







Bahasa
Halaman
Hukum


The structure of the Helicobacter pylori ferric uptakeregulator Fur reveals three functional metal binding sitesmmi_7517 1260..1275
Cyril Dian,1†¶ Sylvia Vitale,2‡¶ Gordon A. Leonard,1
Christelle Bahlawane,2,3 Caroline Fauquant,2§
Damien Leduc,4 Cécile Muller,4 Hilde de Reuse,4
Isabelle Michaud-Soret2* and Laurent Terradot1,5*1Structural Biology Group, European SynchrotronRadiation Facility, BP 220 F-38043 Grenoble cedex 9,France.2CNRS UMR 5249 Laboratoire de Chimie et Biologiedes Métaux; Commissariat à l’Energie Atomique (CEA),Direction des Sciences du Vivant (DSV), Institut deRecherches en Technologies et Sciences pour le Vivant(iRTSV); Université Joseph Fourier, 17 avenue desMartyrs, Grenoble F-38054 cedex 9, France.3Institute of Medical Microbiology and HospitalEpidemiology, Hannover Medical School,Carl-Neuberg-Str. 1, 30625 Hannover, Germany.4Institut Pasteur, Unité Pathogenèse de Helicobacter, 28rue Docteur Roux, 75724 Paris Cedex 15, France.5Institut de Biologie et Chimie des Protéines, BiologieStructurale des Complexes MacromoléculairesBactériens, UMR 5086 CNRS Université de Lyon,IFR128, 7 Passage du Vercors, F-69367 Lyon Cedex07, France.
Summary
Fur, the ferric uptake regulator, is a transcriptionfactor that controls iron metabolism in bacteria.Binding of ferrous iron to Fur triggers a conforma-tional change that activates the protein for binding tospecific DNA sequences named Fur boxes. In Helico-bacter pylori, HpFur is involved in acid response andis important for gastric colonization in model animals.Here we present the crystal structure of a functionallyactive HpFur mutant (HpFur2M; C78S-C150S) bound
to zinc. Although its fold is similar to that of other Furand Fur-like proteins, the crystal structure of HpFurreveals a unique structured N-terminal extension andan unusual C-terminal helix. The structure also showsthree metal binding sites: S1 the structural ZnS4 sitepreviously characterized biochemically in HpFur andthe two zinc sites identified in other Fur proteins.Site-directed mutagenesis and spectroscopy analy-ses of purified wild-type HpFur and various mutantsshow that the two metal binding sites common toother Fur proteins can be also metallated by cobalt.DNA protection and circular dichroism experimentsdemonstrate that, while these two sites influence theaffinity of HpFur for DNA, only one is absolutelyrequired for DNA binding and could be responsiblefor the conformational changes of Fur upon metalbinding while the other is a secondary site.
Introduction
Iron is an essential element for most microorganisms and,although relatively abundant in nature, it is not readilyavailable in the environment because of its insolubility inits oxidized form Fe3+. Therefore, bacteria, and in particu-lar pathogens, have developed high-affinity uptakesystems to acquire the iron necessary for growth and hostcolonization. However, intracellular iron concentrationshave to be tightly controled as overload is highly toxicbecause of the Fenton reaction, which results in the for-mation of hydroxyl radicals. Most Gram-negative (andsome Gram-positive) bacteria control iron homeostasisthrough the activity of the iron-responsive transcriptionalregulator Fur (Ferric uptake regulator protein) (Escolaret al., 1999; Hantke, 2001).
As a result of its involvement in bacteria, in the regula-tion of diverse functions as varied as acid tolerance, oxi-dative stress response, metabolic pathways and virulencefactors, Fur represents a global regulator responding toenvironmental iron concentration (Escolar et al., 1999). InEscherichia coli, Fur controls the expression of more than190 genes and was described as a transcription factorthat binds ferrous iron when the cellular iron concentrationbecomes too high (Hantke, 2001; Andrews et al., 2003;Abed et al., 2007). This binding activates the attachementof Fur to specific DNA sequences in promoter regions
Accepted 14 December, 2010. *For correspondence. E-mail [email protected]; Tel. (+33) 4 72 72 26 52; Fax (+33) 4 72 72 26 04and E-mail [email protected]; Tel (+33) 4 38 78 99 40; Fax (+33) 4 3878 54 87. Present addresses: †Laboratoire de Mécanismes Molécu-laires des Infections et Pathologies, Institut de Biologie Structurale(IBS), CNRS UMR 5075, Université Joseph Fourier, CEA DSV, 41 rueJules Horowitz, Grenoble F-38027, France; ‡Gene Center andDepartment of Biochemistry, Ludwig Maximilians University, Feodor-Lynen-Strasse 25, D-81377 Munich, Germany; §Laboratoired’Enzymologie et Biochimie Structurales UPR3082 CNRS, Gif-sur-Yvette, France. Bât 34,Rue de la terrasse, 91198 Gif sur Yvette,France. ¶These authors contributed equally to this work.
Molecular Microbiology (2011) 79(5), 1260–1275 � doi:10.1111/j.1365-2958.2010.07517.xFirst published online 5 January 2011
© 2011 Blackwell Publishing Ltd

called ‘Fur boxes’, repressing expression of genesinvolved in iron uptake (Bagg and Neilands, 1987; deLorenzo et al., 1987; 1988). In some bacterial species,Fur has been reported to positively regulate the expres-sion of certain genes. In E. coli, most positive regulationby Fur is indirect and takes place at the post-transcriptional level through the repression of an anti-sense non-coding regulatory RNA (Masse andGottesman, 2002; Nandal et al., 2010). In Neisseria men-ingitidis, Fur was demonstrated to act as a direct activatorby gene transcription analysis (Grifantini et al., 2003).
In the widespread human gastric pathogen Helico-bacter pylori, Fur (HpFur) plays an important role in theability of H. pylori to colonize mice and Mongolian gerbils(Bury-Moné et al., 2004; Gancz et al., 2006; Miles et al.,2010a). It was found that HpFur regulates more than 50genes in response to iron (Ernst et al., 2005a; Danielliet al., 2006). HpFur is negatively autoregulated and isrequired for the adaptation to iron limitation and for resis-tance to acid and oxidative stresses (Bijlsma et al., 2002).Intringuingly, HpFur is also able to regulate target genes,such as the sodB superoxide dismutase (repression;Ernst et al., 2005b; Miles et al., 2010b) or the pfr ferritin-like iron storage protein (upregulation: Delany et al.,2001a; Danielli et al., 2006), under its apo-form (i.ewithout iron bound). In this regard, it has been suggestedthat the upregulation of some genes is linked to apo-Furpolymerization along the target DNA (Delany et al.,2001a; 2003).
As well as HpFur, two other transcriptional regulatorswere found to be central to the response of H. pylori toacidity: the nickel responsive regulator HpNikR (Bury-Moné et al., 2004) and the two component regulatoryArsRS system (Pflock et al., 2006). Additionally, a uniqueoverlap, in H. pylori, between nickel, iron and acid-regulated genes, has been reported, including autoregu-lation of the genes encoding HpFur and HpNikR (Delanyet al., 2002; Contreras et al., 2003; Bury-Moné et al.,2004). It has been proposed that such a regulationnetwork is necessary to optimize the response of H. pylorito the variable gastric environment in terms of metal ionsconcentration and pH (Danielli and Scarlato, 2010).However, the molecular mechanisms underlying theseregulations are still poorly understood.
HpFur belongs to the large Fur and Fur-like family com-posed of metal uptake regulators that control the homeo-stasis of iron (Fur from E. coli was the first memberdescribed), zinc (Zur), nickel (Nur) and manganese (Mur)and includes the peroxide-sensing protein PerR. Althoughthese proteins show a wide diversity of metal selectivityand biological functions, they share a similar architecturecomprising two domains: a N-terminal DNA-bindingdomain (~ 80 residues; DBD) containing a winged-helixmotif and a C-terminal dimerization domain (~ 70 resi-
dues), which are connected by an ‘hinge’ loop. Structuralinformation is available for two full-length Fur proteins, thePseudomonas aeruginosa PaFur (Pohl et al., 2003) andthe Vibrio cholerae, VcFur (Sheikh and Taylor, 2009) andfor the DBD of E. coli, EcFur (Pecqueur et al., 2006).Crystal structures of other Fur-like proteins have alsobeen described with those of the Mycobacterium tubercu-losis zinc uptake regulator MtZur (Lucarelli et al., 2007),the Bacillus subtilis peroxide-regulon repressor BsPerR(Traore et al., 2006; Jacquamet et al., 2009) and theStreptomyces coelicolor nickel uptake regulator ScNur(An et al., 2009) confirming the structural homologybetween the members of this family.
The crystal structure of PaFur has served as a model tobetter understand the molecular properties of Fur andFur-like regulators. PaFur contains two Zn2+ binding sites,site 1 and 2. Site 1 is found in the C-terminal dimerizationdomain and is hexa-co-ordinated by two histidines, aglutamic acid, a bidentate aspartic acid and a watermolecule. Site 2 is located in the hinge region between theDBD and dimerization domain and exhibits a tetrahedralgeometry with two histidines and two glutamates asligands. X-ray absorption spectroscopy measurements ofiron substitution in Zn-bound PaFur showed that Fe couldreplace the zinc ion at site 1 but not at site 2. Hence, sites1 and 2 were proposed to act as ‘regulatory’ and ‘struc-tural’ binding sites respectively (Pohl et al., 2003). Thisproposal has been challenged on the basis of a mutagen-esis study of several Fur proteins (Lee and Helmann,2007) and an in silico study of EcFur involving free energycalculations and molecular dynamics (Ahmad et al.,2009), which suggested that site 2 is the regulatory siteand that site 1 plays a subsidiary role that remains to beelucidated. This suggestion was supported by the crystalstructure of VcFur that revealed that the geometry of site2 was conserved while site 1 displayed a tetrahedralco-ordination instead of the octahedral environmentdescribed in PaFur (Sheikh and Taylor, 2009). Finally,another type of metal binding site, occupied by Zn2+, ispresent in several Fur (Jacquamet et al., 1998; Althauset al., 1999) and Fur-like proteins. This site, located in thedimerization domain, is present in the purified proteinsand is necessary to maintain the quaternary structure ofFur dimers (Jacquamet et al., 1998; D’Autreaux et al.,2002) and was thus defined as a structural Zn site. In thissite zinc is bound as a Zn2S2(N/O) moiety [EcFur; (Jac-quamet et al., 1998; Gonzalez de Peredo et al., 1999)] oras a ZnS4 tetrahedral species [BsPerR, MtZur; (Traoreet al., 2006; Lucarelli et al., 2007)].
