
Genome Evolution
77
GENOM EVOUTION MAKALAH Diajukan untuk Memenuhi Tugas Matakuliah Evolusi yang Dibina Oleh Prof. Dr. agr. Mohamad Amin, S.Pd., M.Si Oleh Kelompok 8/Kelas D: Husdiani Ningsih (140341807091) Nuril Maghfiroh (140341807614) W.F Edi Hunsen (140341807914)
-
Upload
nurilmaghfiroh -
Category
Documents
-
view
145 -
download
30
description
makalah evolusi genom
Transcript of Genome Evolution

GENOM EVOUTION
MAKALAH
Diajukan untuk Memenuhi Tugas Matakuliah Evolusi
yang Dibina Oleh Prof. Dr. agr. Mohamad Amin, S.Pd., M.Si
Oleh Kelompok 8/Kelas D:
Husdiani Ningsih (140341807091)
Nuril Maghfiroh (140341807614)
W.F Edi Hunsen (140341807914)
UNIVERSITAS NEGERI MALANG
PASCASARJANA
PROGRAM STUDI PENDIDIKAN BIOLOGI
Maret 2015

BAB I
PENDAHULUAN
A. Latar Belakang
Akhir-akhir ini, genom hanya dapat dipelajari secara tidak langsung,
dengan penerapan sebagian dan terkadang tidak mewakili sekuen genom.
Keadaan mulai berubah dengan cepat sehingga keberadaan sekuen genom lengkap
telah ditemukan. Genom organel adalah yang pertama disekuensing; sekuen
mitokondria lengkap yang pertama (~17.000 bp) dikemukakan pada tahun 1981,
dan genom kloroplas yang pertama (~156.000 bp) pada tahun 1986. Sekuen
genom lengkap pertama pada organisme yang hidup bebas ialah eubacterium
Haemophilus influenzae (~1.830.000 bp), diselesaikan pada tahun 1995, diikuti
perubahan yang cepat dengan ditemukannya sekuen lengkap archaeon,
Methanococcus jannaschii (~1.660.000 bp), dan 16 kromosom ragi uniseluler,
Saccharomyces cerevisiae (~12.000.000 bp). Genom organisme multiseluler
lengkap pertama, pada nematoda Caenirhabditis elegans (~97.000.000 bp), yang
dilaporkan pada tahun 1998, dan proyek genom untuk Drosophila melanogaster,
manusia, tikus, padi, dan jagung diharapkan akan dilengkapi di masa pada masa
yang akan datang. Dalam pembahasan kali ini dikhususkan, untuk mempelajari
evolusi genom secara sederhana dengan menggunakan sekuen genom.
Pembahasan yang akan disajikan berisi empat topik yang berbeda. Topik
yang pertama adalah ukuran genom, yang mana sangat bervariasi antara
organisme. Bagaimana variasi ini dipertahankan, dan mekanisme apa yang dapat
meningkatkan atau menurunkan ukuran genom dalam menghasilkan variasi.
Selain itu, juga dibahas terkait informasi genetik yang termasuk di dalam genom.
Dapatkah genom berisi banyak gen DNA, atau genom terbuat dari sebagian besar
sekuen nongenik. Apakah fraksi nongenik memiliki fungsi, atau tidak. Terdapat
banyak pengulangan sekuen pada genom dan jika demikian apa fungsi dan pola
distribusi kromosom. Topik kedua yaitu masalah keteraturan gen dan dinamika
perubahan evolusi dalam keteraturan gen. Bagaimana distribusi gen di sepanjang
dan di antara kromosom. Mekanisme apa yang bertanggungjawab untuk
pembentukan kembali keteraturan gen selama evolusi. Topik ketiga berkaitan

dengan komposisi nukleotida dalam genom. Apakah ada heterogenitas dalam
komposisi di sepanjang daerah yang berbeda dari genom. Mekanisme apa yang
dapat menimbulkan perbedaan di daerah setempat dalam komposisi nukleotida.
Topik yang terakhir, berkaitan dengan evolusi kode genetik.
B. Rumusan Masalah
1. Bagaimanakah penjelasan adanya variasi ukuran genom di antara organisme?
2. Bagaimanakah penjelasan terkait dengan masalah keteraturan gen dan
dinamika perubahan evolusi dalam hubungannya dengan keteraturan gen?
3. Bagaimanakah penjelasan terkait komposisi nukleotida dalam genom?
4. Bagaimanakah penjelasan terkait dengan adanya peristiwa evolusi pada kode
genetik?
C. Tujuan
1. Menjelaskan adanya variasi ukuran genom di antara organisme.
2. Menjelaskan keteraturan gen dan dinamika perubahan evolusi dalam
hubungannya keteraturan gen.
3. Menjelaskan adanya komposisi nukleotida dalam genom.
4. Menjelaskan adanya peristiwa evolusi pada kode genetik

BAB II
PEMBAHASAN
A. Nilai C
Pada organisme haploid seperti bakteri, ukuran genom ditunjukkan oleh
jumlah total DNA di dalam genom. Pada organisme diploid ataupun poliploid,
ukuran genom didefinisikan sebagai jumlah DNA dalam genom haploid yang
tidak direplikasi, seperti pada inti sperma. Ukuran genom juga disebut dengan
nilai C, dimana C diartikan sebagai “konstan” atau “karakteristik” yang
menunjukkan bahwa ukuran genom haploid menunjukkan variabilitas
intraspesifik yang kecil yang cukup konstan dalam setiap spesies. Sebaliknya,
nilai C memiliki variasi yang luas dari spesies satu ke spesies yang lain baik pada
prokariot maupun eukariot.
Ukuran genom inti pada eukariot biasanya dalam satuan picograms (pg)
dari DNA (1pg=10-12 g). Genom terkecil prokariot umumnya dinyatakan dalam
satuan dalton, suatu unit dari atom relatif atau massa molekul. Ukuran dari genom
yang masih tergolong terkecil, serta ukuran spesifik untaian DNA, lebih sering
dinyatakan dalam base pairs (bp) atau kilobase pairs (Kb) dari DNA atau RNA
untai ganda (1 Kb = 1000 bp). Sekuens genom yang lengkap biasanya dinyatakan
dalam megabase pairs (1Mb = 1000 Kb). Faktor konversi ini ditunjukkan dalam
Tabel 2.1 berikut ini.
Tabel 2.1. Faktor Konversi Ukuran Genom Organisme
UnitFaktor Konversi
Picograms Dalton Base PairsPicogram 1 6,02 x 1011 0,98 x 109
Dalton 1,66 x 10-12 1 1,62 x 10-3
Base Pair 1,02 x 10-9 618 1(Sumber: Graur dan Hsiung Li, 1999)
B. Evolusi Ukuran Genom pada Prokariot
Ukuran genom bakteri bervariasi, berkisar antara 20-30 kali dari ukuran
yang terlecil yaitu 6x105 bp pada beberapa intraseluler parasit obligat, sampai
lebih dari 107 bp pada beberapa spesies cyanobakteri (Tabel 2.2). Mollicutes, yang
tidak memiliki dinding sel dan prokariot terkecil yang hidup bebas dan mampu
melakukan reproduksi sendiri, umumnya memiliki ukuran genom yang sangat

kecil. Kelas Mollicutes terdiri dari enam marga, di antaranya Mycoplasma adalah
yang paling terkenal.
Tabel 2.2 Kisaran Nilai C pada Beberapa Prokariot.Taxon Kisaran ukuran genom
(Kb)Rasio (Tinggi/Rendah)
Bacteria 580 – 13,200 23 Mollicutes 580 – 2,200 4 Gram negativea 650 – 9,500 15 Gram positive (Firmicutes)
1,600 – 11,600 7
Cyanobacteria 3,100 – 13,200 4Archaea 1,600 – 4,100 3
Data diambil dari Caviler-Smith (1985), Romling et al. (1992), Carle et al. (1995), dan sumber lain.aKebanyakan dari kelompok paraphyletic(Sumber: Graur dan Hsiung Li, 1999)
Genom terkecil yang kita ketahui adalah pada patogen urogenital
Mycoplasma genitalium, yang mengandung sekitar 470 gen pengkode protein, 3
gen rRNA spesifik, dan 33 gen tRNA spesifik. Gen pembawa informasi yang
terkandung dalam genom M. genitalium dipercaya hanya sedikit yang lebih besar
dari jumlah minimal yang dibutuhkan untuk hidup. Sejumlah gen pada bakteri lain
kurang lebih pada kisaran 500 hingga 8000 (kira-kira berkisar 20 kali). Dengan
kata lain, variasi gen kira-kira hampir sama dengan variasi pada nilai C.
Rata-rata ukuran gen pengkode protein pada bakteri adalah sekitar 1 Kb,
ukuran fraksi gen pada genom diperkirakan berkisar antara 500 Kb hingga sekitar
104 Kb. Kita dapat menyimpulkan bahwa prokariot tidak mengandung DNA
nongenik dalam jumlah yang besar. Memang, mayoritas sekuen pengkode protein
pada spesies bakteri lebih banyak mencapai 87-94% dari genom, sehingga fraksi
nongenik nampak sedikit lebih kecil. Kecuali hingga sampai saat ini ialah pada
genom intraseluler parasit Rickettsia prowazekii, yang mengandung 24% DNA
noncoding. Untuk eubakteria mempunyai sekuens yang lengkap, ini
memungkinkan untuk memperhitungkan korelasi antara ukuran genom dan
jumlah genom (Gambar 2.1). Korelasi yang hampir sempurna menunjukkan
bahwa variasi pada ukuran genom bakteri dapat sepenuhnya dijelaskan oleh
jumlah gen. Korelasi yang sama nampak pada Archaea, tetapi saat ini data sangat
terbatas untuk menggambarkan kesimpulan pastinya.
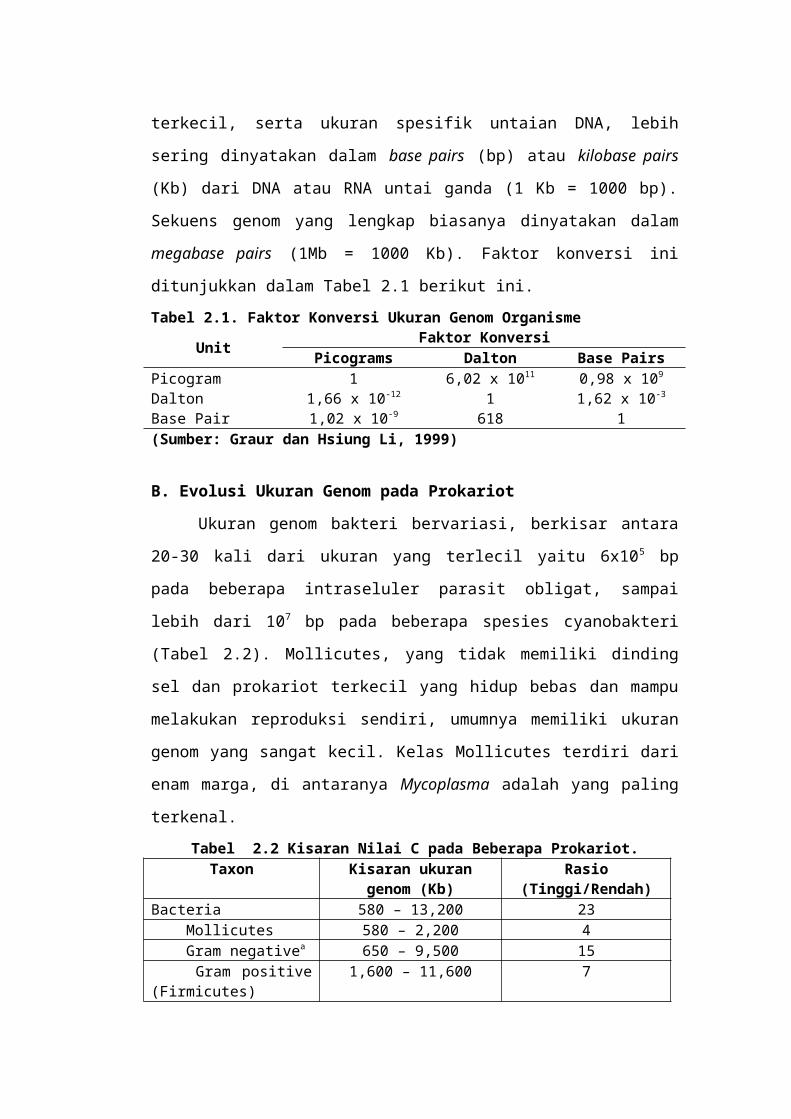
Gambar 2.1 Hubungan antara jumlah gen dan ukuran genom pada sekuen lengkap spesies eubakteria dengan 12 genom sirkuler dan satu genom linier.
(Sumber: Graur dan Hsiung Li, 1999)
Genom bakteri dibagi menjadi 3 fraksi yaitu (1) DNA kromosomal, (2)
DNA yang berasal dari plasmid, dan (3) transposable elements. Fraksi
kromosomal mengandung gen pengkode protein yang dibutuhkan untuk
pertumbuhan dan fungsi metabolisme (90-95%), pengaturan jarak dan jenis sinyal
(~5%), gen spesifik RNA (~1%), dan beberapa sekuen berulang, umumnya pada
urutan panjang beberapa pasang basa. Beberapa bakteri mungkin memiliki
plasmid sebagai elemen genetik ekstrakromosomal. Pada beberapa contoh, gen
diturunkan dari plasmid yang ditemukan menyatu pada koromosom bakteri.
Transposable elements umunya merupakan komponen dari genom bakteri.
Sebagai contoh, wild strain dari Eschericia coli mengandung 1-10 salinan pada
paling sedikit dari 6 tipe yang berbeda dari sekuen insersi (penyisipan). Fraksi
nongenik dari genom (termasuk sekuen insersi, plasmid dan bekteriofag) nampak
pada satu urutan yang ukurannya lebih kecil dari fraksi kromosom. Yang lebih
menarik, pada semua spesies bakteri yang memiliki sekuen genom yang lengkap,
ditemukan petunjuk untuk gen fungsional melalui transfer gen horisontal. Pada
banyak kasus, transfer gen horisontal telah disimpulkan melalui daerah unik pada
isi/kandungan GC dan pemakaian kodon.
Distribusi dari ukuran genom pada bakteri adalah diskontinu,
menunjukkan ujung mayor dengan nilai berkisar antara 0,8 x 106, 1,6 x 106, dan
4,0 x 106 bp, dan beberapa ujung minor pada 7,2 x 106 dan 8,0 x 106 bp. Distribusi
ini membawa Roley dan koleganya untuk mengusulkan bahwa genom yang besar
seperti pada E. coli dapat berkembang dari genom kecil melalui siklus yang

