






Bahasa
Halaman
Hukum


Journal of Fish Biology (1999) 54, 267–286Article No. jfbi.1998.0863, available online at http://www.idealibrary.com on
The North Mediterranean Barbus lineage: phylogenetichypotheses and taxonomic implications based on
allozyme data
C. S. T*†‡, Y. K† P. B**Laboratoire Genome et Populations, UPR 9060, Universite Montpellier II, CC063,
place E. Bataillon, 34095 Montpellier Cedex 05, France†Department of Genetics, School of Biology, Aristotle University of Thessaloniki,
54006 Thessaloniki, Greece
(Received 6 April 1998, Accepted 18 September 1998)
On the basis of distinct ecological and morphological characters, the European Barbus taxahave been clustered in two groups: a fluvio-lacustrine and a rheophilic or strictly riverine one.These two groups (or ecophenotypes) were recognized in different parts of Europe, and formedeither a species assemblage (Barbus barbus group) or a polytypic species (Barbus meridionalis).The hypothesis was that species of the same group belong to the same phylogenetic lineage(clade) and are the result of the same transcontinental colonization event. The analysis, usingallozyme markers, of 10 taxa of the genus Barbus from France, Italy, Greece, the CzechRepublic and Slovakia, showed that the taxa thought to belong to the fluvio-lacustrine and therheophilic groups are not monophyletic. The results suggest that probably in each sub-region,the founding taxon has diverged independently to form species of two different ecophenotypes,one occupying the upstream rivers and the other the lowland rivers. Accordingly, Barbusspecies groups represent clusters of morphologically convergent taxa living in equivalentbiotopes.
? 1999 The Fisheries Society of the British Isles
Key words: Barbus; allozymes; species groups; phylogeny; convergence.
‡Author to whom correspondence should be addressed. Tel.: +33 467 14 37 33; fax: +33 467 14 45 54;email: [email protected]
Deceased November 1995.
INTRODUCTION
The North Mediterranean region is an area where endemism occurs frequently.According to Crivelli & Maitland (1995), 229 endemic fish taxa belonging to 13families have been identified. The genus Barbus Cuvier & Cloquet 1816, istaxonomically very complex comprising at least 800 species (Howes, 1987),spread over three continents (Europe, Asia and Africa) and is one of the mostdiversified members of the Cyprinidae. It also includes diploid, tetraploid andhexaploid species, therefore complicating a standard taxonomy. The taxagrouped in the genus Barbus exhibit very different phenetic features and originsbecause of the poor understanding of the limits between genera in the barbelledcyprinids and the subsequent addition of species with a doubtful genericallocation (Myers, 1960; Doadrio, 1990).
Taxonomists are aware of the necessity to revise the genus. Berrebi et al.(1996) proposed that Barbus sensu stricto is composed of tetraploid species
267
0022–1112/99/020267+20 $30.00/0 ? 1999 The Fisheries Society of the British Isles

268 . . .
(2n=100 chromosomes), that live mainly in the Euro-Mediterranean region, andshare the diagnostic morphological characters defined by Howes (1987) andDoadrio (1990), as well as similar karyotypes, biochemical markers and parasiticfauna. This group is found in Europe, Western Asia and North Africa (fromTunisia to Morocco).
The systematics of Barbus sensu stricto in Europe, prior to 1990, was based onmorphological data and geographical distribution. It was suggested that theEuropean (or ‘ Siberian ’) branch is of Asian origin and reached Europe in theMiddle Miocene (Darlington, 1948; Banarescu, 1973). According to Bianco(1989, 1990), the first penetration of Barbus species in peri-Mediterranean rivers(along with other primary or primary-like freshwater fish) originated from theParatethys and occurred during the freshwater Messinian-Lago Mare phase ofthe Mediterranean (5·5 million years ago), while the present-day distribution ofthese species in Europe is the result of river captures, especially in epicontinentalareas, during glacial marine regressions of the Pleistocene.
Recent studies on Barbus species have included finer discrimination tools, suchas internal osteological characters and molecular markers. Based on the shape ofpharyngeal teeth, Doadrio (1990) proposed that the Spanish and North-WestAfrican species form a group. Subsequently, the allozyme study of Machordom(1990) and Machordom et al. (1995), focused on the Iberian species, confirmedthis grouping of West Mediterranean species. More recent studies have pro-posed species status for B. peloponnesius Valenciennes, 1842, based on morpho-logical (Karakousis et al., 1993) and genetic data (Karakousis et al., 1995;Tsigenopoulos, 1996; Berrebi et al., unpubl.) and revealed differentiation of itspopulations in Greece. However, no complete global analysis is available for theEuropean Barbus species.
From an ecological point of view, the main trait of the distribution of barbs inEurope is the existence of two main ecophenotypes. A rheophilic or strictlyriverine one, which consists of small species (maximum total length LT=30 cm),that frequent mountain streams and the more turbulent stretches of large rivers,and a fluvio-lacustrine one which comprises larger species (LT>50 cm), prefer-ring wide calm rivers, with a regular current, running through the Europeanplains, and sometimes lakes (Economidis, 1989).
The rheophilic barbs are found in all the mountain tributaries of the NorthMediterranean region and they are characterized by speckled large black scaleson the body and a long anal fin which extends further back than the root of thecaudal fin. In this type of species, based mainly on the structure of the last(fourth) soft dorsal ray, the number and size of the lateral line scales, we candistinguish two groups: the meridionalis with an unserrated ray and the cyclolepiswhich has a moderately serrated ray, and larger number of smaller scales on thelateral line (Karaman, 1971). The fluvio-lacustrine species are present in almostall the European rivers and are characterized by a spindle shaped body, smallscales, short anal fin (which does not reach the root of the caudal fin) and atriangular dorsal fin strengthened with an ossified and generally serratedunbranched ray.
Species of the same ecophenotype, when occurring in the same river areallopatric, e.g. rheophilic B. peloponnesius and B. cyclolepis Heckel, 1937 in theAxios River in Greece, while rheophilic and fluvio-lacustrine species may be

