







Bahasa
Halaman
Hukum


Statut et dynamique de Vespula vulgaris,
une guêpe introduite en phase d’invasion sur les
hauts sommets de l’île de Tahiti
(Polynésie française)
Stage réalisé à la Délégation à la Recherche de la Polynésie française
sous la direction du Dr. Jean-Yves Meyer
du 19 mars au 16 mai 2012
Benoit VALLAS 2011/2012
Université Jean Monnet
23, rue Docteur Paul Michelon
42100 Saint Etienne
Délégation à la Recherche
avenue Pouvanaa a Oopa
B.P. 20981 Papeete, Tahiti
Polynésie française
Master 1 « Ecologie/ Ethologie »

Remerciements
Je tiens à remercier toutes les personnes m’ayant apporté une aide durant mon stage. J’adresse tout
particulièrement mes remerciements aux personnes suivantes, pour l’expérience enrichissante et
pleine d’intérêt qu’elles m’ont fait vivre durant ces deux mois au sein de la Délégation à la Recherche
de la Polynésie française :
- Priscille « Tea » Frogier, déléguée à la Recherche, pour son accueil chaleureux, et toutes les
explications apportées qui me permettent aujourd’hui de mieux connaitre le fonctionnement du
gouvernement polynésien. Je la remercie également de m’avoir véhiculé quotidiennement ;
- Jean- Yves Meyer, chargé de recherche, pour m’avoir consacré du temps et donner la possibilité de
traiter un sujet très intéressant. Son encadrement m’a permis d’étendre mes connaissances sur les
écosystèmes insulaires. Je le remercie vivement pour m’avoir guidé durant ce stage et de son aide
précieuse pour la réalisation de ce rapport de stage ;
- Ruth Leng Tang, secrétaire administrative, pour sa bonne humeur quotidienne et toutes ses
explications qui m’ont permis de découvrir la culture polynésienne.
Je remercie également toutes les personnes qui ont eu le courage de m’aider pour prospecter le
Mont Aorai et le Mont Marau : Tiffany Laitame (doctorante à l’Université de la Polynésie française
sur les indicateurs de suivi de la biodiversité), Alexandre Tetuamanuhiri (pompier à la défense et de
la protection civile à Papeete), Marie Fourdrigniez (consultante privée en écologie) et mon
colocataire Léo Agaesse.
Je remercie Rome Quentin (entomologiste au Muséum national d’Histoire naturelle de Paris) pour
son aide concernant l’identification de mes échantillons, Robin Pouteau (The Bio-protection Research
Centre, Lincoln University, Nouvelle Zélande) pour la création de la carte de la répartition potentielle
de la guêpe sur Tahiti, Jérome Marie (ingénieur de recherche, spécialiste en entomologie médicale,
Institut Louis Malardé, Tahiti) pour son accueil au sein du laboratoire d’entomologie de l’I.L.M à
Paea, Christian Malinowski (Association de protection de la nature « Te Rau Ati Ati a Tau a Hiti Noa
Tu ») pour nous avoir accueilli chez lui, Patrick Howell, directeur de l’ILM pour m’avoir permis de
faire une restitution orale de mon stage dans son institut de recherche médicale.
Merci aux personnes qui ont pris le temps de m’envoyer des informations par courrier électronique
afin de m’aider dans mes recherches : Sylvain Charlat (Université de Lyon 1, Laboratoire de
Biométrie et Biologie Evolutive), Peter T Oboyski (University of California, Berkeley), Thibault Ramage
(MNHN de Paris, entomologiste), Hervé Bossin (I.L.M, Tahiti), Erin Wilson (postdoctoral reseach
associate, Department of entomology University of Maryland), Bill Nagle (project coordinator, Pacific
Invasive Initiative, The University of Auckland), Raina Kaholoa’a (biologiste, Haleakala National Park,
Maui, Hawaii), Jean-Luc Renneson (collaborateur scientifique bénévole à l’Institut Royal des sciences
Naturelles de belgique IRSNB), Claire Villemant (Service du patrimoine naturel, MNHN, Paris), Elin
Claridge (anciennement à l’University of California, Berkeley), Hervé Jourdan (IRD, Nouméa).

Glossaire
Allochtone : en écologie, utilisé pour désigner des espèces d’origine étrangère à l’écosystème local.
Attractant spécifique : substance permettant d’attirer une espèce en particulier.
Biocénose : l’ensemble des êtres vivants coexistant dans un espace défini (le biotope).
Choc anaphylactique : réaction allergique exacerbée, entraînant dans la plupart des cas de graves
conséquences et pouvant engager le pronostic vital.
Clypeus : l’un des sclérites (= plaques de chitine formant l’exosquelette) qui compose le « visage »
d’un arthropode.
Déséquilibre taxonomique et fonctionnel : absence de certains groupes biologiques entraînant des
réseaux trophiques simplifiés.
Diurne : « qui vit le jour ».
Ecosystème : un ensemble dynamique d’organisme vivants (plantes, animaux et micro-organismes)
qui interagissent entre eux et avec le milieu ( sol, climat, eau, lumière) dans lequel ils vivent.
Epizootie : maladie frappant dans une région plus ou moins vaste, une espèce animale ou un groupe
d’espèces dans son ensemble.
Espèce clé de voûte : espèce qui joue un rôle important au sein d’une communauté écologique, de
par les relations interspécifiques.
Eusocial : organisation sociale d’un groupe d’individus vivant ensemble, divisé en caste d’invidus
fertiles et non fertiles.
Goulot d’étranglement : phénomène se produisant lorsqu’une population est fortement réduite sur
au moins une génération. Le faible nombre de reproducteurs restants transmet donc seulement une
fraction de la diversité génétique initiale aux générations ultérieures. Il en résulte un
appauvrissement génétique de la population. En conjonction avec l’effet fondateur, les goulots
d’étranglement sont un des mécanismes de l’évolution des espèces.
Holométabole : se dit des insectes chez qui le passage de l’état larvaire à l’état adulte se fait par
l’intermédiaire d’un état nymphal. Les larves et les adultes de ces insectes ont, en général, une
morphologie et des modes de vie très différents.
Intendation : défini un retrait par rapport à la norme. Dans le cas présent, les yeux composés des
guêpes sont en retrait derrières les antennes pour éviter des frottements.
Logiciel R : logiciel de statistique.
Lutte biologique : méthode de lutte contre un organisme (ravageur ou plante adventive) au moyen
d’organismes naturels antagonistes de ceux-ci.
Naturalisation : phase durant laquelle une espèce exotique devient capable de se reproduire dans
un nouvel environnement.

Nid pluriannuel : nid avec un cycle de plusieurs années.
Parasitoïde : organisme qui, pendant une partie seulement de son existence, vit exclusivement aux
dépend d’un autre organisme, qu’il ne détruit généralement que lorsque son propre développement
est terminé.
Plasticité phénotypique : capacité d’un organisme à exprimer différents phénotypes à partir d’un
génotype donné selon des conditions biotiques et/ou abiotiques environnementales.
Prédateur généraliste : organisme pouvant chasser différente espèces animales pour se nourrir.
Ptérygote : sous classe d’insectes possédant des ailes.
Région holarctique : région de l’hémisphère nord.
Super-colonie : colonie d’insectes sociaux formée de multiples reines et de nids. Les ouvrières se
déplacent librement de nids en nids, comme si la population entière était une unique colonie.
Tergides : plaques de chitine formant l’exosquelette des arthropodes sur la face dorsale de
l’abdomen.
Test de Kolmogorov-Smirnov : test de normalité, consistant à calculer le plus grand écart entre les
fréquences cumulées observées et les fréquences cumulées théoriques.
Test Mann-Whitney : test de statistique sui compare deux séries indépendantes sur un critère
ordinal ou quantitatif.
Test non paramétrique : test dont le modèle ne précise pas les conditions que doivent remplir les
paramètres de la population dont a été extrait l’échantillon.
Transect : ligne virtuelle ou physique mis en place pour une étude particulière.
Sex-ratio : taux comparé de mâles et de femelles au sein d’une espèce à reproduction sexuée.
Valeur P : le degré ou niveau de signification d’un test statistique.

