







Bahasa
Halaman
Hukum


Elsevier Editorial System(tm) for Systematic and Applied Microbiology Manuscript Draft Manuscript Number: SAM 3279R1 Title: Vibrio celticus sp. nov., a new Vibrio species belonging to the Splendidus clade with pathogenic potential for clams. Article Type: Full Length Papers Section/Category: Systematics Keywords: Vibrio celticus sp. nov. Splendidus clade MLSA phylogeny. Corresponding Author: Dr Jesus L. Romalde, Ph.D. Corresponding Author's Institution: Universidad de Santiago de Compostela First Author: Roxana Beaz-Hidalgo, Ph.D. Order of Authors: Roxana Beaz-Hidalgo, Ph.D.; Ana L Diéguez, M. Sc.; Ilse Cleewnwerck, Ph.D.; Sabela Balboa, M. Sc.; Alejandra Doce, M. Sc.; Paul de Vos, Ph.D.; Jesus L. Romalde, Ph.D. Abstract: A group of four motile facultative anaerobic marine isolates (Rd 8.15T [= CECT 7224T, = LMG 23850T], Rd 16.13, Rd 6.8 [= LMG 25696] and Rd2L5) were obtained from cultured clams (Ruditapes philippinarum and Venerupis pullastra) in Galicia, Northwestern Spain. They formed a tight phylogenetic group based on sequences of 16S rRNA gene and the four housekeeping genes rpoA (encoding the α−chain of RNA polymerase), rpoD (encoding the sigma factor of RNA polymerase), recA (encoding RecA protein), and atpA (encoding the α-subunit of bacterial ATP synthase). The phylogenies based on these sequences indicated that the four isolates represent a novel species in the genus Vibrio, and more precisely in the Splendidus clade. DNA-DNA hybridizations with the type strains of the species showing more than 98.6% 16S rRNA gene sequence similarity, revealed DNA-DNA relatedness below 70%. The isolates could be differentiated from the phylogenetically related Vibrio species on the basis of several phenotypic features. In addition, strain Rd 8.15T showed potential pathogenic activity for adult clam in the virulence assays. The name Vibrio celticus sp. nov. is proposed for this new taxon, with the type strain being Rd 8.15T (= CECT 7224T, = LMG 23850T).

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
1
2
3
Vibrio celticus sp. nov., a new Vibrio species belonging to the Splendidus clade with 4
pathogenic potential for clam. 5
6
7
Roxana Beaz-Hidalgo1†
, Ana L. Diéguez1, Ilse Cleenwerck
2, Sabela Balboa
1, 8
Alejandra Doce1, Paul de Vos
2 and Jesús L. Romalde
1*. 9
10
1 Departamento de Microbiología y Parasitología. Facultad de Biología. Universidad de 11
Santiago de Compostela. 15782, Santiago de Compostela. Spain. 12
2 BCCM/LMG Bacteria Collection, Laboratory of Microbiology, Ghent University, 13
Ghent, Belgium. 14
15
16
Submitted to: Systematic and Applied Microbiology. June 2010. 17
Revised Ms. SAM 3279 18
19
Running title: Vibrio celticus sp. nov. 20
21
* Corresponding author: 22
Phone: +34-981563100 # 16908 23
Fax: +34-981596904 24
E-mail: [email protected] 25
26
27
† Present address: Departamento de Ciencias Médicas Básicas. Universidad Rovira i 28
Virgili, Reus, Spain. 29
30
R.B.H. and A.L.D. have contributed equally to this work. 31
32
Manuscript

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
2
ABSTRACT 33
34
A group of four motile facultative anaerobic marine isolates (Rd 8.15T
[= CECT 7224T, 35
= LMG 23850T], Rd 16.13, Rd 6.8 [= LMG 25696] and Rd2L5) were obtained from 36
cultured clams (Ruditapes philippinarum and Venerupis pullastra) in Galicia, 37
Northwestern Spain. They formed a tight phylogenetic group based on sequences of 16S 38
rRNA gene and the four housekeeping genes rpoA (encoding the chain of RNA 39
polymerase), rpoD (encoding the sigma factor of RNA polymerase), recA (encoding 40
RecA protein), and atpA (encoding the -subunit of bacterial ATP synthase). The 41
phylogenies based on these sequences indicated that the four isolates represent a novel 42
species in the genus Vibrio, and more precisely in the Splendidus clade. DNA-DNA 43
hybridizations with the type strains of the species showing more than 98.6% 16S rRNA 44
gene sequence similarity, revealed DNA-DNA relatedness below 70%. The isolates 45
could be differentiated from the phylogenetically related Vibrio species on the basis of 46
several phenotypic features. In addition, strain Rd 8.15T showed potential pathogenic 47
activity for adult clam in the virulence assays. The name Vibrio celticus sp. nov. is 48
proposed for this new taxon, with the type strain being Rd 8.15T (= CECT 7224
T, = 49
LMG 23850T). 50
51
Keywords: Vibrio celticus sp. nov., Splendidus clade, MLSA, phylogeny. 52
53
54
55
56
57
58
59
60
61
62
63
64
65
66

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
3
INTRODUCTION 67
68
The genus Vibrio, ubiquitous in the aquatic environment, comprises nowadays more 69
than 70 bacterial species. These species have recently been classified into 14 clades 70
based on multilocus sequence analysis (MLSA) and other molecular techniques [27]. 71
Among these clades, the Splendidus clade contains the highest number of species (>10). 72
Moreover, the species of this clade are the dominant Vibrio species in coastal marine 73
sediments, seawater and bivalves in temperate climates [13, 28]. In addition, some of 74
these species have been associated with mortality of a wide range of marine animals 75
such as molluscs [7, 12, 16, 23, 30, 39], fish [11], shrimps [14] and octopus [5]. 76
77
The Splendidus clade is considered to be a not well resolve group. It shows much 78
phenotypical diversity, which makes species discrimination on the basis of biochemical 79
tests difficult, but also much genetic diversity. The latter was revealed by genotypic 80
studies based on ribotyping, amplified fragment length polymorphism (AFLP), PCR-81
restriction fragment length polymorphism [18, 34, 38], and analysis of housekeeping 82
gene sequences such as recA, gyrB or gapA [27]. On the contrary, 16S rRNA gene 83
sequences are extremely conserved within this group, being not useful for species 84
differentiation. In the last 10 years, up to nine new species have been described within 85
the Splendidus clade including V. crassostreae, V. chagasii, V. cyclitrophicus, V. 86
gallaecicus, V. gigantis, V. kanaloae, V. lentus, V. pomeroyi, and V. tasmaniensis [2, 6, 87
17, 35, 36]. A polyphasic approach including MLSA analysis, DNA-DNA hybridization 88
(DDH), chemotaxonomic techniques, and MALDI-TOFF-MS, is currently considered 89
an appropriate strategy to define new species of the genus Vibrio [27, 37]. 90
91
During a previous study on the diversity of vibrios associated with reared clams in 92
Galicia (Spain), a collection of 145 isolates were analysed by AFLP, a technique that 93
has been proven useful for rapid and reliable species identification and classification of 94
vibrios [1, 34] As a result of that analysis, a group of four strains (cluster 9) could not 95
be ascribed to any known species [1]. This study, describes in detail further taxonomic 96
characterization of these four strains obtained from clams, Ruditapes philippinarum and 97
Venerupis pullastra, cultured in Galicia, and proves they belong to a new species of 98
Vibrio. 99
100