269
present in sympatry or even syntopy, exploiting different parts of the river(Economidis, 1989; Economidis & Banarescu, 1991).
Until now, ichthyologists have assumed that within each group, barbs areclosely related phylogenetically, and this explains why the sub-species statuswas attributed to taxa of the same ecophenotype. Barbus meridionalis Risso1826 has been treated as a polytypic species, with sub-species (B. m. meridionalis,B. m. caninus, B. m. peloponnesius and B. m. petenyi, from France, Italy,Greece and Danubian tributaries, respectively) sharing the same generalmorphology (Almaca, 1984). Karakousis et al. (1995) in an allozyme study ofsome species within this group, placed B. petenyi Heckel, 1852c as a subspeciesof B. peloponnesius, supporting the proposal of Almaca (1984). However,Berrebi et al. (unpubl.) and Tsigenopoulos (1996) indicated the distinctivenessof B. petenyi and B. caninus Bonaparte, 1839 populations, and suggested thatadditional information was required, before definitive conclusions could bereached.
In the fluvio-lacustrine group, generally a species status was attributed todifferent taxa, e.g. B. barbus L., 1758, B. macedonicus Karaman, 1928 and B.plebejus Bonaparte, 1839 (Kottelat, 1997). A common origin for all the speciesbelonging to the fluvio-lacustrine group has not been proposed explicitly, butoften is accepted implicitly (Berrebi et al., unpubl.).
Molecular techniques have proved to be extremely useful in resolving a varietyof systematic questions (Avise, 1994). Under the phylogenetic species concept(Cracraft, 1983), it is possible to carry out systematic analyses using molecularmarkers. In this way, complex species groups which are difficult to distinguishusing morphological data, can be determined by reconstructing molecularphylogenies.
The main objectives of this study are to elucidate the phylogenetic relation-ships among North Mediterranean Barbus species using allozyme data and toinvestigate the assumed monophyly of the rheophilic (or strictly riverine) andfluvio-lacustrine groups.
MATERIALS AND METHODS
SAMPLES AND TISSUE PREPARATIONFifteen populations belonging to 10 morphologically diagnosed taxa of Barbus sensu
stricto, were collected in Greece, Slovakia, the Czech Republic, Italy and France (Fig. 1,Table I). For the fluvio-lacustrine group we used one population of B. plebejus fromItaly, two populations of B. barbus from France and Czech Republic, and threepopulations of B. macedonicus from Greece. For the rheophilic group we used popu-lations of B. meridionalis, B. caninus, B. petenyi, B. peloponnesius, B. cyclolepis and B.prespensis Karaman, 1924. Barbus prespensis is the only species of this genus in Europethat we find mainly in lakes. However, it belongs to the cyclolepis group (Almaca, 1984;Economidis, 1989) and so this study treats it as a rheophilic barb.
Fishes were captured by electrofishing and transported to the laboratory either alive orin ice boxes. Protein extracts were obtained from both muscle and liver. Tissues werehomogenized with a 1 : 2 volume dilution in extraction buffer (0·01 Tris, 0·01 EDTA,pH 6·8). After the addition of 0·5 ml CCl4 in each tube, they were centrifuged at 13 000 gat 4) C for 20 min. The translucent supernatant was extracted and used for starch gelelectrophoresis.
F
.1.
Sam
plin
gsi
tes
ofth
epo
pula
tion
sex
amin
ed.
1,A
ubay
gue
(Her
ault
);2,
Dro
me;
3,Iu
drio
(Iso
nzo)
;4,
Osl
ava
(Dan
ube)
;5,
Tor
ysa
(Dan
ube)
;6,
Pre
spa
Lak
e;7,
Axi
os;
8,A
liakm
on.

T
I.L
isti
ngof
sam
ples
used
inth
est
udy,
thei
rca
ptur
elo
calit
ies,
thei
rec
ophe
noty
pe,w
ith
info
rmat
ion
abou
tthe
pres
ence
ofot
her
Bar
bus
spec
ies
inth
esa
me
basi
nan
dsa
mpl
esi
ze
Spec
ies
Loc
alit
yE
coph
enot
ype
Cou
ntry
Rem
arks
Sam
ple
size
1B
.pe
lopo
nnes
ius
(1)
Axi
osR
.R
heop
hilic
orst
rict
lyri
veri
neG
reec
eN
oot
her
barb
spec
ies
insy
mpa
try
302
B.
pelo
ponn
esiu
s(2
)A
xios
R.
Rhe
ophi
licor
stri
ctly
rive
rine
Gre
ece
Insy
mpa
try
wit
hB
.m
aced
onic
us27
3B
.pe
lopo
nnes
ius
(3)
Alia
kmon
R.
Rhe
ophi
licor
stri
ctly
rive
rine
Gre
ece
No
othe
rba
rbsp
ecie
sin
sym
patr
y20
4B
.pe
lopo
nnes
ius
(4)
Alia
kmon
R.
Rhe
ophi
licor
stri
ctly
rive
rine
Gre
ece
Insy
mpa
try
wit
hB
.m
aced
onic
us20
5B
.m
erid
iona
lisH
erau
ltR
.R
heop
hilic
orst
rict
lyri
veri
neF
ranc
eN
oot
her
barb
spec
ies
insy
mpa
try
106
B.
pete
nyi
Tor
ysa
R.
Rhe
ophi
licor
stri
ctly
rive
rine
Slov
akia
No
othe
rba
rbsp
ecie
sin
sym
patr
y11
7B
.ca
ninu
sIs
onzo
R.
Rhe
ophi
licor
stri
ctly
rive
rine
Ital
yN
oot
her
barb
spec
ies
insy
mpa
try
108
B.
cycl
olep
isA
xios
R.
Rhe
ophi
licor
stri
ctly
rive
rine
Gre
ece
No
othe
rba
rbsp
ecie
sin
sym
patr
y28
9B
.pr
espe
nsis
Pre
spa
Lak
eL
acus
trin
eG
reec
eN
oot
her
barb
spec
ies
insy
mpa
try
22
10B
.m
aced
onic
us(1
)A
xios
R.
Flu
vio-
lacu
stri
neG
reec
eN
oot
her
barb
spec
ies
insy
mpa
try
1111
B.
mac
edon
icus
(2)
Axi
osR
.F
luvi
o-la
cust
rine
Gre
ece
Insy
mpa
try
wit
hB
.pe
lopo
nnes
ius
2012
B.
mac
edon
icus
(3)
Alia
kmon
R.
Flu
vio-
lacu
stri
neG
reec
eIn
sym
patr
yw
ith
B.
pelo
ponn
esiu
s20
13B
.ba
rbus
Dro
me
R.
Flu
vio-
lacu
stri
neF
ranc
eN
oot
her
barb
spec
ies
insy
mpa
try
914
B.
barb
usO
slav
aR
.F
luvi
o-la
cust
rine
Cze
chR
ep.
No
othe
rba
rbsp
ecie
sin
sym
patr
y13
15B
.pl
ebej
usIs
onzo
R.
Flu
vio-
lacu
stri
neIt
aly
No
othe
rba
rbsp
ecie
sin
sym
patr
y14