Sommaire
I. Introduction ………………………………………………………………………………………………………………….…1
I.1. Les invasions biologiques, une menace pour la biodiversité…….……….………….………………..……..1
I.2. Les insectes sociaux envahisseurs.………………………………………………………………………………..……...1
I.2.1. Changements génétiques et environnementaux………..…………………………………..………2
I.2.2. Le frelon asiatique, un exemple célèbre d’invasion…………….…………………………..........2
I.2.3. La guêpe européenne, une espèce invasive dans le Pacifique……………………………..….2
II. Matériel et méthode …......……………………………………………………………………………………………….………3
II.1. Présentation de la Polynésie française et de l’île de Tahiti……..…………………………..…………..…..…3
II.2. Description et écologie de la guêpe européenne…………………..………………………..……………..………3
II.2.1. Critères d’identification…………………………………………………………………………………..………4
II.2.1.1. Différences inter-sexe…………………………………………………………..….……5
II.2.1.2. Différences intra-sexe……………………………………………………………………6
II.3. Sites d’études et dates de collecte…………………………………………………………………………………..……6
II.4. Méthode de collecte…………….………..……………………………………………………………………….….…...……6
II.5. Activité du nid…………………………….…………………………………………………………………………………………8
III. Résultats …….………………………………………………………………………………………………….…….…..……...8
III.1. Recueil d’informations.……………………………………………………………………………………………………….....8
III.2. Date d’introduction et dynamique de l’invasion……….………………………………………………….…..……8
III.3. Efficacité des pièges………………………………………………………………………………………………………....……9
III.4. Répartition altitudinale…………………………………………………………………………………………………….…….9
III.5. Confirmation de la différence de taille des femelles fertiles et stériles…..……....……………..……..9
III.6. Sex-ratio……………………………………………………………………………………………………………………………….11
III.7. Comportement et activité………………………..……………………………………..…………………………………..11
IV. Discussion …………………………………………………………………………………………………………………..…..12
IV .1. Une introduction par l’homme………………………..……………………………………………………………………12
IV.2. Un cycle biologique modifié…………………………………………………………………………………………………12
IV.3 Une dynamique d’invasion favorisée……..………………...…………………………………….…….………..……12
IV.4. Une aire de répartition limitée………….…………………………………………………….…….…………..……13
IV.5. Des impacts potentiels importants……..………………………………………………….…….…………………..…13
V. Conclusion et perspectives de luttes ….…………………….……………………………………………………………14

1
I. Introduction
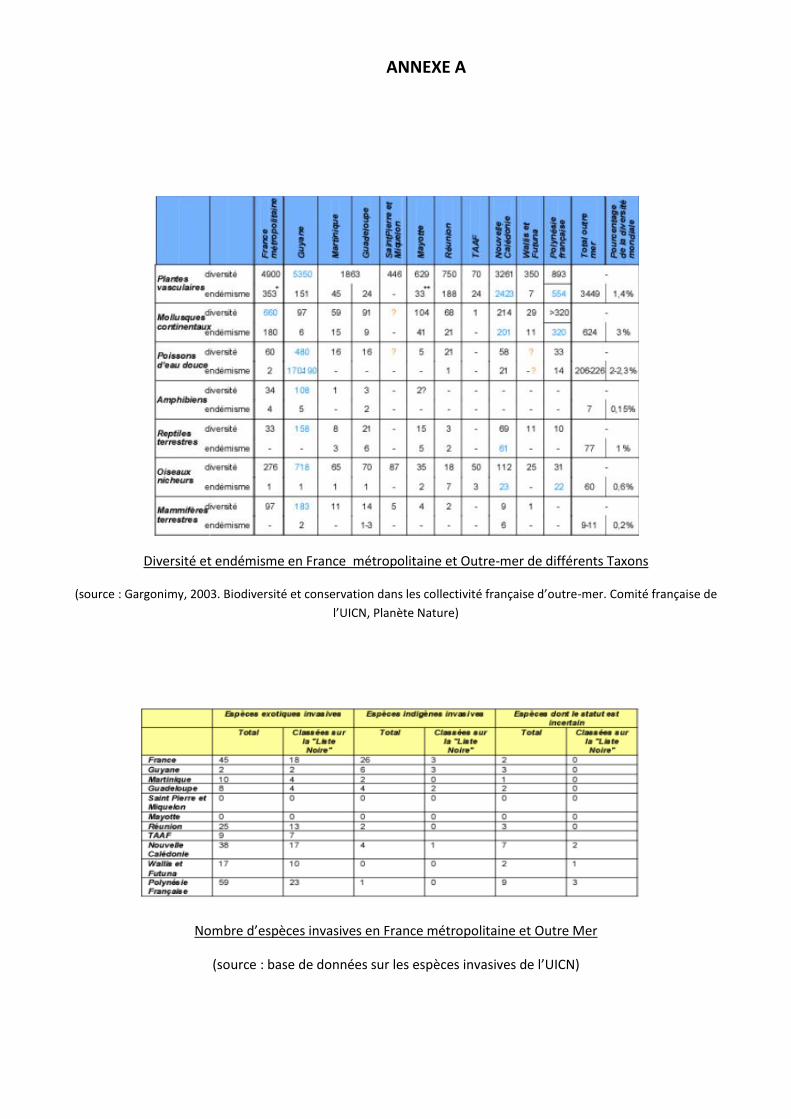
Une évaluation mondiale des écosystèmes, publiée par les Nations Unies en 2005, indique que les
invasions biologiques sont la deuxième cause d’érosion de la biodiversité après la destruction des habitats
(MEA, 2005). Les collectivités françaises d’outre-mer sont particulièrement concernées par ce
phénomène, en particulier les îles tropicales des Caraïbes, de l’Océan Indien et du Pacifique (Soubeyran,
2008). Ainsi 49 des 100 espèces considérées comme les plus envahissantes au monde par l’Union
mondiale pour la Nature (100 World’s Worst Invasive Aliens Species, IUCN) y sont représentées (cf.
ANNEXE A). Ces dernières peuvent avoir une incidence non négligeable sur un panel de secteurs
importants tel que l’environnement, la sécurité sanitaire mais aussi l’économie. A titre d’exemple, leurs
coûts annuels au niveau européen a été estimé entre 9 et 23 milliard d’euros (Kettunen et al., 2008).
I.1. Les invasions biologiques, une menace pour la biodiversité
L’ UICN à estimé que 625 des espèces répertoriées comme menacées au niveau mondial encourent un
danger d’extinction directement lié à l’invasion par des espèces allochtones (Pastoret, 2010). Devant cet
enjeu majeur pour la conservation de la biodiversité, cette thématique est actuellement au cœur des débats
scientifiques. Cependant les introductions d’espèces par l’homme ne sont pas nouvelles : en effet, depuis
le Néolithique, l’homme a été l’acteur de multiples introductions et a transporté de nombreuses espèces
d’une région à une autre afin d’améliorer la productivité alimentaire. La prise de conscience concernant
les risques liés à cette pratique à été rapidement acquise. Par le passé, certaines invasions ont été en effet
été suivies d’épidémies et d’épizooties* (peste bubonique en 1347, variole, rage, etc…).
Néanmoins, bien gérées, les introductions d’espèces permettent aussi de rentabiliser les productions
agricoles, notamment par la méthode de lutte biologique*. L’introduction d’une guêpe parasitoïde*
(Gonatocerus ashmeadi) en Polynésie française à permis une réduction de 90% de la cicadelle
envahissante Homalodisca vitripennis affaiblissant les arbres fruitiers (Grandgirard, 2008). De même, le
champignon pathogène introduit pour lutter contre l’arbre envahissant Miconia calvescens
(Melastomataceae) a permis la régénération des plantes indigènes et endémiques à Tahiti (Meyer et
Fourdrigniez, 2011 ; Meyer et al., 2012) et constitue un moyen de restaurer les milieux naturels envahis.
I.2. Les insectes sociaux envahisseurs
Les insectes sociaux appartenant à la superfamille des Vespoidea tiennent une place importante dans les
invasions animales. Ces derniers présentent une plasticité phénotypique* importante en comparaison aux
insectes solitaires. Leur adaptation peut se faire niveau individuel mais aussi au niveau de la colonie.
Cette organisation permet aussi la mise en place « d’une intelligence en essaim », un ensemble d’entités
individuellement simples au fonctionnement décentralisé capable collectivement de fournir une réponse
* Les mots suivit d’un astérisque sont référencé dans le glossaire

