







Bahasa
Halaman
Hukum


Journal of Experimental Botanydoi:10.1093/jxb/eru371This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details)
ReseaRch PaPeR
WUSCHEL-RELATED HOMEOBOX 8/9 is important for proper embryo patterning in the gymnosperm Norway spruce
Tianqing Zhu*, Panagiotis N. Moschou, José M. Alvarez, Joel J. Sohlberg and Sara von Arnold
Swedish University of Agricultural Sciences, Department of Plant Biology, Uppsala BioCenter, Linnean Center of Plant Biology in Uppsala, PO-Box 7080, SE-75007 Uppsala, Sweden
* To whom correspondence should be addressed. E-mail: [email protected]
Received 29 April 2014; Revised 5 August 2014; Accepted 13 August 2014
Abstract
Proper embryo development is crucial as that is when the primary body axes are established. In Arabidopsis, AtWOX8 and AtWOX9, members of the WUSCHEL-RELATED HOMEOBOX (WOX) gene family, are critical for embryo develop-ment. In Norway spruce, PaWOX8/9, which is expressed in embryos, is the homologue of AtWOX8 and AtWOX9. In this work, it is shown that the transcript abundance of PaWOX8/9 is high during early and late embryogeny and that it decreases when the maturation phase starts. To address the function of PaWOX8/9 during embryo development, RNAi lines were established to down-regulate the transcript level of PaWOX8/9, using both constitutive and induc-ible promoters. Embryos in the PaWOX8/9 RNAi lines show an aberrant morphology caused by disturbed orientation of the cell division plane at the basal part of the embryonal mass during early and late embryogeny. In addition, the transcript level of several key cell-cycle-regulating genes, for example, PaE2FAB-LIKE and PaCYCLIN B-LIKE, are affected in the PaWOX8/9 RNAi lines. Taken together, our results suggest that PaWOX8/9 may perform an evolutionar-ily conserved function as a regulator of the establishment of the apical–basal embryo pattern.
Key words: Apical–basal, cell cycle, division plane, embryogenesis, polarity, spruce, WUSCHEL-RELATED HOMEOBOX.
Introduction
In seed plants, the embryo displays the basic body polari-ties and develops along the apical–basal axis to establish two meristems responsible for post-embryonic growth (Ueda and Laux, 2012). The process that establishes this primary body plan is called embryonic pattern formation. It requires highly regulated spatio-temporal cell division to set up the organ plan and, eventually, the overall shape of the embryo (Berleth and Jurgens, 1993; Traas et al., 1995; Laux et al., 1996). The orientation of the cell division planes is critical as it not only decides the positions but also the fate of the daughter cells (Vandenberg et al., 1995). Knowledge about embryonic pattern formation in plants has, to a large extent, been derived from studies of embryo-defective mutants in the angiosperm model species Arabidopsis (Arabidopsis thali-ana) (Capron et al., 2009; Kanei et al., 2012). By contrast, our knowledge about the molecular regulation of embryo
development in conifers is limited, partly owing to the lack of embryo-defective mutants identified. However, the process of somatic embryogenesis has proved to be a valuable tool for studying the regulation of embryo development in conifers. Somatic embryos have been used for studying gene expression and functions (Tahir et al., 2006; Larsson et al., 2012a, b).
Early embryogenesis in Arabidopsis proceeds through highly regular cell division and gene expression patterns. In Arabidopsis, embryo pattern formation starts with an asym-metric division of the zygote which gives rise to a smaller apical cell and a larger basal cell. In conifers, the zygote undergoes several rounds of nuclear duplication which is not followed by cytokinesis and there is no clear asymmetric cell division defining apical and basal cell lineages (von Arnold et al., 2002). The descendants of the basal daughter of the Arabidopsis zygote only divide transversely to form one file of
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Abbreviations: EE, early embryo; LE, late embryo; ME, mature embryo; PEM, proembryogenic mass; PGR, plant growth regulator.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0/), which permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Experimental Botany Advance Access published September 9, 2014 at Sveriges lantbruksuniversitet on Septem
ber 17, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Page 2 of 10 | Zhu et al.
suspensor cells and the uppermost cell will form the hypophy-sis. BY contrast, the conifer suspensor consists of several files of non-dividing cells, originating from the basal part of the embryonal mass. The embryonal mass and the suspensor are separated by a layer of conifer-specific cells called embryonal tube cells. After the globular stage, the Arabidopsis embryo attains a bilateral symmetry when the two cotyledons start to differentiate. However, in conifers, multiple cotyledons are formed resulting in the embryo retaining radial symmetry. Despite the differences in patterning during embryo develop-ment between gymnosperms and angiosperms, recent results have shown that central parts of the regulatory network are conserved.
WUSCHEL-RELATED HOMEOBOX (WOX) genes form a large gene family in plants and exert important func-tions during all stages of plant development. The role of WOX genes during plant development has been studied in detail in Arabidopsis (Schoof et al., 2000). The WOX gene family members, AtWOX2, AtWOX8 (also named STIMPY-LIKE, STPL), and AtWOX9 (also named STIMPY, STIP) regulate early embryonic patterning in Arabidopsis. AtWOX2 is expressed in descendants of the apical daughter cell of the zygote and AtWOX8 and AtWOX9 are expressed in the basal daughter cell descendants (Haecker et al., 2004; Breuninger et al., 2008; Ueda et al., 2011). AtWOX8 and AtWOX9 share redundant functions but their expression patterns differ to some extent. In the shoot, AtWOX9 functions upstream of the WOX gene AtWUSCHEL, which is required for shoot apical meristem maintenance. AtWOX9 has also been linked to cell cycle regulation, as wox9 loss-of-function mutants are rescued by sucrose which stimulates entry into the cell cycle (Wu et al., 2005). Furthermore, wox8wox9 double mutants show aberrant cell division orientation in the embryo proper and suspensor (Breuninger et al., 2008).
Phylogenetic analyses have divided the WOX gene family into three major clades (the modern clade, the intermediate clade and the ancient clade) (van der Graaff et al., 2009). Eleven WOX genes have previously been cloned in Norway spruce (Picea abies) from both mRNA and genomic DNA and their phylogenetic relationship to other known WOX genes has been examined (Hedman et al., 2013). It was found that Norway spruce WOX genes are represented in all of the three major clades identified. The major diversi-fication within the WOX gene family took place before the split between gymnosperms and angiosperms. Furthermore, there has been a recent expansion within the intermediate clade of the Pinaceae family. The Norway spruce PaWOX8/9 gene, belonging to the intermediate clade, is most similar in sequence to AtWOX8 and AtWOX9 and it is preferentially expressed during embryo development (Palovaara et al., 2010; Hedman et al., 2013). In situ hybridization studies have confirmed that PaWOX8/9 is expressed during early embryo development in Norway spruce (Palovaara et al., 2010).
The functional analysis of PaWOX8/9 during embryo development is presented here. It is shown that PaWOX8/9 is important for the correct orientation of the cell division plane and cell fate determination during early embryo pat-tern formation, which suggests that PaWOX8/9 performs an
evolutionarily conserved function as an important regulator of apical–basal embryo pattern establishment.
Materials and methods
Plant materialThe embryogenic cell line 61:21 of Norway spruce (Picea abies L. Karst) has been used in this study. The cultures were treated as described previously (von Arnold and Clapham, 2008). Briefly, proembryogenic masses (PEMs) were maintained on solidified pro-liferation medium containing the plant growth regulators (PGRs) auxin and cytokinin. To stimulate differentiation of early embryos (EEs), cultures were transferred to pre-maturation medium lacking PGRs for one week. For the development of late embryos (LEs) and mature embryos (MEs), the cultures were transferred to maturation medium supplemented with abscisic acid (ABA). After partial desic-cation the embryos were germinated for two months.
RNA extraction, cDNA synthesis, and quantitative real-time PCRTo study the expression of PaWOX8/9 (accession number: GU944670) during embryo development, samples from PEMs, EEs, LEs and MEs were collected for RNA extraction. Sampling was per-formed at midday and samples were frozen in liquid nitrogen and stored at –80 °C after collection.
Total RNA was isolated using the Spectrum Plant Total RNA kit (Sigma-Aldrich, USA) according to the manufacturer’s instructions. For each sample, 1 µg of total RNA was reverse transcribed with the RevertAid H Minus First Strand cDNA Synthesis Kit (Fermentas, Thermo Scientific, Sweden) using an equimolar ratio of random and oligo-dT primers according to the manufacturer’s instructions.