berurutan pada duplikasi genom. Meskipun begitu, ukuran genom sudah
terakumulasi. Ujung pada distribusi ini cenderung menghilang. Namun, seperti
kebanyakan data ukuran genom yang telah diakumulasi, puncak pada distribusi
cenderung menghilang sebagai kesenjangan dalam distribusi. Dalam serangkaian
studi yang lebih baru, Labedan dan Riley tidak menemukan adanya bukti untuk
duplikasi genom dalam evolusi sejarah E. coli. Saat ini, hanya bakteri gram-
negatif yang menunjukkan distribusi diskontinu.
Semenjak dikemukakan pembahasan antara ukuran genom dan filogeni
bakteri, hal ini mendukung bahwa bertambahnya ukuran genom secara
berkelanjutan terjadi pada garis keturunan bakteri (Wallace dan Morowitz 1973).
Penggunaan filogeni bakteri sebagai dasar untuk membandingkan sekuen rRNA,
Herdman (1985) menghubungkan perubahan dalam ukuran genom yang
dipergunakan dalam sejarah filogeni. Hasil penyelidikan ini mengindikasikan
bahwa bertambahnya ukuran genom terjadi secara independen atau bebas pada
beberapa garis keturunan bakteri. Menariknya, bahwa banyak pertambahan
ukuran genom terjadi secara kebetulan pada beberapa garis keturunan bakteri dan
pada spesifik waktu yang lain pada sejarah evolusi di bumi, yakni pada saat
jumlah oksigen di atmosfer bumi tidak dapat diperkirakan, kira-kira 1,8 milyar
tahun yang lalu.
Distribusi ukuran genom pada bakteri dapat dijelaskan melalui kombinasi
beberapa proses: (1) banyak gen independen dan duplikasi operon, (2) delesi
dalam skala kecil dan insersi, (3) transposisi duplikatif, (4) transfer horisontal gen
terutama dari plasmid dan bakteriofag, dan juga dari spesies lain, dan (5)
hilangnya ujung masif DNA dalam sebagian besar parasit.
C. Genom Minimal
Pencarian genom dari “wujud replikasi autonom terkecil” telah dimulai
pada akhir 1950an oleh Morowitz dan rekannya. Dimulai dengan mempelajari
Mollicutes, yang mana merupakan organisme seluler dengan genom terkecil dan
jumlah gen terkecil di alam. Tidak ada bukti, bagaimanapun juga bahwa 468 gen
pengkode protein dalam M. genitalium benar-benar mewakili kebutuhan minimal
untuk mempertahankan hidup. Terdapat kemungkinan bahwa derajat tertentu
redundansi genetik ada bahkan dalam genom yang paling efisien. Berikut ini akan
dijelaskan dua pendekatan untuk menyimpulkan set gen minimal untuk kehidupan
selular.
1. Pendekatan Analitis
Dasar rasionil di belakang metode anatisis Koonin dan Mushegian (1996)
dan Mushegian dan Koonin (1996a) sangat berterus-terang. Perkiraan awal
komplemen gen minimal dilakukan dengan mengidentifikasi himpunan semua gen
ortolog yang umum untuk sekelompok organisme. Salah satu contohnya,
mengenai perbandingan proteom E. coli, H. influenzae, dan M. genitalium,
ditunjukkan pada Gambar 2.2. Dari perbandingan tersebut, dapat disimpulkan
perkiraan gen minimal ialah 239 gen.
Gambar 2.2 Diagram venn ortolog yang umum untuk gen pengkode protein antara tiga spesies bakteri. M. genitalium dan H. influenzae memiliki 240 kesamaan
orthologs, M. genitalium dan E. coli memiliki 257, dan H. influenzae dan E. coli memiliki 1,128. Terdapat 239 orthologs yang umum untuk ketiga spesies. (Sumber:
Graur dan Hsiung Li, 1999)
Selain gen pengkode protein, beberapa gen vital lain harus disertakan
perangkat minimal. Gen ini tidak dapat diidentifikasi pada tahap pertama analisis
karena adanya fenomena “pemindahan gen nonorthologous”, yang salah satu
bentuk konvergen fungsionalnya terbawa ketika digunakan dalam protein yang
tidak mempunyai hubungan untuk menunjukkan beberapa fungsi yang vital
(Gambar 2.3). Sebagai contoh, fungsi enzim glikolitik phosphoglycerate mutase
dijalankan pada bakteri yang berbeda oleha dua jenis protein yang tidak
mempunyai hubungan satu sama lain. Salah satunya dikode oleh gen gpm dan
merupakan 2,3-biphosphoglycerate-dependent, dan yang lain dikode oleh yibO
dan merupakan 2,3-biphosphoglycerate-independent. Pada M. genitalium fungsi

phosphoglycerate mutase ditunjukkan oleh produk gen yibO, sedangkan dalam H.
influenzae fungsi yang sama ditunjukkan oleh protein yang dikode gen gpm.
Karena dua mutasi phosphoglycerate yang tidak berhubungan ini terletak pada
sekuennya sendiri, perpotongan dua perangkat proteom tidak mengandung
keduanya, meskipun begitu fungsi katalitiknya itu kemungkinan diperlukan untuk
hidup. Kira-kira dua dosin gen diketahui dilibatkan dalam pemindahan gen
nonorthologous, dan itu ditambahkan pada awal set minimal.
Gambar 2.3 Sebuah skenario hilangnya gen diferensial untuk perpindahan gen nonortholog. Berasal dari nenek moyang yang memiliki dua protein (lingkaran dan segitiga) melakukan fungsi serupa. Pengkodean gen salah satu dari mereka hilang dalam keturunan 1, sedangkan yang lainnya hilang dalam keturunan 2. hasilnya
adalah konvergensi fungsional. (Sumber: Graur dan Hsiung Li, 1999)
Akhirnya, gen yang muncul secara khusus pada bakteri parasit atau gen
yang menunjukkan fungsi redundan telah dihapus, ditunjukkan pada set gen
minimal dari 256 hen pada bakteri.
Dari pendekatan ini, set gen minimal yang telah ditemukan mencakup: (1)
sebuah sistem yang hampir sempurna dari translasi; (2) mesin replikasi DNA yang
hampir lengkap; (3) sebuah perangkat dasar dari gen untuk rekombinasi dan
perbaikan DNA; (4) sebuah perangkat transkripsi yang terdiri dari empat unit
RNA polimerase; (5) seperangkat besar protein penjaga; (6) sedikit gen pengkode
protein yang terlibat dalam metabolisme anaerob; (7) beberapa gen yang
mengkode enzim untuk lemak dan biosintesis kofaktor; (8) beberapa protein
transport pada transmembaran; dan (9) satu set dari 18 protein yang tidak
diketahui fungsinya. Yang perlu diperhatikan pada set minimal iniadalah bahwa
tidak terdapat mesin esensial untuk biosintesis asam amino dan nukleotida, yang

sebelumnya dipercaya sudah didapatkan dari lingkungan dalam bentuk “siap
pakai”.
2. Pendekatan Eksperimental
Sebuah pendekatan eksperimental untuk masalah genom minimal telah
dilaksanakan oleh Itaya (1995). Tujuh puluh sembilan lokus pengkode protein
terpilih secara acak pada bakteri gram positif Bacillus subtilis tersingkir melalui
mutagenesis (Gambar 2.4). Mutasi yang hanya pada 6 dari semua lokus membuat
B. subtilis tidak mampu tumbuh dan membentuk koloni, selama mutan istirahat 73
lokus mempertahankan kemampuannya untuk membelah. Hanya tiga dari enam
lokus pengkode protein yang telah diidentifikasi secara jelas fungsinya. Yaitu
dnaA dan dnaB, yang terlibat dalam inisiasi pada replikasi DNA, dan rpoD, yang
merupakan bagian hasil dari sintesis RNA.
Gambar 2.4 Lokasi genomik dari 79 lokus yang dipilih secara acak (baris) dalam Bacillus subtilis yang telah tersingkir oleh mutagenesis. Enam lingkaran yang solid
menunjukkan lokus yang yang harus ada, hanya tiga yang teridentifikasi. Data diambil dari Itaya (1995). (Sumber: Graur dan Hsiung Li, 1999)
Untuk memastikan gen yang tersingkir tersebut tidak mempengaruhi
pertumbuhan yang bukan bagian dari redundan famili multigen, Itaya (1995) juga
mengkonstruk bakteri dengan berbagai mutasi. Menariknya, bahkan ketika 33
lokus yang secara simultan lumpuh, bakteri dan turunannya mempertahankan
kemampuan mereka untuk membentuk koloni. Maka, 73 dari 79 gen diduga
benar-benar tidak diperlukan, sementara hanya sekitar 7,5% genom dianggap
diperlukan. Panjang genom B. subtilis adalah 4,2 x 106 bp, dan diasumsikan
bahwa perbandingan genom yang diperlukan dibanding gen yang tidak diperlukan

adalah sama, panjang genom yang diperlukan diperkirakan mencapai 4,2 x 106 x
0,075 = 3,2 x 105 bp. Dengan menggunakan 1,25 Kb sebagai ukuran rata-rata dari
gen pengkode protein, peroleh sebuah perkiraan set gen minimal dari
320.000/1.250 = 254 gen.
D. Miniaturisasi Genom
Beberapa kesimpulan umum telah dicapai pada pokok bahasan evolusi
morfologi. Pada perbandingannya, salah satu aturan terkecil yang jelas dapat
disimpulkan mencakup pengaruh dari tidak digunakannya tingkatan molekuler:
reduksi drastis pada ukuran genom (miniaturisasi genom) selalu diasosiasikan
dengan kehilangan fungsi. Khususnya, bentuk hidup berupa parasit atau
endosimbiotik yang ditemukan mempengaruhi ukuran genom secara mendalam
dan jika kita lihat sebelumnya, genom bakteri terkecil dimiliki oleh parasit
endoseluler.
Miniaturisasi genom dapat terjadi melalui dua proses: transfer gen atau gen
yang hilang. Pada penjelasan berikutnya akan dibahas mengenai reduksi ukuran
genom yang dikarenakan endosimbiosis dan parasit secara terpisah.
1. Reduksi Ukuran Genom yang Mengiringi Endosimbiosis
Miniaturisasi menyeluruh pada genom mengikuti kejadian endosimbiosis
yang memunculkan peristiwa pada mitokondria dan kloroplas. Beberapa organela
kemungkinan redundan dan hilang tanpa adanya penggantian melalui delesi;
lainnya ditransfer secara massal menuju genom inti. Sebagai contoh, inti genom
yeast mengandung sekitar 300 gen pengkode protein yang fungsinya secara
khusus pada mitokondria. Genom mitokondria ini, hanya mengandung 8 gen
pengkode protein. Kiranya, beberapa gen inti yang menghasilkan fungsi dalam
mitokondria dahulu merupakan bagian genom mitokondria, yang saat ini kapasitas
kodenya sangat terbatas. Meskipun genom mitokondria dengan kapasitas kode
terbesar, pada flagela heterotrop Reclimonas americana, hanya mengandung 62
gen pengkode protein, jauh lebih kecil dari jumlah gen yang dibutuhkan untuk
hidup.
Selain mitokondria dan kloroplas, banyak organela eukariotik lain yang
diturunkan melalui endosimbiosis di antara organisme independen. Margulis, dkk

(1979) mengajukan bahwa flagel, silia, dan organel yang lain dari sel motil
diturunkan dari spirochetes yang lalu diasosiakan bersimbiosis dengan nenek
moyang eukariot. Jika usulan tersebut ternyata benar, maka organel ini pasti telah
mengalami miniaturisasi genom maksimal yaitu, mereka telah kehilangan seluruh
genom mereka.
Contoh menarik reduksi genom yang mengikuti endosimbiosis mencakup
Chlorarachniophyta, sekelompok amoeba berflagel yang memperoleh kapasitas
fotosintesis dengan menelan dan mempertahankan flagel alga hijau (kelas
Ulvophyceae). Alga endosimbian mempertahankan kloroplas, nukleus,
sitoplasma, dan membran plasma. Sisa nukleus, yang disebut nukleomorph,
mengandung tiga kromosom linear kecil dengan jumlah total ukuran genom
haploid sekitar 380.000 bp, yang diketahui sebagai genom eukariot terkecil.
Genom nukleomorph merupakan intisari dari kepadatan: ruang rata-rata antara
gen yang berdekatan lebih 65 bp, beberapa gen overlap dan lainnya ditranskripsi,
dan gen tersebut di-disrupted oleh intron spliceosomal terkecil (18-20 bp) yang
pernah ditemukan. Seperti yang diharapkan, sebagian besar protein dalam
endosimbion akan diimpor dari host.
2. Reduksi Ukuran Genom pada Parasit
Parasitisme melibatkan hubungan yang intim antara dua organisme:
sebuah inang yang menyediakan banyak keperluan metabolik dan fisiologis bagi
yang lain, yaitu yang memparasit. Parasitisme selalu mengakibatkan kehilangan
fungsi genetik pada parasit dan sebagai akibatnya terjadi reduksi pada ukuran
genom. Sebagai contoh, tumbuhan Epiphagus virginiana, sebuah parasit relatif
nonfotosintesis dari lavender, basil, dan catnip, yang mempunyai genom kloroplas
sangat kecil (~70.000 bp) yang mengandung hanya 42 gen. Dapat dipahami,
semua gen untuk fotosintesis dan klororespirasi tidak tersedia. Belum jelas,
mengapa semua kloroplas yang dikode gen RNA polimerase, gen pengkode
protein ribosom dan banyak gen sepesifik tRNA akan hilang.
E. Ukuran Genom pada Eukariot dan Nilai C Paradox
Nilai C pada eukariot biasanya lebih besar daripada prokariot, tetapi ada
pengecualian. Contohnya, yeast S. cerrevisiae mempunyai genome yang

ukurannya hampir sama dengan beberapa bakteri gram positif, seperti
Streptomyces coelicolor dan S. rimosus, dan lebih kecil dari kebanyakan spesies
Cyanobacteria terutama genus Calothrix. Namun, karena genom inti eukariotik
berasal dari replikasi ganda sementara prokariota hanya memiliki satu, eukariota
dapat mengalami replikasi DNA dalam jumlah yang lebih besar dari DNA tiap
satuan waktu daripada prokariota.
Variasi nilai C pada eukariot jauh lebih besar daripada bakteri, dari 8,8 x
106 bp sampai 6,9 x 1011 bp, kira-kira 80.000 kali lipat (Tabel 2.3). Protista
uniseluler, terutama amoeba sarcodine menunjukkan variasi nilai C yang terbesar
melebihi kisaran 20.000 kali lipat. Dalam perbandingannya, rentangan dari nilai C
pada seluruh kingdom animalia, dari porifera sampai manusia, kira-kira hanya
3.000 kali lipat. Tiga kelas amniota (mamalia, burung, dan reptilia) tidak termasuk
diantara eukariot dalam variasi ukuran genom mereka yang kecil (hanya sampai
empat kali lipat). Untuk kelas yang lain, dari data nilai C yang ada, menunjukkan
variasi minimal 100 kali lipat.
Tabel 2.3 Kisaran Nilai C pada Beberapa Kelompok Eukariot
Data dari Sparrow et al. (1972), Cavalier-Smith (1985), dan beberapa sumber lain.(Sumber: Graur dan Hsiung Li, 1999)