2
complexe (webloria.loria.fr/ chevrier//hamlin/swarm.html). Une colonie est donc la plupart du temps un
système sachant s’autoréguler et s’adapter aux variations environnementales. La réussite de l’invasion
des insectes tient souvent à divers facteurs, le plus important étant l’absence de prédateurs, de
compétiteurs ou de parasites (Tsutsui, 2000).
I.2.1. Changements génétiques et environnementaux
Lors d’une introduction, le goulot d’étranglement* conduisant à une perte de diversité génétique peut
parfois être bénéfique pour l’espèce invasive, en sélectionnant des individus ayant subi certains
changements phénotypiques et comportementaux qui seraient plus adaptés à des conditions nouvelles
(Dejean et al. 2011). C’est le cas pour la petite fourmi de feu Wasmannia auropunctata, considérée
comme un insecte « super-envahissant ». Son succès invasif vient d’un changement comportemental : les
ouvriers ne présentent aucune agressivité avec les individus des nids voisins. Cela leur permet de
cohabiter sans gaspiller de l’énergie dans des combats territoriaux. Il en résulte la création, d’une entité
qualifiée de « super-colonie *», pouvant réunir un nombre important de nids (Le Breton et al., 2004).
I.2.2. Le frelon asiatique, un exemple célèbre d’invasion
En France métropolitaine, l’introduction involontaire du frelon asiatique (Vespula velutina nigrithorax )
suscite une inquiétude grandissante. Le frelon serait arrivé en 2004 par voie maritime dans des poteries
en provenance d’Asie. Les abeilles européennes (Apis melliferas et Apis cerena), déjà fragilisées par les
activités anthropiques (pollution chimique, érosion de la diversité floristique), sont maintenant
« prédatées » par le frelon. Les abeilles participent à la pollinisation d’un nombre important d’espèces
végétales (près de 20 000), dont 40 % sont des fruits, légumes ou des oléagineuses ayant une importance
économique. L’apiculture et d’autres activités économiques sont donc touchées, de même que la
biodiversité (www.conservation-nature.fr ; Rome et al., 2011).
I.2.3. La guêpe européenne, une espèce invasive dans le Pacifique
La « guêpe européenne » (également appelée « guêpe commune ») Vespula vulgaris , classée parmi l’une
des espèces les plus invasives au monde par l’ UICN (www.issg.org/bookletF.pdf), est déjà l’auteur d’un
déséquilibre majeur de la faune et de la flore en Nouvelle Zélande (Beggs, 2001). Introduit en 1978, cet
insecte a su se naturaliser* très rapidement dans les forêts naturelles à Nothofagus (Fagaceae). L’espèce
peut y atteindre une densité 10 fois supérieure à la normale (Thomas, 1990).
L’observation récente de V. vulgaris sur les crêtes sommitales de Tahiti représente une menace
importante pour la biocénose* inféodée à ce type d’écosystème encore peu perturbé par l’homme (J.-Y.
Meyer, comm. pers.). Devant ce constat alarmant, il était donc primordial d’évaluer le statut exact de
cette nouvelle espèce allochtone* en Polynésie française.

3
L’objectif de cette étude sera d’étudier l’abondance et la répartition de la guêpe européenne dans l’île de
Tahiti, d’évaluer sa dynamique d’invasion, de comprendre les interactions biocénotiques et de proposer
une stratégie de lutte adaptées pour la préservation du patrimoine naturel spécifique de la Polynésie
française.
II. Matériel et méthode
II.1. Présentation de la Polynésie française et de l’île de Tahiti
Située en plein cœur du Pacifique Sud, la Polynésie française est une collectivité française jouissant d’une
autonomie relativement avancée (article 1er
de la loi organique du 27 février 2004). L’environnement et la
protection de la nature sont par exemple sous la compétence du gouvernement autonome.
La Polynésie française est composée de 5 archipels couvrant une superficie similaire à celle de l’Europe.
Tahiti, d’une surface d’environ 1050 km² est la plus grande des 120 îles composant ce territoire (cf.
ANNEXE B : Carte de la Polynésie française). Le climat est tropical océanique avec l’alternance d’une
saison sèche, de mai à septembre, et une saison humide, d’octobre à avril (météo France, 2004). La
Polynésie française possède une faune et une flore relativement riches, avec des taux d’endémisme
élevés, atteignant par exemple 62% pour la flore vasculaire (Meyer et al., 2010). Les sommets qui
culminent à plus de 2000 m sont dans un état de conservation remarquable et soumis à un climat plus
tempéré (diminution de la température de 0.6°C tous les 100 m). Comme souligné dans une synthèse des
espèces envahissantes dans les collectivités française d’outre-mer, « son long isolement évolutif, sa faible
superficie, sont fort taux d’endémisme et sont déséquilibre taxonomique* et fonctionnel, font de ce
modèle insulaire, un biotope très vulnérable aux invasions biologiques » (Soubeyran, 2008).
II.2. Description et écologie de la guêpe européenne
Vespula vulgaris est une guêpe appartenant à l’ordre des Hyménoptères et à la famille des Vespidae (cf.
ANNEXE B : Arbre phylogénétique des Hyménoptères). Cet insecte holométabole* est bien réparti en
région holarctique* et a été introduit en Australie, à Hawaii et en Nouvelle Zélande. C’est un insecte
eusocial* (Bourke, 1988) vivant dans des nids majoritairement souterrains pouvant atteindre une capacité
de 10 000 individus en Europe (Thomas, 2010).

4
II.2.1. Critères d’identification
Comme la plupart des guêpes du genre Vespula, l’espèce V. vulgaris est relativement difficile à identifier.
De nombreuses espèces de la famille des Vespidae (cf. ANNEXE B : Abdomen de différentes guêpes du
genre Vespula) présentent en effet des critères morphologiques similaires qui peuvent varier intra-
spécifiquement (cf. ANNEXE B : Abdomen de différentes guêpes du genre Vespula). L’observation in
natura aura donc une importance majeur afin d’établir le bon diagnostique.
Les principaux critères d’identifications importants sont la tâche frontale sur le clypeus*, la forme des
mandibules (Figure 1) et les motifs sur les tergides* (Figure 2).
Figure 1 . Face frontale de V. vulgaris (photographie prise avec reflex numérique 55 mm sous loupe binoculaire
10X3).
Figure 2 . Façe dorsale de V. vulgaris (photographie prise avec un reflex numérique 55 mm. L’individu mesurant
210 mm)
*
Aile plaquée contre le corps
Motifs spécifiques répétés sur
les différents tergides

5
II.2.1.1. Différences inter-sexe
L’identification des mâles et femelles est réalisable grâçe aux critères suivant :
- la longueur des antennes, la couleur des tempes, et la forme de l’abdomen (Q. Rome, comm. pers. 2012 ;
J.-L. Renneson, comm. pers.) (Figure 3).
Figure 3 . Critères d’identifications des femelles stériles et des mâles de V.vulgaris chez deux individus de même
taille (photographie prise sous loupe binoculaire avec un appareil reflex numérique)
Les critères morphologiques présentés ci-dessus sont néanmoins insuffisants pour ne pas
confondre cette espèce avec Vespula alascensis.
Il est nécessaire d’effectuer une observation microscopique des génitalias mâles (visibles sur la
photographie 6 en Figure 3) (Carpenter, 2010). La confirmation de l’identification a été réalisée par
Quentin Rome (comm. pers.) à partir d’un échantillon de nos spécimens collectés à Tahiti et
envoyés au Muséum d’Histoire naturelle de Paris lors de ce stage.

