







Bahasa
Halaman
Hukum


ORIGINAL ARTICLE
Validation of metabolomics for toxic mechanism of actionscreening with the earthworm Lumbricus rubellus
Qi Guo Æ Jasmin K. Sidhu Æ Timothy M. D. Ebbels Æ Faisal Rana ÆDavid J. Spurgeon Æ Claus Svendsen Æ Stephen R. Sturzenbaum ÆPeter Kille Æ A. John Morgan Æ Jacob G. Bundy
Received: 12 August 2008 / Accepted: 15 December 2008! Springer Science+Business Media, LLC 2009
Abstract One of the promises of environmental meta-bolomics, together with other ecotoxicogenomic approaches,
is that it can give information on toxic compound mechanism
of action (MOA), by providing a specific response profile orfingerprint. This could then be used either for screening in the
context of chemical risk assessment, or potentially in con-
taminated site assessment for determining what compoundclasses were causing a toxic effect. However for either of
these two ends to be achievable, it is first necessary to know if
different compounds do indeed elicit specific and distinctmetabolic profile responses. Such a comparative study has
not yet been carried out for the earthworm Lumbricus
rubellus. We exposed L. rubellus to sub-lethal concentrationsof three very different toxicants (CdCl2, atrazine, and fluo-
ranthene, representing three compound classes with different
expected MOA), by semi-chronic exposures in a laboratorytest, and used NMR spectroscopy to obtain metabolic pro-
files. We were able to use simple multivariate pattern-
recognition analyses to distinguish different compounds tosome degree. In addition, following the ranking of individual
spectral bins according to their mutual information with
compound concentrations, it was possible to identify bothgeneral and specific metabolite responses to different toxic
compounds, and to relate these to concentration levels
causing reproductive effects in the worms.
Keywords Environmental biomarker ! Atrazine !Cadmium ! Fluoranthene ! Ecotoxicogenomics !Metabonomics
1 Introduction
The major application of metabolomics in the environ-
mental sciences to date has been in ecotoxicology. Within
this, a majority of groups are working in aquatic ecotoxi-cology, recently reviewed by Lin et al. (2006) and Viant
(2007). In contrast, fewer studies have been carried out on
terrestrial models; to date, most of the examples of appli-cation of metabolomics in terrestrial ecotoxicology are in
earthworms. Earthworms have, of course, been widely used
for assessing soil contamination for many years (Spurgeonet al. 2003b; Sanchez-Hernandez 2006). A host of different
biochemical and biomolecular endpoints have been used as
measures of earthworm response, and so it is not surprisingthat metabolite profiles have been added to the arsenal of
molecular biomarkers used by terrestrial ecotoxicologists.
Qi Guo and Jasmin K. Sidhu contributed equally to this paper.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11306-008-0153-z) contains supplementarymaterial, which is available to authorized users.
Q. Guo ! J. K. Sidhu ! T. M. D. Ebbels ! F. Rana !J. G. Bundy (&)Department of Biomolecular Medicine, Division of Surgery,Oncology, Reproductive Biology, and Anaesthetics, Facultyof Medicine, Imperial College London, Sir Alexander FlemingBuilding, London SW7 2AZ, UKe-mail: [email protected]
D. J. Spurgeon ! C. SvendsenCentre for Ecology and Hydrology, Maclean Building, BensonLane, Crowmarsh Gifford, Wallingford OX10 8BB, UK
S. R. SturzenbaumPharmaceutical Science Division, King’s College London,School of Biomedical & Health Sciences, Franklin WilkinsBuilding, Stamford Street, London SE1 9NH, UK
P. Kille ! A. J. MorganUniversity of Cardiff, School of Biosciences, Main Building,Park Place, Cardiff CF10 3TL, UK
123
Metabolomics
DOI 10.1007/s11306-008-0153-z

Recent publications have used NMR-based profiling for
assessing field and semi-field studies on Lumbricus rubel-lus (Bundy et al. 2004, 2007, 2008; Jones et al. 2008). This
species also has EST sequence data available (Owen et al.
2008), meaning that it is suitable for parallel transcriptomicstudies. Furthermore, a project is ongoing to sequence the
entire L. rubellus genome (see www.earthworms.org for
more information). In addition, L. rubellus is widely dis-tributed and can often be found at field sites, making it a
useful choice for both lab and field studies. Eisenia fetida isanother important terrestrial model, because it is required
for regulatory testing (OECD 1984), although as an
extreme epige it is less relevant for soil testing than L.rubellus. Brown et al. (2008) have evaluated different
sample preparation methods with E. fetida which, coupled
with additional genomic information in the form of an ESTlibrary and resultant cDNA microarrays (Gong et al. 2007),
makes it, like L. rubellus, a very suitable organism to
combine future metabolomic research with other post-genomic analyses.
For metabolomics (and other toxicogenomic methods)
to be adopted as a useful monitoring tool in ecotoxicology,it is not enough to do as well as existing methods: sub-
stantial improvements are needed (van Straalen and
Roelofs 2008). Hence an important question is, whatadvantages are there to using ecotoxicogenomic methods in
addition to current approaches based on chemical residue
analysis? If biological response information is needed, whynot make use of a known biomarker response? One pos-
sibility is that omic methods can potentially discriminate
between specific compounds/toxic mode of action (MOA);it then becomes clear that being able to discriminate and
assign MOAs is a critical need for environmental meta-
bolomics studies. Metabolic profiling has been widely usedfor classifying or discriminating MOA in toxicology
(Gartland et al. 1989, 1990, 1991; Ebbels et al. 2007), and
has also been applied to earthworm multiple-toxicantstudies (Bundy et al. 2002). However, there are currently
no studies comparing the metabolic effect of toxicants with
different MOA for the species L. rubellus. It is alsoimportant that enough baseline data are available to give a
context for interpreting metabolic responses—i.e. is a
metabolite change really a biomarker of a specific MOA, oris it instead a more general stress response (van Straalen
and Roelofs 2008)?
Field exposure data are important in ecotoxicology, aslaboratory exposures cannot fully model the complexity of
the natural environment, particularly for a highly hetero-
geneous matrix such as soil. Nonetheless controlledexposures to single toxicants are still essential, in order to
provide a context for field observations. Here, we present
data for NMR-based profiling of tissue extracts ofL. rubellus following chronic soil exposure to three
different toxicants: Cd, fluoranthene (FA), and atrazine
(AZ), i.e. a toxic metal, a common organic pollutant, andan agrochemical (herbicide). We have identified both
general and toxicant-specific biomarkers in the dataset. In
addition, we tested if the profiles could be used to distin-guish the effects of different toxicants, and thus potentially
assign MOA. These data will provide a useful baseline for
future metabolomic experiments using L. rubellus.
2 Materials and methods
2.1 Chemical exposures
A full description of the exposure protocol was given by
Owen et al. (2008), and so we will give only a summaryhere. Briefly, earthworms were exposed in a spiked natural
soil, using an existing 28-day test protocol (Spurgeon et al.
2003a). The toxicants Cd, fluoranthene (FA), and atrazine(AZ) were mixed with a commercially available loam soil
(soil characteristics described by Spurgeon et al. 2004) at a
range of concentrations, using an experimental design withreplicated concentrations (n = 8 for controls, and n = 5
for replicated toxicant concentrations), supplemented by
individual non-replicated exposures at intermediate andadditional concentrations (Table 1). The concentrations
were chosen based on previous experiments to cover the
same sub-lethal effect range for each toxicant, as shownby similar reductions in reproduction for each toxicant
across the concentration ranges used. Each replicate
comprised a group of 8 clitellate earthworms in 1 kg ofsoil. Exposure was for 28 days, and survival, weight
change, and cocoon production at the end of the period
were all recorded. At the end of the 28 days, three indi-viduals from each replicate were then pooled, snap-frozen,
and ground, to give a single sample. In total, 104 samples
were analysed.
