







Bahasa
Halaman
Hukum

UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE BIOLOGIA
DEPARTAMENTO DE BIOQUÍMICA
Fundamentos de Biologia Molecular
Autores:
Profa. Dra. Carmen Veríssima Ferreira (UNICAMP) Profa. Dra. Fernanda Ramos Gadelha (UNICAMP) Prof. Dr. Alexandre D. Martins Cavagis (UFSCAR)
Ms. Daisy Machado Dr. Antonio Hernandes Chaves-Neto
- Campinas (SP) - 2010

Fundamentos de Biologia Molecular 2
BIOLOGIA MOLECULAR
Biologia molecular é a área de conhecimento que estuda diversos aspectos
da função do material genético (genoma) e seus produtos de expressão, as
proteínas. Na Disciplina Fundamentos de Biologia Molecular serão abordadas todas
as etapas da informação gênica, além das aplicações da biologia molecular em
diversos aspectos na área de saúde e finalmente, como alterações no fluxo da
informação gênica podem causar patologias como câncer.
O genoma caracteriza-se por toda a informação hereditária de um organismo
que está codificada em seu DNA (ou, em alguns vírus, no RNA). Isto inclui tanto os
genes como as sequências não-codificadoras (Figura 1 ). No ser humano o genoma
é constituído de 23 cromossomos diferentes. O genoma é transmitido, com
variações individuais, de geração em geração e determina a espécie do ser vivo.
Desta forma, as doenças hereditárias também estão definidas no genoma. No caso
específico dos humanos, a metade do genoma que se herda provém do pai e a outra
metade, da mãe. No contexto químico, o genoma é definido pela seqüência de
desoxiribonucleotídeos na molécula do ácido desoxiribonucléico (DNA).
Figura 1. Representação simplificada de um cromosso mo. O cromossomo é constituído de duas cromátides, cada uma contendo uma dupla fita de ácido desoxiribonucléico. No cromossomo existem seqüências codificantes e não codificantes (como por exemplo, os íntrons)

Fundamentos de Biologia Molecular 3
ÁCIDOS NUCLEICOS
A elucidação da estrutura tridimensional de proteínas, de ácidos nucleicos e
outras biomoléculas bem como a elucidação do fluxo de informação do gene à
proteína, o esclarecimento das vias metabólicas centrais e dos mecanismos de
conversão de energia, tem contribuído muito, nos últimos anos, para a nossa
compreensão da base molecular da vida. Nesta disciplina abordaremos como
organismos tão diferentes quanto à bactéria Escherichia coli e os seres humanos
utilizam os mesmos blocos de construção para fabricar macromoléculas.
As proteínas, diferentemente das outras macromoléculas lipídeos e
polissacarídeos, não são sintetizadas através da ligação pura e simples de seus
monômeros constituintes, os aminoácidos. No caso das proteínas, os aminoácidos
são ligados uns aos outros obedecendo a uma ordem, cuja informação está contida
em uma outra classe de macromoléculas denominadas ácidos nucleicos (DNA e
RNA).
A informação genética para sintetizar uma proteína com uma sequência
correta de aminoácidos está contida no DNA, através da sequência correta de seus
monômeros, os desoxirribonucleotídeos. Esta informação genética ou gênica é
passada de uma geração para outra, através de um processo denominado
replicação. Esta informação passa do DNA para o RNA através de um processo
denominado transcrição. Finalmente, a informação contida no RNA, através da
sequência de seus nucleotídeos constituintes é passada para a proteína, em um
processo denominado tradução, que é a própria síntese protéica (Figura 2).