6
II.2.1.2. Différence intra-sexe
Le critère le plus facilement utilisable pour différencier les femelles stériles et les femelles fertiles est la
taille des individus (Figure 4). Le logiciel R* (pbil.univ-lyon1.fr/R/enseignement.html) a été utilisé afin
d’effectuer un test de comparaison de moyennes* entre notre échantillon de femelles stériles et notre
échantillon de femelles fertiles pour valider l’utilisation de cette différence de taille comme critère
d’identification. Afin de choisir le test adéquat, la normalité et la variance de nos échantillons seront
étudié au préalable. Le risque alpha consenti est 0.05.
II.3. Sites d’études et dates de collecte
En raison de l’absence de routes et pistes permettant l’accès aux hauts sommets de Tahiti, la plupart des
crêtes sommitales de l’île sont difficilement accessibles. Le relief abrupt et la végétation dense empêchent
également le repérage et l’accès aux nids si ces derniers ne sont pas situés sur ou à proximité du sentier
(cf. ANNEXE C : Sentier d’une crête sommitale au Mont Aorai).
Nos prospections ont été effectuées sur le Mont Aorai, culminant à 2070 m, et le Mont Marau avoisinant
les 1500 m (Figure 10 en page 13). Le premier site est accessible par un sentier après 6 h de marche, le
second en véhicule tout-terrain après 45 minutes de route et de piste.
Nous avons effectué nos observations sur le sentier menant au sommet de l’Aorai (latitude S :
17°36’47.3’’ ; longitude O : 149°29’42.6’’) les 6,7, 8 et les 13, 14 avril. Les observations sur le Mont
Marau ont été faites le 3 mai sur la crête est (lat. S : 17°36’33’’ ; long. O : 149°31’58’’) et le 9 mai sur la
crête ouest (lat. S : 17°36’27.8’’ ; long. O : 149°32’00.0’’) entre 1300 et 1400 m.
II.4. Méthodes de collecte
Afin de collecter des spécimens de V. vulgaris, différentes méthodes de capture ont été
utilisées lors de ce stage :
Figure 4. Différence de taille, entre une femelle stérile
(en haut), et une femelle fertile (en bas) de V.vulgaris

7
- un filet à papillon a permis leur récolte durant nos ascensions du mont Aorai, le long d’un transect
altitudinal allant de 600 m à 2070 m. Cette méthode permettra l’étude de répartition de Vespula vulgaris
en fonction de l’altitude. - des
pièges créés avec des bouteilles en plastiques vides de 1.5 litres (Figure 5) contenant une solution sucrée
à 30% (Nollet, 2009 ; Spurr, 1996 ; Seath et al., 1999), laissés durant 24h et suspendus dans des buissons,
permettront de constater la présence ou non de guêpes sur certains sites sans être obligé de rester sur place
durant toute la durée de l’opération.
Nous n’avons pas utilisé d’attractants* spécifiques pour les guêpes, par exemple ceux à base d’heptyl
butyrate (Aldrich, 1986), en raison des difficultés pour se procurer ces produits à Tahiti associés aux
délais imposés par le stage.
Nous avons utilisé ce même type de piège, placé à l’entrée du nid pendant 30 minutes, pour essayer de
capturer un plus grand nombre de guêpes (Figure 6 ; cf. ANNEXE C : Entrée du nid de Vespula vulgaris
au sommet du Mont Aorai).
Figure 5. Elaboration d’un piège à insecte volant
Figure 6. Schéma du piégeage à l’entrée du
nid de V. vulgaris
Bouteille en plastique de 1.5 L
Solution sucrée à 30 %

8
Chaque spécimen capturé sera conservé dans de l’alcool à 70°, observé et identifié à la loupe binoculaire,
et mesuré à l’aide d’une règle double décimètre.
II.5. Activité du nid
L’observation d’un nid situé au sommet de Mont Aorai (2100 m) a permis d’évaluer l’activité des guêpes
au nid durant une journée. Un comptage du nombre d’entrées et sorties des individus a été effectué durant
une minute et réalisé 5 fois, espacé d’un intervalle de 1 minute. L’activité a été mesurée à 9h, 12h30,
16h et 17h (Figure 7).
III. Résultats
Le recueil de données auprès d’informateurs locaux indique la présence de nids au niveau du sommet du
Mont Orohena (2241m) et du Mont Pito Hiti (2110 m) ( J.-Y . Meyer, comm. pers.) et d’individus au
Mont Marau entre 1100 m et 1300 m (S. Charlat, comm. pers.).
La guêpe européenne pourrait être également présente au sommet du Mont Ivirairai (1696 m) (J.-F.
Butaud, comm. pers. 2012) mais son identification reste incertaine en l’absence de photographie ou de
spécimen collecté.
III.2. Date d’introduction et dynamique de l’invasion
L’absence de V. vulgaris dans une collection d’Hyménoptères collectés dans les années 80 et déposée
dans le laboratoire d’entomologie de l’Institut Louis Malardé laisse supposer que la guêpe n’était pas
encore présente à cette époque. Les premières observations de cette guêpe datent du 16/09/1997 ( mais
non identifiée) au sommet du Mont Aorai (Meyer, comm. pers.), avec apparemment l’existence du même
nid que celui actuellement observé. Nous sommes donc en présence d’un guêpier pluriannuel qui
III.1. Recueil d’informations
Figure 7. Méthode de comptage des
entrées et sorties de V. vulgaris à
l’entrée de leur nid
Phase de comptage des entrée et sorties des
individus à l’entrée du nid durant 1 minute
Intervalle de 1 minute entre deux comptages

9
pourrait être âgé de 15 ans. V. vulgaris pourrait avoir été introduite à Tahiti au début des années 1990,
l’origine et les modalités de son introduction restant inconnues.
III.3. Efficacité des pièges
- L’utilisation de pièges contenant une solution sucrée à 30%, disposés pendant 24 h dans les buissons où
le fourragement des guêpes est important, c’est révélée inefficace car aucune guêpe n’a été piégée ;
- Le filet à papillon a permis de capturer quelques spécimens le long du gradient altitudinal et de constater
la présence d’une seule espèce de guêpe Vespula sur le site d’étude du mont Aorai ; -
l‘utilisation d’un piège installé à l’entrée du nid à permis la capture de la majorité des spécimens étudiés.
Au total, ce sont 73 femelles stériles, 10 femelles fertiles (= reines potentielles) et 2 mâles qui ont été
capturés.
III.4. Répartition altitudinale
La présence des guêpes est confirmée le long d’un transect* altitudinal allant de 1000 m à 2070 m sur le
mont Aorai. Aucune guêpe n’a été observée ni capturée en dessous de 1000 m. Nous l’avons également
été observée sur la crête sommitale du Mont Marau entre 1300 et 1400 m. Sa présence ne semble pas
affecter les deux autres espèces de guêpes présentes à Tahiti. Seliphron semantarium n’a pas été observée
au dessus de 1000 m et la « guêpe jaune » Polistes hebraus la plus commune en Polynésie française est
préférentiellement présente à basse altitude (Paulian, 1998), même si nous avons observé quelques
individus jusqu’à 2000 m.
III.5. Confirmation de la différence de taille des femelles fertiles et stériles
Afin de tester la normalité de nos échantillons, un test de Kolmogorov-Smirnov* a été utilisé. Son
utilisation est justifiée par un nombre d’individu très faible dans l’échantillon des femelles fertiles (10
individus). Pour ce test, des distributions normales théoriques ont été crées à partir de nos échantillons.
« normfemelle » est généré en simulant 1000 observations à partir de la moyenne et de l’écart- type de
l’échantillon « femelle stérile » (Figure 8A et Figure 9A). « normreine » est généré en simulant 1000
observations à partir de la moyenne et de l’écart-type de l’échantillon « reines » (Figure 8B et Figure
9B).