2.2 Sample preparation for NMR analysis
We lyophilized ground tissues without allowing them to
thaw, and then stored them at -80"C until extraction. We
extracted the tissues by homogenizing approx. 80 mg dryweight into 4 ml of 70% ice-cold acetonitrile solution,
using a Heidolph SilentCrusher homogenizer. One ml of
each of the extracts was then subjected to solid-phaseextraction, using Strata-X-AW mixed-mode 6 ml car-
tridges with 200 mg adsorbent (Phenomenex, Macclesfield,
UK), in order to remove 2-hexyl-5-ethyl-furan-3-sulfonicacid (HEFS). The extracts were diluted with water, loaded
onto the cartridges, and eluted with 6 ml of HPLC-grade
methanol. The eluate was then dried using a rotary vacuumconcentrator at 45"C, and resuspended in 0.65 ml of NMR
Q. Guo et al.
123
buffer (0.1 M phosphate buffer, pH 7.0; 0.98 mM sodium
trimethylsilyl-2,2,3,3,-2H4-propionate (TSP); made up in
90% v/v 2H2O). This resuspended sample was then filteredthrough a 10 kDa Nanosep centrifugal membrane filter
(VWR, Lutterworth, UK), which had previously been
rinsed three times with 2H2O to remove traces of glycerol.The samples (0.55 ml) were then transferred to 5 mm
NMR tubes.
2.3 NMR analysis
We acquired the spectra on a Bruker Avance DRX600spectrometer (Bruker, Rheinstetten, Germany) with a 14.1
T magnet and a resulting 1H resonance frequency of
600 MHz, which was equipped with a 5 mm triple-axis-inverse probe and BACS tube-changer autosampler. The
samples were loaded onto the autosampler in randomized
blocks, and were held at room temperature while on theautosampler, and at 300 K during acquisition. The spectra
were acquired using previously described parameters for
one-dimensional spectra using the first increment of aNOESY experiment together with solvent suppression on
the water resonance (Beckonert et al. 2007); 256 transients
were acquired per sample, following 8 dummy scans toapproach a steady state. Data were acquired into 32 K
points across a 12 kHz spectral width, with a resultant
acquisition time of 1.36 s. An additional longitudinalrelaxation delay of 3.5 s was included, giving a 5 s recycle
time. We processed the spectra using iNMR 2.5.5 (Nu-
cleomatica, Molfetta, Italy). We performed time-domainfiltering of the residual solvent peak for each spectrum, and
then multiplied each FID by a 0.5 Hz exponential apodi-
zation function, followed by Fourier transformation. Phasevalues and baselines were both adjusted automatically,
using the ‘metabolomic phase correction’ option, and first-
order polynomial baseline correction. The data wereexported at three different resolutions for further analysis:
(i) at the native spectral resolution; (ii) divided into
0.005 ppm bins; and (iii) divided into 148 bins with man-ually selected integral regions, chosen such that, as far as
possible and considering all spectra simultaneously, each
manually selected bin included resonances from a singlemetabolite only, and all resonances in the spectra were
included (i.e. this does not equate to a variable selection
procedure). This reduces the effects of peak shifts betweenspectra (as might be caused by slight pH variation, for
instance), reduces the number of individual resonances
spread across more than one bin, and excludes regionscontaining only noise. The boundaries between the bins are
given in Table S1 (supporting information). Data for
metabolite assignment were taken from previous work,together with additional information from BMRB (Ulrich
et al. 2008).
2.4 Data analysis
All multivariate analyses were carried out using the soft-ware package Simca-P 11.5 (Umetrics UK, Windsor, UK).
Data were pre-treated two ways: (a) for initial multivariateanalyses, i.e. principal components analysis (PCA) of all
data and partial least squares (PLS) regressions of indi-
vidual compounds, we used 3 Hz (0.005 ppm) bins, which
Table 1 Number of replicate microcosms per dose level at differentconcentrations (nominal, mg kg-1 soil) of toxicants dosed toearthworms
Concentration AZ Cd FA
0 8 8 7
5 1
7 1
8 1
9 5
12 1
13 5
14 5
15 1
19 1
20 4
21 1
26 1
29 1
31 1
35 4
43 5
45 1
47 5
59 4
65 1
70 1
76 1
98 1
100 1
105 1
148 5
158 5
222 1
237 1
333 1
356 1
500 5
533 5
750 1
800 1
1200 1
Validation of metabolomics for toxic mechanism
123

have previously been used for pattern-recognition of NMR
data, and considered to give advantages over broaderbins (Warne et al. 2000; Viant 2003). Difference profiles
were then calculated for each of the separate toxicant
groups by subtracting the mean control profile (by ‘dif-ference profile’ we mean the equivalent of a difference
spectrum, but for the binned data, not for the full resolu-
tion spectra).The data were then mean-centred, and analysed both
with and without scaling to unit variance. (b) For SIMCAand PCA of the data for the two highest replicated con-
centration levels for each toxicant (Table 1), the manually
selected bins were used. Difference profiles were scaledsuch that the three independent control groups of the ori-
ginal data would have been converted to unit variance
(Malmendal et al. 2006).In this study, we also used mutual information to
investigate relationships between metabolites and external
variables. Mutual information is a general informationtheoretic approach to measure the statistical dependence
between variables. It has been applied in many areas such
as analysis of gene expression data (Steuer et al. 2002;Daub et al. 2004; Meyer et al. 2008), independent com-
ponent analysis (Hyvarinen et al. 2001), and image
processing (Thevenaz and Unser 2000). The conventionalmethod to quantify the linear dependence between vari-
ables is Pearson correlation. However, a vanishing Pearson
correlation does not imply that two variables are indepen-dent (Steuer et al. 2002). Inter-metabolite relationships are
frequently nonlinear (Camacho et al. 2005), and linear
models are also often inadequate to describe biologicaldose-responses (Calabrese 2008; May and Bigelow 2005).
We previously observed nonlinear responses of metabolites
to copper in L. rubellus (Bundy et al. 2008). Therefore, it ishighly desirable to investigate both the linear and nonlinear
dependence between the metabolites and external vari-
ables, especially those nonlinear relations which cannot bedetected by linear measures. Mutual information is able to
detect any type of functional relationship and extends
conventional methods, such as Pearson correlation.Let A be a system with m finite states {a1, a2, …, am}
and pi be a probability of state ai, then the Shannon entropy
H(A) is given as
H"A# $ %Xm
i$1
p"ai# log p"ai# "1#
The Shannon entropy measures the uncertainty of the stateof the system A. The entropy of the system A is zero if
p(aj) = 1 and p(ai) = 0 for all i = j, whereas the entropy
becomes maximal if pi = pj for all i and j. The jointentropy H(A,B) of two systems A and B (with n finite
states) is defined as
H"A;B# $ %Xm;n
i$1;j$1
p"ai; bj# log p"ai; bj# "2#
So we have
H A;B" #&H A" # ' H B" # "3#
The above equation fulfils equality only if A and B areindependent. The mutual information MI(A,B) can be
defined as
MI A;B" # $ H A" # ' H B" # % H A;B" #( 0 "4#
Since mutual information is defined in terms of discretedata, its application to continuous data, such as metabolo-
mics data, requires a binning procedure. Here, wecalculated MI following the method of Daub et al. (2004)
which is based on B-spline functions, and the results were
visualized by plotting MI against Pearson correlation. Theseplots allow one to visually identify individual variables with
high MI and/or correlation for further inspection.
The statistical significance of the MI was estimatedusing the method proposed by Steuer et al. (2002). A
surrogate dataset is generated by random permutations of
the original data. From the mutual information of the ori-ginal dataset MI(X,Y)data, the average value obtained from
surrogate data\MI(Xsurr, Ysurr)[, and its standard devia-
tion rsurr, the significance S can be given as
S $ MI"X; Y#data %\MI"Xsurr; Ysurr#[rsurr
"5#
In this study, S for each variable was estimated by
resampling the data 30 times and then visualized by colour-
scale.
3 Results and discussion
The NMR spectra showed a typical mixture of smallmolecule metabolites, as expected from extracts of earth-
worm tissue (Fig. S1, supporting information). We have
previously assigned metabolites from proton spectra of L.rubellus tissue extracts (Bundy et al. 2008), so will not here
present extensive data on assignment. We did observe a
novel unassigned compound (with triplet resonances at2.98 and 3.15 ppm) in all samples, that we had not previ-
ously seen in earthworm tissue extracts.