Menariknya, variasi interspesifik yang sangat besar dalam ukuran genom
diantara eukariotik tampaknya tidak berhubungan dengan kekomplekan
organisme atau jumlah kemungkinan gen yang dikode oleh organisme.
Contohnya, beberapa protozoa uniseluler memiliki lebih banyak DNA daripada
mamalia, yang diperkirakan lebih komplek. Organisme yang memiliki kemiripan
morfologi dan anatomi yang komplek (bawang dan lili, Paramecium aurelia dan
P. caudatum) menunjukkan luasnya perbedaan nilai C (Tabel 2.4). Kurangnya
kecocokan antara nilai C dan banyaknya perkiraan dari informasi genetik
membuat genom menjadi lebih dikenal dalam literatur sebagai nilai C paradox.
Nilai C paradox juga terbukti dalam perbandingan beberapa spesies (spesies yang
morfologinya sangat mirip antara yang satu dengan yang lain sehingga tidak dapat
dibedakan fenotipnya). Pada protista, bony fish, amfibi dan tanaman berbunga,
beberapa spesies tertentu memiliki perbedaan nilai C yang besar. Spesies yang
sesaudara memiliki banyak perbedaan dalam nilai C, meskipun menurut definisi
tidak ada perbedaan dalam kompleksitas organismik. Karena itu tidak dapat
diasumsikan bahwa organisme memiliki DNA kurang dari jumlah yang
dibutuhkan untuk fungsi-fungsi vitalnya, harusnya dijelaskan mengapa tampaknya
begitu banyak spesies mengandung kelebihan DNA yang cukup besar.
Tabel 2.4 Nilai C Beberapa Organisme Eukariot yang Diurutkan Berdasarkan Ukuran Genom

Pertanyaan pertama untuk mengklarifikasi apakah ada hubungan antara
ukuran genom dengan jumlah gen. Dengan kata lain, perbedaan khusus dalam
ukuran genom dapat disebabkan oleh DNA genik dan DNA nongenik? Jika
variasi nilai C disebabkan oleh gen, maka variasi nilai C dapat dibedakan ke
dalam 1) Jumlah protein-pengkode gen, 2) Ukuran protein, 3) Ukuran protein-
pengkode gen, 4) Jumlah dan ukuran gen lain dari protein pengkode.
Tentu saja, harus disadari bahwa dalam ketiadaan penentuan sekuen
genom yang sepenuhnya, pemastian jumlah gen dalam spesies menjadi tugas yang
sangat sulit. Pada gen pengkode protein, tugas ini diselesaikan dengan
menggunakan elektroforesis gel dua dimensi, protein dipisahkan oleh tekanan
pada dimensi pertama dan oleh titik isoelektrik (pH pada protein tidak bermuatan)
pada dimensi kedua. Hasilnya adalah kumpulan bintik yang ukurannya berbeda-
beda yang tersebar ke seluruh gel. Jumlah bintik tersebut akan membantu kita
dalam memperkirakan jumlah protein dalam sebuah sel. Pada kenyataannya
pemisahan tersebut sulit terjadi, biasanya bintik yang terbentuk tersebut kurang
jelas atau suram. Jumlah gen yang ditentukan dengan metode ini biasanya
diremehkan. Contohnya, jumlah protein-pengkode gen pada S. cerrevisiae telah
diperkirakan dengan elektroforesis dua dimensi sekitar 3.000. Jumlah protein-
pengkode gen bisanya dikenali dalam unting genom lebih dari dua kali (sekitar
6.200 gen). Meskipun demikian tetap dihunakan perkiraan yang berasal dari
beberapa metode untuk menyamakan tujuan, jumlah tersebut juga dapat
digunakan sebagai indikator relatif dari jumlah gen yang benar.
Jumlah protein pengkode gen pada eukariot biasanya hampir melebihi 50
kali lipat. Variasi ini tidak cukup jelas untuk menjelaskan mengenai 80.000 kali
lipat variasi dalam DNA inti. Jumlah gen berkorelasi positif dengan kompleksitas
sedangkan ukuran genom tidak. Variasi khusus pada rantai molekul mRNA
menjelaskan tentang nilai C paradox. Sementara perbedaan kecil pada daerah
pengkode dan non-pengkode diantara organisne yang berbeda, tidak ada
hubungannya dengan panjang gen dan ukuran genom. Contohnya mRNA hanya
sedikit lebih panjang pada organisme multiseluler daripada pada protista (1.400-
2.200 bp dibanding 1.200-1.500 bp). Meskipun demikian organisme dengan
genom yang lebih besar tidak selalu menghasilkan protein yang lebih besar.

Perbedaan pada ukuran gen (panjang intron dan daerah non-kode lainnya) tidak
dapat menunjukkan jumlah variasi pada ukuran genom. Gen hewan 3-7 kali lebih
panjang dibandingkan panjang rata-rata gen protista dan gen dari vertebrata 2-4
kali lebih besar daripada semua invertebrata, tidak ada hubungan antara ukuran
genome dan rata-rata panjang gen.
Mengenai jenis lain dari DNA genik, berkorelasi positif antara duplikat
dari beberapa RNA-gen spesifik dan ukuran genom. Korelasi tersebut tampak
pada ukuran genom dan jumlah copian dari gen yang tidak diterjemahkan yang
terlibat dalam replikasi kromosom segregasi, dan rekombinasi selama miosis dan
mitosis. Meskipun demikian gen hanya menyusun fraksi dari genom, misalnya
variasi pada jumlah RNA-gen spesifik dan gen yang tidak diterjemahkan tidak
dapat menjelaskan adanya variasi pada ukuran genom.
Cara lain untuk membandingkan jumlah gen antara dua genom adalah
membandingkan polysomal polyadenilated RNA complexity. Panjang total dari
berbagai molekul mRNA dihasilkan oleh suatu jaringan khusus. Perbandingan ini
juga menunjukkan tidak adanya korelasi antara jumlah gen dan ukuran genome.
Contohnya polysomal RNA complexity pada hati ayam adalah 2 x 10 nukleotida,
sedangkan polysomal RNA complexity pada hati tikus adalah setengah dari
jumlah pada hati ayam, walaupun pada kenyataannya ukuran genom pada tikus
lebih dari dua kali ukuran genom ayam.
Ringkasnya, fraksi DNA nongenic sebagai pelaku tunggal untuk nilai C
paradoks. Dengan kata lain, sebagian besar dari genom eukariotik terdiri dari
DNA yang tidak mengandung informasi genetik. Telah diperkirakan bahwa
jumlah DNA nongenic per genom bervariasi pada eukariotik sekitar 3.0 x 103 Kb
sampai 108 Kb (kisaran 300.000 kali lipat) dan tersusun kurang dari 30% sampai
99,998% dari genom.
F. Mekanisme untuk Peningkatan Global pada Ukuran Genom
Untuk menjelaskan keberadaan sejumlah besar DNA non-genic dalam
genom eukariota, terdapat proses yang dapat meningkatkan ukuran genom.
Peningkatan ukuran genom ada dua jenis, yaitu:
1. Peningkatan global, dimana seluruh genom atau kromosom diduplikasi

2. Peningkatan regional, dimana setiap bagian partikel sekuen digandakan untuk
generasi DNA repetitive
1. Poliploidisasi
Karena genom eukariota secara signifikan lebih besar daripada bakteri,
evolusi eukariota dari nenek moyang prokariota terlibat dalam peningkatan ukuran
genom. Ada beberapa mekanisme molekular yang meningkatkan ukuran genom.
Salah satu mekanisme tersebut adalah poliploidisasi, yaitu penambahan satu set
atau lebih kromosom pada set kromosom asli. Suatu organisme dengan sel yang
megandung empat salinan autosom disebut tetraploid, enam salinan disebut
hexaploid dan seterusnya. Gamet dari organisme poliploid tidak haploid, dan
organisme dengan autosom ganjil seperti tanaman pisang domestik triploid (Musa
acuminata) tidak dapat melakukan meiosis dan bereproduksi secara seksual.
Ada dua tipe utama poliploidi: allopoliploidi, yaitu kondisi yang timbul
dari kombinasi set kromosom genetik yang berbeda. Dan autopoliploidi, yaitu
penggandaan satu set kromosom dasar. Autotetraploidi (atau tetraploidi) adalah
duplikasi genom. Duplikasi genom terjadi sebagai konsekuensi dari kurangnya
pemisahan antara semua kromosom anakan hasil replikasi DNA.
Tetraploidi adalah kejadian mutasi umum di alam. Tetraploidi somatik
ditemukan pada hampir seluruh organisme, termasuk protisa, alga, tanaman,
molusca, serangga, dan mamalia. Namun selama sejarah evolusioner, tetraploidi
jarang ditemukan. Karena dalam banyak kasus, tetraploidi merusak. Efek
perusakan meliputi:
a. Perpanjangan waktu pembelahan sel
b. Peningkatan volume nukleus
c. Peningkatan jumlah pemisahan kromosom selama meiosis
d. Ketidakseimbangan genetik, dan
e. Gangguan dengan diferensiasi seksual ketika jenis kelamin organisme
ditentukan oleh salah satu rasio antara jumlah kromosom sex dan jumlah
autosom seperti pada Drosophila atau oleh urutan poliploid pada
Hymenoptera.
Dalam beberapa kasus, tetraploidi (atau tingkatan ploidi yang lebih tinggi)
nampaknya tidk berpengaruh pada fenotip. Sebagai contoh, diploid dan poliploid

spesies Chrysanthemum berbeda dalam kromosom nomor 18-198, namun mereka
hampir tidak bisa dibedakan satu sama lain. Hal serupa terjadi pada mawar
(Rosa), katak leptodaktil (Odontophrynus), dan ikan mas (Carasius). Anehnya
dalam beberapa kasus tetraploidisasi mungkin bermanfaat. Pada tumbuhan
misalnya, poliploidi mengurangi infertilitas hybrid dan menyebabkan hilangnya
ketidakcocokan diri, sehingga tanaman individual di tepi habitat dapat
berkembang biak dengan penyerbukan sendiri.
Dalam tetraploid yang baru terbentuk, tidak ada yang dapat berbicara
tentang peningkatan nilai C, karena nilai ini mengacu pada ukuran genom haploid
dan tidak tergantung pada tingkat ploidi. Namun, dua genom yang mengalami
mutasi, translokasi, penyusunan ulang kromosom, dan perubahan nomor
kromosom akhirnya akan menjadi genom single baru, situasi ini disebut
criptopoliploidi. Dengan kata lain, poliploid tidak akan dibedakan dari diploid.
Criptopoliploidi dapat menjelaskan banyak variasi ukuran genom pada tanaman,
amfibi, dan ikan bertulang.
Distribusi polimodal ukuran genom telah terdaftar pada beberapa
kelompok eukariota. Terutama pada monokotil, dimana ukuran genom
menunjukkan distribusi polimodal dengan puncaknya pada 0,60x106, 1,18x106,
4,51x106, dan 8.53x106 Kb (Gambar 2.5). Distribusi yang sama dapat diamati
pada Echinodermata, insekta, dan fungi, dan pada tingkat lebih rendah pada
amfibi dan ikan bertulang. Dengan demikian, duplikasi genom nampaknya
mnejadi mekanisme utama dalam mekanisme evousi ukuran genom pada
eukariota. Menariknya, masing-masing putaran duplikasi genom hanya
menyebabkan sebagian kecil DNA yang hilang, sehingga jumlah DNA setelah
setiap putaran meningkat dengan sedikir faktor.

Gambar 2.5 Distribusi frekuensi ukuran genom dalam 80 spesies rumput (family Poaceae). Puncak di distribusi multimodal ditandai dengan panah (Sumber: Graur
dan Hsiung Li, 1999)Mengingat bahwa genom mamalia sekitar 1.000 kali lebih besar dari
genom bakteri, dan dengan asumsi bahwa duplikasi genom bertanggung jawab
untuk pembesaran genom, dapat disimpulkan bahwa hanya sekitar sepuluh
putaran duplikasi genom yang diperlukan untuk memperbesar genom dari ukuran
bakteri primodial menjadi ukuran mamalia. Dengan cara lain, duplikasi genom
terjadi rata-rata sekali setiap 300-350 juta tahun. Jika kandungan DNA meningkat
secara terus-menerus dengan penambahan kecil potongan DNA melalui
transposisi atau unequal crossing over, maka tingkat pertumbuhan genom dari
ukuran bakteri menjadi ukuran mamalia seharusnya sekitar 6-7 nukleotida per
tahun.
Selama poliploidisasi, hilangnya duplikat gen trjadi sangat cepat. Sebagai
contoh, gandum Triticum aestivum adalah allohexaploid yang ada sekitar 10.000
tahun lalu. Dalam waktu yang singkat beberapa lokus rangkap tiga menghilang.
Diperkirakan bahwa proporsi enzim yang dihasilkan oleh lokus triplet, duplet, dan
tunggal pada gandum masing-masing 57%, 25%, dan 18%.
Poliploidi merupakan faktor penting dalam spesiasi. Khususunya
reproduksi seksual autotetraploid yang secara otomatis diisolasi dari nenek
moyang diploid karena menghasilkan gamet diploid, yang menggabungkan gamet
haploid dan diploid. Yang menghasilkan keturunan triploid. Organisme dengan
jumlah autosom ganjil tidak dapat bereproduksi secara seksual, sehingga
poliploidi menggambarkan mekanisme yang efektif untuk isolasi reproduksi.