Quantitative real-time PCR (qRT-PCR) was performed on an iCycler iQ PCR Thermal Cycler using the iQ5 Real-Time Detection System and 96-well PCR plates with adhesive seals (Bio-Rad Laboratories, USA). Three reference genes, CELL DIVISION CONTROL2 (PaCDC2), ELONGATION FACTOR 1 (PaEF1), and PHOSPHOGLUCOMUTASE (PaPHOS) were used (Vestman et al., 2011). Experiments were set up as previously described (Hedman et al., 2013). Two or three biological replicates, each with three tech-nical replicates were performed for each test. The primer sequences are presented in Supplementary Table S1 at JXB online. Significance analysis of transcript abundance was determined by t test.
RNA interference vector constructionTo study the function of PaWOX8/9, the coding sequence (CDS) of PaWOX8/9 was amplified from a cDNA library of early somatic embryos of Norway spruce (Hedman et al., 2013). The full-length CDS was subcloned into the pJET1.2/blunt cloning vector using the CloneJETTM PCR Cloning Kit (Fermentas, Thermo Scientific, Sweden). To obtain RNA interference (RNAi), overlapping frag-ments of PaWOX8/9 were amplified and were fused to form a hair-pin structure for PaWOX8/9 (see Supplementary Fig. S1 at JXB online). EcoRI and BamHI enzyme digestion sites were added on forward primers as linkers. The hairpin was confirmed by sequenc-ing. Primers are presented in Supplementary Table S2 at JXB online.
Hairpin structures were introduced into pENTR™/D-TOPO® (Invitrogen, Carlsbad, CA, USA) and then inserted by att site LR recombination into the destination vector pMDC7 [LexA-VP16-ER (XVE) β-estradiol inducible promoter, which is derived from the pER8 vector and contains the estrogen receptor-based transactiva-tor XVE] (Zuo et al., 2000; Brand et al., 2006) or pMDC32 (35S constitutive promoter) (Curtis and Grossniklaus, 2003). As a con-trol, an artificial microRNA (amiR) hairpin structure (Schwab et al., 2006; Weigel et al., 2008), designed to silence GUS, was inserted into pMDC7. Vectors were introduced by electroporation into Agrobacterium tumefaciens strain GV3101.
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

WOX8/9 function in Norway spruce | Page 3 of 10
Transgenic cell linesEmbryogenic cultures were transformed by co-cultivation with A. tumefaciens according to the protocol by Wenck et al. (1999), with modifications. Briefly, after initial co-cultivation in liquid trans-formation medium for 5 h, the cultures were collected on filter paper and placed on solidified proliferation medium. After 48 h, the cul-tures were transferred to proliferation medium containing 400 mg l–1 timentin (Duchefa, The Netherlands) and 250 mg l–1 cefotaxime (Sanofi, France). Four days later, cultures were transferred to prolif-eration medium containing 400 mg l–1 timentin, 250 mg l–1 cefotax-ime, and 15 mg l–1 hygromycin B (Duchefa). Thereafter, the cultures were subcultured to fresh culture medium of the same composition every week.
Stable transformants were selected after 4 weeks, for 35S:WOX8/9i, XVE-WOX8/9i, and XVE-amiRGUS transformed control lines (T-control). Genomic DNA was isolated from PEMs from the selected lines by using the DNeasy Plant Mini Kit (Qiagen, Germany), according to the manufacturer’s instructions. Transformants were confirmed by PCR.
Transcript abundance of PaWOX8/9 in PEMs was analysed by qRT-PCR. In the case of XVE-WOX8/9i lines, the cultures were exposed to β-estradiol (10 µM) for 48 h before analysis. Based on the qRT-PCR data, the following lines were selected for further studies: XVE-WOX8/9i.1, XVE-WOX8/9i.2, XVE-WOX8/9i.3, 35S:WOX8/9i.1, 35S:WOX8/9i.2, 35S:WOX8/9i.4, and 35S:WOX8/9i.6. In addition, two T-control lines XVE-amiRGUS.3 (T.3-control) and XVE-amiRGUS.4 (T.4-control) were selected. For activation of the XVE promoter during maturation the maturation medium was supplemented with 10 µM β-estradiol and the cultures were transferred to fresh medium of the same composition every week.
Morphological analysisTo assess the effect of PaWOX8/9 on embryo morphology dur-ing early embryogeny and at the beginning of late embryogeny, samples were collected from cultures of untransformed control (U-control), XVE-WOX8/9i.3 (non-induced and induced) and lines 35S:WOX8/9i.1, 35S:WOX8/9i.2, 35S:WOX8/9i.4, 35S:WOX8/9i.6. The samples were solidified by mixing with 2 ml of 1.2% (w/v) Seaplaque agarose (FMC BioProducts, USA) in 60 mm Petri dishes.
To study the morphology of EEs (n=26) from U-control and line 35S:WOX8/9i.4, embryos were scanned with a Zeiss 780 confocal microscope (Carl Zeiss AG, Germany) using a 488 nm Argon laser under the 20× objective (NA=0.8).
The developmental pathway for embryos with normal or cone-shaped morphology at the beginning of late embryogeny was followed for 10 d. Embryos from U-control cultures and lines 35S:WOX8/9i.2, 35S:WOX8/9i.4, and 35S:WOX8/9i.6 were sam-pled after 2 weeks on maturation medium and transferred to fresh maturation medium. Time-lapse tracking analyses were performed with about 50 embryos per line. Phenotypes were scored every second day.
For analysing the effect of PaWOX8/9 during embryo maturation, the morphology of embryos in the U-control and XVE-WOX8/9i lines was analysed for up to 10 weeks on maturation medium. β-estradiol (10 µM) was added to the maturation medium either from the start of the maturation treatment or after 2 weeks on matu-ration medium when LEs had developed. The germination rate was estimated after 2 months on germination medium. The data from phenotypic differences of early and late embryos were assessed in the ANOVA Mixed-model (R Development Core Team 2008, ver. 2.15.1).
For histological analysis, mature embryos from line XVE-WOX8/9i.3 as well as from U-control cultures were fixed according to the protocol by Karlgren et al. (2009). The embryos were serial-sectioned (8 µM) using a Zeiss HM 355 microtome. Cell size was measured by image analysis with ImageJ (ver. 1.48g) (Schneider et al., 2012).
Identification and qRT-PCR analyses of cell-cycle-regulating genesTo analyse if PaWOX8/9 affects the expression of cell-cycle-reg-ulating genes, the transcript abundance of cell-cycle-regulating genes was compared in PEMs from the U-control and from line 35S:WOX8/9i.4 by qRT-PCR. Cell-cycle-regulating genes were identified by homology search over the Norway spruce genome database (Nystedt et al., 2013) combined with phylogenetic analy-sis see Supplementary Fig. S3 and the Supplementary Information at JXB online). The accession numbers of the selected genes are presented in Supplementary Table S7 at JXB online. Ten of these genes, PaRETINOBLASTOMA-RELATED PROTEIN-LIKE (PaRBRL), two E2F family genes (PaE2FABL), five CYCLIN-LIKE (PaCYCLs) genes, PaMAP KINASE 6-LIKE (PaMPK6L), and PaEXTRA SPINDLE POLES (PaESP) were selected for qRT-PCR analysis. Three reference genes, PaCDC2, PaEF1, and PaPHOS, were used. The primer sequences are presented in Supplementary Tables S1 and S8 at JXB online.