In a recent study, we have used an active HpFur doublemutant (HpFur2M; Cys78Ser/Cys150Ser) to show thatHpFur dimers contain two identical ZnS4 sites. Thesesites, which we named ‘structural S1’, are essential for thedimerization and are co-ordinated by four cysteines (Cys
Crystal structure of Fur from Helicobacter pylori 1261
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

102, Cys 105, Cys 142 and Cys 145) belonging to twopaired CX2C motifs (Vitale et al., 2009). Here we showthat HpFur2M is active in vitro and in vivo and present itscrystal structure at 1.9 Å resolution. The structure con-firms that HpFur adopts a very similar fold to those ofother Fur and Fur-like proteins and shows that HpFur2Mcontains three metal binding sites, designated S1 (struc-tural, see above), S2 and S3. We used the structuralinformation obtained to generate HpFur2M mutants andshow that S2 is required for DNA-binding activity and thatS3 is dispensable for this function. Our experiments firmlyestablish the occupancy of S2 as being necessary for theconformational changes required for HpFur to bind itsoperator sequence. The S3 site is, for the first time, dem-onstrated as being non-essential for DNA binding butmight be involved in determining the strength of HpFurresponse to excess metal.
Results
In vivo functionality of Fur2M
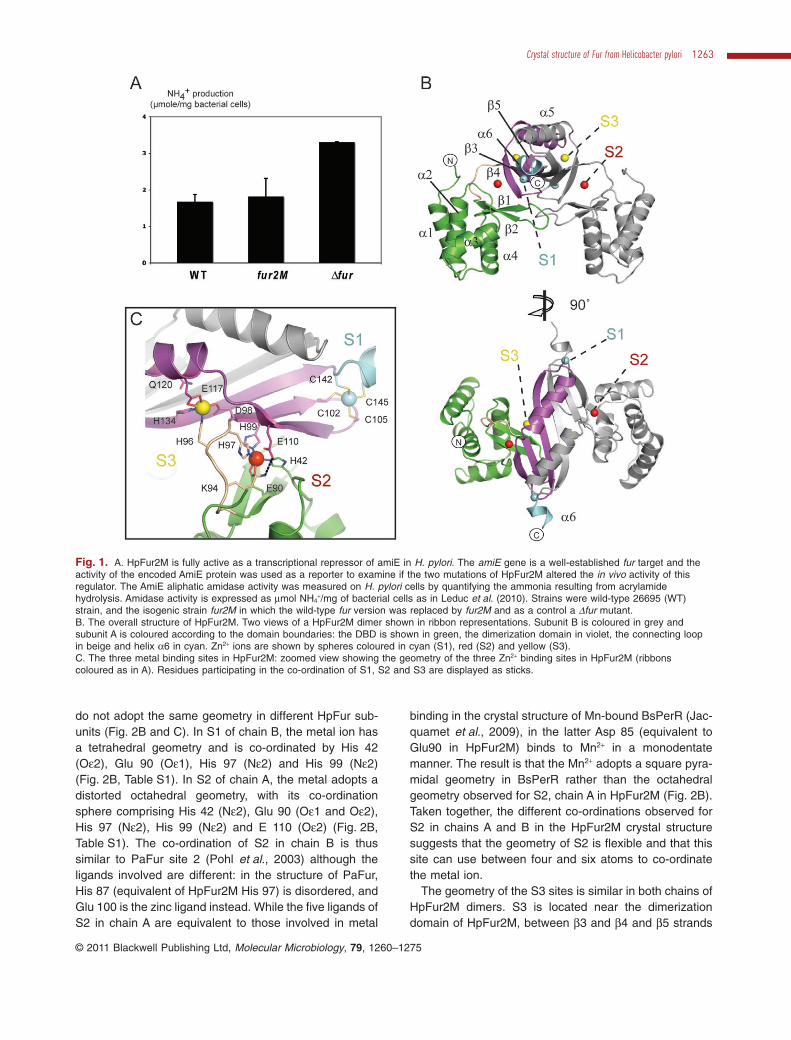
To test the functionality of HpFur2M, an H. pylori strain inwhich the wild-type fur gene was replaced by fur2M wasconstructed. Expression levels of fur and fur2M in the wildtype and mutant strains, respectively, were identical. TheamiE gene (HP0294) is a well-established fur-repressedtarget for which direct binding of Fur to the promoterregion has been demonstrated (van Vliet et al., 2003).amiE encodes an amidase that hydrolyses aliphaticamides to generate the corresponding carboxylic acid andammonia. Acrylamide is one of the best AmiE substrates(Skouloubris et al., 1997). Ammonia produced from acry-lamide hydrolysis was measured in H. pylori wild-typestrain 26695, the isogenic strain 26695-fur2M and, as acontrol, a 26695Dfur mutant (Bury-Moné et al., 2004). TheFur2M strain presented an amidase activity identical tothat of the wild-type strain (Fig. 1A). In contrast, amidaseactivity was enhanced approximately twofold in the Dfurmutant as a result of derepression of the expression of theamiE gene. We also directly measured the amounts ofamiE mRNA in the same three H. pylori strains (Fig. S1).As for the enzymatic activity, we observed that the amountof amiE mRNA expressed in the Fur2M strain was com-parable with that of the wild type while the expression ofamiE was strongly derepressed in the Dfur strain. Theseresults demonstrate that HpFur2M is fully active as atranscriptional repressor in H. pylori.
Crystal structure of HpFur2M
The crystal structure of HpFur2M structure was solved bythe single wavelength anomalous dispersion methodusing the anomalous signal from bound zinc ions and
refined to 1.9 Å resolution. Each asymmetric unit containsfour HpFur2M molecules arranged as two functionaldimers (A/B and C/D). Because the two dimers show onlyvery minor structural variations and contain the samemetal binding sites, the A/B dimer will be used to describethe structure of HpFur2M.
The overall fold of HpFur2M is very similar to that ofother Fur and Fur-like proteins, consisting of a N-terminalDBD (residues 1–92) connected to the C-terminal dimer-ization domain (residues 98–150) by a short hinge region(residues 93–97) (Fig. 1B). The DBD of HpFur2M is com-posed of a winged Helix-Turn-Helix motif (wHTH, helicesa2-a4 and the b1-b2 sheet) with a4 being the DNA rec-ognition helix (Pohl et al., 2003). Unlike other Fur andFur-like proteins, HpFur2M has a ten residues extensionat its N-terminus. This extension results in an elongateda1 helix preceded by five residues that stack against theexternal side of the DBD. The C-terminal dimerizationdomain consists of three antiparallel b-strands (b3, b4 andb5) and two a-helices (a5, a6). This domain is six resi-dues longer than PaFur, and as a result contains the helixa6, a feature previously found only in the structures ofBsPerR and MtZur (Traore et al., 2006; Lucarelli et al.,2007). The dimeric arrangement of HpFur2M is mediatedby b5–b5 interactions between monomers and dimer for-mation results in a six-stranded antiparallel b-sheet beingformed by the association of two HpFur2M dimerizationdomains. Anomalous difference Fouriers coupled withX-ray fluorescence analysis (Leonard et al., 2009) con-firmed the presence of three zinc ions bound per subunit(Fig. S2). Two are embedded in the dimerization domain,while the third is positioned in the hinge region and isco-ordinated by residues from both the DBD and thedimerization domain (Fig. 1B and C).
Metal binding sites of HpFur2M
The structure of HpFur2M contains three Zn2+ bindingsites. One of them, designated S1, was previously iden-tified as a structural site in Vitale et al. (2009). The crystalstructure reported here confirms that at S1 the Zn2+ ion istetra-co-ordinated by two CX2C motifs containing Cys102, Cys 105 and Cys 142, Cys 145 respectively(Fig. 2A). The architecture of S1 in HpFur2M crystal struc-ture corresponds exactly to the ZnS4 site that we previ-ously described and demonstrated to be required for theformation of a HpFur dimers in vitro (Vitale et al., 2009). Inthe crystal structure of HpFur2M, the co-ordination of S1stabilizes the b3-b4-b5 sheet structure found in eachHpFur subunit (Fig. 2A).
The two other Zn2+ binding sites (designated here as S2and S3) observed are homologous to the PaFur zincbinding sites 2 and 1 respectively (Pohl et al., 2003).However, the co-ordination spheres of the Zn2+ ions in S2
1262 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275
do not adopt the same geometry in different HpFur sub-units (Fig. 2B and C). In S1 of chain B, the metal ion hasa tetrahedral geometry and is co-ordinated by His 42(Oe2), Glu 90 (Oe1), His 97 (Ne2) and His 99 (Ne2)(Fig. 2B, Table S1). In S2 of chain A, the metal adopts adistorted octahedral geometry, with its co-ordinationsphere comprising His 42 (Ne2), Glu 90 (Oe1 and Oe2),His 97 (Ne2), His 99 (Ne2) and E 110 (Oe2) (Fig. 2B,Table S1). The co-ordination of S2 in chain B is thussimilar to PaFur site 2 (Pohl et al., 2003) although theligands involved are different: in the structure of PaFur,His 87 (equivalent of HpFur2M His 97) is disordered, andGlu 100 is the zinc ligand instead. While the five ligands ofS2 in chain A are equivalent to those involved in metal
binding in the crystal structure of Mn-bound BsPerR (Jac-quamet et al., 2009), in the latter Asp 85 (equivalent toGlu90 in HpFur2M) binds to Mn2+ in a monodentatemanner. The result is that the Mn2+ adopts a square pyra-midal geometry in BsPerR rather than the octahedralgeometry observed for S2, chain A in HpFur2M (Fig. 2B).Taken together, the different co-ordinations observed forS2 in chains A and B in the HpFur2M crystal structuresuggests that the geometry of S2 is flexible and that thissite can use between four and six atoms to co-ordinatethe metal ion.
The geometry of the S3 sites is similar in both chains ofHpFur2M dimers. S3 is located near the dimerizationdomain of HpFur2M, between b3 and b4 and b5 strands
Fig. 1. A. HpFur2M is fully active as a transcriptional repressor of amiE in H. pylori. The amiE gene is a well-established fur target and theactivity of the encoded AmiE protein was used as a reporter to examine if the two mutations of HpFur2M altered the in vivo activity of thisregulator. The AmiE aliphatic amidase activity was measured on H. pylori cells by quantifying the ammonia resulting from acrylamidehydrolysis. Amidase activity is expressed as mmol NH4
+/mg of bacterial cells as in Leduc et al. (2010). Strains were wild-type 26695 (WT)strain, and the isogenic strain fur2M in which the wild-type fur version was replaced by fur2M and as a control a Dfur mutant.B. The overall structure of HpFur2M. Two views of a HpFur2M dimer shown in ribbon representations. Subunit B is coloured in grey andsubunit A is coloured according to the domain boundaries: the DBD is shown in green, the dimerization domain in violet, the connecting loopin beige and helix a6 in cyan. Zn2+ ions are shown by spheres coloured in cyan (S1), red (S2) and yellow (S3).C. The three metal binding sites in HpFur2M: zoomed view showing the geometry of the three Zn2+ binding sites in HpFur2M (ribbonscoloured as in A). Residues participating in the co-ordination of S1, S2 and S3 are displayed as sticks.
