaim,A. and P. Sztajner. 2005. The opercula ofner itopsid gastropods and theirphylogenetic importance. Journal ofMolluscanStudies 71: 211-219.
DAFfAR PUSTAKA
Proses identifikasi Gastropoda seringdilakukan hanya sebatas karakter lrunci padamorfologi cangkang saja. Seharusnya jugamempertimbangkan karakter anatomi, fisiologi,perilaku, habitat dan lainnya, sertamengkmfirma.sik.andeogan karakter genetilcajikadiperlukan. Selain cangkang, bagian tubublainnya kadang justru menjadi kunci yangmenjadi pembeda spesies, misalnya massa tububdan operkulum. Dengan demikian, dalam prosesidentifikasi bagian tubuh lain tidak bisadiabaikan karena juga menjadi identitas danpenciri dari suatu spesies.




