Results
Transcript abundance of PaWOX8/9 during the development of somatic embryos
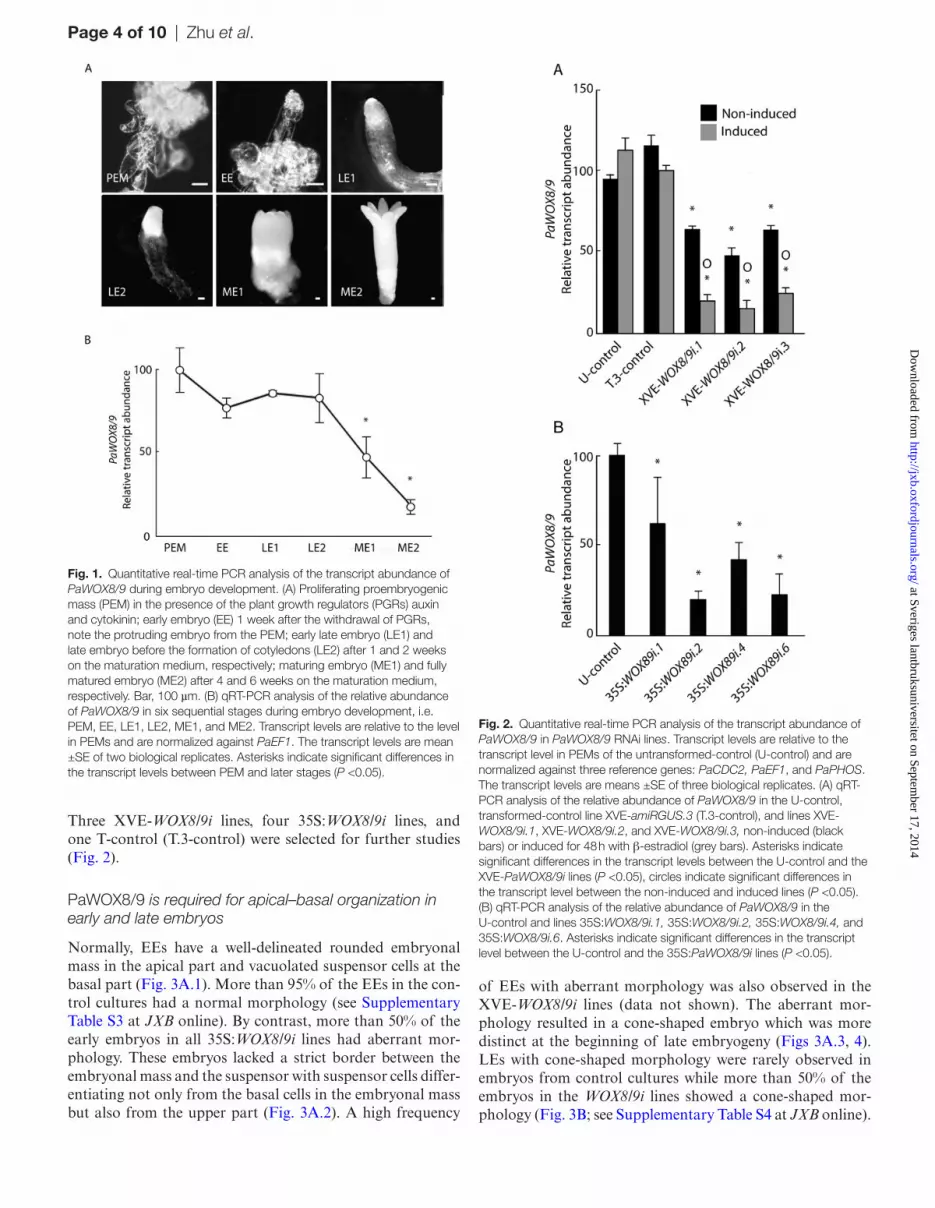
The developmental stages during somatic embryogenesis in Norway spruce have been described (Filonova et al., 2000; Larsson et al., 2012b). The stages used in this study include proliferating proembryogenic masses (PEM), early embryos (EE), early late embryos (LE1), late embryos (LE2), maturing embryos (ME1), and fully matured embryos (ME2) (Fig. 1A).
Transcript levels of PaWOX8/9 were high in PEMs, EEs, LE1s, and LE2s, but significantly lower in ME1s and ME2s (Fig. 1B). PaWOX8/9 is expressed t the beginning of embryo development in Norway spruce (Palovaara et al., 2010; Hedman et al., 2013). The expression of PaWOX8/9 in embryos suggests that PaWOX8/9 is involved in regulating embryo development.
Transcript abundance of PaWOX8/9 in RNAi lines
In order to study the function of PaWOX8/9 during embry-ogenesis, PaWOX8/9 RNAi lines were constructed using both inducible and constitutive promoters. Two controls have been used, an untransformed control (U-control) and a transformed control (T-control; described in the Materials and methods). The transcript levels of PaWOX8/9 in some XVE-WOX8/9i lines decreased by more than 50% in PEMs after 48 h of β-estradiol treatment (see Supplementary Fig. S2A at JXB online). A similar difference in transcript abun-dance of PaWOX8/9 in some XVE-WOX8/9i lines was also observed after 4 weeks exposure to β-estradiol (data not shown). β-estradiol treatment of the control lines did not significantly affect the transcript abundance of PaWOX8/9. Notably, the transcript level of PaWOX8/9 in non-induced XVE-WOX8/9i lines was lower than in the control lines. This implies that the XVE promoter is partly activated in Norway spruce even in the absence of β-estradiol. In most of the 35S:WOX8/9i lines, the transcript abundance of PaWOX8/9 was significantly decreased by c. 60% compared with the U-control (see Supplementary Fig. S2B at JXB online).
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Page 4 of 10 | Zhu et al.
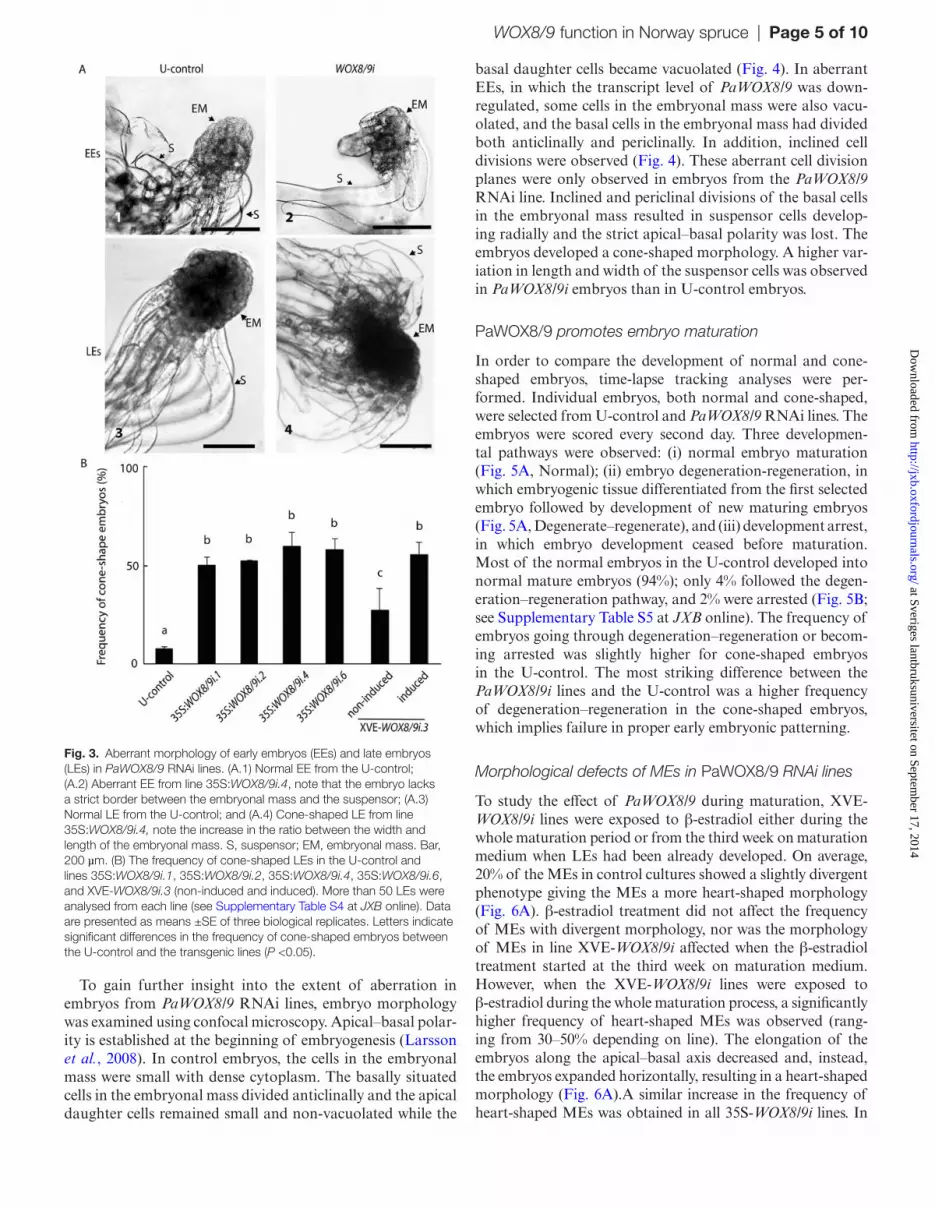
Three XVE-WOX8/9i lines, four 35S:WOX8/9i lines, and one T-control (T.3-control) were selected for further studies (Fig. 2).