272 . . .
STARCH GEL ELECTROPHORESISA total of 265 fishes were examined with 11 enzymatic systems (Table II). Methods
and enzyme staining reactions were employed as described in Pasteur et al. (1987) andMachordom et al. (1995).
The electrophoretic mobility of each allele in every enzymatic system was comparedwith the alleles of B. meridionalis from France, which were taken to be 100, following thestudy of Machordom et al. (1995). Nevertheless, alleles described in this study must notbe confused with alleles mentioned in the article cited above.
CODING TETRAPLOID GENOTYPESAll the European species of the genus Barbus which have been karyotyped are
tetraploids (Collares-Pereira, 1994). The tetraploidization event is ancestral and theinitial tetraploidy evolved, gene by gene, towards functional diploidy where all thechromosomes now work as pairs (M. Castelli, pers. comm.). The species are thusfunctional diploids and ancestral tetraploids. The functional diploidization is a longprocess that destroys the expression of one of the two similar genes by conservation (oraccumulation) of deleterious mutations (Ohno, 1970).
However, for an enzymatic system of genes, the locus that disappeared in a duplicatedpair, may be different in each species analysed. The number of loci, and consequently thenumber of variables for statistical analyses can differ between species. Thus, a specificcoding method must be used. The method is based on the principle that all enzymatic locihave been duplicated and that, when silent, a fixed null allele occurs (Berrebi et al., 1990).The only problem is when two samples share a null allele for a given locus. Here we usedthe maximizing method between species (or sub-species), in which the null alleles areassumed to be of different origin in each taxon, and the minimizing method betweenpopulations of the same taxon, where the null allele is the same shared allele amongpopulations. The minimizing method was in this way limited to the samples of B. barbusfrom France and the Czech Republic and the populations of B. macedonicus andB. peloponnesius from Greece.
DATA ANALYSESIn order to estimate the phylogenetic relationships among the taxa, we applied distance
and parsimony methods to construct phylogenetic trees.
T II. Enzyme systems analyses conditions (tissues and buffers); for enzyme abbrevi-ations see Pasteur et al. (1987)
TissuesBuffers
TC 6.7 TCB 8.7 TME 6.9
Muscle AAT*, AK*, CK*,IDH*, MDH*, PT*
GPI*, LDH*, PGM* EST*
Liver MDH*, SDH* GPI*, LDH* EST*
Distance methodsGene frequencies in populations were estimated using the computer package
GENETIX (Belkhir et al., 1996). Distance measures and all the analyses for treereconstruction were performed with the PHYLIP package (Felsenstein, 1993). In thisstudy we calculated Nei’s (1972) genetic distance using the program GENDIST. Theformula for this distance assumes that changes in allele frequencies occurred via mutationor genetic drift, while the population size remained constant for all populations.
A tree was constructed using these distance estimates with the tree building algorithmneighbour joining (Saitou & Nei, 1987), using the program NEIGHBOR. This method

273
does not assume constant and equal rates of change among lineages (molecular clock),therefore leaving branch lengths unconstrained. We also applied bootstrapping (1000replicates), to have a measure of the support of the branching patterns.
T III. Number of loci per enzymatic system in the tissues used; numbers inparentheses indicate loci shared in the two tissues (to be counted once only)
AAT* AK* CK* EST* GPI* IDH* LDH* MDH* PGM* PT* SDH*
Muscle 3 2 1 (2) 3+(2) 1 (2) 3+(1) 2 1Liver (2) (2) 1+(2) 2+(1) 1
Parsimony methodsIn the tree reconstruction we applied two methods: Wagner’s maximum parsimony
(Kluge & Farris, 1969) and Dollo’s parisomy methods (LeQuesne, 1972; Farris, 1977)using the PHYLIP software (executables MIX and DOLLOP, respectively). In all cases,trees were treated as unrooted without defining any taxon as outgroup.
Based on the presence/absence of each allele in a given taxon, reversion andconvergence are considered equally probable in the Wagner method, while convergence isprohibited in Dollo’s method. The last method is thought to be more realistic, and wellsuited for studies based on allozymic data, where the probability of the independentemergence (convergence, mainly via mutation) of a given character (allele) in differenttaxa will usually be negligible (Bonhomme et al., 1984). In contrast, an allele candisappear (either by genetic drift or by selection) in the same or even differentevolutionary lineages (inducing reversions) with a much higher probability.
The algorithms for these two methods use matrices of coded characters: (1) forpresence and (0) for absence of the character. We have used the matrix constructed underthe assumption that an allele is present in a population when its frequency is higher than5%, in order to avoid errors due to the small size of certain samples (where alleles of lowfrequency have little chance of being present). This matrix was treated further with theexecutable BOOT of the PHYLIP package to generate 1000 new matrices that permittedus to obtain bootstrap values in every node of the topology.
In order to investigate the monophyly of the rheophilic species (B. meridionalis, B.petenyi, B. peloponnesius, B. cyclolepis and B. prespensis), we used the constraint optionin PAUP version 3.1.1. (Swofford, 1993). By comparing the number of events accordingto each constraint, it is possible to estimate the cost of each hypothesis in terms ofparsimony, and thus its probability.
RESULTS
The 11 enzymatic systems analysed in this study revealed a total of 27presumptive loci (Table III) and a total of 132 alleles (Table IV). Only three loci(AK-2*, IDH* and MDH-1*) were monomorphic in all populations examined(loci not shown in Table IV). The most polymorphic loci were MDH-3* with 11alleles and MDH-5* with eight alleles. The B. meridionalis population presentsonly one allele in each locus, and the average number of alleles per locus has itsmaximal value (A=1·76) in the B. peloponnesius population from the AliakmonRiver (Table IV). In the population of the latter species we find also the highestproportion of polymorphic loci (P=52%) and of heterozygotes (H=0·140).
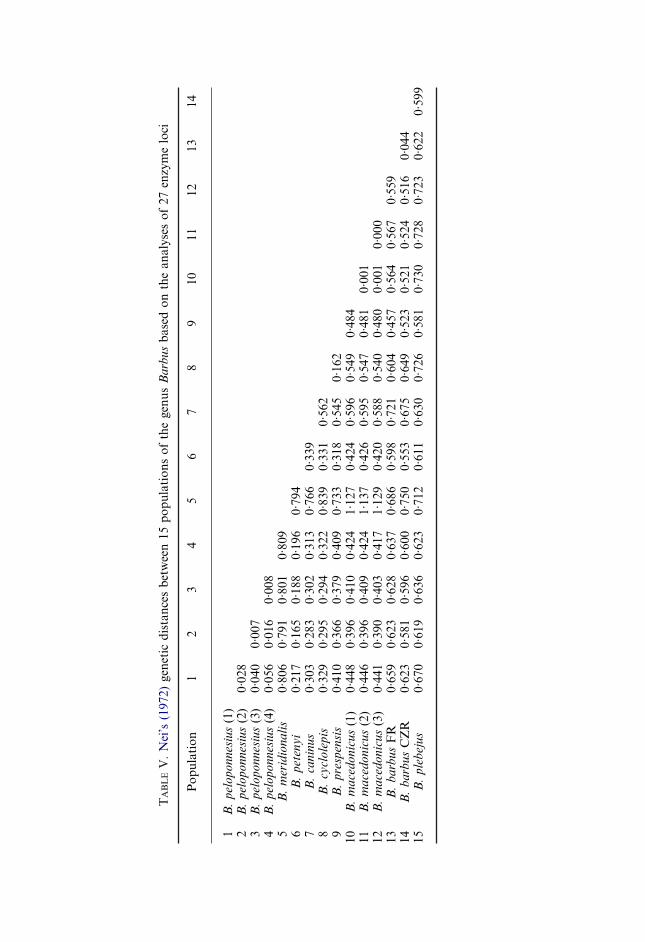
Nei’s (1972) genetic distances between populations of the same species rangefrom a minimum of 0·0003 (between two of B. macedonicus populations) to a