Fundamentos de Biologia Molecular 4
Figura 2. Fluxo da informação gênica . O gene é transcrito dando origem a um
RNA primário, o qual para ter sua função biológica necessita sobre modificações
pós-transcrição se tornando maduro (funcional). Em seguida, o RNAm se translocará
para os ribossomos e será traduzido pela maquinaria responsável pela síntese
protéica. Alguns proteínas após serem sintetizadas já se encontram na forma ativa,
outras necessitam sofrer modificações pós-tradução para poderem exercer a função
biológica.
Transcrito primário
RNAm
Proteína inativa
Proteína ativa
TRANSCRIÇÃO
MODIFICAÇÃO PÓS-TRANSCRIÇÃO
MODIFICAÇÃO PÓS-TRADUÇÃO
TRADUÇÃO

Fundamentos de Biologia Molecular 5
Para se entender melhor todos estes processos, devemos conhecer um
pouco sobre os constituintes dos ácidos nucleicos, os desoxirribonucleotídeos e os
ribonucleotídeos, monômeros constituintes de DNA e de RNA, respectivamente.
I. ESTRUTURA E SÍNTESE DO DNA
Os ácidos nucleicos são requeridos para o armazenamento e expressão da
informação genética. Existem dois tipos quimicamente distintos de ácidos nucleicos:
DNA (ácido desoxirribonucléico) e o RNA (ácido ribonucléico).
O DNA está presente não somente nos cromossomos no núcleo dos
organismos eucarióticos, mas também na mitocôndria e nos cloroplastos de
vegetais. As células procarióticas, que não possuem núcleo, possuem um
cromossomo único, mas também podem conter DNA não-cromossômico em forma
de plasmídeo. O DNA é responsável pela codificação das informações, sendo capaz
não somente de replicar-se precisamente cada vez em que a célula se divide, como
também fazer com que a informação que contém seja expressa seletivamente.
O RNA participa na expressão da informação genética armazenada no DNA.
O RNA difere do DNA por apresentar uma molécula polimérica simples de
mononucleotídeos unidos por ligações fosfodiéster. O DNA é consideravelmente
maior que o RNA e contém o açúcar desoxirribose em vez de ribose e a base
nitrogenada timina em vez de uracil.
O DNA é um polidesoxirribonucleotídeo constituído de
monodesoxirribonucleotídeos covalentemente unidos por ligações 3’,5’-fosfodiéster
(o grupo 5’-hidroxila da desoxipentose de um nucleotídeo liga-se ao grupo 3’-
hidroxila da desoxipentose de outro nucleotídeo através de um grupo fosfato), e as
bases nitrogenadas localizadas junto ao esqueleto de desoxirribose-fosfato, são por
convenção, escrita em sequência do 5’-terminal da cadeia ao 3’-terminal (Figura 3 ).

Fundamentos de Biologia Molecular 6
Ligação de hidrogênio entre as bases nitrogenadas
Figura 3: A Cadeia de DNA dupla fita, com a seqüência de nucleotídeos mostrada
sendo escrita na direção 5’-3’. Uma ligação 3’-5’ fosfodiéster é destacada. As bases
A e T estão ligadas por duas pontes de H, e as bases C e G, por tres pontes de H.
Observar que o terminal 5’ encontra-se fosforilado e o terminal 3’ possui um
grupamento hidroxila (OH) livre.
C 5’
C 3’
C 5’
C 3’
Ligação fosfodiéster
3’
5’
3’
5’

Fundamentos de Biologia Molecular 7
Com exceção de uns poucos vírus que contêm DNA de fita simples, o DNA
existe como uma molécula de fita dupla, formando uma dupla hélice. Na dupla
hélice, as duas cadeias são dobradas em torno de um eixo comum, sendo pareadas
de modo antiparalelo – isto é, o 5’-terminal de uma fita é pareado com o 3’-terminal
de outra fita. As bases de uma fita são pareadas de acordo com as bases da
segunda fita, de modo que uma Adenina (A) é sempre pareada com a Timina (T),
enquanto a Citosina (C) é sempre pareada com a Guanina (G). Assim, uma cadeia
polinucleotídica da dupla hélice de DNA é sempre o complemento da outra (Figura
4).
Figura 4: Dupla Hélice do DNA. A dupla hélice do DNA é mantida por interações fracas tais como: pareamento das bases por ligações de hidrogênio, interações hidrofóbicas, interações iônicas e interações de Van der Waals. Outro ponto que pode ser observado é que as fitas sempre estarão na forma antiparalelas e são complementares (G e C; A eT).