10
Puis nous avons testé deux à deux « normefemelle » / « femelles » et « normreine »/ « reine » pour
vérifier si nos échantillons suivent la même loi (une loi normale dans le cas présent).
Pour les femelles stériles :
La valeur de P* est inférieure au risque alpha de 5%, l’hypothèse Ho est retenue. L’échantillon
« femelle » suit une loi normale.
Pour les reines :
La valeur de P est supérieur au risque alpha, nous rejetons Ho. L’échantillon « reine » ne suit pas une loi
normale.
Les résultats obtenus ci-dessus nous indiquent qu’il est nécessaire d’utiliser un test non-paramétrique*.
La comparaison des moyennes a été effectuée avec un test de Mann-Whitney*.
Question : La taille des reines est-elle plus grande que la taille de femelles stériles ?
Sortie du logiciel R :
La valeur de P est inférieure au seuil alpha de 5 %. Nous rejetons l’hypothèse Ho. Les femelles fertiles
sont significativement plus grandes que les femelles stériles.
D = 0.1954, p-value = 0.01267
D = 0.287, p-value = 0.3885
W = 710, p-value = 9.102e-08
1. Figure 8A. Histogramme
de l’échantillon des
femelles stériles
2. Figure 8B. Histogramme
de l’échantillon des
reines
1 2
3 4
3. Figure 9A. Distribution
normale théorique simulée
à partir des paramètres de
l’échantillon « femelles »
4. Figure 9B. Distribution
normale théorique simulée à
partir des paramètres de
l’échantillon « reine »
11
III.6. Sex-ratio
Les nombreuses captures effectuées sur le Mont Aorai entre 1100 m et 2070 m nous ont également permis
d’établir un sex-ratio*. Ces captures comptabilisent les collectes effectuées avec les pièges mais aussi
avec le filet entre le 7 et 14 avril 2012. Nous avons comptabilisé 73 femelles stériles, 10 femelles fertiles,
2 mâles : Il y a donc 5 femelles fertiles pour 1 mâle.
III.7. Comportement et activité du nid
L’observation du nid situé au sommet du mont Aorai a pu nous donner des informations
comportementales. Le début des activités extérieures de V.vulgaris débute à 5h40 et se termine à 17h45.
Ces horaires correspondent au lever et coucher du soleil à cette période de l’année à Tahiti. C’est donc
une espèce de guêpe à l’activité exclusivement diurne*.
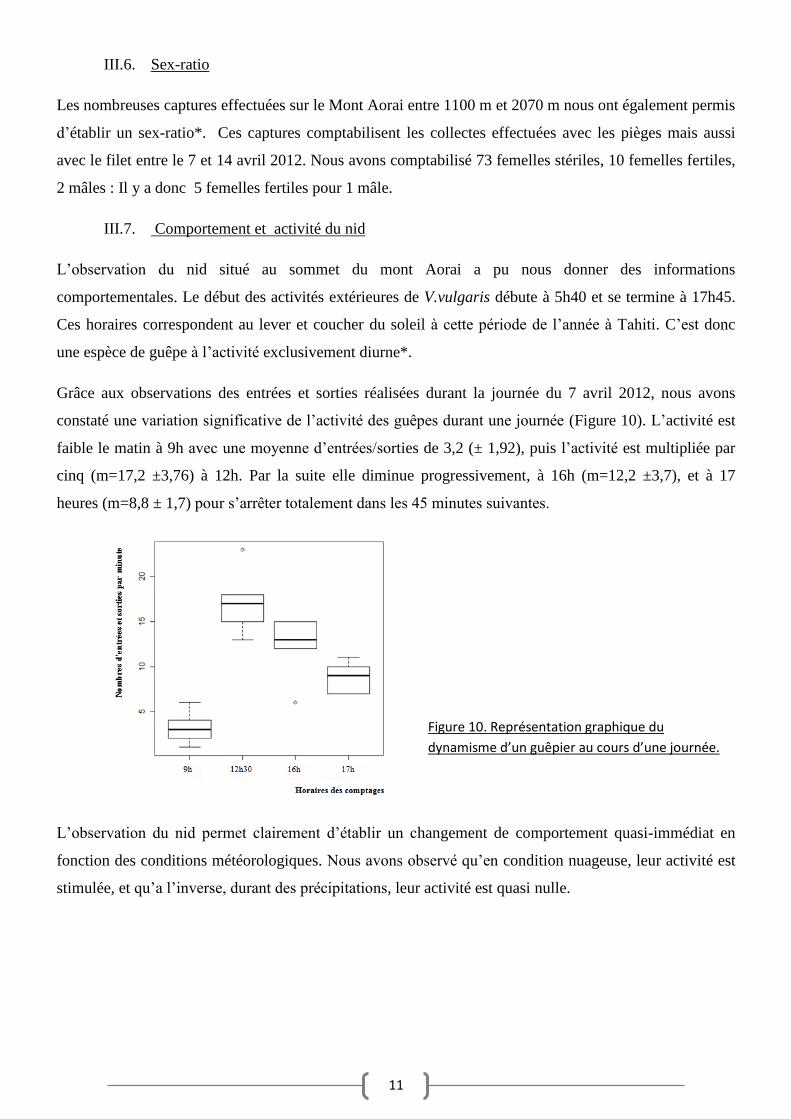
Grâce aux observations des entrées et sorties réalisées durant la journée du 7 avril 2012, nous avons
constaté une variation significative de l’activité des guêpes durant une journée (Figure 10). L’activité est
faible le matin à 9h avec une moyenne d’entrées/sorties de 3,2 (± 1,92), puis l’activité est multipliée par
cinq (m=17,2 ±3,76) à 12h. Par la suite elle diminue progressivement, à 16h (m=12,2 ±3,7), et à 17
heures (m=8,8 ± 1,7) pour s’arrêter totalement dans les 45 minutes suivantes.
L’observation du nid permet clairement d’établir un changement de comportement quasi-immédiat en
fonction des conditions météorologiques. Nous avons observé qu’en condition nuageuse, leur activité est
stimulée, et qu’a l’inverse, durant des précipitations, leur activité est quasi nulle.
Figure 10. Représentation graphique du
dynamisme d’un guêpier au cours d’une journée.