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
4
MATERIALS AND METHODS 101
102
Bacterial strains and phenotypic tests. 103
During a sampling program performed in the years 2004 and 2005, a large collection of 104
marine bacterial isolaes were obtained from healthy clams, R. philippinarum (Adams & 105
Reeve, 1850) and V. pullastra (Montagu, 1803), cultured in different geographical sites 106
on the northwestern coast of Spain as previously described [1]. A representative number 107
of isolates from this collection (namely 145) were analyzed by AFLP and, in the present 108
study, a cluster with 4 strains that could not be identified, consisting of Rd 8.15T (= 109
CECT 7224T = LMG 23850
T), Rd 6.8 (= LMG 25696), and Rd 16.13 from R. 110
decussatus and Rd2L5 from V. pullastra [1], was further investigated. Reference strains 111
used in this study were obtained from Bacterial Culture Collections: V. chagasii LMG 112
21353T, V. crassostreae LMG 22240
T, V. cyclitrophicus, LMG 21359
T, V. gallaecicus 113
CECT 7244T, V. gigantis DSM 18531
T (= LMG 22741
T), V. kanaloae LMG 20539
T, V. 114
lentus CECT 5110T (= LMG 21359
T), V. pomeroyi LMG 20537
T, V. splendidus LMG 115
19031T, and V. tasmaniensis LMG 20012
T. All strains were cultured on plates of Marine 116
agar (MA; Pronadisa, Spain) at 24 ± 1ºC under aerobic conditions. Stock cultures were 117
maintained frozen at -80oC in Marine broth (MB; Oxoid Ltd., UK) supplemented with 118
15% glycerol (v/v). 119
120
The four marine strains from cluster 9 and the reference strains were subjected to the 121
following phenotypic tests as described previously [15, 20, 26, 40]: Gram stain, oxidase 122
activity, cell morphology and motility, oxidation/fermentation test, fermentation and 123
acid production from inositol, mannitol and sucrose, gas and acid production from 124
glucose, indole, methyl red, Voges-Proskauer reaction, utilization of citrate, 125
dihydrolation of arginine, decarboxylation of lysine and ornithine, nitrate reduction, 126
hydrolysis of gelatin, Tween 80, amylase, aesculin and alginate, DNase and urease 127
activities, use of 50 compounds as unique carbon source, salt tolerance tests (0, 0.5, 3, 6 128
and 8% of NaCl), growth at different temperatures (4ºC, 37ºC and 44ºC) and growth on 129
thiosulfate-citrate-bile salts sucrose (TCBS) agar (Oxoid). Sensitivity to the vibriostatic 130
agent O/129 (2,4-diamino-6,7-diisopropylpteridine) (150 µg per disc) and ampicillin (10 131
µg per disc) was determined in Müeller-Hinton (Oxoid) agar. Media were supplemented 132
with 1% NaCl when required. Additional phenotypic characteristics were determined 133
with API 20E and API ZYM (bioMerieux, France) using a saline solution (NaCl 0.85%) 134

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
5
for the bacterial suspension. The study of the acid production from 50 carbon sources 135
was tested using the API 50CH (bioMerieux) with slight modifications [2]. Briefly, 136
bacterial suspensions were prepared in saline solution, adjusted to an optical density of 137
1.0 at 580 nm and mixed (1:90, v/v) with the ZOF medium [15] for inoculation 138
procedures. 139
140
Sequencing of 16S rRNA and housekeeping genes and phylogenetic analyses. 141
Genomic DNA extraction, PCR amplification and sequencing of the 16S rRNA gene 142
and amplification of the genes recA, rpoA, rpoD atpA and pyrH (encoding uridine 143
monophosphate kinase) were carried out as previously described [24, 32, 33, 37, 41]. 144
For the sequencing reactions, the kit GenomeLab DTCS-Quick Start Kit (Beckman 145
Coulter, Germany) was used. Sequence corrections and analysis were performed with 146
DNAstar Seqman program (Lasergene, USA). Sequences of phylogenetically related 147
species were obtained after BLAST searches against the latest EBI releases. 148
Phylogenetic trees were constructed using three different methods, neighbor joining, 149
maximum parsimony (Mega version 4.0) [31] and maximum likelihood (jModelTest, 150
http://darwin.uvigo.es/, FigTree v1.1.2, http://tree.bio.ed.ac.uk/) [25]. 151
152
DNA-DNA hybridization. 153
Genomic DNA for DNA-DNA hybridization experiments and G+C content 154
determinations was extracted as previously described [3]. DNA-DNA hybridizations 155
were performed at 39ºC in a hybridization solution containing 50% of formamide, 156
according to a modification [9] of the microplate method described by Ezaki et al. [4]. 157
Reciprocal reactions (e.g. A xB and B x A) were performed and were generally within 158
the limits of this method [9]. DNA relatedness values (%) reported are the means of 159
minimum four hybridizations. Standard deviations, based on the reciprocal reactions, 160
are included. Additional reactions, including those between the strain Rd 8.15 T
against 161
the other strains in the cluster, were performed by the hydroxyapatite/microtitre plate 162
method [42] with a hybridization temperature (Tm) of 60ºC. The DNA G+C contents 163
were determined using HPLC as previously described [22]. 164
165
MALDI-TOF-MS 166
The protein analysis by MALDI-TOF-MS was performed in the mass unit of the 167
University of Santiago de Compostela. Protein extraction was performed with ethanol, 168

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
6
formic acid and acetonitrile (AN). Processed samples were placed in a 96 well plate, 169
allowed to dry and covered with a matrix solution (-cyano-4-hydroxycinnamic acid; 170
HCCA). Mass spectra were obtained using a MALDI-TOF Autoflex mass spectrometer 171
(Bruker Daltonik GmbH, Germany). The measured mass range of spectra was 2000-172
20,000 Da. Peak comparison was done with the data base of Bruker Daltonik GmbH. 173
The species-limit value considered was 2,300. Identification to genus level was in the 174
range 1,700-1,999. As a positive control, Escherichia coli CECT 433, was included in 175
the analysis and protein profiles were compared with their own profiles. Reproducibility 176
of the results was assessed by repetition of at least ten independent assays. 177
178
Pathogenicity tests. 179
Experimental infections in healthy clams (R. philippinarum and R. decussatus) were 180
performed with strain Rd 8.15T. The adult clams (mean size 30 mm) were kept for 2 181
days in tanks with aerated seawater (T = 19±1ºC; Sal. = 33%o) for acclimatization. 182
Subsequently, three groups of 20 clams of each species were used for the infection 183
experiments. Two groups were bath challenged for 3 h in non-circulating seawater 184
conditions with two different bacterial doses (104 and 10
6 CFU/ml), transferred to 185
empty tanks for 1 h and, finally, to the tanks containing aerated seawater at 19±1ºC. The 186
third group was treated in the same way but adding sterile seawater instead of the Vibrio 187
strain. Clams were monitored for mortality over a 14-day period. Mortalities were 188
attributed to the strains Rd 8.15T only if it could be re-isolated from the tissues of the 189
dead clams. 190
191
RESULTS AND DISCUSSION 192
193
The four clam strains shared the properties of the genus Vibrio. They were motile, 194
facultative anaerobic, Gram-negative and oxidase positive, susceptible to the 195
vibriostatic agent O/129 (150 µg per disc), and capable to reduce nitrates to nitrites. 196
Cells of strains were large regular rods of variable size, 1.3-1.5 µm long and 0.78-0.81 197
µm wide. The four strains showed high phenotypical homogeneity, although variable 198
reactions were observed for the use of D-fructose, D-mannose, D-lactose, L-fucose, 199
gluconic acid, citric acid, trans-aconitic acid, L-leucine, L-arginine, and L-histidine as 200
sole carbon source, and the acid production from L-arabinose, D-melibiose and 201
potassium gluconate (Table S2). The four clam strains showed alkaline phosphatase, 202