We removed the compound HEFS from earthworm tis-sue extracts before NMR analysis, for two reasons: firstly,
as this is found in very high concentration in earthworms,
and has seven separate 1H resonances, it may obscureresponses of other low-concentration metabolites. Sec-
ondly, our preliminary results indicated that, at least for
this set of samples, the tissue extracts were not stable, with
Q. Guo et al.
123

visible changes when the samples were left for several
hours at room temperature (i.e. when running samplesusing an autosampler), including a decrease in HEFS and
increase in resonances from a suspected HEFS degradation
product at 6.60 and 1.16 ppm. We surmised that, given itsamphiphilic properties, HEFS might stabilize macromole-
cules even in the solvent extracts, which could include
degradative enzymes. In addition, we filtered samplesthrough a 10 kDa-cutoff membrane filter. The extracts
treated in this way were stable over the period of acquisi-tion, as observed by NMR.
3.1 The three compounds induced distinct and well-defined metabolic responses at sub-lethal
concentrations
Clear dose-responsive effects were observed for all toxi-
cants at the whole profile level. All exposures were at a
sub-lethal level, but sufficient to cause a significantreduction in an ecologically important endpoint (repro-
duction, as measured by rate of cocoon production).
Pattern-recognition analysis showed that there were bio-logical differences between the control groups for the three
toxicants (data not shown), which was not surprising as,
because of logistical constraints, the three toxicant expo-sures were carried out sequentially rather than in parallel.
Because of this, we used difference spectra for further
comparative analyses.Initially, we analysed the effects of each toxicant sepa-
rately. The supervised method partial least squares (PLS)
regression offers the advantage of modelling the multi-variate response across different concentrations, as
opposed to just class discrimination. All three toxicants had
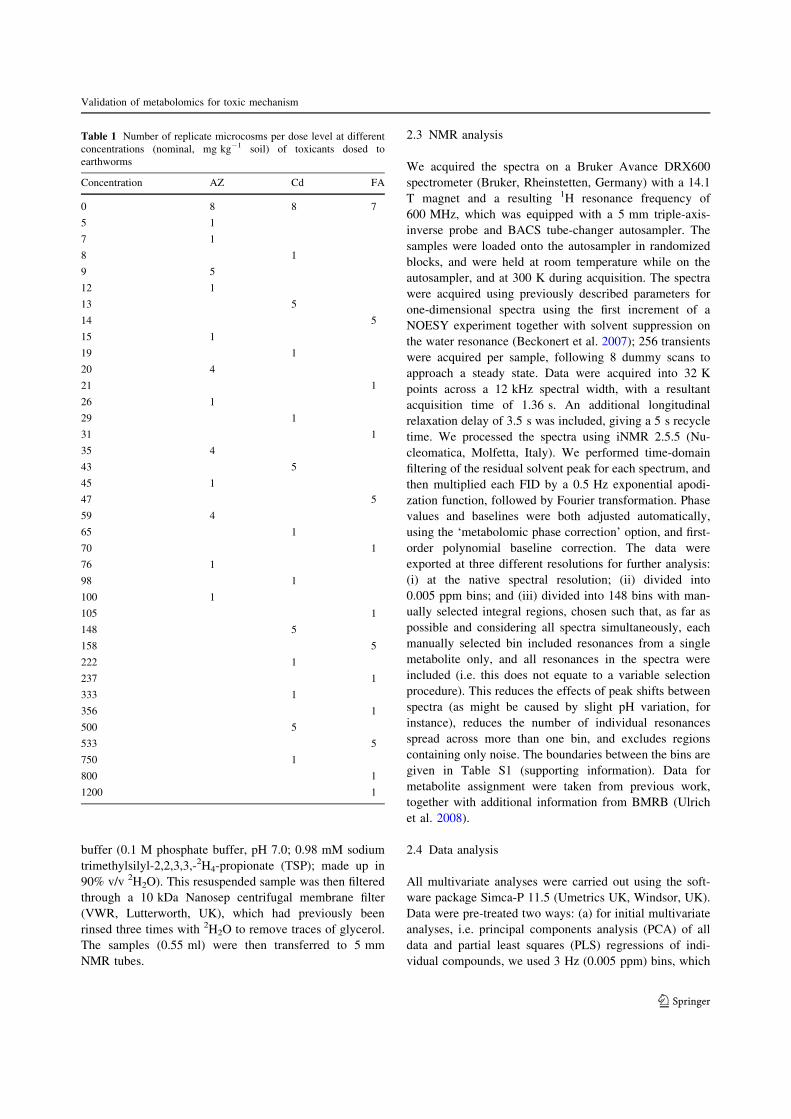
highly significant PLS models for exposure (soil) concen-trations (Fig. 1), for data both scaled and not scaled to unit
variance (not shown). It is important to validate these
models beyond fit alone (Broadhurst and Kell 2006); per-mutation plots show that for each compound, Q2Y
distributions fall to zero as the data are randomized (Fig.
S2, supporting information).In addition to the PLS models for soil toxicant con-
centrations, we fitted models for the tissue concentration
data, shown in Fig. 1d–f as predictions for data from anindependent test set (data for training set are shown in Fig.
S3, supporting information). This was with the aim of
mimicking a more realistic scenario for use with earth-worms sampled from contaminated sites (e.g. during
monitoring of exposure to persistent organic pollutants in
top predators), where (a) tissue concentrations might beused in preference to soil concentrations, because of con-
taminant heterogeneity; and (b) one would want to predict/
classify samples based on existing data. Weak relationshipswere shown for all three toxicants. These were significant
for AZ and Cd (P\ 0.001 and P = 0.009, respectively),
and approached significance at the 5% level for FA(P = 0.076). In our current study, which used well-mixed
soil microcosms, the soil toxicant concentrations were
generally a better indicator of exposure than tissue con-centrations. The tissue levels could have been affected by
either contaminant excretion, for all toxicants, or bio-
transformation, for FA and AZ, either of which phenomenawould underestimate total exposure.
Metabolic profiling can be used as a biomarker discov-ery tool. Environmental biomarkers have great potential as
a tool for effect-based rather than residue-based monitor-
ing—although have also been criticized as not translatingto useful regulatory or monitoring tools (Forbes et al.
2006). Nonetheless, there is interest in ecotoxicogenomic
techniques, including metabolomics, from regulators(Ankley et al. 2006). One reason is that the use of a highly
multivariate profile as an endpoint promises the ability to
distinguish between different chemicals; or, more plausi-bly, different toxicant/MOA classes. Thus, in order to
prove usefulness beyond existing biomarkers, one impor-
tant consideration for environmental metabolomics of L.rubellus is to ask if individual toxicants can be distin-
guished on the basis of their NMR spectral fingerprints.
3.2 Classification of individual samples
An initial unsupervised analysis of all data (PCA) did notseparate the dosed samples into three groups in a
straightforward manner, although there were dose- and
concentration-related effects on different PC axes (sup-porting information, Fig. S4), and so we analysed a sub-set
of the data in more detail. We chose the two highest-con-
centration sets of replicate dosed samples for each toxicant(dose levels given in Table 1), as we assumed the between-
toxicant metabolic differences would be most distinct at the
higher concentrations. A PCA of this reduced dataset didindeed show that the different exposed samples tended to
cluster in toxicant-specific groups (Fig. 2a); in addition, the
higher-concentration dose samples fell further from theorigin than the lower-concentration samples. Given that
PCA alone is not a classification method, we also used
SIMCA (statistical isolinear multiple component analysis;or, soft independent modelling of class analogy (Wold
1974, as cited in Duewer et al. 1975; Wold 1976)) to
perform a simple classification.Our initial SIMCAwas based on a PCA (two components)
of each separate toxicant. Figure 2b shows, unsurprisingly,
that each toxicant model successfully predicted all its ownsamples (with the exception of one AZ sample at the higher
dose level, which fell outside all models, and is not shown in
the figure). Four samples were classed as belonging to allthreemodels (two FA and twoCd, both from the lower dose).