12
IV. Discussion
IV.1. Une introduction par l’homme
Sans connaître la date ni les modalités exactes de son arrivé à Tahiti, nous pouvons avancer l’hypothèse
d’une introduction de par l’intermédiaire d’un vecteur d’origine anthropique. En effet la Polynésie
française est trop éloignée des autres îles et pays où l’espèce est déjà présente (la Nouvelle Zélande est
située à 4000 kilomètre de Tahiti) pour permettre à cette espèce de pouvoir y arriver de façon naturelle.
Une étude génétique à grande échelle permettrait de connaitre l’origine de la population tahitienne de V.
vulgaris et de comprendre son mode d’introduction.
IV.2. Un cycle biologique modifié
En Europe, la reine fertilisée sort de l’hivernage au début du printemps afin d’entreprendre la
construction d’un nid, elle pond rapidement des œufs, afin que des ouvrières stériles (Foster, 2000)
prennent le relais pour parachever le nid et s’occuper de leur souveraine.
A l’automne, il y a la production d’individus mâles et de nouvelles reines pour permettre la reproduction.
Au début de l’hiver les ouvrières et les mâles sont emportés par les premières gelées, seules les nouvelles
reines fécondées hibernent. Ces dernières pourront alors amorcer de manière individuelle la mise en
place d’un guêpier, au début du printemps. Avant leur implantation dans un milieu elles peuvent parcourir
jusqu'à 70 kilomètres.
Dans le cas de Tahiti, île tropicale où les saisons sont peu marquées et sans hiver froid, l’apparition de
nids pluriannuels* a été favorisé, comme cela a déjà été observé à Hawaii (Gambino, 1990). Ces derniers
sont de tailles plus conséquentes avec un nombre plus important d’individus. Le cycle cité ci-dessus est
alors lui aussi modifié. Les résultats obtenus sur le sex-ratio de V. vulgaris nous indique la présence de
mâle (1 mâle pour 5 femelles fertile) durant les mois d’avril et mai, ce qui doit correspondre à une partie
de la saison de reproduction de cette guêpe à Tahiti. Une étude sur un plus long terme ( au minimum une
année) permettrait d’étudier le cycle biologique complet de V . vulgaris afin d’évaluer de manière plus
précise la dynamique des populations et le potentiel invasif de l’espèce.
IV.3. Une dynamique d’invasion favorisée
La présence de nids pluriannuels nous donne des informations sur le potentiel invasif important de
l’espèce.
Ces structures inhabituelles, ayant un cycle plus long qu’une année ont souvent une capacité d’individus
bien supérieure à la moyenne connue en Europe. La concentration des individus autour du nid est donc
plus importante, augmentant ainsi l’impact de ces insectes sur leur environnement.
13
Cela influe aussi sur la dynamique d’invasion de l’espèce : en effet, la présence de nid pluriannuel leur
permet d’accroître leur aire de répartition de manière plus rapide. Puisque les nids restent actifs, les
nouvelles reines n’ont pas d’autres choix que de migrer pour s’implanter ailleurs.
IV.4. Une aire de répartition relativement limitée
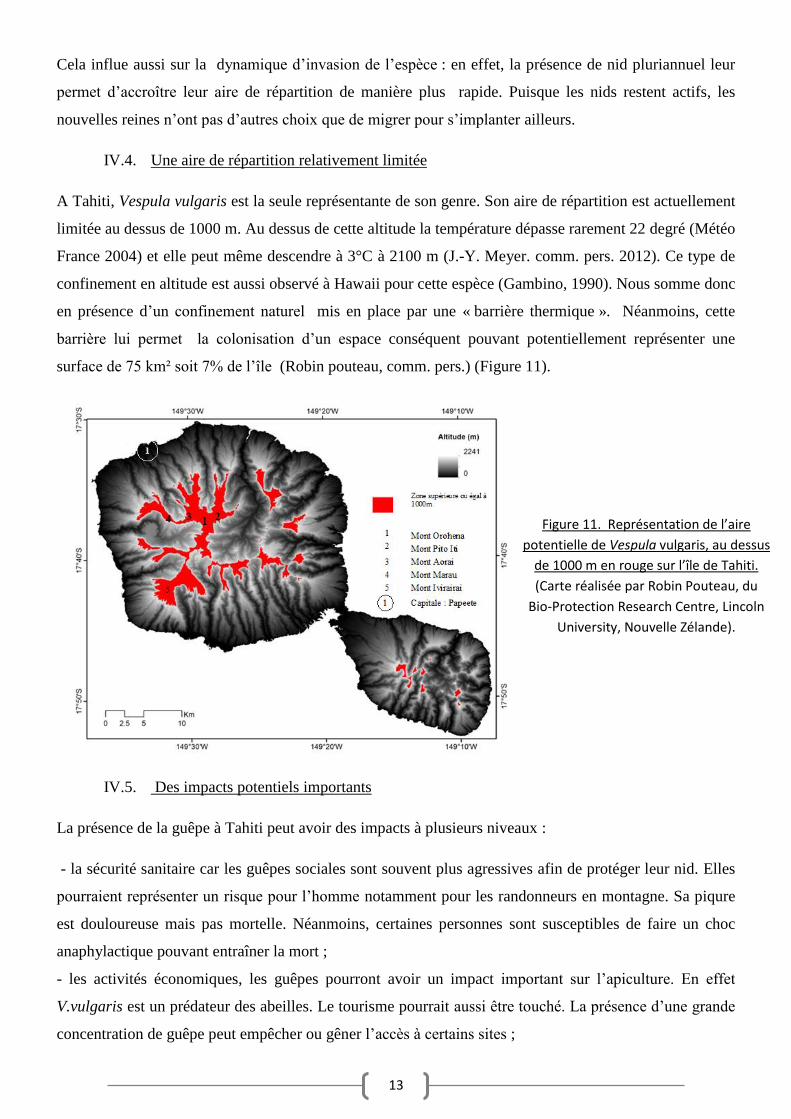
A Tahiti, Vespula vulgaris est la seule représentante de son genre. Son aire de répartition est actuellement
limitée au dessus de 1000 m. Au dessus de cette altitude la température dépasse rarement 22 degré (Météo
France 2004) et elle peut même descendre à 3°C à 2100 m (J.-Y. Meyer. comm. pers. 2012). Ce type de
confinement en altitude est aussi observé à Hawaii pour cette espèce (Gambino, 1990). Nous somme donc
en présence d’un confinement naturel mis en place par une « barrière thermique ». Néanmoins, cette
barrière lui permet la colonisation d’un espace conséquent pouvant potentiellement représenter une
surface de 75 km² soit 7% de l’île (Robin pouteau, comm. pers.) (Figure 11).
IV.5. Des impacts potentiels importants
La présence de la guêpe à Tahiti peut avoir des impacts à plusieurs niveaux :
- la sécurité sanitaire car les guêpes sociales sont souvent plus agressives afin de protéger leur nid. Elles
pourraient représenter un risque pour l’homme notamment pour les randonneurs en montagne. Sa piqure
est douloureuse mais pas mortelle. Néanmoins, certaines personnes sont susceptibles de faire un choc
anaphylactique pouvant entraîner la mort ;
- les activités économiques, les guêpes pourront avoir un impact important sur l’apiculture. En effet
V.vulgaris est un prédateur des abeilles. Le tourisme pourrait aussi être touché. La présence d’une grande
concentration de guêpe peut empêcher ou gêner l’accès à certains sites ;
Figure 11. Représentation de l’aire
potentielle de Vespula vulgaris, au dessus
de 1000 m en rouge sur l’île de Tahiti.
(Carte réalisée par Robin Pouteau, du
Bio-Protection Research Centre, Lincoln
University, Nouvelle Zélande).

14
- enfin un impact écologique fort. Les écosystèmes* naturels peuvent être gravement déséquilibrés V.
vulgaris est en effet une espèce clef de voûte*. En effet, c’est un prédateur généraliste, lui permettant
d’exercer une pression de prédation sur un grand nombre d’invertébrés, susceptibles d’être éliminés les
uns après les autres. Elle se nourrit aussi de ressources sucrées tel que des fruits ou du nectar. La forte
concentration de cette espèce pose donc un problème de compétition d’accès aux ressources alimentaire
pour les autres invertébrés voire certains vertébrés (oiseaux frugivores et nectarivores). Par contre son
impact sur la flore polynésienne est à nuancer. Elle exerce une pression de prédation sur certains insectes
herbivores, les plantes sont donc moins attaquées. C’est un insecte pollinisateur, elle va donc permettre la
reproduction de différentes espèces endémiques, mais aussi amplifier l’invasion de certaines espèces
végétales allochtones. L’étude détaillée du régime alimentaire de l’espèce permettrait de mieux
comprendre les interactions biotiques et d’estimer correctement l’impact de V.vulgaris sur l’entomofaune,
les oiseaux nectarivores, mais aussi la flore tahitienne (cf. Figure 12 présenté en conclusion).
V. Conclusions et perspectives de lutte
Nous avons confirmé la présence et l’identité de V.vulgaris à Tahiti grâce à la collecte au filet de
nombreux spécimens et l’observation d’un nid au Mont Aorai. Cette espèce, probablement introduite par
l’intermédiaire de l’homme dans les années 1990 a su se naturaliser au dessus de 1000 m d’altitude ou les
températures moyennes annuelles sont inférieures à 22°C. L’absence de saison marquée et d’hiver froid à
permis à cette guêpe la création de nids pluriannuels. Ces derniers nous indiquent que la dynamique
d’invasion de l’espèce est potentiellement grande. En effet la présence de nids viables plusieurs années
successives oblige les nouvelles reines à conquérir de nouveaux territoires pour s’implanter. De plus, les
nids pluriannuels intensifient l’impact des guêpes sur leur environnement. Le nombre d’individus par nid
est plus grand, les invertébrés sont donc soumis à plus de prédateurs généralistes mais aussi plus de
compétiteurs aux ressources alimentaires (Figure 12).
Figure 12. Interactions biotiques de V. vulgaris
avec son environnement

