







Bahasa
Halaman
Hukum


Aus der Klinik für Innere Medizin A
(Direktor Univ. Prof. Dr. Markus M. Lerch)
der Universitätsmedizin der Ernst-Moritz-Arndt-Universität Greifswald
The role of keratin 15 in small intestinal
homeostasis
Inaugural - Dissertation zur Erlangung
des akademischen Grades eines Doktors der Medizin
der Universitätsmedizin der Ernst-Moritz-Arndt-Universität Greifswald
2018
Vorgelegt von: Julien Orlando Stephan
Geboren am 01.06.1991 in Bethesda (USA)

Dekan: Prof. Dr. med. Karlhans Endlich
1. Gutachter: Prof. Dr. Barthlen
2. Gutachter: Prof. Dr. Lerch
3: Gutachter: Prof. Dr. Singer
Ort, Raum: Greifswald, Seminarraum Innere Medizin A
Tag der Disputation: 12.12.2019

Diese Arbeit widme ich meinen Eltern, für deren unermüdliche Unterstützung und Liebe.

ZUSAMMENFASSUNG
4
Aus der Klinik und Poliklinik für Innere Medizin A
- Direktor: Univ.Prof. Dr. M. M. Lerch -
der Medizinischen Fakultät der Universität Greifswald
ZUSAMMENFASSUNG
The role of keratin 15 in small intestinal homeostasis - Julien Orlando Stephan-
Die vorliegende Arbeit untersucht die Relevanz des Intermediärfilamentes Zytokeratin 15 (K15)
als potentiellen epithelialen Stammzellmarker im Darm und den Einfluss K15 positiver Zellreihen
auf die Krypta Homöostase.
Zwei Hauptstammzellpools regulieren den schnellen Zellumsatz im Darmepithel. Dies sind
einerseits schnell proliferierende Lgr5 positive Stammzellen, welche zwischen den Paneth-Zellen
an der Krypta-Basis vorgefunden werden. Anderseits gibt es vermutlich langsamer wachsende
Bmi1 positive Zellen, welche sich an der +4-Position oberhalb der Krypta-Basis befinden. Im
Haarfollikel und im Ösophagusepithel stellt das Intermediärfilament K15 einen Marker für
Stammzellen dar, die zur Gewebereparatur beitragen. In dieser Arbeit haben wir gezeigt, dass K15
im Darm langlebige Kryptazellen mit Multipotenz- und Selbsterneuerungspotenzial markiert. K15
positive Krypta-Zellen sind resistent gegen hochdosierte ionisierende Strahlung und tragen zur
Kryptaexpansion bei. Die hier vorgestellten Ergebnisse zeigen nun erstmals im Darm eine
langlebige, multipotente K15 exprimierende Kryptazellpopulation, die eine
Selbsterneuerungskapazität besitzt. Insbesondere führt der Verlust des Tumorsuppressor Gens Apc
in K15 positiven Zellen zur Adenombildung, die potentiell zum Adenokarzinomen fortschreiten
können. Wir erörtern die Hypothese, dass K15 eine Gruppe langlebiger, strahlenresistenter
Stammzellen markiert, die die Homöostase der Krypta und die Regenerationsfähigkeit maßgeblich
beeinflussen.

TABLE OF CONTENTS
5
TABLE OF CONTENTS
ZUSAMMENFASSUNG ............................................................................................................................. 4
TABLE OF CONTENTS ............................................................................................................................ 5
LIST OF FIGURES ..................................................................................................................................... 8
LIST OF ABBREVIATIONS ................................................................................................................... 10
1. ABSTRACT ............................................................................................................................................ 12
2. REVIEW OF THE LITERATURE ..................................................................................................... 13 2.1. OVERVIEW OF INTERMEDIATE FILAMENTS ....................................................................................... 13
2.1.1. Main intermediate filament functions ....................................................................................... 13 2.2. KERATINS ......................................................................................................................................... 14
2.2.1. The role of SEKs in the small intestine ..................................................................................... 14 2.3. OVERVIEW OF THE SMALL INTESTINAL EPITHELIUM ....................................................................... 15
2.3.1. The stem cell niche and their components ................................................................................ 16 2.3.2. The role of stem cells in cancer initiation ................................................................................. 18
3. AIMS OF THE STUDY ........................................................................................................................ 20
4. MATERIAL AND METHODS ............................................................................................................ 21 4.1. CHEMICALS AND REAGENTS ............................................................................................................ 21
4.1.1. Immunohistochemistry and Immunofluorescence ..................................................................... 21 4.1.2. Primary Antibodies ................................................................................................................... 22
4.1.3. Secondary Antibodies ................................................................................................................ 22 4.1.4. PCR primers .............................................................................................................................. 22
4.1.5. Materials used in cell culture .................................................................................................... 23
4.1.6. Instruments and software .......................................................................................................... 23 4.1.7. Consumable supplies ................................................................................................................. 25
4.1.8. Animal care ............................................................................................................................... 25
4.2. ANIMAL EXPERIMENTAL METHODOLOGY ........................................................................................ 26 4.2.1. Origin of transgenic mice used in the experiments ................................................................... 26
4.2.2. Animal care ............................................................................................................................... 26
4.2.3. Genotyping procedure ............................................................................................................... 27 4.2.4. Euthanasia and organ harvest .................................................................................................. 27
4.2.5. Cre / LoxP recombination ......................................................................................................... 27

TABLE OF CONTENTS
6
4.2.6. Experimental strategy for the Krt15-crePR1; R26mT/mG mouse. ................................................ 27 4.2.8. Experimental strategy for Krt15-crePR1; R26Tom mice. ........................................................... 28
4.2.9. Experimental strategy for irradiation of Krt15-crePR1; R26mT/mG and Krt15-/- mice. .............. 29 4.2.10. Experimental strategy for Krt15-CrePR1;Apcfl/fl;R26mT/mG mice ............................................ 29
4.3. TISSUE FIXATION .............................................................................................................................. 30
4.4. IMMUNOHISTOCHEMISTRY (IHC) ..................................................................................................... 30 4.4.1. IHC in combination with the M.O.M.-kit .................................................................................. 31
4.5. IMMUNOFLUORESCENCE (IF) ............................................................................................................ 31
4.5.1. Immunofluorescence using the TSA system .............................................................................. 31
4.6. ALCIAN BLUE STAINING (AB) .......................................................................................................... 32
4.7. SINGLE CELL ISOLATION ................................................................................................................... 32 4.8. CRYPT ISOLATION AND 3D ORGANOID CULTURE ............................................................................. 32
4.9. IRRADIATION ..................................................................................................................................... 33 4.10. HISTOMETRIC ANALYSIS ................................................................................................................. 33
4.10.1. Crypt length analysis .............................................................................................................. 33
4.10.2 Crypt microcolony assay .......................................................................................................... 34 4.10.3. Software and statistical analysis ............................................................................................. 34
5. RESULTS ............................................................................................................................................... 35 5.1. ENDOGENOUS EXPRESSION OF KRT15 IN THE SMALL INTESTINE. ..................................................... 35
5.1.1. Endogenous expression of Krt15 in WT mice ........................................................................... 35 5.1.2. Localization of Krt15+ cells in Krt15-crePR1; R26mT/mG mice. ................................................. 37
5.2. KRT15 EXPRESSION IN VARIOUS EPITHELIAL CELL TYPES PRESENT IN THE INTESTINE. ................... 40
5.2.1. Krt15 expression in enteroendocrine cells. ............................................................................... 41 5.2.2. Krt15 expression in the goblet cell population. ........................................................................ 42
5.2.3. Krt15 expression in the Paneth cell population. ....................................................................... 42
5.2.4. Cell-cycling activity among Krt15+ cells. ................................................................................. 44 5.3. EVALUATION OF PROLIFERATIVE POTENTIAL WITHIN THE KRT15+ CELL POPULATION. .................. 45
5.3.1. Lineage tracing experiment on Krt15-crePR1; R26mT/mG mice. ................................................ 45
5.3.2. Colocalization of Krt15+ cells with proliferating cells in a lineage tracing setting. ................ 49 5.3.3. Colocalization of Krt15+ cells with enteroendocrine cells in a lineage tracing setting. .......... 50
5.3.4. Colocalization of Krt15+ cells with goblet cells in a lineage tracing setting. .......................... 51
5.3.5. Colocalization of Krt15+ cells with Paneth cells in a lineage tracing setting. ......................... 52 5.3.6. Krt15+ cells depict multipotency in 3D organoids .................................................................... 52

TABLE OF CONTENTS
7
5.4. THE ROLE OF KRT15+ CELLS IN RESPONSE TO RADIATION INJURY AND THEIR REGENERATION
CAPABILITIES. .......................................................................................................................................... 56
5.4.1. The role of Krt15+ cells in tissue recovery. ............................................................................... 56 5.4.2. The importance of Krt15 expressing cells in irradiation models compared to Krt15-/- ............ 60
5.4.3. Microcolony assay on Krt15 expressing cells. .......................................................................... 64
5.5. ROLE OF KRT15 IN CANCER INITIATION. ........................................................................................... 66
6. DISCUSSION ......................................................................................................................................... 69
7. CONCLUSION ...................................................................................................................................... 75
8. ACKNOWLEDGEMENT .................................................................................................................... 76
8.1. AUTHOR CONTRIBUTION ................................................................................................................... 76
9. BIBLIOGRAPHY .................................................................................................................................. 77
10. ORIGINAL PAPER ............................................................................................................................ 85
EIDESSTATTLICHE ERKLÄRUNG .................................................................................................... 98
CURRICULUM VITAE ........................................................................................................................... 99
DANKSAGUNG ...................................................................................................................................... 101

LIST OF FIGURES
8
LIST OF FIGURES Figure 1: The regulatory niche of intestinal stem cells ............................................................ 17
Figure 2: Breeding strategy for the Krt15-crePR1; R26mGFP mouse. ..................................... 28
Figure 3: Experimental strategy for Krt15-crePR1; R26mT/mG mouse irradiation. ............... 29
Figure 4:Experimental strategy for 3D organoid structures ................................................... 33
Figure 5: Control of Krt15 knockout in Krt15-/- mice versus WT mice. ................................ 35
Figure 6: Krt15 is expressed in intestinal epithelial cells. ........................................................ 36
Figure 7: Krt15+ cells can consistently be identified in the intestinal epithelium. ................. 37
Figure 8: Krt15-crePR1; R26mT/mG mice reliably label Krt15+ cell in the small intestine. ... 38
Figure 9: Krt15+ cells are predominantly located in the stem cell compartment. ................. 39
Figure 10: Krt15+ cells are predominantly located in the stem cell compartment. ............... 40
Figure 11: Colocalization of Krt15+ cells with enteroendocrine cells. .................................... 41
Figure 12: Krt15+ cells colocalize with goblet cells in isolated events. .................................... 42
Figure 13: Krt15+ cells are located above crypt base, showing no colocalization with secretory
paneth cells. .......................................................................................................................... 43
Figure 14: Krt15+ cells are proliferating cells located at a +4 position. .................................. 44
Figure 15: Timetable depicting experimental setup of the lineage tracing experiment over
150 days. ............................................................................................................................... 45
Figure 16: Evolution of Krt15+ progeny cells within the crypt. .............................................. 46
Figure 17: Lineage tracing over 150 days depicts Krt15+ as proliferative, long-lived epithelial
cells. ....................................................................................................................................... 47
Figure 18: Krt15+ progeny cells colonize the entire crypt as well as adjacent crypt and villi
after 150 days. ...................................................................................................................... 48
Figure 19: Krt15 marks a long-lived consistently expanding crypt population. ................... 49
Figure 20: Krt15+ progeny cells maintain the ability to differentiate in to enteroendocrine
cells. ....................................................................................................................................... 50
Figure 21: Krt15+ progeny cells reliably differentiate into goblet cells over time. ................ 51
Figure 22: Krt15+ cells can differentiate into paneth cells over time. ..................................... 52
Figure 23: live 3D organoids formed by Tomato+(Krt15-derived) ......................................... 53
Figure 24: Fixated 3D organoids formed by Tomato+(Krt15-derived) sf .............................. 53

LIST OF FIGURES
9
Figure 25: Co-staining of Tomato with Ki-67, Mucin 2, Chromogranin A and Lysozyme in
fixated Organoids ................................................................................................................ 54
Figure 26: live 3D organoids formed by isolated single Tomato+(Krt15-derived) cell. ........ 55
Figure 27: GFP marks radioresistant Krt15 positive cells capable of repopulating intestinal
crypts after radiation injury ............................................................................................... 58
Figure 28: Krt15+ cells vitally expand post radiation injury, labeling the majority of the crypt
population. ............................................................................................................................ 59
Figure 29: Proliferative capacity of Krt15+ progeny cells within labeled crypts, post radiation
injury. ................................................................................................................................... 59
Figure 30: Depressed ileal crypt rehabilitation in Krt15-/- mice, post irradiation injury. .... 61
Figure 31: Discrepancies in crypt length in the ileum of WT mice and Krt15-/- mice 5 days
post radiation injury. ........................................................................................................... 61
Figure 32: Jejunal crypts unchallenged post irradiation in Krt15-/- mice. ............................. 62
Figure 33: Discrepancies in crypt length between WT mice and Krt15-/- mice 5 days post
radiation injury. ................................................................................................................... 63
Figure 34: Krt15-/- mice, portrayed a significant reduction of proliferating ileal crypt cells.
............................................................................................................................................... 64
Figure 35: Significant reduction in absolute number of live cells per crypt in wildtype and
Krt15-/- mice. ......................................................................................................................... 65
Figure 36: Number of microcolonies per 50 crypts, in wildtype vs. Krt15-/- mice post
irradiation. ........................................................................................................................... 65
Figure 37: Timetable depicting experimental setup for the cancer initiation model ............ 66
Figure 38: Representative image of intestinal tumors in Krt15-CrePR1;Apcfl/fl;R26mT/mG
mice ....................................................................................................................................... 67
Figure 39: Representative histology of flat adenoma and invasive adeno carcinoma in Krt15-
CrePR1;Apcfl/fl;R26mT/mG mice. .......................................................................................... 67

LIST OF ABBREVIATIONS
10
LIST OF ABBREVIATIONS -/- Knock-out
+/- Heterozygote
+/+ Wild type
AAALCA Association for Assessment and Accreditation of Laboratory Animal Care
AB Alcian Blue
ABSL Animal Biosafety Level
BMP bone morphogenetic protein
bp Base pair
BrdU Bromodeoxyuridine
BSA Bovine serum albumin
C Colon
CBC Crypt based columnar
cDNA Complementary deoxyribonucleic acid
Chr A Chromogranin A
CO2 Carbon dioxide
CTL Control
CY Cyanine dye
DAB Diaminobenzene
DAPI Diamidino-2-Phenylindole
DMEM Dulbecco's Modification of Eagle's Medium
DNA Deoxyribonucleic acid
DPBS Dulbecco's Phoasphate buffered Saline
DT Diphtheria toxin
DTR Diphtheria toxin receptor
EGF epidermal growth factor
EtOH Ethanol
GFP Green fluorescent protein
H2O Water
HRP Horseradish peroxidase
iDTR Inducible diphtheria toxin receptor

LIST OF ABBREVIATIONS
11
IF Immunofluorescence
IFL Intermediate filament
IgG Immunoglobulin G
IHC Immunohistochemistry
IP Intra peritoneal
ISC Intestinal stem cell
JIR Jackson Immune Research
K15 Cytokeratin / Keratin 15 (protein)
K19 Cytokeratin / Keratin 19
KO Knock out mice
Krt 15 Keratin 15 promoter (DNA)
Lgr5 Leucine-rich repeat-containing G-protein coupled receptor 5
LYZ Lysozyme
milliQ H2O Purified water
Muc 2 Mucin 2
PBS Phosphate buffered saline
PBT Phosphate buffered saline with 1% Tween 20
PCR Polymerase chain reaction
PFA Paraformaldehyde
qPCR Quantitative polymerase chain reaction
R26 ROSA 26 locus
Rb Rabbit
RT Room temperature
SEK Simple epithelial Keratins
SI Small intestine
SOX9 Sex-determining region y-box 9
TSA Tyramide signal amplification
ULAR University Laboratory Animal Resource
WT Wild type mice

1. ABSTRACT
12
1. ABSTRACT Two principal stem cell pools orchestrate the fast cell turnover in the intestinal epithelium. Rapidly
cycling Lgr5+ stem cells are intercalated between the Paneth cells at the crypt base (CBCs) and a
putative slower cycling Bmi1+ cells are located at the +4 position above the crypt base. In the hair
follicle and the esophageal epithelium, the intermediate filament Keratin 15 (Krt15) marks stem
cells contributing to tissue repair. Herein, we demonstrated that Krt15 labels long-lived crypt cells
harboring multipotency and self-renewing potential. Krt15+ crypt cells are resistant to high-dose
radiation and contribute to crypt expansion following injury. These results suggest that Krt15
annotate a long-lived, multipotent crypt cell population harboring self-renewal capacity. Notably,
Apc loss in Krt15+ cells lead to adenoma formation that can occasionally progress to
adenocarcinoma.

2. REVIEW OF THE LITERATURE
13
2. REVIEW OF THE LITERATURE
2.1. Overview of intermediate filaments The eukaryotic cytoskeleton is composed of three major protein families, i.e. microfilaments,
microtubules and intermediate filaments (IFL) which can be differentiated from one another by
size and function. IFL have an average size of 10 nm and are thereby ranged in size between
microfilaments (7nm) and microtubule (25nm). In contrast to microfilaments and microtubules,
IFL have a widespread expression profile formed by the transcription of 70 functional genes with
alternative splicing and post-transcriptional modifications (Szeverenyi et al., 2008). These
modifications bear extremely versatile protein structure with individualized properties. Whereas
microfilaments and microtubule are polymers of a single type of protein, IFL are heteropolymers
composed of a variety of proteins. More than 50 different proteins have been identified and
classified into 6 types of IFL, based on similarities in their primary protein structure (Cooper,
2000). Type I and II are keratins, expressed in epithelial cells and divided into hard epithelial
keratins and soft epithelial keratins (SEK). Hard keratins are present in structures such as hairs,
nails and horns, whereas SEK are contained in the cytoplasm of epithelial cells. Type III IFLs
include vimentin and desmin and are especially expressed in fibroblast und muscle tissue. Type
IV IFLs consist mainly of neurofilaments expressed in neural cells. Type V IFLs are nuclear
laminins, forming an orthogonal meshwork that is crucial for nuclear stability. Type VI IFLs
mainly consist of nestine proteins expressed in neural stem cells.
2.1.1. Main intermediate filament functions
During the 1990s, the main role of IFL was believed to be maintaining the structural and
mechanical integrity of cells. Only recent studies unraveled IFL as a highly dynamic structure
involved in organizing various cellular processes (Eriksson et al., 2009). Diseases associated with
isolated IFL mutations are linked to a broad range of symptoms, unraveling the vast impact of
these underestimated proteins. IFLs can act as functional determinants signaling pathways and
posttranslational protein targeting. The cytoprotective effects of IFLs are related to their
mechanical properties facilitating cells to cope with mechanical and non-mechanical stresses.

2. REVIEW OF THE LITERATURE
14
However, cytoprotection may also arise from the capacity of IFs to interact with signaling
pathways involved in determining cell survival and cell fate (Sahlgren et al., 2003).
2.2. Keratins Keratins are type I and II IFLs expressed in epithelial cells of all vertebrates. Like all types of IFL,
they play a crucial role in cellular integrity, cell signaling, protein targeting, apoptosis and
protection against stresses. Keratins form obligate heteropolymers (one type I and one type II) and
share a common structure that consists of a central coiled-coil α-helical rod domain that is flanked
by non-α-helical head and tail domains. Different types of keratins can be distinguished, regarding
their biochemical constitution or according to keratin-producing cells and tissues. Epithelial cells
of in simple epithelia regularly synthesize K5 or K14, whereas K8 and K18 predominate in
stratified epithelia. These epithelial cells can also produce other keratins in addition to or instead
of the primary keratins and these keratins are referred to as secondary keratins, such as K7/K19 in
simple epithelia or K15 and K6/K16 in stratified epithelia (Bragulla and Homberger, 2009). For
example, Krt15 is expressed in the basal layers of the stratified epithelium as well as in
keratinoblasts of the hair follicle. The distinction between keratin types can help distinguish
between various levels of differentiation of epithelial cells. Recent findings link Krt15 expression
patterns to undifferentiated stem cells in the bulge of the hair follicle as well as in epidermal cell
layers (Bose et al., 2013; Liu et al., 2003). Krt15+ cells in the bulge of the hair follicle are long-
lived and possess a greater colony-forming ability, establishing them as putative follicular stem
cells (Inoue et al., 2009). Krt15+ cells have also been shown to contribute in epidermal renewing
and repair after wounding (Ito et al., 2005; Li et al., 2013). Furthermore, our team identified Krt15
expressing cells to mark a long-lived subpopulation of basal cells in the mouse esophagus capable
of generating all states of squamous lineages (Giroux et al., 2017).
2.2.1. The role of SEKs in the small intestine
One of the challenges in modern biomedicine is to explore the functional significance of keratins
in the intestine as various keratin types appear limited to the type of tissue and cell differentiation.
Specific keratins seem to predominate in undifferentiated epithelial cells, presumably providing a
selection advantage. A collaborating group investigated the role of Krt19 as potential stem cell
marker in the colon epithelium. Their findings indicated that Krt19 marks a line of radiation-

2. REVIEW OF THE LITERATURE
15
resistant stem cell located in the intestinal crypt (Asfaha et al., 2015a). Similar to K15, K19 was
reported as biochemical marker for cutaneous stem cells and is believed to be distinctly expressed
in undifferentiated progenitor cells (Driskell et al., 2015; Kloepper et al., 2008; Michel et al.,
1996). Furthermore, stem cells located in the bulge of the hair follicle were shown to express Krt19
especially in the wounded epidermis. Based on these observations, both K15 and K19 could be
used as markers of stem cells to segregate them from the differentiated cell types. Nonetheless, the
role of keratins in the intestinal homeostasis remains mainly unchartered.
2.3. Overview of the Small intestinal epithelium The gastrointestinal tract is the largest organ in the human organism and covers more than 32
square meters of epithelial surface (Helander and Fändriks, 2014). Specialized cells form a highly
complex and adaptable barrier between the exterior world and the mammalian host. The small
intestinal epithelium is formed by a single layer of columnar epithelial cells organized in villi and
crypts. These folded structures as well as the microvilli present at the surface of the enterocytes
generate a large contact surface enabling the organism to absorb a maximum amount of nutrients
and water. The gastrointestinal tract is colonized by commensal bacteria that aid in digestion and
influence the development and function of the mucosal immune system. However, microbial
colonization carries the risk of infection and inflammation as well as any potential damage by
toxins. Epithelial homeostasis is maintained by an interaction of the mucosal immune system and
the intestinal epithelial cells. Epithelial cells can be categorized into absorptive (enterocytes) and
secretory lineages (goblet cells, enteroendocrine cells and Paneth cells). Most epithelial cells
consist of absorptive enterocytes essential for nutrient and water uptake. Mucin-producing goblet
cells are mostly located in the villi and are responsible for covering the epithelium with a protective
layer of mucus. Enteroendocrine cells compose only a small proportion of the differentiated cells,
but play a crucial role by producing active hormones and cytokines acting locally and systemically.
At the bottom of each small intestinal crypt can be found Paneth cells. These cells are
extraordinarily versatile exocrine cells responsible for producing antimicrobial factors, such as
alpha-defensine and locally active mediators. These cells are believed to be essential to uphold the
microenvironment necessary for the crypt-based stem cell population. Residing at the base of the
crypt are the intestinal stem cells (ISCs) responsible for continuous turnover of epithelial cells
every 4 to 7 days. ISCs give rise to transient amplifying cells which will then proliferate and

2. REVIEW OF THE LITERATURE
16
terminally differentiate while undergoing an upward migration. Differentiated cells migrate along
the crypt/villi axis to the top of the villi where they undergo anoikis. This rapidly cycling system
makes the small intestine the ideal location to study stem cell activity.
2.3.1. The stem cell niche and their components
The term stem cell niche exists since 1978 (Schofield, 1978) and describes the unique
microenvironment crucial for stem cell survival. This stem cell niche is a complex multicellular
construct essential for stem cell maintenance, differentiation and proliferation. The ISC niche
consists of different cellular components, namely endothelial cells, Paneth cells, myofibroblasts,
neural cells, lymphocytes and adjacent smooth muscle tissue. Each component contributes by
producing unique cell-cell ligands, local growth factors or cytokines (Barker, 2014; Sailaja et al.,
2016). Wnt-signaling, bone morphogenetic protein (BMP), Notch and epidermal growth factor
(EGF) signaling pathways have been reported to regulate stem cell activity and are gradually
expressed along the length of the villi and crypts (Carulli et al., 2015; Kuhnert et al., 2004;
Pellegrinet et al., 2011; Vries et al., 2010). At this point, two main stem cell states have been
categorized. The rapidly proliferating population of crypt base columnar (CBC) cells are
intercalated between the Paneth cells. CBC cells express Lgr5, are highly proliferative pluripotent
radio-sensitive stem cells (Barker et al., 2007). CBC give rise to transient amplifying cells, which
terminally differentiate into secretory and absorptive lineages. Alongside Lgr5 a variety of
different CBC cell markers were found consisting of: Ascl2, Olfm4, Sox9-EGFP(lo), Rnf43, Znrf3,
Smoc2, Troy, Prom1, and Msi1 (Fafilek et al., 2013; Hao et al., 2012; Muñoz et al., 2012; Roche
et al., 2015; Schuijers et al., 2014, 2015; Snippert et al., 2009). Yet Lgr5 has been predominantly
studied, due to the robust expression in CBC cells and the availability of reliable tracing systems.
The second stem cell state, the reserve stem cells are located at the +4 position, meaning 4 cell
positions above the crypt base. A variety of different stem cell markers for the +4 position were
found, comprising: p-PTEN, Bmi1, mTert, Hopx and Lrig1 (He et al., 2007; Montgomery et al.,
2011; Powell et al., 2012; Sangiorgi and Capecchi, 2008a; Wong et al., 2012). The first functional
validation of +4 stem cells was achieved via lineage tracing, using an in vivo mouse model.
Contrary to Lgr5+ CBC cells, Bmi1 labeled cells are slow cycling and radioresistant cells. Bmi1+
stem cells seem to be mainly insensitive to Wnt-signaling variations proliferating only weekly
under homeostatic conditions (Yan et al., 2012). However, ablation of CBC following radiation

2. REVIEW OF THE LITERATURE
17
activates quiescent +4 stem cells causing them to rapidly proliferate and repopulate the entire crypt
population. Experimental studies of Lgr5+ cells ablation were designed using Lgr5-DTR mice,
thus specifically deleting Lgr5 expressing cells (Yan et al., 2012). Lgr5-DTR mice express the
diphtheria toxin receptor under the control of the Lgr5 promoter, allowing targeted deletion of
Lgr5+ cells. After ablation of CBC cells, +4 stem cells rapidly proliferated, replacing vacant crypt
spots with Bmi1+ progeny cells. Remarkably, Bmi1+ progeny cells replaced ablated CBC cells and
began to express Lgr5, confirming the potential of Bmi1+ stem cells to differentiate into all
intestinal cell lineages.
Figure 1: The regulatory niche of intestinal stem cells
Picture derived from an illustration taken out of The Journal of Physiology, Volume 594, Issue 17, pages 4827-4836, 28 JUL 2016
DOI: 10.1113/JP271931.
The stem cells, consisting of CBC and +4 stem cells, reside at the bottom of the crypt. Rapidly dividing transit-amplifying cells
arise from these stem cells and differentiate into absorptive lineages (enterocytes) or secretory lineages (enteroendocrine cells,
goblet cells and Paneth cells). The niche consists of multiple components and cell types, including extracellular matrix, fibroblasts,

2. REVIEW OF THE LITERATURE
18
myofibroblasts, smooth muscle cells, neural cells, endothelial cells, lymphocytes and macrophages along with various secreted
factors. Wnt, BMP, Notch, Hedgehog, and EGF signaling pathways are regulating clonogenic activity (Sailaja et al., 2016).
A collaborating group identified Keratin 19 as a marker of a new type of stem cell located from
the +4 position and reaching up to the crypt isthmus. This novel cell type is Lgr5-, radio resistant
and gives rise to all epithelial cell lineages, including Lgr5+ CBC cells (Asfaha et al., 2015a). It
could also be associated with cancer initiation of radioresistant neoplastic tumor cells. These
findings were particularly interesting for our purpose, as K19 marks the first intestinal stem cell
marker labeled by intermediate filaments. K19 and K15 are putative biomarkers in the epidermal
tissue of the hair follicle, however their involvement in the intestinal epithelium has yet to be
sufficiently identified (Inoue et al., 2009; Ma et al., 2004).
2.3.2. The role of stem cells in cancer initiation
Cancer of the gastrointestinal tract, such as the colorectal cancer, is one of the most common cancer
worldwide and carries the second highest mortality rate in developed countries (Jemal et al., 2011).
The traditional view of cancer initiation argues that all mutated cells have the equal potential to
proliferate and drive tumor growth, based on the principal of stochastic probability. However, the
traditional cancer model has recently been challenged by the cancer stem cell model. The cancer
stem cell model postulates that tumors are organized hierarchically, with cancer stem cells as a
proliferative driving force (Dick, 2009). Adult stem cells possess essential characteristics such as
self-renewal capacity and long-term replication ability, bearing similitudes to cancer cells. During
normal homeostasis, these capacities are tightly regulated by diverse growth factors and signaling
pathways such as Wnt signaling, mutated stem cells are far more likely to transform to cancer stem
cells, than any other cell (Espersen et al., 2015). These theories fueled recent studies to identify
putative intestinal stem cell markers and their potential role in cancer initiation. Some intestinal
stem cell markers are now believed to be linked with colorectal cancer initiation including SOX9,
Bmi1 and Lgr5 (Espersen et al., 2015; Seshagiri et al., 2012; Takahashi et al., 2011). In addition,
the first proven stem cell marker Lgr5 has been extensively studied and increased expression in
adenomas as well as invasive colorectal cancer was confirmed in several studies (Fan et al., 2010,
2010; Takeda et al., 2011). High expression of Lgr5 is usually linked with Wnt signaling. R-
spondin, a Wnt-signaling agonist was linked to increased Lgr5 expression in colorectal cancer via
positive feedback loops (de Lau et al., 2011; Seshagiri et al., 2012). Next to Lgr5+ CBC cells,

2. REVIEW OF THE LITERATURE
19
Bmi1+ stem cells are located at the +4 position and are functionally distinct by their radioresistant
and quiescent role in the intestinal crypt. The exact implication of Bmi1 in colon cancer initiation
remains unclear, yet studies report some overexpression of Bmi1 in human colon cancer (Li et al.,
2010; Reinisch et al., 2006; Tateishi et al., 2006). Induction of b-catenin in Bmi1+ ISC has shown
to generate rapidly growing adenomas (Sangiorgi and Capecchi, 2008a). Nonetheless, the exact
implication of stem cells on homeostasis and potential impact on cancer initiation has yet to be
clarified.