T
IV.
Alle
licfr
eque
ncie
sin
the
15po
pula
tion
san
alys
ed,
expe
cted
hete
rozy
gosi
ty(H
),pr
opor
tion
ofpo
lym
orph
iclo
ci(P
)an
dm
ean
num
ber
ofal
lele
spe
rlo
cus
(A);
the
num
bers
for
popu
lati
ons
corr
espo
ndto
thos
eof
Tab
leI;
AK
-2*,
IDH
*an
dM
DH
-1*
wer
em
onom
orph
icin
all
popu
lati
ons
exam
ined
;nu
llal
lele
sar
em
arke
dw
ith
anas
teri
sk(*
)
Loc
us1
23
45
67
89
1011
1213
1415
AA
T-1
*82
——
——
——
——
—0·
250·
130·
13—
——
981·
001·
001·
001·
00—
1·00
1·00
1·00
1·00
0·75
0·88
0·88
1·00
1·00
1·00
100
——
——
1·00
——
——
——
——
——
AA
T-2
*1*
0·03
——
——
——
——
——
——
——
920·
971·
001·
001·
00—
1·00
1·00
1·00
1·00
1·00
1·00
1·00
1·00
1·00
1·00
100
——
——
1·00
——
——
——
——
——
AA
T-3
*65
——
——
——
——
——
——
——
0·21
75—
——
——
——
——
——
——
—0·
7910
01·
001·
001·
001·
001·
001·
001·
001·
001·
001·
001·
001·
001·
001·
00—
AK
-1*
1*—
——
—1·
00—
——
——
——
——
—10
01·
001·
001·
001·
00—
1·00
1·00
1·00
1·00
1·00
1·00
1·00
1·00
1·00
—24
0—
——
——
——
——
——
——
—1·
00C
K*
691·
001·
001·
001·
00—
1·00
1·00
1·00
1·00
——
——
——
100
——
——
1·00
——
——
1·00
1·00
1·00
1·00
1·00
1·00
ES
T-1
*87
0·50
0·31
0·33
0·23
——
——
—0·
951·
000·
95—
——
93—
——
——
—1·
00—
——
——
——
—96
0·50
0·69
0·68
0·78
—1·
00—
1·00
—0·
05—
0·05
—0·
050·
1110
0—
——
—1·
00—
——
1·00
——
—1·
000·
55—
103
——
——
——
——
——
——
—0·
410·
89

T
IV.
Con
tinu
ed
Loc
us1
23
45
67
89
1011
1213
1415
ES
T-3
*94
1·00
0·89
1·00
0·98
——
0·35
1·00
——
—0·
03—
——
100
——
——
1·00
——
——
——
——
——
102
—0·
11—
0·03
—1·
000·
30—
1·00
1·00
1·00
0·98
0·83
0·85
1·00
110
——
——
——
0·35
——
——
—0·
170·
05—
GP
I-1*
1*—
——
——
——
—0·
36—
——
——
—2*
——
——
——
0·10
——
——
——
4*—
——
——
——
——
——
0·33
0·08
—10
01·
001·
000·
950·
951·
001·
000·
901·
000·
641·
001·
001·
000·
670·
921·
0020
0—
—0·
050·
05—
——
——
——
——
——
GP
I-2*
1*0·
470·
220·
210·
10—
——
——
——
——
——
2*—
——
——
1·00
——
——
——
——
3*—
——
——
—1·
00—
——
——
——
4*—
——
——
——
——
——
0·22
0·15
—10
0—
——
—1·
00—
——
——
——
——
—10
30·
380·
630·
710·
90—
——
——
1·00
1·00
1·00
0·78
0·85
1·00
110
0·15
0·15
0·08
——
——
1·00
1·00
——
——
——
GP
I-3*
1*—
——
——
——
—0·
05—
——
——
—10
01·
000·
941·
000·
951·
001·
000·
251·
000·
95—
——
1·00
0·92
—10
3—
0·04
—0·
03—
——
——
——
——
——
106
—0·
02—
——
—0·
75—
——
——
——
0·61
112
——
—0·
03—
——
——
1·00
1·00
1·00
—0·
080·
1811
7—
——
——
——
——
——
——
—0·
21