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
7
esterase, esterase lipase, leucine arylamidase, trypsin, and acid phosphatase activities in 203
the API ZYM tests. Variable results were obtained for valine arylamidase (negative for 204
strain Rd2L5), and Naphthol-AS-BI- phosphohydrolase (negative for strains Rd2L5 and 205
Rd 8.15T). 206
207
Nearly complete 16S rRNA gene sequences were determined for the four clam strains. 208
Phylogenetic analysis based on these sequences, using the neighbor joining, maximum 209
parsimony (data not shown) and maximum likelihood methods, confirmed their position 210
in the genus Vibrio, and allocated them to the Splendidus clade (Figs. 1 and S1). The 211
four strains showed 99.9-100% 16S rRNA gene sequence similarity to each other, and 212
more than 98.6% similarity [29] with eight species of the Splendidus clade (Table S1). 213
These results support those previously reported by other authors [2, 6, 10, 19, 30, 34] on 214
the extreme conservation of the 16S rRNA gene within the Splendidus clade, and its 215
uselessness for differentiation among these species. 216
217
MLSA has been proposed as a valuable technique for the identification of vibrios, and 218
for studying their phylogeny [2, 4, 17, 37]. Partial sequences of rpoA, rpoD, recA, atpA, 219
and pyrH were determined for the four strains, and sequence similarities above 99.9%, 220
99.6%, 98.7%, 99.9% and 96.4% were obtained for each of these genes, respectively 221
(Table S1). As these values are higher than the limits for species delineation previously 222
described for these genes [37], these results indicated that the four strains probably 223
represent a single novel species in the genus Vibrio, supporting hereby the previous 224
AFLP data. Phylogenetic trees based on the housekeeping genes recA, rpoA, rpoD, and 225
atpA separately (Fig. S2), confirm the close relationship of the four clam strains. In 226
these trees, the strains form a tight group, whereas in the pyrH gene gene sequence 227
based tree the four strains are split over two subgroups (Fig. S2e). The latter tree 228
topology can be explained by recombination events, which have been detected using the 229
RDP3 beta41 software [8, 21]. 230
Phylogenetic trees based on concatenated sequences (4060 nt) of the 16S rRNA gene 231
and the 4 housekeeping genes recA, rpoA, rpoD, and atpA also confirmed the close 232
relationship between the four strains, and support their distinction from the closest 233
phylogenetic neighbours based on 16S rRNA gene sequences (Fig. S3). When pyrH 234
gene sequences were included (concatenated sequences of 4477 nt), the tight 235
relationship among the four clam strains was confirmed but the tree topology was 236

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
8
somewaht distorted (Figs. 2 and S4) because of the detected recombination events in the 237
pyrH gene. 238
239
Levels of DNA relatedness were determined between the four clam strains, and between 240
strain Rd 8.15T and the type strains of the Splendidus clade to which it showed more 241
than 98.5% 16S rRNA sequence similarity [29]. The DNA relatedness values among the 242
four clam strains ranged from 88% to 94.2%, while DNA relatedness values between 243
strain Rd 8.15T and the tested type strains were below 70%, namely, 52±19% with V. 244
crassostreae LMG 22240T, 55±4% with V. cyclitrophicus LMG 21359
T, 50±13% with 245
V. gigantis LMG 22741T, 63±6% with V. kanaloae LMG 20539
T, 50±18% with V. 246
lentus LMG 21359T, 46±8% with V. pomeroyi LMG 20537
T, 53±2% with V. 247
psplendidus LMG 19031T, and 46±8% with V. tasmaniensis LMG 20012
T. These values 248
are in the range of those obtained between other species of the Splendidus clade [2, 6, 249
17, 19, 35, 36], and prove that the four strains represent a single novel species in the 250
genus Vibrio. The DNA G+C content of strain Rd 8.15T is 46.6 mol%. 251
252
Several differentiating phenotypic characteristics were found between the presumptive 253
novel species and closely related Vibrio species (Table 1). Useful phenotypic traits to 254
differentiate the four clam isolates from other species within the Splendidus clade 255
include the positive reaction for the Voges Proskauer test, their ability to produce acid 256
from L-rhamnose and D-lactose and their inability to produce acid from potassium 257
gluconate or to use N-acetyl-D-glucosamine as a sole carbon source. 258
259
MALDI-TOFF-MS for strain Rd 8.15T resulted in a protein profile that was different 260
from the profiles of the type strains of closely related species present in the data base 261
(Fig. S5). The closest species defined by MALDI-TOF-MS were V. gigantis (score = 262
2,140) and V. pomeroyi (score = 2,000). For the other species of the Splendidus clade 263
values ranged from 1,403 and 1,885. A correct identification of E. coli CECT 433 was 264
obtained, with profiles always having a score above 2,300. 265
266
In the virulence assays, strain Rd 8.15T produced cummulative mortalities of 100% of 267
the inoculated adult clams, regardless the dose assayed or the bivalve species (R. 268
philippinarum or R. decussatus). In the control tanks mortalities were always lower than 269
15%. Strain Rd 8.15T were re-isolated from all the dead inoculated animals, but not 270