PaWOX8/9 is required for apical–basal organization in early and late embryos
Normally, EEs have a well-delineated rounded embryonal mass in the apical part and vacuolated suspensor cells at the basal part (Fig. 3A.1). More than 95% of the EEs in the con-trol cultures had a normal morphology (see Supplementary Table S3 at JXB online). By contrast, more than 50% of the early embryos in all 35S:WOX8/9i lines had aberrant mor-phology. These embryos lacked a strict border between the embryonal mass and the suspensor with suspensor cells differ-entiating not only from the basal cells in the embryonal mass but also from the upper part (Fig. 3A.2). A high frequency
of EEs with aberrant morphology was also observed in the XVE-WOX8/9i lines (data not shown). The aberrant mor-phology resulted in a cone-shaped embryo which was more distinct at the beginning of late embryogeny (Figs 3A.3, 4). LEs with cone-shaped morphology were rarely observed in embryos from control cultures while more than 50% of the embryos in the WOX8/9i lines showed a cone-shaped mor-phology (Fig. 3B; see Supplementary Table S4 at JXB online).
Fig. 2. Quantitative real-time PCR analysis of the transcript abundance of PaWOX8/9 in PaWOX8/9 RNAi lines. Transcript levels are relative to the transcript level in PEMs of the untransformed-control (U-control) and are normalized against three reference genes: PaCDC2, PaEF1, and PaPHOS. The transcript levels are means ±SE of three biological replicates. (A) qRT-PCR analysis of the relative abundance of PaWOX8/9 in the U-control, transformed-control line XVE-amiRGUS.3 (T.3-control), and lines XVE-WOX8/9i.1, XVE-WOX8/9i.2, and XVE-WOX8/9i.3, non-induced (black bars) or induced for 48 h with β-estradiol (grey bars). Asterisks indicate significant differences in the transcript levels between the U-control and the XVE-PaWOX8/9i lines (P <0.05), circles indicate significant differences in the transcript level between the non-induced and induced lines (P <0.05). (B) qRT-PCR analysis of the relative abundance of PaWOX8/9 in the U-control and lines 35S:WOX8/9i.1, 35S:WOX8/9i.2, 35S:WOX8/9i.4, and 35S:WOX8/9i.6. Asterisks indicate significant differences in the transcript level between the U-control and the 35S:PaWOX8/9i lines (P <0.05).
Fig. 1. Quantitative real-time PCR analysis of the transcript abundance of PaWOX8/9 during embryo development. (A) Proliferating proembryogenic mass (PEM) in the presence of the plant growth regulators (PGRs) auxin and cytokinin; early embryo (EE) 1 week after the withdrawal of PGRs, note the protruding embryo from the PEM; early late embryo (LE1) and late embryo before the formation of cotyledons (LE2) after 1 and 2 weeks on the maturation medium, respectively; maturing embryo (ME1) and fully matured embryo (ME2) after 4 and 6 weeks on the maturation medium, respectively. Bar, 100 µm. (B) qRT-PCR analysis of the relative abundance of PaWOX8/9 in six sequential stages during embryo development, i.e. PEM, EE, LE1, LE2, ME1, and ME2. Transcript levels are relative to the level in PEMs and are normalized against PaEF1. The transcript levels are mean ±SE of two biological replicates. Asterisks indicate significant differences in the transcript levels between PEM and later stages (P <0.05).
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
WOX8/9 function in Norway spruce | Page 5 of 10
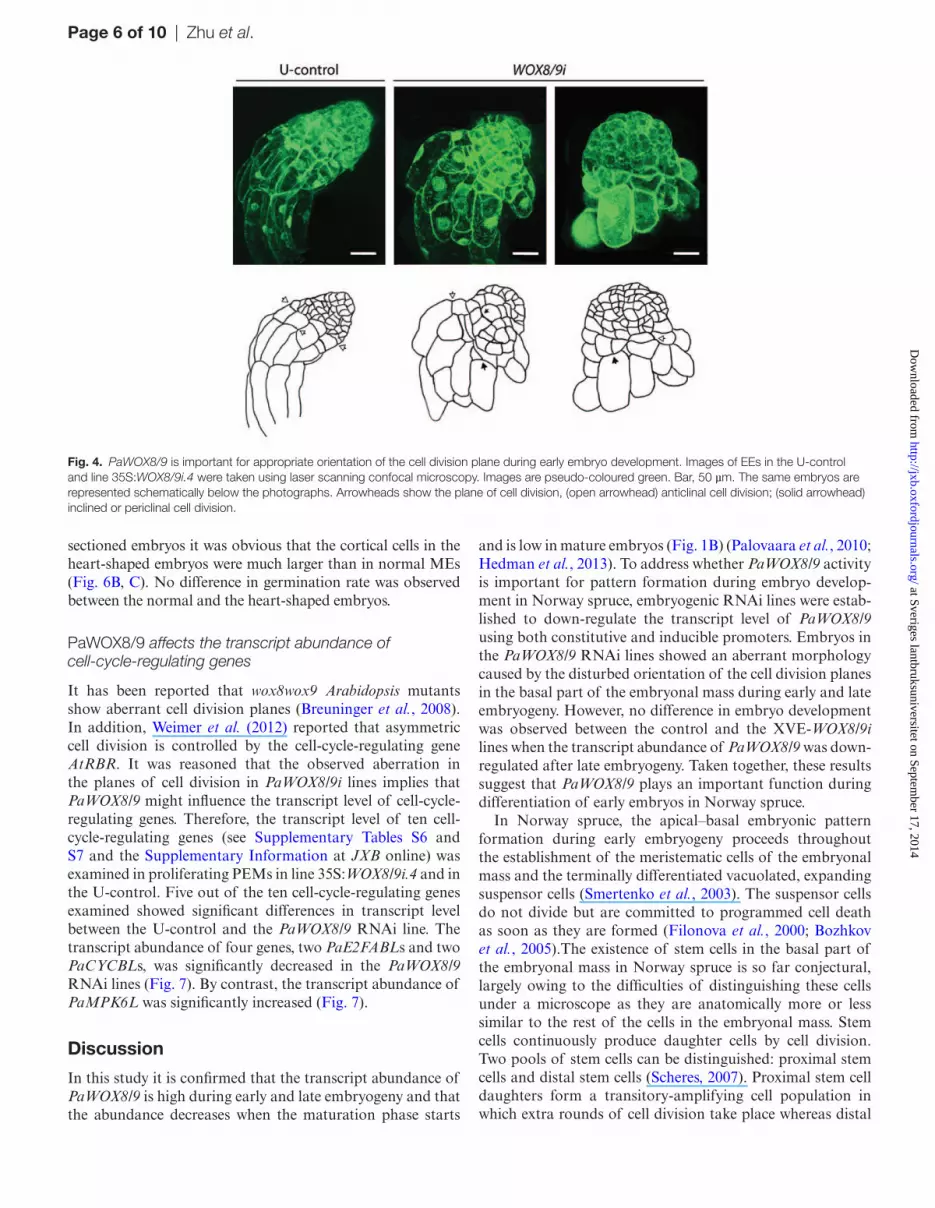
To gain further insight into the extent of aberration in embryos from PaWOX8/9 RNAi lines, embryo morphology was examined using confocal microscopy. Apical–basal polar-ity is established at the beginning of embryogenesis (Larsson et al., 2008). In control embryos, the cells in the embryonal mass were small with dense cytoplasm. The basally situated cells in the embryonal mass divided anticlinally and the apical daughter cells remained small and non-vacuolated while the
basal daughter cells became vacuolated (Fig. 4). In aberrant EEs, in which the transcript level of PaWOX8/9 was down-regulated, some cells in the embryonal mass were also vacu-olated, and the basal cells in the embryonal mass had divided both anticlinally and periclinally. In addition, inclined cell divisions were observed (Fig. 4). These aberrant cell division planes were only observed in embryos from the PaWOX8/9 RNAi line. Inclined and periclinal divisions of the basal cells in the embryonal mass resulted in suspensor cells develop-ing radially and the strict apical–basal polarity was lost. The embryos developed a cone-shaped morphology. A higher var-iation in length and width of the suspensor cells was observed in PaWOX8/9i embryos than in U-control embryos.