2. Polisomi
Aneuploidi adalah kondisi dimana jumlah kromosom dalam sel bukan
merupakan kelipatan integral dari set haploid untuk spesies. Euploidi mengacu
pada jumlah kromosom yang merupakan kelipatan dari kromosom haploid.
Terkait dengan mekanisme yang bertanggung jawab untuk peningkatan ukuran
genom, ada dua tipe aneuploidi: duplikasi kromosom lengkap (polisomi), dan
duplikasi bagian utama dari kromosom (polisomi parsial).
Polisomi paling sering merugikan. Pada mamalia misalnya, terkait dengan
letalitas dan infertilitas. Pada manusia, contoh dari polisomi termasuk anomaly
seperti syndrome Down (trisomi 21), dan trisomi 18. Kerusakan parah dikaitkan
dengan polisomi parsial (misalnya cat-eyes syndrome). Oleh karena itu, duplikasi
kromosom lengkap atau sebagian tidak memberikan kontribusi yang signifikan
untuk meningkatkan ukuran genom.
3. Duplikasi pada Genom Yeast, Tetraploidi atau Regional
Saccharomyces cerevisiae telah lama diduga sebagai sebuah
cryptotetraploid. Secara sistematik, proteome yeast dicari untuk daerah duplikasi
(Gambar 2). Kriteria yang digunakan untuk mendefinisikan dua daerah duplikasi
adalah:
a. Kesamaan sekuen antara dua daerah yang bergabung dengan kemungkinan
lebih kecil dari 10-18 secara kebetulan
b. Setidaknya ada tiga gen yang sama, dengan jarak antar gen kurang dari 50
Kb, dan
c. Konservasi sekuen gen orientasi relative gen. Menurut kriteria ini, telah
diidentifikasi 54 pasang daerah duplikasi non-overlapping mencakup sekitar
50% genom yeast (Gambar 2.6).

Gambar 2.6. Lokasi 54 daerah duplikasi non-overlapping (kotak padat) pada genom yeast. Dua salinan dari masing-masing daerah diplikasi diberi nomor yang sama di
bawah kotak masing-masing. Nomor tercantum dalam terjadinya kromosom. Jumlah gen homolog di masing-masing daerah duplikasi tercantum di atas kotak tersebut. Jumlah kromosom ditulis dalam angka romawi. (Sumber: Graur dan
Hsiung Li, 1999)
Ada dua kemungkinan penjelasan untuk pengamatan tersebut:
a. Daerah duplikasi dibentuk secara mandiri dengan banyak duplikasi regional
yang terjadi pada waktu yang berbeda selama evolusi S. cerevisiae, atau
b. Daerah duplikasi dibentuk secara bersamaan oleh kejadian tetraploidisasi,
dilanjutkan dengan penyusunan ulang genom dan hilangnya banyak gen
duplikat. Ada dua alasan untuk mendukung model yang terakhir; pertama, 50
daerah duplikasi mempertahankan orientasi yang sama dengan mengarah ke
sentromer. Kedua, berdasarkan distribusi poisson, 54 daerah duplikasi
diharapkan dapat menghasilkan sekitar tujuh daerah triplikasi.

S. cerevisiae merupakan tetraploid kuno, yang dibentuk melalui fusi dari
dua nenek moyang genom yeast diploid, masing-masing berisi sekitar 5.000 gen.
tetraploidisasi terjadi sekitar 100 juta tahun yang lalu pada nenek moyang empat
spesies Saccharomyces setelah penyimpangan dari S. kluyveri. Spesies baru itu
menjadi cryptotetraploid, dan sekitar 92% salinan duplikat gen hilang melalui
delesi atau penghapusan sekuen. Terdapat 70-100 gangguan pemetaan (misalnya
translokasi secara regional) yang disimpulkan dapat menjelaskan distribusi
kromosom yang terjadi saat duplikasi gen (Gambar 2.7).
Gambar 2.7 Skema nomor gen dan urutan evolusi gen dalam duplikasi genom seperti yeast. Sebuah genom skematis ditunjukkan dengan dua kromosom (satu kotak) dan 26 gen (A sampai Z). Huruf besar dan huruf kecil digunakan untuk
membedakan antara dua set asli kromosom. Pada tahap terakhir, efek dari peristiwa rekombinasi dalam dua gen paralogous ditampilkan. Kejadian ini
menghasilkan dua gen hybrid baru (E dan e ') dan urutan gen baru. (Sumber: Graur dan Hsiung Li, 1999)

4. Poliploidi dari Genom Vertebrata
Vertebrata memiliki gen lebih besar dari invertebrata. Gen tunggal
invertebrate biasanya berhubungan hingga empat gen vertebrata pada kromosom
yang berbeda. Nampaknya sekuen dari keempat salinan memiliki jarak yang sama
satu dengan yang lain. Pola ini pertama kali diamati untuk kelompok gen Hox,
tetapi menurut Spring (1977), fenomena ini adalah umum. Ia mengajukan
hipotesis, menurutnya munculnya vertebrata ini dimungkinkan oleh dua putaran
tetraploidisasi, sehingga terbentuk duplikasi empat genom. Dengan demikian,
vertebrata mungkin sebenarnya termasuk cryptoploid
G. Pemeliharaan DNA Non-genik
Berbagai upaya telah dilakukan untuk memecahkan paradox nilai C.
Berikut ini adalah empat hipotesis dan bukti empiris yang bersangkutan:
1. Hipotesis
a. Hipotesis seleksionis, menegaskan bahwa DNA non-genik melakukan fungsi
penting, seperti regulasi global ekspresi gen. Menurut hipotesis ini, kelebihan
DNA adalah jelas, dan DNA sepenuhnya fungsional. Akibatnya, jika terjadi
delesi pada DNA akan merusak organisme.
b. Hipotesis netralis, menyatakan bahwa fraksi DNA non-genik pada genom
aukariotik kurang berfungsi secara genetik dan fisiologis. Ohno (1972)
menyebut DNA ini sebagai DNA sampah untuk menjelaskan ketidak
berfungsiannya. Menurut pandangan hipotesis ini, DNA non-genik hanya
merupakan hasil kebetulan semata selama proses evolusi dan tidak
mempengaruhi kemampuan organisme, tetapi ini akan diteruskan dari
generasi ke generasi yang tak terbatas.
c. Hipotesis seleksionis intragenomik, menganggap DNA non-genik sebagai
“parasit fungsional” atau “simbion genetik” yang terakumulasi dalam genom
dan secara aktif dikelola oleh seleksi intragenomik karena tingginya tingkat
reproduksi dibandingkan dengan fraksi genom. Hal itu umum terjadi untuk
menemukan istilah DNA selfish yang diterapkan untuk fraksi non-genik.
DNA selfish memiliki dua sifat yang berbeda:

1) Muncul ketika sekuen DNA menyebar dengan membentuk salinan diri
tambahan dalam genom, dan
2) Tidak membuat kontribusi khusus untuk kesehatan orgnisme inang, atau
bahkan merugikan
Mekanisme utama untuk mengikat DNA selfish adalah duplikasi transposisi
dan jenis DNA selfish yang paling banyak adalah elemen transposabel dan
retrotransposabel. Perbedaan krusial antara DNA selfish dengan DNA sampah
adalah DNA selfish mampu melakukan amplifikasi sendiri, sedangkan DNA
sampah melakukannya secara pasif dalam genom. Dengan demikian, DNA
sampah dipertahankan dalam populasi dengan pergeseran genetik acak,
sedangkan DNA selfish dikelola dengan tipe insersi-kuasi delesi-equilibrium,
dimana proses eliminasi dengan pemilihan DNA selfish terlalu lambat untuk
mengimbangi laju akumulasi. DNA selfish cenderung mengalami
peningkatan dalam genom. Namun tidak dapat meningkat tanpa batas waktu,
karena organisme dengan kelebihan jumlah DNA non-genik akan mengalami
metabolisme.
d. Hipotesis Nukleotipik, menghubungkan fungsi struktural untuk DNA non-
genik, yaitu fungsi yang tidak berhubungan dengan sifatnya yang membawa
informasi genetik. Salah satu skema nukleotipik tersebut telah diusulkan oleh
Cavalier-Smith (1978,1985a), yang berpendapat bahwa harus ada suatu
"kekuatan evolusi besar" yang mempertahankan genom besar. Hipotesis ini
menyatakan bahwa DNA bertindak sebagai "nukleoskeleton" yang
mempertahankan volume nukleus pada ukuran proporsional dengan volume
sitoplasma. Karena sel yang lebih besar membutuhkan inti yang lebih besar,
seleksi untuk volume sel tertentu secara sekunder akan menghasilkan pilihan
untuk ukuran genom tertentu. Menurut skema ini, kelebihan DNA
dipertahankan oleh seleksi, tetapi komposisi nukleotida dapat berubah secara
acak. Banyak fungsi nukleotipik tambahan telah dikaitkan dengan fraksi non-
genik, tapi semua hipotesis nukleotipik memiliki satu kesamaan; mereka
semua menganggap genom sebagai unit struktural dari arsitektur nukleus-
sebuah blok bangunan yang terbuat dari asam nukleat, bukan sekedar
pembawa informasi genetik.

2. Bukti
Sangat sedikit sekali bukti tentang hipotesis seleksionis. Bahkan,
kebanyakan indikasi menjelaskan bahwa sebagian besar apa yang sekarang
dianggap DNA non-genik memang tidak memiliki informasi genetik, dan dapat
dihapus tanpa efek fenotipik yang jelas. Oleh karena itu, kelebihan DNA pada
eukariota tidak menghasilkan sistem metabolisme sampai batas yang signifikan,
dan kebutuhan (misalnya, energi dan nutrisi) akan mempertahankan dan
mereplikasi sejumlah besar DNA non-genik yang tidak berlebihan. Namun, ada
beberapa kelemahan dalam mempertahankan sejumlah besar DNA non-genik.
Pertama, genom yang besar telah ditemukan menunjukkan sensitivitas yang lebih
besar untuk mutagen dari genom yang kecil. Kedua, memelihara dan mereplikasi
sejumlah besar DNA non-genik mempersulit atau membebani organisme tertentu,
terutama ketika sebagian besar genom adalah non-genik. Oleh karena itu dapat
diterima bahwa DNA nongenik hanya dapat terkumpul sampai kebutuhan untuk
organisme bereplikasi menjadi signifikan.
Sulit untuk membedakan antara hipotesis seleksionis intra genomik dan
hipotesis netralis dalam tingkatan konseptual, apalagi untuk menguji berdasarkan
data yang empirik. DNA selfish menjadi kontributor utama dari DNA non-genik,
meskipun ada mekanisme penting lainnya untuk menghasilkan DNA tersebut.
Namun, sebagian besar fraksi non-genik dari genom berasal dari DNA selfish
tidak lagi diterima. Banyak yang saat ini mengalami kondisi degenerasi elemen
transposabel, dimana dihadapkan pada kematian apabila tidak lagi mampu
melakukan transposisi.
Membedakan antara eksperimen DNA sampah dan penjelasan
nucleoskeletal memang cukup sulit, Pagel dan Johnstone (1992) mengusulkan dua
ekspektasi yang berasal dari masing-masing dua teori, bahwa harga utama dari
DNA sampah adalah waktu yang diperlukan untuk melakukan penggandaan.
Organisme yang berkembang lebih lambat karena itu mungkin bisa
"mentoleransi" jumlah yang lebih besar dari DNA sampah, dan dengan demikian
korelasi negatif di seluruh spesies antara ukuran genom dan tingkat perkembangan
akan diperkirakan. Sebaliknya, perkiraan hipotesis nucleoskeletal adalah untuk
korelasi positif antara ukuran genom dan ukuran sel. Sayangnya, organisme

dengan sel yang besar juga cenderung berkembang secara perlahan, sedangkan
organisme yang lebih cepat tumbuh biasanya memiliki sel lebih kecil. Jadi,
menurut hipotesis DNA skeletal korelasi negatif antara tingkat perkembangan dan
nilai C juga diharapkan. Namun, menurut hipotesis nucleotypic, hubungan antara
tingkat perkembangan dan ukuran genom terjadi kemudian, sebagai akibat dari
hubungan antara tingkat perkembangan dan ukuran sel.
3. Spesies yang Sama Memiliki Ukuran Genom yang Berbeda
Terdapat perbedaan dalam ukuran genom antara organisme yang terkait erat,
dimana paradoks nilai C tidak dapat dijelaskan dengan menerapkan fungsi
nukleotipik, karena tidak adanya perbedaan nukleotipik. Yang tersisa hanyalah
dua kemungkinan mekanistik: baik ada perbedaan dalam tingkat akumulasi DNA
sampah, atau ada perbedaan dalam tingkat organisme berbeda yang
menghilangkan DNA sampah.
Untuk waktu yang cukup lama telah diketahui bahwa genom spesies
Drosophila mengandung pseudogen yang sangat sedikit (Vanin 1985;Weiner
dkk1986;. Wildf 1986). Baru-baru ini, Fetrovdkk (1996) dan Petrov dan Hartl
(1998) menemukan bahwa kematian Helenaretroposons akibat kehilangan DNA
pada tingkat yang luar biasa tinggi selama evolusi. Mereka menempatkan dua dan
keduanya, serta menyarankan bahwa maraknya pengahapusan daerah DNA yang
tidak mengikuti tingkat kendala selektif, dan mereka lebih lanjut terekstrapolasi
pada tingkat penghapusan yang berbeda, bukan tingkat akumulasi, yang dapat
menyebabkan perbedaan dalam ukuran genom antara taksa. Asumsi mereka
adalah bahwa tingginya tingkat penghapusan tidak terbatas pada elemen Helena
sendiri, tetapi bahwa fenomena tersebut yang berlaku umum untuk semua wilayah
seleksi yang tidak terbatas.
Untuk menguji asumsi ini, mereka membandingkan ukuran intron di antara
dua spesies Drosophila. D.virilis memiliki genom dua kali lebih besar dari D.
melanogaster (Moriyama etal. 1998). Perbedaan ini dapat dikaitkan dengan
heterokromatin, tetapi bahkan jika faktor ini diperhitungkan, genom D.virilis
masih sekitar 36% lebih besar dari D melanogaster. Dalam perbandingannya 115
intron lengkap dikumpulkan dari 42 gen ortolog, mereka menemukan bahwa
perbedaan panjang intron antara kedua spesies Drosophila yang signifikan secara