3. AIMS OF THE STUDY
20
3. AIMS OF THE STUDY The main objective of this study is to unravel the roles of keratin 15 in intestinal homeostasis.
Specific aims are as follow:
A. Determine the endogenous expression of K15 in the mouse small intestinal epithelium
B. Identify which small intestinal cell types express K15
C. Evaluate the proliferative potential of the K15 positive cell population
D. Investigate the role of K15 expressing cells in an injury model and the regeneration
capability of these cells
E. Determine whether K15 cells are crucial for small intestinal homeostasis
F. Determine the implication of K15 in cancer initiation

4. MATERIAL AND METHODS
21
4. MATERIAL AND METHODS 4.1. Chemicals and Reagents 4.1.1. Immunohistochemistry and Immunofluorescence
Zinc Formalin Fixative
Polyscience Inc. 21516-3.75
Paraffin: TissuePrep embedding compound Fisher Scientific T565
Ethanol 200 Proof 100%
Decon Laboratories #2701
Ethanol 150 Proof 95%
Decon Laboratories #2801
Xylene
VWR 89370-088
10X PBS
Growcells MRGF-6235
Hydrogen peroxide solution 30% in H2O Sigma-Aldrich 216763
Citric Acid Monohydrate
Fisher Scientific A104-500
Starting Block T20
Fisher Scientific PI-37539
Diamidino-2-Phenylindole (DAPI) Fisher Scientific D1306
Grill III Hematoxylin
Leica Surgipath 3801540
M.O.M. Kit
Vector BMK-2202
Triton X-100
Fisher Scientific BP151-500
Tween 20 - was part of the washing buffer Amresco M147-1L
Mounting media for IF
KPL 71-00-16
Cytoseal 60
Richard-Allan Corp 8310-4
3 % Acetic Acid
Self-made -
1% Alcian Blue in 3% Acetic acid
R&D Corp S111A-8OZ
Nuclear fast RED Kernechtrot 0.1% R&D Corp S248-8OZ
Normal Donkey Serum
JIR 017-000-121
BSA - Bovine Serum Albumin Fract V Fisher Biotech CAS 9048-46-8
Biotin
Sigma-Aldrich B4501
Avidin from egg
Sigma-Aldrich A9275
Reagent A (Avidin DH)
Vectastain PK-6100
Reagent B (Biotinylated HRP)
Vectastain PK-6100
TSA
Perkin Elmer NEL700A001K
Biotinyl tyramide reagent
Perkin Elmer FP1019

4. MATERIAL AND METHODS
22
Streptavidin-HRP
Perkin Elmer FP1047
4.1.2. Primary Antibodies
Lysozyme Rabbit Diagnostic Biosystems RP-028
Mucin 2 Glycoprotein Mouse Vector Labs VP-M656
Mucin 2 Glycoprotein Rabbit Abcam ab133555
SP-1 Chromogranin A Rabbit ImmunoStar 20085
Ki67 Rabbit Abcam ab16667
BrdU (Bu 20a) mouse IgG1 cell signaling 5292S
GFP Goat pAb Abcam ab6673
K15 Mouse Vector Labs VP-C411
K15 Rabbit Abcam ab52816
4.1.3. Secondary Antibodies
Biotinylated Anti-RB IgG Goat Vector BA-1000
Biotinylated Anti-Mouse IgG Horse Vector BA-2001
Biotinylated Anti-Goat IgG Rabbit Vector BA-5000
CY 3-conjugated Anti-Rabbit Donkey JIR 711-165-152
CY 3-conjugated Anti-mouse Donkey JIR 715-165-150
CY 2-conjugated Anti-Rabbit Donkey JIR 711-225-152
CY 2- conjugated Streptavidin Donkey JIR 016-220-084
CY 3- conjugated Streptavidin Donkey JIR 016-160-084
CY 3-conjugated Anti-Chicken Donkey JIR 703-165-155
4.1.4. PCR primers
Cre-neu-LP (F): 5’-CAGGGTGTTATAAGCAATCCC-3’
Cre-neu-UP (R): 5’-CCTGGAAAATGCTTCTGTCCG-3’
R26RFP wt (F): 5’-AAGGGAGCTGCAGTGGAGTA-3’
R26RFP wt (R): 5’-CCGAAAATCTGTGGGAAGTC-3’
GFP Common: 5’-AAAGTCGCTCTGAGTTGTTAT-3’

4. MATERIAL AND METHODS
23
GFP Wildtype: 5-GGAGCGGGAGAAATGGATATG-3’
Krt15 wt (F): 5’-GCTGGTATTGGTGTCAGAGAAG-3’
Krt15 wt (R): 5’-CCTGCACCAGACACTTAGATTT-3’
4.1.5. Materials used in cell culture
Biolite 6 Well Thermo Scientific 130184
Biolite 12 Well Thermo Scientific 130185
Biolite 24 Well Thermo Scientific 930186
Sterile filter - Puradisc 25 mm Whatman 6780-2504
Sterile filter - Minisart 0.2 µm sartorius stedim 16532
DPBS Life technologies 14190-136
DMEM Corning cellgro 10-013-CV
0.05% Trypsin-EDTA Life technologies 25300-054
Penicillin Streptomycin Life technologies 15140-122
CMF-HBSS - Hank's Balanced Salt Solution Life technologies 88284
GlutamaxTM Thermo Scientific g1870127
Corning® Matrigel® matrix Corning® 356234
N-2 Supplement Thermo Scientific 17502048
B-27 Supplement Thermo Scientific 17504044
recombinant mEGF R&D Systems 2028-EG
CHIR99021 Cayman Chemical 13122
4.1.6. Instruments and software
Microscope - Eclipse E600
Nikon E600
Mecury Lamp power supply
Nikon C-SHG1
MAC5000 power base - manual shutter LUDL electronics 73005001
Camera 12bit
Q IMAGING 01-QIC-M12C
NIS - Imaging Software
Nikon
iVision - Imaging Software
Bio Vision Technologies
Tissue-Tek VIP 5
Sakura
Thermo Shandon Histocentre 2
Sakura RM2255
4. MATERIAL AND METHODS
24
Automated Microtome
Leica 22-047-239
TBS Tissue Floating Bath
Fisher Scientific 6846
Isotemp Incubator
Fisher Scientific 11-690-637D
pH Meter - accumet basic
Fisher Scientific AB15
pH Buffer 7.00
Thermo Scientific Orion 910107
pH Buffer 4.01
Thermo Scientific Orion 910104
Microwave oven
General Electric Co. JES1460DN1
Pressure cooker-Antigenretriever
Aptum Biologics #R2100US
Isotemp Incubator
Fisher Scientific 11-690-650D
Millipore UV
Synergy SYNSV0000
Nano Drop 2000 Thermo Scientific E112352
Universal Hood ii (visualizing gels) BIO RAD 721BR08505
Real-Time PCR System life technologies 272002766
2720 Thermal Cycler life technologies 4359659
Centrifuge Kendro Laboratory 75004377
Hoshizaki ice maker Hoshizaki F344-HAF
UV-VIS spectrophotometer Beckmann Coulter DU 640
Freezer -120 Thermo Electron Corp. 7404
Electrophoresis System Fisher Scientific FB 105
Millipore UV Synergy SYNSV0000
Balance Mettler Toledo PM4000
Small incubator for single cell suspension Fisher Scientific Isotemp 202
PIPETTMANN P2 Gilson F144801
PIPETTMANN P10 Gilson F144802
PIPETTMANN P20 Gilson F123600
PIPETTMANN P100 Gilson F123615
PIPETTMANN P1000 Gilson F123602

4. MATERIAL AND METHODS
25
4.1.7. Consumable supplies
25 mL Tube Thermo Scientific 339652
50 mL Tube Thermo Scientific 339650
GeneJET Plasmid Miniprep Kit Thermo Scientific K0503
GeneJET Gel Extraction Kit Thermo Scientific K0692
GeneJET Genomic DNA Purification Kit Thermo Scientific K0721
GeneJET PCR Purification Kit Thermo Scientific K0702
GeneJET RNA Purification Kit Thermo Scientific K0732
Microcentrifuge Tube DOT scientific inc. RN1700-OMT
1-10 µl Pipet Tips DOT scientific inc. ESN-P10XLGS
1-200 µl Pipet Tips DOT scientific inc. ESN-0200NBG
Clear Adhesive Film Micro Amp 43011
High sided Low profile PCR Plate Thermo Scientific AB-1900
Fast optical 96-well reaction plate Life technologies 4346906
TaqMan Reverse Transcriptase Fisher Scientific N8080234
Power SYBR Green Master Mix Life technologies 4367659
4.1.8. Animal care
Plastic mouse cage Ancare AN75
Mouse cage lid Ancare N10
Stainless steel card holder Ancare HH35
Micro filter top Ancare R20MBT
Rodent diet Pico Labs #5053
Irradiator RS2000 RAD source

4. MATERIAL AND METHODS
26
4.2. Animal experimental methodology The Institutional Animal Care and Use Committee of the University of Pennsylvania approved all
animal studies. All experiments were designed and executed in respect to mouse protocol #805215.
4.2.1. Origin of transgenic mice used in the experiments
• Krt15-crePR1 mice were provided by Dr. George Cotsarelis, a collaborating partner at the
University of Pennsylvania and recently published in (Morris et al., 2004)
• Krt15-/- mice were provided by Dr. George
• C57BL/J6 wild-type mice (WT) were purchased from Jackson Laboratories, Lot Nr.
000664 and recently published in (Mouse Genome Sequencing Consortium et al., 2002)
• Rosa26mTomato/mGFP (R26mT/mG) mice were purchased from Jackson Laboratories, Lot Nr.
007576 and recently published in (Lozano et al., 2012)
• Apcfl/fl mice were obtained from the NCI Mouse Repository (Kuraguchi et al., 2006)
• Rosa26LSL-td Tomato(R26Tom) were kindly provided by Dr. Christopher Lenger (University of
Pennsylvania)
4.2.2. Animal care
Krt15-crePR1 and Krt15-/- mice were generated by a collaborating team at the Transgenic and
Chimeric Mouse Facility of the University of Pennsylvania School of Medicine and described
previously by a collaborating team (Morris et al., 2004). C57BL/J6 wild-type mice (WT),
Rosa26mTomato/mGFP (R26mT/mG) reporter mice and inducible DTR Rosa26iDTR (R26iDTR) mice were
obtained from Jackson Laboratories. All animals were held in the animal care facility of the
biomedical department of the University of Pennsylvania. The facility fits ABSL2 standard and
was managed by the AAALAC approved University Laboratory Animal Resource (ULAR)
division. The animals were subject of 12-hour night and day cycle and had free access to water
and food. The mice were housed in AN75 Mouse Cages offering 75 square inches of floor space
on dust free softwood kindling. The room temperature was of 22 ± 2 °C und the relative air humidity
set to 55%.

4. MATERIAL AND METHODS
27
4.2.3. Genotyping procedure
Tissue samples were collected from the tail of the mice and placed on ice. We extracted DNA from
the tissue using the GeneJET Genomic DNA Purification Kit. Individual PCR was performed to
genotype each allele. Each sample DNA was tested via PCR against a positive and negative
control.
4.2.4. Euthanasia and organ harvest
Mice were sacrificed at different time points using a CO2 inhalation chamber. Following CO2
inhalation, we ensured the successful sacrifice via cervical dislocation. We harvested tongue,
esophagus, stomach, small intestine and colon via sagittal incision from the abdomen to the thorax.
Harvested tissue was immediately prepped for tissue fixation.
4.2.5. Cre / LoxP recombination
The Cre / LoxP recombination is a common method in molecular biology used for gene insertion,
deletion or translocation at a specific LoxP site in genome. The Cre recombinase is an enzyme
derived from P1 Bacteriophages (Abremski and Hoess, 1984) catalyzing DNA recombination
between two short DNA recognition sequences. Those sequences are called loxP (locus of {X}
over P1) and consist of 34 base pairs (bp) specific for each loxP site. Each LoxP site consists of
two 13-bp palindromic sequences that are aligned on both sides of an asymmetric 8-bp spacer
region (Gierut et al., 2014). The basic strategy for Cre/LoxP-directed gene knockout experiments
is to flank, or ‘‘flox’’ the target gene with two loxP sites; the orientation of the two loxP sites
determines whether the DNA sequence is being excised or inverted by the Cre. By using ligand
depended Cre such as CreERT2 or CrePR1 in vivo, inducibility can be achieved. Inducible Cres
are commonly linked to an estrogen or progesterone receptor and inactive until induction.
Synthetic receptor ligands such as RU486 for the CrePR1 can penetrate the cell nucleus and
activating the inducible Cre.
4.2.6. Experimental strategy for the Krt15-crePR1; R26mT/mG mouse.
Krt15-crePR1 mice were bred with R26mT/mG and 6-week old mice were used for lineage tracing
experiments. Cre recombination was induced in Krt15-crePR1; R26mT/mG mice by intraperitoneal
(IP) injection of 0.5 mg of RU486 (Sigma-Aldrich). RU486 was dissolved in ethanol and then

4. MATERIAL AND METHODS
28
diluted in peanut oil. RU486 was administered daily for five continuous days and mice were then
sacrificed at different time points. Tongue, esophagus, forestomach and intestinal tissues were
removed and transferred into zinc formalin fixative. Appropriate control was achieved with
untreated Krt15-crePR1; R26mT/mG mice harvested on the same time points as the experimental
mice.
Figure 2: Breeding strategy for the Krt15-crePR1; R26mGFP mouse.
Krt15-crePR1 mice were bred with the Rosa2 mTomato/mGFP reporter mice. At 6 weeks of age the mice were injected with RU486, a
synthetic progesterone receptor agonist also known as Mifepristone. RU486 induces the ligand dependent CrePR1 which would
then excise the R26 mTomato Sequence at the loxP site. After deletion of the mTomato cassette only recombined Cells with active
Krt15 promoter will express mGFP, thus enabling lineage tracing.
4.2.8. Experimental strategy for Krt15-crePR1; R26Tom mice.
Krt15-CrePR1 mice were bred with R26Tom mice and the progeny mice were used for 3D organoid
culture. Cre recombination was induced in six- to eight-week old Krt15-CrePR1;R26Tom mice by
R26 mTomato R26 mGFPCre PR1Krt15 PromoterLoxP LoxP
Cre PR1Krt15 Promoter R26 mTomato R26 mGFP
Inject with RU486
Cre PR1Krt15 Promoter R26 mGFP

4. MATERIAL AND METHODS
29
a single I.P. injection of 0.5mg RU486. Mice were sacrificed 24 hr. later and the small intestines
were harvested. Tissues were processed as described below for crypt culture and/or single cell
sorting.
4.2.9. Experimental strategy for irradiation of Krt15-crePR1; R26mT/mG and Krt15-/- mice.
Krt15-crePR1; R26mT/mG were injected with 0.5 mg of RU486 to induce recombination. C57BL/J6
wild-type mice and Krt15-/- knockout mice did not receive Ru486 treatment. Furthermore, all mice
were administered one injection of BrdU (Sigma-Aldrich) 1,5 h prior to sacrifice (Kalabis et al.,
2008). After 5 consecutive days of RU486 treatment we irradiated the mice with a 12 Gy whole
body dose of radiation. Mice were sacrificed 2 and 5 days after irradiation treatment. The gut was
removed and transferred to zinc formalin fixative. After following fixation protocol, the tissues
were embedded in paraffin and cut using the microtome in sections 5 μm of debts for IF and IHC
staining.
Figure 3: Experimental strategy for Krt15-crePR1; R26mT/mG mouse irradiation.
The Krt15-crePR1; R26mT/mG mice as well as the WT mice were treated with RU486 for 5 days to ensure high recombination rate.
After 5 days the mice were irradiated at 12 Grey ablating most radiosensitive intestinal stem cells. Following irradiation, the mice
were sacrificed at 2 and 5 days post treatment.
4.2.10. Experimental strategy for Krt15-CrePR1;Apcfl/fl;R26mT/mG mice
Krt15-crePR1; R26mT/mG were bred with Apcfl/fl mice to obtain Krt15-CrePR1;Apcfl/fl;R26mT/mG
mice. Cre recombination was induced using daily injections of 0.5mg of RU486 for five
consecutive days. Experimental mice were sacrificed 150 days post recombination or at prior time
point in concert with severe weight loss.
Inject with RU486for 5 days
Cre PR1Krt15 Promoter R26 mGFP
Irradiationwith 12 Grey
Sacrifice and analysis 2 & 5 days following irradiation

4. MATERIAL AND METHODS
30
4.3. Tissue fixation Extracted tissues were splayed open and sandwiched between pieces of filter paper to fix flat. The
tissues, wedged in between filter paper, were then placed in small tissue fixation cassettes and
emerged in a 4% paraformaldehyde (PFA) solution at 4 °C overnight. The following day, we
washed the tissue samples with 1X PBS for 60 minutes and then transferred the samples to a 70%
Ethanol solution at 4°C. The cassettes could then be processed with Tissue-TEK VIP for paraffin
embedding. The paraffin embedded tissue could then be cut in sections 5 μm of debts, using the
microtome. Each section was placed on a glass slide and left to dry at 60°C over night.
4.4. Immunohistochemistry (IHC) To dewax the slides, we preheated them using a 60°C incubator for 15 minutes, after which the
slides were placed in a Xylene bath. The slides were kept in Xylene for 3 minutes and then moved
to a second Xylene bath for the same time-period. Subsequently, the slides were then moved into
100% Ethanol twice for two minutes per interval. Following this step, the slides were then moved
from 95% Ethanol to 80% and then to 70% every 1 minute. Finally, the slides were rinsed in milliQ
water for 1 minute before antigen retrieval. Antigen retrieval was performed by pressure cooking,
immersing the slides in citric acid (pH 6.0) buffer. We incubated the slides for 20 minutes within
the pressure cooker and then left them to cool for 40 minutes. Once the slides cooled to room
temperature, we gently rinsed them with milliQ water for 5 minutes. After that, the slides were
quenched in 30% hydrogen peroxide for 15 minutes, subsequently eliminating the endogenous
peroxidase reaction. Before blocking, slides were rinsed off with milliQ water and 1 X PBS for 5
minutes each. We used a hydrophobic pen to restrict the volume of the reagents applied to each
section. For IHC blocking we used a Biotin/Avidin system. This system is based on the fact that
the rather large Glycoprotein Avidin depicts a strong affinity to the Biotin molecule, thus forming
Avidin/Biotin structures on non-specific antibody loci (Hsu et al., 1981). Avidin D blocking
reagent (Vecta) was applied first for a time-period of 15 minutes and then rinsed off with 1 X PBS
for 5 minutes. Following Avidin D we applied Biotin blocking reagent (Vecta) for another 15
minutes and rinsed with 1 X PBS. As final step before antibody treatment we used unspecific
Protein Blocking Agent for 10 min. Primary antibodies were diluted in PBT and incubated
overnight at 4°C. Following 3 washing steps with 1 X PBS of 5 minutes each slide was then

4. MATERIAL AND METHODS
31
incubated with biotinylated secondary antibodies diluted in PBT for 30 min at 37°C. Next we used
the ABC Reagent (Vector Laboratories) for an additional 30 min at 37°C. Finally, slides were
treated with DAB substrate, making sure that consistent exposure time within each slide was set
and counterstained with hematoxylin for 5 seconds.
4.4.1. IHC in combination with the M.O.M.-kit
The M.O.M.™ immunodetection Kit was specifically designed by Vector®, to reduce endogenous
mouse IgG staining when using mouse primary antibodies on mouse tissue (He et al., 2013).
Specific mouse horse radish peroxidase (HRP) polymers significantly reduce background from
mouse antigens and thereby allow us to use mouse K15 antibodies on our WT and K15-/- mice.
4.5. Immunofluorescence (IF) Sections were rehydrated in the same fashion as described in 2.4 and underwent the same antigen
retrieval steps. Contrary to the IHC slides, IF sections were blocked using an adapted blocking
buffer with 1% BSA, 0,3% Triton X and 5% Donkey serum in 1X PBS for 1h at room temperature.
Following blocking steps, the slides were rinsed off with milliQ Water and incubated with primary
antibodies diluted in 1% BSA overnight at 4°C. Sections were then incubated with secondary
antibodies for 1h at room temperature and counterstained with DAPI for 1 minute.
4.5.1. Immunofluorescence using the TSA system
The Tyramide Signal Amplification (TSA™) is a HRP-mediated method that was developed in
the late 1990s to detect proteins and polynucleotides in situ (van Gijlswijk et al., 1996). Sensitivity
levels using TSA systems are greatly higher than the common HRP based methods (Liu et al.,
2006). TSA Biotin Kits use HRP to catalyze covalent deposition of biotin labels directly adjacent
to the immobilized enzyme. The labeling reaction is quick (less than 10 minutes) and deposited
labels can be detected with streptavidin conjugates for imaging in brightfield or fluorescence
microscopy. After overnight incubation with primary antibodies and consecutive washing steps
with PBS, we added biotinylated secondary antibodies, incubating them for 30 minutes at 37°C.
Following washing, we added the streptavidin-HRP (Perkin Elmer) for 30 minutes at RT. Then,
we washed the slides with PBS and added the biotinylated Tyramide Reagent (Perkin Elmer) for
10 minutes at RT. We then added the CY2 conjugated Streptavidin binding the Tyramide.

4. MATERIAL AND METHODS
32
Subsequently, this enabled us to stain another marker using a tertiary antibody from another
species and CY3 conjugated for antibodies.
4.6. Alcian Blue staining (AB) Sections were rehydrated in the same fashion as described in 4.4. and underwent the same antigen
retrieval step. The slides were then quenched with hydrogen peroxide and blocked with Avidin,
Biotin and BSA as described in 4.4. Primary antibodies were diluted in PBT and applied overnight
at 4°C. Following the washing steps with 1 X PBS, the slides were then incubated with biotinylated
secondary antibodies diluted in PBT for 30 min at 37°C. Next we used the ABC Reagent (Vector
Laboratories) for an additional 30 min at 37°C. The slides were developed with DAB substrate for
10 seconds and immediately washed in milliQ H2O. The slides were then placed in 3% acetic acid
for 3 minutes. The samples were then treated with 1% Alcian Blue in 3% acetic acid (pH2.5) for
30 minutes. After a 10-minute wash in milliQ water we counterstained the samples with Nuclear
fast RED 0.1% for 40 seconds. After counterstaining, we dehydrated the slides and placed them in
Xylene for mounting.
4.7. Single cell isolation Crypt single cells were isolated as described previously by our collaborators (Hamilton et al.,
2015). Briefly, the small intestine was opened longitudinally and washed with cold PBS. Tissue
was incubated 20 min on ice in PBS-EDTA-DTT. Tissues were then incubated in PBS-EDTA at
37°C. Crypts were isolated from the epithelial fraction using a 70μm filter. Crypts were then
dissociated to a single cell suspension using PBS/dispase at 37°C.
4.8. Crypt isolation and 3D organoid culture Crypts were isolated from the mouse small intestine. Tissues were chilled in Ca2+-Mg2+-free HBSS
(CMF-HBSS) with 1mM N-acetyl-cysteine (NAC). Tissues were then incubated in CMF-
HBSS/1mM NAC/10mM EDTA for 45min at 4°C. Epithelial cells were then dissociated through
vortexing/resting cycles. Crypts were separated from epithelial dissociation with 70μm filter.
Crypts were pelleted and resuspended in Basal Media (Advanced DMEM/F12, 2mM GlutamaxTM,
10mM HEPES, 1X Penicillin/Streptomycin, 5μM CHIR99021 (Cayman Chemical), 1mM NAC,

4. MATERIAL AND METHODS
33
1x N2 Supplement (Gibco) and 1X B27 Supplement (Gibco)). Approximately 500 crypts were
then embedded in 80% Matrigel/20% ENR (Basal Media, 50ng/ml recombinant mEGF(R&D
Systems) and 1% Noggin/R-Spondin conditioned media). Enteroids were cultured in ENR.
Figure 4:Experimental strategy for 3D organoid structures
Krt15-CrePR1;R26mTom/mGFP mice were injected with 1 dose of RU486 sacrificed 24h post injection. Intestinal tissue was then
extracted and crypts were isolated following the Hamilton protocol. Isolated crypts were then transferred into matrigel matrix for
3D organoid culturing.
4.9. Irradiation Six to eight-week old mice were subjected to 12Gy whole-body gamma-irradiation (Gammacell
40 Cesium 137 Irradiation Unit). Radiation was used as the cytotoxic model, as dose delivery
throughout the epithelial tissue is known to induce stem cell renewal (Otsuka et al., 2013). Mice
were sacrificed 5 days later and the small intestines were harvested and fixed for histology.
Surviving crypts, crypt length and villi height were measured on H&E stained ileum sections.
Twenty-five high magnification fields were analyzed for each mouse and a minimum of 100 crypts
or villi was measured for each mouse.
4.10. Histometric analysis 4.10.1. Crypt length analysis
Measurements were performed on small intestinal crypts and villi using an Eclipse E600
microscope with NIS imaging software. For each individual mouse, crypt depth was measured in
10–15 intestinal circumferences. An intestinal circumference is a convenient section of intestinal
epithelium representing a relative standard length and width of the individual mouse (Booth et al.,
2004). Out of those 10–15 circumferences we measured 120 crypts per animal. Where possible,
the crypts and villi measured were continuously indicating a true longitudinal section. Crypt and
Tissue extraction &crypt isolation
Transfer isolated crypts into 3D organotypic culture