PaWOX8/9 promotes embryo maturation
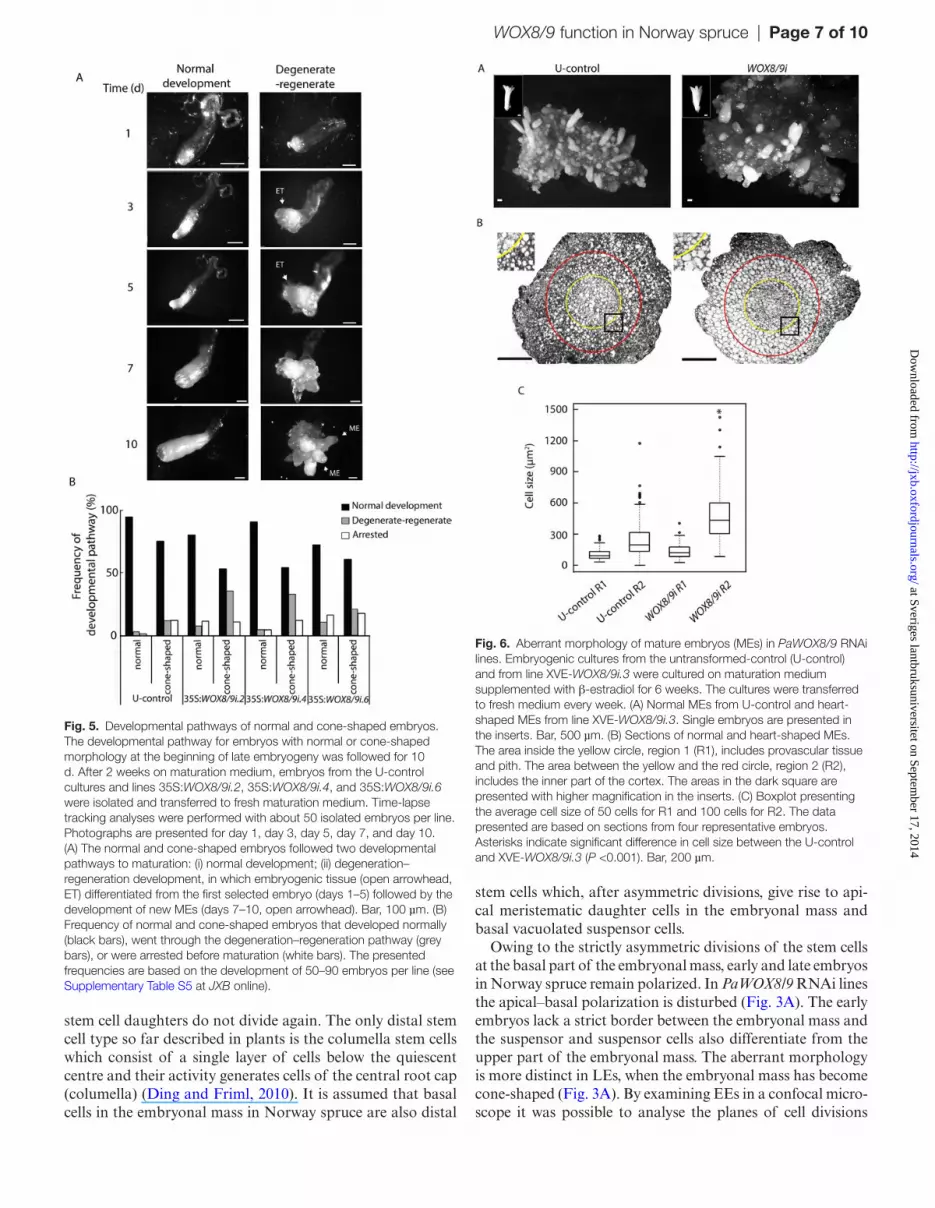
In order to compare the development of normal and cone-shaped embryos, time-lapse tracking analyses were per-formed. Individual embryos, both normal and cone-shaped, were selected from U-control and PaWOX8/9 RNAi lines. The embryos were scored every second day. Three developmen-tal pathways were observed: (i) normal embryo maturation (Fig. 5A, Normal); (ii) embryo degeneration-regeneration, in which embryogenic tissue differentiated from the first selected embryo followed by development of new maturing embryos (Fig. 5A, Degenerate–regenerate), and (iii) development arrest, in which embryo development ceased before maturation. Most of the normal embryos in the U-control developed into normal mature embryos (94%); only 4% followed the degen-eration–regeneration pathway, and 2% were arrested (Fig. 5B; see Supplementary Table S5 at JXB online). The frequency of embryos going through degeneration–regeneration or becom-ing arrested was slightly higher for cone-shaped embryos in the U-control. The most striking difference between the PaWOX8/9i lines and the U-control was a higher frequency of degeneration–regeneration in the cone-shaped embryos, which implies failure in proper early embryonic patterning.
Morphological defects of MEs in PaWOX8/9 RNAi lines
To study the effect of PaWOX8/9 during maturation, XVE-WOX8/9i lines were exposed to β-estradiol either during the whole maturation period or from the third week on maturation medium when LEs had been already developed. On average, 20% of the MEs in control cultures showed a slightly divergent phenotype giving the MEs a more heart-shaped morphology (Fig. 6A). β-estradiol treatment did not affect the frequency of MEs with divergent morphology, nor was the morphology of MEs in line XVE-WOX8/9i affected when the β-estradiol treatment started at the third week on maturation medium. However, when the XVE-WOX8/9i lines were exposed to β-estradiol during the whole maturation process, a significantly higher frequency of heart-shaped MEs was observed (rang-ing from 30–50% depending on line). The elongation of the embryos along the apical–basal axis decreased and, instead, the embryos expanded horizontally, resulting in a heart-shaped morphology (Fig. 6A).A similar increase in the frequency of heart-shaped MEs was obtained in all 35S-WOX8/9i lines. In
Fig. 3. Aberrant morphology of early embryos (EEs) and late embryos (LEs) in PaWOX8/9 RNAi lines. (A.1) Normal EE from the U-control; (A.2) Aberrant EE from line 35S:WOX8/9i.4, note that the embryo lacks a strict border between the embryonal mass and the suspensor; (A.3) Normal LE from the U-control; and (A.4) Cone-shaped LE from line 35S:WOX8/9i.4, note the increase in the ratio between the width and length of the embryonal mass. S, suspensor; EM, embryonal mass. Bar, 200 µm. (B) The frequency of cone-shaped LEs in the U-control and lines 35S:WOX8/9i.1, 35S:WOX8/9i.2, 35S:WOX8/9i.4, 35S:WOX8/9i.6, and XVE-WOX8/9i.3 (non-induced and induced). More than 50 LEs were analysed from each line (see Supplementary Table S4 at JXB online). Data are presented as means ±SE of three biological replicates. Letters indicate significant differences in the frequency of cone-shaped embryos between the U-control and the transgenic lines (P <0.05).
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Page 6 of 10 | Zhu et al.
sectioned embryos it was obvious that the cortical cells in the heart-shaped embryos were much larger than in normal MEs (Fig. 6B, C). No difference in germination rate was observed between the normal and the heart-shaped embryos.
PaWOX8/9 affects the transcript abundance of cell-cycle-regulating genes
It has been reported that wox8wox9 Arabidopsis mutants show aberrant cell division planes (Breuninger et al., 2008). In addition, Weimer et al. (2012) reported that asymmetric cell division is controlled by the cell-cycle-regulating gene AtRBR. It was reasoned that the observed aberration in the planes of cell division in PaWOX8/9i lines implies that PaWOX8/9 might influence the transcript level of cell-cycle-regulating genes. Therefore, the transcript level of ten cell-cycle-regulating genes (see Supplementary Tables S6 and S7 and the Supplementary Information at JXB online) was examined in proliferating PEMs in line 35S:WOX8/9i.4 and in the U-control. Five out of the ten cell-cycle-regulating genes examined showed significant differences in transcript level between the U-control and the PaWOX8/9 RNAi line. The transcript abundance of four genes, two PaE2FABLs and two PaCYCBLs, was significantly decreased in the PaWOX8/9 RNAi lines (Fig. 7). By contrast, the transcript abundance of PaMPK6L was significantly increased (Fig. 7).