Fundamentos de Biologia Molecular 8
Desnaturação de DNA - Efeito hipercrômico
O DNA pode ser desnaturado pela adição de ácidos, bases, remoção de
Mg2+, ou pelo calor. Na desnaturação do DNA pelo calor, as duas fitas se separam,
no entanto, caso haja resfriamento, as duas fitas podem voltar a se parear (processo
chamado de renaturação) (Figura 5A ).
A desnaturação de uma molécula de DNA pela temperatura pode ser
acompanhada por um parâmetro denominado por temperatura de fusão (Tm), que
corresponde a ponto de equivalência na curva de fusão do DNA (gráfico de A260 em
função da temperatura), a absorbância a 260 nm (A260) também aumenta, mostrando
um perfil semelhante ao de uma curva de titulação ácido-base. Este comportamento
consiste no chamado efeito hipercrômico, onde a A260 do DNA dupla fita nativo é
menor que a soma das A260 dos nucleotídeos constituintes, devido às interações
(van der Waals e hidrofóbicas) entre as bases empilhadas no DNA desnaturado,
promovendo nuvens eletrônicas distintas das existentes no DNA nativo (Figura 5B ).
Devido aos diferentes conteúdos dos DNAs temos diferentes valores de
temperaturas de fusão. Assim, por exemplo, a Tm de um DNA viral pode ser maior
que a Tm de um DNA de bactérias. Como a desnaturação do DNA consiste na
abertura das duas fitas, que são mantidas pareadas por pontes de hidrogênio, e
sabendo-se que o pareamento A-T envolve duas pontes e G-C, três pontes, um DNA
com maior conteúdo de G e C, terá uma temperatura de fusão maior. A
determinação da Tm de um DNA pode fornecer uma estimativa de sua composição
de bases. Com aumento controlado da temperatura é possível abrir uma região rica
em A-T, sem comprometer a abertura de trechos ricos em G-C. A Figura 5C mostra
diferentes perfis de desnaturação do DNA da E. coli e do M. phlei. De acordo com as
curvas, podemos supor que o conteúdo de C e G do M. phlei é maior que o da E.
coli, pois a temperatura necessária para que o DNA deste último seja desnaturado, é
maior.

Fundamentos de Biologia Molecular 9
Figura 5. Desnaturação do DNA. (a) Desnaturação e renaturação do DNA; (B)
Efeito hipercrômico; (C) DNAs de fontes diferentes podem requerer temperaturas
diferentes para causar desnaturação dos mesmos. As curvas mostras em (C)
sugerem que o conteúdo de C e G do M. phlei é maior que na E. coli, por isto é
necessária uma temperatura maior, para observamos aumento da absorbância a
260 nm.
Calor
Resfriamentolento
DNA nativo (dupla fita) DNA desnaturado
A
B C
Calor
Resfriamentolento
DNA nativo (dupla fita) DNA desnaturado
A
B C

Fundamentos de Biologia Molecular 10
REPLICAÇÃO DO DNA
Os mecanismos que regulam a replicação do DNA e a divisão celular estão
relacionados. A replicação e o ciclo celular devem ser conectados de tal forma que a
frequência dos ciclos de replicação se ajuste à velocidade do crescimento da célula
e o término da replicação seja relacionado à divisão celular. Devem existir, portanto,
sinais celulares para o crescimento da célula, tais como velocidade de síntese
protéica, resposta à falta de aminoácidos ou fontes de carbono, que induzem os
ciclos de eventos da divisão celular.
Em células eucarióticas, o processo de replicação do DNA ocorre durante a
fase do ciclo celular denominada período de síntese (S). A replicação apresenta
algumas características típicas: a) a replicação ocorre de forma semiconservativa; b)
requer um grande consumo de energia, na forma de nucleotídeos trifosfatados (ATP,
GTP, CTP e TTP); c) a reação catalisada pela DNA polimerase (enzima responsável
para unir um nucleotídeo ao outro) se dá no sentido 5’ para 3’.
Replicação é semiconservativa
A hipótese Watson-Crick propõe que cada fita de dupla hélice do DNA seja
usada como um molde para a replicação das fitas-filhas complementares. Desta
forma, duas moléculas de fitas duplas de DNA, idênticas ao DNA parental, serão
formadas, cada uma contendo uma fita intacta do DNA parental (Figura 6 ).
Figura 6 : Replicação Semiconservativa do DNA. Após a replicação, a dupla hélice
formada conterá uma fita parental pareada com a fita recém-sintetizada (fita nova)