Validation of metabolomics for toxic mechanism
123
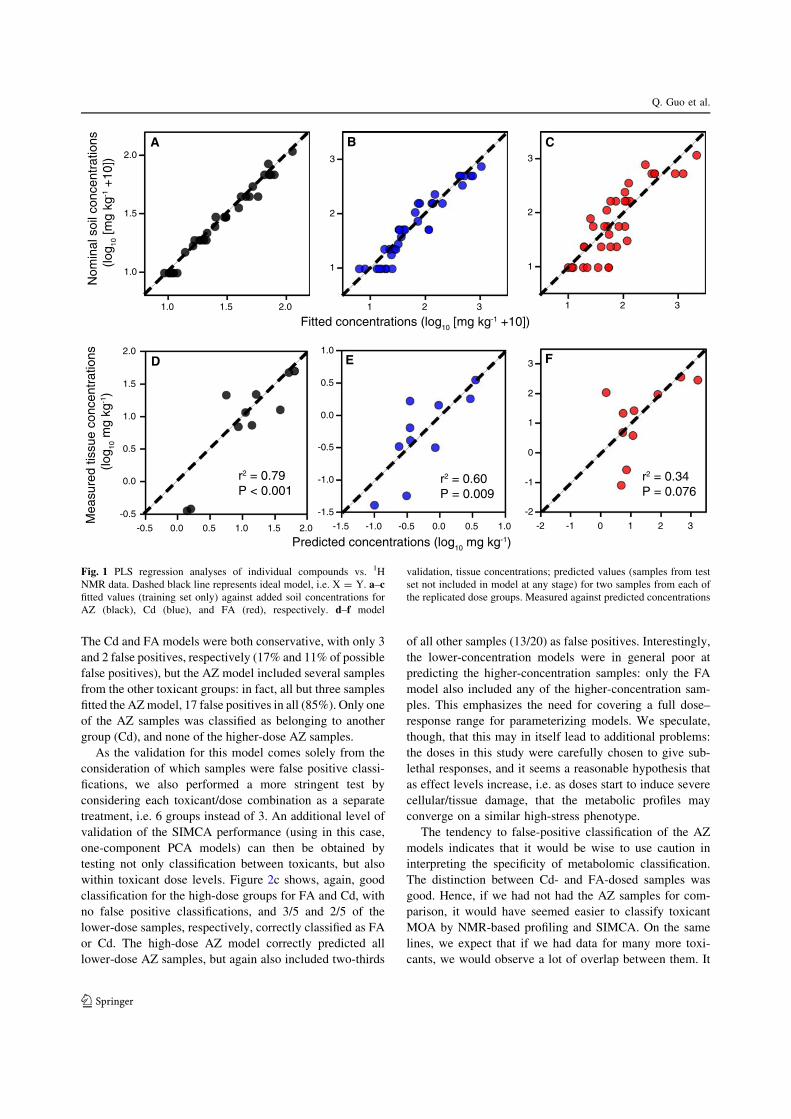
The Cd and FA models were both conservative, with only 3
and 2 false positives, respectively (17% and 11% of possiblefalse positives), but the AZ model included several samples
from the other toxicant groups: in fact, all but three samples
fitted the AZmodel, 17 false positives in all (85%). Only oneof the AZ samples was classified as belonging to another
group (Cd), and none of the higher-dose AZ samples.
As the validation for this model comes solely from theconsideration of which samples were false positive classi-
fications, we also performed a more stringent test by
considering each toxicant/dose combination as a separatetreatment, i.e. 6 groups instead of 3. An additional level of
validation of the SIMCA performance (using in this case,
one-component PCA models) can then be obtained bytesting not only classification between toxicants, but also
within toxicant dose levels. Figure 2c shows, again, good
classification for the high-dose groups for FA and Cd, withno false positive classifications, and 3/5 and 2/5 of the
lower-dose samples, respectively, correctly classified as FA
or Cd. The high-dose AZ model correctly predicted alllower-dose AZ samples, but again also included two-thirds
of all other samples (13/20) as false positives. Interestingly,
the lower-concentration models were in general poor atpredicting the higher-concentration samples: only the FA
model also included any of the higher-concentration sam-
ples. This emphasizes the need for covering a full dose–response range for parameterizing models. We speculate,
though, that this may in itself lead to additional problems:
the doses in this study were carefully chosen to give sub-lethal responses, and it seems a reasonable hypothesis that
as effect levels increase, i.e. as doses start to induce severe
cellular/tissue damage, that the metabolic profiles mayconverge on a similar high-stress phenotype.
The tendency to false-positive classification of the AZ
models indicates that it would be wise to use caution ininterpreting the specificity of metabolomic classification.
The distinction between Cd- and FA-dosed samples was
good. Hence, if we had not had the AZ samples for com-parison, it would have seemed easier to classify toxicant
MOA by NMR-based profiling and SIMCA. On the same
lines, we expect that if we had data for many more toxi-cants, we would observe a lot of overlap between them. It
1.0 1.5 2.0
1.0
1.5
2.0
Nom
inal
soi
l con
cent
ratio
ns (
log 10
[mg
kg-1 +
10])
1 2 3
1
2
3
-1.5 -1.0 -0.5 0.0 0.5 1.0-1.5
-1.0
-0.5
0.0
0.5
1.0
1 2 3
1
2
3
-2 -1 0 1 2 3-2
-1
0
1
2
3
Fitted concentrations (log10 [mg kg-1 +10])
Mea
sure
d tis
sue
conc
entr
atio
ns (
log 10
mg
kg-1)
Predicted concentrations (log10 mg kg-1)
D
A
E
B
F
C
r2 = 0.60P = 0.009
r2 = 0.34P = 0.076
r2 = 0.79P < 0.001
-0.5 0.0 0.5 1.0 1.5 2.0-0.5
0.0
0.5
1.0
1.5
2.0
Fig. 1 PLS regression analyses of individual compounds vs. 1HNMR data. Dashed black line represents ideal model, i.e. X = Y. a–cfitted values (training set only) against added soil concentrations forAZ (black), Cd (blue), and FA (red), respectively. d–f model
validation, tissue concentrations; predicted values (samples from testset not included in model at any stage) for two samples from each ofthe replicated dose groups. Measured against predicted concentrations
Q. Guo et al.
123
should be borne in mind that the models presented herewere based on relatively small numbers of samples, and
thus the results should be interpreted with care. For this
particular study, the models for the AZ-dosed samples
tended to misclassify the other toxicant-dosed samples asfalse positives. However it would not be safe to assume that
this would be a widely applicable inference about the
effects of atrazine on L. rubellus biochemistry: more data,from repeated biological experiments using different soil
types etc., would be required before we could draw such
general conclusions.We also note that SIMCA may not be the best choice for
future real-world compound classification: simpler models,e.g. Fisher’s discriminant analysis (Fisher 1936), ideally
based on a few selected biomarker compounds, offer
advantages such as model transparency and transferabilitybetween different labs and users; and, conversely,
approaches which do not assume specific distributions can
more fully model the data complexity (e.g. Flaherty et al.2005; Ebbels et al. 2007; Maere et al. 2008). However,
SIMCA provided proof-of-principle for multivariate com-
pound MOA classification, based on a simple model thatuses all spectral data.
3.3 Responses of specific compounds(biomarker identification)
In addition to the sample-centred pattern-recognitionanalyses described above, we were also interested in a
variable-centred analysis to determine which metabolites
were changing as a result of compound exposure. PLS, likemany data modelling approaches, assumes a linear
dependence of response on predictor variables. However,
in reality, many biomolecular responses can be expected tobe non-linear. Hence we also calculated the mutual infor-
mation (MI) and Pearson correlation (r) between each
variable (i.e. NMR spectral region bin) and the compoundconcentration for that sample, using both nominal and
measured tissue concentrations. The MI gives a measure of
statistical dependence between two variables, which can beuseful where there is a biological relationship which has
zero or low r (Steuer et al. 2002), and has been used for
assessing relationships between metabolites for urinaryprofile data (Bang et al. 2008). Plotting r against MI is an
efficient way to display the results for highly multivariate
data (Daub et al. 2004), as it can be seen at a glance whichvariables represent high MI and also high r, or else high MI
but low r (probable non-linear relationship). Individual
variables can then be selected for more detailed examina-tion of their relationship to the external factor (e.g.