4. MATERIAL AND METHODS
34
villus height were measured via ImageJ and the data presented as a group which means a change
in size from WT control.
4.10.2 Crypt microcolony assay
The term microcolony was first used in 1970 (Withers and Elkind, 1970) to describe regenerative
crypts post injury which were believed to repopulate the intestinal epithelium. The number of
surviving and regenerating crypts per intestinal circumference was scored and the average per
mouse and per group determined. A surviving crypt was defined as one that had 10 or more BrdU+
cells. Only regions that were orientated correctly and did not contain Peyers patches were scored
(Booth et al., 2015; Jones et al., 2015; Potten et al., 1994). Cytotoxic damage caused by ionizing
radiation accelerated stem cell renewal, thus accumulating BrdU + proliferating cells (Roe et al.,
1996).
4.10.3. Software and statistical analysis
All tables presented in this dissertation were analyzed for statistical relevance, using the unpaired
student T-Test. Calculations were done via GraphPad Prism® 6.0, applying a confidence interval
of 99%.
5. RESULTS
35
5. RESULTS 5.1. Endogenous expression of Krt15 in the small intestine. Our first aim is directed at proving Krt15 expression in the intestinal epithelium of the small
intestine. Furthermore, we were interested in the exact location of Krt15 expressing cells within
the structure of the intestinal crypt and villi. To minimize the risk of false positive data, we planned
two separate experiments targeted at the same inquiry. On one hand, we looked at the endogenous
expression of Krtr15 in WT mice via IHC and on the other hand, we inspected the expression of
GFP via IF in Krt15-crePR1; R26mT/mG mice.
5.1.1. Endogenous expression of Krt15 in WT mice
We sacrificed the 4 mice at the age of 6 months, harvested the small intestine and conducted an
IHC staining as described previously. We stained directly for K15 using a rabbit antibody created
by Abcam (ab52816). As a negative control, we used a Krt15 knockout mouse provided by Dr.
George Cotsarelis. To ensure validity of our antibodies, we used sections of mouse esophagus as
positive controls.
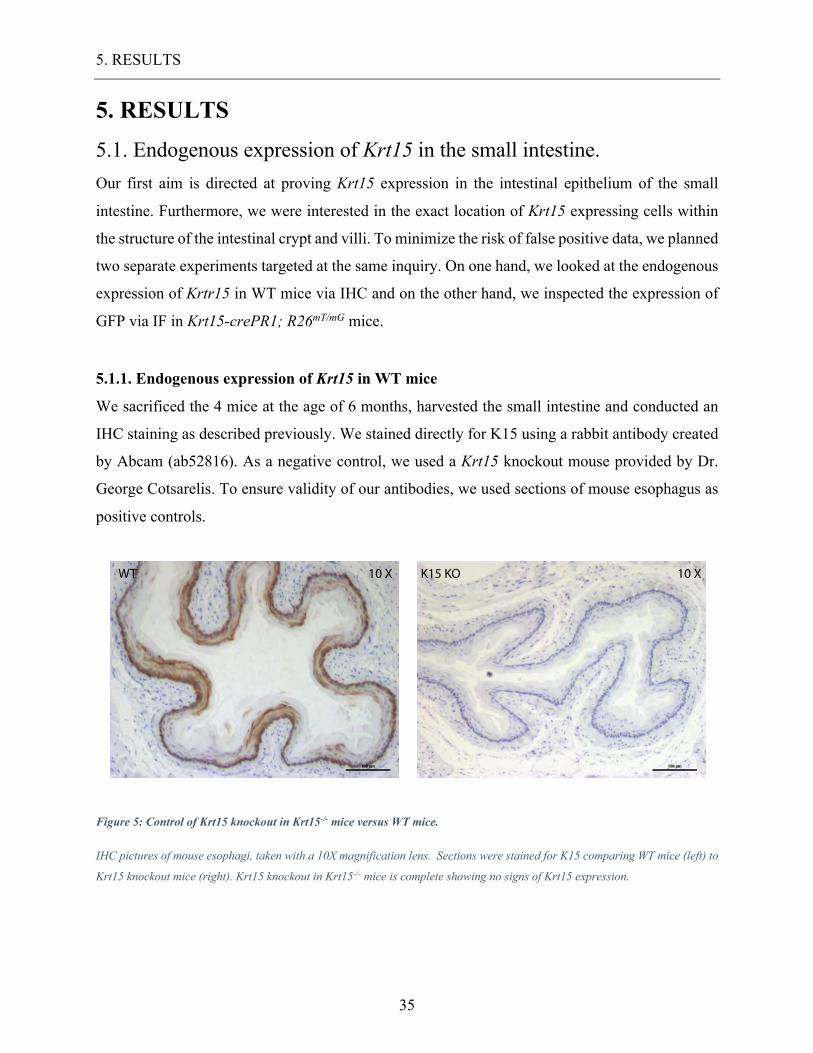
Figure 5: Control of Krt15 knockout in Krt15-/- mice versus WT mice.
IHC pictures of mouse esophagi, taken with a 10X magnification lens. Sections were stained for K15 comparing WT mice (left) to
Krt15 knockout mice (right). Krt15 knockout in Krt15-/- mice is complete showing no signs of Krt15 expression.
10 X K15 KO 10 XWT
5. RESULTS
36
Krt15-/- mice demonstrated a complete knockout of Krt15 contrary to the consistent expression of
Krt15 in basal compartment of the esophageal epithelium. We concluded that the antibody used
specifically recognize K15. Small intestinal staining demonstrated similar specificity apart from
background staining in the stroma of the villi. These results demonstrate the existence of Krt15
expressing cells in the intestinal epithelium. Krt15+ cells were found at two different locations
within the intestinal epithelium: One group of Krt15 expressing cells was found in the villi, the
second group of Krt15 expressing cells in the crypt.
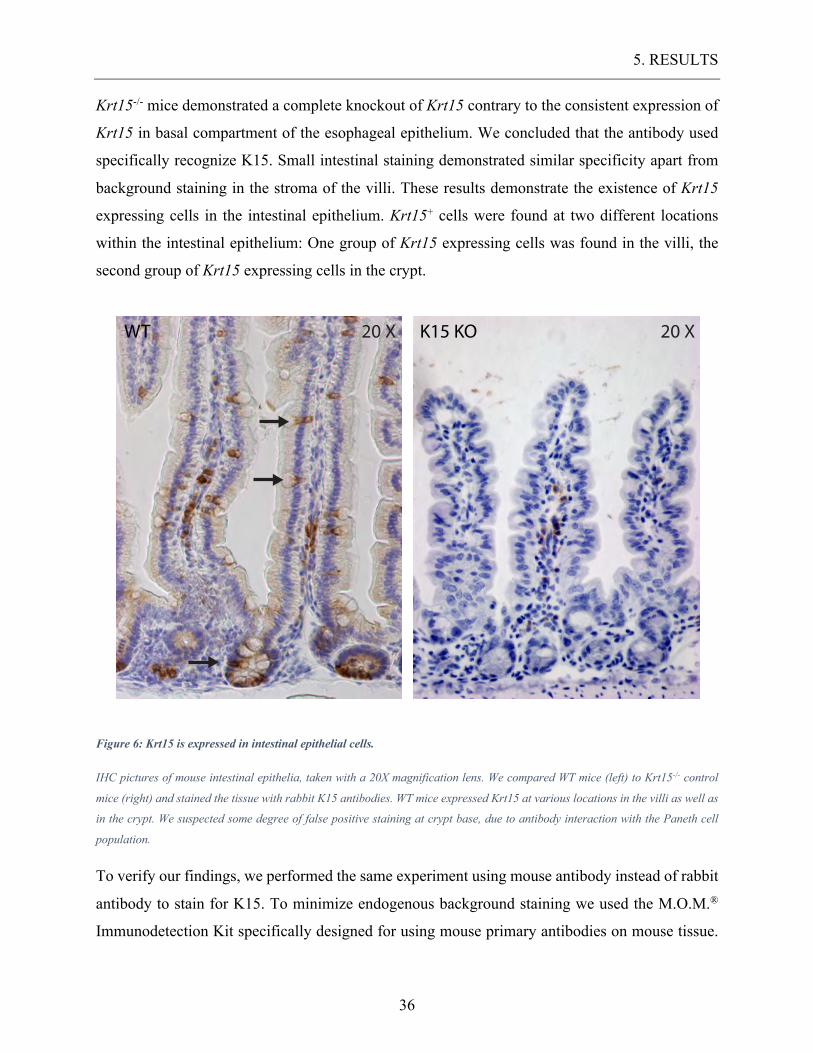
Figure 6: Krt15 is expressed in intestinal epithelial cells.
IHC pictures of mouse intestinal epithelia, taken with a 20X magnification lens. We compared WT mice (left) to Krt15-/- control
mice (right) and stained the tissue with rabbit K15 antibodies. WT mice expressed Krt15 at various locations in the villi as well as
in the crypt. We suspected some degree of false positive staining at crypt base, due to antibody interaction with the Paneth cell
population.
To verify our findings, we performed the same experiment using mouse antibody instead of rabbit
antibody to stain for K15. To minimize endogenous background staining we used the M.O.M.®
Immunodetection Kit specifically designed for using mouse primary antibodies on mouse tissue.
K15 KO 20 X20 XWT
5. RESULTS
37
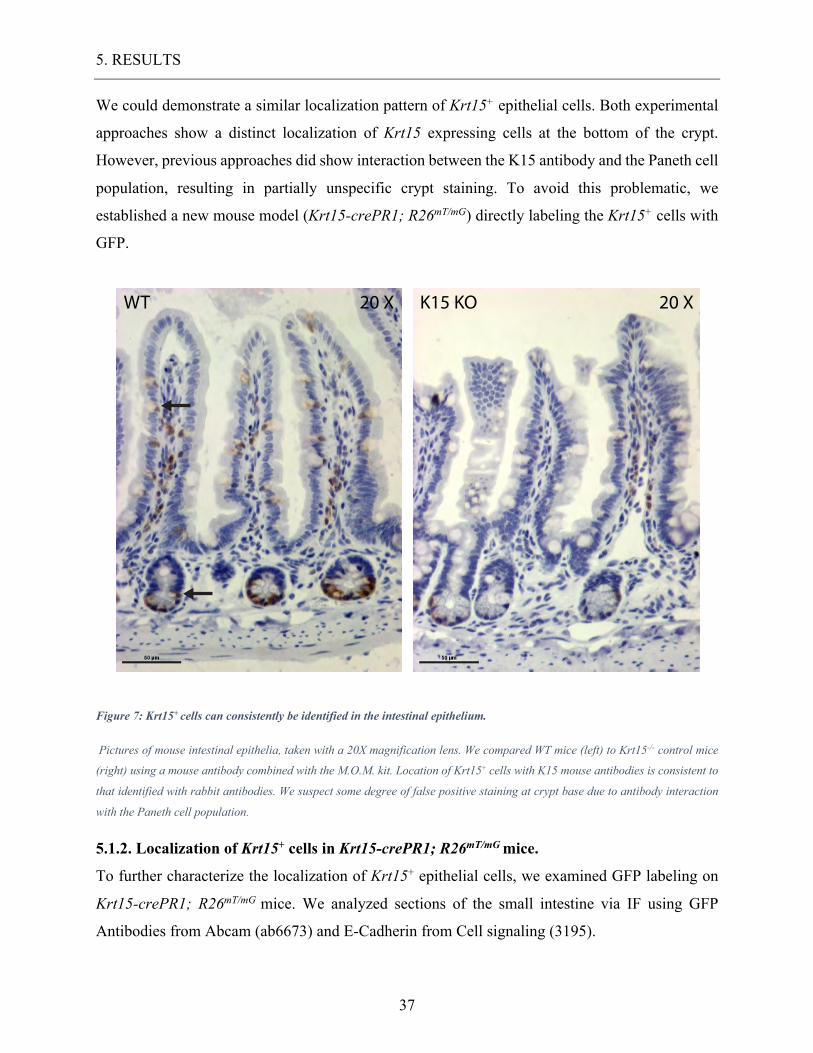
We could demonstrate a similar localization pattern of Krt15+ epithelial cells. Both experimental
approaches show a distinct localization of Krt15 expressing cells at the bottom of the crypt.
However, previous approaches did show interaction between the K15 antibody and the Paneth cell
population, resulting in partially unspecific crypt staining. To avoid this problematic, we
established a new mouse model (Krt15-crePR1; R26mT/mG) directly labeling the Krt15+ cells with
GFP.
Figure 7: Krt15+ cells can consistently be identified in the intestinal epithelium.
Pictures of mouse intestinal epithelia, taken with a 20X magnification lens. We compared WT mice (left) to Krt15-/- control mice
(right) using a mouse antibody combined with the M.O.M. kit. Location of Krt15+ cells with K15 mouse antibodies is consistent to
that identified with rabbit antibodies. We suspect some degree of false positive staining at crypt base due to antibody interaction
with the Paneth cell population.
5.1.2. Localization of Krt15+ cells in Krt15-crePR1; R26mT/mG mice.
To further characterize the localization of Krt15+ epithelial cells, we examined GFP labeling on
Krt15-crePR1; R26mT/mG mice. We analyzed sections of the small intestine via IF using GFP
Antibodies from Abcam (ab6673) and E-Cadherin from Cell signaling (3195).
K15 KO 20 XWT 20 X
5. RESULTS
38
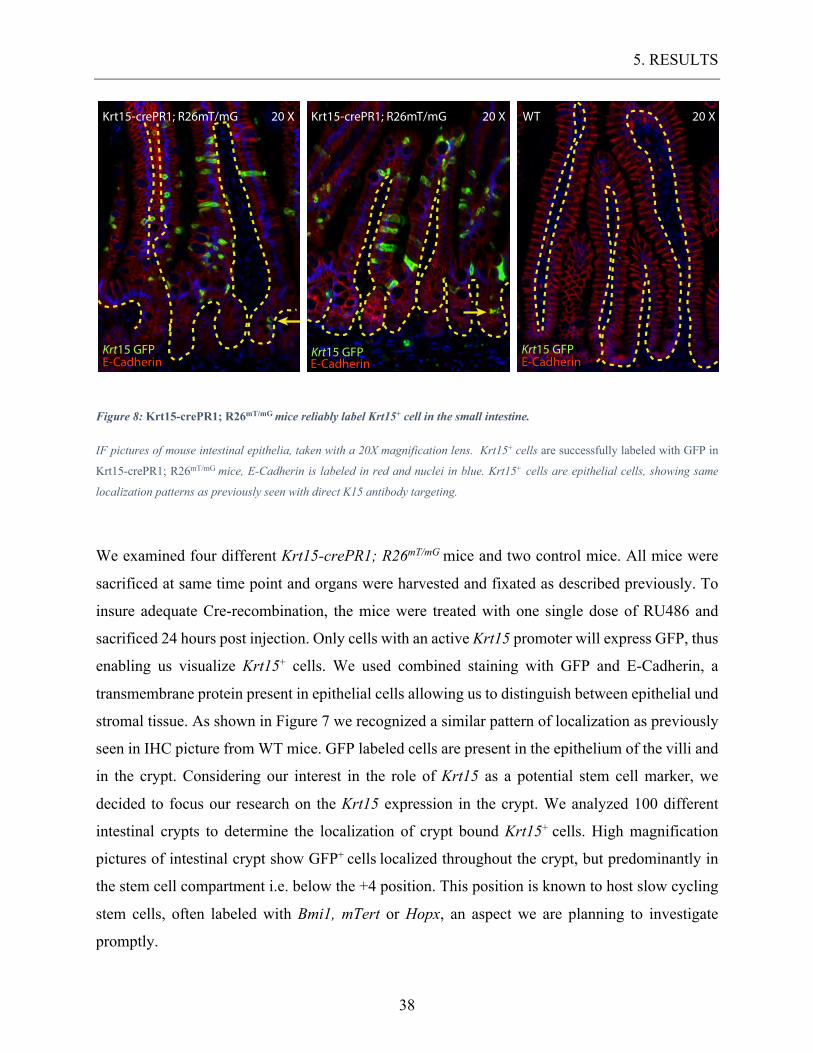
Figure 8: Krt15-crePR1; R26mT/mG mice reliably label Krt15+ cell in the small intestine.
IF pictures of mouse intestinal epithelia, taken with a 20X magnification lens. Krt15+ cells are successfully labeled with GFP in
Krt15-crePR1; R26mT/mG mice, E-Cadherin is labeled in red and nuclei in blue. Krt15+ cells are epithelial cells, showing same
localization patterns as previously seen with direct K15 antibody targeting.
We examined four different Krt15-crePR1; R26mT/mG mice and two control mice. All mice were
sacrificed at same time point and organs were harvested and fixated as described previously. To
insure adequate Cre-recombination, the mice were treated with one single dose of RU486 and
sacrificed 24 hours post injection. Only cells with an active Krt15 promoter will express GFP, thus
enabling us visualize Krt15+ cells. We used combined staining with GFP and E-Cadherin, a
transmembrane protein present in epithelial cells allowing us to distinguish between epithelial und
stromal tissue. As shown in Figure 7 we recognized a similar pattern of localization as previously
seen in IHC picture from WT mice. GFP labeled cells are present in the epithelium of the villi and
in the crypt. Considering our interest in the role of Krt15 as a potential stem cell marker, we
decided to focus our research on the Krt15 expression in the crypt. We analyzed 100 different
intestinal crypts to determine the localization of crypt bound Krt15+ cells. High magnification
pictures of intestinal crypt show GFP+ cells localized throughout the crypt, but predominantly in
the stem cell compartment i.e. below the +4 position. This position is known to host slow cycling
stem cells, often labeled with Bmi1, mTert or Hopx, an aspect we are planning to investigate
promptly.
20 XWT
Krt15 GFPE-Cadherin
Krt15-crePR1; R26mT/mG 20 X
Krt15 GFPE-Cadherin
Krt15-crePR1; R26mT/mG 20 X
Krt15 GFPE-Cadherin
5. RESULTS
39
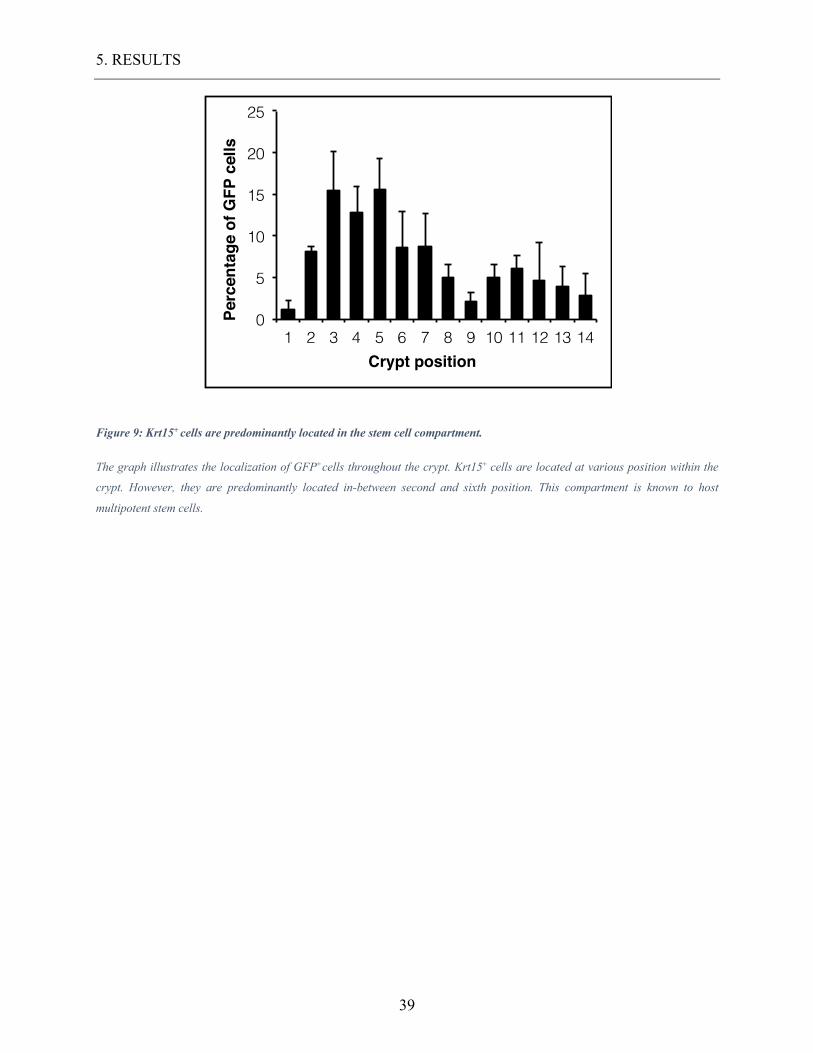
Figure 9: Krt15+ cells are predominantly located in the stem cell compartment.
The graph illustrates the localization of GFP+cells throughout the crypt. Krt15+ cells are located at various position within the
crypt. However, they are predominantly located in-between second and sixth position. This compartment is known to host
multipotent stem cells.
0
5
10
15
20
25
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Perc
enta
ge o
f GFP
cel
ls
Crypt position
5. RESULTS
40
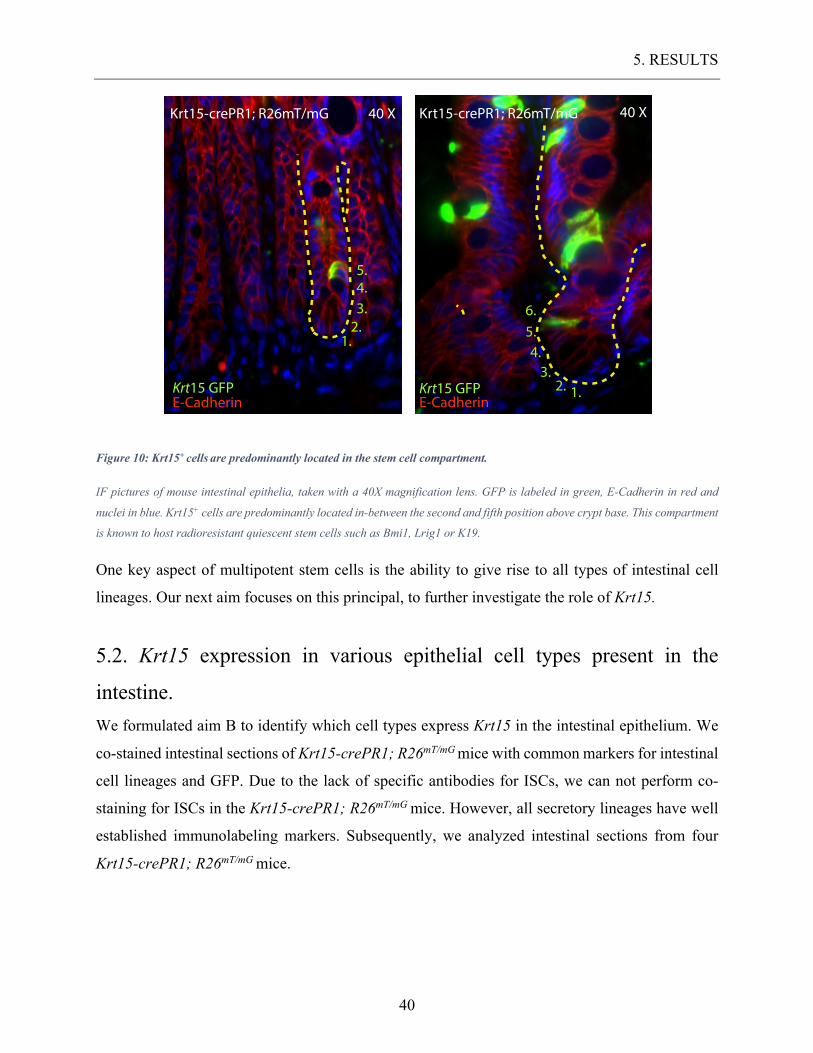
Figure 10: Krt15+ cells are predominantly located in the stem cell compartment.
IF pictures of mouse intestinal epithelia, taken with a 40X magnification lens. GFP is labeled in green, E-Cadherin in red and
nuclei in blue. Krt15+ cells are predominantly located in-between the second and fifth position above crypt base. This compartment
is known to host radioresistant quiescent stem cells such as Bmi1, Lrig1 or K19.
One key aspect of multipotent stem cells is the ability to give rise to all types of intestinal cell
lineages. Our next aim focuses on this principal, to further investigate the role of Krt15.
5.2. Krt15 expression in various epithelial cell types present in the
intestine. We formulated aim B to identify which cell types express Krt15 in the intestinal epithelium. We
co-stained intestinal sections of Krt15-crePR1; R26mT/mG mice with common markers for intestinal
cell lineages and GFP. Due to the lack of specific antibodies for ISCs, we can not perform co-
staining for ISCs in the Krt15-crePR1; R26mT/mG mice. However, all secretory lineages have well
established immunolabeling markers. Subsequently, we analyzed intestinal sections from four
Krt15-crePR1; R26mT/mG mice.
Krt15-crePR1; R26mT/mG 40 X
Krt15 GFPE-Cadherin
1.2.3.4.
Krt15-crePR1; R26mT/mG 40 X
Krt15 GFPE-Cadherin
1.2.3.
4.5.
5.
6.
5. RESULTS
41
5.2.1. Krt15 expression in enteroendocrine cells.
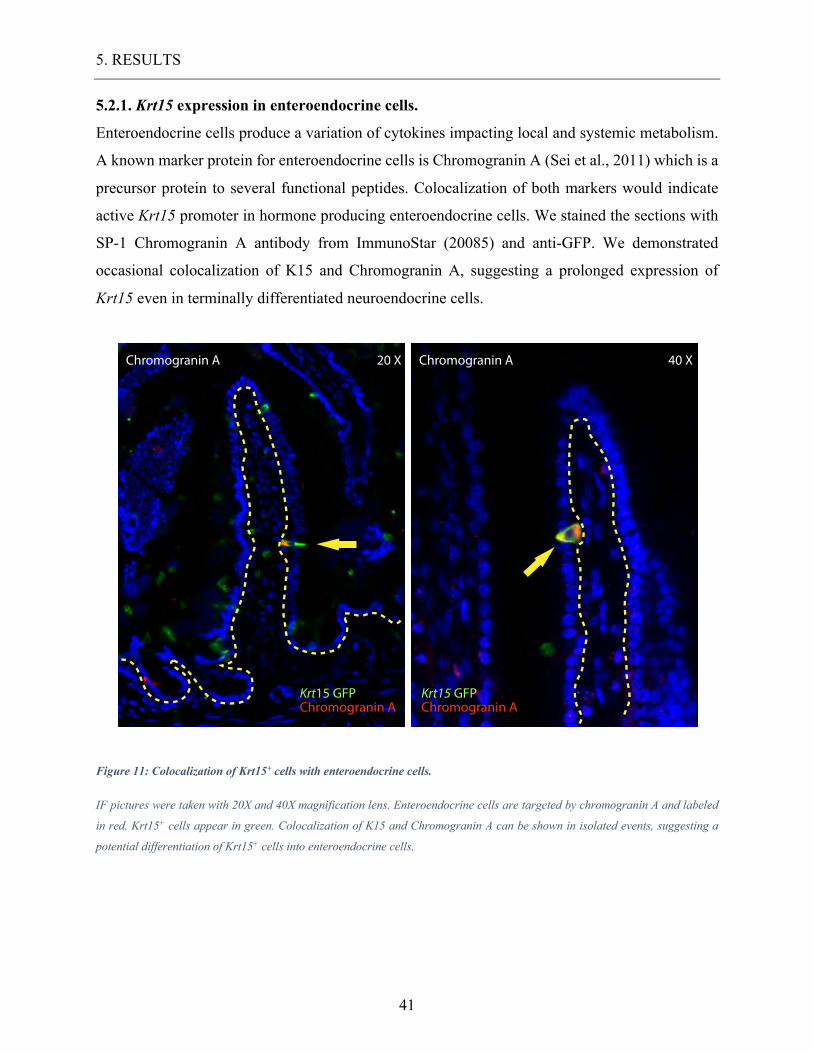
Enteroendocrine cells produce a variation of cytokines impacting local and systemic metabolism.
A known marker protein for enteroendocrine cells is Chromogranin A (Sei et al., 2011) which is a
precursor protein to several functional peptides. Colocalization of both markers would indicate
active Krt15 promoter in hormone producing enteroendocrine cells. We stained the sections with
SP-1 Chromogranin A antibody from ImmunoStar (20085) and anti-GFP. We demonstrated
occasional colocalization of K15 and Chromogranin A, suggesting a prolonged expression of
Krt15 even in terminally differentiated neuroendocrine cells.
Figure 11: Colocalization of Krt15+ cells with enteroendocrine cells.
IF pictures were taken with 20X and 40X magnification lens. Enteroendocrine cells are targeted by chromogranin A and labeled
in red, Krt15+ cells appear in green. Colocalization of K15 and Chromogranin A can be shown in isolated events, suggesting a
potential differentiation of Krt15+ cells into enteroendocrine cells.
20 XChromogranin A 40 XChromogranin A
Krt15 GFPChromogranin A
Krt15 GFPChromogranin A
5. RESULTS
42
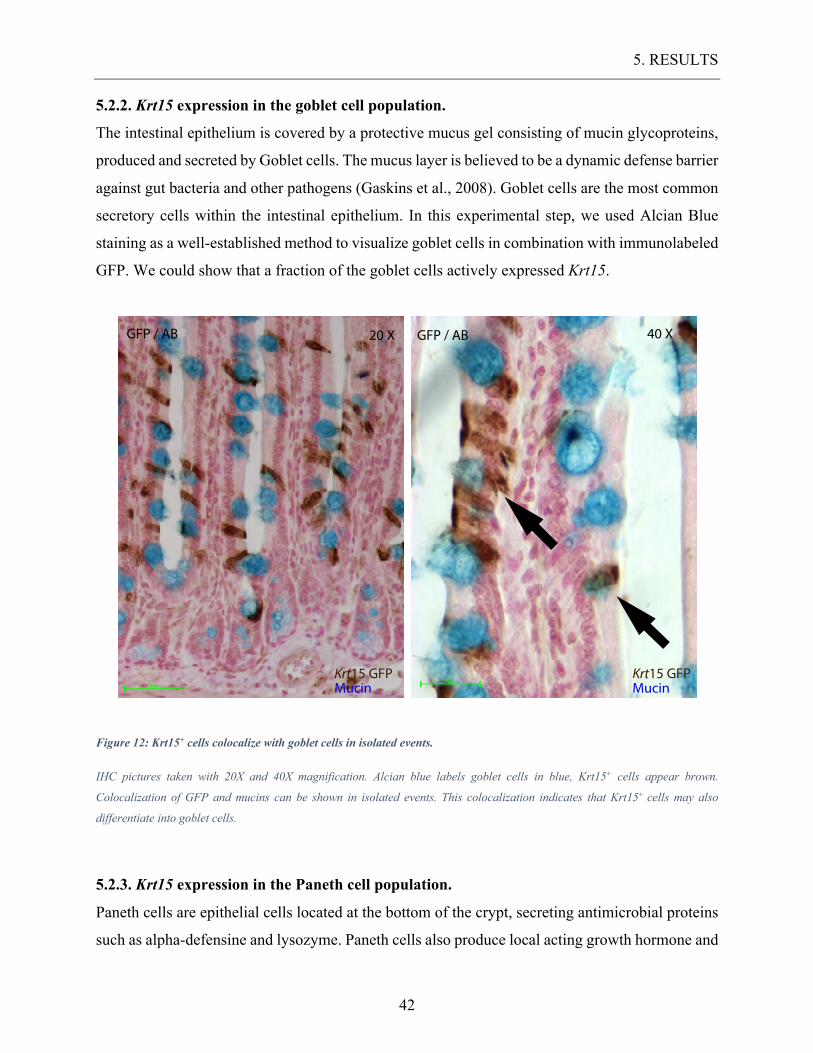
5.2.2. Krt15 expression in the goblet cell population.
The intestinal epithelium is covered by a protective mucus gel consisting of mucin glycoproteins,
produced and secreted by Goblet cells. The mucus layer is believed to be a dynamic defense barrier
against gut bacteria and other pathogens (Gaskins et al., 2008). Goblet cells are the most common
secretory cells within the intestinal epithelium. In this experimental step, we used Alcian Blue
staining as a well-established method to visualize goblet cells in combination with immunolabeled
GFP. We could show that a fraction of the goblet cells actively expressed Krt15.
Figure 12: Krt15+ cells colocalize with goblet cells in isolated events.
IHC pictures taken with 20X and 40X magnification. Alcian blue labels goblet cells in blue, Krt15+ cells appear brown.
Colocalization of GFP and mucins can be shown in isolated events. This colocalization indicates that Krt15+ cells may also
differentiate into goblet cells.
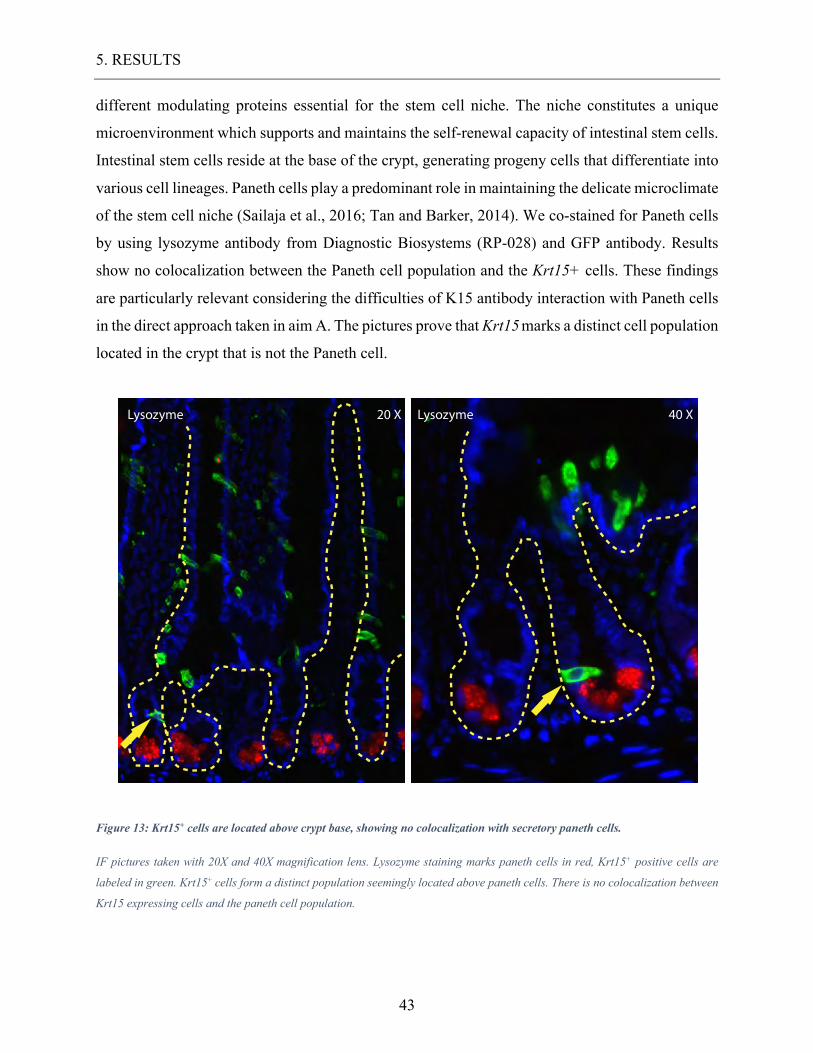
5.2.3. Krt15 expression in the Paneth cell population.
Paneth cells are epithelial cells located at the bottom of the crypt, secreting antimicrobial proteins
such as alpha-defensine and lysozyme. Paneth cells also produce local acting growth hormone and
GFP / AB 20 X GFP / AB 40 X
Krt15 GFPMucin
Krt15 GFPMucin
5. RESULTS
43
different modulating proteins essential for the stem cell niche. The niche constitutes a unique
microenvironment which supports and maintains the self-renewal capacity of intestinal stem cells.
Intestinal stem cells reside at the base of the crypt, generating progeny cells that differentiate into
various cell lineages. Paneth cells play a predominant role in maintaining the delicate microclimate
of the stem cell niche (Sailaja et al., 2016; Tan and Barker, 2014). We co-stained for Paneth cells
by using lysozyme antibody from Diagnostic Biosystems (RP-028) and GFP antibody. Results
show no colocalization between the Paneth cell population and the Krt15+ cells. These findings
are particularly relevant considering the difficulties of K15 antibody interaction with Paneth cells
in the direct approach taken in aim A. The pictures prove that Krt15 marks a distinct cell population
located in the crypt that is not the Paneth cell.
Figure 13: Krt15+ cells are located above crypt base, showing no colocalization with secretory paneth cells.
IF pictures taken with 20X and 40X magnification lens. Lysozyme staining marks paneth cells in red, Krt15+ positive cells are
labeled in green. Krt15+ cells form a distinct population seemingly located above paneth cells. There is no colocalization between
Krt15 expressing cells and the paneth cell population.
40 XLysozyme20 XLysozyme
5. RESULTS
44
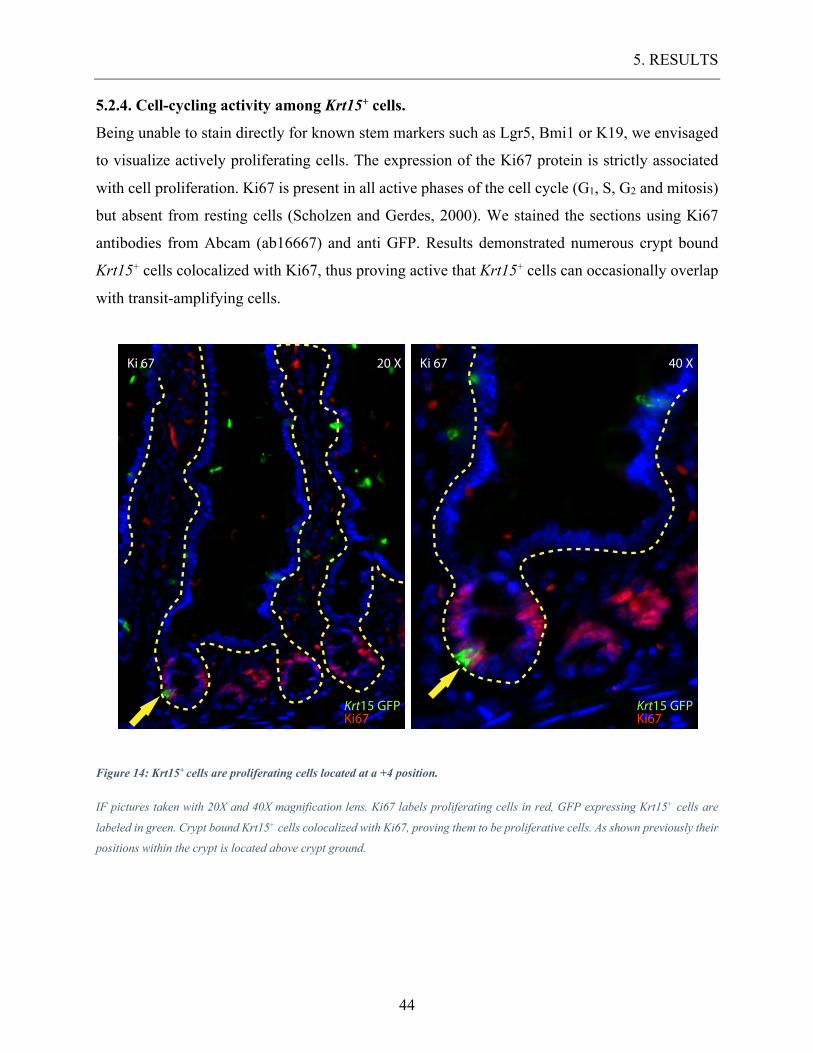
5.2.4. Cell-cycling activity among Krt15+ cells.
Being unable to stain directly for known stem markers such as Lgr5, Bmi1 or K19, we envisaged
to visualize actively proliferating cells. The expression of the Ki67 protein is strictly associated
with cell proliferation. Ki67 is present in all active phases of the cell cycle (G1, S, G2 and mitosis)
but absent from resting cells (Scholzen and Gerdes, 2000). We stained the sections using Ki67
antibodies from Abcam (ab16667) and anti GFP. Results demonstrated numerous crypt bound
Krt15+ cells colocalized with Ki67, thus proving active that Krt15+ cells can occasionally overlap
with transit-amplifying cells.
Figure 14: Krt15+ cells are proliferating cells located at a +4 position.
IF pictures taken with 20X and 40X magnification lens. Ki67 labels proliferating cells in red, GFP expressing Krt15+ cells are
labeled in green. Crypt bound Krt15+ cells colocalized with Ki67, proving them to be proliferative cells. As shown previously their
positions within the crypt is located above crypt ground.
40 XKi 6720 XKi 67
Ki67Krt15 GFP
Ki67Krt15 GFP