statistik. Perbedaan panjang rata-rata antara intron D.virilis dan D.melanogaster
(masing-masing 394 dan 283; bp) adalah 39%, yang mengherankan dekat dengan
ukuran yang berbeda dalam fraksinonrepetitive antara genom. Dengan demikian,
tampaknya bahwa beberapa organisme lebih efisien dalam "membuang sampah"
dari yang lain (Petrov dan Hartl 1997).
H. Struktur Urutan yang Berulang dari Genom Eukariotik
Genom eukariotik ditandai dengan dua fitur utama, yaitu pengulangan
sekuen, dan komposisi kompartementalisasi menjadi fragmen yang berbeda
ditandai dengan komposisi nukleotida spesifik.
DNA berulang terdiri dari sekuen nukleotida dari berbagai panjang dan
komposisi yang terjadi beberapa kali dalam genom, baik bersama-sama atau
secara tersebar. Segmen DNA yang tidak berulang yang disebut sebagai salinan
tunggal atau DNA unik. Proporsi genom diambil oleh sekuens berulang sangat
bervariasi antara taksa. Dalam ragi, proporsi ini berjumlah sekitar 20% dari
genom. Pada hewan, proporsinya berkisar dari sekitar 5% pada nyamuk yang
tidak menggigit Chironomustetans untuk menuju 90% pada kadal
Necturusmasculosus. Pada mamalia, hingga 60% dari DNA adalah berulang. Pada
tumbuhan, proporsinya bisa melebihi 80%, dan nilai-nilai yang jauh lebih tinggi
juga telah terdaftar (Flavell, 1986).
Studi klasik kinetika reaksi reasosiasi DNA dengan Britten dan Kohne
(1968) menunjukkan bahwa genom eukariota tingkat tinggi dapat dibagi secara
kasar ke dalam empat fraksi (Gambar 2.8). Fraksi pertama disebut DNA foldback,
dan terdiri dari urutan palindromik yang dapat membentuk jepitan beruntai
struktur ganda segera setelah DNA terdenaturasi yang kemudian diizinkan untuk
renaturasi. Fraksi DNA foldback biasanya sangat kecil, meskipun di beberapa
organisme mungkin mencapai nilai lebih dari 10%.

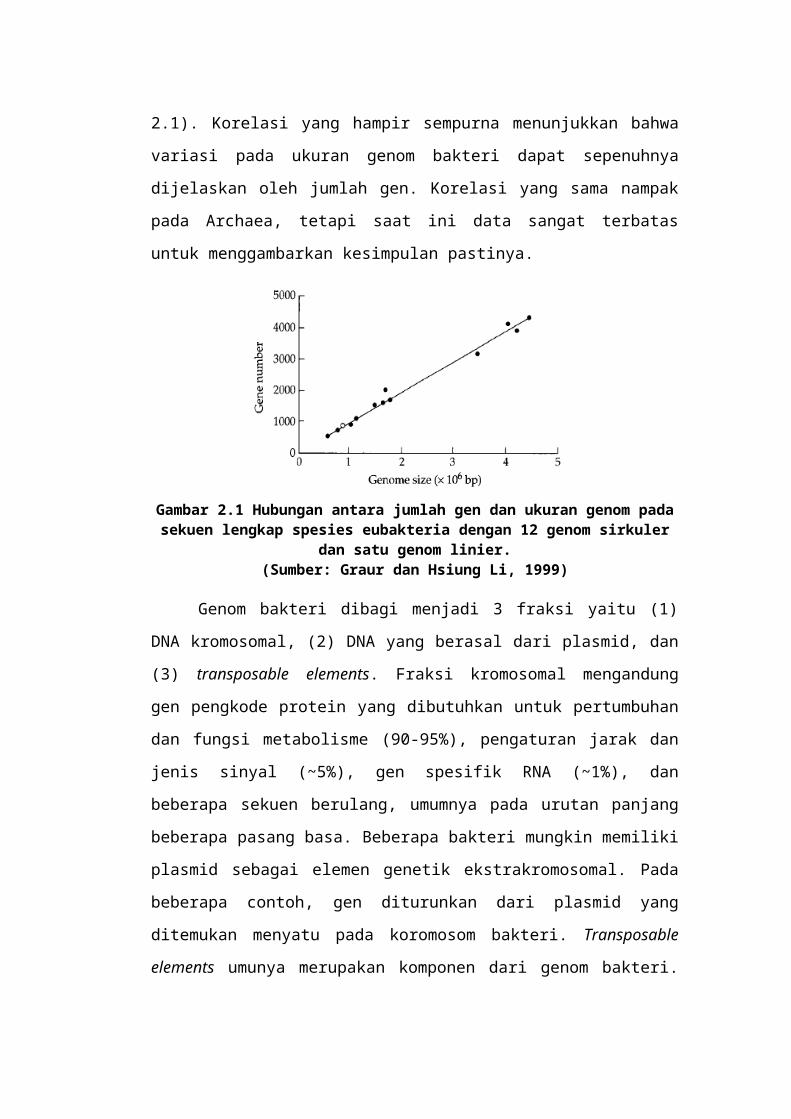
Gambar 2.8 Sebuah profil reasosiasi DNA mamalia. DNA dimurnikan, dipotong, dilelehkan dengan panas ke dalam untai tunggal, dan kemudian dibiarkan
reasosiasi melalui pendinginan bertahap. Persentase reasosiasi DNA untai ganda pada sumbu vertikal ditunjukkan sebagai fungsi dari produk konsentrasi DNA dan
waktu (C0t) pada sumbu horisontal. (Sumber: Graur dan Hsiung Li, 1999)
Beberapa DNA hanya reanneals pada nilai C0t tinggi (dibaca "cot"). Fraksi
ini terdiri dari satu salinan sekuen, dan karena sifat pewarnaan dalam persiapan
karyological, kadang-kadang disebut sebagai eukhromatin. Di antara kurang lebih
dua definisi komponen genom, terdapat sekuens DNA yang reanneal sebesar nilai
C0t menengah. Ini adalah kebiasaan untuk membagi urutan ini ke dalam DNA
yang sangat repetitif dan DNA berulang menengah. Fraksi sangat berulang terdiri
dari urutan pendek, dari beberapa ratusan panjang nukleotida, yang diulang ribuan
bahkan jutaan kali. Dalam persiapan karyological, fraksi sangat berulang tampak
gelap dan sangat bernoda dan disebut heterokhromatin. Fraksi berulang tengah
terdiri lebih dari ratusan atau ribuan urutan pasangan basa rata-rata, yang muncul
dalam genom hingga ratusan kali. Terdapat suatu rangkaian dari kedua ukuran
pengulangan dan nomorpengulangan dalam genom. Oleh karena itu, istilah DNA
sangat repetitif dan DNA berulang tidak mewakili kelas DNA benar-benar
berbeda.
Pada pola pokok penyebaran pengulangan, fraksi berulang ditemukan terdiri
dari dua jenis families: lokasi sekuen berulang dan penyebaran sekuen berulang.
1. Lokasi Sekuen Berulang
Kebanyakan genom eukariotik mengandun gurutan DNA berulang secara
acak. Dalam beberapa spesies, lokasi pengulangan sekuen DNA dapat
menjelaskan keutamaan DNA dalam genom. Sebagai contoh, pada tikus kanguru,
Dipodomysordii, lebih dari 50% dari genom terdiri dari tiga sekuens berulang:

AAG2, 4x 109kali ;TTAGGG, 2,2 x109 kali, dan ACACAGCGGG, 1,2 x 109kali.
Tentu saja, families ini tidak sepenuhnya homogen tetapi berisi banyak varian
yang berbeda dari urutan konsensus dalam satu atau dua nukleotida. Sebagai
contoh, beberapa urutan dalam family "TTAGGG" sebenarnya TTAGAG.
Bahkan genom yang jauh lebih kecil mungkin berisi sebagian besarurutan
yang sangat berulang. Sebagai contoh, 40% dari genom Drosophilavirilis terdiri
dari tiga urutan yang sangat berulang: ACAAACT1,1x 107kali;ATAAACT, 3,6x
106kali dan ACAAATT, 3,6x 106 kali. Anehnya, 35% genom dari kode
pencetakan uniseluler, Absidiaglauca, yang hanya sembilan kali lebih besar dari
E. coli, tersusun dari DNA berulang.
Banyaknya lokasi sekuen berulang memiliki sebuah komposisi nukleotida
yang seragam yang menunjukkan bahwa, pada saat fraksionalisasi DNA genomik
dan pemisahan dengan gradien kerapatan, mereka membentuk satu atau lebih pita
tebal yang jelas dibedakan dari apusan yang diciptakan oleh fragmen DNA
lainnya dengan banyak komposisi heterogen. Pita yang berukuran jauh lebih berat
atau lebih ringan dari urutan genom lain, yang disebut DNA satelit. Beberapa
DNA satelit mungkin sangat kaya G+ C atau sangat kaya A +T; GC dalam
rentang satelit dari yang terendah 1% pada kepiting Cancergracilis dan C.
antenarius, sampai mencapai 73% pada patogen trypanasomal
Leishmaniainfantum dan nyamuk Chironomusplumosus. Genom mamalia
biasanya terdiri dari DNA satelit 5-30%. Jumlah DNA satelit pada tanaman dapat
mencapai 40% dari genom total.
Dalam beberapa spesies, urutan berulang yang tersusun secara tandem
ditemukan pada semua kromosom, sementara lainnya dibatasi pada lokasi
kromosom tertentu. Sebagai contoh, lebih dari 60% dari genom
Drosophilanasutoides terdiri dari DNA satelit, dan sebagian besar terlokalisasi
pada salah satu dari empat autosom dan kromosom Y (Gambar 2.9), yang
tampaknya mengandung dalam jumlah sedikit (Miklos 1985). Tidak semua lokasi
pengulangan DNA terdiri dari pengulangan pendek. Misalnya, paus pembunuh,
Orcinusorca, mengandung sekitar setengah juta kopi dari sekuen panjang
1.579bp, terhitung sekitar 15% dari genom (Widegren et al.1985).

Gambar 2.9 Sekuens DNAyang sangat repetitif (daerah hitam) yang sebagian besar terlokalisasi paling besar dari tiga autosom dan kromosom Y. (Sumber: Graur dan
Hsiung Li, 1999)
Berdasarkan bukti yang ada pada saat ini, dimungkinkanbahwa lokasi
sekuen berulang adalah tanpafungsi.Selain itu, adalah mungkin bahwa jumlah
lokasi sekuen berulang tidak menurunkan atau meningkatkan ketahanan individu.
Akibatnya, evolusi sekuens tersebut tidak dipengaruhi oleh seleksi alam. Jumlah
dan komposisi ini terulang secara bervariasi melalui mutasi seperti konversigen
dan pindah silang yang tidak merata, dan fiksasi dalam populasi terjadi melalui
hanyutan genetik secara acak. Konversi gen dan pindah silang yang tidak
merataakan menghasilkan dua hasil untuk sekuen ini: (1) urutan homogenitas,dan
(2) jumlah fluktuasi dari waktu ke waktu (Charlesworth et.al,1986). Ini juga telah
menyarankan bahwa tingkat pergantian lokasi sekuen berulang yaitu, susunan
yang ada akan dihapus oleh pindah silang yang tidak merata, sedangkan susunan
baru dapat terus menerus diciptakan oleh proses duplikasi DNA (Walsh,1987).
Usulan bahwa pengulangan sekuen secara tandem pada DNA sampah pada
dasarnya menunjukkan tidak adanya efek fenotipik. Selain itu, diasumsikan
bahwa kehadiran mereka atau tidak dalam jumlah yang bervariasi tidak
mempengaruhi keberadaan operator. Meskipun ini mungkin benar dalam
kebanyakan kasus, ada bukti yang berkaitan dengan serangkaian sekuen berulang
tertentu yang menunjukkan bahwa hal ini tidak selalu terjadi. Responden lokus
(Rsp) dalam populasi alami Drosophila melanogaster terdiri dari 20-2,500 salinan
dari sekuen kaya AT, panjang 120-bp- (Wu et al,1988). Dalam sebuah kompetisi
percobaan yang melibatkan populasi campuran yang terdiri dari lalat dengan 700
salinan pengulangan dan lalat dengan 20 copian, diamati bahwa frekuensi dari