T
IV.
Con
tinu
ed
Loc
us1
23
45
67
89
1011
1213
1415
GP
I-4*
1*0·
470·
300·
300·
05—
——
——
——
2*—
——
1·00
——
——
——
3*—
——
——
—0·
82—
——
——
—4*
——
——
—0·
40—
——
——
5*—
——
——
——
——
1·00
1·00
0·95
—6*
——
——
——
——
——
——
0·44
0·23
—81
0·05
0·07
0·03
0·18
——
——
——
——
——
—10
00·
330·
560·
500·
551·
000·
601·
00—
0·18
——
0·05
0·56
0·77
1·00
118
0·15
0·07
0·18
0·23
——
——
——
——
——
—G
PI-
5*1*
0·27
0·11
0·30
0·55
——
——
——
——
——
—2*
——
——
1·00
——
——
——
——
——
3*—
——
——
0·27
——
——
——
——
4*—
——
——
0·40
——
——
——
——
5*—
——
——
——
——
——
—0·
08—
6*—
——
——
——
——
——
——
—1·
0010
00·
600·
670·
680·
35—
0·73
0·60
1·00
1·00
1·00
1·00
1·00
1·00
0·92
—10
70·
130·
220·
030·
10—
——
——
——
——
——
LD
H-1
*81
——
0·03
0·03
——
——
——
——
——
—10
01·
000·
930·
950·
931·
000·
941·
001·
001·
001·
001·
001·
001·
001·
001·
0014
5—
——
0·03
——
——
——
——
——
—18
5—
0·07
0·03
0·03
——
——
——
——
——
—19
0—
——
——
0·06
——
——
——
——
—L
DH
-2*
10—
——
0·03
——
——
——
——
——
—10
01·
001·
001·
000·
981·
001·
001·
001·
001·
001·
001·
001·
001·
001·
001·
00

T
IV.
Con
tinu
ed
Loc
us1
23
45
67
89
1011
1213
1415
LD
H-3
*5
——
—0·
10—
——
——
——
—1·
000·
96—
83—
—0·
05—
——
—0·
961·
001·
001·
001·
00—
——
100
1·00
1·00
0·95
0·90
1·00
1·00
1·00
0·04
——
——
—0·
041·
00M
DH
-2*
95—
——
——
——
——
——
—1·
001·
00—
100
——
——
1·00
——
——
——
——
——
120
1·00
1·00
1·00
1·00
—1·
001·
001·
001·
001·
001·
001·
00—
—1·
00M
DH
-3*
1*—
0·04
——
——
——
——
——
——
—59
0·13
0·63
0·98
0·95
——
——
——
——
——
—65
0·27
0·11
——
——
——
——
——
——
—76
——
——
——
——
——
——
——
1·00
78—
——
——
——
1·00
1·00
——
——
——
82—
——
——
1·00
——
——
——
——
—93
——
—0·
05—
——
——
——
——
——
96—
——
——
——
——
——
—1·
001·
00—
100
——
——
1·00
—1·
00—
——
——
——
—11
00·
600·
220·
03—
——
——
——
——
——
—11
2—
——
——
——
——
1·00
1·00
1·00
——
—M
DH
-4*
1*—
——
——
——
——
——
—1·
001·
00—
100
——
——
1·00
——
——
——
——
——
106
1·00
1·00
1·00
1·00
—1·
00—
1·00
1·00
1·00
1·00
1·00
——
—11
6—
——
——
——
——
——
——
—1·
0013
0—
——
——
—1·
00—
——
——
——
—

T
IV.
Con
tinu
ed
Loc
us1
23
45
67
89
1011
1213
1415
MD
H-5
*1*
——
——
—0·
09—
——
——
——
——
741·
001·
000·
900·
93—
——
——
——
——
——
86—
——
——
0·91
——
——
——
——
—93
——
——
——
1·00
——
——
——
——
100
——
0·10
0·08
1·00
——
1·00
1·00
——
—0·
94—
1·00
103
——
——
——
——
——
——
—0·
73—
109
——
——
——
——
—1·
001·
001·
00—
——
113
——
——
——
——
——
——
0·06
0·27
—M
DH
-6*
1*0·
870·
350·
420·
38—
——
——
——
——
——
2*—
——
——
——
0·04
——
——
——
—3*
——
——
1·00
——
——
——
——
—4*
——
——
—1·
00—
——
——
——
—5*
——
——
——
——
——
——
1·00
1·00
—6*
——
——
——
——
——
——
—1·
0010
00·
130·
650·
580·
62—
1·00
—0·
961·
001·
001·
001·
00—
——
PG
M-1
*90
0·03
—0·
05—
——
——
—0·
410·
500·
48—
——
100
0·97
1·00
0·93
1·00
1·00
1·00
1·00
0·43
0·82
0·59
0·50
0·53
1·00
1·00
0·96
104
——
——
——
—0·
11—
——
——
——
108
——
——
——
—0·
46—
——
——
——
110
——
——
——
——
0·18
——
——
——
115
——
——
——
——
——
——
——
0·04
116
——
0·03
——
——
——
——
——
——

T
IV.
Con
tinu
ed
Loc
us1
23
45
67
89
1011
1213
1415
PG
M-2
*1*
0·03
—0·
05—
——
——
——
——
——
—2*
——
——
——
—0·
18—
——
——
——
3*—
——
——
——
—0·
14—
——
——
—4*
——
——
——
——
——
——
1·00
0·89
—54
——
——
——
——
0·14
——
——
——
63—
——
——
——
0·82
——
——
——
—65
——
——
——
——
——
——
—0·
11—
75—
——
——
——
—0·
73—
——
——
1·00
840·
971·
000·
951·
00—
1·00
1·00
——
1·00
1·00
1·00
——
—10
0—
——
—1·
00—
——
——
——
——
—P
T-3
*10
01·
001·
001·
001·
001·
000·
931·
001·
001·
00—
0·04
——
—1·
0020
0—
——
——
0·07
——
—1·
000·
961·
001·
001·
00—
SD
H*
100
——
——
1·00
0·33
—0·
961·
00—
——
1·00
1·00
1·00
250
1·00
1·00
1·00
1·00
—0·
671·
000·
04—
1·00
1·00
1·00
——
—(H
)0·
134
0·14
00·
128
0·12
10·
000
0·06
90·
068
0·04
40·
063
0·03
80·
031
0·03
80·
067
0·09
40·
046
(P)
0·24
0·36
0·44
0·44
0·00
0·24
0·16
0·08
0·16
0·08
0·08
0·16
0·20
0·36
0·16
(A)
1·60
1·64
1·72
1·76
1·00
1·24
1·20
1·24
1·24
1·12
1·12
1·20
1·20
1·44
1·20