contaminant concentration), and so this approach, as we
have employed it here, is best thought of as a screeningtool. As increasing the spectral resolution increases the
number of points on this type of plot and consequently
hinders interpretation, we used the manually selected binsto minimize the number of variables. Using data on
AZ Cd
FA
B
C
A
AZ
59
AZ
35
Cd
500
Cd
148
FA 5
33
FA 1
58
AZ 59
AZ 35
Cd 500
Cd 148
FA 533
FA 1580
0.25
0.5
0.75
1
-30 -15 0 15 30
PC 1 scores (24% variance explained)
-20
-10
0
10
20
PC
2 s
core
s (1
7% v
aria
nce
expl
aine
d)
Fig. 2 a Principal components analysis of two highest replicateddose groups for three compounds. Small symbol size = lower doselevel, and large symbol size = higher dose level. Black = AZ,blue = Cd, red = FA. b SIMCA analysis of three groups (AZ, Cd,FA). Sample within the appropriate ellipse indicates sample wasclassified as belonging to that compound group. Symbols are the sameas for panel a. c SIMCA analysis of six groups (AZ, Cd, and FA, atboth higher and lower doses). Y axis indicates model, X axis indicatessample. Both size and colour of point indicates proportion of samplesclassified as belonging to that model (1 = all belonging, 0 = nonebelonging), i.e. point indicates the proportion of samples in the jthcolumn belonging to the model in the ith row. Thick black linesindicate within-compound comparisons, i.e. an ‘ideal’ metabolicresponse would have been 100% classification for squares within thethick black lines, and 0% for squares outside the lines
Validation of metabolomics for toxic mechanism
123
individual fitted compounds would have been better still
(Wishart 2008), but was not practicable in the currentstudy. It should also be borne in mind that more data are
required to calculate MI accurately than Pearson r, andideally we would have had more samples, especially for thesingle-toxicant analyses (Camacho et al. 2005; Khan et al.
2007). However, our use of the permutation procedure in
the MI calculation protects against spuriously high valuesof the MI being interpreted as a significant relationship.
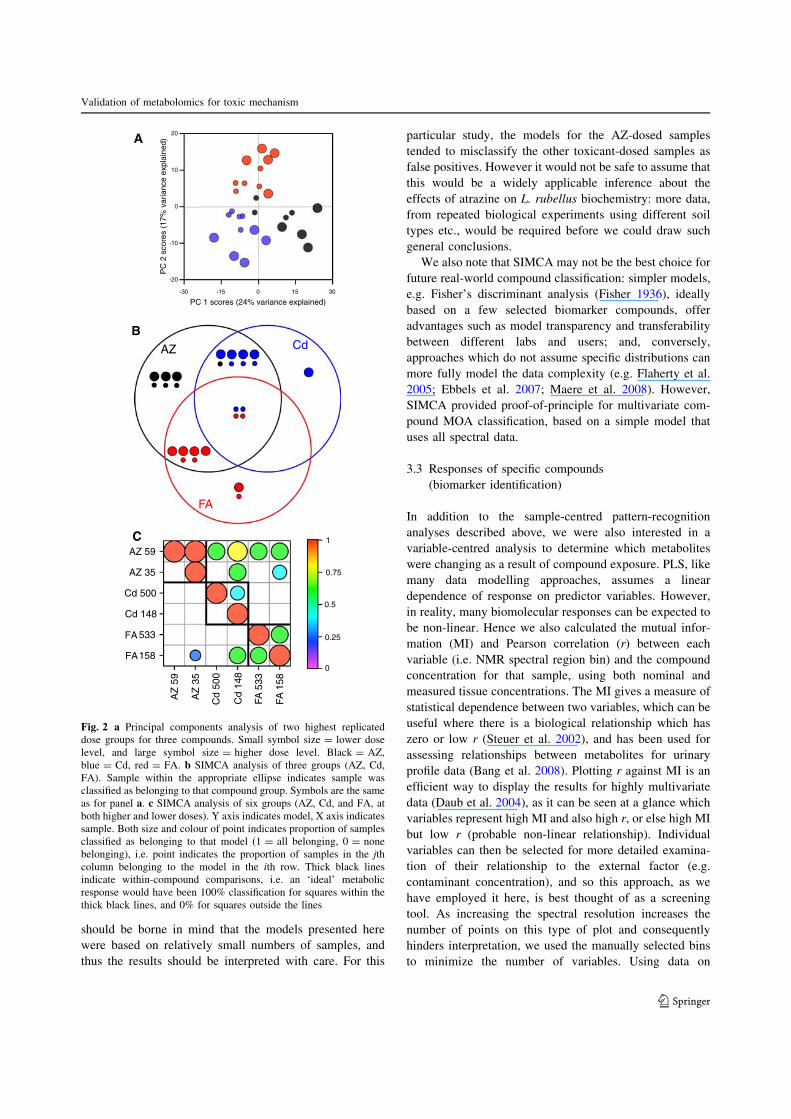
The MI plots show that, when calculated against eithersoil or tissue concentrations, there were some highly sig-
nificant metabolic responses to the three compounds, but
these were largely linear relationships (Fig. 3). For bothtissue and soil AZ concentrations, a variable centred at
1.20 ppm was the most significant; inspection of the
original NMR spectra shows that this region contains a
doublet resonance. We have tentatively assigned this as b-hydroxybutyrate, on the basis of chemical shift, multi-
plicity, and J-coupling. In addition, fumarate (variable at
6.52 ppm) was identified as a significantly responsivemetabolite (Fig. 3a). Atrazine exposure causes upregula-
tion of transcripts associated with the citric acid cycle and
oxidative phosphorylation in L. rubellus, and also a verysignificant upregulation of transcripts related to protein
turnover (Owen et al. 2008). It is possible that the changesobserved here represent some kind of biochemical starva-
tion response induced by atrazine. Cadmium induced two
very clear biochemical responses, a reduction in succinate(variable at 2.41 ppm), and an increase in nicotinic acid
(variable at 8.72 ppm). The responses to FA included a
!-hydroxybutyrate
Fumarate
Succinate2.39
Betaine
CTP
Lactate
CTP
Lactate
A B
C D
E F
Asn
Lys
Asp
DMH
Glucose
Lys
3.39
2.39
3.553.74
3.79
!-hydroxybutyrate
Fumarate
3.79
Malate
mutual information
mutual information mutual information
mutual information
mutual informationmutual information
Pear
son’
s co
rr. c
oeffi
cien
tPe
arso
n’s
corr
. coe
ffici
ent
Pear
son’
s co
rr. c
oeffi
cien
t
Pear
son’
s co
rr. c
oeffi
cien
tPe
arso
n’s
corr
. coe
ffici
ent
Pear
son’
s co
rr. c
oeffi
cien
t
Fig. 3 Linear and non-linearrelationships of metabolites totoxic chemicals: mutualinformation (abscissa) plottedagainst Pearson correlation r(ordinate). Parabola indicatesthe MI expected for linearrelationship; points with highMI but low correlation are likelyto show strong non-linearrelationships againstcontaminants. Colour scaleindicates significance of MI. aAZ (soil concentrations). b AZ(tissue concentrations). c Cd(soil concentrations). d Cd(tissue concentrations). e FA(tissue concentrations). f FA(soil concentrations)