lalat dengan 20 kali menurun seiring waktu (Wu et al,1989). Oleh karena itu,
disimpulkan bahwa lalat dengan 700 kopian memiliki keberadaan lebih tinggi dari
lalat dengan hanya 20 kopian. Kecuali untuk perannya dalam sistem distorsi
segregasi, fungsi lokus Rsp saat ini tidak diketahui, tetapi jelas bukan DNA
sampah, karena ketiadaan mempengaruhi keberadaan organisme. Namun, kami
tidak mengetahui adanya kasus lain di mana sekuen berulang secara tandem
ditunjukkan untuk mempengaruhi ketahanan.
2. Penyebaran Sekuen Berulang
Kelas keduadari pengulangan DNA terdiri dari urutan yang tersebar di
seluruh genom.Salinan dari penyebaran sekuen berulang ditemukan diintron,
mengapit daerah gen/daerah antargen, dan DNA nongenik.
Terdapat dua kategori utama dari penyebaran sekuen berulang: pengulangan
sekuens berupa tandem yang sederhana dan pengulangan yang berseling. Tabel
2.5 menunjukkan klasifikasi pengulangan sekuen berupa tandem yang sederhana
sesuai dengan ukuran dari unit yang berulang, jumlah tiap susunan unit berulang,
dan lokasi genom dari susunan tandem. Perhatikan lokasi sekuen berulang pada
sebagian besar satelit dan mini satellites, meskipun sebagian kecil dari
minisatellites tersebar. Telah diperkirakan bahwa terdapat 300.000 trinucleotide
dan tetra nucleotide pengulangan tandem pendek pada genom manusia atau satu
susunan setiap 10 Kb genom DNA (Beckmann dan Weber,1992). Umumnya
mikrosatelit manusia terdiri dari pengulangan dinukleotida CA.Terdapat sekitar
50.000 salinan mikrosatelit dalam genom manusia yaitu, satu susunan setiap 30
Kb (Hudson et al,1992).
Tabel 2.5 Klasifikasi Pengulangan Sekuen
Genom manusia juga berisi empat kelas utama pengulangan yang
berseling: (1) SINEs, (2) LINEs, (3) seperti retrovirus dan elemen

retrotransposon, dan (4) DNA yang dimediasifosil transposabel. Kelimpahan dan
distribusi relatif genom dari kelas-kelas pengulangan yang berseling ditunjukkan
pada gambar 2.10.
Gambar 2.10 Kelimpahan relatif dan distribusi genom manusia melalui kelas pengulangan yang berseling pada daerah dengan kandungan GC.Distribusi hampir
komplementer dengan pengulangan dari Alu dan LINE1. (Sumber: Graur dan Hsiung Li, 1999)
Genom manusia mengandung dua families, LINE1 (LI) dan LINE2 (L2).
Terdapat sekitar 600.000 pengulangan LI dalam genom manusia, atau sekitar 15%
dari genom. Family LI telah aktif dalam genom mamalia sebelum terjadinya
perbedaan antara marsupial dan placentals. Asal dari family L2 jauh lebih kecil
(~271.000 pengulangan) yang mungkin sangat kuno, kemungkinan besar
terjadinya perbedaan amfibi dari vertebrata amniote. Sekitar 95% dari semua
urutan LI tersebut dipotong di ujung 5 dan tidak ditranskripsi atau retrotransposed.
Tingkat perbedaan urutan LI antara spesies jauh lebih besar dari pada derajat
perbedaan antara salinan LI yang sejenis. Sebagai contoh, urutan LI dari tikus dan
manusia rata-rata berbeda satu sama lain sekitar 30%, dibandingkan dengan
perbedaan sebuah sekuen dari sekitar 4% dalam tikus (Hutchison et al,1989).
Elemen L1 yang rusak berkembang jauh lebih cepat dari pada elemen yang
masih utuh. Selain itu, garis keturunan evolusi dari sekuen L1 yang rusak tidak
mengandung cabang, yang menunjukkan bahwa elemen-elemen tidak mampu
melakukan replikasi transposisi. Kemudian berbentuk pseudogen dari retroposons,
di mana kendala fungsional tidak lagi beroperasi, dan dengan demikian mengikuti

asimilasi komposional dan lamanya pembatasan sampai mereka tidak lagi dikenal
sebagai LINEs. Faktanya bahwa sebagian sekuen L1 yang rusak menyiratkan
bahwa penyebaran elemen L1 dalam genom tergantung pada sejumlah kecil
elemen sumber. Akibatnya, elemen L1 dalam genom sangat homogen dan tingkat
pergantian sekuens sangat tinggi. Memang, pada hewan pengerat telah
diperkirakan bahwa lebih dari setengah dari elemen L1 hanya 3 juta tahun atau
bahkan lebih muda.
Genom manusia juga mengandung dua families SINE, 7SL yang diturunkan
family Alu, dengan sekitar 1.100.000 salinan atau 10% dari genom, dan tRNA
yang diturunkan family MIR, dengan sekitar 400.000 salinan. Pada daftar
pengulangan berseling yang lengkap dalam genom manusia juga harus disebutkan
elemen-retrovirus dan retrotransposon (~5% dari genome), sisa-sisa elemen DNA
transposabel (~2%), dan sekitar 60.000 salinan tidak terklasifikasi dari
pengulangan berseling (~1%). Kesimpulannya, lebih dari sepertiga dari genom
manusia berasal dari mobile elements dari beberapa families. Keutamaan
pengulangan sekuen berseling tersebut, tidak lagi memiliki kemampuan untuk
berpindah.
3. Urutan yang Berulang: Penyebab Variasi dalam Ukuran Genom
Seperti disebutkan sebelumnya, komponen utama dari paradoks nilai C
adalah kenyataan bahwa organisme yang secara morfologis dan anatomis mirip
menunjukkan nilai C yang sangat berbeda. Ini lebih jelas dari pada dalam
perbandingan antara spesies yang termasuk dalam genus yang sama. Perbedaan
dalam ukuran genom dapat dijelaskan oleh perbedaan dalam pecahan berulang.
Dari hewan pengerat seperti Ctenomys (tuco-tucos), untuk tanaman seperti Avena
(gandum), dan Hylobates (gibbon) sampai Drosophila, setiap spesies congeneric
berbeda satu sama lain nilai C-nya, perbedaan dapat sepenuhnya dijelaskan oleh
pengulangan fraksinongenic dari genom, sering pula dengan perbedaan dalam
jumlah pengulangan tandem sederhana. Selain itu, setiap kali takson ditemukan di
mana ukuran genom jauh lebih kecil dari taksa yang terkait, kami selalu
menemukan bahwa perbedaan adalah sepenuhnya karena sekuens berulang.
Sebagai contoh, beberapa kelelawar memiliki genom yang sekitar 50% ukuran
mamalia eutherian lainnya. Perbedaan tersebut disebabkan oleh kurangnya

mikrosatelit AT dan GC, yang pada mamalia lain tersedia cukup. Demikian juga,
kurangnya variasi ukuran genom relatif pada burung (Tabel 8.3) dapat disebabkan
kelangkaan mikrosatelit pada genom burung.
I. Mekanisme Untuk Meningkatkan Daerah Dalam Ukuran Genom
Peningkatan regional dalam ukuran genom dapat dijelaskan dengan
beberapa mekanisme. Duplikasi transposition adalah salah satu mekanisme yang
telah diketahui yang bisa menghasilkan sekuens berulang yang terpisah.
Mekanisme lainnya menghasilkan lokasi sekuen berulang. Telah disarankan
bahwa seluruh pengulangan fraksi DNA pertengahan pada eukariotik berasal dari
elemen transposable. Sebagian besar elemen tidak lagi bisa berpindah karena telah
mengalami kerusakan akibat mutasi atau insersi pada elemen yang lain.
Peristiwa pindah silang yang tidak merata kemungkinan merupakan
mekanisme yang bertanggungjawab terhadap peningkatan dan jumlah salinan dari
satelit dan minisatelit. Meskipun demikian, fakta peristiwa pindah silang yang
tidak merata ini biasanya menghasilkan sekuens yang terdiri dari pengulangan
panjang. Dilain pihak, beberapa lokasi sekuen berulang seperti mikrosatelit dan
pengulangan tandem yang pendek.
Ditemukan adanya bukti bahwa jumlah salinan pada lokus minisatelit bisa
mengalamai peningkatan dengan cepat. Contohnya pada manusia, sebuah lokus
MS32 terdiri dari 600 pengulangan. Sedangkan pada monyet purba, lokus
homolog terdiri dari 3-4 pengulangan. Karakter terakhir agaknya mewakili
keadaan nenek moyang dan jumlah ulangan yang tinggi pada manusia mewakili
keadaan sekarang. Amplifikasi DNA mengacu pada setiap mekanisme yang
meningkatkan jumlah salinan gen atau sekuen DNA untuk tingkat karakteristik
organisme. Khususnya, amplifikasi DNA yang mengacu pada peristiwa yang
terjadi dalam kehidupan suatu organisme dan menyebabkan peningkatan secara
tiba-tiba dalam jumlah salinan dari sekuen DNA. Dalam hal ini dibedakan
menjadi 2 amplifikasi, yaitu amplifikasi vertikal dan amplifikasi horisontal.
Amplifikasi vertikal mengacu pada proses yang melalui pelipat gandaan sekuen
tertentu di luar kromosom. Amplifikasi horisontal mengacu pada proses

penciptaan beberapa salinan dari sekuen DNA tertentu dan penggabungannya
dalam genom yang diwariskan dari organisme.
Salah satu metode yang dapat menjelaskan mekanisme amplifikasi ialah
model rolling circle dari replikasi DNA (Gambar 2.11). Tipe replikasi ini
digunakan dalam amplifikasi gen rRNA pada oosit Amphibi. Dalam hal ini,
amplifikasi melibatkan pembentukan salinan extrachromosomal sirkuler sekuen
DNA, yang kemudian dapat menghasilkan banyak unitextrachromosomal
tambahan yang mengandung pengulangan tandem dari urutan asli. Jika unit
tersebut menjadi terintegrasi kembali ke dalam kromosom, akan ada tambahan
genom yang terdiri dari urutan berulang yang identik.
Gambar 2.11. Model Rolling Circle dari Amplifikasi Gen pada Oosit Amphibi. rRNA kromosomal disusun dalam susunan tandem yang berisi bagian transkripsi (hitam) dan daerah nontranskripsi (putih). Amplifikasi melibatkan pembentukan salinan ekstrakromosom sirkuler yang berisi jumlah variabel pengulangan, yang
kemudian diamplifikasi melalui beberapa putaran dari replikasi rolling circle. Keperiodikan akan berubah mengikuti amplifikasi rolling circle. (Sumber: Graur
dan Hsiung Li, 1999)

J. Distribusi Gen
Kita hanya berhubungan dengan porsi DNA yang mungkin atau mungkin
juga tidak memiliki fungsi, tetapi jika tetap bekerja, fungsi tersebut pasti bukan
protein- pengkode satu.Dimana protein yang mengkode gen? Kita akan membahas
5 isu yang berhubungan: (1) Jumlah gen, (2) lokasi gen genom, (3) Kepadatan
gen, (4) variabilitas jumlah kromosom, dan (5) Proses evolusi mempengaruhi
urutan gen.
1. Berapa banyak gen yang ada, dimana letaknya dan apakah kita
membutuhkannya?
Tiga organisme eukariotik yang digunakan untuk menjawab pertanyaan
tersebut :Yeast roti, Saccharomyces ceerevisiae, dan nematoda Caenorhabditis
elegans yang seluruh genomnya telah disekuensing.bagaimanapun organisme ini
tidak mewakili keseluruhan organisme eukariotik semenjak genom mereka dipilih
untuk disekuensing karena ukurannya kecil. Saccharomyces cerevisiae memiliki
lebih dari 6000 gen-gen pengkode protein yang menyebar merata diantara 16
kromosom, contohnya jumlah gen-gen pada tiap kromosom sebanding dengan
panjangnya (Gambar 2.12).
Gambar 2.12 Hubungan antara jumlah gen dan panjang kromosom pada Saccharomyces cerevisiae. Sedikit variasi disekitar garis regresi mengindikasikan bahwa gen didistribusikan merata
diantara 16 kromosom. (Sumber: Graur dan Hsiung Li, 1999)
Disisi lain tidak terjadi distribusi gen sepanjang kromosom. Terdapat daerah
dengan densitas gen yang tinggi dan rendah (Gambar 2.13). Pada Caenorhabditis
elegans terdapat lebih dari 19.000 gen terdistribusi antara 6 kromosom dengan

panjang total sekitar 97 Mb. Distribusi kromosom kurang seragam daripada yeast,
dengan kromosom X memiliki kepadatan gen terendah daripada kromosom yang
lain.
Gambar 2.13 Periodisitas kerapatan gen sepanjang kromosom 11 pada Saccharomyces cerevisiae.
(Sumber: Graur dan Hsiung Li, 1999)
Pengetahuan mengenai genom organisme multiseluler sangatlah
terbatas.Bagaimanapun juga sebagian besar genom belum tentu berisi informasi
pengkode protein.Jika kita mengurangi dari panjang semua genome sekuen
berulang, semua pseudogen, semua intron dan semua daerah intergenik sangat
sedikit yang tersisa. Pada manusia, eksperimen hibridisasi RNA-DNA masa
lampau telah menunjukkan bahwa banyak gen yang bukan pengkode protein di
dalam fraksi berulang dari genom dan bahkan di dalam fraksi DNA yang unik
hanya sekitar 3% dari DNA yang ditranskripsikan. Dengan menggunakan data
pemetaan transkripsi, Gardiner memperkirakan bahwa kurang dari 10% genom
manusia merupakan gen.
Distribusi gen pengkode protein diantara kromosom manusia sangat tidak
merata. Beberapa kromosom seperti kromosom 1, 19 dan 20 diprediksi sangat
kaya akan gen, lainnya kromosom 4 dan 18 mungkin tersusun banyak sparser
informasi genetik. Contohnya Kromosom 19 yang kaya akan gen diperkirakan
mengandung 2000 gen di dalam daerah eukromatik sekitar 60 juta pasang basa.
Kepadatan gen mencapai 0,03 gen/Kb. Kita mencatat bahwa nilai ini terlalu
tinggi bahkan untuk kromosom 19, apalagi untuk kromosom yang lain. Terdapat 3