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
9
from dead control clams. These results suggest the pathogenic potential of strain Rd 271
8.15T for clams. 272
273
In conclusion, this polyphasic study clearly demonstrates that the four isolates represent 274
an undescribed species of the genus Vibrio potentially pathogenic for clam. The name 275
Vibrio celticus sp. nov. is proposed with Rd 8.15T (=CECT 7224
T =LMG 23850
T) as the 276
type strain. 277
278
279
280
Description of Vibrio celticus sp nov. 281
Vibrio celticus [cel’ti.cus. L. masc. adj. celticus, pertaining to the celtics, here pertaining 282
to the pre-romanic inhabitants of Galicia]. 283
284
Cells are Gram-negative rods, motile and facultative anaerobic. Colonies are round with 285
smooth margins, beige in color and non-swarming on MA plates. Strains grow on TCBS 286
agar forming yellow colonies after incubation for 1 day at 24ºC. Glucose metabolism is 287
fermentative without gas production. Strains reduce nitrates to nitrites. Oxidase and 288
catalase tests are positive. Strains are sensitive to the vibriostatic agent O/129 (150µg 289
per disc). All strains grow at 4ºC and require NaCl for growth. The optimal NaCl 290
concentration for growth is 1 to 3% (wt/vol) and strains are able to grow at 6% NaCl but 291
not at higher salinity concentrations. Not luminescent. All strains are positive for 292
arginine dihidrolase, and negative for lysine and ornitine decarboxylases, ONPG, use of 293
citrate and hydrolysis of urea. Indole, Voges-Proskauer, and methyl red reactions are 294
positive. All strains hydrolyse starch, gelatin, aesculin and Tween 80. Acid is produced 295
from D-lactose and L-rhamnose, but not from potassium gluconate (except strain 296
Rd2L5). None of the strains utilized N-acetyl-D-glucosamine as sole carbon source. The 297
strains showed alkaline phosphatase, esterase, esterase lipase, leucine arylamidase, 298
trypsin and acid phosphatase activities. Other properties are given in Tables 1 and S2. 299
300
The type strain Rd 8.15T (=CECT 7224
T, =LMG 23850
T) was isolated from clam 301
Ruditapes philippinarum in northwestern coast of Spain. Its DNA G+C content is 44.6 302
mol%. 303
304

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
10
305
The GenBank accession numbers for the 16S rRNA, recA, rpoA, rpoD, atpA and pyrH 306
gene sequences of Vibrio celticus sp. nov. strains, for rpoD gene sequence of V. 307
gallaecius CECT 7244T, and for pyrH gene sequence of V. kanaloae 20539
T are listed 308
in Supplementary Table S3. 309
310
311
ACKNOWLEDGEMENTS 312
This work was supported in part by a grants AGL2003-09307-C02-01 and AGL2006-313
13208-C02-01, from the Ministerio de Ciencia y Tecnología and grant 314
PGIDIT04PXIC20001PN from the Xunta de Galicia (Spain). R.B.H. and S.B. 315
acknowledge the Ministerio de Ciencia y Tecnología (Spain), and A.D. the Xunta de 316
Galicia (Spain) for research fellowships. The authors thank E. Guitián from the RIAIDT 317
(USC) for the mass spectrometry analysis and Bruker Daltonik GmbH for the data-base 318
availability. The resarch was also supported bt the Prime Minister’s Services, Federal 319
Office for Scientific, Technical and Culture Affairs, Belgium. The authors wish to thank 320
Katrien Engelbeen for performing the DNA-DNA hybridizations. 321
322
323

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
11
REFERENCES 324
325
[1] R. Beaz-Hidalgo, I. Cleenwerck, S. Balboa, M. De Wachter, F. Thompson, J. 326
Swings, P. De Vos, J.L. Romalde, Diversity of vibrios associated with reared clams 327
in Galicia (NW of Spain). Syst. Appl. Microbiol. 31 (2008) 215-222. 328
[2] R. Beaz-Hidalgo, A. Doce, J. Pascual, A.E.Toranzo, J.L.Romalde, Vibrio 329
gallaecicus sp. nov. isolated from cultured clams in north-western Spain. Syst. Appl. 330
Microbiol. 32 (2009) 111–117. 331
[3] I. Cleenwerck, K. Vandemeulebroecke, D. Janssens, J. Swings, Re-examination of 332
the genus Acetobacter, with descriptions of Acetobacter cerevisiae sp. nov. and 333
Acetobacter malorum sp. nov. Int. J. Syst. Bacteriol. 52 (2002) 1551-1558. 334
[4] T. Ezaki, Y. Hashimoto, E. Yabuuchi, Fluorometric deoxyribonucleic acid-335
deoxyribonucleic acid hybridization in microdilution wells as an alternative to 336
membrane filter hybridization in which radioisotopes are used to determine genetic 337
relatedness among bacterial strains. Int. J. Syst. Bacteriol. 39 (1989) 224-229. 338
[5] R. Farto, S.P. Armada, M. Montes, J. Guisande, M.J. Pérez, T.P. Nieto, Vibrio lentus 339
associated with diseased wild octopus (Octopus vulgaris). J Invert. Pathol. 83 340
(2003)149-156. 341
[6] N. Faury, D.Saulnier, F.L. Thompson, M. Gay, J. Swings, F. Le Roux, Vibrio 342
crassostreae sp. nov isolated from the haemolymph of oysters (Crassostrea gigas). 343
Int. J. Syst. Evol. Microbiol. 54 (2004) 2137-2140. 344
[7] M. Gay, T. Renault, A. Pons, F. Le Roux, Two Vibrio splendidus related strains 345
collaborate to kill Crassostrea gigas: taxonomy and host alterations. Dis. Aquat. 346
Org. 62 (2004) 65-74. 347
[8] M.J. Gibbs, J.S. Armstrong, A.J. Gibbs, Sister-scanning: a Monte Carlo procedure 348
for assessing signals in recombinant sequences. Bioinformatics 16 (2000) 573-582. 349
[9] J. Goris, K. Suzuki, P. De Vos, T. Nakase, K. Kersters, Evaluation of a microplate 350
method DNA-DNA hybridization method compared with the initial renaturation 351
method. Can. J. Microbiol. 44 (1998) 1148-1153. 352
[10] B.P. Hedlund, J.T. Staley, Vibrio cyclotrophicus sp. nov., a polycyclic aromatic 353
hydrocarbon (PAH) degrading marine bacterium. Int. J. Syst. Evol. Microbiol. 51 354
(2001) 61-66. 355
356
357

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
12
[11] S. Jensen, O.B. Samuelsen, K. Andersen, L. Torkildsen, C. Lambert, G. Choquet, 358
C. Paillard, O. Berg, Characterization of strains of Vibrio splendidus and V. tapetis 359
isolated from corwing wrasse Symphodus melops suffering vibriosis. Dis. Aquat. 360
Org. 53 (2003) 25-31. 361
[12] A. Lacoste, S. Jalabert, S. Malham, A. Cueff, F. Gelebart, C. Cordevant, M. Lange, 362
S. A. Poulet, A Vibrio splendidus strain is associated with summer mortality of 363
juvenile oysters Crassostrea gigas in the Bay of Morlaix (North Brittany, France). 364
Dis. Aquat. Org. 46 (2001) 139-145. 365
[13] C. Lambert, J.L. Nicolas, V. Cilia, S. Corre, Vibrio pectenicida sp. nov., a 366
pathogen of scallop (Pecten maximus) larvae. Int. J. Syst. Bacteriol. 48 (1998) 481-367
487. 368
[14] E.M. Leano, C.R. Lavilla-Pitogo, M.G. Paner, Bacterial flora in the hepatopancreas 369
of pond reared Penaeus monodon juveniles with luminous vibriosis. Aquaculture 370
164 (1998) 367-374. 371
[15] M.L. Lemos, A.E. Toranzo, J.L. Barja, Modified medium for the oxidation-372
fermentation test in the identification of marine bacteria. Appl. Environ. Microbiol. 373
49 (1985) 1541-1543. 374
[16] F. Le Roux, M. Gay, C. Lambert, M. Waechter, S. Poubalanne, B. Chollet, J.L. 375
Nicolas, F. C. J. Berthe, Comparative analysis of Vibrio splendidus- related strains 376
isolated during Crassostreae gigas mortality events. Aquat. Living Resour. 15 (2002) 377
251-258. 378
[17] F. Le Roux, A. Goubet, F.L. Thompson, N. Faury, M. Gay, J. Swings, D. Saulnier, 379
Vibrio gigantis sp. nov. isolated from the haemolymph of cultured oysters 380
(Crassostrea gigas). Int. J. Syst. Evol. Microbiol. 55 (2005) 2251-2255. 381
[18] M.C. Macián, E. Garay, F. Gonzalez- Candelas, M.J. Pujalte, R. Aznar, Ribotyping 382
of vibrio populations associated with cultured oysters (Ostrea edulis). Sys. Appl. 383
Microbiol. 23 (2000) 409-417. 384
[19] M.C. Macián, W. Ludwig, R. Aznar, P.A.D Grimont, K.H. Schleifer, E. Garay, M.J. 385
Pujalte, Vibrio lentus sp. nov., isolated from Mediterranean oysters. Int. J. Syst. 386
Evol. Microbiol. 51 (2001) 1449-1456. 387