5. RESULTS
45
5.3. Evaluation of proliferative potential within the Krt15+ cell population.
To determine the growth of Krt15+ cells, it is necessary to examine Krt15+ progeny cells. A
common characteristic of adult stem cells it that they are multipotent, i.e. able to give rise to all
differentiated lineages throughout a tissue. The setting of a lineage tracing experiment would
enable us to investigate the Krt15+ population and all progeny cells over a long period of time.
5.3.1. Lineage tracing experiment on Krt15-crePR1; R26mT/mG mice.
We induced LoxP recombination via Ru486 injections in 6-week-old mice and sacrificed the mice
at different time points. Experimental mice were injected with Ru486 for 5 consecutive days to
insure maximum recombination. We sacrificed a total number of 49 mice over a period of 150
days at different time points (Days: 1,2,3,5,7,10,14,21,28,56,150).
This setup enables us to determine the fate of the Krt15+ population at day one post injections up
to their progeny cells at the latest time point, 150 days post induction. Considering that the small
intestinal epithelium is renewed every 3 days in the mouse except for the Paneth cells, all GFP
expressing cells past this day constitute Krt15+ progeny cells. Krt15+ progeny cells are cells arising
from the Krt15+ origin cells. Any labeled cells beyond a week shows that Krt15+ have self-
renewing capacity and are long-lived cells.
Figure 15: Timetable depicting experimental setup of the lineage tracing experiment over 150 days.
Experimental mice were treated during 5 consecutive days with RU486 to maximize induction rate. After successful induction, mice
were sacrificed at specific time points for IF contemplation. The timeframe is set to determine the vivacity and endurance of the
Krt15+ population and evaluate the fate of any daughter cells.
At early time points (day: 1,2,3,5), we observed a small population of Krt15+ cells located within
the crypt. Following time points show a graduate expansion of GFP expressing Krt15+ progeny
cells. Day 14 marks the first date on which crypts with Krt15+ cells consist of more than 50 percent
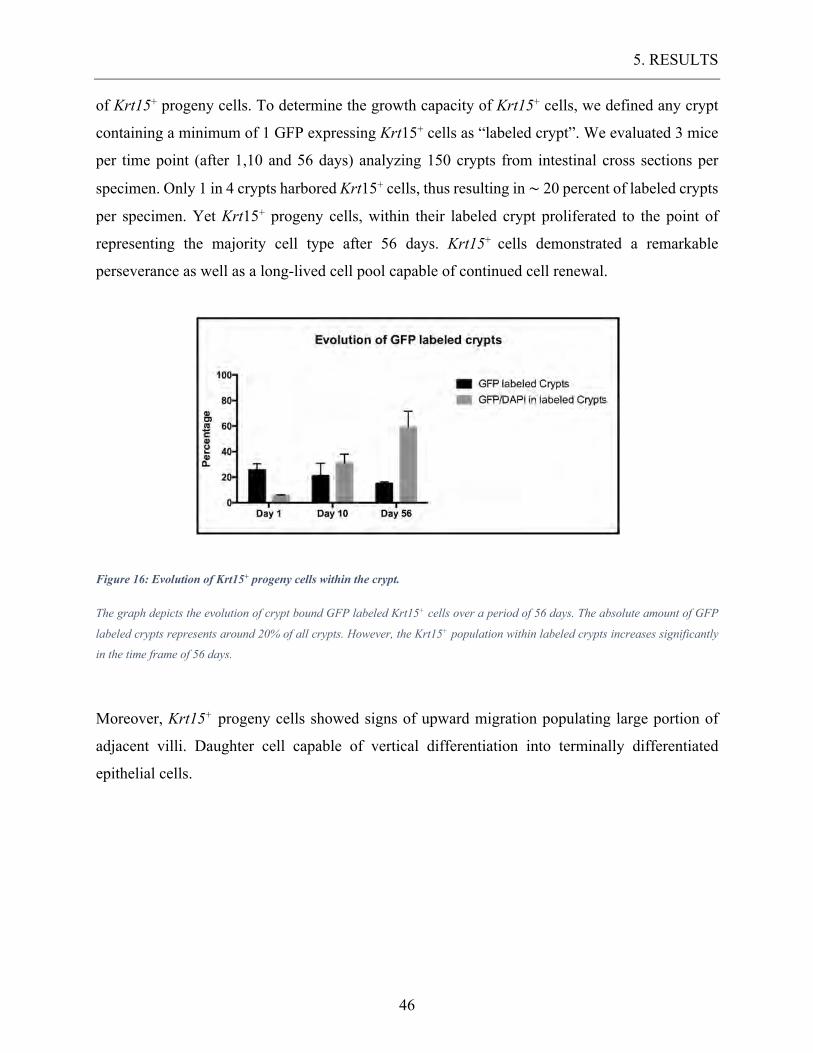
5. RESULTS
46
of Krt15+ progeny cells. To determine the growth capacity of Krt15+ cells, we defined any crypt
containing a minimum of 1 GFP expressing Krt15+ cells as “labeled crypt”. We evaluated 3 mice
per time point (after 1,10 and 56 days) analyzing 150 crypts from intestinal cross sections per
specimen. Only 1 in 4 crypts harbored Krt15+ cells, thus resulting in ~ 20 percent of labeled crypts
per specimen. Yet Krt15+ progeny cells, within their labeled crypt proliferated to the point of
representing the majority cell type after 56 days. Krt15+ cells demonstrated a remarkable
perseverance as well as a long-lived cell pool capable of continued cell renewal.
Figure 16: Evolution of Krt15+ progeny cells within the crypt.
The graph depicts the evolution of crypt bound GFP labeled Krt15+ cells over a period of 56 days. The absolute amount of GFP
labeled crypts represents around 20% of all crypts. However, the Krt15+ population within labeled crypts increases significantly
in the time frame of 56 days.
Moreover, Krt15+ progeny cells showed signs of upward migration populating large portion of
adjacent villi. Daughter cell capable of vertical differentiation into terminally differentiated
epithelial cells.
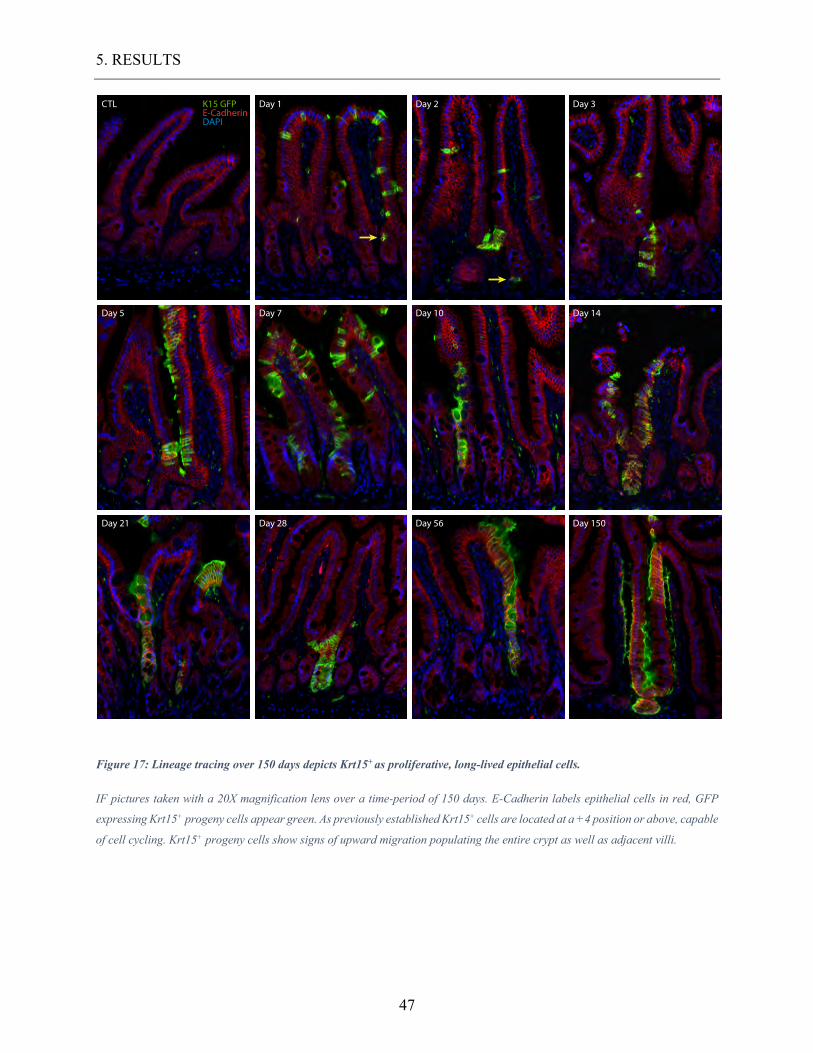
5. RESULTS
47
Figure 17: Lineage tracing over 150 days depicts Krt15+ as proliferative, long-lived epithelial cells.
IF pictures taken with a 20X magnification lens over a time-period of 150 days. E-Cadherin labels epithelial cells in red, GFP
expressing Krt15+ progeny cells appear green. As previously established Krt15+ cells are located at a +4 position or above, capable
of cell cycling. Krt15+ progeny cells show signs of upward migration populating the entire crypt as well as adjacent villi.
Figure for dissertation
CTL Day 1 Day 2 Day 3
Day 5 Day 7 Day 10 Day 14
Day 21 Day 28 Day 56 Day 150
K15 GFPE-CadherinDAPI
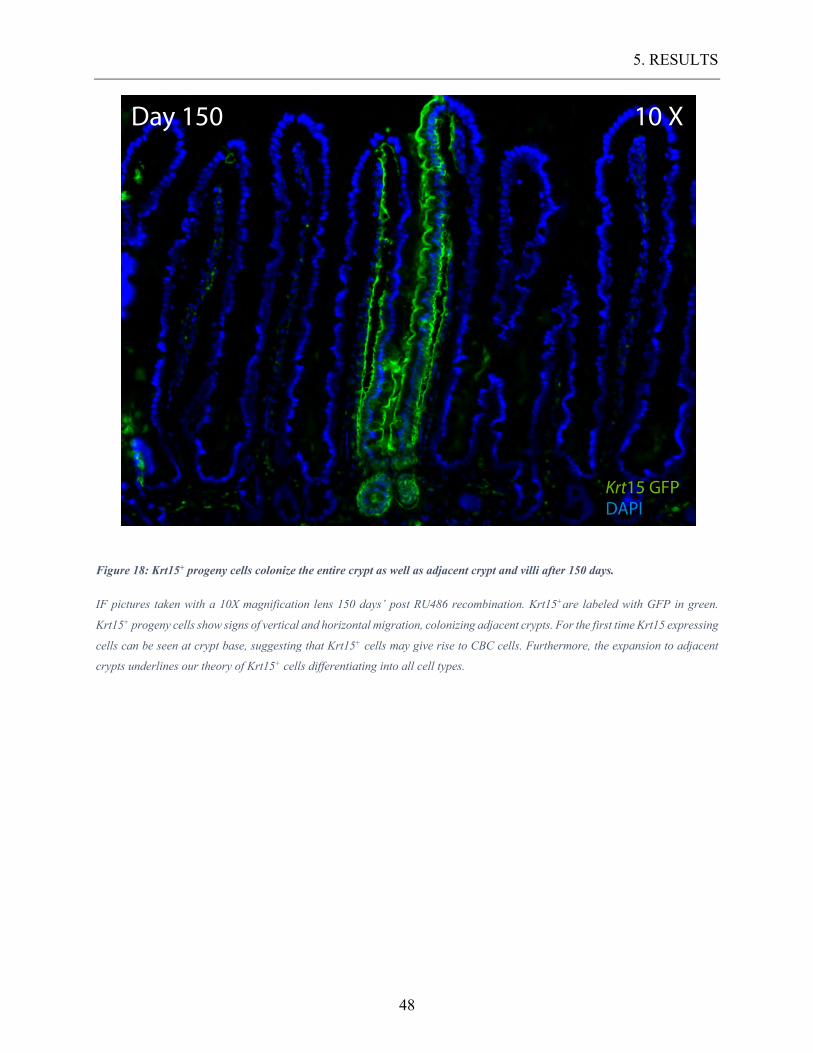
5. RESULTS
48
Figure 18: Krt15+ progeny cells colonize the entire crypt as well as adjacent crypt and villi after 150 days.
IF pictures taken with a 10X magnification lens 150 days’ post RU486 recombination. Krt15+are labeled with GFP in green.
Krt15+ progeny cells show signs of vertical and horizontal migration, colonizing adjacent crypts. For the first time Krt15 expressing
cells can be seen at crypt base, suggesting that Krt15+ cells may give rise to CBC cells. Furthermore, the expansion to adjacent
crypts underlines our theory of Krt15+ cells differentiating into all cell types.
Day 150 10 X
Krt15 GFPDAPI
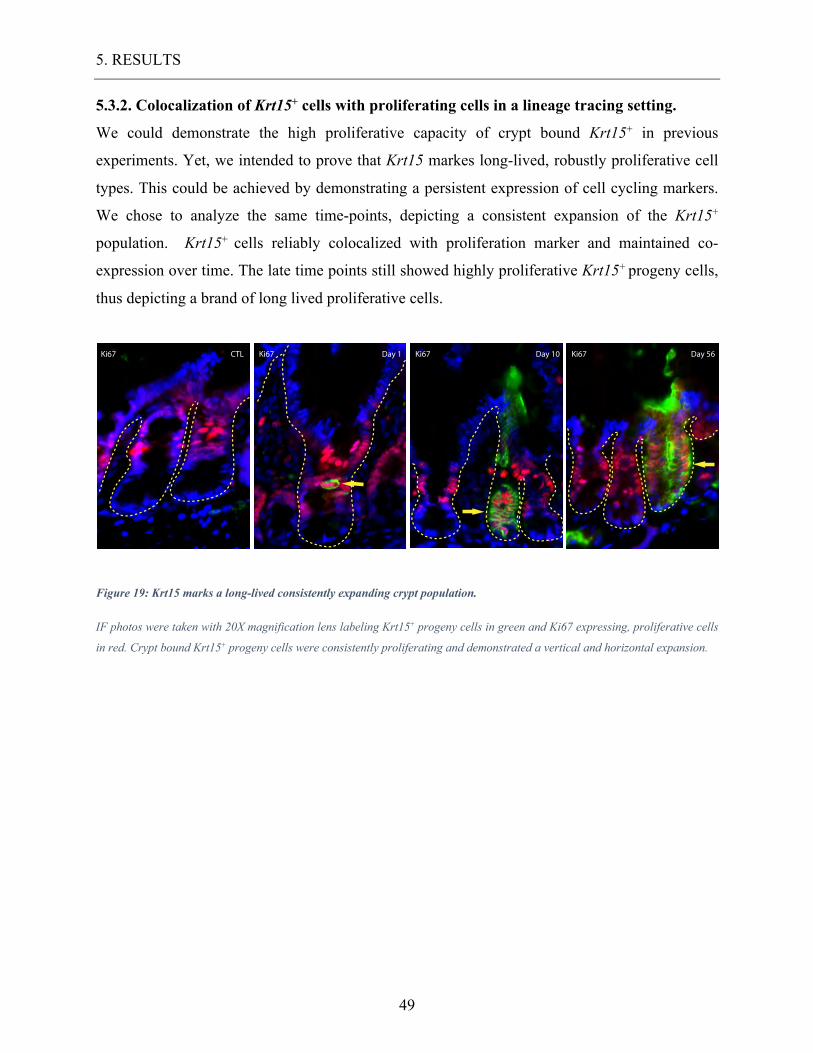
5. RESULTS
49
5.3.2. Colocalization of Krt15+ cells with proliferating cells in a lineage tracing setting.
We could demonstrate the high proliferative capacity of crypt bound Krt15+ in previous
experiments. Yet, we intended to prove that Krt15 markes long-lived, robustly proliferative cell
types. This could be achieved by demonstrating a persistent expression of cell cycling markers.
We chose to analyze the same time-points, depicting a consistent expansion of the Krt15+
population. Krt15+ cells reliably colocalized with proliferation marker and maintained co-
expression over time. The late time points still showed highly proliferative Krt15+ progeny cells,
thus depicting a brand of long lived proliferative cells.
Figure 19: Krt15 marks a long-lived consistently expanding crypt population.
IF photos were taken with 20X magnification lens labeling Krt15+ progeny cells in green and Ki67 expressing, proliferative cells
in red. Crypt bound Krt15+ progeny cells were consistently proliferating and demonstrated a vertical and horizontal expansion.
Ki67 Ki67 Ki67 Ki67CTL Day 1 Day 10 Day 56
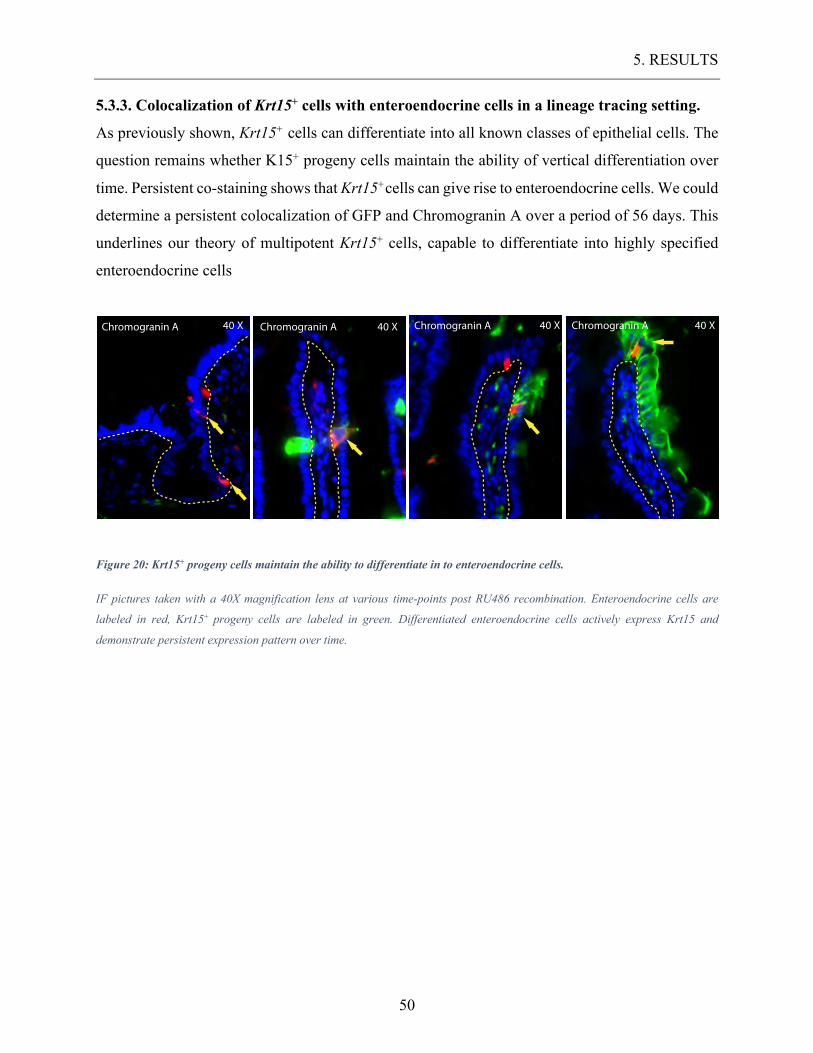
5. RESULTS
50
5.3.3. Colocalization of Krt15+ cells with enteroendocrine cells in a lineage tracing setting.
As previously shown, Krt15+ cells can differentiate into all known classes of epithelial cells. The
question remains whether K15+ progeny cells maintain the ability of vertical differentiation over
time. Persistent co-staining shows that Krt15+cells can give rise to enteroendocrine cells. We could
determine a persistent colocalization of GFP and Chromogranin A over a period of 56 days. This
underlines our theory of multipotent Krt15+ cells, capable to differentiate into highly specified
enteroendocrine cells
Figure 20: Krt15+ progeny cells maintain the ability to differentiate in to enteroendocrine cells.
IF pictures taken with a 40X magnification lens at various time-points post RU486 recombination. Enteroendocrine cells are
labeled in red, Krt15+ progeny cells are labeled in green. Differentiated enteroendocrine cells actively express Krt15 and
demonstrate persistent expression pattern over time.
Chromogranin A 40 X Chromogranin A 40 XChromogranin A 40 XChromogranin A 40 X
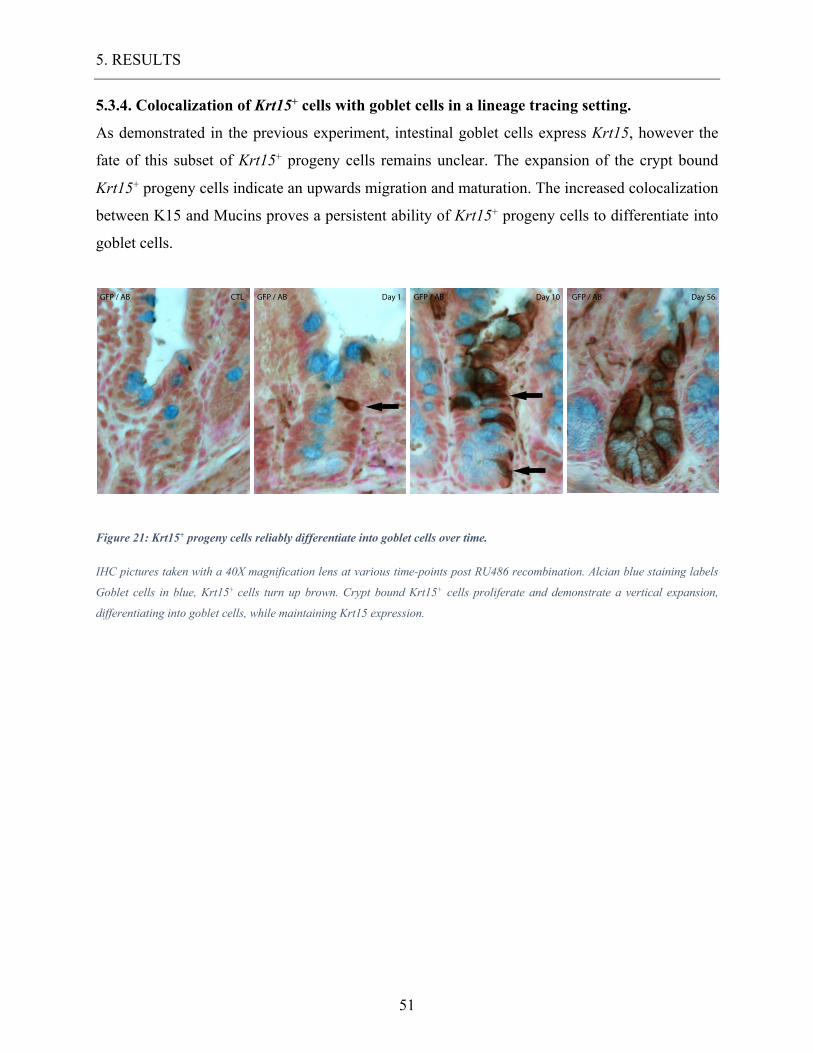
5. RESULTS
51
5.3.4. Colocalization of Krt15+ cells with goblet cells in a lineage tracing setting.
As demonstrated in the previous experiment, intestinal goblet cells express Krt15, however the
fate of this subset of Krt15+ progeny cells remains unclear. The expansion of the crypt bound
Krt15+ progeny cells indicate an upwards migration and maturation. The increased colocalization
between K15 and Mucins proves a persistent ability of Krt15+ progeny cells to differentiate into
goblet cells.
Figure 21: Krt15+ progeny cells reliably differentiate into goblet cells over time.
IHC pictures taken with a 40X magnification lens at various time-points post RU486 recombination. Alcian blue staining labels
Goblet cells in blue, Krt15+ cells turn up brown. Crypt bound Krt15+ cells proliferate and demonstrate a vertical expansion,
differentiating into goblet cells, while maintaining Krt15 expression.
GFP / AB GFP / AB GFP / ABGFP / ABCTL Day 1 Day 10 Day 56
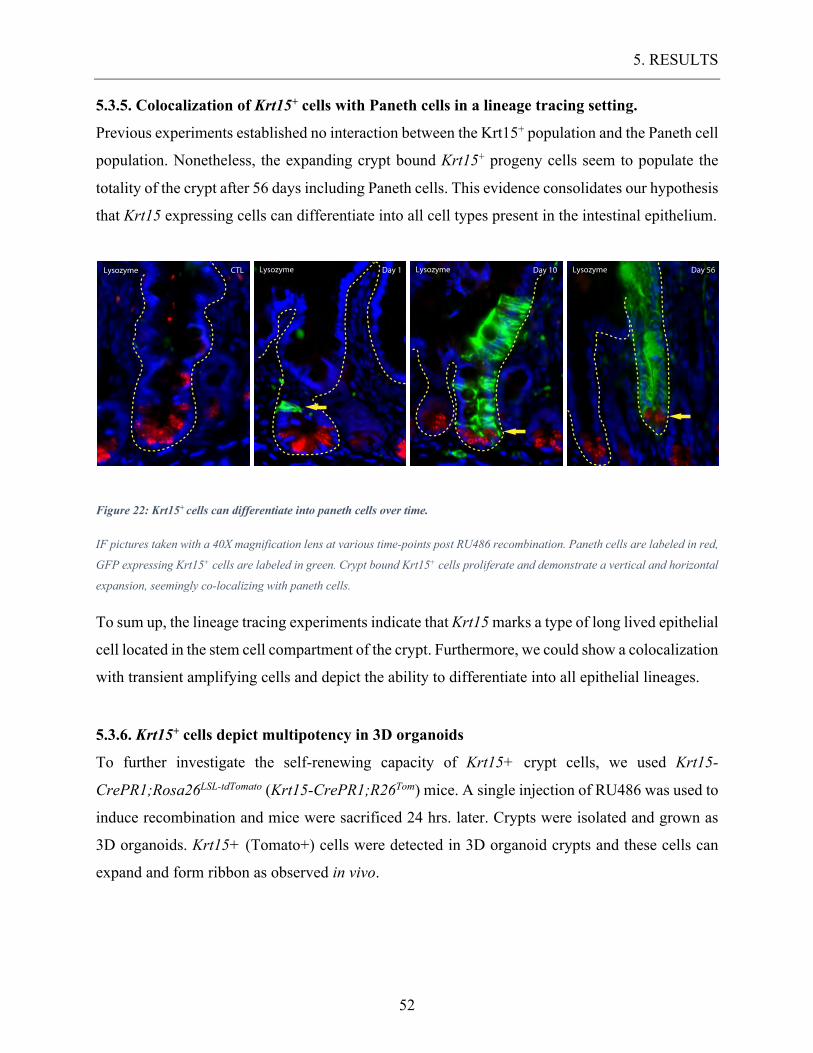
5. RESULTS
52
5.3.5. Colocalization of Krt15+ cells with Paneth cells in a lineage tracing setting.
Previous experiments established no interaction between the Krt15+ population and the Paneth cell
population. Nonetheless, the expanding crypt bound Krt15+ progeny cells seem to populate the
totality of the crypt after 56 days including Paneth cells. This evidence consolidates our hypothesis
that Krt15 expressing cells can differentiate into all cell types present in the intestinal epithelium.
Figure 22: Krt15+ cells can differentiate into paneth cells over time.
IF pictures taken with a 40X magnification lens at various time-points post RU486 recombination. Paneth cells are labeled in red,
GFP expressing Krt15+ cells are labeled in green. Crypt bound Krt15+ cells proliferate and demonstrate a vertical and horizontal
expansion, seemingly co-localizing with paneth cells.
To sum up, the lineage tracing experiments indicate that Krt15 marks a type of long lived epithelial
cell located in the stem cell compartment of the crypt. Furthermore, we could show a colocalization
with transient amplifying cells and depict the ability to differentiate into all epithelial lineages.
5.3.6. Krt15+ cells depict multipotency in 3D organoids
To further investigate the self-renewing capacity of Krt15+ crypt cells, we used Krt15-
CrePR1;Rosa26LSL-tdTomato (Krt15-CrePR1;R26Tom) mice. A single injection of RU486 was used to
induce recombination and mice were sacrificed 24 hrs. later. Crypts were isolated and grown as
3D organoids. Krt15+ (Tomato+) cells were detected in 3D organoid crypts and these cells can
expand and form ribbon as observed in vivo.
Lysozyme Lysozyme Lysozyme LysozymeCTL Day 1 Day 10 Day 56
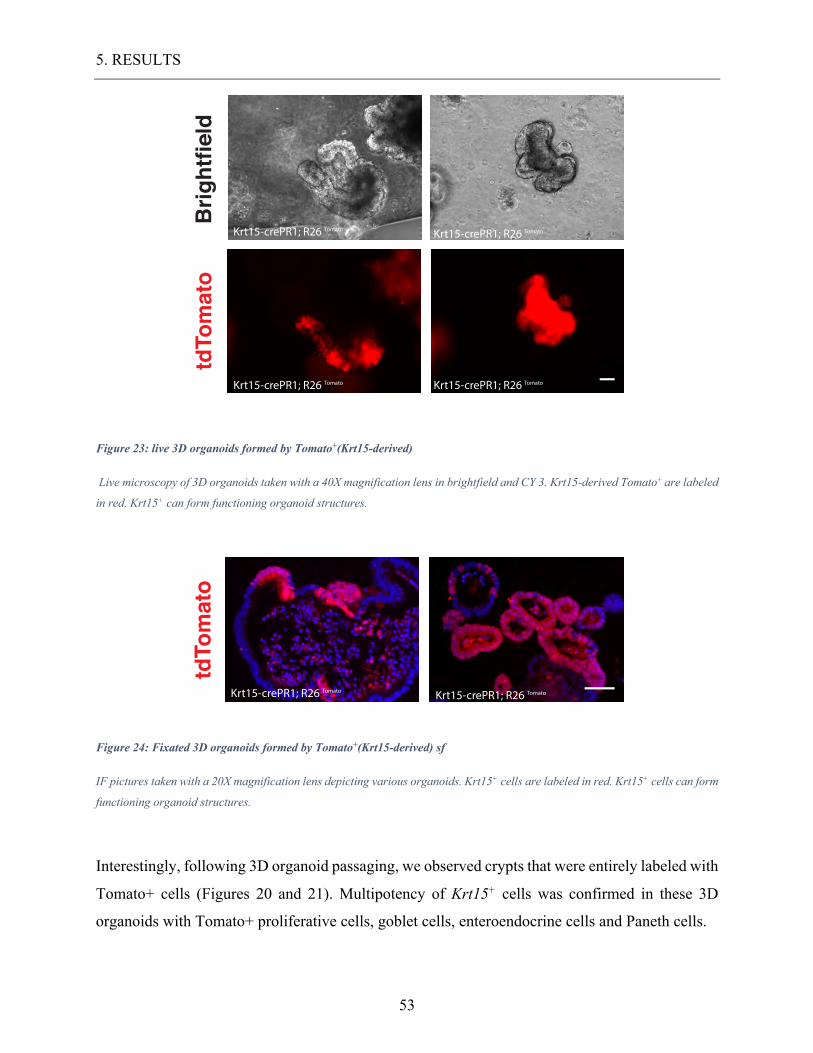
5. RESULTS
53
Figure 23: live 3D organoids formed by Tomato+(Krt15-derived)
Live microscopy of 3D organoids taken with a 40X magnification lens in brightfield and CY 3. Krt15-derived Tomato+ are labeled
in red. Krt15+ can form functioning organoid structures.
Figure 24: Fixated 3D organoids formed by Tomato+(Krt15-derived) sf
IF pictures taken with a 20X magnification lens depicting various organoids. Krt15+ cells are labeled in red. Krt15+ cells can form
functioning organoid structures.
Interestingly, following 3D organoid passaging, we observed crypts that were entirely labeled with
Tomato+ cells (Figures 20 and 21). Multipotency of Krt15+ cells was confirmed in these 3D
organoids with Tomato+ proliferative cells, goblet cells, enteroendocrine cells and Paneth cells.
tdTomato
Brightfield
Krt15-crePR1; R26 TomatoKrt15-crePR1; R26 Tomato
Krt15-crePR1; R26 Tomato Krt15-crePR1; R26 Tomato
tdTomato
Krt15-crePR1; R26 Tomato Krt15-crePR1; R26 Tomato
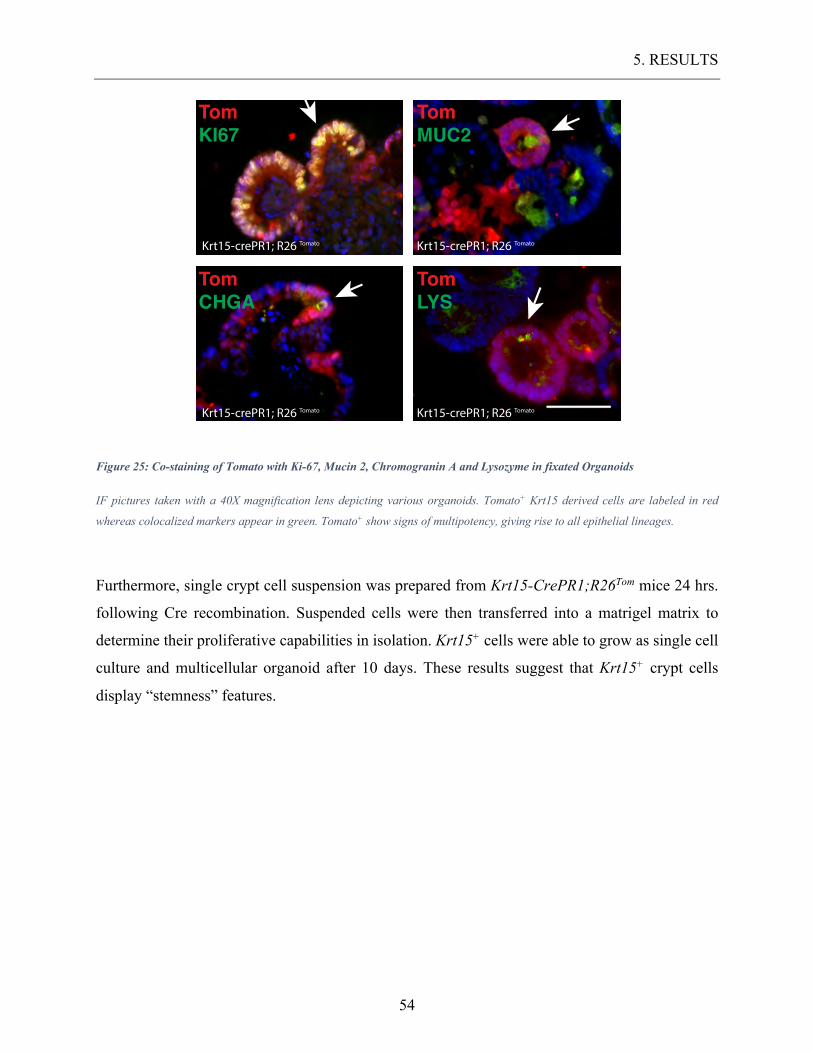
5. RESULTS
54
Figure 25: Co-staining of Tomato with Ki-67, Mucin 2, Chromogranin A and Lysozyme in fixated Organoids
IF pictures taken with a 40X magnification lens depicting various organoids. Tomato+ Krt15 derived cells are labeled in red
whereas colocalized markers appear in green. Tomato+ show signs of multipotency, giving rise to all epithelial lineages.
Furthermore, single crypt cell suspension was prepared from Krt15-CrePR1;R26Tom mice 24 hrs.
following Cre recombination. Suspended cells were then transferred into a matrigel matrix to
determine their proliferative capabilities in isolation. Krt15+ cells were able to grow as single cell
culture and multicellular organoid after 10 days. These results suggest that Krt15+ crypt cells
display “stemness” features.
TomKI67
TomMUC2
TomLYS
TomCHGA
Krt15-crePR1; R26 Tomato Krt15-crePR1; R26 Tomato
Krt15-crePR1; R26 Tomato Krt15-crePR1; R26 Tomato
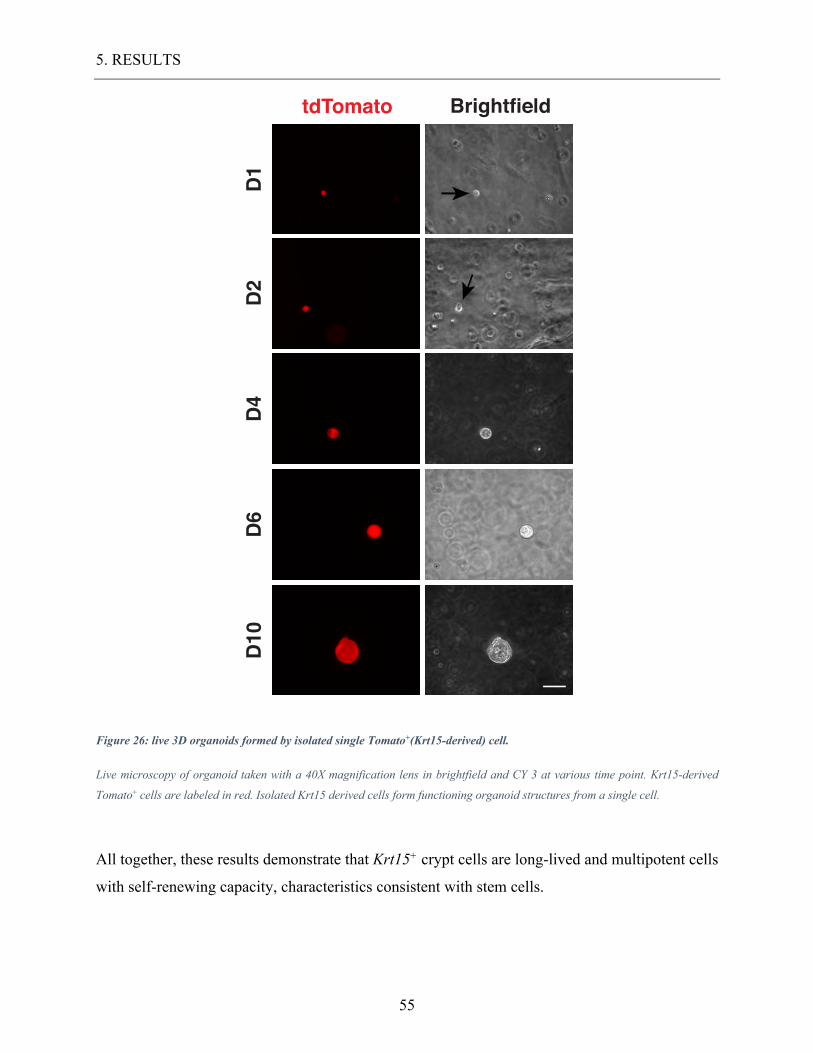
5. RESULTS
55
Figure 26: live 3D organoids formed by isolated single Tomato+(Krt15-derived) cell.
Live microscopy of organoid taken with a 40X magnification lens in brightfield and CY 3 at various time point. Krt15-derived
Tomato+ cells are labeled in red. Isolated Krt15 derived cells form functioning organoid structures from a single cell.
All together, these results demonstrate that Krt15+ crypt cells are long-lived and multipotent cells
with self-renewing capacity, characteristics consistent with stem cells.
D10
D6
D4
D2
D1
tdTomato Brightfield

5. RESULTS
56
5.4. The role of Krt15+ cells in response to radiation injury and their
regeneration capabilities. Our earlier findings demonstrated the proliferative capacities of crypt bound Krt15+ cells, yet it
remains unclear to what extend this influences epithelial homeostasis. Radiation injury is known
to initiate stem cell proliferation as epithelial repair becomes necessary (May et al., 2008; Yan et
al., 2012). The remarkably quick turnover of the intestinal epithelium is driven by multipotent stem
cells, crucial for upholding epithelial homeostasis. Durability and resistance to extracorporeal
noxae is a key feature for stem cells and is predominantly expressed in quiescent +4 positioned
stem cells. Lgr5+ CBC stem cells seem to be radiosensitive to the point of stem cell sterilization
(van Es et al., 2012; Yan et al., 2012), whereas K19+ and Bmi1+ stem cells are radioresistant
(Asfaha et al., 2015a; Yan et al., 2012). Past experiments have shown dormant +4 positioned stem
cells capable of replacing ablated CBC cells after irradiation. We hypothesized K15 would label a
type of radioresistant cell type.
5.4.1. The role of Krt15+ cells in tissue recovery.
We sought to investigate the self-renewal properties of Krt15+ regarding their sensitivity to
radiation. The key aspect was to investigate the intestinal epithelium in the process of
convalescence, following radiation injury. Thus, we treated a total of 12 adult Krt15-crePR1;
R26mT/mG mice with RU486 for 5 consecutive days. The mice were then divided into two groups,
one receiving a 12 Gy whole-body irradiation, the other being the control group. This dose is
sufficient to cause severe crypt destruction and stem cell ablation in the majority of the crypts
(May et al., 2008). Both experimental groups consisted of 6 mice each, which we then sacrificed
after 2 and 5 days post irradiation. As depicted in figure 19, we compared irradiated and non-
irradiated Krt15-crePR1; R26mT/mG mice 2 and 5 days post treatment. It appears that two days post
irradiation, the effect of epithelial cell damage still predominates and only little cell proliferation
can be seen. Nonetheless, self-renewal activity peaks 5 days post irradiation treatment and crypts
fill with rapidly proliferating cells. Injured epithelial cells are rapidly replaced by cells emerging
from crypt stem cell populations. Crypts in the process of recovery appear enlarged and deepened
containing a predominant number of proliferating cells. Experiments suggest that the vivacity of
the epithelial tissue after injury correlates with the crypt structure (Al Alam et al., 2015; Kantara

5. RESULTS
57
et al., 2015). Krt15+ cells prove to be resilient to radiation injury and persist in their crypt location.
The expeditious expansion of Krt15 progeny cells after radiation injury demonstrates the
implication of Krt15 expressing stem cells in the process of recovery. To quantify our findings,
we conducted a count of crypt cells, establishing a ratio of GFP+ cells to all crypt cells. We counted
50 of intact crypts per specimen, focusing on GFP labeled crypts consequently concentrating on
crypts with Krt15+ cells, which constituted only 10% of all crypts. Table 2 depicts the significant
increase of the Krt15+ population 5 days post irradiation underlining their resilience to radiation
damage and their capability to repopulate the crypt. We compared the percentage of Krt15+ crypt
cells 2 and 5 days post irradiation and standardized it to non-irradiated experimental mice. Two
days post treatment the percentage of GFP+ cells in both irradiated and non-irradiated mice
remained around 18%. Five days post treatment, the number of GFP+ cells drastically increased in
the irradiated mice to around 90 % of the crypt population. Moreover, we investigated the
proliferation rate via Ki67 depicted in table 3. We determined the amount of KI67+ cells within
the Krt15+ cell population, by establishing a ratio of GFP+ and Ki67+ co-stained crypt cells versus
all Ki67+ crypt cells. Five days post irradiation an overwhelming majority of all dividing cells co-
stained with GFP, thus supporting our thesis of Krt15+ cells being involved in tissue regeneration.
Furthermore, the comparison of irradiated intestinal tissue to non-irradiated tissue depicted a
disparity of proliferation patterns of Krt15+ cells. In healthy tissue Krt15+ cells appeared to divide
at relatively moderate pace. However, irradiation seemingly eliminating most rivalry stem cells
causes a major influx in Krt15+ propagation.
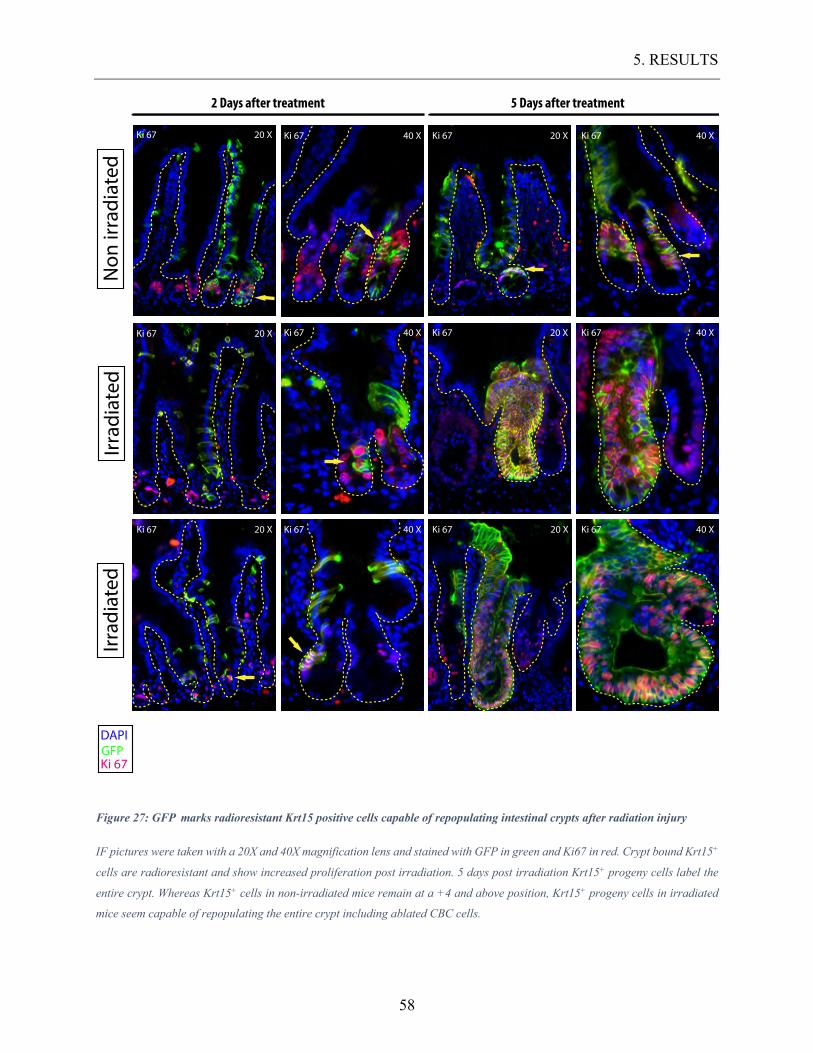
5. RESULTS
58
Figure 27: GFP marks radioresistant Krt15 positive cells capable of repopulating intestinal crypts after radiation injury
IF pictures were taken with a 20X and 40X magnification lens and stained with GFP in green and Ki67 in red. Crypt bound Krt15+
cells are radioresistant and show increased proliferation post irradiation. 5 days post irradiation Krt15+ progeny cells label the
entire crypt. Whereas Krt15+ cells in non-irradiated mice remain at a +4 and above position, Krt15+ progeny cells in irradiated
mice seem capable of repopulating the entire crypt including ablated CBC cells.
Non
irra
diat
ed2 Days after treatment 5 Days after treatment
20 XKi 67 40 XKi 67 20 XKi 67
40 XKi 67 20 XKi 67 40 XKi 67
40 XKi 67
20 XKi 67
Irrad
iate
d
DAPIGFPKi 67
Irrad
iate
d
Ki 67 40 XKi 67 20 X Ki 67 20 X Ki 67 40 X
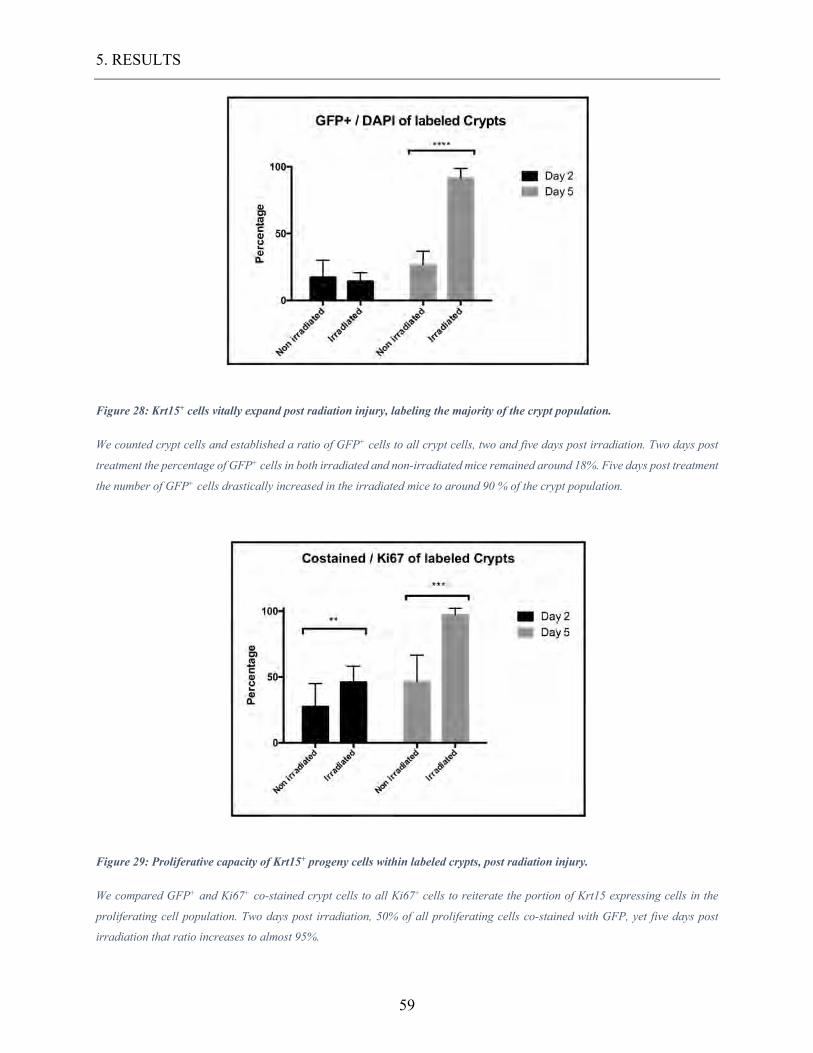
5. RESULTS
59
Figure 28: Krt15+ cells vitally expand post radiation injury, labeling the majority of the crypt population.
We counted crypt cells and established a ratio of GFP+ cells to all crypt cells, two and five days post irradiation. Two days post
treatment the percentage of GFP+ cells in both irradiated and non-irradiated mice remained around 18%. Five days post treatment
the number of GFP+ cells drastically increased in the irradiated mice to around 90 % of the crypt population.
Figure 29: Proliferative capacity of Krt15+ progeny cells within labeled crypts, post radiation injury.
We compared GFP+ and Ki67+ co-stained crypt cells to all Ki67+ cells to reiterate the portion of Krt15 expressing cells in the
proliferating cell population. Two days post irradiation, 50% of all proliferating cells co-stained with GFP, yet five days post
irradiation that ratio increases to almost 95%.