Crystal structure of Fur from Helicobacter pylori 1263
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

and helix a5 and corresponds to site 1 in PaFur (Pohlet al., 2003). However, in HpFur2M this Zn2+ ion has atetrahedral geometry and is co-ordinated by His 96 (Ne2),Asp 98 (Od2), Glu 117 (Oe2) and His 134 (Ne2) (Fig. 2C).
In PaFur the Zn2+ ion in this site is also co-ordinated byequivalent residues (His 86, Asp 88, Glu 107, His 124) buthas a distorted octahedral environment because Asp 88 isbidentate and the co-ordination sphere is completed by a
Fig. 2. Comparison of the Zn2+ binding sites in HpFur2M with those of other Fur and Fur-like proteins.A. Detailed representation of the Zn2+ S1 binding site of HpFur2M that modulates dimer formation and stabilization (left). This site is notconserved in PaFur (right, pdb code 1MZB) that displays a salt bridge instead.B. Detailed comparison of S2 in chains A (bottom left) and B (top left) of HpFur2M with the equivalent site in PaFur (top right) and theregulatory metal binding site of BsPerR (Bottom right, pdb code 3F8N).C. Comparison of S3 of HpFur2M (left) with the equivalent site in PaFur (right) showing identical ligands in both sites but with a differentgeometry of the Zn2+ ion. HpFur2M, PaFur and BsPerR dimer are represented as ribbons. Subunits of HpFur PaFur and BsPerR are colouredaccording to the domain boundaries. The DBD is shown in green, the connecting loop in yellow and the dimerization domain in blue. Forclarity, the dimerization domain of subunit B of HpFur is coloured in pink to highlight differences with subunit A. Important residues arerepresented as sticks and coloured by atom type (carbon coloured as for the domains; oxygen, red; nitrogen, blue; sulphur, orange). Spherescoloured in grey, green and red indicate Zn2+, Mn2+ and water respectively. Hydrogen bonds are presented as black dashed lines.D. Sequence alignment of Fur and Fur-like proteins. The amino acid sequences of Fur proteins from H. pylori strain 26695 (HpFur), V.cholerae (VcFur), P. aeruginosa (PaFur), E. coli (EcFur) and of the Fur-like Zur from M. tuberculosis (MtZur), Nur from S. coelicolor (Nur) andB. subtilis PerR (PerR) were aligned using clustalW (Thompson et al., 1994). Secondary structure elements are indicated above HpFursequence and coloured according to Fig. 1B). Residues involved in S1 (or equivalent) are shaded in blue, S2 in red, S3 in yellow. Stars aboveHpFur sequence indicate the two cysteines (C78 and C150) that have been mutated into serines in HpFur2M. Other residues mutated in thisstudy are indicated by dots coloured in red (HpFur2MDS2) or yellow (HpFur2MDS3).
1264 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275
water molecule bound to Gln 110 (Fig. 2C). In HpFur2M,the side-chain of Gln 120 (equivalent of PaFur Gln 110)points towards the Zn2+ ion in S3 (compare the position ofthese residues in Fig. 2C). This observation indicates thatin HpFur2M, S3 might also be able to adopt an octahedralgeometry, which is typical of Mn2+, Co2+ or Fe2+ ions.
Metal binding of HpFur WT, HpFur2M, HpFurDS2 andHpFurDS3
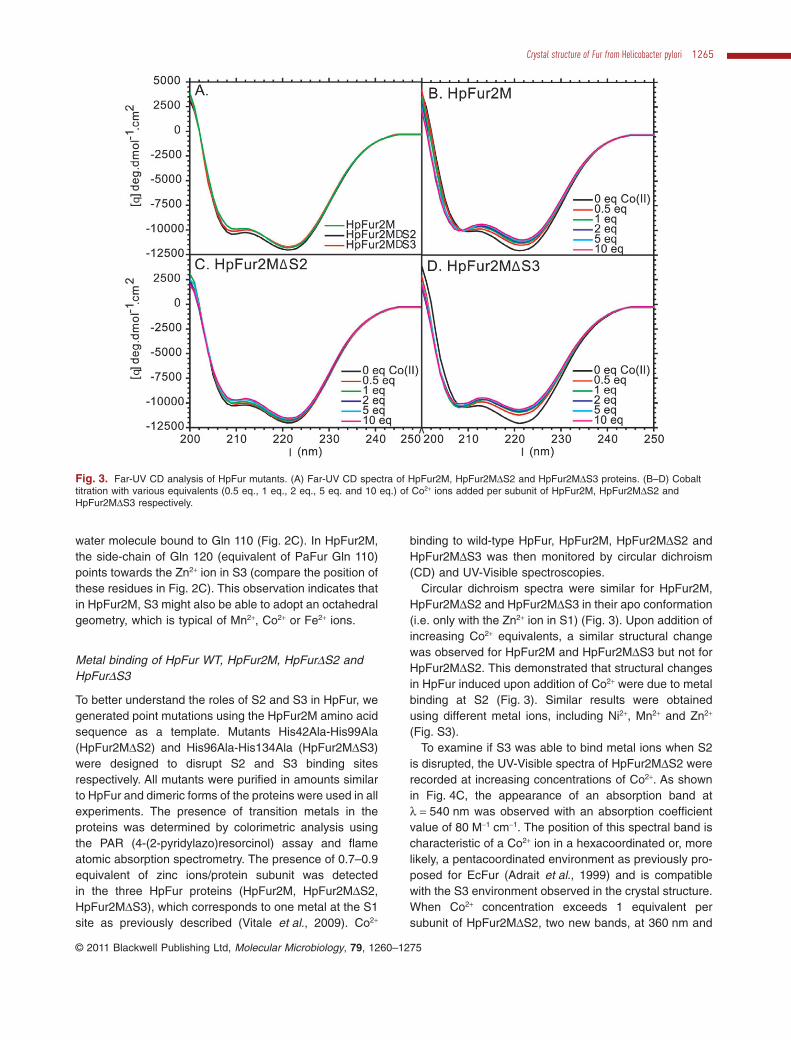
To better understand the roles of S2 and S3 in HpFur, wegenerated point mutations using the HpFur2M amino acidsequence as a template. Mutants His42Ala-His99Ala(HpFur2MDS2) and His96Ala-His134Ala (HpFur2MDS3)were designed to disrupt S2 and S3 binding sitesrespectively. All mutants were purified in amounts similarto HpFur and dimeric forms of the proteins were used in allexperiments. The presence of transition metals in theproteins was determined by colorimetric analysis usingthe PAR (4-(2-pyridylazo)resorcinol) assay and flameatomic absorption spectrometry. The presence of 0.7–0.9equivalent of zinc ions/protein subunit was detectedin the three HpFur proteins (HpFur2M, HpFur2MDS2,HpFur2MDS3), which corresponds to one metal at the S1site as previously described (Vitale et al., 2009). Co2+
binding to wild-type HpFur, HpFur2M, HpFur2MDS2 andHpFur2MDS3 was then monitored by circular dichroism(CD) and UV-Visible spectroscopies.
Circular dichroism spectra were similar for HpFur2M,HpFur2MDS2 and HpFur2MDS3 in their apo conformation(i.e. only with the Zn2+ ion in S1) (Fig. 3). Upon addition ofincreasing Co2+ equivalents, a similar structural changewas observed for HpFur2M and HpFur2MDS3 but not forHpFur2MDS2. This demonstrated that structural changesin HpFur induced upon addition of Co2+ were due to metalbinding at S2 (Fig. 3). Similar results were obtainedusing different metal ions, including Ni2+, Mn2+ and Zn2+
(Fig. S3).To examine if S3 was able to bind metal ions when S2
is disrupted, the UV-Visible spectra of HpFur2MDS2 wererecorded at increasing concentrations of Co2+. As shownin Fig. 4C, the appearance of an absorption band atl = 540 nm was observed with an absorption coefficientvalue of 80 M-1 cm-1. The position of this spectral band ischaracteristic of a Co2+ ion in a hexacoordinated or, morelikely, a pentacoordinated environment as previously pro-posed for EcFur (Adrait et al., 1999) and is compatiblewith the S3 environment observed in the crystal structure.When Co2+ concentration exceeds 1 equivalent persubunit of HpFur2MDS2, two new bands, at 360 nm and
Fig. 3. Far-UV CD analysis of HpFur mutants. (A) Far-UV CD spectra of HpFur2M, HpFur2MDS2 and HpFur2MDS3 proteins. (B–D) Cobalttitration with various equivalents (0.5 eq., 1 eq., 2 eq., 5 eq. and 10 eq.) of Co2+ ions added per subunit of HpFur2M, HpFur2MDS2 andHpFur2MDS3 respectively.