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
13
[20] J.F. MacFaddin, Pruebas bioquímicas para la identificación de bacterias de 388
importancia clínica. The William & Wilkins Company. Baltimore. (1993). 389
Translation by Médica Panamericana S.A. 390
[21] D.P. Martin, C. Williamson, D. Posada, RDP2: recombination detection and 391
analysis from sequence alignments. Bioinformatics 21(2005) 260-262. 392
[22] M. Mesbah, U. Premachandran, W.B. Whitman, Precise measurement of the G+C 393
content of deoxyribonucleic acid by high-performance liquid chromatography. Int. J. 394
Syst. Bacteriol. 39 (1989) 159-167. 395
[23] J.L. Nicolas, S. Corre, G. Gauthier, R. Robert, D. Ansquer, Bacterial problems 396
associated with scallop Pecten maximus larval culture. Dis. Aquat. Org. 27 (1996) 397
67-76. 398
[24] C.R. Osorio, M.D. Collins, A.E. Toranzo, J.L. Romalde, 16S rRNA gene sequence 399
analysis of Photobacterium damselae and nested-PCR method for rapid detection of 400
fish pasteurellosis. Appl. Environ. Microbiol. 65 (1999) 2942-2946. 401
[25] D. Posada, jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 25 (2008) 402
1253-1256. 403
[26] J.L. Romalde, A.E. Toranzo, Evaluation of the API 20E system for the routine 404
diagnosis of the enteric redmouth disease. Bull. Eur. Ass. Fish. Pathol. 11 (1991) 405
147-149. 406
[27] T. Sawabe, K. Kita-Tsukamoto, F.L. Thompson, Inferring the evolutionary history 407
of vibrios by means of multilocus sequence analysis. J. Bacteriol. 189 (2007) 7932-408
7936. 409
[28] P.A. Sobecky, T.J. Mincer, M.C. Chang, A. Toukdarian, D.R. Helinski, Isolation of 410
broad- host- range replicons from marine sediment bacteria. Appl. Environ. 411
Microbiol. 64 (1998) 2822-2830. 412
[29] E. Stackebrandt, J. Ebers, Taxonomic parameters revisited: tarnished gold 413
standards. Microbiol. Today. 33 (2006) 152-155. 414
[30] G. Sugumar, T. Nakai, Y. Hirata, D. Matsubara, K. Muroga, Vibrio splendidus 415
biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea 416
gigas larvae. Dis. Aquat. Org. 33 (1998) 111-118. 417
[31] K. Tamura, J. Dudley, M. Nei, S. Kumar, MEGA4: Molecular Evolutionary 418
Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24 (2007) 1596-419
1599. 420

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
14
[32] C.C. Thompson, F.L. Thompson, K. Vandemeulebroecke, B. Hoste, P. Dawyndt, J. 421
Swings Use of recA as an alternative phylogenetic marker in the family 422
Vibrionaceae. Int. J. Syst. Evol. Microbiol. 54 (2004) 919-929. 423
[33] C.C. Thompson, F.L. Thompson, A.C. Vicente. J. Swings, Phylogenetic analysis of 424
vibrios and related species by means of atpA gene sequences. Int. J. Syst. Evol. 425
Microbiol. 57 (2007) 2480-2484. 426
[34] F.L. Thompson, B. Hoste, K. Vandemeulebroecke, J. Swings, Genomic diversity 427
amongst Vibrio isolates from different source determined by fluorescent amplified 428
fragment length polymorphism. System. Appl. Microbiol. 24 (2001) 520-538. 429
[35] F.L. Thompson, C.C. Thompson, J. Swings, Vibrio tasmaniensis sp. nov. isolated 430
from Atlantic salmon (Salmo salar L.). Syst. Appl. Microbiol. 26 (2003a) 65-69. 431
[36] F.L. Thompson, C.C. Thompson, Y. Li, B. Gomez-Gil, J. Vanderberghe, B. Hoste, 432
J. Swings, Vibrio kanaloae sp. nov., Vibrio pomeroyi sp. nov. and Vibrio chagasii 433
sp. nov., from sea water and marine animals. Int. J. Syst. Evol. Microbiol. 53 (2003b) 434
753-759. 435
[37] F.L. Thompson, D. Gevers, C.C. Thompson, P. Dawyndt, S. Naser, B. Hoste, C.B. 436
Munn, J. Swings, Phylogeny and molecular identification of Vibrios on the basis of 437
Multilocus Sequence Analysis. Appl. Environ. Microb. 71 (2005) 5107-5115. 438
[38] H. Urakawa, K. Kita-Tsukamoto, K. Ohwada, 16S rDNA restriction fragment 439
length polymorphism analysis of psychrotophic vibrios from Japanese coastal water. 440
Can. J. Microbiol. 45 (1999) 1001-1007. 441
[39] M. Waechter, F. Le Roux, J.L. Nicolas, E. Marissal, F. Berthe, Characterization of 442
pathogenic bacteria of the cupped oyster Crassostrea gigas. C. R. Biol. 325 (2002) 443
231-238. 444
[40] P.A. West, P. R. Brayton, T.N. Bryant, R.R. Colwell, Numerical taxonomy of 445
vibrios isolated from aquatic environments. Int. J. Syst. Bacteriol. 36 (1986) 531-446
543. 447
[41] S. Yamamoto, H. Kasai, D.L. Arnold, R.W. Jackson, A. Vivian, S. Harayama. 448
Phylogeny of the genus Pseudomonas: intrageneric structure reconstructed from the 449
nucleotide sequences of gyrB and rpoD genes. Microbiology 146 (2000) 2385-2394. 450
[42] F. Ziemke, M.G. Hofle, J. Lalucat, R. Roselló-Mora, Reclassification of 451
Shewanella putrefaciens Owen´s genomic group II as Shewanella baltica sp. nov. 452
Int. J. Syst. Bacteriol. 48 (1998) 179-186. 453