Fundamentos de Biologia Molecular 11
Etapas na Síntese do DNA
1) Início da Replicação
A. Separação das duas fitas de DNA complementares:
Para que as duas fitas da dupla hélice parental de DNA sejam replicadas,
elas devem ser primeiramente separadas, pois as polimerases usam somente DNA
fita simples como molde. Nos procarióticos, a replicação do DNA inicia em uma
sequência de nucleotídeos específica que representa a origem única de replicação.
Nos eucarióticos, a replicação inicia em múltiplos sítios (origens de replicação) ao
longo da hélice de DNA. Estes sítios incluem sequência curtas de pares de bases
AT.
B. Formação da zona de replicação
A medida que as duas fitas se desenrolam e se separam, formam uma
região parecida com um “V” designada forquilha de replicação, onde as proteínas
que participarão da síntese da fita nova, incluindo a DNA polimerase, irão se ligar. A
partir da origem, a replicação prossegue ao longo da fita de DNA. A replicação do
DNA de dupla fita é bidirecional e a forquilha de replicação se move em ambas as
direções a partir da origem.
Algumas proteínas são requeridas para o reconhecimento da origem de
replicação e/ forquilha de replicação, formando um Complexo. Proteínas estas que
são responsáveis por manter a separação das fitas parentais (fitas que serão
utilizadas como molde) e por desenrolar a dupla hélice. Estas proteínas incluem
(Figura 7 ):
- Proteína DNA: é responsável pela separação das fitas, formando regiões
localizadas de DNA de fita simples.
- DNA Helicases: Enzimas que tem como função a quebra das pontes de
hidrogênio entre as bases, separando as duas fitas de DNA. As helicases requerem
energia, fornecida pelo ATP.
- Proteínas SSB ou proteínas que se ligam à DNA fita simples: Também
denominadas de proteínas desestabilizadoras da hélice, ligam-se somente em DNA

Fundamentos de Biologia Molecular 12
fita simples impedindo o “repareamento” da dupla fita. A presença de proteínas SSB,
durante a replicação, é extremamente importante, pois evitam que regiões
despareadas sofram torções. Desta forma, as SSBs garantem uma conformação do
DNA ideal para a replicação e pareamento da fita recém sintetizada, além de
proteger as fitas simples de degradação por nucleases.
- Topoisomerases: são enzimas que promovem quebras transitórias nas
ligações fosfodiésteres, gerando uma forma intermediária, onde a proteína
permanece ligada covalentemente ao DNA, permitindo que as fitas do DNA passem
umas sobre as outras, alterando, assim, o superenrolamento da molécula.
Figura 7: Proteínas responsáveis por manter separação das fitas parentais e
desenrolar da hélice dupla adiante da zona de replicação avançada.
5'
5'
3'
3'
DNA Girase
Helicase
Direção do movimento da
forquilha de replicação
Proteínas que se ligam ao
DNA simples fita (SSB)
SSB
Fita líder (contínua)Fita seguidora
(descontínua)