Crystal structure of Fur from Helicobacter pylori 1265
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275
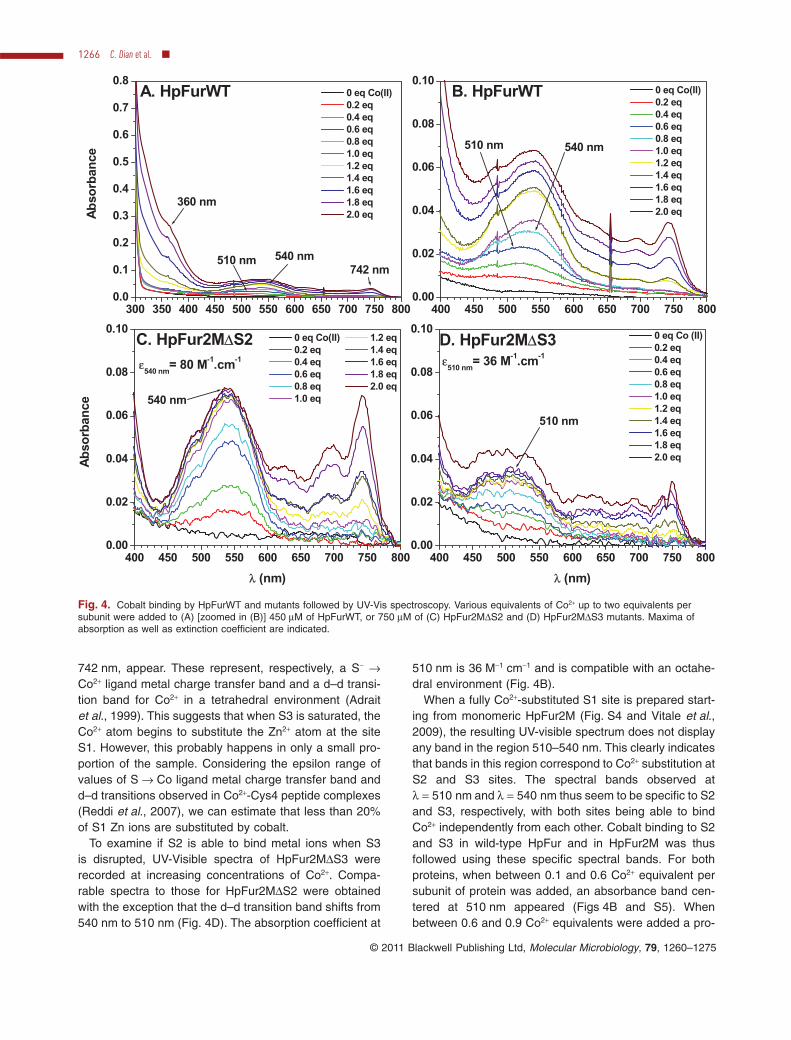
742 nm, appear. These represent, respectively, a S- →Co2+ ligand metal charge transfer band and a d–d transi-tion band for Co2+ in a tetrahedral environment (Adraitet al., 1999). This suggests that when S3 is saturated, theCo2+ atom begins to substitute the Zn2+ atom at the siteS1. However, this probably happens in only a small pro-portion of the sample. Considering the epsilon range ofvalues of S → Co ligand metal charge transfer band andd–d transitions observed in Co2+-Cys4 peptide complexes(Reddi et al., 2007), we can estimate that less than 20%of S1 Zn ions are substituted by cobalt.
To examine if S2 is able to bind metal ions when S3is disrupted, UV-Visible spectra of HpFur2MDS3 wererecorded at increasing concentrations of Co2+. Compa-rable spectra to those for HpFur2MDS2 were obtainedwith the exception that the d–d transition band shifts from540 nm to 510 nm (Fig. 4D). The absorption coefficient at
510 nm is 36 M-1 cm-1 and is compatible with an octahe-dral environment (Fig. 4B).
When a fully Co2+-substituted S1 site is prepared start-ing from monomeric HpFur2M (Fig. S4 and Vitale et al.,2009), the resulting UV-visible spectrum does not displayany band in the region 510–540 nm. This clearly indicatesthat bands in this region correspond to Co2+ substitution atS2 and S3 sites. The spectral bands observed atl = 510 nm and l = 540 nm thus seem to be specific to S2and S3, respectively, with both sites being able to bindCo2+ independently from each other. Cobalt binding to S2and S3 in wild-type HpFur and in HpFur2M was thusfollowed using these specific spectral bands. For bothproteins, when between 0.1 and 0.6 Co2+ equivalent persubunit of protein was added, an absorbance band cen-tered at 510 nm appeared (Figs 4B and S5). Whenbetween 0.6 and 0.9 Co2+ equivalents were added a pro-
400 450 500 550 600 650 700 750 8000.00
0.02
0.04
0.06
0.08
0.10
510 nm 540 nm
B. HpFurWT 0 eq Co(II)0.2 eq0.4 eq0.6 eq0.8 eq1.0 eq1.2 eq1.4 eq1.6 eq1.8 eq2.0 eq
300 350 400 450 500 550 600 650 700 750 8000.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
510 nm 540 nm
360 nm
742 nm
A. HpFurWT 0 eq Co(II)0.2 eq0.4 eq0.6 eq0.8 eq1.0 eq1.2 eq1.4 eq1.6 eq1.8 eq2.0 eqA
bso
rban
ce
400 450 500 550 600 650 700 750 8000.00
0.02
0.04
0.06
0.08
0.10
ε540 nm
= 80 M-1.cm-1
540 nm
C. HpFur2MΔS2
Ab
sorb
ance
λ (nm)
0 eq Co(II) 1.2 eq0.2 eq 1.4 eq0.4 eq 1.6 eq0.6 eq 1.8 eq0.8 eq 2.0 eq1.0 eq
400 450 500 550 600 650 700 750 8000.00
0.02
0.04
0.06
0.08
0.10
ε510 nm
= 36 M-1.cm-1
510 nm
0 eq Co (II)0.2 eq0.4 eq0.6 eq0.8 eq1.0 eq1.2 eq1.4 eq1.6 eq1.8 eq2.0 eq
D. HpFur2MΔS3
λ (nm)
Fig. 4. Cobalt binding by HpFurWT and mutants followed by UV-Vis spectroscopy. Various equivalents of Co2+ up to two equivalents persubunit were added to (A) [zoomed in (B)] 450 mM of HpFurWT, or 750 mM of (C) HpFur2MDS2 and (D) HpFur2MDS3 mutants. Maxima ofabsorption as well as extinction coefficient are indicated.
1266 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275
gressive shift of the maximal absorbance from 510 nm to540 nm transition was observed. Beyond 1 Co2+ equiva-lent, the two absorbances at 360 nm and 742 nm appear,showing that the Co2+ atom begins to replace the Zn2+
atom in structural site S1, while the absorbance at 540 nmcontinues sligthly to increase. These results suggest thatin wild-type HpFur and in HpFur2M Co2+ binds first to S2and then to S3 and to S1.
Role of S2 and S3 in HpFur DNA-binding activity
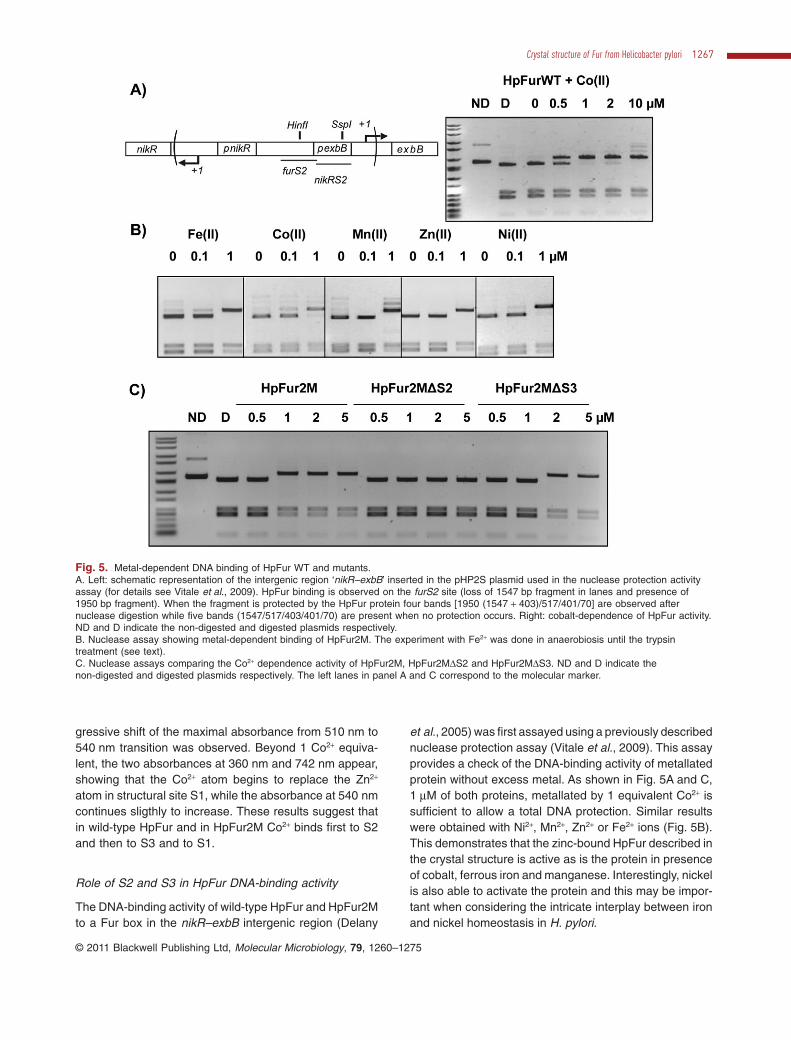
The DNA-binding activity of wild-type HpFur and HpFur2Mto a Fur box in the nikR–exbB intergenic region (Delany
et al., 2005) was first assayed using a previously describednuclease protection assay (Vitale et al., 2009). This assayprovides a check of the DNA-binding activity of metallatedprotein without excess metal. As shown in Fig. 5A and C,1 mM of both proteins, metallated by 1 equivalent Co2+ issufficient to allow a total DNA protection. Similar resultswere obtained with Ni2+, Mn2+, Zn2+ or Fe2+ ions (Fig. 5B).This demonstrates that the zinc-bound HpFur described inthe crystal structure is active as is the protein in presenceof cobalt, ferrous iron and manganese. Interestingly, nickelis also able to activate the protein and this may be impor-tant when considering the intricate interplay between ironand nickel homeostasis in H. pylori.
Fig. 5. Metal-dependent DNA binding of HpFur WT and mutants.A. Left: schematic representation of the intergenic region ‘nikR–exbB’ inserted in the pHP2S plasmid used in the nuclease protection activityassay (for details see Vitale et al., 2009). HpFur binding is observed on the furS2 site (loss of 1547 bp fragment in lanes and presence of1950 bp fragment). When the fragment is protected by the HpFur protein four bands [1950 (1547 + 403)/517/401/70] are observed afternuclease digestion while five bands (1547/517/403/401/70) are present when no protection occurs. Right: cobalt-dependence of HpFur activity.ND and D indicate the non-digested and digested plasmids respectively.B. Nuclease assay showing metal-dependent binding of HpFur2M. The experiment with Fe2+ was done in anaerobiosis until the trypsintreatment (see text).C. Nuclease assays comparing the Co2+ dependence activity of HpFur2M, HpFur2MDS2 and HpFur2MDS3. ND and D indicate thenon-digested and digested plasmids respectively. The left lanes in panel A and C correspond to the molecular marker.
Crystal structure of Fur from Helicobacter pylori 1267
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

In order to identify the roles of S2 and S3 sites in DNAbinding, the DNA-binding profiles of purified HpFur2MDS2and HpFur2MDS3 were also analysed. HpFur2MDS2 wastotally inactive while HpFur2MDS3 showed a twofoldreduction in DNA-binding activity (Fig. 5C). Metal bindingat S2 thus appears to be essential for DNA binding whilethe binding at S3 is important, but not necessary, forHpFur DNA-binding activity.