5. RESULTS
60
5.4.2. The importance of Krt15 expressing cells in irradiation models compared to Krt15-/-
In our previous experiments, we established K15 as a marker for radiation resistant stem-cells,
capable of repopulating intestinal crypt after injury. Nonetheless, the question remains whether
Krt15+ stem-cells are required for crypt homeostasis and injury management. To this purpose, we
compared 4 adult Krt15-/- mice to 3 adult Wildtype mice. Prenatal deletion of Krt15 in mice did
not disrupt intestinal epithelial homeostasis and all specimens were phenotypically unobtrusive.
Manifestly, the organisms were capable of replacing Krt15+ expressing cells, thus upholding
intestinal homeostasis. To examine the injury management of Krt15-/- mice, we designed a similar
experiment as described earlier, irradiating the mice and screening for intestinal tissue damage.
Mice were treated with 12 Gy or ionizing radiation at the age of 8 weeks and dissected 5 days post
treatment. Small intestinal crypts were then examined, analyzing 100 intact crypts per specimen.
We analyzed crypt-length of the ileum and jejunum, as a parameter of tissue rehabilitation in Krt15-
/- mice and Wildtype. Histometric analysis of the crypt length showed significant differences in
crypt length of treated Krt15-/- and Wildtype mice, as demonstrated in figure 20. We compared the
average crypt length of all measurements and ran a T-test, detecting a highly significant length
difference. The considerably shortened crypt and dysmorphic villi of the Krt15-/- mice indicated
an inferior regenerative capability. The loss of radiation resistant stem-cells caused delayed injury
recovery.
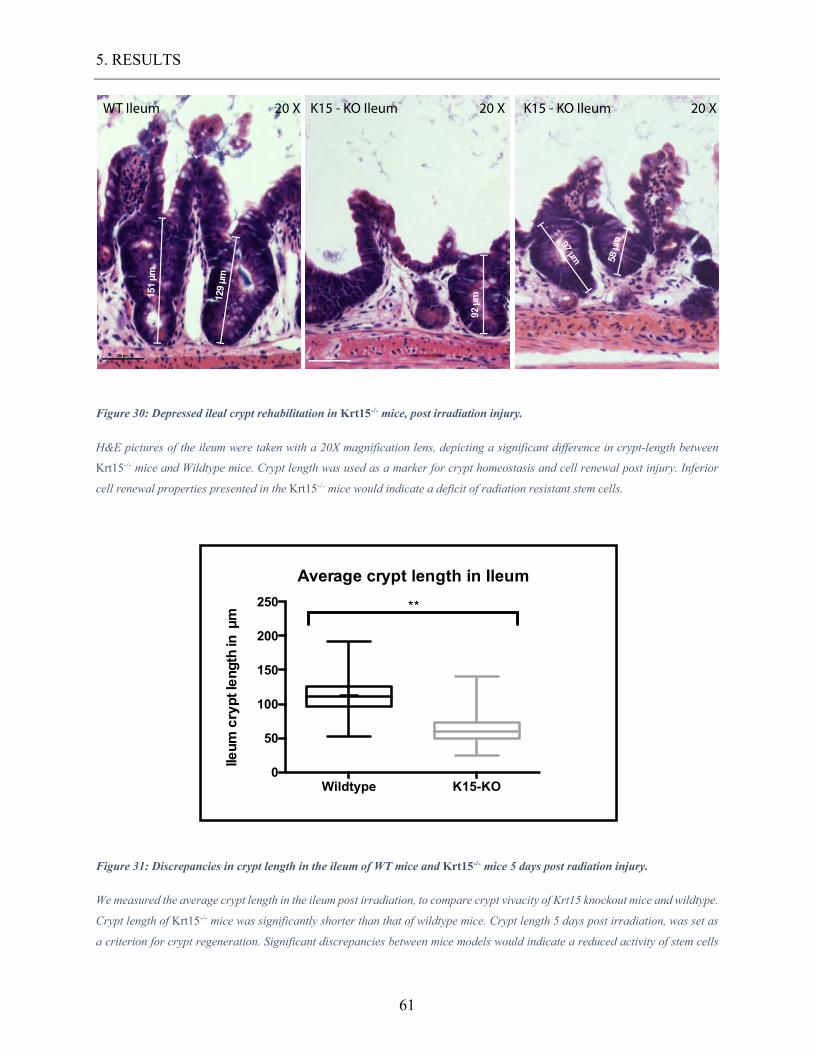
5. RESULTS
61
Figure 30: Depressed ileal crypt rehabilitation in Krt15-/- mice, post irradiation injury.
H&E pictures of the ileum were taken with a 20X magnification lens, depicting a significant difference in crypt-length between
Krt15-/- mice and Wildtype mice. Crypt length was used as a marker for crypt homeostasis and cell renewal post injury. Inferior
cell renewal properties presented in the Krt15-/- mice would indicate a deficit of radiation resistant stem cells.
Figure 31: Discrepancies in crypt length in the ileum of WT mice and Krt15-/- mice 5 days post radiation injury.
We measured the average crypt length in the ileum post irradiation, to compare crypt vivacity of Krt15 knockout mice and wildtype.
Crypt length of Krt15-/- mice was significantly shorter than that of wildtype mice. Crypt length 5 days post irradiation, was set as
a criterion for crypt regeneration. Significant discrepancies between mice models would indicate a reduced activity of stem cells
K15 - KO Ileum 20 XK15 - KO Ileum 20 XWT Ileum 20 X
151
µm
92 µ
m
97 µm 58 µ
m
129
µm
Wildtype K15-KO0
50
100
150
200
250
Ileum
cry
pt le
ngth
in µ
m
Average crypt length in Ileum
**
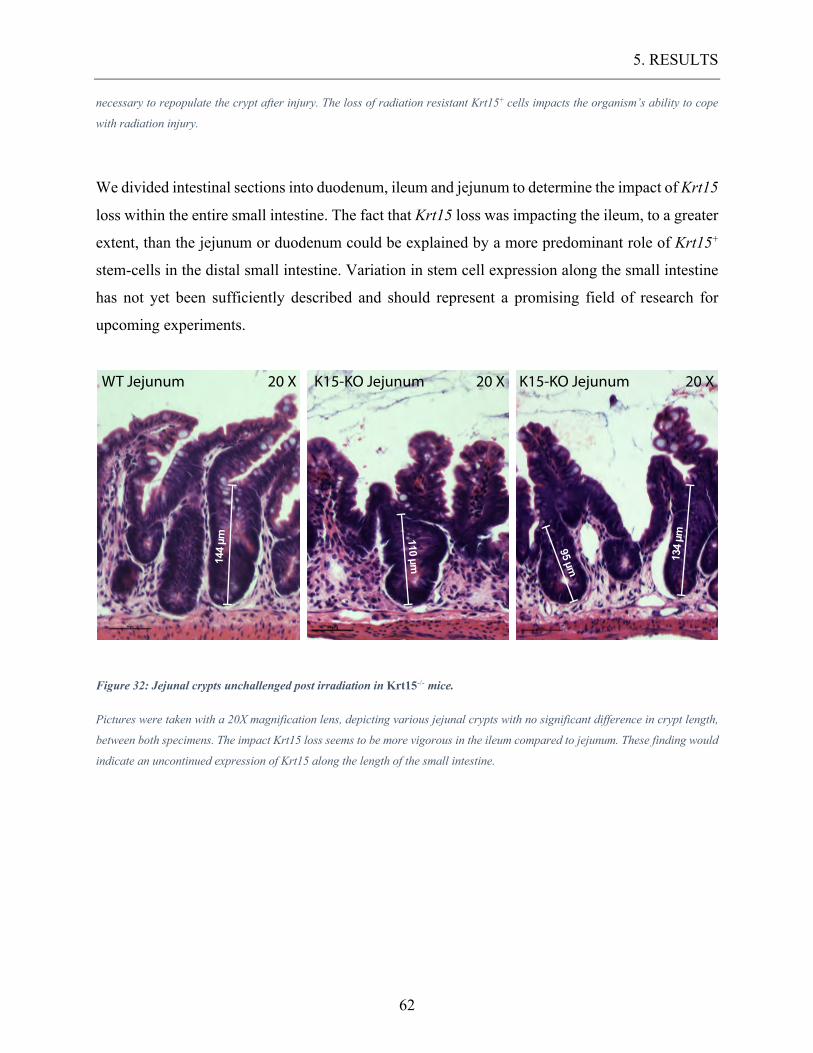
5. RESULTS
62
necessary to repopulate the crypt after injury. The loss of radiation resistant Krt15+ cells impacts the organism’s ability to cope
with radiation injury.
We divided intestinal sections into duodenum, ileum and jejunum to determine the impact of Krt15
loss within the entire small intestine. The fact that Krt15 loss was impacting the ileum, to a greater
extent, than the jejunum or duodenum could be explained by a more predominant role of Krt15+
stem-cells in the distal small intestine. Variation in stem cell expression along the small intestine
has not yet been sufficiently described and should represent a promising field of research for
upcoming experiments.
Figure 32: Jejunal crypts unchallenged post irradiation in Krt15-/- mice.
Pictures were taken with a 20X magnification lens, depicting various jejunal crypts with no significant difference in crypt length,
between both specimens. The impact Krt15 loss seems to be more vigorous in the ileum compared to jejunum. These finding would
indicate an uncontinued expression of Krt15 along the length of the small intestine.
WT Jejunum 20 X K15-KO Jejunum 20 XK15-KO Jejunum 20 X
144
µm 110 µm
95 µm 134
µm

5. RESULTS
63
Figure 33: Discrepancies in crypt length between WT mice and Krt15-/- mice 5 days post radiation injury.
We measured the average crypt length in the jejunum post irradiation, to compare crypt vivacity of Krt15 knockout mice and
wildtype. There was no significant difference in crypt length between wildtype mice and Krt15-/- mice. These findings would indicate
a subordinated role of Krt15 in the jejunum compared to the ileum.
WT K15-KO0
50
100
150
200
250
Cry
pt le
ngth
in µ
m
Average crypt length in the Jejenum
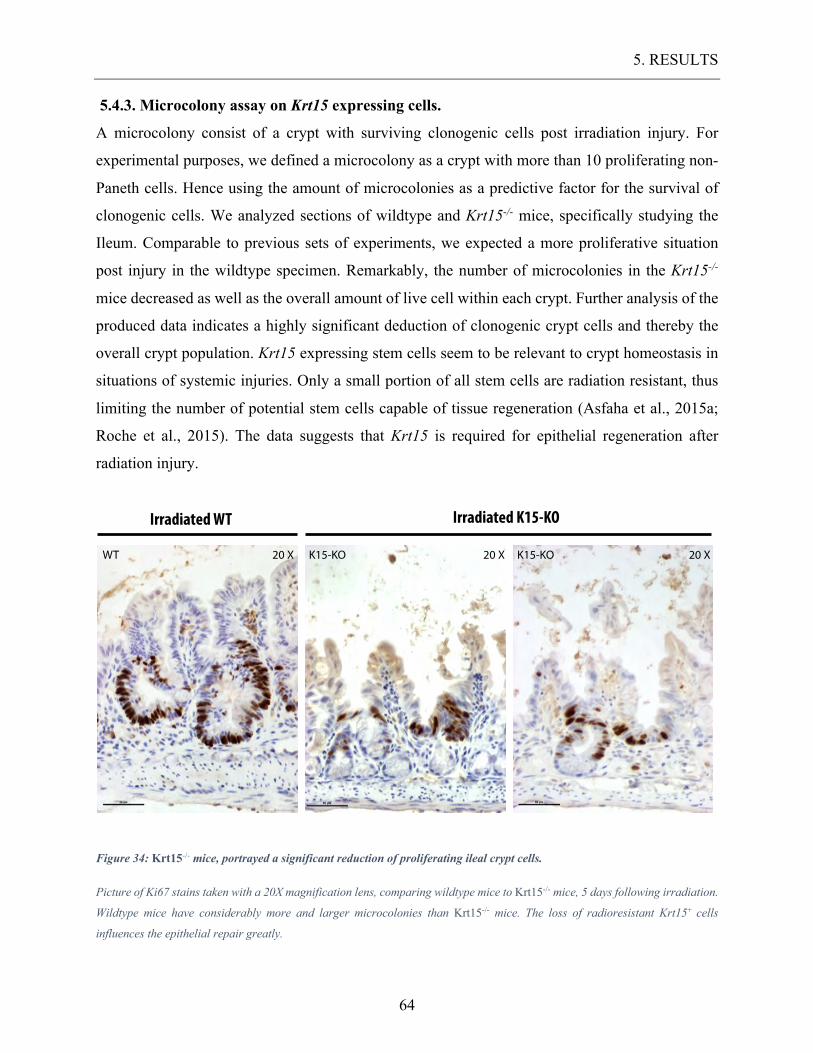
5. RESULTS
64
5.4.3. Microcolony assay on Krt15 expressing cells.
A microcolony consist of a crypt with surviving clonogenic cells post irradiation injury. For
experimental purposes, we defined a microcolony as a crypt with more than 10 proliferating non-
Paneth cells. Hence using the amount of microcolonies as a predictive factor for the survival of
clonogenic cells. We analyzed sections of wildtype and Krt15-/- mice, specifically studying the
Ileum. Comparable to previous sets of experiments, we expected a more proliferative situation
post injury in the wildtype specimen. Remarkably, the number of microcolonies in the Krt15-/-
mice decreased as well as the overall amount of live cell within each crypt. Further analysis of the
produced data indicates a highly significant deduction of clonogenic crypt cells and thereby the
overall crypt population. Krt15 expressing stem cells seem to be relevant to crypt homeostasis in
situations of systemic injuries. Only a small portion of all stem cells are radiation resistant, thus
limiting the number of potential stem cells capable of tissue regeneration (Asfaha et al., 2015a;
Roche et al., 2015). The data suggests that Krt15 is required for epithelial regeneration after
radiation injury.
Figure 34: Krt15-/- mice, portrayed a significant reduction of proliferating ileal crypt cells.
Picture of Ki67 stains taken with a 20X magnification lens, comparing wildtype mice to Krt15-/- mice, 5 days following irradiation.
Wildtype mice have considerably more and larger microcolonies than Krt15-/- mice. The loss of radioresistant Krt15+ cells
influences the epithelial repair greatly.
WT 20 X K15-KO 20 X K15-KO 20 X
Irradiated WT Irradiated K15-KO
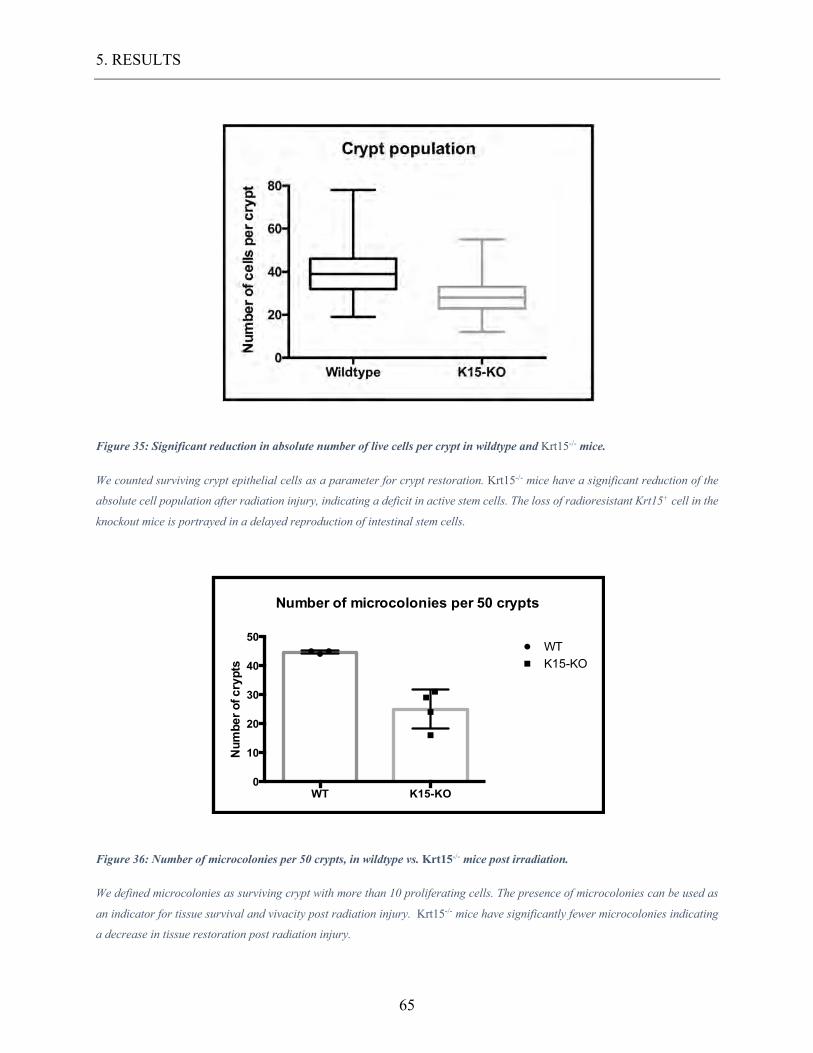
5. RESULTS
65
Figure 35: Significant reduction in absolute number of live cells per crypt in wildtype and Krt15-/- mice.
We counted surviving crypt epithelial cells as a parameter for crypt restoration. Krt15-/- mice have a significant reduction of the
absolute cell population after radiation injury, indicating a deficit in active stem cells. The loss of radioresistant Krt15+ cell in the
knockout mice is portrayed in a delayed reproduction of intestinal stem cells.
Figure 36: Number of microcolonies per 50 crypts, in wildtype vs. Krt15-/- mice post irradiation.
We defined microcolonies as surviving crypt with more than 10 proliferating cells. The presence of microcolonies can be used as
an indicator for tissue survival and vivacity post radiation injury. Krt15-/- mice have significantly fewer microcolonies indicating
a decrease in tissue restoration post radiation injury.
WT K15-KO0
10
20
30
40
50
Num
ber o
f cry
pts
WTK15-KO
Number of microcolonies per 50 crypts

5. RESULTS
66
5.5. Role of Krt15 in cancer initiation. Tumor-initiating capacity is an important feature in intestinal stem cells. Lgr5+ cells, Lrig1+ cells
and Krt19+ cells have been described as the cell-of-origin for intestinal tumorigenesis following
Apc loss (Asfaha et al., 2015a; Barker et al., 2009; Powell et al., 2012). Also, Bmi1+ cells can form
adenomas following induction of a stable form of b-catenin in these cells (Sangiorgi and Capecchi,
2008b). In order to determine if Krt15+ cells can initiate tumor formation in the intestine, we bred
Apcfl/fl mice with our Krt15-CrePR1;R26mT/mG mice. Cre recombination was induced using daily
injections of 0.5mg RU486 for five consecutive days. The control mice consisted of two WT mice
and one Krt15-CrePR1;R26mT/mG who did not receive any RU486 injections. All mice (n=11 mice)
were sacrificed 6 months after Cre recombination or at prior time points in concert with severe
weight loss.
Figure 37: Timetable depicting experimental setup for the cancer initiation model
Krt15-CrePR1;Apcfl/fl;R26mT/mG mice were injected daily with 0.5mg RU486 for five consecutive days to induce Cre recombination
and mice were sacrificed when sick or 150 days following Cre induction.
Gross lesions were observed in all Krt15-CrePR1;Apcfl/fl;R26mT/mG mice but not in control mice.
All mice developed lesions in the small intestine, while colon lesions were observed in 37 % of
the mice. GFP labeling confirmed that the observed lesions were derived from Krt15+ cells (Figure
28).
0-1-2-3-4 150Time (d)
RU486
Krt15-CrePR1;Apcfl/fl;R26mT/mG
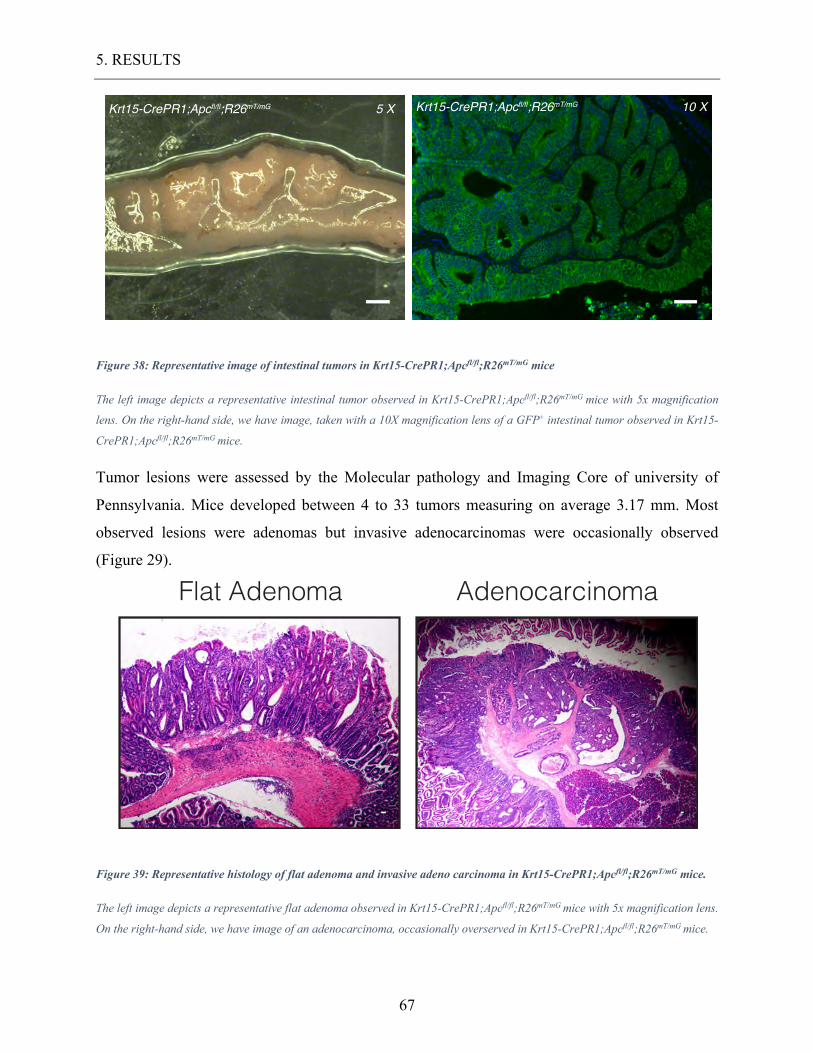
5. RESULTS
67
Figure 38: Representative image of intestinal tumors in Krt15-CrePR1;Apcfl/fl;R26mT/mG mice
The left image depicts a representative intestinal tumor observed in Krt15-CrePR1;Apcfl/fl;R26mT/mG mice with 5x magnification
lens. On the right-hand side, we have image, taken with a 10X magnification lens of a GFP+ intestinal tumor observed in Krt15-
CrePR1;Apcfl/fl;R26mT/mG mice.
Tumor lesions were assessed by the Molecular pathology and Imaging Core of university of
Pennsylvania. Mice developed between 4 to 33 tumors measuring on average 3.17 mm. Most
observed lesions were adenomas but invasive adenocarcinomas were occasionally observed
(Figure 29).
Figure 39: Representative histology of flat adenoma and invasive adeno carcinoma in Krt15-CrePR1;Apcfl/fl;R26mT/mG mice.
The left image depicts a representative flat adenoma observed in Krt15-CrePR1;Apcfl/fl;R26mT/mG mice with 5x magnification lens.
On the right-hand side, we have image of an adenocarcinoma, occasionally overserved in Krt15-CrePR1;Apcfl/fl;R26mT/mG mice.
Krt15-CrePR1;Apcfl/fl;R26mT/mGKrt15-CrePR1;Apcfl/fl;R26mT/mG 5 X 10 X
Flat Adenoma Adenocarcinoma

5. RESULTS
68
These results suggest that Krt15+ crypt cells are tumor-initiating and lesions derived from Krt15+
cells can even progress to invasive adenocarcinoma, which would appear to distinguish such cells
from other ISCs.

6. DISCUSSION
69
6. DISCUSSION Krt15 has been described as a putative epidermal stem cell marker and is thought to play a pivotal
role in cell homeostasis and carcinogenesis of the cutaneous epithelium. Krt15+ hair bulge cells
are slow cycling long-lasting stem cells, preferentially proliferating in combined expression with
Krt19. Recent publication of Krt19 expression in the intestinal epithelium, lead to the theory that
both keratins could be used as stem cell markers in the intestine. Our first approach was to prove
Krt15 expression in the intestine by directly targeting Krt15+ cells. We examined the small
intestine of WT-mice with K15 antibodies using Krt15-/- as a negative control. Although our
control mice depicted an absolute Krt15 knockout in the esophagus, we encountered occasional
false positive staining at crypt ground. These false positives could be explained by an interaction
of K15 antibodies with unknown elements of the Paneth cell population. To confirm these
observations, we designed a transgenic mouse model Krt15-crePR1; R26mT/mG, enabling us to label
Krt15 expressing cells directly. By repeated Cre induction via RU486, we established a high
recombination rate, allowing us to trace the Krt15+ population. This unique mouse model permitted
us to determine the fate of any recombined Krt15+ cell and all daughter cells. In a series of
experiments on Krt15-crePR1; R26mT/mG mice, we established a robust expression of Krt15 in the
small intestine at various locations. The intestinal epithelium can be divided functionally into two
compartments: the crypt, containing long-lived stem cells and mainly undifferentiated cells and
the villi consisting of highly differentiated short-lived cells. We identified Krt15+ cells in the crypt
mainly between the second and fifth position, which is consistent with the location of most ISCs.
Nevertheless, a series of differentiated cells expressed Krt15 in the villi which was unexpected
since Krt15 expression in the epidermal epithelium is mostly restricted to undifferentiated basal
epithelial cells. Vertical differentiation involves the process of progressive terminal differentiation
of basal cells in the epidermal epithelium as well as transient amplifying cell in the intestine.
Intestinal daughter cells undergo a vertical migration and simultaneous differentiation.
Collaborating groups proposed Krt15 regulation by two main putative mechanisms in
keratinocytes: one that drives its expression in the basal layer, mediated primarily by FOXM1, and
another that induces its expression in the suprabasal layers, involving PKC/AP-1 signaling (Bose
et al., 2013). Diverse Krt15 regulation would explain the presence in terminally differentiated
epithelial cells. Another possible explanation could be the age at which we sacrificed the mice (6-
week-old). Suprabasal Krt15 expression has been reported in squamous epithelium of the

6. DISCUSSION
70
esophagus of neonatal mice and sheep, adult specimens however, presented Krt15 expression
confined to the basal layer (Porter et al., 2000; Waseem et al., 1999). Consistent with that theory,
intestinal probes taken from adult mice in the lineage tracing experiment did not show any sign of
isolated Krt15+ cells in the villi. Therefore, suggesting an alternate role of Krt15 in the adult
epithelia compatible with selective expression pattern. As yet, the exact regulation of Krt15 in
adult epithelia is not understood. Although 6-week-old mice seemingly expressed Krt15 both the
villi and the crypt, proliferative Krt15+ cells were restricted to the crypt. Krt15+ cells could reliably
be detected between second fifth crypt position, known to form the stem cell compartment.
Consequently, we decided to trace our Krt15+ population to determine their fate and differentiation
capability. The mouse model with the inducible Cre-LoxP allowed clear recognition of Krt15+
progeny cells. Robust Cre-recombination was achieved by consecutive Ru486 administration for
5 days on 49 mice, sacrificing them at various time points up to 150 days. After recombination,
only Krt15+ progeny cells would express GFP, thus labeling all daughter cells. We quantified
Krt15+ crypt analysis, selecting 4 mice per time point (1, 10 and 56 days) and counting crypt cells
of 150 intact crypts per specimen. The number of 150 analyzed crypts per time point mitigated the
risk of statistical variation. Less than 20% of all crypts contain Krt15+ cells after 56 days, yet they
constitute the majority of cells in those crypts. ISC markers are known to label only a small portion
of all intestinal crypts, suggesting distinct gene variation of ISCs with otherwise similar cell
functions. A stem cell marker reliably labeling all CBC or +4 positioned stem cells has yet to be
found.
Continued progression of the Krt15+ population marks a long-lasting type of stem cell. Although
Krt15+ cells appear to be slow cycling, they proliferate to the extent of labeling entire crypts and
villi. We could also establish that Krt15+ progeny cells differentiate into secretory and absorptive
cell lines. To prove multipotency in Krt15+ cells we designed an experimental approach
transferring in vivo gained crypt into in vitro 3D organoid. We extracted intestinal crypts from
Krt15-CrePR1;Rosa26LSL-tdTomato mice and transferred set crypts into a matrigel matrix. Krt15+
crypt bound cells proved capable of creating organoids as well as giving rise to all cell lineages in
an in vitro setting. Moreover, the single cell suspension demonstrates that isolated Krt15+ cells can
form new organoids, without the support of a stem cell niche.
Regrettably, we could not elaborate any interaction between Krt15+ cells and other known stem
cell, since there exist no adequate antibodies reliably targeting Lgr5+ CBC stem cells (Yamazaki

6. DISCUSSION
71
et al., 2015). Nonetheless, it would be most relevant to cross our Krt15-crePR1; R26mT/mG mouse
with an Lgr5 reporter mice, to demonstrate the implication of Krt15 in CBC cell homeostasis.
Krt15+ cells were shown to colocalize with Lgr5+ cells found in the mouse epidermal bulge of hair
follicle which acted as stem cells regenerating new hair follicles and maintaining all cell lineages
of the follicle (Jaks et al., 2008). Yet another approach could be to cell sort Krt15+ and run a qPCR
to establish mRNA expression level of known stem cell markers. These experimental approaches
remain however future projects.
Similar markers such as Krt19 label a radioresistant slow cycling stem cell, capable of crypt
regeneration after irradiation. The position of Krt15+ cells and their slow cycling quiescent nature
puts forth the theory that they could have similar properties. As experimental approach for this
aim, we chose an injury model irradiating lineage tracing mice at 12 Gy. 12 Gy irradiation was
previously established as sufficiently strong, for complete ablation of radiosensitive Lgr5+ CBC
stem cells (Asfaha et al., 2015a; van Es et al., 2012). Irradiation damage and the loss of highly
proliferate Lgr5+ stem cells severely impact epithelial homeostasis initiating intestinal stem cell
division during epithelial repair (May et al., 2008; Yan et al., 2012). Hence, it appears to be the
ideal setting to examine the influence of Krt15+ on epithelial homeostasis and injury response. We
addressed the question at hand with a cohort of 12 Krt15-crePR1; R26mT/mG mice, irradiating half
and using the other half as non-irradiated control group. 3 specimens out of each cohort were then
sacrificed after 2 and 5 days, ensuring an adequate number of treated animals for each time point.
Irradiation effect was then quantified by crypt analysis via cell counting and crypt length analysis.
Results of crypt analysis were highly significant and depicted a radioresistant Krt15+ cell
population with highly increased cell division. The first time-point (2 days post irradiation)
illustrates a severally damaged epithelial structure, with limited proliferation of Krt15+ cells. After
5 days however, cell division of Krt15+ cells is increased to the point of labeling entire crypts and
adjacent villi. Such rapid cell proliferation admirably exhibits the important role of Krt15+ cells
on epithelial homeostasis after irradiation injury. Comparable epidermal injury models show an
initial upregulation of Krt15 in the basal layer expanding up to suprabasal layers of the follicular
epidermis, followed by Krt15 down-regulation and re-establishing basal specific expression (Troy
et al., 2011). Presumably, Krt15+ cells are capable of restoring crypt-integrity by replacing ablated
radiosensitive stem cells. Krt15+ cells were shown to colocalize with Lgr5+ cells found in the
mouse epidermal bulge of hair follicle, which acted as stem cells regenerating new hair follicles

6. DISCUSSION
72
and maintaining all cell lineages of the follicle (Jaks et al., 2008). To prove this assumption, we
need to confirm interaction between Krt15+ cells and CBC stem cells, before and after irradiation.
Thus, demonstrating the ability of Krt15+ cells to give rise to new CBC stem cells after crypt
sterilization via irradiation. Since Lgr5+ CBC cells cannot be visualized by normal
immunohistochemistry due to a lack of adequate antibodies, this could be realized by insito
Hybridization targeting Lgr5 expression or other known CBC stem cell markers. Yet, this remains
an experimental approach for future projects.
To verify the impact of Krt15 on epithelial homeostasis, we designed an irradiation assay on Krt15-
/- mice comparing them to WT mice. Early systemic loss of Krt15 can commonly be compensated
by other keratins and result in significant disruption of epidermal epithelial function. The impact
on the intestinal epithelium however, has not yet been identified. Epithelial conditions were
quantified 5 days post irradiation, comparing Krt15-/- mice to WT mice. Normal uninjured
intestinal epithelium was not disrupted by loss of Krt15, neither was the crypt nor villi structure
altered nor morphologically aberrant. However, epithelial repair after irradiation injury appeared
segmentally impaired by Krt15 loss. Similarly, epithelial repair in the ileum was significantly
impaired by Krt15 deletion, as measured by microcolony assay and crypt-length analysis.
Microcolony assay is a commonly used method to quantify epithelial repair after irradiation. It was
first established in the 1970s and has been used as a widely accepted experimental tool ever since.
To minimize detection-bias, we analyzed 150 intact ileal crypts per specimen repeating the same
procedure for jejunal crypts. Furthermore, we evaluated crypt length, as a parameter of successful
epithelial repair, using the same n as in the previous approach. Both approaches produced similar
significant results suggesting predominant expression of Krt15 in the ileum compared to the
jejunum. Presumably the impact of Krt15 on epithelial homeostasis could variate along the length
of the small intestine. Graduate expression of Krt15 along the length of the small intestine has not
yet been reported, however considering the stretch of the intestinal epithelium, localized
expression of any marker could be considered as biological diversity. qPCR analysis of epithelial
probes, taken at various sections of the intestine, could directly target this question and display
differentiated expression patterns. In any case, the reduced epithelial repair due to Krt15 knockout
supports our theory of Krt15 playing a crucial role for epithelial homeostasis.
Although substantial research has been done to identify intestinal stem cells, the reliability of stem
cell markers in clinical application remains debatable. A possible clinical application in the field

6. DISCUSSION
73
of stem cell research focuses on defining cancer initiating stem cells. Tumor growth is mainly
independent from external growth factors, either producing growth factors autonomously or by
inadequate overexpression of hormone receptors. Intestinal as well as epidermal stem cells are
predominantly regulated by Wnt signaling. The presence of Wnt signaling is one of the driving
forces of CBC cell proliferation (Haegebarth and Clevers, 2009). Loss of the tumor suppressor
gene APC results in adenomatous transformation of the epithelium, driven by Wnt-signaling and
is considered to be the principal cause of colon cancer in humans (Korinek et al., 1997). The role
of Wnt-signaling further underlines differences between +4 and CBC stem cells. The Bmi1+ stem
cell population was found inherently insensitive to global gain- and loss-of-function modulation
compared to Lgr5+ CBC stem cells (Yan et al., 2012). The cancer stem cell theory assigns cancer
specific attributes, such as inadequate growth and pluripotency to mutated stem cell forming the
origin of cancer genesis. Many studies were conducted to identify the cell-of-origin of intestinal
cancers. Apc loss in Lgr5+ cells (Asfaha et al., 2015a; Barker et al., 2009), Lrig1+ cells (Powell et
al., 2012, 2014) and Krt19+ cells(Asfaha et al., 2015b) lead to adenoma formation. Adenomas are
also observed following induction of stable β-catenin in Bmi1+ cells (Sangiorgi and Capecchi,
2008a). Yet none of these lesions have been reported to progress to adenocarcinoma. Herein, we
observed that mice harboring Apc loss in Krt15+ cells develop adenomas as well as occasionally
invasive adenocarcinomas. These results suggest that Krt15+ cells are tumor-initiating cells that
also have the capacities to progress to more invasive phenotypes. Krt15-CrePR1;Apcfl/fl mice
might also represent a new mouse model for intestinal cancer.
It would be interesting to analyze cell growth in the context of hormone signaling such as Wnt
signaling. One of the most elegant ways to determine Wnt signaling sensibility is by creating 3D
entero-organoids. Our next experimental step would consist of sorting Krt15+ cells from Krt15-
crePR1; R26mT/mG mice via FACS and transferring them to a matrigel in vitro structure, thus
establishing an elaborate entero-genesis assay. The in vitro assay would enable us to determine the
potency of isolated Krt15+ cells while depicting relevant implications of known growth signaling
pathways such as Wnt-signaling and Notch-signaling. By variating external growth factors or
niche signaling supplemented by Paneth cells, we could determine dependency of Krt15+ cells on
external parameters. We assume that Krt15+ cells compete with different stem cells and may play
a subordinate role in normal crypt homeostasis. The role of Krt15+ cells could rather be explained
by that of quiescent resilient stem cells vital for restoring homeostasis after injury. It is certainly

6. DISCUSSION
74
possible that Krt15 may only mark a subset of quiescent stem cells and our results do not exclude
overlapping expression with populations identified by other intestinal stem cell markers.