Fundamentos de Biologia Molecular 13
2) Adição do primer
As DNA polimerases (enzimas responsáveis pela síntese da nova fita do
DNA) não podem iniciar a síntese de uma fita complementar através de um molde
totalmente composto de fita simples. Elas requerem um primer ou iniciador que são
regiões curtas de RNA sintetizadas pela DNA primase ou iniciase (uma RNA
polimerase), estes primers são complementares e antiparalelos ao molde de DNA. A
fita contínua necessita somente de um primer inicial enquanto a fita descontínua
utiliza vários primers (Figura 7 ).
3) Elongação
DNA polimerase é a enzima que faz a síntese de uma nova fita de DNA,
possuindo a capacidade de adicionar nucleotídeos na extremidade 3’OH,
possibilitando o crescimento da cadeia no sentido 5’-3’. São constituídas de três
tipos: DNA polimerase I, DNA polimerase II e DNA polimerase III.
- DNA poli I: possui atividades de síntese (DNA polimerase) e atividade de
degradação 5’-3’ exonuclease, sendo capaz de remover hidroliticamente o primer de
RNA e atividade 3’-5’ exonuclease, importante para que a replicação ocorra livre de
erro (Figura 6).
- DNA poli II: sua principal função em extratos celulares está relacionado aos
processos de reparo do DNA.
- DNA poli III: é a principal enzima responsável pelo elongamento da cadeia
do DNA em procariotos, possuindo atividade de síntese e degradação.
As DNA polimerases responsáveis por copiar o molde de DNA somente são
capazes de ler as seqüências de nucleotídeos no sentido 3’-5’, polimerizando a fita
em direção oposta ao sentido da zona de replicação 5’-3’(continua). Esta fita que
está sendo copiada na direção da zona de replicação é denominada Fita Líder e a
outra fita que está sendo sintetizada na direção oposta (5’- 3’) de modo descontínuo,
é Fita secundária. A fita secundária é sintetizada descontinuamente, em fragmentos
curtos de DNA, denominados de Fragmentos de Okazaki (Figura 8 ).
O alongamento da cadeia de DNA é catalisada pela DNA polimerase III,
utilizando o grupo 3’-hidroxila do primer de RNA como aceptor do primeiro
desoxirribonucleotídeo. A DNA polimerase III começa a adicionar os nucleotídeos ao