The ability of purified HpFur, HpFur2M, HpFur2MDS2and HpFur2MDS3 to bind to a hpn2 promoter fragmentusing EMSA was then tested (Fig. S6). Hpn is a nickelstorage protein which production is under Fur control(Ernst et al., 2005a; Gancz et al., 2006). We used a60 bp fragment (-81 to -21 bp relative to the transcrip-tion start site of hpn2) that contains the proposed Furbox (Ernst et al., 2005a). As shown in Fig. S6, HpFurWT,HpFur2M and HpFur2MDS3 but not HpFur2MDS2 areactive in vitro and bind to the 60 bp DNA duplex. Thedissociation constants for HpFurWT, HpFur2M andHpFur2MDS3 were estimated to be of 10.9 � 2, 4 � 1.5and 18.2 � 1.3 nM respectively. HpFur2M exhibits aslightly higher affinity for the target DNA than HpFur,which might be due to its better stability at higher con-centrations. In spite of relatively high variations betweenthe different experiments, as indicated in Fig. S6C bythe error bars, the HpFurD2MS3 protein displayed con-sistently less affinity for the hpn2 duplex than HpFurWTor HpFur2M. This suggests a possible involvement ofthe S3 site in the tuning of the DNA-binding ability ofHpFur. The absence of DNA-binding activity of theHpFur2MDS2 protein confirms the key regulatory role ofthe S2 site.
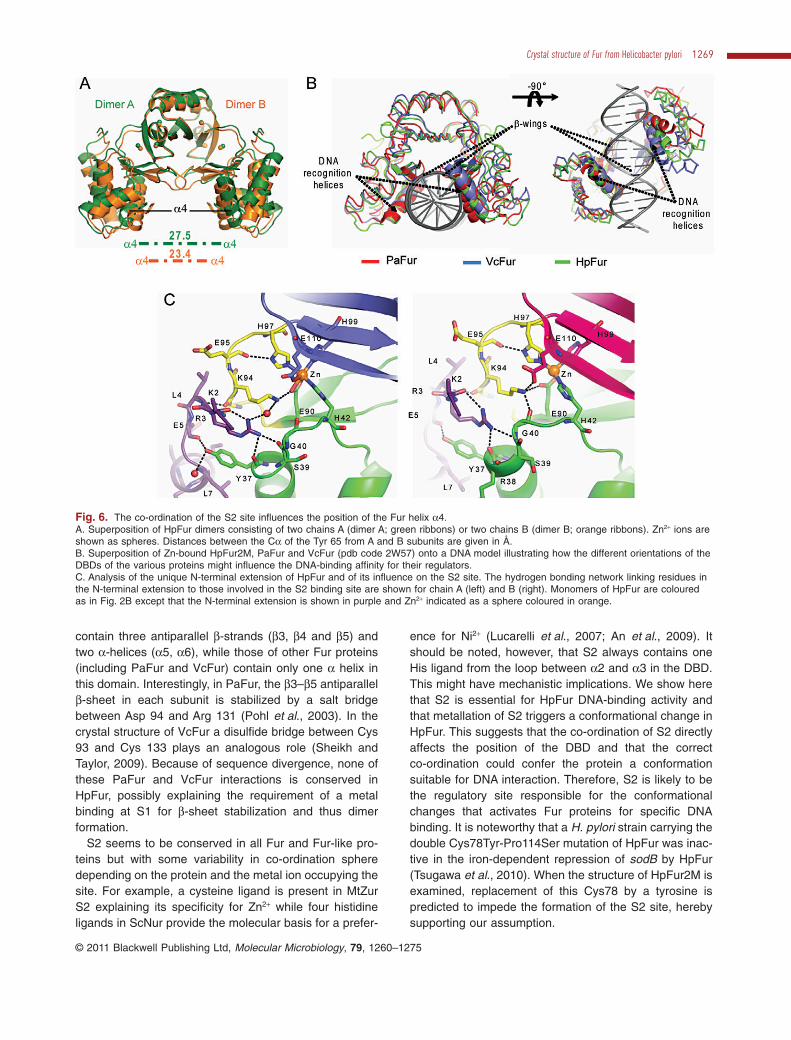
The co-ordination of S2 affects the position of the DBDs
As has been shown above, metal binding at S2 is neces-sary for the DNA-binding activity of HpFur. In the crystalstructure, the conformation observed for the HpFur2Mdimer thus corresponds to the active form of the protein.The HpFur protein dimer has a V shape with the twowHTH motifs forming the arms that act as tweezers on thedsDNA and place a4 recognition helices in contact withtwo major grooves (Fig. 6A and B) (Ahmad et al., 2009;An et al., 2009). In such a conformation, Fur–DNA inter-actions are probably completed by the insertion of theb-wings of the DBD into the DNA minor grooves (Fig. 6B).
In the crystal structure of HpFur2M there is a significantdifference of the organization of the S2 binding sites inchains A and B of the HpFur2M dimer. To examine theimplications of this in detail, models of functional dimersconsisting either of two A chains (AA) or two B chains (BB)were generated and superimposed using solely thedimerization domains as a reference. As shown in Fig. 6A,helices a4 of the two model dimers have different posi-
tions and orientations. Similar differences have beenobserved in the crystal structures of other Fur proteinsthat display heterogeneity in ligand binding within S2(Pohl et al., 2003; Sheikh and Taylor, 2009). Indeed, whendifferent Fur dimers are superimposed using only onesubunit as a reference, a significant difference of a4–a4distances as well as their relative orientation is apparent(Fig. 6B). This suggests that the variations in the archi-tecture of S2 influence the position of the Fur DBDs andpossibly regulate the DNA-binding affinity of Fur proteinsfor the Fur box.
Discussion
In the human pathogen H. pylori, two metalloregulators,NikR and Fur, play a central role in the pH-adaptiveresponse that allows the bacterium to thrive in thestomach (Bury-Moné et al., 2004; van Amsterdam et al.,2006). HpFur regulates an impressive number of genes[59 as described in (Danielli et al., 2006)], upon iron limi-tation and this control is important for the ability of thebacteria to colonize animal models (Bury-Moné et al.,2004; Danielli et al., 2006; Gancz et al., 2006). In addition,HpFur is involved in oxidative stress response (Delanyet al., 2001b; Tsugawa et al., 2010), which adds to thecomplexity of the existing links between the responses todifferent stress conditions. The molecular and structuralproperties of Fur proteins have been investigated, but therole of the metal binding sites identified is still poorlyunderstood. In this work, we have solved and refined thecrystal structure of the active HpFur2M and have per-formed functional assays that, taken together demon-strate the different roles of metal binding sites in Furproteins.
Roles of metal binding sites S1, S2 and S3
Our study provides for the first time, the crystal structureof a Fur protein containing three metal binding sites. Thestructural analysis also highlights a poor conservation ofthese sites in other Fur and Fur-like proteins. In HpFur, S1is a tetrahedral ZnS4 structural site, which stabilizesthe b3-b4-b5 sheet, a crucial element for HpFur dimerformation. Thus, in terms of metal binding, the dimeriza-tion domain of HpFur2M is more similar to BsPerR andMtZur (that also display a S1 site) than to other Furproteins. A Zn2+ site similar to S1 may exist in EcFur butwith a different architecture, as this protein contains onlyone CxxC motif and the nature of this site is ZnS2(N/O)2
and with one histidine as ligand (Jacquamet et al., 1998;Gonzalez de Peredo et al., 1999). The dimerizationdomain of HpFur2M is also structurally more similar tothose of BsPerR and MtZur than to other Fur and Fur-likeproteins. The dimerization domains of these proteins
1268 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275
contain three antiparallel b-strands (b3, b4 and b5) andtwo a-helices (a5, a6), while those of other Fur proteins(including PaFur and VcFur) contain only one a helix inthis domain. Interestingly, in PaFur, the b3–b5 antiparallelb-sheet in each subunit is stabilized by a salt bridgebetween Asp 94 and Arg 131 (Pohl et al., 2003). In thecrystal structure of VcFur a disulfide bridge between Cys93 and Cys 133 plays an analogous role (Sheikh andTaylor, 2009). Because of sequence divergence, none ofthese PaFur and VcFur interactions is conserved inHpFur, possibly explaining the requirement of a metalbinding at S1 for b-sheet stabilization and thus dimerformation.
S2 seems to be conserved in all Fur and Fur-like pro-teins but with some variability in co-ordination spheredepending on the protein and the metal ion occupying thesite. For example, a cysteine ligand is present in MtZurS2 explaining its specificity for Zn2+ while four histidineligands in ScNur provide the molecular basis for a prefer-
ence for Ni2+ (Lucarelli et al., 2007; An et al., 2009). Itshould be noted, however, that S2 always contains oneHis ligand from the loop between a2 and a3 in the DBD.This might have mechanistic implications. We show herethat S2 is essential for HpFur DNA-binding activity andthat metallation of S2 triggers a conformational change inHpFur. This suggests that the co-ordination of S2 directlyaffects the position of the DBD and that the correctco-ordination could confer the protein a conformationsuitable for DNA interaction. Therefore, S2 is likely to bethe regulatory site responsible for the conformationalchanges that activates Fur proteins for specific DNAbinding. It is noteworthy that a H. pylori strain carrying thedouble Cys78Tyr-Pro114Ser mutation of HpFur was inac-tive in the iron-dependent repression of sodB by HpFur(Tsugawa et al., 2010). When the structure of HpFur2M isexamined, replacement of this Cys78 by a tyrosine ispredicted to impede the formation of the S2 site, herebysupporting our assumption.
Fig. 6. The co-ordination of the S2 site influences the position of the Fur helix a4.A. Superposition of HpFur dimers consisting of two chains A (dimer A; green ribbons) or two chains B (dimer B; orange ribbons). Zn2+ ions areshown as spheres. Distances between the Ca of the Tyr 65 from A and B subunits are given in Å.B. Superposition of Zn-bound HpFur2M, PaFur and VcFur (pdb code 2W57) onto a DNA model illustrating how the different orientations of theDBDs of the various proteins might influence the DNA-binding affinity for their regulators.C. Analysis of the unique N-terminal extension of HpFur and of its influence on the S2 site. The hydrogen bonding network linking residues inthe N-terminal extension to those involved in the S2 binding site are shown for chain A (left) and B (right). Monomers of HpFur are colouredas in Fig. 2B except that the N-terminal extension is shown in purple and Zn2+ indicated as a sphere coloured in orange.
Crystal structure of Fur from Helicobacter pylori 1269
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

S3 is present in PaFur, VcFur and MtZur proteins butabsent in BsPerR and ScNur. In contrast to S2, the S3 siteis dispensable for DNA binding by HpFur but its disruptionsignificantly reduces HpFur DNA-binding affinity. Thissuggests that the binding of a metal ion at S3 couldstrengthen the DNA-binding affinity of HpFur for DNA inmetal excess. Indeed, some residues from S2 and S3belong to the same b3 strand, thus binding of metal ionsat both sites could increase the stability of the b-sheet.Further work will be necessary to determine how metalbinding at S3 affects HpFur DNA-binding affinity and/oroperator sequence selectivity. It is also noteworthy thatHpFur responds to many different metal ions in vitro sug-gesting its ability to be activated by Ni2+ as well as Fe2+.This lack of specificity of HpFur for Fe2+ is consistent withthe intricate regulation network existing between the Furand NikR metalloregulators.