Q. Guo et al.
123
variable at 4.11 ppm (lactate; the methyl resonance at
1.33 ppm was not here identified as significantly different,presumably because at neutral pH this overlaps with a
resonance from threonine in 1D NMR spectra), and a group
of variables at 6.00, 6.13, 7.94, and 7.96 ppm. (The indi-vidual relationships for these variables listed here are all
shown in supporting information, Fig. S5.) These last four
variables correspond to three resonances (7.94 and 7.96represent two halves of a doublet, binned separately
because of partial overlap with another resonance): thechemical shifts, multiplicities, relative intensities, and
J-couplings are all consistent with cytidine triphosphate
(CTP), and the statistical correlation between these threeresonances across all spectra is high, indicating they belong
to a single compound, and so we have assigned this as
CTP. (The 7.4 Hz J-coupling of the resonance at 6.13 ppmis particularly diagnostic, indicating this is not an anomeric
ribose proton resonance.) This analysis clearly shows that
the response of L. rubellus to the three toxicants does notjust involve the same metabolites in each case, again
supporting the future potential of metabolomics for MOA
classification in earthworms.Multivariate analysis of sample differences (pattern
recognition) is especially useful as a screening tool for
metabolic biomarker discovery. However, actual imple-mentation of the results may well be better done through
selection of a small number of robust biomarkers, rather
than using the entire profile (although there may, of course,also be advantages to using the whole profile). Here, a
simple scatterplot of just two metabolites identified from
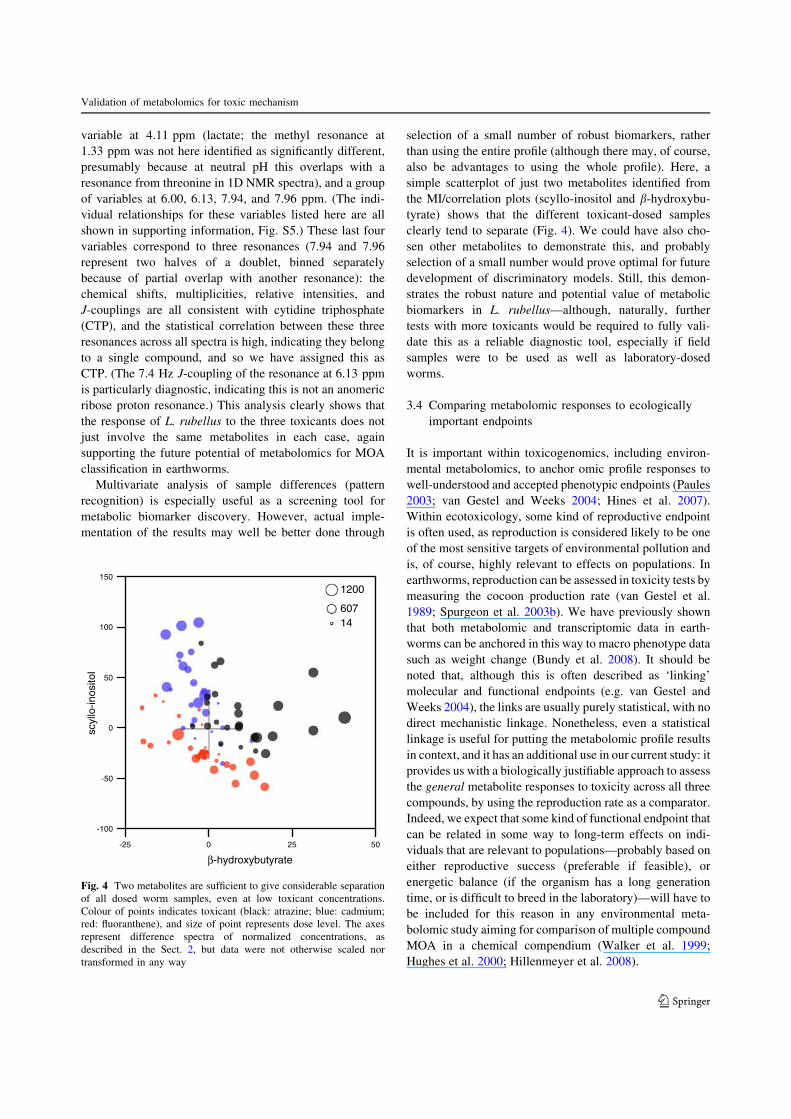
the MI/correlation plots (scyllo-inositol and b-hydroxybu-tyrate) shows that the different toxicant-dosed samples
clearly tend to separate (Fig. 4). We could have also cho-
sen other metabolites to demonstrate this, and probablyselection of a small number would prove optimal for future
development of discriminatory models. Still, this demon-strates the robust nature and potential value of metabolic
biomarkers in L. rubellus—although, naturally, further
tests with more toxicants would be required to fully vali-date this as a reliable diagnostic tool, especially if field
samples were to be used as well as laboratory-dosed
worms.
3.4 Comparing metabolomic responses to ecologically
important endpoints
It is important within toxicogenomics, including environ-
mental metabolomics, to anchor omic profile responses towell-understood and accepted phenotypic endpoints (Paules
2003; van Gestel and Weeks 2004; Hines et al. 2007).
Within ecotoxicology, some kind of reproductive endpointis often used, as reproduction is considered likely to be one
of the most sensitive targets of environmental pollution and
is, of course, highly relevant to effects on populations. Inearthworms, reproduction can be assessed in toxicity tests by
measuring the cocoon production rate (van Gestel et al.
1989; Spurgeon et al. 2003b). We have previously shownthat both metabolomic and transcriptomic data in earth-
worms can be anchored in this way to macro phenotype data
such as weight change (Bundy et al. 2008). It should benoted that, although this is often described as ‘linking’
molecular and functional endpoints (e.g. van Gestel and
Weeks 2004), the links are usually purely statistical, with nodirect mechanistic linkage. Nonetheless, even a statistical
linkage is useful for putting the metabolomic profile results
in context, and it has an additional use in our current study: itprovides us with a biologically justifiable approach to assess
the general metabolite responses to toxicity across all three
compounds, by using the reproduction rate as a comparator.Indeed, we expect that some kind of functional endpoint that
can be related in some way to long-term effects on indi-
viduals that are relevant to populations—probably based oneither reproductive success (preferable if feasible), or
energetic balance (if the organism has a long generation
time, or is difficult to breed in the laboratory)—will have tobe included for this reason in any environmental meta-
bolomic study aiming for comparison of multiple compound
MOA in a chemical compendium (Walker et al. 1999;Hughes et al. 2000; Hillenmeyer et al. 2008).
-25 0 25 50
!-hydroxybutyrate
-100
-50
0
50
100
150
scyl
lo-in
osito
l
1200
60714
Fig. 4 Two metabolites are sufficient to give considerable separationof all dosed worm samples, even at low toxicant concentrations.Colour of points indicates toxicant (black: atrazine; blue: cadmium;red: fluoranthene), and size of point represents dose level. The axesrepresent difference spectra of normalized concentrations, asdescribed in the Sect. 2, but data were not otherwise scaled nortransformed in any way
Validation of metabolomics for toxic mechanism
123
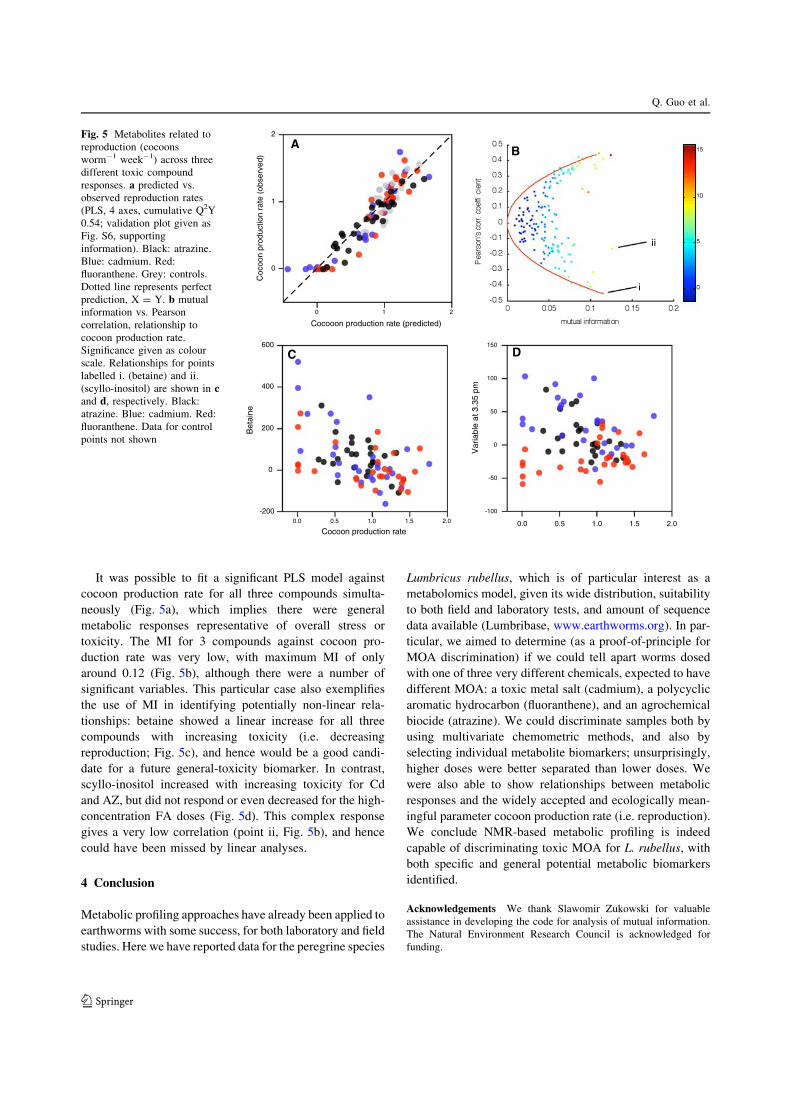
It was possible to fit a significant PLS model againstcocoon production rate for all three compounds simulta-
neously (Fig. 5a), which implies there were general
metabolic responses representative of overall stress ortoxicity. The MI for 3 compounds against cocoon pro-
duction rate was very low, with maximum MI of only
around 0.12 (Fig. 5b), although there were a number ofsignificant variables. This particular case also exemplifies
the use of MI in identifying potentially non-linear rela-
tionships: betaine showed a linear increase for all threecompounds with increasing toxicity (i.e. decreasing
reproduction; Fig. 5c), and hence would be a good candi-
date for a future general-toxicity biomarker. In contrast,scyllo-inositol increased with increasing toxicity for Cd
and AZ, but did not respond or even decreased for the high-
concentration FA doses (Fig. 5d). This complex responsegives a very low correlation (point ii, Fig. 5b), and hence
could have been missed by linear analyses.