Discussion
In this study it is confirmed that the transcript abundance of PaWOX8/9 is high during early and late embryogeny and that the abundance decreases when the maturation phase starts
and is low in mature embryos (Fig. 1B) (Palovaara et al., 2010; Hedman et al., 2013). To address whether PaWOX8/9 activity is important for pattern formation during embryo develop-ment in Norway spruce, embryogenic RNAi lines were estab-lished to down-regulate the transcript level of PaWOX8/9 using both constitutive and inducible promoters. Embryos in the PaWOX8/9 RNAi lines showed an aberrant morphology caused by the disturbed orientation of the cell division planes in the basal part of the embryonal mass during early and late embryogeny. However, no difference in embryo development was observed between the control and the XVE-WOX8/9i lines when the transcript abundance of PaWOX8/9 was down-regulated after late embryogeny. Taken together, these results suggest that PaWOX8/9 plays an important function during differentiation of early embryos in Norway spruce.
In Norway spruce, the apical–basal embryonic pattern formation during early embryogeny proceeds throughout the establishment of the meristematic cells of the embryonal mass and the terminally differentiated vacuolated, expanding suspensor cells (Smertenko et al., 2003). The suspensor cells do not divide but are committed to programmed cell death as soon as they are formed (Filonova et al., 2000; Bozhkov et al., 2005).The existence of stem cells in the basal part of the embryonal mass in Norway spruce is so far conjectural, largely owing to the difficulties of distinguishing these cells under a microscope as they are anatomically more or less similar to the rest of the cells in the embryonal mass. Stem cells continuously produce daughter cells by cell division. Two pools of stem cells can be distinguished: proximal stem cells and distal stem cells (Scheres, 2007). Proximal stem cell daughters form a transitory-amplifying cell population in which extra rounds of cell division take place whereas distal
Fig. 4. PaWOX8/9 is important for appropriate orientation of the cell division plane during early embryo development. Images of EEs in the U-control and line 35S:WOX8/9i.4 were taken using laser scanning confocal microscopy. Images are pseudo-coloured green. Bar, 50 µm. The same embryos are represented schematically below the photographs. Arrowheads show the plane of cell division, (open arrowhead) anticlinal cell division; (solid arrowhead) inclined or periclinal cell division.
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
WOX8/9 function in Norway spruce | Page 7 of 10
stem cell daughters do not divide again. The only distal stem cell type so far described in plants is the columella stem cells which consist of a single layer of cells below the quiescent centre and their activity generates cells of the central root cap (columella) (Ding and Friml, 2010). It is assumed that basal cells in the embryonal mass in Norway spruce are also distal
stem cells which, after asymmetric divisions, give rise to api-cal meristematic daughter cells in the embryonal mass and basal vacuolated suspensor cells.
Owing to the strictly asymmetric divisions of the stem cells at the basal part of the embryonal mass, early and late embryos in Norway spruce remain polarized. In PaWOX8/9 RNAi lines the apical–basal polarization is disturbed (Fig. 3A). The early embryos lack a strict border between the embryonal mass and the suspensor and suspensor cells also differentiate from the upper part of the embryonal mass. The aberrant morphology is more distinct in LEs, when the embryonal mass has become cone-shaped (Fig. 3A). By examining EEs in a confocal micro-scope it was possible to analyse the planes of cell divisions
Fig. 5. Developmental pathways of normal and cone-shaped embryos. The developmental pathway for embryos with normal or cone-shaped morphology at the beginning of late embryogeny was followed for 10 d. After 2 weeks on maturation medium, embryos from the U-control cultures and lines 35S:WOX8/9i.2, 35S:WOX8/9i.4, and 35S:WOX8/9i.6 were isolated and transferred to fresh maturation medium. Time-lapse tracking analyses were performed with about 50 isolated embryos per line. Photographs are presented for day 1, day 3, day 5, day 7, and day 10. (A) The normal and cone-shaped embryos followed two developmental pathways to maturation: (i) normal development; (ii) degeneration–regeneration development, in which embryogenic tissue (open arrowhead, ET) differentiated from the first selected embryo (days 1–5) followed by the development of new MEs (days 7–10, open arrowhead). Bar, 100 µm. (B) Frequency of normal and cone-shaped embryos that developed normally (black bars), went through the degeneration–regeneration pathway (grey bars), or were arrested before maturation (white bars). The presented frequencies are based on the development of 50–90 embryos per line (see Supplementary Table S5 at JXB online).
Fig. 6. Aberrant morphology of mature embryos (MEs) in PaWOX8/9 RNAi lines. Embryogenic cultures from the untransformed-control (U-control) and from line XVE-WOX8/9i.3 were cultured on maturation medium supplemented with β-estradiol for 6 weeks. The cultures were transferred to fresh medium every week. (A) Normal MEs from U-control and heart-shaped MEs from line XVE-WOX8/9i.3. Single embryos are presented in the inserts. Bar, 500 µm. (B) Sections of normal and heart-shaped MEs. The area inside the yellow circle, region 1 (R1), includes provascular tissue and pith. The area between the yellow and the red circle, region 2 (R2), includes the inner part of the cortex. The areas in the dark square are presented with higher magnification in the inserts. (C) Boxplot presenting the average cell size of 50 cells for R1 and 100 cells for R2. The data presented are based on sections from four representative embryos. Asterisks indicate significant difference in cell size between the U-control and XVE-WOX8/9i.3 (P <0.001). Bar, 200 µm.
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Page 8 of 10 | Zhu et al.
(Fig. 4). In the normal, control embryos, the stem cells at the basal part of the embryonal mass divide anticlinally to give rise to one cell which remains in the embryonal mass and one vacuolated suspensor cell. By contrast, in embryos with decreased transcript levels of PaWOX8/9, the basal stem cells of the embryonal mass divide anticlinally, periclinally, and are also inclined. After inclined and periclinal divisions, it appears that both daughter cells remain in the basal part of the embryonal mass. It is likely that the periclinal cell divisions result in radial growth of the embryonal mass, which gives the LEs a cone-shaped morphology. Inclined and periclinal cell divisions might also explain why vacuolated suspensor cells not only develop along the apical–basal axis but also develop radially. It has been shown that stem-cell daughters can adopt different fates depending on their position, and that plant cell fate is spatially determined and requires highly regulated cell division (Scheres et al., 1995; Vandenberg et al., 1995). Our results suggest that (i) the shape of early embryos in Norway spruce depends on the orientation of cell division planes in the stem cells in the basal part of the embryonal mass, and (ii) PaWOX8/9 directly or indirectly regulates cell division orien-tation and is important for the cell fate determination required for normal apical–basal organization in early embryos. A sim-ilar function has been shown in Arabidopsis where AtWOX8 and AtWOX9 regulate cell morphology and division pattern in both the basal and the apical lineages from the single-cell embryo stage (Breuninger et al., 2008).
By tracking the development of normal and cone-shaped LEs in control and PaWOX8/9 RNAi lines, about 30% of the cone-shaped embryos with down-regulated transcript levels of PaWOX8/9 develop abnormally (Fig. 5). Initially, new embryogenic tissues differentiate from the LEs and, after 1 week, new maturing embryos develop from the embryogenic tissue. It is known that a strict balance between cell division and cell differentiation is essential for normal embryo devel-opment and that randomization of cell division planes in embryos leads to drastic morphological defects (Berleth and Jurgens, 1993; Traas et al., 1995; Laux et al., 1996). Although it has not been possible to trace from which cells in the cone-shaped embryos the embryogenic tissue differentiate, it is likely that the division-plane alterations in the stem cells at the basal part of the embryonal mass change cell identity and that some cells have retained meristematic identity.
Decreased transcript abundance of PaWOX8/9 during the differentiation of EEs and the development of LEs gives a high frequency of cone-shaped LEs (Fig. 3A.4) and heart-shaped MEs (Fig. 6A). However, if the transcript level of PaWOX8/9 is down-regulated later during the maturation process, the embryos develop into normal cotyledonary embryos. This suggests that PaWOX8/9 is important during early embryo development but not at late embryogeny. The heart-shaped MEs observed in the PaWOX8/9 RNAi lines (Fig. 6A) resemble those of the AtWOX9 mutant stip-1 in Arabidopsis which fail to elongate along the apical–basal axis and cells expand horizontally (Wu et al., 2007).