A putative role for the unique N-terminal extension ofHpFur in DNA binding
One intriguing feature of HpFur compared with the othermembers of the Fur family is that it can bind to differentpromoters, including Ppfr and Pfur, in the absence of itsiron cofactor (Delany et al., 2001a; 2002; 2003). Theresulting activity of HpFur (i.e. activation or repression)seems to depend on both the position and the sequenceof the given operator (Delany et al., 2003; Lee andHelmann, 2007) and a possible oligomerization of HpFurmight also be involved in this apo-HpFur regulation(Delany et al., 2001a; 2003).
In the crystal structure described here, HpFur displays adense hydrogen bonding network that connects the uniqueN-terminal extension of HpFur to the a2-a3 loop and thehinge region (Fig. 6C). Interestingly, this network is absentin the structures of other Fur proteins. Remarkably, as aconsequence of the variations in S2 co-ordination, thisnetwork is slightly different in the two HpFur subunitsmaking up a dimer. Because of its tetrahedral co-ordination, S2 in subunit B of HpFur dimer is unlikely torepresent the iron-bound form. Nevertheless, in thissubunit, the dimerization domain still connects to thehydrogen bonding network (via Glu 110 and Glu 90 thatinteract with Lys 94). This suggests that, in absence of ironand possibly of any metal, a hydrogen bond networkconnecting the N-terminal extension of HpFur to the a2-a3loop may still form and constrain the hinge position suffi-ciently to stabilize an active conformation, able to bindsome promoter regions, of HpFur. Gene regulation byapo-Fur has not been identified in any other bacteriumand, contrarily to the iron-bound regulation, could notbe complemented by Fur from other species that do notcontain the N-terminal extension (Miles et al., 2010b).Although the role of this unique N-terminal HpFur exten-
sion should be questioned in relation with pH variation andmetal concentration, it is tempting to speculate that it mayindirectly participate in the DNA-binding activity of apo-HpFur, possibly by favouring the V-shape conformation ofHpFur in the absence of metal ion at the regulatory S2 site.
Experimental procedures
Chemicals
Trizma, Bis-Tris Propane, MOPS, and HEPES buffers,Trypsin and metal ion salts were purchased from Sigma-Aldrich.
Cloning and site-directed mutagenesis
The cloning of fur into the expression vector pET11a hasbeen described previously (Vitale et al., 2009). This constructwas used to produce pfur2M plasmid, in which cysteines 78and 150 were mutated to serines (Vitale et al., 2009). The twodouble mutants, His42Ala-His99Ala (HpFur2MDS2) andHis96Ala-His134Ala (HpFur2MDS3), were constructed usingthe QuickChange site-directed mutagenesis kit (Stratagene),using pfur2M as a template with the following primers(mutated base underlined): primers 42f (5′-CGCAGCGGCACAGCCCTAAGCCCTGAAG-3′), 42r (5′-CTTCAGGGCTTAGGGCTGTGCCGCTGCG-3′), 99f (5′-GCGGCTAAAGAACACCATGATGCCATCATTTGTTTGCATTGCGG-3′) and 99r(5′-CCGCAATGCAAACAAATGATGGCATCATGGTGTTCTTTAGCCGC-3′) for HpFur2MDS2, and primers 96f (5′-GCGGCTAAAGAAGCCCATGATCACATCATTTG-3′), 96r (5′-CAAATGATGTGATCATGGGCTTCTTTAGCCGC-3′), 134f (5′-GCCAAGCTGATTAGCGCTGACATGAAAATGTTTGTGTGG-3′) and 134r (5′-CCACACAAACATTTTCATGTCAGCGCTAATCAGCTTGGC-3′) for HpFur2MDS3 mutant. The resultingplasmids were sequenced by Cogenics (Meylan, France).
Protein overexpression and purification
HpFur2MDS2 and HpFur2MDS3 were overexpressed andpurified in the same manner as HpFur and HpFur2M (Vitaleet al., 2009). Protein concentration was determined byUV-Vis spectroscopy at l = 280 nm and the theoreticalabsorption coefficient of 11 460 M-1 cm-1 (http://expasy.org/tools/protparam.html). Approximately 8 mg of pure proteinwas obtained per litre of bacterial culture. All the proteinconcentrations are indicated in subunit concentration. Thepresence of transition metals in purified HpFur proteins wasdetermined by colorimetric analysis using the PAR assay(4-(2-pyridylazo)resorcinol) and by flame atomic absorptionspectrometry (Perkin Elmer 560 Norwalk) using an externalcalibration curve as described in Vitale et al. (2009).
Crystallization, data collection, data processing,structure determination and refinement
Initial crystallization conditions were obtained from the Peg/Ion screen from Hampton Research using the hanging-dropvapour-diffusion method at 20°C. First hits were grown fromcondition N°46 by mixing 1 ml of HpFur2M (30 mg ml-1 of
1270 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

protein in 20 mM HEPES pH 7.4, 200 mM NaCl) previouslymetallated with 1 eq. of ZnSO4/subunit and 1 ml of reservoirsolution. Metallation was carried out in steps of 0.2 eq. toavoid precipitation, which was observed above 1 eq/subunit.Optimized crystals suitable for diffraction data collection grewafter 1 month diffusing against a reservoir solution comprising20% PEG 3350; 100 mM Na citrate. The HpFur2M crystalsobtained belonged to the space group P1 (Table 1) andcontain four molecules per asymmetric unit. Two datasetwere collected at 100 K from a single crystal at the ESRFID29 Beamline; a medium resolution dataset at the peak ofzinc K-absorption (l = 1.2818 Å) used for structure determi-nation; a second high-resolution dataset collected usingX-ray of (l = 0.9762 Å) used for structure refinement. Diffrac-tion images were integrated with either MOSFLM (Leslie,1992) or XDS (Kabsch, 1993). In the case that MOSFLM wasused, intensity data were merged using SCALA (Collabora-tive Computational Project-4, 1994; Evans, 2006). Intensitieswere converted to structure factors and anomalous differ-ences were derived using the program TRUNCATE (Frenchand Wilson, 1978). ShelXD (Schneider and Sheldrick, 2002)was used to find the positions of the Zn2+ ions in the asym-metric unit. These were then used in SHARP (de La Fortelleand Bricogne, 1997) to obtain experimental phases at 2.15 Åresolution using the single wavelength anomalous dispersiontechnique. The experimental phases were improved by auto-matic density modification in the program DM. ARP/wARP(Perrakis et al., 1999) was then used for automatic buildingand tracing (307 residues out of a possible 592) into the
resulting electron density map. The model was completedmanually using COOT (Emsley and Cowtan, 2004) and thensubmitted to TLS restrained refinement using two TLS group(chains A, B and chains C, D) with PHENIX.REFINE (Afonineet al., 2005) (Table 1) This model was used for molecularreplacement using PHASER (McCoy et al., 2007) in the highresolution dataset at 1.85 Å. The final model at high resolu-tion was obtained using a refinement strategy similar to thatused for the starting model. Figures were generated withPymol Software (DeLano, 2002). The co-ordinates and struc-ture factors of HpFur2M have been deposited in the PDB(entry code 2XIG).
HpFur cobalt binding followed by UV/Vis spectroscopy
UV/Vis spectra were acquired using a Hitachi U-3010spectrophotometer. Co2+ binding experiments were per-formed in a 70 ml quartz cuvette (1 cm path length) usingprotein concentrations of 450 mM for HpFur2M and 750 mMfor HpFur2MDS2 and HpFur2MDS3 (in 20 mM HEPESpH 7.4, 500 mM NaCl) and adding small volumes (1 ml) ofCoSO4 at 150 mM. Spectra were recorded every 10 min afterequilibration.
Nuclease protection assay
HpFur binding to the HpFur box in the nikR–exbB intergenicregion were assayed by protection of three engineered HinfI,EcoRI and SspI restriction sites inserted in this region as
Table 1. Data collection, phasing and refinement statistics.
Dataset Peak Remote
ESRF Beamlines ID29 ID29Wavelength (Å) 1.2818 0.9762Number of unique reflections 32 314 51 362Space group P1 P1Cell dimensions a = 47.77 b = 48.01 c = 72.15 a = 48.15 b = 48.2 c = 72.25
a = 83.21° b = 77.79° g = 88.10° a = 83.32° b = 77.82° g = 87.65°Resolution range (Å) 70.29–2.15 (2.21–2.15) 47.96–1.85 (1.95–1.85)Rsymm (%) 8.2 (32.6) 9.8 (38.8)<I/s(I)> 6.5 (2.1) 4.5 (1.8)Completeness (%) 94.7 (94.7) 95.2 (94.8)Multiplicity 5.8 (3.7) 4.1 (4.1)Figure of Merit (DM) 0.846
Refined model compositionSubunit A 2–148 1–22
25–148Subunit B 1–150 1–148Subunit C 2–148 1–150Subunit D 1–148 1–148Zinc ions 12 12
Model quality indicatorsRwork/Rfree (%) 19.45/24.28 19.81/23.39Rmsd bonds (Å)/angle (deg) 0.007/1.030 0.005/0.870Estimated co-ordinate error (Å) 0.57 0.21
Ramachandran AnalysisResidues in Favored Regions [%] 559 [96.21] 563 [96.24]Residues in Generously allowed Regions [%] 20 [3.44] 16 [2.76]Residues in Disallowed Regions [%] 2 [0.34] –
Values in parenthesis refer to the highest resolution shell.Figure of merit: FOM = |Fhkl
best| /|Fhkl|.
Crystal structure of Fur from Helicobacter pylori 1271
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

previously described (Vitale et al., 2009). HpFur proteins at500 mM were incubated with 1 equivalent of various metaldications (Co2+, Mn2+, Fe2+, Zn2+ or Ni2+) per subunit, thendiluted to varying concentrations. The 10 ml reaction mixturecontained varying concentrations of metal substituted-HpFur(0.5 mM, 1 mM, 2 mM or 5 mM) and 200 ng (10 nM final con-centration) of pHP2G plasmid in the 20 mM BisTrisPropanepH 7.5, 100 mM KCl, 5 mM MgCl2 binding buffer. The mixturewas incubated for 20 min at room temperature. An additionalincubation of 1 h at 37°C was then carried out in presence oftwo units of HinfI. After heat inactivation of the enzyme, thereaction mixture was treated with Trypsin (Sigma) at 0.25 mMfor 2 h at 37°C, then mixed with 2 ml of sample loading solu-tion (0.25% bromophenol blue, 0.25% xylene cyanol and30% glycerol). The samples were immediately loaded on a0.8% agarose gel in TAE containing ethidium bromide andthe electrophoresis was run at 100 V for 30 min.