Fundamentos de Biologia Molecular 14
longo do molde de fita única, que especifica a sequência de bases da cadeia recém-
sintetizada (Figura 9 ). À medida que o nucleotídeo está sendo adicionado, a DNA
polimerase III faz correção (atividade 3’-5’ exonuclease). É importante para a
sobrevivência dos organismos que a sequência de nucleotídeos seja replicada com
mínimo de erros, para evitar mutações.
A DNA polimerase III continua a sintetizar o DNA até ser bloqueada pela
proximidade ao primer de RNA. Quando isso ocorre, o RNA é excisado e a lacuna
preenchida pela DNA polimerase I que remove os nucleotídeos de RNA. A ligação
fosfodiéster final entre o grupo 5’-hidroxila na cadeia de DNA sintetizada pela DNA
polimerase III e o grupo 3’-hidroxila da cadeia feita pela DNA polimerase I é
catalizada pela DNA-ligase. A junção destas duas fitas de DNA requer energia, a
qual, em seres humanos, é fornecida pela clivagem de ATP em AMP+2Pi.
Figura 8: Alongamento das fitas líder e secundária.
5'
5'
3'
3'
3'5'
DNA Girase
Helicase
Direção do movimento da
forquilha de replicação
Proteínas que se ligam ao
DNA simples fita (SSB)
SSBDNA primase
RNA iniciador
RNA iniciador
DNA polimer.III
DNA Polimer. I
DNA ligase
Fita líder (contínua)Fita seguidora
(descontínua)
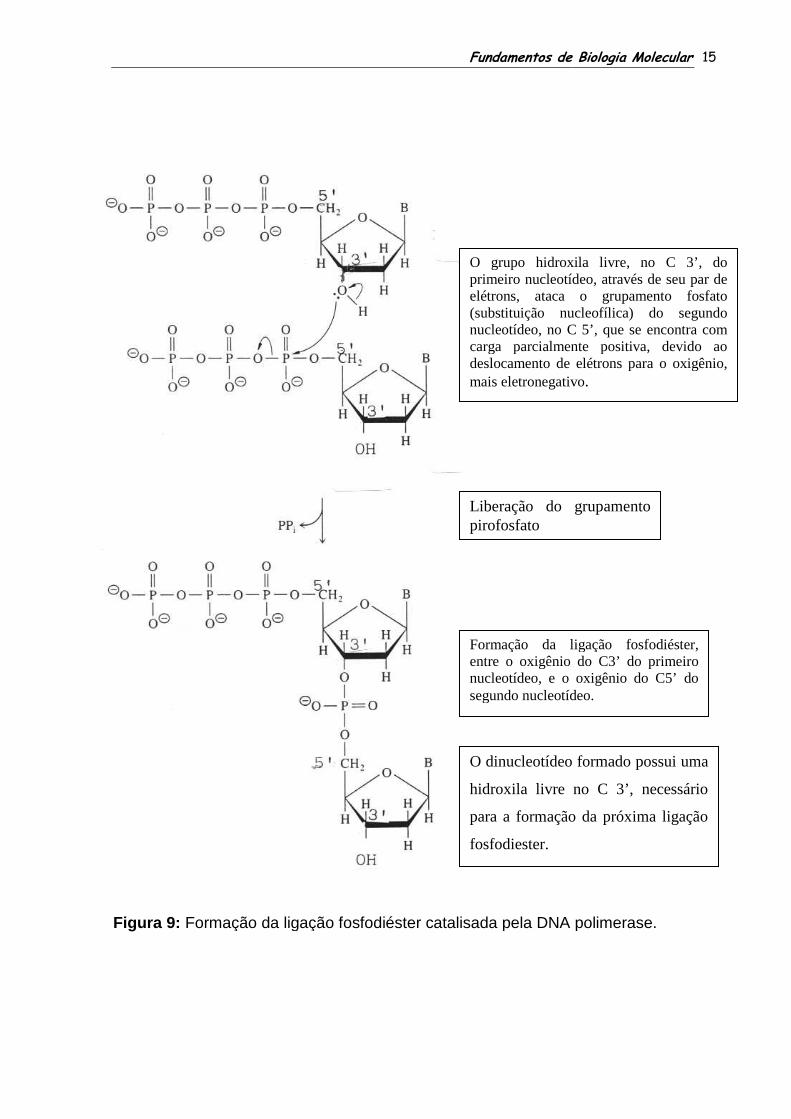
Figura 9: Formação da ligação fosfodiéster catalisada pela DNA polimerase.
Fundamentos de Biologia Molecular
Formação da ligação fosfodiéster catalisada pela DNA polimerase.
O grupo hidroxila livre, no C 3’, do primeiro nucleotídeo, através de seu par de elétrons, ataca o grupamento fosfato (substituição nucleofílica) donucleotídeo, no C 5’, que se encontra com carga parcialmente positiva, devido ao deslocamento de elétrons para o oxigênio, mais eletronegativo
Liberação do grupamento pirofosfato
Formação da ligação fosfodiéentre o oxigênio do C3’ do primeiro nucleotídeo, e o oxigênio do C5’ do segundo nucleotídeo.
O dinucleotídeo formado possui uma
hidroxila livre no C 3’, necessário
para a formação da próxima ligação
fosfodiester.
Fundamentos de Biologia Molecular 15
Formação da ligação fosfodiéster catalisada pela DNA polimerase.
O grupo hidroxila livre, no C 3’, do primeiro nucleotídeo, através de seu par de elétrons, ataca o grupamento fosfato (substituição nucleofílica) do segundo nucleotídeo, no C 5’, que se encontra com carga parcialmente positiva, devido ao deslocamento de elétrons para o oxigênio, mais eletronegativo.
Liberação do grupamento
Formação da ligação fosfodiéster, entre o oxigênio do C3’ do primeiro nucleotídeo, e o oxigênio do C5’ do segundo nucleotídeo.
O dinucleotídeo formado possui uma
hidroxila livre no C 3’, necessário
para a formação da próxima ligação
Top Related
Copyright © 2022 FDOKUMEN