7. CONCLUSION
75
7. CONCLUSION Undifferentiated epidermal cells selectively express Krt15 as dominating intermediate filaments
subsequently evoking a link between the level of cell differentiation and cytoskeletal components
(Bose et al., 2013). As K15 is one of the basal-specific cytoskeletal proteins, its presence in
intestinal crypts may indicate the necessity of structural integrity for developing cells. In contrast
to the squamous epithelium, the involvement of cytoskeletal proteins in proliferation and
homeostasis in the intestine has scarcely been identified. We proved the presence of K15 in small
intestine and demonstrated a robust expression of Krt15 in intestinal crypts and villi. Furthermore,
we showed that Krt15, a known stem cell and differentiation marker of the epidermal epithelium,
is expressed in long-lived, radioresistant, proliferating epithelial cells in the small intestine. Krt15
expressing+ cells survive well beyond 150 days and show lineage tracing capacity even after
irradiation. We could also prove multipotency in 3D organotypic organoids. In which Krt15+ cells
could recreate crypt like formation in vitro even from a single isolated Krt15+ cell. Due to the
location of Krt15+ cells at the bottom of crypt we expect a certain overlap between the Lgr5+ and
Krt15+ population, however both cell types are functionally distinct by their relative
radioresistance. The direct interaction of Krt15+ cells with the Lgr5+ CBC cell population is yet to
be analyzed. The lineage tracing assays indicated that Krt15+ progeny cells can differentiate into
all known cell types with continuously active Krt15 promoter. Moreover, loss of Apc the Krt15+
cells seems to trigger cell degeneration and cancer initiation. Krt15-CrePR1;Apcfl/fl mice all
developed adenomas as well as occasional progress to adenocarcinoma. Since it is believed that
most intestinal carcinomas origin from mutated stem cell, a more in depth knowledge of specific
markers becomes crucial. We propose that Krt15 marks a type of long-lived radioresistant stem
cells, inherently influencing crypt homeostasis and regenerative capacity

8. ACKNOWLEDGEMENT
76
8. ACKNOWLEDGEMENT I am grateful for the continued support and scientific training by Prof. Dr. Markus M. Lerch as
well as all members of Lerch lab (University of Greifswald). Special thanks goes to Dr. Anil K.
Rustgi and PhD Veronique Giroux for the substantial support. Furthermore, I am grateful to Dr.
Christopher Lengner and Dr. Ning Li as well as members of the Rustgi lab for discussions and
comments in the manuscript. We thank the Molecular Pathology and Imaging Core, Human
Microbial Analytic Depository Core, Cell Culture and iPS Core, Genetic and Modified Mouse
Core and FACS/sorting Core facilities (University of Pennsylvania). We thank Dr. George
Cotsarelis (University of Pennsylvania) for Krt15-CrePR1 and Krt15-/- mice and Dr. Christopher
Lengner (University of Pennsylvania) for RosaLSL-tdTomato mice. This work was supported by NCI
P01-CA098101 (VG, AJKS, AKR), NIH/NIDDK P30-DK050306 Center of Molecular Studies in
Digestive and Liver Diseases, American Cancer Society (AKR), Fonds de recherche en santé du
Québec P-Giroux-27692 and P-Giroux-31601 (VG), NIH NIDDK K01-DK100485 (KEH) and
Crohn’s and Colitis Foundation Career Development Award (KEH).
8.1. Author contribution Veronique Giroux (VG), Katheryn E. Hamilton (KEH), Anil K. Rustgi (AKR) and Julien Stephan
(JS) designed the study. Veronique Giroux (VG) performed the 3D organoid experiments as well
as the cancer initiation model, while all other experiments were performed by Julien Stephan (JS).
Ben Rhoades (BR) maintained the mouse colony.

9. BIBLIOGRAPHY
77
9. BIBLIOGRAPHY Abremski, K., and Hoess, R. (1984). Bacteriophage P1 site-specific recombination. Purification and properties of the Cre recombinase protein. J. Biol. Chem. 259, 1509–1514.
Al Alam, D., Danopoulos, S., Schall, K., Sala, F.G., Almohazey, D., Fernandez, G.E., Georgia, S., Frey, M.R., Ford, H.R., Grikscheit, T., et al. (2015). Fibroblast growth factor 10 alters the balance between goblet and Paneth cells in the adult mouse small intestine. Am. J. Physiol. - Gastrointest. Liver Physiol. 308, G678–G690.
Asfaha, S., Hayakawa, Y., Muley, A., Stokes, S., Graham, T.A., Ericksen, R.E., Westphalen, C.B., von Burstin, J., Mastracci, T.L., Worthley, D.L., et al. (2015a). Krt19+/Lgr5− Cells Are Radioresistant Cancer-Initiating Stem Cells in the Colon and Intestine. Cell Stem Cell 16, 627–638.
Asfaha, S., Hayakawa, Y., Muley, A., Stokes, S., Graham, T.A., Ericksen, R.E., Westphalen, C.B., von Burstin, J., Mastracci, T.L., Worthley, D.L., et al. (2015b). Krt19(+)/Lgr5(-) Cells Are Radioresistant Cancer-Initiating Stem Cells in the Colon and Intestine. Cell Stem Cell 16, 627–638.
Barker, N. (2014). Adult intestinal stem cells: critical drivers of epithelial homeostasis and regeneration. Nat. Rev. Mol. Cell Biol. 15, 19–33.
Barker, N., van Es, J.H., Kuipers, J., Kujala, P., van den Born, M., Cozijnsen, M., Haegebarth, A., Korving, J., Begthel, H., Peters, P.J., et al. (2007). Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 449, 1003–1007.
Barker, N., Ridgway, R.A., van Es, J.H., van de Wetering, M., Begthel, H., van den Born, M., Danenberg, E., Clarke, A.R., Sansom, O.J., and Clevers, H. (2009). Crypt stem cells as the cells-of-origin of intestinal cancer. Nature 457, 608–611.
Booth, C., Booth, D., Williamson, S., Demchyshyn, L.L., and Potten, C.S. (2004). Teduglutide ([Gly2]GLP-2) protects small intestinal stem cells from radiation damage. Cell Prolif. 37, 385–400.
Booth, C., Tudor, G.L., Katz, B.P., and MacVittie, T.J. (2015). The Delayed Effects of Acute Radiation Syndrome: Evidence of long-term functional changes in the clonogenic cells of the small intestine. Health Phys. 109, 399–413.
Bose, A., Teh, M.-T., Mackenzie, I.C., and Waseem, A. (2013). Keratin K15 as a Biomarker of Epidermal Stem Cells. Int. J. Mol. Sci. 14, 19385–19398.
Bragulla, H.H., and Homberger, D.G. (2009). Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 214, 516–559.
Carulli, A.J., Keeley, T.M., Demitrack, E.S., Chung, J., Maillard, I., and Samuelson, L.C. (2015). Notch receptor regulation of intestinal stem cell homeostasis and crypt regeneration. Dev. Biol. 402, 98–108.

9. BIBLIOGRAPHY
78
Cooper, G.M. (2000). The Cell (Sinauer Associates).
Dick, J.E. (2009). Looking ahead in cancer stem cell research. Nat. Biotechnol. 27, 44–46.
Driskell, I., Oeztuerk-Winder, F., Humphreys, P., and Frye, M. (2015). Genetically Induced Cell Death in Bulge Stem Cells Reveals Their Redundancy for Hair and Epidermal Regeneration. Stem Cells Dayt. Ohio 33, 988–998.
Eriksson, J.E., Dechat, T., Grin, B., Helfand, B., Mendez, M., Pallari, H.-M., and Goldman, R.D. (2009). Introducing intermediate filaments: from discovery to disease. J. Clin. Invest. 119, 1763–1771.
van Es, J.H., Sato, T., van de Wetering, M., Lyubimova, A., Yee Nee, A.N., Gregorieff, A., Sasaki, N., Zeinstra, L., van den Born, M., Korving, J., et al. (2012). Dll1+ secretory progenitor cells revert to stem cells upon crypt damage. Nat. Cell Biol. 14, 1099–1104.
Espersen, M.L.M., Olsen, J., Linnemann, D., Høgdall, E., and Troelsen, J.T. (2015). Clinical Implications of Intestinal Stem Cell Markers in Colorectal Cancer. Clin. Colorectal Cancer 14, 63–71.
Fafilek, B., Krausova, M., Vojtechova, M., Pospichalova, V., Tumova, L., Sloncova, E., Huranova, M., Stancikova, J., Hlavata, A., Svec, J., et al. (2013). Troy, a tumor necrosis factor receptor family member, interacts with lgr5 to inhibit wnt signaling in intestinal stem cells. Gastroenterology 144, 381–391.
Fan, X.-S., Wu, H.-Y., Yu, H.-P., Zhou, Q., Zhang, Y.-F., and Huang, Q. (2010). Expression of Lgr5 in human colorectal carcinogenesis and its potential correlation with beta-catenin. Int. J. Colorectal Dis. 25, 583–590.
Gaskins, H.R., Croix, J.A., Nakamura, N., and Nava, G.M. (2008). Impact of the Intestinal Microbiota on the Development of Mucosal Defense. Clin. Infect. Dis. 46, S80–S86.
Gierut, J.J., Jacks, T.E., and Haigis, K.M. (2014). Strategies to Achieve Conditional Gene Mutation in Mice. Cold Spring Harb. Protoc. 2014, 339–349.
van Gijlswijk, R.P., Wiegant, J., Vervenne, R., Lasan, R., Tanke, H.J., and Raap, A.K. (1996). Horseradish peroxidase-labeled oligonucleotides and fluorescent tyramides for rapid detection of chromosome-specific repeat sequences. Cytogenet. Cell Genet. 75, 258–262.
Giroux, V., Lento, A.A., Islam, M., Pitarresi, J.R., Kharbanda, A., Hamilton, K.E., Whelan, K.A., Long, A., Rhoades, B., Tang, Q., et al. (2017). Long-lived keratin 15+ esophageal progenitor cells contribute to homeostasis and regeneration. J. Clin. Invest. 127.
Giroux, V., Stephan, J., Chatterji, P., Rhoades, B., Wileyto, E.P., Klein-Szanto, A.J., Lengner, C.J., Hamilton, K.E., and Rustgi, A.K. (2018). Mouse Intestinal Krt15+ Crypt Cells Are Radio-Resistant and Tumor Initiating. Stem Cell Rep.

9. BIBLIOGRAPHY
79
Haegebarth, A., and Clevers, H. (2009). Wnt Signaling, Lgr5, and Stem Cells in the Intestine and Skin. Am. J. Pathol. 174, 715–721.
Hamilton, K.E., Crissey, M.A.S., Lynch, J.P., and Rustgi, A.K. (2015). Culturing Adult Stem Cells from Mouse Small Intestinal Crypts. Cold Spring Harb. Protoc. 2015, pdb.prot078303.
Hao, H.-X., Xie, Y., Zhang, Y., Charlat, O., Oster, E., Avello, M., Lei, H., Mickanin, C., Liu, D., Ruffner, H., et al. (2012). ZNRF3 promotes Wnt receptor turnover in an R-spondin-sensitive manner. Nature 485, 195–200.
He, J., Stewart, K., Kinnell, H.L., Anderson, R.A., and Childs, A.J. (2013). A developmental stage-specific switch from DAZL to BOLL occurs during fetal oogenesis in humans, but not mice. PloS One 8, e73996.
He, X.C., Yin, T., Grindley, J.C., Tian, Q., Sato, T., Tao, W.A., Dirisina, R., Porter-Westpfahl, K.S., Hembree, M., Johnson, T., et al. (2007). PTEN-deficient intestinal stem cells initiate intestinal polyposis. Nat. Genet. 39, 189–198.
Helander, H.F., and Fändriks, L. (2014). Surface area of the digestive tract - revisited. Scand. J. Gastroenterol. 49, 681–689.
Hsu, S.M., Raine, L., and Fanger, H. (1981). Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: a comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. Off. J. Histochem. Soc. 29, 577–580.
Inoue, K., Aoi, N., Sato, T., Yamauchi, Y., Suga, H., Eto, H., Kato, H., Araki, J., and Yoshimura, K. (2009). Differential expression of stem-cell-associated markers in human hair follicle epithelial cells. Lab. Investig. J. Tech. Methods Pathol. 89, 844–856.
Ito, M., Liu, Y., Yang, Z., Nguyen, J., Liang, F., Morris, R.J., and Cotsarelis, G. (2005). Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat. Med. 11, 1351–1354.
Jaks, V., Barker, N., Kasper, M., van Es, J.H., Snippert, H.J., Clevers, H., and Toftgård, R. (2008). Lgr5 marks cycling, yet long-lived, hair follicle stem cells. Nat. Genet. 40, 1291–1299.
Jemal, A., Bray, F., Center, M.M., Ferlay, J., Ward, E., and Forman, D. (2011). Global cancer statistics. CA. Cancer J. Clin. 61, 69–90.
Jones, J.W., Tudor, G., Li, F., Tong, Y., Katz, B., Farese, A.M., MacVittie, T.J., Booth, C., and Kane, M.A. (2015). Citrulline as a biomarker in the murine total-body irradiation model: correlation of circulating and tissue citrulline to small intestine epithelial histopathology. Health Phys. 109, 452–465.
Kalabis, J., Oyama, K., Okawa, T., Nakagawa, H., Michaylira, C.Z., Stairs, D.B., Figueiredo, J.-L., Mahmood, U., Diehl, J.A., Herlyn, M., et al. (2008). A subpopulation of mouse esophageal basal cells has properties of stem cells with the capacity for self-renewal and lineage specification.

9. BIBLIOGRAPHY
80
Kantara, C., Moya, S.M., Houchen, C.W., Umar, S., Ullrich, R.L., Singh, P., and Carney, D.H. (2015). Novel Regenerative Peptide TP508 Mitigates Radiation-Induced Gastrointestinal Damage By Activating Stem Cells and Preserving Crypt Integrity. Lab. Investig. J. Tech. Methods Pathol. 95, 1222–1233.
Kloepper, J.E., Tiede, S., Brinckmann, J., Reinhardt, D.P., Meyer, W., Faessler, R., and Paus, R. (2008). Immunophenotyping of the human bulge region: the quest to define useful in situ markers for human epithelial hair follicle stem cells and their niche. Exp. Dermatol. 17, 592–609.
Korinek, V., Barker, N., Morin, P.J., van Wichen, D., de Weger, R., Kinzler, K.W., Vogelstein, B., and Clevers, H. (1997). Constitutive transcriptional activation by a beta-catenin-Tcf complex in APC-/- colon carcinoma. Science 275, 1784–1787.
Kuhnert, F., Davis, C.R., Wang, H.-T., Chu, P., Lee, M., Yuan, J., Nusse, R., and Kuo, C.J. (2004). Essential requirement for Wnt signaling in proliferation of adult small intestine and colon revealed by adenoviral expression of Dickkopf-1. Proc. Natl. Acad. Sci. U. S. A. 101, 266–271.
Kuraguchi, M., Wang, X.-P., Bronson, R.T., Rothenberg, R., Ohene-Baah, N.Y., Lund, J.J., Kucherlapati, M., Maas, R.L., and Kucherlapati, R. (2006). Adenomatous Polyposis Coli (APC) Is Required for Normal Development of Skin and Thymus. PLOS Genet. 2, e146.
de Lau, W., Barker, N., Low, T.Y., Koo, B.-K., Li, V.S.W., Teunissen, H., Kujala, P., Haegebarth, A., Peters, P.J., van de Wetering, M., et al. (2011). Lgr5 homologues associate with Wnt receptors and mediate R-spondin signalling. Nature 476, 293–297.
Li, D., Tang, H., Fan, J., Yan, D., Zhou, C., Li, S., Wang, X., and Peng, Z. (2010). Expression level of Bmi-1 oncoprotein is associated with progression and prognosis in colon cancer. J. Cancer Res. Clin. Oncol. 136, 997–1006.
Li, S., Park, H., Trempus, C.S., Gordon, D., Liu, Y., Cotsarelis, G., and Morris, R.J. (2013). A keratin 15 containing stem cell population from the hair follicle contributes to squamous papilloma development in the mouse. Mol. Carcinog. 52, 751–759.
Liu, G., Amin, S., Okuhama, N.N., Liao, G., and Mingle, L.A. (2006). A quantitative evaluation of peroxidase inhibitors for tyramide signal amplification mediated cytochemistry and histochemistry. Histochem. Cell Biol. 126, 283–291.
Liu, Y., Lyle, S., Yang, Z., and Cotsarelis, G. (2003). Keratin 15 Promoter Targets Putative Epithelial Stem Cells in the Hair Follicle Bulge. J. Invest. Dermatol. 121, 963–968.
Lozano, R., Naghavi, M., Foreman, K., Lim, S., Shibuya, K., Aboyans, V., Abraham, J., Adair, T., Aggarwal, R., Ahn, S.Y., et al. (2012). Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380, 2095–2128.
Ma, D.R., Yang, E.N., and Lee, S.T. (2004). A review: the location, molecular characterisation and multipotency of hair follicle epidermal stem cells. Ann. Acad. Med. Singapore 33, 784–788.

9. BIBLIOGRAPHY
81
May, R., Riehl, T.E., Hunt, C., Sureban, S.M., Anant, S., and Houchen, C.W. (2008). Identification of a Novel Putative Gastrointestinal Stem Cell and Adenoma Stem Cell Marker, Doublecortin and CaM Kinase-Like-1, Following Radiation Injury and in Adenomatous Polyposis Coli/Multiple Intestinal Neoplasia Mice. STEM CELLS 26, 630–637.
Michel, M., Török, N., Godbout, M.J., Lussier, M., Gaudreau, P., Royal, A., and Germain, L. (1996). Keratin 19 as a biochemical marker of skin stem cells in vivo and in vitro: keratin 19 expressing cells are differentially localized in function of anatomic sites, and their number varies with donor age and culture stage. J. Cell Sci. 109 ( Pt 5), 1017–1028.
Montgomery, R.K., Carlone, D.L., Richmond, C.A., Farilla, L., Kranendonk, M.E.G., Henderson, D.E., Baffour-Awuah, N.Y., Ambruzs, D.M., Fogli, L.K., Algra, S., et al. (2011). Mouse telomerase reverse transcriptase (mTert) expression marks slowly cycling intestinal stem cells. Proc. Natl. Acad. Sci. 108, 179–184.
Morris, R.J., Liu, Y., Marles, L., Yang, Z., Trempus, C., Li, S., Lin, J.S., Sawicki, J.A., and Cotsarelis, G. (2004). Capturing and profiling adult hair follicle stem cells. Nat. Biotechnol. 22, 411–417.
Mouse Genome Sequencing Consortium, Waterston, R.H., Lindblad-Toh, K., Birney, E., Rogers, J., Abril, J.F., Agarwal, P., Agarwala, R., Ainscough, R., Alexandersson, M., et al. (2002). Initial sequencing and comparative analysis of the mouse genome. Nature 420, 520–562.
Muñoz, J., Stange, D.E., Schepers, A.G., van de Wetering, M., Koo, B.-K., Itzkovitz, S., Volckmann, R., Kung, K.S., Koster, J., Radulescu, S., et al. (2012). The Lgr5 intestinal stem cell signature: robust expression of proposed quiescent “+4” cell markers. EMBO J. 31, 3079–3091.
Otsuka, K., Hamada, N., Magae, J., Matsumoto, H., Hoshi, Y., and Iwasaki, T. (2013). Ionizing radiation leads to the replacement and de novo production of colonic Lgr5(+) stem cells. Radiat. Res. 179, 637–646.
Pellegrinet, L., Rodilla, V., Liu, Z., Chen, S., Koch, U., Espinosa, L., Kaestner, K.H., Kopan, R., Lewis, J., and Radtke, F. (2011). Dll1- and dll4-mediated notch signaling are required for homeostasis of intestinal stem cells. Gastroenterology 140, 1230-1240.e1-7.
Porter, R.M., Lunny, D.P., Ogden, P.H., Morley, S.M., McLean, W.H., Evans, A., Harrison, D.L., Rugg, E.L., and Lane, E.B. (2000). K15 expression implies lateral differentiation within stratified epithelial basal cells. Lab. Investig. J. Tech. Methods Pathol. 80, 1701–1710.
Potten, C.S., Merritt, A., Hickman, J., Hall, P., and Faranda, A. (1994). Characterization of radiation-induced apoptosis in the small intestine and its biological implications. Int. J. Radiat. Biol. 65, 71–78.
Powell, A.E., Wang, Y., Li, Y., Poulin, E.J., Means, A.L., Washington, M.K., Higginbotham, J.N., Juchheim, A., Prasad, N., Levy, S.E., et al. (2012). The Pan-ErbB Negative Regulator Lrig1 Is an Intestinal Stem Cell Marker that Functions as a Tumor Suppressor. Cell 149, 146–158.

9. BIBLIOGRAPHY
82
Powell, A.E., Vlacich, G., Zhao, Z.-Y., McKinley, E.T., Washington, M.K., Manning, H.C., and Coffey, R.J. (2014). Inducible loss of one Apc allele in Lrig1-expressing progenitor cells results in multiple distal colonic tumors with features of familial adenomatous polyposis. Am. J. Physiol. Gastrointest. Liver Physiol. 307, G16-23.
Reinisch, C., Kandutsch, S., Uthman, A., and Pammer, J. (2006). BMI-1: a protein expressed in stem cells, specialized cells and tumors of the gastrointestinal tract. Histol. Histopathol. 21, 1143–1149.
Roche, K.C., Gracz, A.D., Liu, X.F., Newton, V., Akiyama, H., and Magness, S.T. (2015). SOX9 Maintains Reserve Stem Cells and Preserves Radioresistance in Mouse Small Intestine. Gastroenterology 149, 1553-1563.e10.
Roe, D.J., Alberts, D.S., Wargovich, M.J., Bostick, R.M., Garewal, H.S., Einspahr, J., Fosdick, L., Ramsey, L., Woods, C., and McGee, D.L. (1996). Reproducibility of the measurement of colonic proliferation using bromodeoxyuridine across laboratories. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 5, 349–353.
Sahlgren, C.M., Mikhailov, A., Vaittinen, S., Pallari, H.-M., Kalimo, H., Pant, H.C., and Eriksson, J.E. (2003). Cdk5 regulates the organization of Nestin and its association with p35. Mol. Cell. Biol. 23, 5090–5106.
Sailaja, B.S., He, X.C., and Li, L. (2016). The regulatory niche of intestinal stem cells. J. Physiol. 594, 4827–4836.
Sangiorgi, E., and Capecchi, M.R. (2008a). Bmi1 is expressed in vivo in intestinal stem cells. Nat. Genet. 40, 915–920.
Sangiorgi, E., and Capecchi, M.R. (2008b). Bmi1 is expressed in vivo in intestinal stem cells. Nat. Genet. 40, 915–920.
Schofield, R. (1978). The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells 4, 7–25.
Scholzen, T., and Gerdes, J. (2000). The Ki-67 protein: from the known and the unknown. J. Cell. Physiol. 182, 311–322.
Schuijers, J., van der Flier, L.G., van Es, J., and Clevers, H. (2014). Robust Cre-Mediated Recombination in Small Intestinal Stem Cells Utilizing the Olfm4 Locus. Stem Cell Rep. 3, 234–241.
Schuijers, J., Junker, J.P., Mokry, M., Hatzis, P., Koo, B.-K., Sasselli, V., van der Flier, L.G., Cuppen, E., van Oudenaarden, A., and Clevers, H. (2015). Ascl2 acts as an R-spondin/Wnt-responsive switch to control stemness in intestinal crypts. Cell Stem Cell 16, 158–170.
Sei, Y., Lu, X., Liou, A., Zhao, X., and Wank, S.A. (2011). A stem cell marker-expressing subset of enteroendocrine cells resides at the crypt base in the small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 300, G345-356.

9. BIBLIOGRAPHY
83
Seshagiri, S., Stawiski, E.W., Durinck, S., Modrusan, Z., Storm, E.E., Conboy, C.B., Chaudhuri, S., Guan, Y., Janakiraman, V., Jaiswal, B.S., et al. (2012). Recurrent R-spondin fusions in colon cancer. Nature 488, 660–664.
Snippert, H.J., van Es, J.H., van den Born, M., Begthel, H., Stange, D.E., Barker, N., and Clevers, H. (2009). Prominin-1/CD133 marks stem cells and early progenitors in mouse small intestine. Gastroenterology 136, 2187-2194.e1.
Szeverenyi, I., Cassidy, A.J., Chung, C.W., Lee, B.T.K., Common, J.E.A., Ogg, S.C., Chen, H., Sim, S.Y., Goh, W.L.P., Ng, K.W., et al. (2008). The Human Intermediate Filament Database: comprehensive information on a gene family involved in many human diseases. Hum. Mutat. 29, 351–360.
Takahashi, H., Ishii, H., Nishida, N., Takemasa, I., Mizushima, T., Ikeda, M., Yokobori, T., Mimori, K., Yamamoto, H., Sekimoto, M., et al. (2011). Significance of Lgr5+ve Cancer Stem Cells in the Colon and Rectum. Ann. Surg. Oncol. 18, 1166–1174.
Takeda, K., Kinoshita, I., Shimizu, Y., Matsuno, Y., Shichinohe, T., and Dosaka-Akita, H. (2011). Expression of LGR5, an intestinal stem cell marker, during each stage of colorectal tumorigenesis. Anticancer Res. 31, 263–270.
Tan, D.W.-M., and Barker, N. (2014). Chapter Three - Intestinal Stem Cells and Their Defining Niche. In Current Topics in Developmental Biology, M. Rendl, ed. (Academic Press), pp. 77–107.
Tateishi, K., Ohta, M., Kanai, F., Guleng, B., Tanaka, Y., Asaoka, Y., Tada, M., Seto, M., Jazag, A., Lianjie, L., et al. (2006). Dysregulated expression of stem cell factor Bmi1 in precancerous lesions of the gastrointestinal tract. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 12, 6960–6966.
Troy, T.-C., Arabzadeh, A., and Turksen, K. (2011). Re-Assessing K15 as an Epidermal Stem Cell Marker. Stem Cell Rev. Rep. 7, 927–934.
Vries, R.G.J., Huch, M., and Clevers, H. (2010). Stem cells and cancer of the stomach and intestine. Mol. Oncol. 4, 373–384.
Waseem, A., Dogan, B., Tidman, N., Alam, Y., Purkis, P., Jackson, S., Lalli, A., Machesney, M., and Leigh, I.M. (1999). Keratin 15 expression in stratified epithelia: downregulation in activated keratinocytes. J. Invest. Dermatol. 112, 362–369.
Withers, H.R., and Elkind, M.M. (1970). Microcolony survival assay for cells of mouse intestinal mucosa exposed to radiation. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 17, 261–267.
Wong, V.W.Y., Stange, D.E., Page, M.E., Buczacki, S., Wabik, A., Itami, S., van de Wetering, M., Poulsom, R., Wright, N.A., Trotter, M.W.B., et al. (2012). Lrig1 controls intestinal stem-cell homeostasis by negative regulation of ErbB signalling. Nat. Cell Biol. 14, 401–408.
Yamazaki, M., Kato, A., Zaitsu, Y., Watanabe, T., Iimori, M., Funahashi, S., Kitao, H., Saeki, H., Oki, E., and Suzuki, M. (2015). Intensive Immunofluorescence Staining Methods for Low

9. BIBLIOGRAPHY
84
Expression Protein: Detection of Intestinal Stem Cell Marker LGR5. Acta Histochem. Cytochem. 48, 159–164.
Yan, K.S., Chia, L.A., Li, X., Ootani, A., Su, J., Lee, J.Y., Su, N., Luo, Y., Heilshorn, S.C., Amieva, M.R., et al. (2012). The intestinal stem cell markers Bmi1 and Lgr5 identify two functionally distinct populations. Proc. Natl. Acad. Sci. U. S. A. 109, 466–471.

10. ORIGINAL PAPER
85
10. ORIGINAL PAPER
Mouse Intestinal Krt15+ Crypt Cells Are Radio-Resistant and
Tumor Initiating
Giroux, V., Stephan, J., Chatterji, P., Rhoades, B., Wileyto, E.P., Klein-Szanto, A.J., Lengner,
C.J., Hamilton, K.E., and Rustgi, A.K. (2018).
Stem Cell Reports: 2018 Jun 5; 10(6): 1947–1958.

10. ORIGINAL PAPER
86
Stem Cell ReportsArticle
Mouse Intestinal Krt15+Crypt Cells Are Radio-Resistant and Tumor Initiating
Veronique Giroux,1,2 Julien Stephan,1,2 Priya Chatterji,1,2 Ben Rhoades,1,2 E. Paul Wileyto,3
Andres J. Klein-Szanto,4 Christopher J. Lengner,5 Kathryn E. Hamilton,1,2,6 and Anil K. Rustgi1,2,7,*1Division of Gastroenterology, Department of Medicine, University of Pennsylvania, Perelman School of Medicine, 951 BRBII/III, 421 Curie Boulevard,Philadelphia, PA 19104, USA2Abramson Cancer Center, University of Pennsylvania, Philadelphia, PA 19104, USA3Department of Biostatistics and Epidemiology, University of Pennsylvania, Philadelphia, PA 19104, USA4Department of Pathology and Cancer Biology Program, Fox Chase Cancer Center, Philadelphia, PA 19111, USA5Department of Biomedical Sciences, School of Veterinary Medicine, Institute for Regenerative Medicine, University of Pennsylvania, Philadelphia,PA 19104, USA6Department of Pediatrics, Division of Gastroenterology, Hepatology and Nutrition, Children’s Hospital of Philadelphia, Philadelphia, PA 19104, USA7Department of Genetics, University of Pennsylvania, Philadelphia, PA 19104, USA*Correspondence: [email protected]://doi.org/10.1016/j.stemcr.2018.04.022
SUMMARY
Two principal stem cell pools orchestrate the rapid cell turnover in the intestinal epithelium. Rapidly cycling Lgr5+ stem cells are inter-
calated between the Paneth cells at the crypt base (CBCs) and injury-resistant reserve stem cells reside above the crypt base. The inter-
mediate filament Keratin 15 (Krt15) marks either stem cells or long-lived progenitor cells that contribute to tissue repair in the hair follicle
or the esophageal epithelium. Herein, we demonstrate that Krt15 labels long-lived and multipotent cells in the small intestinal crypt by
lineage tracing.Krt15+ crypt cells display self-renewal potential in vivo and in 3Dorganoid cultures.Krt15+ crypt cells are resistant to high-
dose radiation and contribute to epithelial regeneration following injury. Notably, loss of the tumor suppressorApc inKrt15+ cells leads to
adenoma and adenocarcinoma formation. These results indicate thatKrt15marks long-lived,multipotent, and injury-resistant crypt cells
that may function as a cell of origin in intestinal cancer.
INTRODUCTION
Epithelial stem cells are multipotent cells with self-renewal capacity that ensure normal epithelial renewaland tissue regeneration in response to injury (e.g., radia-tion). The intestinal epithelium is highly proliferative, be-ing renewed every few days in the mouse (Bjerknes andCheng, 1999; Wong et al., 1999; Barker et al., 2012).Two main intestinal stem cell (ISC) pools orchestratemaintenance of the intestinal homeostatic epithelium(Beumer and Clevers, 2016). First, an actively proliferativeLgr5+ crypt base columnar cell population (CBC) is drivenby the activity of the canonical Wnt pathway (Barkeret al., 2007; Kretzschmar and Clevers, 2017). CBCs arealso characterized by the expression of Ascl2, Olfm4 (vander Flier et al., 2009), and Smoc2 (Munoz et al., 2012). Sec-ond, a slower cycling reserve crypt stem cell population islocated around the +4 position above the crypt base andlacks regulation by the canonical WNT signaling pathway(Sangiorgi and Capecchi, 2008). Specifically, reserve ISCsare marked by CreER insertions into the Bmi1 (Sangiorgiand Capecchi, 2008) or Hopx loci (Takeda et al., 2011),as well as by a Tert-CreER transgene mouse (Montgomeryet al., 2011). Reserve ISCs were originally associatedwith label-retention capacities (Potten et al., 1978). Theidentity and function of intestinal label-retaining cells(LRCs) remain to be fully understood, but recent workshows that intestinal LRCs are secretory precursors of Pan-
eth and enteroendocrine cells, located in the crypt andexpress Lgr5 (Buczacki et al., 2013). Subsequent workshowed the label-retaining secretory precursor cells to bea distinct population from the reserve ISCs labeled byCreER knockin reporters (Li et al., 2016).While a body of work has illuminated the distinct
nature of these two populations, certain controversiespersist. For example, in contrast to Bmi1-CreER+ cells,Bmi1-GFP+ cells may represent an enteroendocrine pro-genitor cell population (Jadhav et al., 2017). Furthermore,the heterogeneity of these populations makes interpreta-tion of genetic labeling challenging at times. For example,the RNA binding protein Mex3a marks a subpopulationof Lgr5+ cells displaying characteristics consistent withreserve-like stem cells (Barriga et al., 2017). Other allelescan broadly mark several cell types; for example, Lrig1marks Lgr5+ cells (Wong et al., 2012) and reserve ISCs (Po-well et al., 2012). However, the populations marked byLrig1 can vary greatly depending on whether the readoutis endogenous mRNA, protein (which may be antibodydependent), or reporter alleles (Poulin et al., 2014; Powellet al., 2012; Wong et al., 2012). The Sox9-CreER allelealso marks reserve ISCs and CBCs (Roche et al., 2015).The transcripts of certain reserve stem cell markers areexpressed in other crypt cells, notably CBCs, therebycomplicating analysis (Li et al., 2014; Munoz et al.,2012; Grun et al., 2015). Nevertheless, single-cell profilinghas revealed that Bmi1-CreER+ cells and Hopx-CreER+ cells
Stem Cell Reports j Vol. 10 j 1947–1958 j June 5, 2018 j ª 2018 The Authors. 1947This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).