alasan utama untuk statemen tersebut yaitu: (1) hanya derah eukromatin yang
telah diperhitungkan, (2) beberapa gen mungkin kenyataannya berupa pseudogen,
dan (3) seperti dikatakan sebelumnya, kromosom 19 merupakan kromosom
dengan kepadatan gen yang tinggi.
Kepadatan gen dan pemanjangan fraksi gen, terlihat berkorelasi negatif
dengan ukuran genom (Gambar 2.14). Pada Mycoplasma genitalium 0.8 gen/Kb.
Kerapatan menurun menjadi 0.6 gen/Kb pada E. coli, yang memiliki genom 8 kali
lebih besar. Pada Eukariota, kerapatan mendekati 0.5 gen/Kb pada yeast dan 0.2
gen/Kb pada Caenorhabditis yang memiliki genom 8 kali lebih besar. Perkiraan
kita kerapatan gen pada organsime lain adalah kurang pasti, tetapi jelas memiliki
kecenderungan yang sama. Contoh kerapatan gen pada Arabidopsis thaliana
adalah 0.2 gen/Kb dalam daerah kaya gen pada kromosom 1, tetapi hanya 0.03
gen/Kb pada eukromatin pada sebagian besar kromosom kaya gen pada manusia.
Tetapi nilai terakhir tidak cukup baik untuk membandingkan dengan perkiraan
kepadatan Alu pada kromosom yang sama (1,1 elemen/Kb).
Gambar 2.14 Hubungan antara fraksi gen dan ukuran genom. (Sumber: Graur dan Hsiung Li,
1999)
Pada umumnya genom tumbuhan seperti beras, jagung, dan (gandum)
barley sebagian besar protein pengkode gen dikelompokkan dalam segmen DNA

yang panjang (secara kolektif disebut gen space) yang mewakili sebagian kecil
(12-24%) dari genom inti, dipisahkan oleh hamparan luas daerah kosong-gen.
K. Evolusi Kromosomal
Tidak ada ukuran umum mengenai kompleksitas ukuran biologi.dua
kemungkinan kandidat adalah sejumlah preotein kode-gen dan “kekayaan dan
variasi morfologi serta tingkah laku”. Tidak ada alasan untuk menduga bahwa
sejumlah gen meningkat sejalan dengan waktu evolusinya. Bagaimanapun bukti
empirik mengindikasikan bahwa pada beberapa garis keturunan/silsilah
kompleksitas gen meningkat secara pesat. Ini menunjukkan bahwa jumlah gen
tidak meningkat terus-menerus selama evolusi, tetapi meningkat dalam tahap-
tahap yang berbeda. Tahap terbesar terjadi pada saat transisi dari prokariot
menjadi eukariot dan saat transisi dari invertebrata menjadi vertebrata.Tahap
pertama dianggap telah difasilitasi oleh penemuan dari nukleosom, mengingat
pada tahap kedua dengan penyebaran metilasi gen sebagai mekanisme untuk
mengontrol ekspresi gen pada seluruh genom.
Akhir-akhir ini perkiraan yang dapat dipercaya mengenai jumlah gen
berdasarkan sampling sekuens/urutan besar dikumpulkan. Data ini
mengindikasikan bahwa sejumlah gen memang meningkat dan terjadi pada tahap
quantum. Pada faktanya terdapat suatu kejadian yang dimungkinkan secara tepat
merupakan waktu dimana tahap quantum terjadi. Pada hewan terjadi “lompatan ke
depan yang besar” pada jumlah gen yang kadang-kadang terjadi pada masa
silurian sebelum terjadinya perbedaan vertebrata tetapi setelah perbedaan kordata
(invertebrata) (Gambar 2.15).

Gambar 2.15 Perkiraan jumlah gen pada spesies bakteri, fungi, invertebrata, dan vertebrata. (Sumber: Graur dan Hsiung Li, 1999)
Meskipun kenyataannya sitogenetik merupakan sebuah disiplin ilmu yang
lebih tua dari pada bio molekuler. Kita tahu sangat sedikit tentang evolusi
kromosom melebihi deskriptif fenomenologi. Bagaimanapun dengan kedatangan
era genomik, kita memulai untuk mengumpulkan beberapa wawasan ke dalam
isu-isu seperti evolusi sejumlah kromosom dan dinamika dari penataan kembali
urutan gen.
1. Kromosom, Plasmid dan Episom
Organisme dan organel tersusun atas dua jenis materi genetik: kromosom
dan elemen ekstrakromosom. DNA kromosomal berisi gen-gen yang sedikitnya
beberapa darinya tidak penting. Elemen ekstrakromosomal disisi lain berisi
informasi genetik meskipun mungkin memiliki efek fenotip yang penting, tidak
dibutuhkan pada semua kondisi. Antara efek 2 fenotipik yang diketahui paling
baik dari elemen ekstrakromosomal adalah : 1. Antibiotik, logam berat dan tahan
panas, 2. Virulensi dan patogenitas, 3 . autotrofi,dan 4. Kelenturan antigen.
Kelas utama dari elemen2 ekstrakromosomal adalah pllasmid dan episom.
Plasmid adalah molekul ekstrakromosomal yang secara sendiri bereplikasi dari
genom ekstrakromosomal. Plasmid ada semata-mata dalam sebuah tempat sendiri
dan bereplikasi secara independen dari kromosom.Rata-rata replikasinya mungkin
lebih tinggi dari DNA kromosom.
Genom plasmid dapat berupa sirkuler atau linier, dan ukuran yang berbeda-
beda dari 1000 nukleotida yang disebut kriptik plasmid hingga 400 kb (giant
plasmid). Episom berisi hanya informasi genetik non esensial, tetapi mereka
mampu berada diantara dua tempat: secara independen bereplikasi dalam sebuah
sel atau terintegrasi dalam sebuah kromosom. Contoh episom : profage virus.
2. Evolusi sejumlah kromosom pada Prokariot
Sebagian besar bakteri tersusun atas kromosom tunggal.Genus Brucella
merupakan kelompok bakteri gram negatif yang bersifat patoggen terhadap
manusia dan hewan yang salah satu spesiesnya kita temukan mengandung
kromosom tunggal atau ganda. Contohnya B. Melitensis yang patogen pada biri-
biri/domba dan kambing dan memiliki kromosom sirkular berukuran 2,100 Kb

dan 1,150 Kb. Pada spesies Brucella yang lain yang memiliki dua kromosom,
ukurannya mungkin berbeda misalnya 1,850 Kb dan 1,350 Kb pada B. Suis.
Sangat menarik bahwa ukuran kromosom tunggal pada rantai kromosom hampir
sama seperti ukuran total kromosom untuk dua rantai kromosom.
Kondisi yang hampir sama juga ditemukan pada anggota lain dari subdivisi
α proteobakteria. Rhodobacter sphaeroides, bakteri fotosintetik fakultatif
ditemukan memiliki dua kromosom sirkulaar yang sesungguhnya (3,000 dan
900Kb) yang masing-masing mengandung gen yang penting untuk fungsi
metabolik.
Sangat menarik, genom dari Methanococcus jannaschii, archaeon pertama
yang diurutkan secara lengkap ditemukan tersusun atas tiga elemen fisik yang
berbeda: (1) Kromosom sirkular besar sekitar 1,700 Kb berisi 1,700 gen pengkode
protein, (2) elemen besar 60 Kb, berisi 43 daerah yang diprediksi pengkode
protein, dan (3) elemen kecil sekitar 17 nKb dengan kapasitas pengkode 12
protein. Kita tidak mengetahui apakah dua elemen terakhir tersebut merupakan
kromosomal atau ekstrakromosomal.
Dengan menggunakan protokol laboratorium yang rumit, Itaya dan Tanaka
berehasil membagi kromosom bakteri menjadi dua subgenom replikasi
independen. Penemuan ini mengindikasikan bahwa evolusi sejumlah kromosom
pada bakteri mungkin dipotong oleh adanya mutasi daripada seleksi terhadap
multikromosom.
3. Variasi Sejumlah Kromosom pada Eukariotik
Pada serangga, n bervariasi antara 1 (semut Australia_Myrmecia pilosula)
hingga hampir 250 pada kupu-kupu Lysandra atlantica.Pada tumbuhan, dalam
famili tunggal (Asteraceae) kita menemukan bahwa n bervariasi dari 2 pada
Haplopappus gracilis hingga kira-kira 90 pada Senecio robertii-friesii. Pada
mamalia yang memiliki jarak ukuran genom yang sangat sempit (tabel 8.3),
nbervariasi dari 5 pada Ctenomys steinbachi hingga 102 pada Tympanoctomys
barrerae. Anehnya jumlah kromosom tidak semuanya berkorelasi pada konten
DNA.
L. Mekanisme Perubahan Urutan dan Distribusi Gen diantara Kromosom
Berdasar posisi seentromer, kromosom eukariotik dibedakan menjadi 3
jenis: Telosentrik, Akrosentrik dan Metasentrik (Gambar 2.16).
Gambar 2.16 Klasifikasi kromosom eukariotik berdasarkan posisi sentromer (lingkaran). (Sumber: Graur dan Hsiung Li, 1999)
Banyak proses yang menyebabkan perubahan urutan gen (Gambar 2.17).
Pembalikan kromosom termasuk rotasi segmen 180o, dengan hasil bahwa urutan
gen untuk segmen merupakan kebalikan dengan mematuhi urutan aslinya.
Terdapat dua jenis inversi/pembalikan: Perisentrik dan parasentrik. Pada awalnya,
segmen mengalami pembalikan termasuk sentromer. Delesi kromosom mungkin
terjadi secara teminal atau interstisial.Sebagai alternatifnya, bagian kromosom
mungkin di duplikasi. Suatu proses yan dahulu kita sebut sebagai bagian polisomi.
Kromosom mungkin juga dieliminasi, suatu proses yang hanya terjadi pada sel
somatik arthropoda. Pada akhirnya kromosom berpisah menjadi dua dan tiap
kromosom hasil pemisahan memiliki kehidupan independen. Proses ini hanya
dapat terjadi jika kromosom berdifusi dengan sentromer, contohnya jika selama
mitosis dan meiosis,benang spindel menarik dan mengaitkan pada beberapa situs
sepanjang ukuran kromosom.
Gambar 2.17 Sebuah kromosom dengan 5 gen yan mungkin menyebabkan beberapa proses penataan ulang gen. (Sumber: Graur dan Hsiung Li, 1999)

Kromosom yang berbeda mungkin mengubah informasi genetik melalui
proses yang demikian seperti translokasi resiprok dan non resiprokal, dan fusi
sentrik (Gambar 2.18). berdasarkan tipe pembalikan kromosom dalam prosesnya,
fusi sentrik mungkin atau juga mungkin tidak disertai hilangnya gen. Reduksi
sejumlah kromosom oleh fusi terlihat seperti kejadian evolusi berulang.
Gambar 2.18 Contoh perubahan informasi genetik antara 2 kromosom nonhomolog. (Sumber: Graur dan Hsiung Li, 1999)
1. Penghitungan kejadian penataan Ulang Urutan Gen
Untuk mempelajari evolusi panataan ulang urutan gen kita harus mampu
memperkirakan sejumlah peristiwa seperti inversi, transposisi, dan delesi yang
penting dalam merubah urutan gen pada genom hingga yang lainnya. Metode
sederhana yang disebut Metode Reduksi Penjajaran, kita melakukian
penghitungan sehingga disebut Jarak Pemeriksaan Evolusi (dilambangkan E)
antara dua genom, A dan B. E memiliki dua komponen: Jarak Delesi (D) dimana
nomor terendah delesi atau insersi diperlukan oleh genom A dan B untuk
memiliki set gen yang serupa, sekalipun pada urutan yang berbeda, dan Jarak
Penataan ulang (R), contohnya sejumlah kecil inversi dan transposisi diperlukan
untuk merubah urutan gen A menjadi urutan gen B.
E = D + R
Untuk perkiraan E, kita menggunakan tiga prosedur sederhana geometrik:
Delesi, Bundling (ikat) dan inversi (Gambar 2.19). pertama kita hubungkan gen
yang homolog dengan suatu garis. Pada tahap ini kita membedakan antara
pasangan homolog yang memiliki persamaan orientasi dan relatif diinversikan

pada satu dan lainnya. Prosedur Delesi: semua gen yang tidak muncul pada salah
satu dari dua genom dihilangkan. Dengan demikian, D sama dengan jumlah
segmen yang dipindahkan. Pada kasus ini kita menghilangkan 5 gen tetapi hanya
2 segmen, sehingga D = 2.
Gambar 2.19 Tiga prosedur geometrik dasar yang menyertakan metode reduksi penjajaran untuk menyimpulkan sejumlah penataan ulang urutan gen antara dua
genom. (Sumber: Graur dan Hsiung Li, 1999)
2. Penataan Ulang Urutan Gen pada Bakteri
Karena bakteri sungguh dibatasi dalam rekombinasi dan karena banyak gen
mereka berfungsi sebagai unit (operon), evolusi bakteri dikarakterisasikan oleh
stabilitas spasial dan temporal dari urutan gen. Dengan penyelesaian sekuensing
genom bakteri pertama, bagaimanapun ini menjadi jelas bahwa urutan gen bakteri
adalah bebas/apa saja tetapi dikonservasi. Untuk menggambarkan kekurangan
dari konservasi dibandingkan urutan gen antara haemphilus influenzae dan
Mycoplasma genitalium. 184 gen ortolog antara 2 spesies.
3. Penataan Ulang Urutan Gen pada Eukariotik
Mempelajari penataan ulang urutan gen pada eukariotik lebih lengkap dan
tidak terbatas dari pada bakteri untuk beberapa alasan. Pertama, genom eukariotik
mengandung banyak gen berulang, dan memutuskan apakah dua gen dari dua
organisme tersebut merupakan ortolog atau paralog sungguh lengkap. Kedua,
penataan ulang urutan gen termasuk perpindahan dan perubahan informasi antara
kromosom sangat baik seperti di dalam kromosom. Ketiga, urutan gen terlihat
sangat tidak stabil. Contohnya: gen dari kromosom 1 manusia ditemukan pada 9
kromosom yang berbeda pada tikus. Eukariot memiliki sejumlah besar gen dari
pada prokariot.
Perbandingan antara multikromosom organisme, sangat tepat dengan
mendefinisikan beberapa istilah yang mungkin membantu kita memecahkan

masalah perpindahan gen diantara kromosom. Synteny merupakan kejadian dua
atau lebih gen dalam kromosom yang sama. Conserved Synteny merupakan dua
atau lebihgen homolog yang disintesis pada dua spesies.Conserved Linkaged
menyinggung pada konservasi keduanya, baik synteny dan urutan gen dalam gen
homolog antar spesies.Disrupted Synteny merupakan kasus dimana dua gen
ditempatkan dalam kromosom yang sama dalam satu spseies tetapi ortolog
mereka ditempatkan pada kromosom yang berbeda pada spesies kedua. Jadi jelas
bahwa disrupted synteny juga akan dihitung sebagai disrupted linkage.
4. Urutan Gen sebagai Karakter Filogenetik
Ahli biologi evolusioner menggambarkan peningkatan keistimewaan
struktural dari genom seperti urutan gen sebagai penanda filogenetik. Tetapi,
peningkatan nilai penataan ulang urutan gen dan nilai variasi antara keturunan
evolusi merupakan berita buruk untuk rekonstruksi filogenetik berdasarkan urutan
gen. Contohnya dalam mempelajarai urutan gen mitokondria pada 137 spesies
burung mennunjukkan bahwa evolusi paralel dalam urutan gen merupakan
kejadian yang biasa dan urutan gen yang sama mungkin muncul secara
independen. Karena ketidak sempurnaan properti filogenetik dan sulitnya dalam
merekonstruksi urutan pada kejadian awal penataan ulang urutan gen, pohon
filogeni didasarkan pada seringnya urutan gen mengadung kekeliruan yang jelas.
M. Konten GC pada Bakteria
Antara genom eubakteria (termasuk genom organel), memiliki persentase
guanin dan sitosin/konten GC yang bervariasi antara 25% hingga 75%.Pada
banyak kasus, konten GC terlihat berhubungan dengan filogeni, dengan hubungan
dengan bakteri memiliki konten GC yang serupa (Gambar 2.20).