Cell division is regulated through a complex array of molecular events. In Arabidopsis, these events involve fac-tors like sucrose and auxin, which stimulate the expression of CYCD. CYCD together with cyclin-dependent kinase (CDK) phosphorylates RBR1 which consequently is released from the E2F/dimerization partner (DP) complex. The E2F/DP complex is then activated to promote (E2FA or E2FB) or inhibit (E2FC) the entrance into the S phase. The RBR/E2F/DP pathway is conserved as both animals and plants use it to control the G1-S transition (Cruz-Ramirez et al., 2012). During the S phase, CYCA forms a complex with CDKA. CYCA is then replaced by CYCB, which stimulates entrance into mitosis (Inzé and De Veylder, 2006; Nieuwland et al., 2009). ESP promotes the metaphase to anaphase transition and cell polarity acquisition (Wu et al., 2010; Moschou et al., 2013). Furthermore, MPK6 is linked to the execution of developmental programmes involving embryogenesis (Hord et al., 2008; Wang et al., 2008).
Our results suggest that PaWOX8/9 directly or indirectly controls the transcript level of some cell-cycle-regulating genes. The most conspicuous change is the decrease in tran-script levels of PaE2F and PaCYCBL in PaWOX8/9 RNAi lines (Fig. 7). Interestingly, expression of AtCYCBs is under the control of AtE2Fs (del Pozo et al., 2006). AtCYCBs reg-ulate both the G2-M transition and intra-M-phase control (Breyne and Zabeau, 2001; Potuschak and Doerner, 2001).
Over-expression of AtE2FA or AtE2FB activates mito-sis, resulting in more cell proliferation with more cells but of smaller size (Magyar et al., 2012). By contrast, in multi-cellular organisms, a decrease in cell number triggers an
Fig. 7. Quantitative real-time PCR analysis of the transcript abundance of cell-cycle-regulating genes in the untransformed control (black bars) and line 35S:WOX8/9i.4 (grey bars). The transcript level of cell-cycle-regulating genes was analysed in proliferating PEMs in the U-control and in line 35S:WOX8/9i.4. The transcript level of the following genes was analysed (for gene accession numbers and primer sequences, see Supplementary Tables S7 and S8 at JXB online): PaRBRL, PaE2FABL1, PaE2FABL2, and PaCYCAL1, PaCYCAL2, PaCYCAL3, PaCYCBL1, PaCYCBL2, PaMPK6L, and PaESP. The main functional stages during the cell cycle are illustrated by the lines below. Transcript values are relative to the transcript level of PaE2FABL1 in PEMs in the U-control and are normalized against three reference genes: PaCDC2, PaEF1, and PaPHOS. The transcript levels presented are means ±SE of two biological replicates. Asterisks indicate significant differences in transcript level between the U-control and the 35S:WOX8/9i.4 line (P <0.05).
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

WOX8/9 function in Norway spruce | Page 9 of 10
increase in mature cell size to compensate for reduced organ size (Ferjani et al., 2007). In accordance, the decrease in the transcript abundance of PaE2Fs in PaWOX8/9 RNAi lines might have caused a decrease in cell number by repressing mitosis. The frequency of heart-shaped MEs is higher in PaWOX8/9 RNAi lines, and in these embryos, the cortical cells are larger than those in normal-shaped MEs (Fig. 6B). It is assumed that the increased frequency of heart-shaped MEs in PaWOX8/9 RNAi lines is caused by a disturbed cell divi-sion pattern during early embryo development.
In conclusion, our results suggest that PaWOX8/9 acts as an important regulator for establishing the apical–basal embryo pattern and that this function is evolutionarily con-served between gymnosperms and angiosperms. This func-tion is accomplished by controlling the orientation of the cell division planes and cell fate determination during early embryonic pattern formation. In addition, PaWOX8/9 is active upstream of several important cell-cycle regulators. Future studies will reveal the precise function of PaWOX8/9 in the regulation of the complex gene regulatory network that determines the symmetry of cell division and how this infor-mation is integrated at the whole organismal level to attain proper shape.
Supplementary data
Supplementary data can be found at JXB online.Supplementary Table S1. Primer sequences used for qRT-
PCR of PaWOX8/9 and corresponding reference genes.Supplementary Table S2. Primer sequences used for vector
construction of PaWOX8/9 interference.Supplementary Table S3. Frequency of aberrant early
embryos (EEs) in PaWOX8/9 RNAi lines.Supplementary Table S4. Frequency of cone-shaped late
embryos (LEs) in WOX8/9 RNAi lines.Supplementary Table S5. Tracking of the developmental
pathway of normal and cone-shaped late embryos (LEs).Supplementary Table S6. List of the selected cell-cycle-reg-
ulating genes from Arabidopsis thaliana.Supplementary Table S7. List of the selected cell-cycle-reg-
ulating genes from Norway spruce.Supplementary Table S8. Primer sequences used for qRT-
PCR of the cell-cycle-regulating genes from Norway spruce.Supplementary Fig S1. Schematic illustration of the CDS
of PaWOX8/9.Supplementary Fig S2. Quantitative real-time PCR analy-
sis of the transcript abundance of PaWOX8/9 in PaWOX8/9 RNAi lines.
Supplementary Fig. S3. Phylogenetic trees of PaE2FAB-LIKE genes (A) and PaCYCLIN-LIKE genes (B).
Supplementary Information. Identification and selection of cell-cycle-regulating genes in Norway spruce.
AcknowledgementsThe Swedish governmental research programme Trees and Crops for the Future (TC4F) is acknowledged for financial support. Dr Ueli Grossniklaus
and Dr Nam-Hai Chua are acknowledged for providing the vector pMDC7. Dr Ueli Grossniklaus is acknowledged for providing the vector pMDC32. The authors are grateful to Dr David Clapham for critical reading of the manuscript.
ReferencesBerleth T, Jurgens G. 1993. The role of the monopteros gene in organizing the basal body region of the Arabidopsis embryo. Development 118, 575–587.
Bozhkov PV, Filonova LH, Suarez MF. 2005. Programmed cell death in plant embryogenesis. Current Topics in Developmental Biology 67, 135–179.
Brand L, Horler M, Nuesch E, Vassalli S, Barrell P, Yang W, Jefferson RA, Grossniklaus U, Curtis MD. 2006. A versatile and reliable two-component system for tissue-specific gene induction in Arabidopsis. Plant Physiology 141, 1194–1204.
Breuninger H, Rikirsch E, Hermann M, Ueda M, Laux T. 2008. Differential expression of WOX genes mediates apical–basal axis formation in the Arabidopsis embryo. Developmental Cell 14, 867–876.
Breyne P, Zabeau M. 2001. Genome-wide expression analysis of plant cell cycle modulated genes. Current Opinion in Plant Biology 4, 136–142.
Capron A, Chatfield S, Provart N, Berleth T. 2009. Embryogenesis: pattern formation from a single cell. The Arabidopsis Book 7, e0126.
Cruz-Ramirez A, Diaz-Trivino S, Blilou I, et al. 2012. A bistable circuit involving SCARECROW-RETINOBLASTOMA integrates cues to inform asymmetric stem cell division. Cell 150, 1002–1015.
Curtis MD, Grossniklaus U. 2003. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiology 133, 462–469.
del Pozo JC, Diaz-Trivino S, Cisneros N, Gutierrez C. 2006. The balance between cell division and endoreplication depends on E2FC-DPB, transcription factors regulated by the ubiquitin-SCFSKP2A pathway in Arabidopsis. The Plant Cell 18, 2224–2235.
Ding Z, Friml J. 2010. Auxin regulates distal stem cell differentiation in Arabidopsis roots. Proceedings of the National Academy of Sciences, USA 107, 12046–12051.
Ferjani A, Horiguchi G, Yano S, Tsukaya H. 2007. Analysis of leaf development in fugu mutants of Arabidopsis reveals three compensation modes that modulate cell expansion in determinate organs. Plant Physiology 144, 988–999.
Filonova LH, Bozhkov PV, von Arnold S. 2000. Developmental pathway of somatic embryogenesis in Picea abies as revealed by time-lapse tracking. Journal of Experimental Botany 51, 249–264.