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49
15
Table 1. Phenotypic characteristics for distinguishing V. celticus from phenotypically and phylogenetically related Vibrio species.
Characteristic 1 2 3 4 5 6 7 8 9 10 11
ADH + + – + + – – + + – –
ONPG* – – + – + – + – – + –
Voges Proskauer + – – – – + + – – – –
Acid from:
L-rhamnose*
D-lactose*
Potassium gluconate*
+
+
V– (1)
–
–
+
+
–
+
–
–
+
–
+
+
–
–
+
–
–
–
–
–
+
–
ND
ND
–
+
+
–
–
+
Susceptibility to:
O/129 (150µg)
+
+
+
+
+
+
–
V
+
+
+
Growth at:
35ºC
6% NaCl
8% NaCl
–
+
–
–
–
–
V
–
V
–
+
–
–
+
+
+
V
–
–
+
–
–
+
V
–
+
V
+
+
+
–
–
–
Hydrolysis of:
Gelatin
Tween 80*
+
+
+
+
+
+
+
+
+
–
–
–
+
+
+
+
+
+
+
+
+
+
Use as sole carbon source of:
N-acetyl-D-glucosamine
–
+
ND
+
+
+
ND
+
ND
+
ND
+, positive; –, negative; V, variable; V–, variable but the type strain is negative. The number in parenthesis indicates the number of strains giving a positive
result. Taxa are indicated as: 1, V. celticus (4 strains); 2. V. gigantis DSM 18531T; 3, V. splendidus, CECT 528
T; 4, V. crassostreae, LMG 22240
T; 5, V,
pomeroyi, LMG 20537T; 6, V. tasmaniensis, LMG 20012
T; 7, V. lentus CECT 5110
T; 8, V. kanaloae LMG 20539
T; 9, V. chagasii LMG 21353
T; 10, V.
cyclitrophicus, LMG 21359T; 11, V. gallaecicus LMG 7244
T. Data for the reference strains are taken from the literature [2, 4, 10, 17, 19, 35, 36], except when
indicated by *. Those data derive from tests performed in the frame of this study on the type strains. All taxa are negative for lysine and ornithine
decarboxylase, acid from inositol and D–sorbitol. All taxa are positive for indole and hydrolysis of starch.

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
16
Figure legends
Fig. 1. Phylogenetic tree based on partial 16S rRNA gene sequences, and obtained by
the Neighbor joining method. Vibrio cholerae ATCC 14035T
was used as out–group.
GeneBank sequence accession numbers are given in parentheses. Numbers at the nodes
show the percentages bootstrap values.
Fig. 2. Phylogenetic tree based on concatenated sequences of the housekeeping genes
recA, rpoA, rpoD, atpA, and pyrH, and the 16S rRNA gene, and obtained by the
Neighbor joining method. Numbers at the nodes show the percentages bootstrap values.
GeneBank sequence accession numbers for individual gene sequences are given in
Table S3 and Fig. S2.

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
17
Fig. 1.- Beaz-Hidalgo et al.

1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65
18
Fig. 2.- Beaz-Hidalgo et al.

Vibrio celticus sp. nov., a new Vibrio species belonging to the
Splendidus clade with pathogenic potential for clams.
Roxana Beaz-Hidalgo1†
, Ana L. Diéguez1, Ilse Cleenwerck
2, Sabela Balboa
1,
Alejandra Doce1, Paul de Vos
2 and Jesús L. Romalde
1*.
Supplementary Material
Supplementary Material
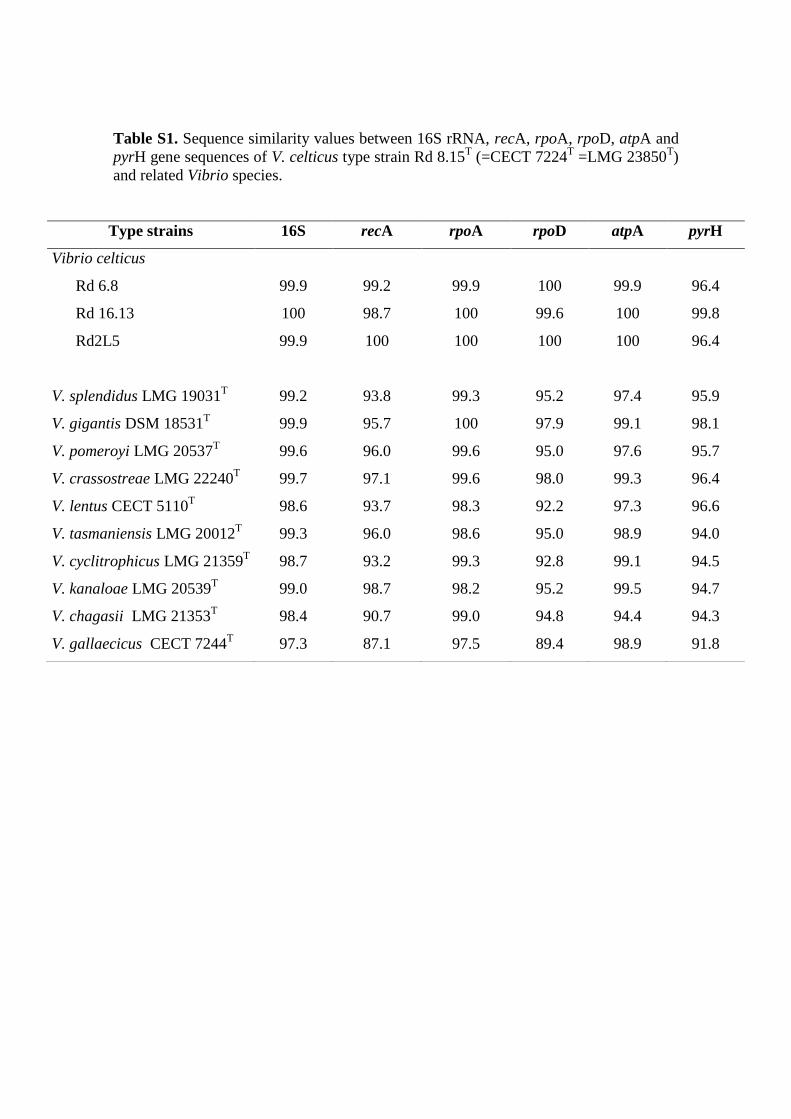
Table S1. Sequence similarity values between 16S rRNA, recA, rpoA, rpoD, atpA and
pyrH gene sequences of V. celticus type strain Rd 8.15T (=CECT 7224
T =LMG 23850
T)
and related Vibrio species.
Type strains 16S recA rpoA rpoD atpA pyrH
Vibrio celticus
Rd 6.8
Rd 16.13
Rd2L5
V. splendidus LMG 19031T
V. gigantis DSM 18531T
V. pomeroyi LMG 20537T
V. crassostreae LMG 22240T
V. lentus CECT 5110T
V. tasmaniensis LMG 20012T
V. cyclitrophicus LMG 21359T
V. kanaloae LMG 20539T
99.9
100
99.9
99.2
99.9
99.6
99.7
98.6
99.3
98.7
99.0
99.2
98.7
100
93.8
95.7
96.0
97.1
93.7
96.0
93.2
98.7
99.9
100
100
99.3
100
99.6
99.6
98.3
98.6
99.3
98.2
100
99.6
100
95.2
97.9
95.0
98.0
92.2
95.0
92.8
95.2
99.9
100
100
97.4
99.1
97.6
99.3
97.3
98.9
99.1
99.5
96.4
99.8
96.4
95.9
98.1
95.7
96.4
96.6
94.0
94.5
94.7
V. chagasii LMG 21353T 98.4 90.7 99.0 94.8 94.4 94.3
V. gallaecicus CECT 7244T 97.3 87.1 97.5 89.4 98.9 91.8