15
Enfin, à l’aide des résultats obtenus sur le statut et la dynamique d’invasion de V. vulgaris, il est
important que les gestionnaires locaux prennent une décision concernant le devenir de cette espèce et
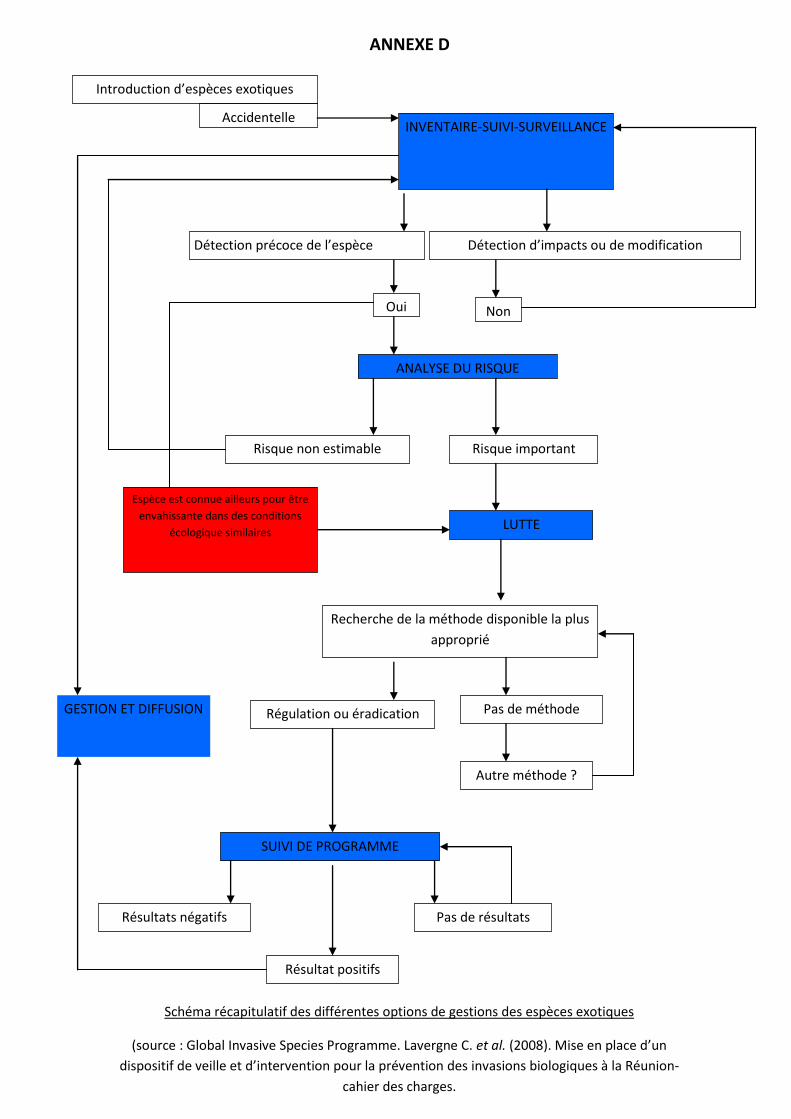
mettent en place une stratégie de gestion adaptée : éradication totale ou rcontrôle des populations (cf.
ANNEXE D).
V.vulgaris est peut être en période de latence (« lag phase ») car les nids semblent encore en faible
nombre, mais une explosion démographique est possible dans les années à venir. Cela à été observé chez
deux autres espèces invasives à Tahiti, l’arbre Miconia calvescens et l’oiseau Pycnonotus cafer qui ont
mis environ 30 ans après leur introduction avant d’envahir l’île (J. -Y. Meyer, comm. pers. ).
Les méthodes de lutte possibles sont :
- la régulation (ou contrôle) des populations en utilisant la méthode de lutte biologique. L’utilisation du
parasitoïde Sphecophaga vesparum burra spécifique des guêpes Vespula est possible, néanmoins les
premiers résultats sont visibles 20 ans après son introduction comme observé en Nouvelle Zélande (Harris
et al., 1999).
- l’élimination des nids existants au moyen de méthodes chimiques avec un insecticide, des appâts
empoisonnés au sulfuramide (Spurr, 1993) ou injection d’un gaz
Un classement de Vespula vulgaris dans la liste des « espèces menaçants la biodiversité de la Polynésie
française » selon le Code de l’Environnement en vigueur dans ce territoire (www.environnement.pf)
serait pertinent pour initier des actions contre cette espèce. Afin d’anticiper et de prévenir la possible
invasion de la guêpe européenne, il est important de sensibiliser le grand publique à Tahiti (cf. ANNEXE
E), mais aussi les îles pouvant être envahie par cette espèce telles que les îles arquises présentant un relief
important, ou les îles Australes régies par un climat sub-tempéré qui permettrait la colonisation de
V.vulgaris sur l’intégralité de ces îles.

Bibliographie
Aldrich J. R., Lusby W. R., Kochansky J. P. (1986). Identification of a new
predaceous stink bug pheromone and its attractiveness to the eastern
yellowjacket. Cellular and Molecular Life Sciences 42(5): 583-585
Beggs J. (2001). The ecological consequences of social wasps invading an ecosystem that has an abundant carbohydrate resource. Biological Conservation, 99: 17-28.
Bourke A. F. G. (1988). Worker reproduction in the higher eusocial hymenoptera. The Quartely Review of Biology, 63(3): 291-311.
Carpenter J. M., Glare T. R. (2010). Misidentification of Vespula alascensis as V.vulgaris in North America ( Hymenoptera : Vespidae ; Vespinae). American Museum Natural, 3690: 1-7.
Dejean A., Céréghino R., Carpenter J. M., Corbara B., Hérault B., Rossi V., Leponce M. , Orivel J. , Bonal D. (2011). Climate change impact on Neotropical social wasps. PloS ONE, 6(11). DOI:10.1371/journal.pone.0027004.
Foster K. R., Ratnieks F. L. W. (2000). Convergent evolution of worker policing by egg eating in the honeybee and common wasp. The Royal Society London. B., 268(1463): 169-174.
Gambino P., Medeiros A. C., Loope L. L. (1990). Invasion and colonization of upper elevations on East Maui by Vespula pensylvanica. Annals of the Entomological Society of America, 83(6): 1088-1095.
Grandgirard J., Hoddle M. S., Petit J. N., Roderick G. K., Davies N. (2008). Engineering an invasion: classical biological control of the glassy-winged sharpshooter, Homalodisca vitripennis, by egg parasitoid Gonatocerus ashmeadi in Tahiti and Moorea, French Polynesia. Biological invasions, 10(2): 135-148.
Harris R. J., Read P. E. C., (1999). Enhanced biological control of wasps. Science for Conservation 115 Department of Conservation, New Zealand, 39 p.
Kettunen M., Genovesi P., Gollasch S., Pagad S., Starfinger U., Brink P.T., Shine C. (2008). Technical support to EU strategy on invasive species (IS)- Assessment of the impacts of IS in Europe and the EU. Institute for European Environmental Policy (IEEP), Brussels, 40 p.

Le Breton J., Delabie J. H. C., Chazeau J., Dejean A., Jourdan H. (2004). Experimental evidence of large-scale unicoloniality in the tramp ant Wasmannia auropunctata. Journal of Insect Behavior, 17(2): 263-271
Meyer J.-Y., Fourdrigniez M. (2011). Conservation benefits of biological control: The recovery of a threatened plant subsequent to the introduction of a pathogen to contain an invasive tree species. Biological Conservation 144: 106-113,
Meyer J.-Y., Fourdrigniez M., Taputuarai R. (2012). Restoring habitat for native and endemic plant through the introduction of fungal pathogen to control the alien invasive tree Miconia calvescens in the island of Tahiti. BioControl DOI: 10.1007/s10526-011-9402-6
Meyer J.-Y., Salvat B. (2009). French Polynesia: biology. Pp 332-338 in Gillespie R. G., Clague D. A. (eds,.) Encyclopedia of Islands. University of California Press, Berkeley, 1074 p.
Météo France (2004). Atlas climatologique de la Polynésie française. Météo-
France- Direction interregional de Polynésie française, Papeete, 113 p.
Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-being,
Biodiversity Synthesis. World Ressources Institute, Washington, DC .
Nollet F. B., Gomes B. (2009). An improved bait method for collecting Hymenoptera, especially social wasps. Neotropical Entomology, 38(4): 477-481.
Pastoret P.-P., Moutou F. (2010). Espèces envahissantes. Rev. Sci. Tech. Off. Int. Epiz., 29(1) : 15-18.
Paulian R. (1998). Les Insectes de Tahiti. Société nouvelle des Editions Boubée. Paris, 331 p.
Rome Q., Perrard A., Muller F., Villemant C. (2011). Monitoring and contol modalities of a honeybee predator, the yellow-legged hornet Vespa velutina nigrithorax. Aliens: The Invasive Species Bulletin 31: 7-15.
Seath C. J., Crescent T., Newmills, Dunfermline, Fife, Scotland (1999). Wasp trapping with carbohydrates. Proceeding of the 3rd International Conference on Urban Pests. Robinson H., Rettich F., Rambo G. W. (eds).
Soubeyran, Y (2008). Espèces exotiques envahissantes dans les collectivités françaises d’outre-mer. Etat des lieux et recommandations. Comité français de l’UICN. Paris, 202 p.
Spurr E. B. (1996). Carbohydrate bait preferences of wasps. New Zealand Journal of Zoology, 23: 315-324.