Gambar 2.20. Konten GC pada beberapa genom eubakteria. (Sumber: Graur dan
Hsiung Li, 1999)
Terdapat dua jenis hipotesis yang penting untuk menjelaskan variasi konten
GC pada bakteri.Gambaran seleksi memandang konten GC sebagai bentuk
adaptasi dengan kondisi lingkungan.Contohnya , pasangan G:C lebih stabil dari
pada pasangan A:T karena penambahan ikatan hidrogen, oleh karena itu mungkin
hubungan antara konten GC dan suhu pada bakteri yang mengarahkan. Studi awal
rupanya untuk mengindikasi bahwa pada bakteri termofilik yan mendiami relung
sangat panas, terdapat pemakaian yang istimewa dari asam amino yang dikode
oleh kodon yang kaya akan GC (misalnya: alanin dan arginin) dan tidak
menggunakan asam amino yang dikode oleh kodon yang kekurangan GC seperti
serin dan lisin. Pada studi yang ekstensive tidak ditemukan hubungan antara
konten GC dengan pertumbuhan temperatur yang optimal.
Skenario seleksi yang lain dengan menggunakan radiasi UV sebagai
tenaga selektif. Sejak dimer T-T sensitif terhadap radiasi, mikroorganisme pada
lapisan permukaan atas tanah, yang mengarahkan pada cahaya matahari,

seharusnya memiliki konten GC yan kebih tinggi daripada bakteri yang tidak
diarahkan, misalnya bakteri intestinal.
Gambaran mutasi melibatkan dugaan pola mutasi untuk menjelaskan
variasi konten GC.Berdasarkan gambaran ini, konten GC spesies bakteri
menunjukkan keseimbangan antara (1) rata-rata substitusi dari G/C menuju T/A,
ditunjukkan sebagai u; dan (2) rata-rata substitusi dari A/T menuju G/C,
ditunjukkan sebagai v.
N. Chirochores
Perbedaan dalam arah untai leading dan untai lagging dari DNA yang
direplikasi bisa dihasilkan dalam untai dengan pola mutasi bebas.Dalam ketiadaan
dari beberapa seleksi kecenderungan diantara dua untai, perbedaan dalam pola
dari kesalahan replikasi mungkin awal perbedaan dari pola substitusi. Selama
waktu evolusi, perbedaan mungkin terakumulasi ke frekuensi kesetimbangan
dalam masing-masing untai yang berbeda dari frekuensi harapan dibawah kondisi
kecenderungannya, misalnya f A=f T dan f C=f G (Bab 4). Standart deviasi dari
kecepatan mutasi yang sama antara dua untai diukur dengan menggunakan
sebuah variabel yang disebut skew, Sx=y, yang merupakan sebuah ukuran dari
ketidak samaan antara frekuensi dari nukleotida X dan Y dalam sebuah untai.
Perhitungannya adalah:
Sx=y=fx−fyfx+ fy
Lobry (1996c) memeriksa tiga genom bakteri dan menemukan penyimpangan
yang sangat jelas dari f C= f G. Hal ini tidak mengejutkan. Hal yang mengejutkan
adalah distribusi ruang dari skew. Penyimpangan SC=G secara tiba-tiba berubah
arah dari asal dan ujung dari replikasi.
Perubahan dalam nilai SC=G pada awal dari replikasi membuktikan sebuah
kejituan yang umum dalam bakteri yang mana mereka telah digunakan dengan
sukses untuk mengidentifikasi asal dari replikasi dalam spesies yang kurang dari
persetujuan sekuen yang dijaga ketepatan identifikasinya sebelumnya.Genom
bakteri dibagi menjadi 2 segmen dengan sebuah celah simetri dalam sebuah
bagian rangkaian dari nukleotida komplemen, sebagian besar C dan G, di antara
dua untai.Dalam kesamaan dengan isochores yang disebutchirochores.Dua
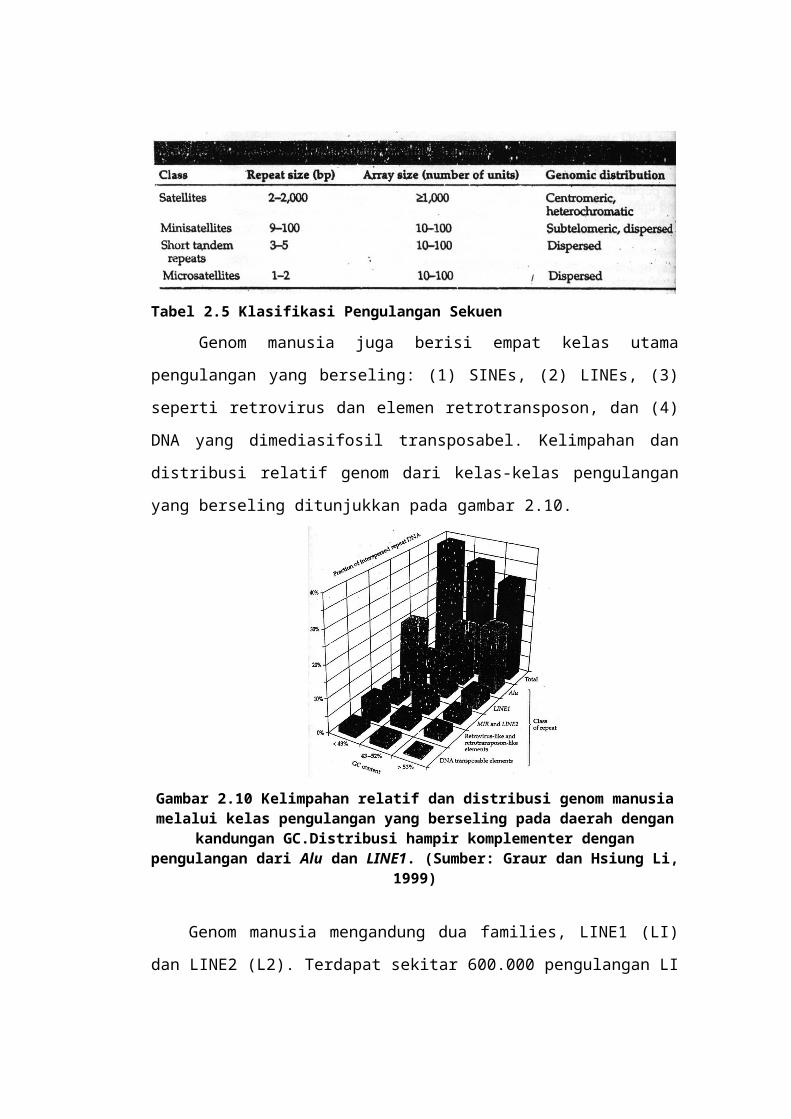
kemungkinan penjelasan-penjelasan untuk menjelaskan keberadaan dari
chirochores.
Hipotesis seleksionis dari Forsdyke yang menyatakan bahwa ada tekanan
selektif global pada genom yang serupa dari oligonukleotid-oligonukleotid
komplemen yang dekat yang mungkin berasal dari struktur sekunder, seperti
‘jepit’ untai ganda, untuk menjaga genom melawan temperatur yang dapat
merusak. Bagaimanapun, dalam sebuah studi dar Gltier dan Lobry menemukan
tidak ada bukti untuk tekanan selektif yang mungkin mempengaruhi struktur
chorichores.Penjelasan yang paling masuk akal sepertinya ada kecenderungan
mutasi.Ada dua bukti untuk pernyataan ini. Pertama, batasan antara chirochores-
chirochores bertepatan dengan asal atau akhir dari replikasi, dan hal ini memberi
kesan sebuah hubungan dengan proses dari replikasi dan perbaikan DNA. Yang
kedua, kecenderungan relatif dari S =G = 0 yang lebih besar dari daerah antara dan
posisi kodon ketiga kemudian untuk posisi pertama dan kedua kodon,
sebagaimana diharapkan jika sebuah perkiraan adalah mutational.
O. PERISTIWA EVOLUSI PADA KODE GENETIK
1. Distribusi Gen dan Eklemen Genetik lainnya diantara Isokor
Posisi isokor manusia dan vertebrata lain pada banyak genom ditentukan
banyak metode. Hampir 30% semua gen manusia merupakan komponen berat
(H3) yg hanya mewakili 3-5% genom. Sangat jarang gen panjang pada isokor
yang kaya GC. Perbedaan daerah gen yang kaya dan miskin GC hampir
seluruhnya dijelaskan oleh intron, yang rata 3x lebih panjang di daerah miskin
GC.Banyak kasus, terjadi penempelan gen pada fragmen DNA yang memiliki
konten GC mirip dengan gen itu sendiri.Melalui degradasi acak fragmen membuat
preparasi DNA, jarak sempit dari komposisi fragmen pembawa gen menunjukkan
bahwa komposisi basa nitrogennya homogen seperti fragmen mereka sendiri.
Observasi ini untuk isolasi gen dan pengelompokan gen, mengindikasikan bahwa
isokor itu berukuran besar jika dibandingkan dengan klaster gen yg diperiksa,
beberapa berukuran 40Kb atau lebih.Ini mnunjukkan bahwa isokor lebih besar
dari 300 Kb. Kadang gen ditemukan pada fragmen yang menutupi jarak level GC
yang lebar. Ini bisa terjadi jika gen pemeriksa berada dekat dengan batas antara

dua isokor, sehingga kerusakan acak menghasilkan fragmen pembawa gen dengan
komposisi berbeda.
Korelasi positif antara level GC pada gen, ekson dan intron dan level GC
pada daerah DNA yang besar dimana mereka menempel. Berlawanan dengan
klaster α dan β globin pada manusia berisi GC rendah.β dan gen globin seperti β
mengandung GC rendah dan menempel pada daerah yang miskin GC. Sedangkan
αdan gen globin seperti α, pada sisi yang lain merupakan dearah kaya GC dan
menempel pada daerah tersebut. Konten GC yangberada pada derah pengkode
memelihara lebih tinggi daripada yang berada di daerah sisi (gb. 8.32).level GC
pada posisi kodon ketiga lebih merata daripada yang berada pada daerah intron,
yang berbelok lebih tinggi daripada yang berada di daerah sisi ujung 5 dan 3.
2. Asal Dari Isokor
Asal dari isokor yang kaya GC masih sebagai misteri atau kontroversi.
Cenderung merupakan segmen DNA panjang (300Kb atau lebih) salah satu dari
GC yang kaya /GC miskin, tidak ditempatkan pada variasi GC, seperti yang
diperiksa diantara daerah variasi gen. Bernardi dkk, mengusulkan bahwa isokor
muncul sebagai keuntungan yang fungsional. Ini telah diklaim bahwa pada darah
cacing, sebuah peningkatan konten GC dapat melindungi DNA, RNA, dan protein
dari kerusakan pemanasan.In imerupakan hipotesis seleksionis.
Wolfe dkk, mengusulkan bahwa isokor muncul dari dugaan mutasi yang
berkaitan dengan perubahan komposisi pada “pool” prekursor nukleotida selama
replikasi DNA germline. Isokor yang kaya GC dibawa pada replicon yang
mereplikasi pada awal siklus sel germline, selama prekursor “pool” memiliki
konten GC yang tinggi, dengan demikian memiliki kecenderungan untuk mutasi
menjadi GC. Isokor yang kaya AT, disisi lain direplikasi lebih lambat dalam
siklus sel, ketika prekursor nukleotida memiliki konten AT yang tinggi. Ini
disebut hipotesis Mutasi.

BAB III
PENUTUP
A. Kesimpulan
1. Variasi ukuran genom di antara organisme sangat bervariasi. Variasi ini dapat
dilihat dari nilai C yang menentukan ukuran genom tiap-tiap organisme.
2. Keteraturan gen dan dinamikan perubahan evolusi dalam hubungannya
dengan keteraturan gen melibatkan berbegai hal, yaitu jumlah gen, lokasi gen
genom, kepadatan gen, variabilitas jumlah kromosom, dan bagaimana
mekansime evolusi yang mempengaruhi urutan gen.
3. Kelebihan DNA dipertahankan oleh seleksi, tetapi komposisi nukleotida
dapat berubah secara acak. Banyak fungsi nukleotipik tambahan telah
dikaitkan dengan fraksi non-genik, tapi semua hipotesis nukleotipik memiliki
satu kesamaan; mereka semua menganggap genom sebagai unit struktural
dari arsitektur nukleus- sebuah blok bangunan yang terbuat dari asam nukleat,
bukan sekedar pembawa informasi genetik.
4. Peristiwa evolusi pada kode genetik melibatkan distribusi gen dan elemen
genetik lainnya diantara isokor. Posisi isokor manusia dan vertebrata lain
pada banyak genom ditentukan banyak metode. Hampir 30% semua gen
manusia merupakan komponen berat (H3) yg hanya mewakili 3-5% genom.




