10. ORIGINAL PAPER
87
are transcriptionally distinct from Lgr5-CreER + cells andgenerate CBCs under homeostatic conditions ( Li et al.,2014 ). Also, it has been demonstrated that Bmi1-CreER-labeled cells could replenish Lgr5+ stem cell population af-ter diphtheria toxin (DT)-mediated ablation ( Tian et al.,2011 ). Hopx-CreER -labeled cells could also give rise toCBCs ( Takeda et al., 2011 ).Interestingly, Lgr5+ cells are sensitive to DNA damage
and largely ablated with high-dose irradiation ( Yan et al.,2012; Hua et al., 2012; Metcalfe et al., 2014; Tao et al.,2015 ), whereas Bmi1-CreER cells ( Yan et al., 2012 ), Hopx-CreER cells ( Yousefi et al., 2016 ), and Lrig1-CreER cells ( Po-well et al., 2012 ) are resistant to high-dose radiation injury.Following radiation, reserve ISCs can give rise to CBCs(Montgomery et al., 2011; Yan et al., 2012; Yousefi et al.,2016 ). Although Lgr5+ cells are sensitive to injury, ablationof Lgr5+ cells concomitant with or following radiationresults in failed regeneration, suggesting that de novogeneration of new Lgr5+ cells is required for efficient tissuerepair ( Metcalfe et al., 2014 ). Interestingly, despite the exis-tence of Wnt-negative, injury-resistant reserve ISCs thatcontribute to intestinal epithelial regeneration, evidenceexists for plasticity in more differentiated intestinal cells.For example, Dll1+ secretory progenitor cells can revert toa stem cell state and give rise to Lgr5+ cells ( van Es et al.,2012 ). More recently, Asfaha et al. ( 2015 ) identifiedKrt19+/Lgr5- radio-resistant and cancer-initiating cells inthe small intestine located above the crypt base. Similarly,alkaline-phosphatase-positive transit-amplifying cells canregenerate CBCs after their genetic ablation with Lgr5-DTR ; however, it remains unknown whether this mecha-nism is employed in DNA damaging injury under physio-logical conditions, which might be expected to also ablatethe very rapidly cycling transit-amplifying cells ( Tettehet al., 2016 ).In the present study, we describe the role of Keratin 15
(Krt15 )-labeled cells in the mouse small intestinal epithe-lium. Krt15 -labeled cells were characterized initially asstem cells in the hair follicle bulge contributing to woundhealing and the development of squamous papilloma(Morris et al., 2004; Ito et al., 2005 ; Li et al., 2013 ). Wedescribed recently a long-lived Krt15+ progenitor cell pop-ulation in the mouse esophageal epithelium ( Giroux et al.,2017 ). Herein, we identify and describe a long-lived Krt15+cell population in the small intestinal crypt using geneticlineage tracing in mice. Krt15+ crypt cells give rise to allthe intestinal lineages and have self-renewal capacity. Ra-dio-resistant Krt15+ cells contribute to tissue regenerationafter radiation-mediated injury. Interestingly, Apc loss inKrt15+ cells leads to adenoma and adenocarcinoma forma-tion in the small intestine, as well as occasional adenomaformation in the colon, demonstrating the tumor-initi-ating potential of these cells.
RESULTS
Krt15 Marks Proliferating Cells in the Small IntestinalCryptKrt15 + stem/progenitor cells were described originally inthe bulge of the hair follicle ( Morris et al., 2004 ) and,more recently, within the basal compartment of the esoph-ageal squamous epithelium ( Giroux et al., 2017 ). Takentogether, this suggests an important role for Krt15+ cellsin the maintenance of squamous epithelia and append-ages. In contrast to the multi-layered squamous epitheliumin the skin and esophagus, a single layer of the columnarepithelium is present in the mammary gland, pancreas,and intestine. We hypothesized that Krt15+ cells in thesmall intestine might annotate a unique subpopulationof cells with properties that contribute to tissue mainte-nance and regeneration.First, we noted that endogenous K15 protein is detected
in some crypt cells and occasionally in cells within the villi(Figure 1 A) of the small intestine. K15 protein can be de-tected in all small intestinal segments with slightly higherexpression in the ileum. We confirmed these results using amouse expressing a progesterone receptor (PR)-fused Crerecombinase downstream of Krt15 promoter ( Krt15-CrePR1mouse) ( Morris et al., 2004 ) crossed with a reporter mouseexpressing the mTomato-LSL-mGFP construct from theRosa26 locus ( R26 mT/mG ) (Muzumdar et al., 2007 ). A singleinjection of the PR agonist, RU486, was used for Cre induc-tion and mice were sacrificed 24 hr later to visualize Krt15+(GFP+) cells. Krt15+ cells were observed in some crypts andoccasionally in the villi of the small intestine ( Figure 1 B). Inthe villi, 10% of the GFP+ cells also stained positive forAlcian blue, a goblet cell marker, and 3 % also expressedchromogranin A (CHGA), an enteroendocrine cell marker(Figures S1 A and S1B), suggesting that the majority ofKrt15+ cells in the villi are enterocytes. Since the Krt15+villi cells could result from migration of a recombinationevent in the top of the crypt, we injected Krt15-CrePR1;R26 mT/mG mice with a single dose of RU486 andmice were sacrificed 8 hr later to minimize the potentialfor migration oflabeled cells. We observed that mostlabeled cells were located in the lower third of the villi,thereby suggesting that recombination happened in thecrypt or at the crypt/villi junction ( Figure S1 C).Interestingly, approximately 40% of Krt15 -labeled cells
in the crypt are localized in the stem cell compartment(i.e., +4 position and below) ( Figures 1 C and S1D). Weconfirmed that the GFP+ cells located between positions +1and +4 from the crypt base are not Paneth cells by lysozymestaining ( Figures 1 D and 1E). GFP can also be detected insome Ki-67 + transit-amplifying cells as a result of possiblestem cell division and/or residual Krt15 promoter activity
1948 Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018
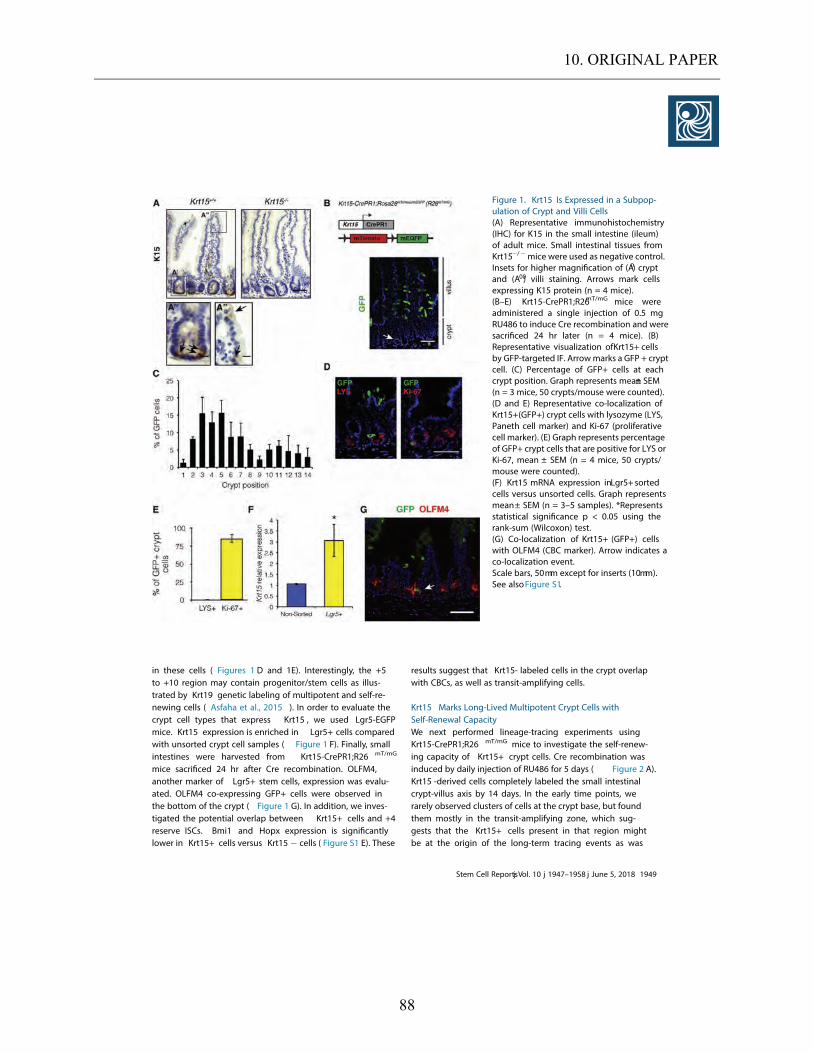
10. ORIGINAL PAPER
88
in these cells ( Figures 1 D and 1E). Interestingly, the +5to +10 region may contain progenitor/stem cells as illus-trated by Krt19 genetic labeling of multipotent and self-re-newing cells ( Asfaha et al., 2015 ). In order to evaluate thecrypt cell types that express Krt15 , we used Lgr5-EGFPmice. Krt15 expression is enriched in Lgr5+ cells comparedwith unsorted crypt cell samples ( Figure 1 F). Finally, smallintestines were harvested from Krt15-CrePR1;R26 mT/mG
mice sacrificed 24 hr after Cre recombination. OLFM4,another marker of Lgr5+ stem cells, expression was evalu-ated. OLFM4 co-expressing GFP+ cells were observed inthe bottom of the crypt ( Figure 1 G). In addition, we inves-tigated the potential overlap between Krt15+ cells and +4reserve ISCs. Bmi1 and Hopx expression is significantlylower in Krt15+ cells versus Krt15 cells ( Figure S1 E). These
results suggest that Krt15- labeled cells in the crypt overlapwith CBCs, as well as transit-amplifying cells.
Krt15 Marks Long-Lived Multipotent Crypt Cells withSelf-Renewal CapacityWe next performed lineage-tracing experiments usingKrt15-CrePR1;R26 mT/mG mice to investigate the self-renew-ing capacity of Krt15+ crypt cells. Cre recombination wasinduced by daily injection of RU48 6 for 5 days ( Figure 2 A).Krt15 -derived cells completely labeled the small intestinalcrypt-villus axis by 14 days. In the early time points, werarely observed clusters of cells at the crypt base, but foundthem mostly in the transit-amplifying zone, which sug-gests that the Krt15+ cells present in that region mightbe at the origin of the long-term tracing events as was
Figure 1. Krt15 Is Expressed in a Subpop-ulation of Crypt and Villi Cells(A) Representative immunohistochemistry(IHC) for K15 in the small intestine (ileum)of adult mice. Small intestinal tissues fromKrt15 / mice were used as negative control.Insets for higher magnification of (A0) cryptand (A00) villi staining. Arrows mark cellsexpressing K15 protein (n = 4 mice).(B–E) Krt15-CrePR1;R26mT/mG mice wereadministered a single injection of 0.5 mgRU48 6 to induce Cre recombination and weresacrificed 2 4 hr later (n = 4 mice). (B)Representative visualization ofKrt15+ cellsby GFP-targeted IF. Arrowmarks a GFP + cryptcell. (C) Percentage of GFP+ cells at eachcrypt position. Graph represents mean± SEM(n = 3 mice, 50 crypts/mouse were counted).(D and E) Representative co-localization ofKrt15+(GFP+) crypt cells with lysozyme (LYS,Paneth cell marker) and Ki-67 (proliferativecell marker). (E) Graph represents percentageof GFP+ crypt cells that are positive for LYS orKi-67 , mean ± SEM (n = 4 mice, 50 crypts/mouse were counted).(F) Krt15 mRNA expression inLgr5+ sortedcells versus unsorted cells. Graph representsmean± SEM (n = 3–5 samples). *Representsstatistical significance p < 0.05 using therank-sum (Wilcoxon) test.(G) Co-localization of Krt15+ (GFP+) cellswith OLFM4 (CBC marker). Arrow indicates aco-localization event.Scale bars, 50mm except for inserts (10mm).See alsoFigure S1.
Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018 1949
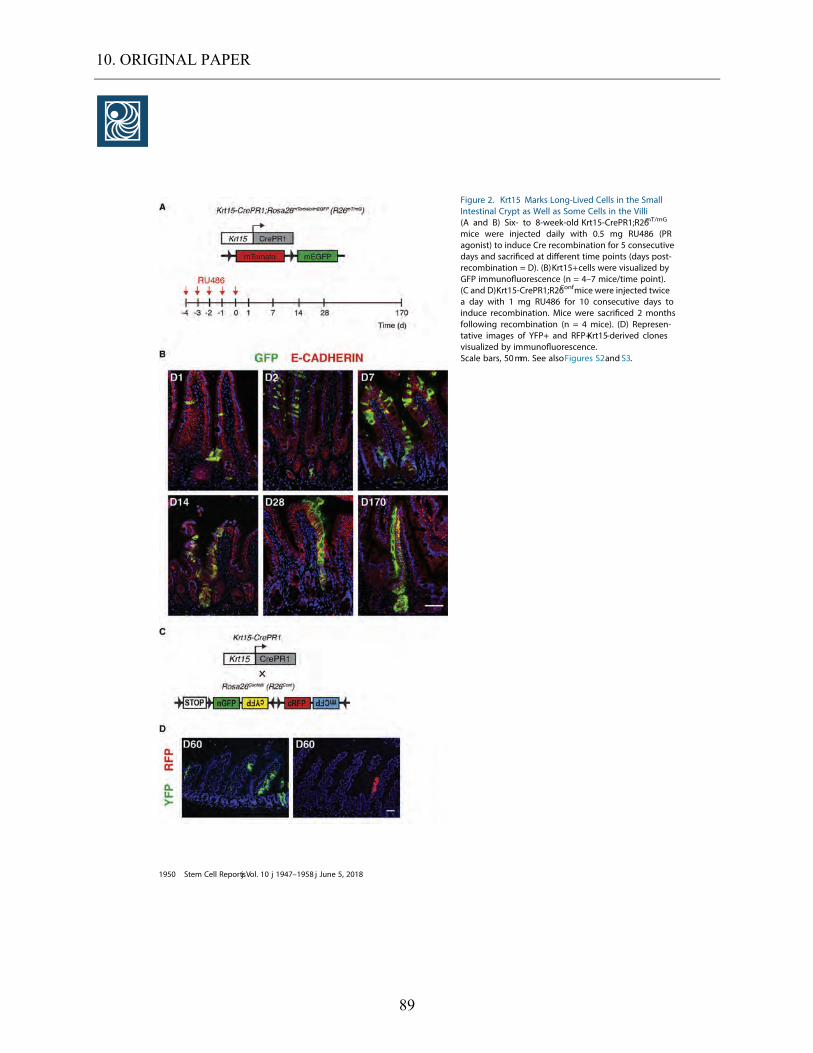
10. ORIGINAL PAPER
89
Figure 2. Krt15 Marks Long-Lived Cells in the SmallIntestinal Crypt as Well as Some Cells in the Villi(A and B) Six- to 8-week-old Krt15-CrePR1;R26mT/mG
mice were injected daily with 0.5 mg RU486 (PRagonist) to induce Cre recombination for 5 consecutivedays and sacrificed at different time points (days post-recombination = D). (B)Krt15+cells were visualized byGFP immunofluorescence (n = 4–7 mice/time point).(C and D)Krt15-CrePR1;R26Confmice were injected twicea day with 1 mg RU486 for 1 0 consecutive days toinduce recombination. Mice were sacrificed 2 monthsfollowing recombination (n = 4 mice). (D) Represen-tative images of YFP+ and RFP+Krt15-derived clonesvisualized by immunofluorescence.Scale bars, 50mm. See alsoFigures S2 and S3 .
1950 Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018

10. ORIGINAL PAPER
90
observed with Krt19+/Lgr5 cells ( Asfaha et al.,2015 ). La-beling persists beyond 6 months,thereby suggesting thatsome Krt15+ crypt cells are long-lived stem cells ( Figures2B and S2A). Notably,7% of the crypts were labeled 1 dayafter the last RU186 injection and 3% after 2 months ( Fig-ure S2 B). Co-localization of GFP with Ki-67,Alcian blue,chromogranin A,and lysozyme demonstrate that Krt15+cells can give rise to proliferative,transit-amplifying cells,goblet cells,enteroendocrine cells,and Paneth cells,respec-tively ( Figures S3 A–S3D). Krt15-CrePR1 mice were alsocrossed with RosaConfetti mice. Every Krt15 -labeled ribbonobserved wasmonochromatic,suggesting that the progenyof Krt15+ cells may arise frommonoclonal units ( Figures 2 Cand 2D). Interestingly, Krt15 marked clones in the colonepithelium as well ( Figure S3 E). Taken together,these re-sults indicate that Krt15 can mark long-lived and multipo-tent crypt stem cells.To investigate the self-renewing capacity of Krt15+ crypt
cells, we used Krt15-CrePR1;Rosa26 LSL-tdTomato (Krt15-CrePR1;R26 Tom) mice. A single injection of RU486 wasused to induce recombination and mice were sacrificed24 hr later. Crypts were isolated and grown as 3D organo-ids. Krt15+ (Tomato+) cells were detected in 3D organoidcrypts and these cells expanded to form lineage-tracedribbons as observed in vivo (Figure 3 A). Interestingly,following 3D organoid passaging,we observed organoidsconsisting entirely of Tomato+ cells ( Figures 3 A and 3B).The multipotency of Krt15+ crypt cells was confirmed inthese 3D organoids with Tomato+ proliferative cells,gobletcells,enteroendocrine cells,and Paneth cells ( Figure 3 C).Furthermore,single crypt cell suspensions were preparedfrom Krt15-CrePR1;R26 Tom mice 24 hr following Cre recom-bination. Krt15+ cells were able to grow as single cell cul-tures,suggesting that Krt15+ crypt cells may display ‘‘stem-ness’’features. The organoid formation efficiency of Krt15+crypt cells is 0.01%,which is less than the 6% efficiency ofLgr5+ cells originally reported ( Sato et al.,2011 ),but similarto the 0.01%–1.2% efficiency reported by other groups(Xian et al.,2017;Qi et al.,2017 ). Of note,these differencesin organoid formation capacity of Krt15+ and Lgr5+ cellsmay not correlate with distinct properties in vivo . Takentogether, these results demonstrate that Krt15 markssome long-lived and multipotent crypt cells with self-re-newing capacity,characteristics consistent with stem cells.
Krt15+ Cells Are Radio-Resistant and Expand inResponse to High-Dose RadiationReserve stem cells are believed to be radio-resistant andcontribute to tissue regeneration following high-dose radi-ation ( Sangiorgi and Capecchi,2008; Yan et al., 2012;Takeda et al., 2011; Yousefi et al., 2016 ). Interestingly,Krt15 mRNA expression is increased in small intestinal cellsfollowing irradiation ( Figure S4 A). Therefore,we investi-
gated the contribution of Krt15+ cells to tissue regenerationfollowing high-dose g-irradiation. First,we used Krt15-CrePR1;R26 mT/mG mice to determine if Krt15+ cells are resis-tant to high-dose radiation. Following Cre recombination,mice were subjected to whole-body g-irradiation (12 Gy)(Figure 4 A). After irradiation, Krt15+ cells were present inthe highly proliferative crypts that underwent regenera-tion and designated as microcolonies ( Figure 4 B). Fivedays following irradiation,the percentage of GFP+ cellsper crypt was increased in the irradiated mice whencompared with non-irradiated mice ( Figure 4 C). Further-more, Krt15+ crypt cells were more proliferative in irradi-ated animals versus non-irradiated mice ( Figures 4 D and4E). These results are consistent with an expansionof Krt15+ cells in response to irradiation. Finally, weinduced Cre recombination in Krt15-CrePR1;R26 mT/mG
mice following whole-body irradiation and noticed thatKrt15+ cells expanded in response to injury in these condi-tions as well ( Figures S4 B and S4C). Thus,we demonstratethat Krt15 -labeled cells are radio-resistant and expand inresponse to high-dose irradiation.Due to themultiple roles of keratins in cell migration and
proliferation,we sought to characterize the potential roleof K15 protein itselfin tissue regeneration following injury.We subjected Krt15 +/+ and Krt15 / mice to whole-bodyg-irradiation (12 Gy) and sacrificed them 5 days later ( Fig-ure 5A). Regeneration of the ileum from Krt15 / micewas impaired following irradiation ( Figure 5 B). First,fewerresidual crypts were observed in the ileum from Krt15 /
compared with Krt15 +/+ irradiated mice ( Figure 5 C). Sec-ond,crypt length and villus height were both significantlyreduced in Krt15 -deficient mice when compared withKrt15 +/+mice ( Figures 5 D and 5E). Third,crypt regenerativecapacity was determined using bromodeoxyuridine (BrdU)incorporation. Crypts containing >10 BrdU+ cells wereconsidered as microcolonies. Krt15 deficiency significantlyreduced the percentage of microcolonies present in theileum 5 days following irradiation ( Figures 5 F and 5G).Finally,the percentage of BrdU+ cells per crypt was lowerin Krt15 / versus Krt15 +/+ ileum. These results suggest apossible role for K15 protein in intestinal tissue regenera-tion. We appreciate this is likely different from the proper-ties of the Krt15+ cells.
Apc Loss in Krt15+ Cells Leads to Adenoma andAdenocarcinoma Formation in the Small Intestineand ColonTumor-initiating capacity is an important feature of ISCs.Lgr5+, Lrig1+, and Krt19+/Lgr5 populations have allbeen described to contain a cell of origin for intestinaltumorigenesis following Apc loss ( Barker et al.,2009;Powellet al.,2012;Asfaha et al.,2015 ). Also, Bmi1-CreER+ cells canform adenomas following induction of a stable form of
Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018 1951
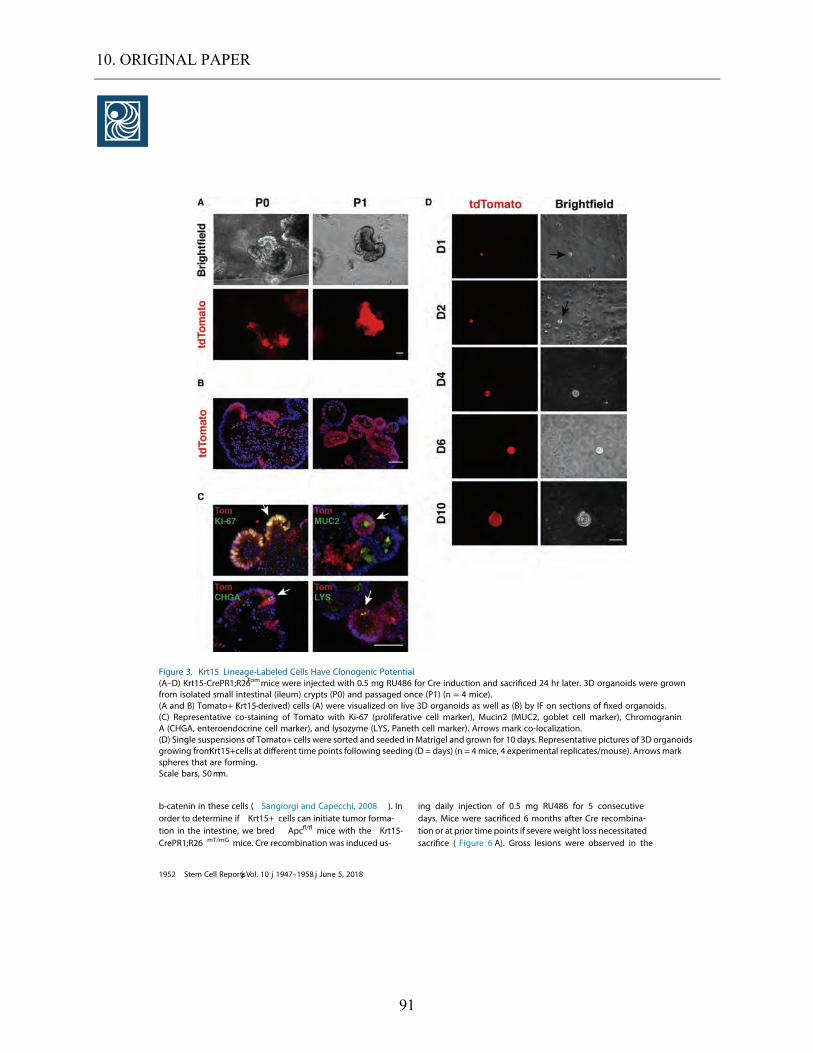
10. ORIGINAL PAPER
91
b-catenin in these cells ( Sangiorgi and Capecchi, 2008 ). Inorder to determine if Krt15+ cells can initiate tumor forma-tion in the intestine, we bred Apcfl/fl mice with the Krt15-CrePR1;R26 mT/mG mice. Cre recombination was induced us-
ing daily injection of 0.5 mg RU48 6 for 5 consecutivedays. Mice were sacrificed 6 months after Cre recombina-tion or at prior time points if severe weight loss necessitatedsacrifice ( Figure 6 A). Gross lesions were observed in the
Figure 3. Krt15 Lineage-Labeled Cells Have Clonogenic Potential(A–D) Krt15-CrePR1;R26Tommice were injected with 0.5 mg RU48 6 for Cre induction and sacrificed 2 4 hr later. 3 D organoids were grownfrom isolated small intestinal (ileum) crypts (P0) and passaged once (P1 ) (n = 4 mice).(A and B) Tomato+ (Krt15-derived) cells (A) were visualized on live 3 D organoids as well as (B) by IF on sections of fixed organoids.(C) Representative co-staining of Tomato with Ki-6 7 (proliferative cell marker), Mucin2 (MUC2 , goblet cell marker), ChromograninA (CHGA, enteroendocrine cell marker), and lysozyme (LYS, Paneth cell marker). Arrows mark co-localization.(D) Single suspensions of Tomato+ cells were sorted and seeded in Matrigel and grown for 1 0 days. Representative pictures of 3 D organoidsgrowing fromKrt15+cells at different time points following seeding (D = days) (n = 4mice, 4 experimental replicates/mouse). Arrowsmarkspheres that are forming.Scale bars, 50mm.
1952 Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018
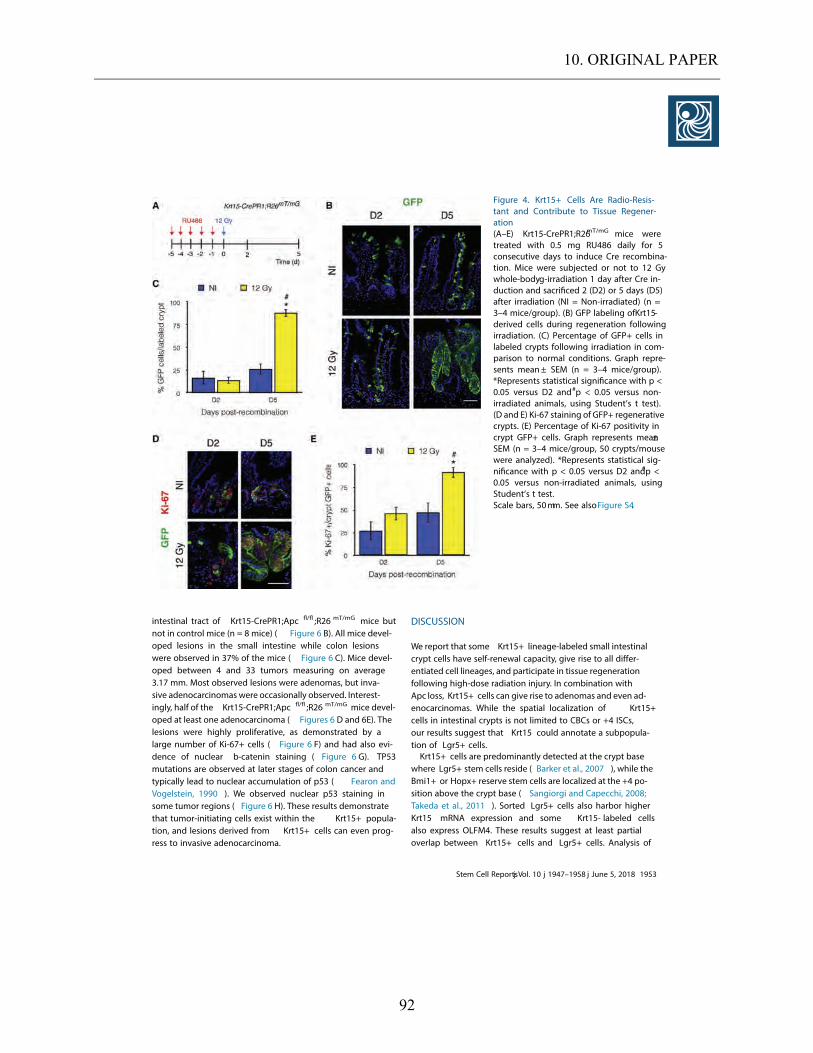
10. ORIGINAL PAPER
92
intestinal tract of Krt15-CrePR1;Apc fl/fl ;R26 mT/mG mice butnot in control mice (n = 8 mice) ( Figure 6 B). All mice devel-oped lesions in the small intestine while colon lesionswere observed in 37 % of the mice ( Figure 6 C). Mice devel-oped between 4 and 33 tumors measuring on average3.17 mm. Most observed lesions were adenomas, but inva-sive adenocarcinomas were occasionally observed. Interest-ingly, half of the Krt15-CrePR1;Apc fl/fl ;R26 mT/mG mice devel-oped at least one adenocarcinoma ( Figures 6 D and 6E). Thelesions were highly proliferative, as demonstrated by alarge number of Ki-67 + cells ( Figure 6 F) and had also evi-dence of nuclear b-catenin staining ( Figure 6 G). TP53mutations are observed at later stages of colon cancer andtypically lead to nuclear accumulation of p5 3 ( Fearon andVogelstein, 1990 ). We observed nuclear p5 3 staining insome tumor regions ( Figure 6 H). These results demonstratethat tumor-initiating cells exist within the Krt15+ popula-tion, and lesions derived from Krt15+ cells can even prog-ress to invasive adenocarcinoma.
DISCUSSION
We report that some Krt15+ lineage-labeled small intestinalcrypt cells have self-renewal capacity, give rise to all differ-entiated cell lineages, and participate in tissue regenerationfollowing high-dose radiation injury. In combination withApc loss, Krt15+ cells can give rise to adenomas and even ad-enocarcinomas. While the spatial localization of Krt15+cells in intestinal crypts is not limited to CBCs or +4 ISCs,our results suggest that Krt15 could annotate a subpopula-tion of Lgr5+ cells.Krt15+ cells are predominantly detected at the crypt base
where Lgr5+ stem cells reside ( Barker et al., 2007 ), while theBmi1+ or Hopx+ reserve stem cells are localized at the +4 po-sition above the crypt base ( Sangiorgi and Capecchi, 2008;Takeda et al., 2011 ). Sorted Lgr5+ cells also harbor higherKrt15 mRNA expression and some Krt15- labeled cellsalso express OLFM4 . These results suggest at least partialoverlap between Krt15+ cells and Lgr5+ cells. Analysis of
Figure 4. Krt15+ Cells Are Radio-Resis-tant and Contribute to Tissue Regener-ation(A–E) Krt15-CrePR1;R26mT/mG mice weretreated with 0.5 mg RU48 6 daily for 5consecutive days to induce Cre recombina-tion. Mice were subjected or not to 1 2 Gywhole-bodyg-irradiation 1 day after Cre in-duction and sacrificed 2 (D2 ) or 5 days (D5)after irradiation (NI = Non-irradiated) (n =3–4 mice/group). (B) GFP labeling ofKrt15-derived cells during regeneration followingirradiation. (C) Percentage of GFP+ cells inlabeled crypts following irradiation in com-parison to normal conditions. Graph repre-sents mean± SEM (n = 3–4 mice/group).*Represents statistical significance with p <0.05 versus D2 and#p < 0.05 versus non-irradiated animals, using Student’s t test).(D and E) Ki-6 7 staining of GFP+ regenerativecrypts. (E) Percentage of Ki-6 7 positivity incrypt GFP+ cells. Graph represents mean±SEM (n = 3–4 mice/group, 50 crypts/mousewere analyzed). *Represents statistical sig-nificance with p < 0.05 versus D2 and#p <0.05 versus non-irradiated animals, usingStudent’s t test.Scale bars, 50mm. See alsoFigure S4.
Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018 1953
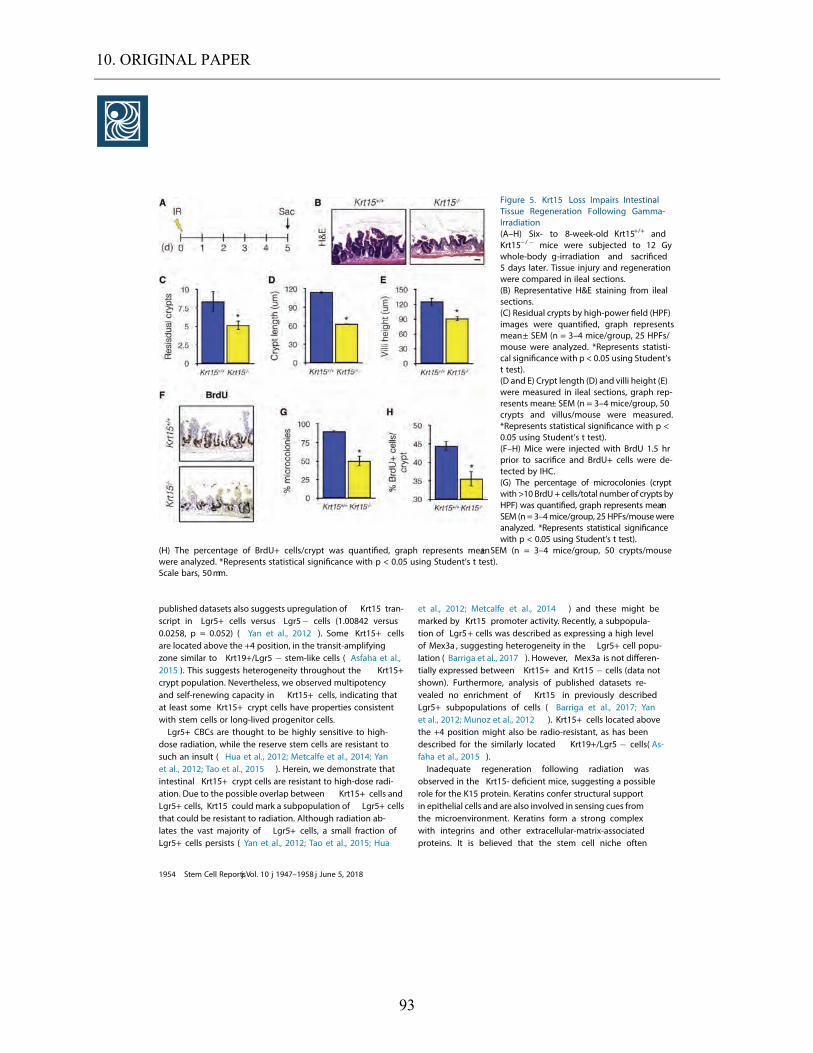
10. ORIGINAL PAPER
93
published datasets also suggests upregulation of Krt15 tran-script in Lgr5+ cells versus Lgr5 cells (1.0084 2 versus0.0258, p = 0.052) ( Yan et al., 2012 ). Some Krt15+ cellsare located above the +4 position, in the transit-amplifyingzone similar to Krt19+/Lgr5 stem-like cells ( Asfaha et al.,2015 ). This suggests heterogeneity throughout the Krt15+crypt population. Nevertheless, we observed multipotencyand self-renewing capacity in Krt15+ cells, indicating thatat least some Krt15+ crypt cells have properties consistentwith stem cells or long-lived progenitor cells.Lgr5+ CBCs are thought to be highly sensitive to high-
dose radiation, while the reserve stem cells are resistant tosuch an insult ( Hua et al., 2012; Metcalfe et al., 2014 ; Yanet al., 2012; Tao et al., 2015 ). Herein, we demonstrate thatintestinal Krt15+ crypt cells are resistant to high-dose radi-ation. Due to the possible overlap between Krt15+ cells andLgr5+ cells, Krt15 could mark a subpopulation of Lgr5+ cellsthat could be resistant to radiation. Although radiation ab-lates the vast majority of Lgr5+ cells, a small fraction ofLgr5+ cells persists ( Yan et al., 2012; Tao et al., 2015; Hua
et al., 2012; Metcalfe et al., 2014 ) and these might bemarked by Krt15 promoter activity. Recently, a subpopula-tion of Lgr5+ cells was described as expressing a high levelof Mex3a , suggesting heterogeneity in the Lgr5+ cell popu-lation ( Barriga et al., 2017 ). However, Mex3a is not differen-tially expressed between Krt15+ and Krt15 cells (data notshown). Furthermore, analysis of published datasets re-vealed no enrichment of Krt15 in previously describedLgr5+ subpopulations of cells ( Barriga et al., 2017 ; Yanet al., 2012; Munoz et al., 2012 ). Krt15+ cells located abovethe +4 position might also be radio-resistant, as has beendescribed for the similarly located Krt19+/Lgr5 cells( As-faha et al., 2015 ).Inadequate regeneration following radiation was
observed in the Krt15- deficient mice, suggesting a possiblerole for the K15 protein. Keratins confer structural supportin epithelial cells and are also involved in sensing cues fromthe microenvironment. Keratins form a strong complexwith integrins and other extracellular-matrix-associatedproteins. It is believed that the stem cell niche often
Figure 5. Krt15 Loss Impairs IntestinalTissue Regeneration Following Gamma-Irradiation(A–H) Six- to 8-week-old Krt15+/+ andKrt15 / mice were subjected to 12 Gywhole-body g-irradiation and sacrificed5 days later. Tissue injury and regenerationwere compared in ileal sections.(B) Representative H&E staining from ilealsections.(C) Residual crypts by high-power field (HPF)images were quantified, graph representsmean± SEM (n = 3–4 mice/group, 2 5 HPFs/mouse were analyzed. *Represents statisti-cal significance with p < 0.05 using Student’st test).(D and E) Crypt length (D) and villi height (E)were measured in ileal sections, graph rep-resents mean± SEM (n = 3–4 mice/group, 50crypts and villus/mouse were measured.*Represents statistical significance with p <0.05 using Student’s t test).(F–H) Mice were injected with BrdU 1.5 hrprior to sacrifice and BrdU+ cells were de-tected by IHC.(G) The percentage of microcolonies (cryptwith >10 BrdU+ cells/total number of crypts byHPF) was quantified, graph represents mean±SEM (n= 3–4 mice/group, 2 5 HPFs/mousewereanalyzed. *Represents statistical significancewith p < 0.05 using Student’s t test).
(H) The percentage of BrdU+ cells/crypt was quantified, graph represents mean± SEM (n = 3–4 mice/group, 50 crypts/mousewere analyzed. *Represents statistical significance with p < 0.05 using Student’s t test).Scale bars, 50mm.
1954 Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018
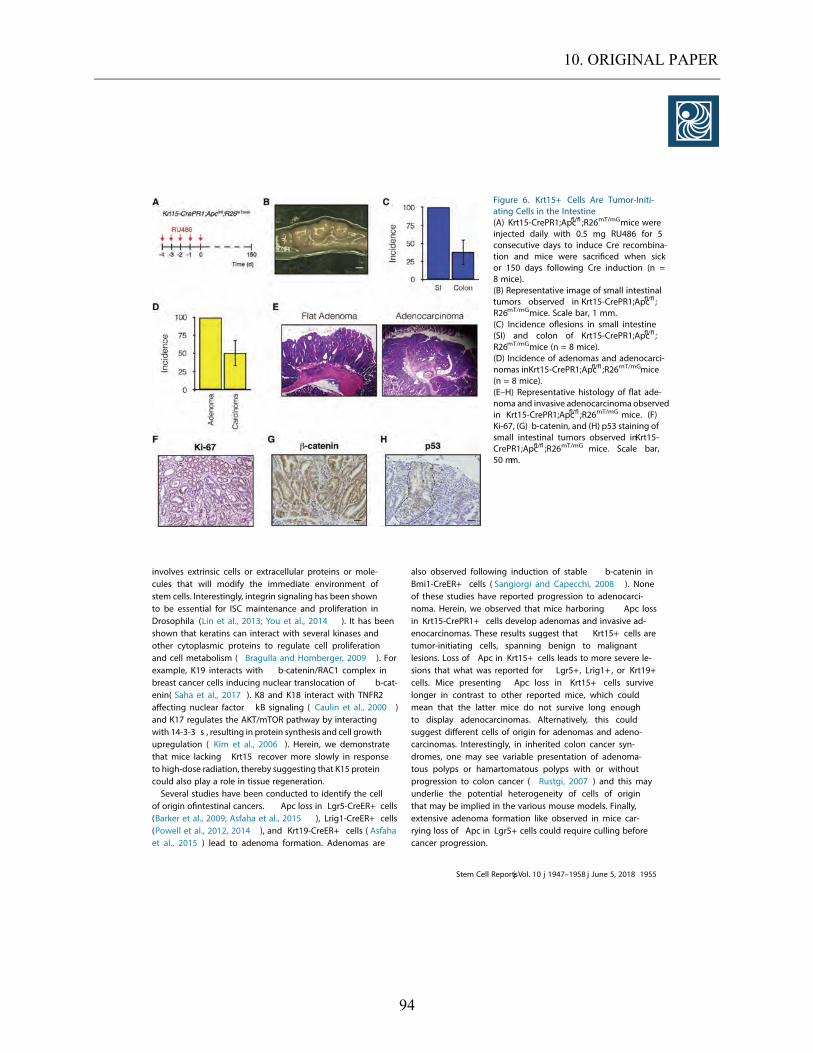
10. ORIGINAL PAPER
94
involves extrinsic cells or extracellular proteins or mole-cules that will modify the immediate environment ofstem cells. Interestingly, integrin signaling has been shownto be essential for ISC maintenance and proliferation inDrosophila (Lin et al., 2013; You et al., 2014 ). It has beenshown that keratins can interact with several kinases andother cytoplasmic proteins to regulate cell proliferationand cell metabolism ( Bragulla and Homberger, 2009 ). Forexample, K19 interacts with b-catenin/RAC1 complex inbreast cancer cells inducing nuclear translocation of b-cat-enin( Saha et al., 2017 ). K8 and K18 interact with TNFR2affecting nuclear factor kB signaling ( Caulin et al., 2000 )and K17 regulates the AKT/mTOR pathway by interactingwith 14-3-3 s , resulting in protein synthesis and cell growthupregulation ( Kim et al., 2006 ). Herein, we demonstratethat mice lacking Krt15 recover more slowly in responseto high-dose radiation, thereby suggesting that K15 proteincould also play a role in tissue regeneration.Several studies have been conducted to identify the cell
of origin ofintestinal cancers. Apc loss in Lgr5-CreER+ cells(Barker et al., 2009 ; Asfaha et al., 2015 ), Lrig1-CreER+ cells(Powell et al., 2012, 2014 ), and Krt19-CreER+ cells ( Asfahaet al., 2015 ) lead to adenoma formation. Adenomas are
also observed following induction of stable b-catenin inBmi1-CreER+ cells ( Sangiorgi and Capecchi, 2008 ). Noneof these studies have reported progression to adenocarci-noma. Herein, we observed that mice harboring Apc lossin Krt15-CrePR1+ cells develop adenomas and invasive ad-enocarcinomas. These results suggest that Krt15+ cells aretumor-initiating cells, spanning benign to malignantlesions. Loss of Apc in Krt15+ cells leads to more severe le-sions that what was reported for Lgr5+, Lrig1+ , or Krt19+cells. Mice presenting Apc loss in Krt15+ cells survivelonger in contrast to other reported mice, which couldmean that the latter mice do not survive long enoughto display adenocarcinomas. Alternatively, this couldsuggest different cells of origin for adenomas and adeno-carcinomas. Interestingly, in inherited colon cancer syn-dromes, one may see variable presentation of adenoma-tous polyps or hamartomatous polyps with or withoutprogression to colon cancer ( Rustgi, 2007 ) and this mayunderlie the potential heterogeneity of cells of originthat may be implied in the various mouse models. Finally,extensive adenoma formation like observed in mice car-rying loss of Apc in Lgr5+ cells could require culling beforecancer progression.
Figure 6. Krt15+ Cells Are Tumor-Initi-ating Cells in the Intestine(A) Krt15-CrePR1;Apcfl/fl ;R26mT/mGmice wereinjected daily with 0.5 mg RU48 6 for 5consecutive days to induce Cre recombina-tion and mice were sacrificed when sickor 150 days following Cre induction (n =8 mice).(B) Representative image of small intestinaltumors observed in Krt15-CrePR1;Apcfl/fl ;R26mT/mGmice. Scale bar, 1 mm.(C) Incidence oflesions in small intestine(SI) and colon of Krt15-CrePR1;Apcfl/fl ;R26mT/mGmice (n = 8 mice).(D) Incidence of adenomas and adenocarci-nomas inKrt15-CrePR1;Apcfl/fl ;R26mT/mGmice(n = 8 mice).(E–H) Representative histology of flat ade-noma and invasive adenocarcinoma observedin Krt15-CrePR1;Apcfl/fl ;R26mT/mG mice. (F)Ki-67 , (G) b-catenin, and (H) p53 staining ofsmall intestinal tumors observed inKrt15-CrePR1;Apcfl/fl ;R26mT/mG mice. Scale bar,50 mm.
Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018 1955