280 . . .
PHYLOGENYThe tree constructed by the neighbour joining method and based on Nei’s
distances (Fig. 2) shows three principal characteristics: (i) the monophyly of B.cyclolepis and B. prespensis, supported by a relatively high bootstrap value(77%), (ii) the high divergence between the three fluvio-lacustrine species(B. barbus, B. plebejus and B. macedonicus), that do not form a monophyleticgroup and (iii) the close relationship between B. peloponnesius and the rheophilicspecies from Italy (B. caninus) and Slovakia (B. petenyi) that form a poorlysupported monophyletic group, while B. meridionalis occupies a basal position.
Using the Wagner parsimony method, we obtained two most parsimonioustrees that require 102 steps. These two trees differ only in the clustering of twoof the four B. peloponnesius populations. In Fig. 3, we present the unrootedconsensus tree obtained after 1000 bootstraps of the matrix with the codedcharacters. The constraint’s option in PAUP’s parsimony, shows that thegrouping of the strictly rheophilic species is the least probable, requiring 109steps, i.e. seven steps more.
With the Dollo parsimony method the consensus tree (of two most parsi-monious trees) is very comparable to that of Wagner’s parsimony (Fig. 3), inthat the populations B. peloponnesius and B. macedonicus form a monophy-letic group. The only difference is that B. caninus and B. petenyi populationsare inserted between the branches leading to B. barbus and B. meridionalispopulations (tree not shown).
The major difference between the consensus trees obtained using the twoparsimony methods and the neighbor joining tree (Fig. 2), is that in theparsimony trees B. macedonicus forms a poorly supported monophyletic groupwith B. peloponnesius (bootstrap value in the order of 32% in the maximumparsimony tree). Bootstrap supports for the nodes are generally poor and therobust nodes (>85%) are mainly for populations belonging to the same species,i.e. for B. barbus, B. macedonicus and B. peloponnesius.
DISCUSSION
TAXONOMIC AND EVOLUTIONARY INFERENCESThe French species B. meridionalis is generally very different from the other
rheophilic species of the meridionalis group (B. peloponnesius, B. petenyi and
maximum of 0·056 (between the populations of B. peloponnesius) (Table V).However, genetic distances between populations belonging to different speciesvary from 0·162 (between B. prespensis and B. cyclolepis) to 1·137 (betweenB. macedonicus and B. meridionalis).
For the species that belong to the same ecophenotype, in the fluvio-lacustrineone distances range from 0·516 (between the Danubian population of B. barbusand B. macedonicus-3) to 0·73 (between B. plebejus and B. macedonicus-1), whilein the rheophilic group, B. meridionalis seems to be very distant from all theothers (average distance of 0·79). If this population is excluded, the distancesamong the rheophilic species range from 0·162 (between B. cyclolepis and B.prespensis populations) to 0·562 (between B. prespensis and B. caninus), while forthe meridionalis group genetic distances vary from 0·165 to 0·339.
T
V.
Nei
’s(1
972)
gene
tic
dist
ance
sbe
twee
n15
popu
lati
ons
ofth
ege
nus
Bar
bus
base
don
the
anal
yses
of27
enzy
me
loci
Pop
ulat
ion
12
34
56
78
910
1112
1314
1B
.pe
lopo
nnes
ius
(1)
2B
.pe
lopo
nnes
ius
(2)
0·02
83
B.
pelo
ponn
esiu
s(3
)0·
040
0·00
74
B.
pelo
ponn
esiu
s(4
)0·
056
0·01
60·
008
5B
.m
erid
iona
lis0·
806
0·79
10·
801
0·80
96
B.
pete
nyi
0·21
70·
165
0·18
80·
196
0·79
47
B.
cani
nus
0·30
30·
283
0·30
20·
313
0·76
60·
339
8B
.cy
clol
epis
0·32
90·
295
0·29
40·
322
0·83
90·
331
0·56
29
B.
pres
pens
is0·
410
0·36
60·
379
0·40
90·
733
0·31
80·
545
0·16
210
B.
mac
edon
icus
(1)
0·44
80·
396
0·41
00·
424
1·12
70·
424
0·59
60·
549
0·48
411
B.
mac
edon
icus
(2)
0·44
60·
396
0·40
90·
424
1·13
70·
426
0·59
50·
547
0·48
10·
001
12B
.m
aced
onic
us(3
)0·
441
0·39
00·
403
0·41
71·
129
0·42
00·
588
0·54
00·
480
0·00
10·
000
13B
.ba
rbus
FR
0·65
90·
623
0·62
80·
637
0·68
60·
598
0·72
10·
604
0·45
70·
564
0·56
70·
559
14B
.ba
rbus
CZ
R0·
623
0·58
10·
596
0·60
00·
750
0·55
30·
675
0·64
90·
523
0·52
10·
524
0·51
60·
044
15B
.pl
ebej
us0·
670
0·61
90·
636
0·62
30·
712
0·61
10·
630
0·72
60·
581
0·73
00·
728
0·72
30·
622
0·59
9

282 . . .
D = 0.1
B. peloponnesius (4)
B. peloponnesius (3)
B. peloponnesius (2)
B. peloponnesius (1)
B. caninus
B. petenyi
B. macedonicus (1)
B. macedonicus (2)
B. macedonicus (3)
B. cyclolepis
B. prespensis
B. barbus FR
B. barbus CZR
B. meridionalis
B. plebejus
Neighbour joining
60
64
99
77
35
37
4197
40
71
69
100
F. 2. Unrooted consensus tree built using the neighbour joining algorithm and based on Nei’s (1972)genetic distances (D). Underlined taxa belong to the rheophilic species group. Numbers at nodesindicate bootstrap values (1000 replicates).
B.caninus) (Table V and Figs 2 and 3). The Italian B. caninus and Danubian B.petenyi appear to be close to each other and also closer to B. peloponnesius thanto B. meridionalis. This result supports an earlier study by Bianco (1995b) inwhich B. caninus showed distinct morphological characters when compared withB. meridionalis from France. Concerning the Danubian B. petenyi, Karakousiset al. (1993, 1995), reported that Romanian populations of this species are veryclose to B. peloponnesius populations from Northern Greece and so B. petenyimay be regarded as sub-species of B. peloponnesius. However, in our study thesame populations of B. peloponnesius are well differentiated from the populationof B. petenyi and are always separated in the phylogenetic trees by B. caninus.
As we can see in the distance matrix (Table V), B. meridionalis is geneticallyvery distant from the other species of rheophilic barbs (either of meridionalis orof cyclolepis groups) and in the phylogenetic trees it occupies a basal position(Figs 2 and 3). This position of B. meridionalis may be the reason that we do notobserve a monophyletic clustering of the meridionalis-like species. Barbusmeridionalis is thought to have gone through a severe population bottleneck,which resulted in a loss of heterozygosity (Berrebi et al., 1988; Persat & Berrebi,