Table S2.- Variable phenotypic characteristics of Vibrio celticus sp. nov.a
a Acid is produced from: D-glucose, D-fructose, D-galactose, D-mannose, D-ribose, D-mannitol, glycerol,
N-acetyl-glucosamine, D-cellobiose, D-maltose, D-trehalose, glycogen, starch, D-sucrose, D-lactose, L-
rhamnose, and D-xylose. None of the strains produce acid from erythritol, D-arabinose, L-xylose, D-
adonitol, methyl-D-xylopranoside, L-sorbose, dulcitol, inositol, D-sorbitol, methyl-D-
mannopyranoside, methyl-D-glucopyranoside, amygdalin, arbutin, aesculin, salicin, inulin, D-
melezitose, D-raffinose, xylitol, gentiobiose, D-turanose, D-lyxose, D-tagatose, D-fucose, L-fucose, D-
arabitol, L-arabitol, potassium 2-ketogluconate and potassium 5-ketogluconate. All strains utilized as sole
carbon source D-glucose, D-trehalose, D-maltose, D-cellobiose, D-sucrose, D-mannitol, glycerol, sodium
acetate, propionic acid, succinic acid, glycine, L-serine, L-threonine, L-glutamic acid, D-alanine, L-
aspartic acid, -ketoglutaric acid, fumaric acid, and N-acetyl-D-glucosamine, but not D-ribose, L-
arabinose, D-xylose, D-galactose, L-rhamnose, D-melibiose, D-salicin, D-amygdaline, D-glucuronic acid,
D-galacturonic acid, D-sorbitol, myo-inositol, butiric acid, L-tirosine, L-lysine, L-ornitine, L-citruline,
amino-N-butiric acid, and N-acetyl-D-glucosamine.
TEST Strain
Rd 8.15T Rd 6.8 Rd 16.13 Rd2L5 Production of acid from:
L- arabinose – – – + D-melibiose + – – – Potassium gluconate – – – +
Use as sole carbon source: + + + + D-fructose + + – + D-mannose + + – + D-lactose – – – + L-fucose – – – + Gluconic acid – – – + Citric acid + + – + Trans-aconitic acid + – – – L-leucine – – – + L-arginine + – + + L-histidine – – – +

Table S3. GenBank accession numbers of the 16S rRNA, recA, rpoA, rpoD, atpA, and
pyrH gene sequences of the four strains of Vibrio celticus sp. nov. investigated in this
study.
Strain 16rRNA recA rpoA rpoD atpA pyrH
RD 8.15T EF599162 EU541590 EU541570 FN582243 FN582232 FN582244
Rd6.8 FN582228 FN582236 FN582237 FN582242 FN582231 FN582245
Rd16.13 FN582227 FN582234 FN582239 FN582240 FN582230 FN582247
Rd2L5 FN582229 FN582235 FN582238 FN582241 FN582233 FN582246
V. gallaecicus
CECT 7244T
FN908850
V. kanaloae
LMG 20539T
FN908851

Fig. S1. Phylogenetic tree based on partial 16S rRNA gene sequences, and obtained by
the maximum likelihood method (GTR+I+G model). The Vibrio choleare ATCC
14035T
was used as out-group. GeneBank sequence accession numbers are given in
parenthesis. Numbers above the branches show the phylogenetic distance.
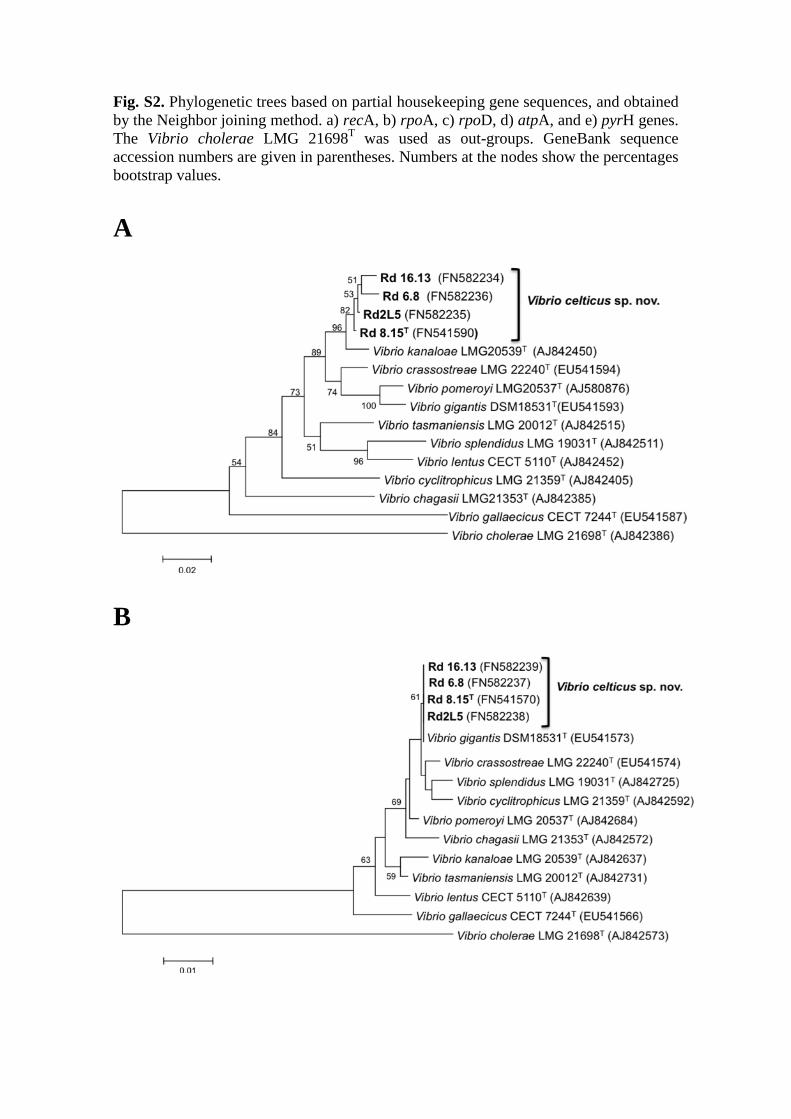
Fig. S2. Phylogenetic trees based on partial housekeeping gene sequences, and obtained
by the Neighbor joining method. a) recA, b) rpoA, c) rpoD, d) atpA, and e) pyrH genes.
The Vibrio cholerae LMG 21698T was used as out-groups. GeneBank sequence
accession numbers are given in parentheses. Numbers at the nodes show the percentages
bootstrap values.
A
B
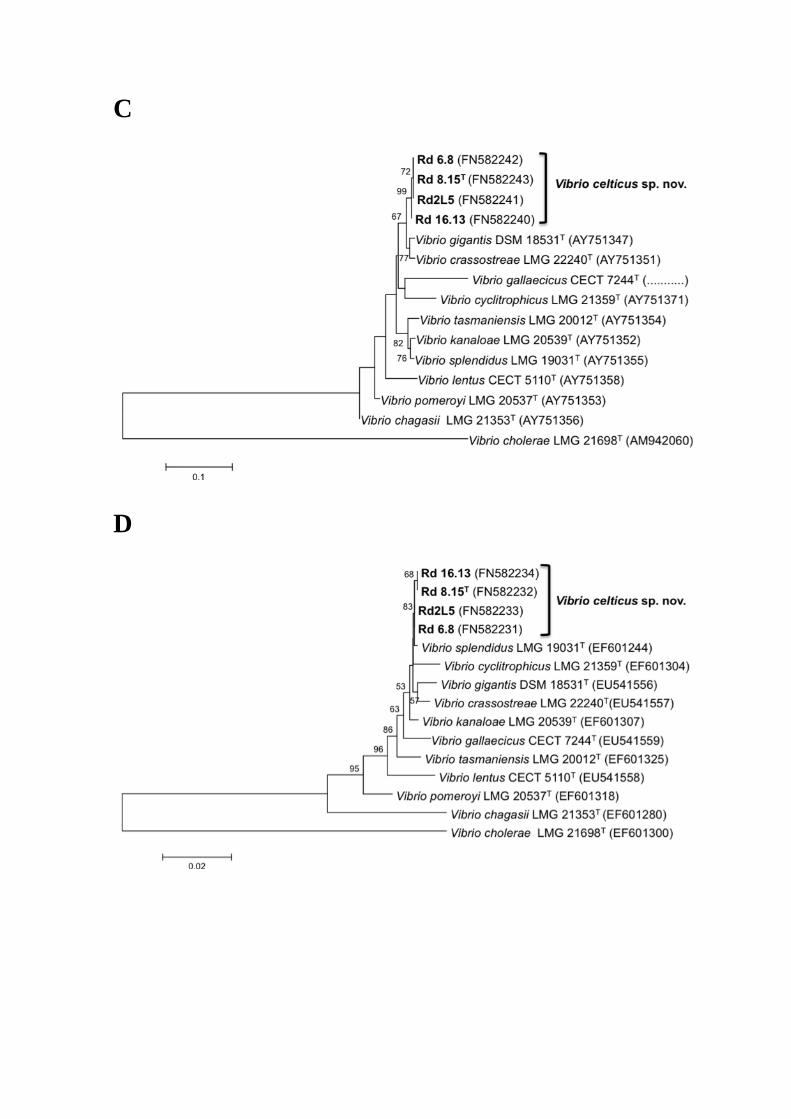
C
D