Haecker A, Gross-Hardt R, Geiges B, Sarkar A, Breuninger H, Herrmann M, Laux T. 2004. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development 131, 657–668.
Hedman H, Zhu T, von Arnold S, Sohlberg JJ. 2013. Analysis of the WUSCHEL-RELATED HOMEOBOX gene family in the conifer Picea abies reveals extensive conservation as well as dynamic patterns. BMC Plant Biology 13, 89.
Hord CL, Sun YJ, Pillitteri LJ, Torii KU, Wang H, Zhang S, Ma H. 2008. Regulation of Arabidopsis early anther development by the mitogen-activated protein kinases, MPK3 and MPK6, and the ERECTA and related receptor-like kinases. Molecular Plant 1, 645–658.
Inzé D, De Veylder L. 2006. Cell cycle regulation in plant development. Annual Review of Genetics 40, 77–105.
Kanei M, Horiguchi G, Tsukaya H. 2012. Stable establishment of cotyledon identity during embryogenesis in Arabidopsis by ANGUSTIFOLIA3 and HANABA TARANU. Development 139, 2436–2446.
Karlgren A, Carlsson J, Gyllenstrand N, Lagercrantz U, Sundstrom JF. 2009. Non-radioactive in situ hybridization protocol applicable for Norway spruce and a range of plant species. Journal of Visualized Experiments .
Larsson E, Sitbon F, Ljung K, von Arnold S. 2008. Inhibited polar auxin transport results in aberrant embryo development in Norway spruce. New Phytologist 177, 356–366.
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Page 10 of 10 | Zhu et al.
Larsson E, Sitbon F, von Arnold S. 2012a. Differential regulation of Knotted1-like genes during establishment of the shoot apical meristem in Norway spruce (Picea abies). Plant Cell Reports 31, 1053–1060.
Larsson E, Sundstrom JF, Sitbon F, von Arnold S. 2012b. Expression of PaNAC01, a Picea abies CUP-SHAPED COTYLEDON orthologue, is regulated by polar auxin transport and associated with differentiation of the shoot apical meristem and formation of separated cotyledons. Annals of Botany 110, 923–934.
Laux T, Mayer KFX, Berger J, Jurgens G. 1996. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 122, 87–96.
Magyar Z, Horvath B, Khan S, Mohammed B, Henriques R, De Veylder L, Bako L, Scheres B, Bogre L. 2012. Arabidopsis E2FA stimulates proliferation and endocycle separately through RBR-bound and RBR-free complexes. EMBO Journal 31, 1480–1493.
Moschou PN, Smertenko AP, Minina EA, Fukada K, Savenkov EI, Robert S, Hussey PJ, Bozhkov PV. 2013. The caspase-related protease separase (EXTRA SPINDLE POLES) regulates cell polarity and cytokinesis in Arabidopsis. The Plant Cell 25, 2171–2186.
Nieuwland J, Scofield S, Murray JAH. 2009. Control of division and differentiation of plant stem cells and their derivatives. Seminars in Cell and Developmental Biology 20, 1134–1142.
Nystedt B, Street NR, Wetterbom A, et al. 2013. The Norway spruce genome sequence and conifer genome evolution. Nature 497, 579–584.
Palovaara J, Hallberg H, Stasolla C, Hakman I. 2010. Comparative expression pattern analysis of WUSCHEL-related homeobox 2 (WOX2) and WOX8/9 in developing seeds and somatic embryos of the gymnosperm Picea abies. New Phytologist 188, 122–135.
Potuschak T, Doerner P. 2001. Cell cycle controls: genome-wide analysis in Arabidopsis. Current Opinion in Plant Biology 4, 501–506.
Scheres B. 2007. Stem-cell niches: nursery rhymes across kingdoms. Nature Reviews Molecular Cell Biology 8, 345–354.
Scheres B, Dilaurenzio L, Willemsen V, Hauser MT, Janmaat K, Weisbeek P, Benfey PN. 1995. Mutations affecting the radial organization of the Arabidopsis root display specific defects throughout the embryonic axis. Development 121, 53–62.
Schneider CA, Rasband WS, Eliceiri KW. 2012. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9, 671–675.
Schoof H, Lenhard M, Haecker A, Mayer KF, Jurgens G, Laux T. 2000. The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100, 635–644.
Schwab R, Ossowski S, Riester M, Warthmann N, Weigel D. 2006. Highly specific gene silencing by artificial microRNAs in Arabidopsis. The Plant Cell 18, 1121–1133.
Smertenko AP, Bozhkov PV, Filonova LH, von Arnold S, Hussey PJ. 2003. Re-organisation of the cytoskeleton during developmental programmed cell death in Picea abies embryos. The Plant Journal 33, 813–824.
Tahir M, Law DA, Stasolla C. 2006. Molecular characterization of PgAGO, a novel conifer gene of the ARGONAUTE family expressed in
apical cells and required for somatic embryo development in spruce. Tree Physiology 26, 1257–1270.
Traas J, Bellini C, Nacry P, Kronenberger J, Bouchez D, Caboche M. 1995. Normal differentiation patterns in plants lacking microtubular preprophase bands. Nature 375, 676–677.
Ueda M, Laux T. 2012. The origin of the plant body axis. Current Opinion in Plant Biology 15, 578–584.
Ueda M, Zhang Z, Laux T. 2011. Transcriptional activation of Arabidopsis axis patterning genes WOX8/9 links zygote polarity to embryo development. Developmental Cell 20, 264–270.
Vandenberg C, Willemsen V, Hage W, Weisbeek P, Scheres B. 1995. Cell fate in the Arabidopsis root-meristem determined by directional signaling. Nature 378, 62–65.
van der Graaff E, Laux T, Rensing SA. 2009. The WUS homeobox-containing (WOX) protein family. Genome Biology 2009, 10:248.
Vestman D, Larsson E, Uddenberg D, Cairney J, Clapham D, Sundberg E, von Arnold S. 2011. Important processes during differentiation and early development of somatic embryos of Norway spruce as revealed by changes in global gene expression. Tree Genetics and Genomes 7, 347–362.
von Arnold S, Clapham D. 2008. Spruce embryogenesis. Methods in Molecular Biology 427, 31–47.
von Arnold S, Sabala I, Bozhkov P, Dyachok J, Filonova L. 2002. Developmental pathways of somatic embryogenesis. Plant Cell Tissue and Organ Culture 69, 233–249
Wang HC, Liu YD, Bruffett K, Lee J, Hause G, Walker JC, Zhang SQ. 2008. Haplo-insufficiency of MPK3 in MPK6 mutant background uncovers a novel function of these two MAPKs in Arabidopsis ovule development. The Plant Cell 20, 602–613.
Weigel D, Ossowski S, Schwab R. 2008. Gene silencing in plants using artificial microRNAs and other small RNAs. The Plant Journal 53, 674–690.
Weimer AK, Nowack MK, Bouyer D, Zhao X, Harashima H, Naseer S, De Winter F, Dissmeyer N, Geldner N, Schnittger A. 2012. RETINOBLASTOMA RELATED 1 regulates asymmetric cell divisions in Arabidopsis. The Plant Cell 24, 4083–4095.
Wenck AR, Quinn M, Whetten RW, Pullman G, Sederoff R. 1999. High-efficiency Agrobacterium-mediated transformation of Norway spruce (Picea abies) and loblolly pine (Pinus taeda). Plant Molecular Biology 39, 407–416.
Wu S, Scheible WR, Schindelasch D, Van Den Daele H, De Veylder L, Baskin TI. 2010. A conditional mutation in Arabidopsis thaliana separase induces chromosome non-disjunction, aberrant morphogenesis and cyclin B1;1 stability. Development 137, 953–961.
Wu X, Chory J, Weigel D. 2007. Combinations of WOX activities regulate tissue proliferation during Arabidopsis embryonic development. Developmental Biology 309, 306–316.
Wu XL, Dabi T, Weigel D. 2005. Requirement of homeobox gene STIMPY/WOX9 for Arabidopsis meristem growth and maintenance. Current Biology 15, 436–440.
Zuo J, Niu QW, Chua NH. 2000. Technical advance: an estrogen receptor-based transactivator XVE mediates highly inducible gene expression in transgenic plants. The Plant Journal 24, 265–273.
at Sveriges lantbruksuniversitet on September 17, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Top Related
Copyright © 2022 FDOKUMEN