4 Conclusion
Metabolic profiling approaches have already been applied to
earthworms with some success, for both laboratory and field
studies. Here we have reported data for the peregrine species
Lumbricus rubellus, which is of particular interest as ametabolomics model, given its wide distribution, suitability
to both field and laboratory tests, and amount of sequence
data available (Lumbribase, www.earthworms.org). In par-ticular, we aimed to determine (as a proof-of-principle for
MOA discrimination) if we could tell apart worms dosed
with one of three very different chemicals, expected to havedifferent MOA: a toxic metal salt (cadmium), a polycyclic
aromatic hydrocarbon (fluoranthene), and an agrochemical
biocide (atrazine). We could discriminate samples both byusing multivariate chemometric methods, and also by
selecting individual metabolite biomarkers; unsurprisingly,
higher doses were better separated than lower doses. Wewere also able to show relationships between metabolic
responses and the widely accepted and ecologically mean-
ingful parameter cocoon production rate (i.e. reproduction).We conclude NMR-based metabolic profiling is indeed
capable of discriminating toxic MOA for L. rubellus, withboth specific and general potential metabolic biomarkersidentified.
Acknowledgements We thank Slawomir Zukowski for valuableassistance in developing the code for analysis of mutual information.The Natural Environment Research Council is acknowledged forfunding.
0 1 2
Cocooon production rate (predicted)
0
1
2
Coc
oon
prod
uctio
n ra
te (
obse
rved
)
0.0 0.5 1.0 1.5 2.0
Cocoon production rate
-200
0
200
400
600
Bet
aine
0.0 0.5 1.0 1.5 2.0
-100
-50
0
50
100
150
Var
iabl
e at
3.3
5 pm
A B
C D
Fig. 5 Metabolites related toreproduction (cocoonsworm-1 week-1) across threedifferent toxic compoundresponses. a predicted vs.observed reproduction rates(PLS, 4 axes, cumulative Q2Y0.54; validation plot given asFig. S6, supportinginformation). Black: atrazine.Blue: cadmium. Red:fluoranthene. Grey: controls.Dotted line represents perfectprediction, X = Y. b mutualinformation vs. Pearsoncorrelation, relationship tococoon production rate.Significance given as colourscale. Relationships for pointslabelled i. (betaine) and ii.(scyllo-inositol) are shown in cand d, respectively. Black:atrazine. Blue: cadmium. Red:fluoranthene. Data for controlpoints not shown
Q. Guo et al.
123

References
Ankley, G. T., et al. (2006). Toxicogenomics in regulatory ecotoxi-cology. Environmental Science and Technology, 40, 4055–4065.doi:10.1021/es0630184.
Bang, J. W., et al. (2008). Integrative top-down system metabolicmodeling in experimental disease states via data-driven Bayesianmethods. Journal of Proteome Research, 7, 497–503. doi:10.1021/pr070350l.
Beckonert, O., et al. (2007). Metabolic profiling, metabolomic andmetabonomic procedures for NMR spectroscopy of urine,plasma, serum and tissue extracts. Nature Protocols, 2, 2692–2703. doi:10.1038/nprot.2007.376.
Broadhurst, D. I., & Kell, D. B. (2006). Statistical strategies for avoidingfalse discoveries in metabolomics and related experiments. Meta-bolomics, 2, 171–196. doi:10.1007/s11306-006-0037-z.
Brown, S. A., Simpson, A. J., & Simpson, M. J. (2008).Evaluation of sample preparation methods for nuclear mag-netic resonance metabolic profiling studies with Eiseniafetida. Environmental Toxicology and Chemistry, 27, 828–836. doi:10.1897/07-412.1.
Bundy, J. G., et al. (2002). Metabonomic assessment of toxicity of 4-fluoroaniline, 3,5-difluoroaniline and 2-fluoro-4-methylaniline tothe earthworm Eisenia veneta (Rosa): Identification of newendogenous biomarkers. Environmental Toxicology and Chem-istry, 21, 1966–1972. doi:10.1897/1551-5028(2002)021\1966:MAOTOF[2.0.CO;2.
Bundy, J. G., et al. (2004). Environmental metabonomics: Applyingcombination biomarker analysis in earthworms at a metalcontaminated site. Ecotoxicology (London, England), 13, 797–806. doi:10.1007/s10646-003-4477-1.
Bundy, J. G., et al. (2007). Metabolic profile biomarkers of metalcontamination in a sentinel terrestrial species are applicableacross multiple sites. Environmental Science and Technology,41, 4458–4464. doi:10.1021/es0700303.
Bundy, J. G., et al. (2008). ‘Systems toxicology’ approach identifiescoordinated metabolic responses to copper in a terrestrial non-model invertebrate, the earthworm Lumbricus rubellus. BMCBiology, 6, 25. doi:10.1186/1741-7007-6-25.
Calabrese, E. J. (2008). Hormesis: Why it is important to toxicologyand toxicologists. Environmental Toxicology and Chemistry, 27,1451–1474. doi:10.1897/07-541.1.
Camacho, D., de la Fuente, A., & Mendes, P. (2005). The origin ofcorrelations in metabolomics data. Metabolomics, 1, 53–63. doi:10.1007/s11306-005-1107-3.
Daub, C. O., Steuer, R., Selbig, J., & Kloska, S. (2004). Estimatingmutual information using B-spline functions—an improvedsimilarity measure for analysing gene expression data. BMCBioinformatics, 5, 118. doi:10.1186/1471-2105-5-118.
Duewer, D. L., Kowalski, B. R., & Schatzki, T. F. (1975). Sourceidentification of oil spills by pattern recognition analysis ofnatural elemental composition. Analytical Chemistry, 47, 1573–1583. doi:10.1021/ac60359a051.
Ebbels, T. M., et al. (2007). Prediction and classification of drugtoxicity using probabilistic modeling of temporal metabolic data:The consortium on metabonomic toxicology screening approach.Journal of Proteome Research, 6, 4407–4422. doi:10.1021/pr0703021.
Fisher, R. A. (1936). The use of multiple measures in taxonomicproblems. Annals of Eugenics, 7, 179–188.
Flaherty, P., Giaever, G., Kumm, J., Jordan, M. I., & Arkin, A. P.(2005). A latent variable model for chemogenomic profiling.Bioinformatics (Oxford, England), 21, 3286–3293. doi:10.1093/bioinformatics/bti515.
Forbes, V. E., Palmqvist, A., & Bach, L. (2006). The use and misuseof biomarkers in ecotoxicology. Environmental Toxicology andChemistry, 25, 272–280. doi:10.1897/05-257R.1.