283
10 changes
B. peloponnesius (1)
B. peloponnesius (2)
B. peloponnesius (4)
B. peloponnesius (3)
B. petenyi
B. macedonicus (1)
B. macedonicus (2)
B. macedonicus (3)
B. cyclolepis
B. prespensis
B. barbus FR
B. barbus CZR
B. meridionalis
B. plebejus
Maximum parsimony
45
50
95
73
35
20
32
9683
99
B. caninus
5483
F. 3. Unrooted consensus tree obtained using the Wagner’s parsimony method. Underlined taxa andnumbers at nodes correspond to rheophilic species and bootstrap values (1000 replicates)respectively. Branch lengths are proportional to the number of changes between taxa (see scalebar).
1990). Consequently, it is probable that this species may have lost a number ofalleles, which may have been important in linking it with the other membersof the same group.
For the reasons mentioned above, if we exclude B. meridionalis, the otherrheophilic species of the meridionalis group (B. peloponnesius, B. petenyi and B.caninus) are very closely related (genetic distances from 0·165 to 0·339) and in theneighbour joining tree they produce a monophyletic assemblage. The clusteringof these three species may indicate dispersion by river captures, which occurredin moderately high altitudes of mountain valleys from the Danube to the AxiosRiver via the Morava River of the Danube basin (Banarescu, 1960 inEconomidis & Banarescu, 1991) and to northern Italian and Slovenian rivers.
The populations of B. peloponnesius analysed in this study, present a higherdegree of heterozygosity and genetic polymorphism (Table IV), compared to thepopulations of B. petenyi and B. caninus. This was also observed in an earlierstudy (Karakousis et al., 1995). An explanation may be that these populationswere less affected by glacial extensions and therefore retained a more stablepopulation size.
Barbus prespensis and B. cyclolepis form the single monophyletic group ofspecies presented in both phylogenetic trees (Figs 2 and 3). This resultwas expected since these two species belong to the cyclolepis group (Almaca,1984; Economidis, 1989). Nevertheless, this group never gives a monophyletic

284 . . .
assemblage with the species of the meridionalis group, so as to produce amonophyletic cluster of rheophilic taxa.
In the fluvio-lacustrine group, the three species (B. barbus, B. macedonicus andB. plebejus) are genetically very different (with genetic distances that range from0·516 to 0·73). Distances are almost two- to threefold greater than distancesbetween species in the meridionalis and cyclolepis groups (from 0·165 to 0·339and 0·162, respectively). Bianco (1995a) considered that these species are able todisperse using river confluences in the lower parts of the basins (for instanceduring glacial maxima that produced marine regressions) and regarded them asrelicts of the Messinian-Lago Mare phase of the Mediterranean.
The two populations of B. barbus from France and the Czech Republic aregenetically very close (Table IV, and Figs 2 and 3). In a previous study,Machordom et al. (1995), reported that populations of this species from Franceand Belgium presented smaller genetic distance (0·008), compared to that foundin our study (0·044) between the French and the Czech populations. TheDanubian population also shows a high percentage of polymorphic loci andheterozygotes (Table IV) compared to the French one. This may reflect that theDanube represented a refuge for a stock that eventually gave rise to a widespreadand relatively homogeneous species in Central and North Europe, after theretreat of glaciers in the Holocene (Persat & Berrebi, 1990).
The present study shows that there are no monophyletic groups clustering therheophilic or the fluvio-lacustrine species (Figs 2 and 3). In each ecophenotype,species appear to be paraphyletic, suggesting that the concept of monophyleticspecies’ groups (or ecophenotypes) is probably wrong and the postulatedhypothesis of two main invasions (and so two main lineages: one adapted to theplains biotope and the other to upstream rivers) is not supported.
To explain this set of results, two main hypotheses can be proposed.(1) We cannot exclude the possibility that allozymes are not appropriate
markers to classify the taxa studied here. The important differentiation betweenspecies (high number of diagnostic loci) is most likely due to the isolationbetween species. Under that hypothesis, common ancestor allozymic characters(synapomorphic) may be rare, as after long and continuous differentiation, thesecharacters may have evolved in different ways in each isolated population.
(2) The second hypothesis, which is considered the most probable, is theexistence of independent species in Europe, rather than species belonging tospecies groups. The reason that we did not find monophyletic assemblages thatgroup each of the rheophilic and the fluvio-lacustrine barbs, may be that speciesof a given ecophenotype are paraphyletic. The fluvio-lacustrine species are neverfound to be part of a monophyletic clade, whereas the rheophilic species, withthe tree reconstruction methods used here, present only one well-supportedmonophyletic group, that comprises B. cyclolepis and B. prespensis.
That hypothesis may suggest convergence. For instance, the adaptation tomountain life could produce, from independent taxa, a similar phenotype byconvergence: the small size, the brown marbled colour, the scales and the shapeof the body and the fins. Such a hypothesis needs to be confirmed using othermarkers and other samples, but at this time, we consider it to be the mostprobable explanation. The present data conflict with the idea of twomonophyletic lineages of barbs which had dispersed across the European