Spurr E. B (1993). The effectiveness of sulfuramid in sardine bait for control of wasps. Proceedings of the 46th New Zealand Plant Protection Conference: 307-312.
Stratégie Nationale pour la biodiversité 2011-2010 (2011). Ministère de l’Ecologie, du Développement durable, des Transports et du Logement, Paris.
Thomas C. D., Moller H., Plunkett G. M., Harris R. J (1990). The prevalence of introduced Vespula vulgaris wasps in a New Zealand beech forest community. New Zealand Journal of Ecology, 13: 63-72
Tsutsui N. D., Suarez A. V. , Holway D. A., Case T. J. (2000). Reduced genetic variation and the success of an invasive species. Proceeding of the National Academy of sciences of the United States of America, 23 ; 97 (11): 5948-53
Sites internet
www.issg.org
www.conservation-nature.fr, GB, Invasion biologique, Frelon asiatique
www.webloria.loria.fr
www.environement.pf
www.uinc.fr
pbil.univ-lyon1.fr/R/enseignement.html
ANNEXE A
Diversité et endémisme en France métropolitaine et Outre-mer de différents Taxons
(source : Gargonimy, 2003. Biodiversité et conservation dans les collectivité française d’outre-mer. Comité française de
l’UICN, Planète Nature)
Nombre d’espèces invasives en France métropolitaine et Outre Mer
(source : base de données sur les espèces invasives de l’UICN)
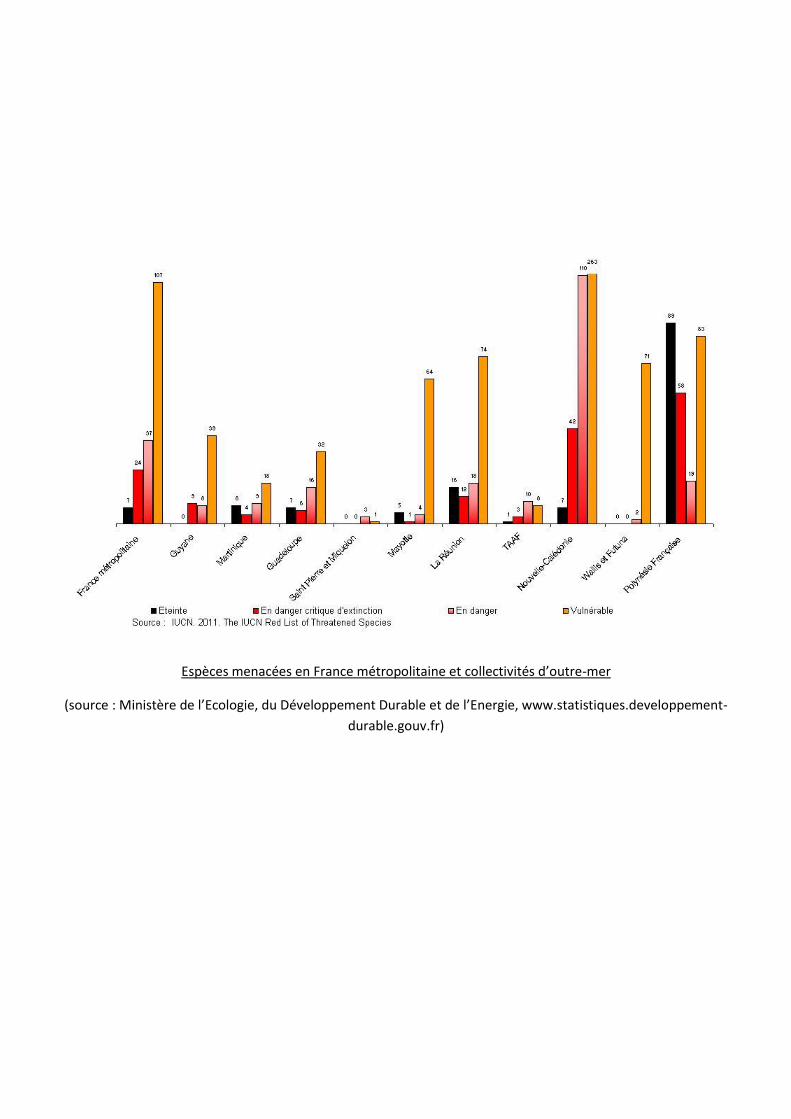
Espèces menacées en France métropolitaine et collectivités d’outre-mer
(source : Ministère de l’Ecologie, du Développement Durable et de l’Energie, www.statistiques.developpement-
durable.gouv.fr)

ANNEXE B
Carte de la Polynésie française

Abdomen de différentes guêpes du genre Vespula
(source : Canadian Journal of Arthropod Identification.
www.biology.ualberta.ca/bsc/ejournal/bmc_05/key_vespula.html)
ANNEXE C
Arbre phylogénétique des Hyménoptères

ANNEXE C
Sentier d’une crête sommitale au Mont Aorai
Entrée du nid de Vespula vulgaris au sommet du Mont Aorai (2070 m)
Introduction d’espèces exotiques
Accidentelle INVENTAIRE-SUIVI-SURVEILLANCE
Détection précoce de l’espèce Détection d’impacts ou de modification
Espèce est connue ailleurs pour être
envahissante dans des conditions
écologique similaires
ANALYSE DU RISQUE
Oui Non
Risque important
LUTTE
Résultats négatifs
Autre méthode ?
Régulation ou éradication
Recherche de la méthode disponible la plus
approprié
SUIVI DE PROGRAMME
Pas de méthode
Risque non estimable
Résultat positifs
Pas de résultats
GESTION ET DIFFUSION
ANNEXE D
Schéma récapitulatif des différentes options de gestions des espèces exotiques
(source : Global Invasive Species Programme. Lavergne C. et al. (2008). Mise en place d’un
dispositif de veille et d’intervention pour la prévention des invasions biologiques à la Réunion-
cahier des charges.

ANNEXE E
Fiche d’identification de Vespula vulgaris, une guêpe introduite en phase d’invasion
(Fiche réalisé durant le stage, afin de sensibiliser le grand public et les gestionnaires locaux)

Restitution orale des résultats du stage aux autorités locales

Résumé
Les invasions biologiques sont considérées comme l’une des principales menaces pour la biodiversité
dans les îles, en particulier celles de la Polynésie française (Pacifique Sud). La guêpe européenne Vespula
vulgaris, déjà envahissante à Hawaii, en Australie et en Nouvelle-Zélande, a réussi à se naturaliser sur les
plus hauts sommets de l’île de Tahiti. Le recueil de données indiquerait une introduction possible dans
les années 90. L’observation et la capture de 85 spécimens le long d’un gradient altitudinal au mont
Aorai (entre 600 et 2100 m) montrent que son aire de répartition est exclusivement située au dessus de
1000 m où la température moyenne annuelle est inférieure à 22 °C. L’absence de saisons marquées à
Tahiti, sans hiver froid, permettrait la formation de nids pluriannuels, dont un seul a été trouvé à l’Aorai.
Le sex-ratio (1 mâle pour 5 femelles fertile) indiquerait que la guêpe est en période de reproduction
durant le mois d’avril. L’activité du nid s’étale entre 5h45 et 17h45 avec un pic à 12h30, mais varie avec
les conditions climatiques. Cette guêpe omnivore étant un prédateur généraliste, son impact potentiel
sur tous les invertébrés présents au dessus de 1000 m, mais aussi la pression de compétition exercée
pour les ressources alimentaires, pourrait affecter le fonctionnement de l’écosystème montagnard de
l’île. Afin de mieux évaluer les impacts de V. vulgaris et tenter son éradication ou le contrôle de ses
populations, il serait important de mieux connaître son cycle biologique tout au long de l’année mais
également son régime alimentaire.
Mots clés : biodiversité, compétition, espèce envahissante, guêpe, île, insectes sociaux, Polynésie
française, prédation, sex-ratio, Vespula vulgaris
Abstract
Biological invasions are considered one of the main threat to biodiversity in islands, including those of
French Polynesia (South Pacific). The European wasp Vespula vulgaris, already invasive in Hawaii,
Australia and New Zealand, is now naturalized on the highest summits of Tahiti island. Data collection
indicate a possible introduction in the 90’s. Observation and capture of 85 specimens along an elevation
gradient on mont Aorai (between 600 and 2,100 m) demonstrate that the wasp is exclusively found
above 1,000 m elevation where mean annual temperature is below 22 °C. The lack of pronounced
seasons, without a cold winter, may allow the formation of multi-annual nests, only one being found on
Aorai. Sex-ratio (1 male for 5 fertile females) indicates that reproduction occurs in April. Nest activity is
between 5:45 am to et 5:45 pm with a peak at 12:30 am, but varies with climatic conditions. This
omnivorous wasp is a generalist predator. Its potential impact on all invertebrates living above 1,000 m
elevation, as well as competition with their food sources, may alter the functioning of the island unique
montane ecosystem. To better assess the impacts of V. vulgaris and try to eradicate the species or
control its populations, it would be important to improve the knowledge on its life cycle but also on its
diet.
Key words : biodiversity, competition, French Polynesia, island, invasive species, predation, social
insects, sex-ratio, Vespula vulgaris, wasp
Top Related
Copyright © 2022 FDOKUMEN