E

Fig. S3. Phylogenetic tree based on concatenated sequences of the housekeeping genes,
recA, rpoA, rpoD, and atpA and the 16S rRNA gene, and obtained by Neighbor Joining
(A) the maximum likelihood (GTR+I+G model)(B) methods. GeneBank accession
numbers of individual gene sequences are given in Table S3 and Fig. S2. Numbers in
nodes in A indicate bootstrap values and above branches in B show the phylogenetic
distance.
A
B

Fig. S4. Phylogenetic tree based on concatenated sequences of the housekeeping genes,
recA, rpoA, rpoD, atpA, and pyrH and the 16S rRNA gene, and obtained by the
maximum likelihood method (GTR+I+G model). GeneBank accession numbers of
individual gene sequences are given in Table S3 and Fig. S2. Numbers above branches
show the phylogenetic distance.

Fig S5. Protein profile of type strain of V. celticus sp. nov., Rd 8.15T obtained by the
technique MALDI-TOF-MS.

Dr. Ramón Roselló-Mora
Executive editor
Systematic and Applied Microbiology
Dear Dr. Roselló-Mora:
Querido Ramón:
Thank you very much for the useful comments on the manuscript SAM 3279 entitled
“Vibrio celticus sp. nov., a new Vibrio species belonging to mthe Splendidus clade qith
pathogenic potential for clams", by Beaz-Hidalgo et al. We are submitting a revised
version, which was modified taking into account the reviewers’ suggestions and your
comments.
Answers to editor:
1. The meaning of the adjective “celticus –a –um” was changed according to your
suggestions to .”cel’ti.cus. L. masc. adj. celticus, pertaining to the celtics, here
pertaining to the pre-romanic inhabitants of Galicia”
Answers to reviewer # 1:
1. Vibrio kanaloae, V. chagasii and V. gallaecicus were included in MLSA
analysis and the correspondent independent and concatenated trees. New
sequences were obtained for some genes and deposited in the GenBank, and the
accession numbers included in Table S3. To avoid a hugh table, the inclusion of
all the accession numbers was not perfoemed. Instead size source in tress was
increased to make easy their reading.
2. The number of Vibrio species was modified (p. 3, line 71) and V. cyclitrophicus
and V. lentus included in the paragraph (p. 3, lines 86-88).
3. The list of genes was corrected (p. 7, line 218).
4. Line 258 (p. 8), the sentence was modified to clarify its meaning.
*Detailed Response to Reviewers

5. At the end of the description, reference was made to the tables to complete the
characteristics of the new species.
6. Table 1 footnote was corrected.
7. The reviewer was right indicating the mistakes in the similarities included in
Table S1. All similarity values were checked out and corrected if needed. The
corrected values match well with the tree topologies.
Answers to reviewer # 2:
1. Regarding t the pathogenicity tests, this approach has been used widely in the
literature. The doses utilized are the usual for this type of experiment. Although
not included in this work, the pathogenicity tests were performed for a high
number of vibrios in paralell using the same doses, and for ost of them no
mortality was detected, excluding therefore the septic shock mentioned by the
reviewer. The transfer of the clams to a empty container for 1 h is to avoid the
dilution factor that would happen if the clams were put straight into the water.
Since the clams are intertidal organisms this period has no effects on them.
The reviewer is right in the fact that disease signs are not clear in molluscs, but
in most works the re-isolation of the inoculated bacterium is enough to prove the
pathogenic potential, if the bacterium is not detected in the controls. It is also
true that V. celticus was isolated from periodical samplings and theoretically
healthy clams, but as many other vibrios it can be an opportunistic pathogen
from the normal microbiota of the clams, which what we want to pointed out in
the manuscript.
2. Line 75, we agree with the reviewer and the sentence was changed
3. Lines 79, 93, 96 and 97, all the suggestions were accepted and the text modified
acordingly.
4. The taxonomic authorities for clam species were included in the text.
5. The other typographical and style corrections suggested by the reviewer were
included in the text.

We hope that you find this revision suitable for publication.
Sincerely,
Dr. Jesus L. Romalde
Top Related
Copyright © 2022 FDOKUMEN