285
continent and led to new species due to geographical isolation. We can imaginethat an ancestral species was genetically homogeneous, just like the presentspecies B. barbus, widespread in Central and Northern Europe. From thathomogeneous ancestor, local divergence may have occurred independently andconsequently led to convergent morphological evolution.
The authors thank C. D. Triantaphyllidis and P. S. Economidis for help in samplingthe Greek species and for fruitful discussions; M. Specchi for collecting the Italiansamples; and V. Slechta for the Danubian samples. The study was supported with ECgrant EV5VCT920097, subcontract CIPDCT925119 and CST by a grant from the GreekFoundation for State Scholarships.
References
Almaca, C. (1984). Notes on some species of Western Palearctic Barbus (Cyprinidae,Pisces). Arqivos do Museu Bocage, serie C II, 1–76.
Avise, J. C. (1994). Molecular Markers, Natural History and Evolution. London:Chapman & Hall.
Banarescu, P. (1973). Origin and affinities of the freshwater fish fauna of Europe.Ichthyologia 5, 1–8.
Belkhir, K., Borsa, P., Goudet, J. & Bonhomme, F. (1996). Genetix, logiciel sousWindows TM pour la genetique des populations. Montpellier: Laboratoire Genomeet Populations, CNRS UPR 9060, University Montpellier II.
Berrebi, P., Lamy, G., Cattaneo-Berrebi, G. & Renno, J. F. (1988). Variabilite genetiquede Barbus meridionalis Risso (Cyprinidae): une espece quasi monomorphe. BulletinFrancais de Peche et de Pisciculture 310, 77–84.
Berrebi, P., Leveque, C., Cattaneo-Berrebi, G., Agnese, J.-F., Guegan, J.-F. &Machordom, A. (1990). Diploid and tetraploid african Barbus (Osteichtyes,Cyprinidae): on the coding of differential gene expression. Aquatic LivingResources 3, 313–323.
Berrebi, P., Kottelat, M., Skelton, P. & Rab, P. (1996). Systematics of Barbus: state ofthe art and heuristic comments. Folia Zoologica 45 (Suppl. 1), 5–12.
Bianco, P. G. (1989). Some hypotheses on the origin of primary freshwater fishes insouthern Europe. Biologia Gallo-hellenica 15, 59–64.
Bianco, P. G. (1990). Potential role of the paleohistory of the Mediterranean andParatethys basins on the early dispersal of Euro-Mediterranean freshwater fishes.Ichthyological Exploration of Freshwaters 1, 167–184.
Bianco, P. G. (1995a). Factors affecting the distribution of freshwater fishes especially inItaly. Cybium 19, 241–259.
Bianco, P. G. (1995b). A revision of the Italian Barbus species (Cypriniformes:Cyprinidae). Ichthyological Exploration of Freshwaters 6, 305–324.
Bonhomme, F., Catalan, J., Britton-Davidian, J., Chapman, V. M., Moriwaki, D., Nevo,E. & Thaler, L. (1984). Biochemical diversity and evolution in the genus Mus.Biological Genetics 22, 275–303.
Collares-Peirera, M. J. (1994). Revision caryologique des barbines et hypothesesconcernant la plesiomorphie possible de l’etat polyploıde chez les cyprinides.Bulletin Francais de Peche et de Pisciculture 334, 191–199.
Cracraft, J. (1983). Species concept and speciation analysis. In Current Ornithology(Johnston, R. E., ed.), pp. 159–187. New York: Plenum Press.
Crivelli, A. J. & Maitland, P. S. (1995). Future prospects for the freshwater fish fauna ofthe north Mediterranean region. Biological Conservation 72, 335–337.
Darlington, P. J. (1948). The geographical distribution of cold-blooded vertebrates.Quarterly Review of Biology 23, 105–123.
Doadrio, I. (1990). Phylogenetic relationships and classification of western palearcticspecies of the genus Barbus (Osteichtyes, Cyprinidae). Aquatic Living Resources 3,265–282.

286 . . .
Economidis, P. S. (1989). Distribution pattern of the genus Barbus (Pisces: Cyprinidae)in the freshwaters of Greece. Travaux du Museum d’Histoire Naturelle GrigoreAntipa—Bucarest XXX, 223–229.
Economidis, P. S. & Banarescu, P. M. (1991). The distribution and origins of freshwaterfishes in the Balkan peninsula, especially in Greece. Internationale Revue GesamtenHydrobiologie 76, 257–283.
Farris, J. S. (1977). Phylogenetic analysis under Dollo’s laws. Systematic Zoology 26,77–88.
Felsenstein, J. (1993). PHYLIP 3.5 (Phylogeny Inference Package). Seattle: Universityof Washington.
Howes, G. J. (1987). The phylogenetic position of the Yugoslavian Cyprinid fish genusAulopyge Heckel, 1841, with an appraisal of the genus Barbus Cuvier & Cloquet,1816, and the subfamily Cyprinidae. Bulletin of the British Museum of NaturalHistory (Zoology) 52, 165–196.
Karakousis, Y., Peios, C., Economidis, P. S. & Triantaphyllidis, C. (1993). Multivariateanalysis of the morphological variability among Barbus peloponnesius (Cyprinidae)populations from Greece and two populations of B. meridionalis meridionalis andB. meridionalis petenyi. Cybium 17, 229–240.
Karakousis, Y., Machordom, A., Doadrio, I. & Economidis, P. S. (1995). Phylogeneticrelationships of Barbus peloponesius Valenciennes, 1842 (Osteichthyes: Cyprinidae)from Greece with other species of Barbus as revealed by allozyme electrophoresis.Biochemical Systematics and Ecology 23, 365–375.
Karaman, M. (1971). Susswasserfische der Turkei. 8 Teil. Revision der BarbenEuropas, Vorderasien und Nordafrikas. Mitteilungen aus den HamburgischenZoologischen Museum und Institut 67, 175–254.
Kluge, A. G. & Farris, J. S. (1969). Quantitative phyletics and the evolution of anurans.Systematic Zoology 18, 1–32.
Kottelat, M. (1997). European freshwater fishes. An heuristic checklist of the fresh-water fishes of Europe (exclusive of former USSR), with an introduction fornon-systematists and comments on nomenclature and conservation. Biologia,Bratislava 52 (Suppl. 5), 1–271.
LeQuesne, W. J. (1972). Further studies based on the uniquely derived characterconcepts. Systematic Zoology 21, 281–288.
Machordom, A. (1990). Filogenia y evolution del genero Barbus en la peninsula ibericamediante marcadores moleculares. Ph.D. Thesis, Universidad Complutence deMadrid, Spain.
Machordom, A., Doadrio, I. & Berrebi, P. (1995). Phylogeny and evolution of the genusBarbus in the Iberian Peninsula as revealed by allozyme electrophoresis. Journal ofFish Biology 47, 211–236.
Myers, G. S. (1960). Preface to any future classification of the cyprinid fishes of the genusBarbus. Stanford Ichthyological Bulletin 7, 212–215.
Nei, M. (1972). Genetic distance between populations. American Naturalist 106,283–292.
Ohno, S. (1970). Evolution by Gene Duplication. New York: Springer-Verlag.Pasteur, N., Pasteur, G., Bonhomme, F., Catalan, J. & Britton-Davidian, J. (1987).
Manuel de genetique par electrophorese des proteines. Paris: Lavoisier.Persat, H. & Berrebi, P. (1990). Relative ages of present populations of Barbus barbus
and Barbus meridionalis (Cyprinidae) in southern France: preliminary consider-ations. Aquatic Living Resources 3, 253–263.
Saitou, N. & Nei, M. (1987). The neighbor-joining method: a new method forreconstructing phylogenetic trees. Molecular Biology and Evolution 4, 406–425.
Swofford, D. L. (1993). PAUP: Phylogenetic Analysis Using Parsimony. Champaign,Ill.: Illinois Natural History Survey.
Tsigenopoulos, C. (1996). Phylogeographie mitochondriale du genre Barbus au nord dela Mediterranee. Une approche de la speciation des poissons d’eau douced’Europe occidentale, DEA dissertation, University Montpellier II, France, pp. 29.
Top Related

Copyright © 2022 FDOKUMEN