10. ORIGINAL PAPER
95
The lesions formed by Krt15+ cells most likely originatefrom the cells located at the crypt base and/or those locatedin the transit-amplifying zone. We also cannot excludethe possibility that Krt15+ villi cells might have given riseto the lesions since it has been demonstrated that cells inthe villi can initiate tumorigenesis ( Davis et al.,2015 ). Inthat model,aberrant expression of the BMP antagonist,GREM1,led to formation ofintestinal tumors histologicallysimilar to polyps observed in hereditary mixed polyposissyndrome. Nevertheless, Krt15-CrePR1;Apc fl/fl mice mightrepresent a newmousemodel for intestinal cancer as a foun-dation for understanding the interrelationship betweennormal intestinal cells and malignant transformation.Overall,we demonstrate that Krt15+ crypt cells are self-
renewing,multipotent, radio-resistant,and tumor initi-ating in concert with Apc loss. Krt15+ cells could representa radio-resistant subpopulation of Lgr5+ cells. RNAprofiling of Krt15+ esophageal long-lived progenitor cellsrevealed enrichment for gene sets associated with Wnt/b-catenin pathway and DNA repair ( Giroux et al.,2017 ).It is possible,if not likely,that the Wnt/ b-catenin pathwayis involved in the proliferative capacity of Krt15+ intestinalcrypt cells and their tumor-initiating capacities. Further-more,Krt15+ cells could also display higher DNA repair ca-pacity to ensure genomic stability in response to radiationby example.
EXPERIMENTAL PROCEDURES
Mouse Experimental DesignKrt15-CrePR1 (Morris et al.,2004 ) and Krt15 / mice were pro-vided by Dr. George Cotsarelis (University of Pennsylvania).Rosa26mTomato/mGFP (R26mT/mG ) mice ( Muzumdar et al.,2007 ) werepurchased from Jackson Laboratories. Rosa26Confetti (R26Conf) micewere provided by Dr. Ben Stanger (University of Pennsylvania).Rosa26LSL-tdTomato (R26Tom) mice were provided by Dr. ChristopherLengner (University of Pennsylvania). Apcfl/fl mice were obtainedfrom the NCI Mouse Repository ( Kuraguchi et al.,2006 ). All exper-iments were performed with 6- to 10-week-old mice. Intraperito-neal injection of PR agonist,RU486,was performed to induce Crerecombination. The Institutional Animal Care and Use Committeeof the University of Pennsylvania approved all animal studies.Detailed information regarding mouse experimental design andtreatment are in the Supplemental Information .
IrradiationSix- to eight-week-old mice were subjected to 12 Gy whole-bodygamma-irradiation (Gammacell 40 Cesium 137 Irradiation Unit).Mice were sacrificed 5 days later,and the small intestines were har-vested and fixed for histology. Surviving crypts,crypt length,andvilli height were measured on H&E-stained ileum sections.Twenty-five high-magnification fields were analyzed for eachmouse and a minimum of 100 crypts or villi were measured foreach mouse.
Single-Cell IsolationCrypt single cells were isolated as described previously ( Hamiltonet al.,2015 ). Briefly,the small intestine was opened longitudinallyand washed with cold PBS. Tissue was incubated 20 min on ice inPBS-EDTA-DTT. Tissues were then incubated in PBS-EDTA at 37 C.Crypts were isolated from the epithelial fraction using a 70- mm fil-ter. Crypts were then dissociated to a single-cell suspension usingPBS/dispase at 37 C.
Crypt Isolation and 3D Organoid CultureCrypts were isolated from themouse small intestinal ileum. Tissueswere chilled in Ca 2+-Mg2+-free Hank’s balanced salt solution (CMF-HBSS) with 1mMN-acetyl-cysteine (NAC). Tissues were then incu-bated in CMF-HBSS/1 mM NAC/10 mM EDTA for 45 min at 4 C.Epithelial cells were then dissociated through vortexing/restingcycles. Crypts were separated from epithelial dissociation with a70-mm filter. Crypts were pelleted and resuspended in Basal Media(Advanced DMEM/F12,2 mM Glutamax,10 mM HEPES,1 3 peni-cillin/streptomycin,5 mM CHIR99021 [Cayman Chemical],1 mMNAC, 1 3 N2 Supplement [Gibco], and 1 3 B27 Supplement[Gibco]). Approximately 500 crypts were then embedded in 80%Matrigel/20% ENR (Basal Media;50 ng/mL recombinant mouseepidermal growth factor [R&D Systems] and 1% Noggin/R-Spon-din conditioned media). Enteroids were cultured in ENR.
Fluorescence-Activated Cell SortingCrypt single-cell suspension was prepared from Krt15-CrePR1;R26Tom small intestines as described above. The fluorescence-acti-vated cell sorting sorter Jazz (BD Biosciences) was used to sortTomato+ and Tomato-crypt cells. Sorting was conducted at theUniversity of Pennsylvania Flow Cytometry and Cell Sorting Facil-ity. Cells were sorted in Advanced DMEM/F12 media supple-mented with 1 3 penicillin-streptomycin, 1 3 GlutaMAX, 1 3HEPES,and 10 mM Rock inhibitor Y27632. Cells were then usedfor 3D organoid formation or RNA extraction.Details on mouse experimental design,immunohistochemistry
and immunofluorescence,RNA extraction,qPCR,and statisticalanalyses are available in Supplemental Information .
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental ExperimentalProcedures,four figures,and two tables and can be found withthis article online at https://doi.org/10.1016/j.stemcr.2018.04.022 .
AUTHOR CONTRIBUTIONS
V.G.,K.E.H.,C.J.L.,and A.K.R. designed the study. V.G.,J.S.,andP.C. performed the experiments. B.R. maintained the mouse col-ony. E.P.W. assisted with statistical analysis. A.J.K.-S. assisted withhistology analysis. V.G. and A.K.R. wrote the manuscript. J.S. iscurrently affiliated with University Medicine Greifswald (Depart-ment of Medicine).
ACKNOWLEDGMENTS
We are grateful to members of the Rustgi lab for discussions andcomments on the manuscript. We thank the Molecular Pathology
1956 Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018

10. ORIGINAL PAPER
96
and Imaging Core, Human Microbial and Analytic DepositoryCore, Cell Culture and iPS Core, Genetic and Modified MouseCore, and FACS/sorting Core facilities (University of Pennsylva-nia). We thank Dr. George Cotsarelis (University of Pennsylvania)for Krt15-CrePR1 and Krt15 / mice, Dr. Ben Stanger (University ofPennsylvania) for RosaConfetti mice, and Dr. Christopher Lengner(University of Pennsylvania) for RosaLSL-tdTomato mice. We thankMaryAnn Crissey and Dr. John Lynch for providing Lgr5+ sortedcells. This work was supported by NCI P01-CA098101 (V.G.,A.J.K.-S., and A.K.R.), NIH/National Institute of Diabetes andDigestive and Kidney Diseases (NIDDK) P30-DK050306 Center ofMolecular Studies in Digestive and Liver Diseases, NIH R01-DK056645 (A.K.R.), American Cancer Society (A.K.R.), Fonds de re-cherche en sante´ du Que bec P-Giroux-27692 and P-Giroux-31601(V.G.), NIH NIDDK K01-DK100485 (K.E.H.), and Crohn’s and Co-litis Foundation Career Development Award (K.E.H.).
Received: July 7, 2017Revised: April 23, 2018Accepted: April 24, 2018Published: May 24, 2018
REFERENCES
Asfaha, S., Hayakawa, Y., Muley, A., Stokes, S., Graham, T.A., Erick-sen, R.E., Westphalen, C.B., von Burstin, J., Mastracci, T.L., Worth-ley, D.L.,, et al. (2015). Krt19(+)/Lgr5(-) cells are radioresistant can-cer-initiating stem cells in the colon and intestine. Cell Stem Cell16 , 627–638 .
Barker, N., Ridgway, R.A., van Es, J.H., van de Wetering, M.,Begthel, H., van den Born, M., Danenberg, E., Clarke, A.R., San-som, O.J., and Clevers, H. (2009). Crypt stem cells as the cells-of-origin ofintestinal cancer. Nature 457 , 608–611 .
Barker, N., van Es, J.H., Kuipers, J., Kujala, P., van den Born, M., Co-zijnsen, M., Haegebarth, A., Korving, J., Begthel, H., Peters, P.J.,et al. (2007). Identification of stem cells in small intestine and co-lon by marker gene Lgr5. Nature 449 , 1003–1007 .
Barker, N., van Oudenaarden, A., and Clevers, H. (2012). Identi-fying the stem cell of the intestinal crypt: strategies and pitfalls.Cell Stem Cell 11 , 452–460 .
Barriga, F.M., Montagni, E., Mana, M., Mendez-Lago, M., Her-nando-Momblona, X., Sevillano, M., Guillaumet-Adkins, A., Ro-driguez-Esteban, G., Buczacki, S.J.A., Gut, M., et al. (2017).Mex3a marks a slowly dividing subpopulation of Lgr5+ intestinalstem cells. Cell Stem Cell 20 , 801–816.e7 .
Beumer, J., and Clevers, H. (2016). Regulation and plasticity ofin-testinal stem cells during homeostasis and regeneration. Develop-ment 143 , 3639–3649 .
Bjerknes, M., and Cheng, H. (1999). Clonal analysis of mouse in-testinal epithelial progenitors. Gastroenterology 116 , 7–14 .
Bragulla, H.H., and Homberger, D.G. (2009). Structure and func-tions of keratin proteins in simple, stratified, keratinized and corni-fied epithelia. J. Anat. 214 , 516–559 .
Buczacki, S.J., Zecchini, H.I., Nicholson, A.M., Russell, R., Vermeu-len, L., Kemp, R., and Winton, D.J. (2013). Intestinal label-retain-
ing cells are secretory precursors expressing Lgr5. Nature 495 ,65–69 .
Caulin, C., Ware, C.F., Magin, T.M., and Oshima, R.G. (2000). Ker-atin-dependent, epithelial resistance to tumor necrosis factor-induced apoptosis. J. Cell Biol. 149 , 17–22 .
Davis, H., Irshad, S., Bansal, M., Rafferty, H., Boitsova, T., Bardella,C., Jaeger, E., Lewis, A., Freeman-Mills, L., Giner, F.C., et al. (2015).Aberrant epithelial GREM1 expression initiates colonic tumorigen-esis from cells outside the stem cell niche. Nat. Med. 21 , 62–70 .
Fearon, E.R., and Vogelstein, B. (1990). A genetic model for colo-rectal tumorigenesis. Cell 61 , 759–767 .
Giroux, V., Lento, A.A., Islam, M., Pitarresi, J.R., Kharbanda, A.,Hamilton, K.E., Whelan, K.A., Long, A., Rhoades, B., Tang, Q.,et al. (2017). Long-lived keratin 15+ esophageal progenitor cellscontribute to homeostasis and regeneration. J. Clin. Invest. 127 ,2378–2391 .
Grun, D., Lyubimova, A., Kester, L., Wiebrands, K., Basak, O., Sa-saki, N., Clevers, H., and van Oudenaarden, A. (2015). Single-cellmessenger RNA sequencing reveals rare intestinal cell types. Na-ture 525 , 251–255 .
Hamilton, K.E., Crissey, M.A., Lynch, J.P., and Rustgi, A.K. (2015).Culturing adult stem cells frommouse small intestinal crypts. ColdSpring Harb. Protoc. 2015 , 354–358 .
Hua, G., Thin, T.H., Feldman, R., Haimovitz-Friedman, A., Clevers,H., Fuks, Z., and Kolesnick, R. (2012). Crypt base columnar stemcells in small intestines of mice are radioresistant. Gastroenter-ology 143 , 1266–1276 .
Ito, M., Liu, Y., Yang, Z., Nguyen, J., Liang, F., Morris, R.J., and Cot-sarelis, G. (2005). Stem cells in the hair follicle bulge contribute towound repair but not to homeostasis of the epidermis. Nat. Med.11 , 1351–1354 .
Jadhav, U., Saxena, M., O’Neill, N.K., Saadatpour, A., Yuan, G.C.,Herbert, Z., Murata, K., and Shivdasani, R.A. (2017). Dynamic reor-ganization of chromatin accessibility signatures during dedifferen-tiation of secretory precursors into Lgr5+ intestinal stem cells. CellStem Cell 21 , 65–77.e5 .
Kim, S., Wong, P., and Coulombe, P.A. (2006). A keratin cytoskel-etal protein regulates protein synthesis and epithelial cell growth.Nature 441 , 362–365 .
Kretzschmar, K., and Clevers, H. (2017). Wnt/beta-cateninsignaling in adult mammalian epithelial stem cells. Dev. Biol.428 , 273–282 .
Kuraguchi, M., Wang, X.P., Bronson, R.T., Rothenberg, R., Ohene-Baah, N.Y., Lund, J.J., Kucherlapati, M., Maas, R.L., and Kucherla-pati, R. (2006). Adenomatous polyposis coli (APC) is required fornormal development of skin and thymus. PLoS Genet. 2, e146 .
Li, N., Nakauka-Ddamba, A., Tobias, J., Jensen, S.T., and Lengner,C.J. (2016). Mouse label-retaining cells are molecularly and func-tionally distinct from reserve intestinal stem cells. Gastroenter-ology 151 , 298–310.e7 .
Li, N., Yousefi, M., Nakauka-Ddamba, A., Jain, R., Tobias, J., Ep-stein, J.A., Jensen, S.T., and Lengner, C.J. (2014). Single-cell anal-ysis of proxy reporter allele-marked epithelial cells establishesintestinal stem cell hierarchy. Stem Cell Reports 3 , 876–891 .
Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018 1957

10. ORIGINAL PAPER
97
Li, S., Park, H., Trempus, C.S., Gordon, D., Liu, Y., Cotsarelis, G.,and Morris, R.J. (2013). A keratin 15 containing stem cell popula-tion from the hair follicle contributes to squamous papillomadevelopment in the mouse. Mol. Carcinog. 52 , 751–759 .
Lin, G., Zhang, X., Ren, J., Pang, Z., Wang, C., Xu, N., and Xi, R.(2013). Integrin signaling is required for maintenance and prolifer-ation ofintestinal stem cells in Drosophila . Dev. Biol. 377 , 177–187 .
Metcalfe, C., Kljavin, N.M., Ybarra, R., and de Sauvage, F.J. (2014).Lgr5+ stem cells are indispensable for radiation-induced intestinalregeneration. Cell Stem Cell 14 , 149–159 .
Montgomery, R.K., Carlone, D.L., Richmond, C.A., Farilla, L., Kra-nendonk, M.E., Henderson, D.E., Baffour-Awuah, N.Y., Ambruzs,D.M., Fogli, L.K., Algra, S., et al. (2011). Mouse telomerase reversetranscriptase (mTert) expression marks slowly cycling intestinalstem cells. Proc. Natl. Acad. Sci. USA 108 , 179–184 .
Morris, R.J., Liu, Y., Marles, L., Yang, Z., Trempus, C., Li, S., Lin, J.S.,Sawicki, J.A., and Cotsarelis, G. (2004). Capturing and profilingadult hair follicle stem cells. Nat. Biotechnol. 22 , 411–417 .
Munoz, J., Stange, D.E., Schepers, A.G., van de Wetering, M., Koo,B.K., Itzkovitz, S., Volckmann, R., Kung, K.S., Koster, J., Radulescu,S., et al. (2012). The Lgr5 intestinal stem cell signature: robustexpression of proposed quiescent ’+4’ cell markers. EMBO J. 31 ,3079–3091 .
Muzumdar, M.D., Tasic, B., Miyamichi, K., Li, L., and Luo, L.(2007). A global double-fluorescent cre reporter mouse. Genesis45 , 593–605 .
Potten, C.S., Hume, W.J., Reid, P., and Cairns, J. (1978). The segre-gation of DNA in epithelial stem cells. Cell 15 , 899–906 .
Poulin, E.J., Powell, A.E., Wang, Y., Li, Y., Franklin, J.L., and Coffey,R.J. (2014). Using a new Lrig1 reporter mouse to assess differencesbetween two Lrig1 antibodies in the intestine. Stem Cell Res. 13 ,422–430 .
Powell, A.E., Vlacich, G., Zhao, Z.Y., McKinley, E.T., Washington,M.K., Manning, H.C., and Coffey, R.J. (2014). Inducible loss ofone Apc allele in Lrig1-expressing progenitor cells results inmultiple distal colonic tumors with features offamilial adenoma-tous polyposis. Am. J. Physiol. Gastrointest. Liver Physiol. 307 ,G16–G23 .
Powell, A.E., Wang, Y., Li, Y., Poulin, E.J., Means, A.L., Washington,M.K., Higginbotham, J.N., Juchheim, A., Prasad, N., Levy, S.E., et al.(2012). The pan-ErbB negative regulator Lrig1 is an intestinalstem cell marker that functions as a tumor suppressor. Cell 149 ,146–158 .
Qi, Z., Li, Y., Zhao, B., Xu, C., Liu, Y., Li, H., Zhang, B., Wang, X.,Yang, X., Xie, W., et al. (2017). BMP restricts stemness ofintestinalLgr5(+) stem cells by directly suppressing their signature genes.Nat. Commun. 8, 13824 .
Roche, K.C., Gracz, A.D., Liu, X.F., Newton, V., Akiyama, H., andMagness, S.T. (2015). SOX9 maintains reserve stem cells and pre-serves radioresistance in mouse small intestine. Gastroenterology149 , 1553–1563.e10 .
Rustgi, A.K. (2007). The genetics of hereditary colon cancer. GenesDev. 21 , 2525–2538 .
Saha, S.K., Choi, H.Y., Kim, B.W., Dayem, A.A., Yang, G.M., Kim,K.S., Yin, Y.F., and Cho, S.G. (2017). KRT19 directly interacts
with beta-catenin/RAC1 complex to regulate NUMB-dependentNOTCH signaling pathway and breast cancer properties. Onco-gene 36 , 332–349 .
Sangiorgi, E., and Capecchi, M.R. (2008). Bmi1 is expressed in vivoin intestinal stem cells. Nat. Genet. 40 , 915–920 .
Sato, T., van Es, J.H., Snippert, H.J., Stange, D.E., Vries, R.G., vanden Born, M., Barker, N., Shroyer, N.F., van de Wetering, M., andClevers, H. (2011). Paneth cells constitute the niche for Lgr5stem cells in intestinal crypts. Nature 469 , 415–418 .
Takeda, N., Jain, R., LeBoeuf, M.R., Wang, Q., Lu, M.M., and Ep-stein, J.A. (2011). Interconversion between intestinal stem cellpopulations in distinct niches. Science 334 , 1420–1424 .
Tao, S., Tang, D., Morita, Y., Sperka, T., Omrani, O., Lechel, A., Sakk,V., Kraus, J., Kestler, H.A., Kuhl, M., et al. (2015). Wnt activity andbasal niche position sensitize intestinal stem and progenitor cellsto DNA damage. EMBO J. 34 , 624–640 .
Tetteh, P.W., Basak, O., Farin, H.F., Wiebrands, K., Kretzschmar, K.,Begthel, H., van den Born, M., Korving, J., de Sauvage, F., van Es,J.H., et al. (2016). Replacement oflost Lgr5-positive stem cellsthrough plasticity of their enterocyte-lineage daughters. CellStem Cell 18 , 203–213 .
Tian, H., Biehs, B., Warming, S., Leong, K.G., Rangell, L., Klein,O.D., and de Sauvage, F.J. (2011). A reserve stem cell populationin small intestine renders Lgr5-positive cells dispensable. Nature478 , 255–259 .
van der Flier, L.G., van Gijn, M.E., Hatzis, P., Kujala, P., Haegebarth,A., Stange, D.E., Begthel, H., van den Born, M., Guryev, V., Oving,I., et al. (2009). Transcription factor achaete scute-like 2 controlsintestinal stem cell fate. Cell 136 , 903–912 .
van Es, J.H., Sato, T., van de Wetering, M., Lyubimova, A., Nee,A.N., Gregorieff, A., Sasaki, N., Zeinstra, L., van den Born, M., Korv-ing, J., et al. (2012). Dll1+ secretory progenitor cells revert to stemcells upon crypt damage. Nat. Cell Biol. 14 , 1099–1104 .
Wong, M.H., Stappenbeck, T.S., and Gordon, J.I. (1999). Living andcommuting in intestinal crypts. Gastroenterology 116 , 208–210 .
Wong, V.W., Stange, D.E., Page, M.E., Buczacki, S., Wabik, A., Itami,S., van de Wetering, M., Poulsom, R., Wright, N.A., Trotter, M.W.,et al. (2012). Lrig1 controls intestinal stem-cell homeostasis bynegative regulation of ErbB signalling. Nat. Cell Biol. 14 , 401–408 .
Xian, L., Georgess, D., Huso, T., Cope, L., Belton, A., Chang, Y.T.,Kuang, W., Gu, Q., Zhang, X., Senger, S., et al. (2017). HMGA1 am-plifies Wnt signalling and expands the intestinal stem cellcompartment and paneth cell niche. Nat. Commun. 8, 15008 .
Yan, K.S., Chia, L.A., Li, X., Ootani, A., Su, J., Lee, J.Y., Su, N., Luo,Y., Heilshorn, S.C., Amieva, M.R., et al. (2012). The intestinal stemcell markers Bmi1 and Lgr5 identify two functionally distinct pop-ulations. Proc. Natl. Acad. Sci. USA 109 , 466–471 .
You, J., Zhang, Y., Li, Z., Lou, Z., Jin, L., and Lin, X. (2014).Drosophila perlecan regulates intestinal stem cell activity via cell-matrix attachment. Stem Cell Reports 2, 761–769 .
Yousefi, M., Li, N., Nakauka-Ddamba, A., Wang, S., Davidow, K.,Schoenberger, J., Yu, Z., Jensen, S.T., Kharas, M.G., and Lengner,C.J. (2016). Msi RNA-binding proteins control reserve intestinalstem cell quiescence. J. Cell Biol. 215 , 401–413 .
1958 Stem Cell Reportsj Vol. 10 j 1947–1958 j June 5, 2018

Eidesstattliche Erklärung
98
Eidesstattliche Erklärung
Hiermit erkläre ich, dass ich die vorliegende Dissertation selbständig verfasst und keine anderen
als die angegebenen Hilfsmittel benutzt habe.
Die Dissertation ist bisher keiner anderen Fakultät, keiner anderen wissenschaftlichen Einrichtung
vorgelegt worden.
Ich erkläre, dass ich bisher kein Promotionsverfahren erfolglos beendet habe und dass eine
Aberkennung eines bereits erworbenen Doktorgrades nicht vorliegt.
Datum: Unterschrift:
CURRICULUM VITAE
99
CURRICULUM VITAE
Personal data:
Name Julien Orlando STEPHAN
Primary Address Baustraße 12, 17489 Greifswald (Germany)
Telephone +49 17661314872
Email [email protected]
Birthday and -place 1st June 1991 in Bethesda, Maryland (USA)
Nationality USA, Germany, France
School career:
08/1997 – 07/1998 Primary School Bogenstraße, Solingen (Germany)
08/1998 – 07/2001 Primary School Gräfin-Imma, Bochum (Germany)
08/2001 – 07/2007 Gymnasium Schiller-Schule, Bochum (Germany)
07/2007 – 07/2008 Collegio Loretto, La Paz (Bolivia)
08/2008 – 06/2010 Gymnasium John-Lennon, Berlin (Germany)
Abitur 2010 Graduation score: 1.9 focuses on Math and Biology (US equivalent: high school + one year)
University career:
10/2011 – 11/2018 School of Medicine, Universität Greifswald (Germany)
10/2014 – 10/2015 School of Medicine, University of Pennsylvania (USA)
Received scholarship:
08/2007 – 07/2008 Rotary Scholarship, La Paz (Bolivia)
International experiences:
08/2007 – 07/2008 Rotary Scholarship, La Paz (Bolivia)
08/2011 – 11/2011 School of Medicine, Université de Nantes (France)
10/2014 – 11/2015 School of Medicine, University of Pennsylvania (USA)

CURRICULUM VITAE
100
Laboratory experience:
02/2014 – 10/2014 Department of Gastroenterology, University of Greifswald (Germany)
10/2014 – 11/2015 Department of Gastroenterology, University of Pennsylvania (USA)
Publication:
Giroux, V., Stephan, J., Chatterji, P., Rhoades, B., Wileyto, E.P., Klein-Szanto, A.J., Lengner,
C.J., Hamilton, K.E., and Rustgi, A.K. (2018). Mouse Intestinal Krt15+ Crypt Cells Are Radio-
Resistant and Tumor Initiating. Stem Cell Rep.
Volunteering:
08/2007 – 07/2008 Social worker in humanitarian affairs, La Paz (Bolivia)
10/2010 – 10/2011 Vivantes Hospital, Berlin (Germany)
Work experiences:
02/2012 – 06/2014 Schlothauer & Wauer (Germany & Austria)
- Student assistant in civil engineering
02/2014 – 10/2014 Department of gastroenterology, University of Greifswald
- Laboratory technician
05/2016 – 03/2017 Department of pharmacology, University of Greifswald
- Medical assistant in clinical trials
since 01/2019 Department of gynaecology, University of Greifswald
- Resident physician
Language skills:
German fluent, mother tongue
French fluent, mother tongue
English fluent
Spanish very good working knowledge
Datum: Unterschrift:

DANKSAGUNG
101
DANKSAGUNG Dank gebührt Prof. Dr. Markus. M. Lerch und den Mitgliedern der wissenschaftlichen
Arbeitsgruppe der Inneren Medizin A für meine Einarbeitung in die erforderlichen
Labormethoden, meiner kontinuierlichen Unterstützung, wie auch meiner Entsendung in die
Arbeitsgruppe von Dr. Anil K. Rustgi. Desweiteren danke ich Dr. Anil K. Rustgi und PhD
Veronique Giroux, sowie allen Mitgliedern des Rustgi Labors für die wissenschaftliche Anleitung
und stetige Begleitung dieser Experimente. Darüber hinaus danke ich den Mitarbeiter dem
Molecular Pathology and Imaging Core, Human Microbial Analytic Depository Core, Cell Culture
and iPS Core, Genetic, sowie dem Modified Mouse Core der University of Pennsylvania.
Besonderer Dank gilt Dr. George Cotsarelis (University of Pennsylvania) für die Bereitstellung
von Krt15-CrePR1 und Krt15-/- Mäusen, wie auch Dr. Christopher Lengner (University of
Pennsylvania) für RosaLSL-tdTomato Mäuse.
Top Related
Copyright © 2022 FDOKUMEN