HpFur cobalt binding followed by CD spectroscopy
Far-UV CD spectra (195–250 nm) of the WT HpFur, HpFur2M,HpFur2MDS2 and HpFur2MDS3 mutants were recorded on aChirascan Circular Dichroism Spectrometer (Applied photo-physics) at 25°C in a buffer consisting of 20 mM MOPS pH 7,40 mM KCl. A 1 mm path length cell was used for the mea-surement, and parameters were set as followed: bandwidth,1 nm; step resolution, 1 nm; scan speed, 50 nm min-1;response time, 1 s. Each spectrum was obtained from anaverage of four scans. The protein concentration was 10 mMand increasing amounts of CoSO4 were added: 1/1/2 ml at1.5 mM and 3/5 ml at 3 mM in order to add 0.5/1/2/5 and 10equivalents/subunit of protein respectively. Spectra wererecorded every 10 min after equilibration. Before the calcula-tion of the mean residue molar ellipticity, all of the spectra werecorrected by subtracting buffer contributions.
Bacterial strains and growth conditions
Helicobacter pylori strains were grown on blood agar base 2(Oxoid) plates supplemented with 10% defibrinated horseblood and with a antibiotic–fungicide mix consisting ofvancomycin (final concentration 12.5 mg ml-1), polymyxin B(0.31 mg ml-1), amphotericin B (2.5 mg ml-1) and trimethoprim(6.25 mg ml-1). H. pylori was incubated at 37°C undermicroaerobic conditions. The H. pylori mutant was obtainedby natural transformation as described previously (Schaueret al., 2007) with 2 mg of PCR product. For selection of H.pylori transformants, kanamycin was added to the growthmedium at concentrations of 20 mg ml-1. For amidase activitymeasurements, liquid cultures were in Brain Heart Infusion(Oxoid) supplemented with 4% decomplemented fetal calfserum (Eurobio). H. pylori strains used in this study werewild-type strain 26695 (Tomb et al., 1997), and two isogenicmutants, the previously constructed 29965Dfur (Bury-Monéet al., 2004) and strain 26695 fur2M that was constructed asdetailed below.
Construction of the H. pylori fur2M mutant
The fur2M version was introduced into the chromosome of H.pylori strain 26695 in replacement of the wild-type fur gene byallelic exchange. For this, using the ‘three steps PCR’ proce-
dure described in Stingl et al. (Stingl et al., 2008), we gener-ated a PCR fragment that was composed of three fragmentsin the following order: (i) the entire fur2M gene amplified frompeT11A-Hpfur2M with primers CM9 (CTTTGGAATCCATTT-TAGAGCGC) and CM15 (GTTAGTCACCCGGGTACT-TAACTTTCACTCTCTTGGCA), (ii) a non-polar kanamycinresistance cassette (Menard et al., 1993) and (iii) a fragmentof 600 bp downstream from the fur gene that was amplifiedwith primers CM14 (AGATTGGTGAGTCTTGTTATAGTT)and CM16 (CTGGATGAATTGTTTTAGAAGATTTTAAAAAAGAAGCTTAGA) from DNA of H. pylori 26695. The completePCR product was used to directly transform H. pylori bynatural transformation as previously described (Bury-Monéet al., 2004; Schauer et al., 2007). Correct chromosomalinsertion of the non-polar kanamycin and introduction of thefur2M version on the H. pylori chromosome were verified bysequencing.
Measurements of AmiE amidase activity
The AmiE amidase activity of H. pylori was measured usingthe ammonia assay kit (Sigma) on culture supernatants stan-dardized as follows. H. pylori strains were amplified on platesfor 24 h, grown overnight in liquid medium, and inoculated inliquid culture at OD600 nm = 0.2 and left to grow for 5 h untilOD600 nm = 0.5. These fresh log-phase cells were harvested,washed once in PBS (Roche), and 3 ¥ 108 cfu bacteria (cor-responding to 0.135 mg dry mass of bacterial cells) wereresuspended in 1 ml of buffer with 100 mM of acrylamide, asubstrate of AmiE (Skouloubris et al., 1997). After 30 minincubation at 37°C, bacteria were centrifuged. The NH3 con-centration of the supernatant was measured immediately withthe ammonia assay kit (Sigma) according to the manufactur-er’s recommendations. This assay is based on the followingreaction: in the presence of NH3, a-ketoglutaric acid andNADPH, the enzyme glutamate dehydrogenase producesGlu and NADP+. The oxidation of NADPH to NADP+ results ina decrease in the absorbance at 340 nm that is proportionalto the concentration of NH3. The samples were diluted whennecessary in order to measure NH3 amounts in the region oflinearity of the assay according to the manufacturer’s recom-mendations and the NH3 production was calculated from astandard curve.
Acknowledgements
This work was funded by the ESRF and ANR program ANR-07-BLAN-0083 with funding for the postdoctoral stays of C.Dian, C. Bahlawane and C. Muller. S. Vitale and C. Fauquantwere supported by the CNRS, the CEA and the SFB 900/A forCB. LT is supported by a joint ATIP-Avenir and Ligue Natio-nale Contre le Cancer program. We thank Catherine-MarieDelay for assistance with the in vivo mutagenesis study.
References
Abed, N., Bickle, M., Mari, B., Schapira, M., Sanjuan-Espana,R., Robbe Sermesant, K., et al. (2007) A comparativeanalysis of perturbations caused by a gene knock-out, a
1272 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

dominant negative allele, and a set of peptide aptamers.Mol Cell Proteomics 6: 2110–2121.
Adrait, A., Jacquamet, L., Le Pape, L., Gonzalez de Peredo,A., Aberdam, D., Hazemann, J.L., et al. (1999) Spectro-scopic and saturation magnetization properties of themanganese- and cobalt-substituted Fur (ferric uptake regu-lation) protein from Escherichia coli. Biochem 38: 6248–6260.
Afonine, P.V., Grosse-Kunstleve, R.W., and Adams, P.D.(2005) A robust bulk-solvent correction and anisotropicscaling procedure. Acta Crystallogr D Biol Crystallogr 61:850–855.
Ahmad, R., Brandsdal, B.O., Michaud-Soret, I., and Willas-sen, N.P. (2009) Ferric uptake regulator protein: bindingfree energy calculations and per-residue free energydecomposition. Proteins 75: 373–386.
Althaus, E.W., Outten, C.E., Olson, K.E., Cao, H., andO’Halloran, T.V. (1999) The ferric uptake regulation (Fur)repressor is a zinc metalloprotein. Biochem 38: 6559–6569.
van Amsterdam, K., van Vliet, A.H., Kusters, J.G., and vander Ende, A. (2006) Of microbe and man: determinants ofHelicobacter pylori-related diseases. FEMS Microbiol Rev30: 131–156.
An, Y.J., Ahn, B.E., Han, A.R., Kim, H.M., Chung, K.M., Shin,J.H., et al. (2009) Structural basis for the specialization ofNur, a nickel-specific Fur homolog, in metal sensing andDNA recognition. Nucleic Acids Res 37: 3442–3451.
Andrews, S.C., Robinson, A.K., and Rodriguez-Quinones, F.(2003) Bacterial iron homeostasis. FEMS Microbiol Rev27: 215–237.
Bagg, A., and Neilands, J.B. (1987) Ferric uptake regulationprotein acts as a repressor, employing iron (II) as a cofactorto bind the operator of an iron transport operon in Escheri-chia coli. Biochem 26: 5471–5477.
Bijlsma, J.J., Waidner, B., Vliet, A.H., Hughes, N.J., Hag, S.,Bereswill, S., et al. (2002) The Helicobacter pylori homo-logue of the ferric uptake regulator is involved in acidresistance. Infect Immun 70: 606–611.
Bury-Moné, S., Thiberge, J.M., Contreras, M., Maitournam,A., Labigne, A., and De Reuse, H. (2004) Responsivenessto acidity via metal ion regulators mediates virulence in thegastric pathogen Helicobacter pylori. Mol Microbiol 53:623–638.
Collaborative Computational Project-4 (1994) The CCP4suite: programs for protein crystallography. Acta Crystal-logr D50: 760–763.
Contreras, M., Thiberge, J.M., Mandrand-Berthelot, M.A.,and Labigne, A. (2003) Characterization of the roles ofNikR, a nickel-responsive pleiotropic autoregulator of Heli-cobacter pylori. Mol Microbiol 49: 947–963.
D’Autreaux, B., Touati, D., Bersch, B., Latour, J.M., andMichaud-Soret, I. (2002) Direct inhibition by nitric oxide ofthe transcriptional ferric uptake regulation protein vianitrosylation of the iron. Proc Natl Acad Sci USA 99:16619–16624.
Danielli, A., and Scarlato, V. (2010) Regulatory circuits inHelicobacter pylori: network motifs and regulators involvedin metal-dependent responses. FEMS Microbiol Rev 34:738–752.
Danielli, A., Roncarati, D., Delany, I., Chiarini, V., Rappuoli,
R., and Scarlato, V. (2006) In vivo dissection of the Heli-cobacter pylori Fur regulatory circuit by genome-wide loca-tion analysis. J Bacteriol 188: 4654–4662.
De Lorenzo, V., Herrero, M., Giovannini, F., and Neilands,J.B. (1988) Fur (ferric uptake regulation) protein and CAP(catabolite-activator protein) modulate transcription of furgene in Escherichia coli. Eur J Biochem 173: 537–546.
DeLano, W.L. (2002) The PyMOL Molecular GraphicsSystem [WWW document]. URL http://www.pymol.org/
Delany, I., Spohn, G., Rappuoli, R., and Scarlato, V. (2001a)The Fur repressor controls transcription of iron-activatedand -repressed genes in Helicobacter pylori. Mol Microbiol42: 1297–1309.
Delany, I., Pacheco, A.B., Spohn, G., Rappuoli, R., and Scar-lato, V. (2001b) Iron-dependent transcription of the frpBgene of Helicobacter pylori is controlled by the Fur repres-sor protein. J Bacteriol 183: 4932–4937.
Delany, I., Spohn, G., Pacheco, A.B., Ieva, R., Alaimo, C.,Rappuoli, R., and Scarlato, V. (2002) Autoregulation ofHelicobacter pylori Fur revealed by functional analysis ofthe iron-binding site. Mol Microbiol 46: 1107–1122.