Gartland, K. P., Beddell, C. R., Lindon, J. C., & Nicholson, J. K.(1990). A pattern recognition approach to the comparison ofPMR and clinical chemical data for classification of nephrotox-icity. Journal of Pharmaceutical and Biomedical Analysis, 8,963–968. doi:10.1016/0731-7085(90)80151-E.
Gartland, K. P., Beddell, C. R., Lindon, J. C., & Nicholson, J. K.(1991). Application of pattern recognition methods to theanalysis and classification of toxicological data derived fromproton nuclear magnetic resonance spectroscopy of urine.Molecular Pharmacology, 39, 629–642.
Gartland, K. P., Bonner, F. W., & Nicholson, J. K. (1989).Investigations into the biochemical effects of region-specificnephrotoxins. Molecular Pharmacology, 35, 242–250.
Gong, P., et al. (2007). Toxicogenomic analysis provides new insightsinto molecular mechanisms of the sublethal toxicity of 2,4,6-trinitrotoluene in Eisenia fetida. Environmental Science andTechnology, 41, 8195–8202. doi:10.1021/es0716352.
Hillenmeyer, M. E., et al. (2008). The chemical genomic portrait ofyeast: Uncovering a phenotype for all genes. Science, 320, 362–365. doi:10.1126/science.1150021.
Hines, A., Oladiran, G. S., Bignell, J. P., Stentiford, G. D., & Viant,M. R. (2007). Direct sampling of organisms from the field andknowledge of their phenotype: Key recommendations forenvironmental metabolomics. Environmental Science and Tech-nology, 41, 3375–3381. doi:10.1021/es062745w.
Hughes, T. R., et al. (2000). Functional discovery via a compendiumof expression profiles. Cell, 102, 109–126. doi:10.1016/S0092-8674(00)00015-5.
Hyvarinen, A., Karhunne, J., & Oja, E. (2001). IndependentComponent Analysis. New York: Wiley.
Jones, O. A., Spurgeon, D. J., Svendsen, C., & Griffin, J. L. (2008). Ametabolomics based approach to assessing the toxicity of thepolyaromatic hydrocarbon pyrene to the earthworm Lumbricusrubellus.Chemosphere, 71, 601–609. doi:10.1016/j.chemosphere.2007.08.056.
Khan, S., et al. (2007). Relative performance of mutual informationestimation methods for quantifying the dependence among shortand noisy data.Physical Review E: Statistical, Nonlinear, and SoftMatter Physics, 76, 026209. doi:10.1103/PhysRevE.76.026209.
Lin, C. Y., Viant, M. R., & Tjeerdema, R. S. (2006). Metabolomics:Methodologies and applications in the environmental sciences.Journal of Pesticide Science, 31, 245–251. doi:10.1584/jpestics.31.245.
Maere, S., Van Dijck, P., & Kuiper, M. (2008). Extracting expressionmodules from perturbational gene expression compendia. BMCSystems Biology, 2, 33. doi:10.1186/1752-0509-2-33.
Malmendal, A., et al. (2006). Metabolomic profiling of heat stress:Hardening and recovery of homeostasis in Drosophila. AmericanJournal of Physiology: Regulatory, Integrative and Compara-tive Physiology, 291, R205–R212. doi:10.1152/ajpregu.00867.2005.
May, S., & Bigelow, C. (2005). Modeling nonlinear dose–responserelationships in epidemiologic studies: Statistical approaches andpractical challenges. Dose Response, 3, 474–490. doi:10.2203/dose-response.003.04.004.
Meyer, P. E., Lafitte, F., & Bontempi, G. (2008). minet: A R/Bioconductor package for inferring large transcriptional net-works using mutual information. BMC Bioinformatics, 9, 461.doi:10.1186/1471-2105-9-461.
OECD. (1984). Guidelines for the testing of chemicals. 207.Earthworm acute toxicity tests. Paris: OECD.
Validation of metabolomics for toxic mechanism
123

Owen, J., et al. (2008). Transcriptome profiling of developmental andxenobiotic responses in a keystone soil animal, the oligochaeteannelid Lumbricus rubellus. BMC Genomics, 9, 266. doi:10.1186/1471-2164-9-266.
Paules, R. (2003). Phenotypic anchoring: Linking cause and effect.Environmental Health Perspectives, 111, A338–A339.
Sanchez-Hernandez, J. C. (2006). Earthworm biomarkers in ecologicalrisk assessment. Reviews of Environmental Contamination andToxicology, 188, 85–126. doi:10.1007/978-0-387-32964-2-3.
Spurgeon, D. J., Svendsen, C., Kille, P., Morgan, A. J., & Weeks, J.M. (2004). Responses of earthworms (Lumbricus rubellus) tocopper and cadmium as determined by measurement of juveniletraits in a specifically designed test system. Ecotoxicology andEnvironmental Safety, 57, 54–64. doi:10.1016/j.ecoenv.2003.08.003.
Spurgeon, D. J., Svendsen, C., Weeks, J. M., Hankard, P. K.,Stubberud, H. E., & Kammenga, J. E. (2003a). Quantifyingcopper and cadmium impacts on intrinsic rate of populationincrease in the terrestrial oligochaete Lumbricus rubellus.Environmental Toxicology and Chemistry, 22, 1465–1472.doi:10.1897/1551-5028(2003)22\1465:QCACIO[2.0.CO;2.
Spurgeon, D. J., Weeks, J. M., & van Gestel, C. A. M. (2003b). Asummary of eleven years progress in earthworm ecotoxicology.Pedobiologia, 47, 588–606.
Steuer, R., Kurths, J., Daub, C. O., Weise, J., & Selbig, J. (2002). Themutual information: Detecting and evaluating dependenciesbetween variables. Bioinformatics (Oxford, England), 18(Suppl2), S231–S240.
Thevenaz, P., & Unser, M. (2000). Optimization of mutual informa-tion for multiresolution image registration. IEEE Transactionson Image Processing, 9, 2083–2099. doi:10.1109/83.887976.
Ulrich, E. L., et al. (2008). BioMagResBank. Nucleic Acids Research,36, D402–D408. doi:10.1093/nar/gkm957.
van Gestel, C. A., van Dis, W. A., van Breemen, E. M., &Sparenburg, P. M. (1989). Development of a standardized
reproduction toxicity test with the earthworm species Eiseniafetida andrei using copper, pentachlorophenol and 2,4-dichloro-aniline. Ecotoxicology and Environmental Safety, 18, 305–312.doi:10.1016/0147-6513(89)90024-9.
van Gestel, C. A., & Weeks, J. M. (2004). Recommendations of the3rd international workshop on earthworm ecotoxicology, Aar-hus, Denmark, August 2001. Ecotoxicology and EnvironmentalSafety, 57, 100–105.
van Straalen, N. M., & Roelofs, D. (2008). Genomics technology forassessing soil pollution. Journal of Biology (Online), 7, 19. doi:10.1186/jbiol80.
Viant, M. R. (2003). Improved methods for the acquisition andinterpretation of NMR metabolomic data. Biochemical andBiophysical Research Communications, 310, 943–948. doi:10.1016/j.bbrc.2003.09.092.
Viant, M. R. (2007). Metabolomics of aquatic organisms: The new‘omics’ on the block. Marine Ecology Progress Series, 332,301–306. doi:10.3354/meps332301.
Walker, M. G., Volkmuth, W., Sprinzak, E., Hodgson, D., & Klingler,T. (1999). Prediction of gene function by genome-scale expres-sion analysis: Prostate cancer-associated genes. GenomeResearch, 9, 1198–1203. doi:10.1101/gr.9.12.1198.
Warne, M. A., Lenz, E. M., Osborn, D., Weeks, J. M., & Nicholson, J.K. (2000). An NMR-based metabonomic investigation of thetoxic effects of 3-trifluoromethyl-aniline on the earthwormEisenia veneta. Biomarkers, 5, 56–72. doi:10.1080/135475000230541.
Wishart, D. S. (2008). Quantitative metabolomics using NMR. Trendsin Analytical Chemistry, 27, 228–237. doi:10.1016/j.trac.2007.12.001.
Wold, S. (1976). Pattern recognition by means of disjoint principalcomponents models. Pattern Recognition, 8, 127–139. doi:10.1016/0031-3203(76)90014-5.
Q. Guo et al.
123
Top Related
Copyright © 2022 FDOKUMEN