Delany, I., Spohn, G., Rappuoli, R., and Scarlato, V. (2003)An anti-repression Fur operator upstream of the promoteris required for iron-mediated transcriptional autoregulationin Helicobacter pylori. Mol Microbiol 50: 1329–1338.
Delany, I., Ieva, R., Soragni, A., Hilleringmann, M., Rappuoli,R., and Scarlato, V. (2005) In vitro analysis of protein-operator interactions of the NikR and fur metal-responsiveregulators of coregulated genes in Helicobacter pylori.J Bacteriol 187: 7703–7715.
Emsley, P., and Cowtan, K. (2004) Coot: model-building toolsfor molecular graphics. Acta Crystallogr D Biol Crystallogr60: 2126–2132. Epub 2004 Nov 2126.
Ernst, F.D., Bereswill, S., Waidner, B., Stoof, J., Mader, U.,Kusters, J.G., et al. (2005a) Transcriptional profiling ofHelicobacter pylori Fur- and iron-regulated geneexpression. Microbiology 151: 533–546.
Ernst, F.D., Homuth, G., Stoof, J., Mader, U., Waidner, B.,Kuipers, E.J., et al. (2005b) Iron-responsive regulation ofthe Helicobacter pylori iron-cofactored superoxide dismu-tase SodB is mediated by Fur. J Bacteriol 187: 3687–3692.
Escolar, L., Perez-Martin, J., and de Lorenzo, V. (1999)Opening the iron box: transcriptional metalloregulation bythe Fur protein. J Bacteriol 181: 6223–6229.
Evans, P. (2006) Scaling and assessment of data quality.Acta Crystallogr D 62: 72–82.
French, S., and Wilson, K. (1978) On the treatment of nega-tive intensity observations. Acta Crystallogr A 34: 517–525.
Gancz, H., Censini, S., and Merrell, D.S. (2006) Iron and pHhomeostasis intersect at the level of Fur regulation in thegastric pathogen Helicobacter pylori. Infect Immun 74:602–614.
Gonzalez de Peredo, A., Saint-Pierre, C., Adrait, A., Jacqua-met, L., Latour, J.M., Michaud-Soret, I., and Forest, E.(1999) Identification of the two zinc-bound cysteines in theferric uptake regulation protein from Escherichia coli:chemical modification and mass spectrometry analysis.Biochem 38: 8582–8589.
Grifantini, R., Sebastian, S., Frigimelica, E., Draghi, M.,Bartolini, E., Muzzi, A., et al. (2003) Identification ofiron-activated and -repressed Fur-dependent genes by
Crystal structure of Fur from Helicobacter pylori 1273
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

transcriptome analysis of Neisseria meningitidis group B.Proc Natl Acad Sci USA 100: 9542–9547.
Hantke, K. (2001) Iron and metal regulation in bacteria. CurrOpin Microbiol 4: 172–177.
Jacquamet, L., Aberdam, D., Adrait, A., Hazemann, J.L.,Latour, J.M., and Michaud-Soret, I. (1998) X-ray absorptionspectroscopy of a new zinc site in the fur protein fromEscherichia coli. Biochem 37: 2564–2571.
Jacquamet, L., Traore, D.A., Ferrer, J.L., Proux, O., Teste-male, D., Hazemann, J.L., et al. (2009) Structural charac-terization of the active form of PerR: insights into the metal-induced activation of PerR and Fur proteins for DNAbinding. Mol Microbiol 73: 20–31.
Kabsch, W. (1993) Automatic processing of rotation diffrac-tion data from crystals of initially unknown symmetry andcell constants. J Appl Crystallogr 26: 795–800.
de La Fortelle, E., and Bricogne, G. (1997) Maximum-likelihood heavy-atom parameter refinement in the MIRand MAD methods.
Leduc, D., Gallaud, J., Stingl, K., and de Reuse, H. (2010)Coupled amino acid deamidase-transport systems essen-tial for Helicobacter pylori colonization. Infect Immun 78:2782–2792.
Lee, J.W., and Helmann, J.D. (2007) Functional specializa-tion within the Fur family of metalloregulators. Biometals20: 485–499.
Leonard, G.A., Sole, V.A., Beteva, A., Gabadinho, J.,Guijarro, M., McCarthy, J., et al. (2009) Online collectionand analysis of X-ray fluorescence spectra on the macro-molecular crystallography beamlines of the ESRF. J ApplCrystallogr 42: 333–335.
Leslie, A.G.W. (1992) Recent changes to the MOSFLMpackage for processing film and image plate data. JointCCP4 and ESF-EACMB Newsletter on Protein Crystallog-raphy No26 Daresbury Laboratory, Warrington, UK.
de Lorenzo, V., Wee, S., Herrero, M., and Neilands, J.B.(1987) Operator sequences of the aerobactin operon ofplasmid ColV-K30 binding the ferric uptake regulation (fur)repressor. J Bacteriol 169: 2624–2630.
Lucarelli, D., Russo, S., Garman, E., Milano, A., Meyer-Klaucke, W., and Pohl, E. (2007) Crystal structure andfunction of the zinc uptake regulator FurB from Mycobac-terium tuberculosis. J Biol Chem 282: 9914–9922.
McCoy, A.J., Grosse-Kunstleve, R.W., Adams, P.D., Winn,M.D., Storoni, L.C., and Read, R.J. (2007) Phaser crystal-lographic software. J Appl Crystallogr 40: 658–674.
Masse, E., and Gottesman, S. (2002) A small RNA regulatesthe expression of genes involved in iron metabolism inEscherichia coli. Proc Natl Acad Sci USA 99: 4620–4625.
Menard, R., Sansonetti, P.J., and Parsot, C. (1993) Nonpolarmutagenesis of the ipa genes defines IpaB, IpaC, and IpaDas effectors of Shigella flexneri entry into epithelial cells.J Bacteriol 175: 5899–5906.
Miles, S., Piazuelo, M.B., Semino-Mora, C., Washington,M.K., Dubois, A., Peek, R.M., et al. (2010a) Detailed in vivoAnalysis of the Role of Helicobacter pylori Fur in Coloniza-tion and Disease. Infect Immun 48: 378–386.
Miles, S., Carpenter, B.M., Gancz, H., and Merrel, D.S.(2010b) Helicobacter pylori apo-Fur regulation appearsunconserved across species. J Microbiol 48: 378–386.
Nandal, A., Huggins, C.C., Woodhall, M.R., McHugh, J.,
Rodriguez-Quinones, F., Quail, M.A., et al. (2010) Induc-tion of the ferritin gene (ftnA) of Escherichia coli by Fe(2+)-Fur is mediated by reversal of H-NS silencing and is RyhBindependent. Mol Microbiol 75: 637–657.
Pecqueur, L., D’Autreaux, B., Dupuy, J., Nicolet, Y., Jacqua-met, L., Brutscher, B., et al. (2006) Structural changes ofEscherichia coli ferric uptake regulator during metal-dependent dimerization and activation explored by NMRand X-ray crystallography. J Biol Chem 281: 21286–21295.
Perrakis, A., Morris, R., and Lamzin, V.S. (1999) Automatedprotein model building combined with iterative structurerefinement. Nat Struct Biol 6: 458–463.
Pflock, M., Finsterer, N., Joseph, B., Mollenkopf, H., Meyer,T.F., and Beier, D. (2006) Characterization of the ArsRSregulon of Helicobacter pylori, involved in acid adaptation.J Bacteriol 188: 3449–3462.
Pohl, E., Haller, J.C., Mijovilovich, A., Meyer-Klaucke, W.,Garman, E., and Vasil, M.L. (2003) Architecture of a proteincentral to iron homeostasis: crystal structure and spectro-scopic analysis of the ferric uptake regulator. Mol Microbiol47: 903–915.
Reddi, A.R., Guzman, T.R., Breece, R.M., Tierney, D.L., andGibney, B.R. (2007) Deducing the energetic cost ofprotein folding in zinc finger proteins using designedmetallopeptides. J Am Chem Soc 129: 12815–12827.
Schauer, K., Gouget, B., Carriere, M., Labigne, A., and deReuse, H. (2007) Novel nickel transport mechanism acrossthe bacterial outer membrane energized by the TonB/ExbB/ExbD machinery. Mol Microbiol 63: 1054–1068.
Schneider, T.R., and Sheldrick, G.M. (2002) Substructuresolution with SHELXD. Acta Crystallogr D Biol Crystallogr58: 1772–1779.
Sheikh, M.A., and Taylor, G.L. (2009) Crystal structure of theVibrio cholerae ferric uptake regulator (Fur) revealsinsights into metal coordination. Mol Microbiol 72: 1208–1220.
Skouloubris, S., Labigne, A., and De Reuse, H. (1997) Iden-tification and characterization of an aliphatic amidase inHelicobacter pylori. Mol Microbiol 25: 989–998.
Stingl, K., Schauer, K., Ecobichon, C., Labigne, A., Lenor-mand, P., Rousselle, J.C., et al. (2008) In vivo interactomeof Helicobacter pylori urease revealed by tandem affinitypurification. Mol Cell Proteomics 7: 2429–2441.
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994)CLUSTAL W: improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting,position-specific gap penalties and weight matrix choice.Nucleic Acids Res 22: 4673–4680.
Tomb, J.F., White, O., Kerlavage, A.R., Clayton, R.A., Sutton,G.G., Fleischmann, R.D., et al. (1997) The completegenome sequence of the gastric pathogen Helicobacterpylori. Nature 388: 539–547.
Traore, D.A., El Ghazouani, A., Ilango, S., Dupuy, J., Jac-quamet, L., Ferrer, J.L., et al. (2006) Crystal structure ofthe apo-PerR-Zn protein from Bacillus subtilis. Mol Micro-biol 61: 1211–1219.
Tsugawa, H., Suzuki, H., Satoh, K., Hirata, K., Matsuzaki, J.,Saito, Y., et al. (2010) Two amino acids mutation of Ferricuptake regulator (Fur) determines Helicobacter pylori resis-tance to metronidazole. Antioxid Redox Signal 14: 15–23.
1274 C. Dian et al. �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275

Vitale, S., Fauquant, C., Lascoux, D., Schauer, K., Saint-Pierre, C., and Michaud-Soret, I. (2009) A ZnS(4) structuralzinc site in the Helicobacter pylori ferric uptake regulator.Biochem 48: 5582–5591.
van Vliet, A.H., Stoof, J., Poppelaars, S.W., Bereswill, S.,Homuth, G., Kist, M., et al. (2003) Differential regulation ofamidase- and formamidase-mediated ammonia productionby the Helicobacter pylori fur repressor. J Biol Chem 278:9052–9057.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
Crystal structure of Fur from Helicobacter pylori 1275
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 79, 1260–1275
Top Related
Copyright © 2022 FDOKUMEN

