







Bahasa
Halaman
Hukum


1
The NAMPT Promoter is Regulated by Mechanical Stress, STAT5, and ARDS-
Associated Genetic Variants
Xiaoguang Sun1*, Venkateswaran Ramamoorthi Elangovan2,3*, Brandon Mapes1*, Sara M.
Camp2, Saad Sammani1, Laleh Saadat1, Ermelinda Ceco1, Shwu-Fan Ma4, Carlos Flores5,
Matthew S. MacDougall1, Hector Quijada2, Bin Liu1, Carrie L. Kempf2, Ting Wang2, Eddie T.
Chiang2, Joe G. N. Garcia1,2,3§
1Department of Medicine, University of Illinois at Chicago; 2Arizona Respiratory Center and Department of Medicine, The University of Arizona; 3 Department of Pharmacology, University of Illinois at Chicago.
4Department of Medicine, University of Chicago; 5Centro de Investigación Biomédica en Red de Enfermedades Respiratorias de Enfermedades Respiratorias, Instituto de Salud Carlos III §Corresponding Author: Joe G. N. Garcia, MD Senior Vice President for Health Sciences Arizona Health Sciences Center University of Arizona (520) 626-1197 [email protected] *Contributed equally to this work **Funding: This work was supported by the NIH/NHLBI grant R01-HL73994 (JGNG) ***Running Title: Regulation of NAMPT promoter activity in ARDS ****Keywords: ARDS, cyclic stretch, MSIR, NAMPT, PBEF, STAT5
Page 1 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

2
ABSTRACT
Rationale: Increased nicotinamide phosphoribosyltransferase (NAMPT) transcription is
mechanistically linked to ventilator-induced inflammatory lung injury (VILI), with VILI severity
attenuated by reduced NAMPT bioavailability. The molecular mechanisms of NAMPT promoter
regulation in response to excessive mechanical stress remain poorly understood. Objective: To
define the contribution of specific transcription factors, ARDS-associated SNPs, and promoter
demethylation to NAMPT transcriptional regulation in response to mechanical stress. Methods:
In vivo NAMPT protein expression levels were examined in mice exposed to high tidal volume
mechanical ventilation. In vitro NAMPT expression levels were examined in human pulmonary
artery endothelial cells (EC) exposed to 5% or 18% cyclic stretch (CS), with NAMPT promoter
activity assessed using NAMPT promoter luciferase reporter constructs with a series of nested
deletions. In vitro NAMPT transcriptional regulation was further characterized by measuring
luciferase activity, DNA demethylation, and chromatin immunoprecipitation. Results: VILI–
challenged mice exhibited significantly increased NAMPT expression in BAL leukocytes and in
lung endothelium. A mechanical stress-inducible region (MSIR) was identified in the NAMPT
promoter from −2428 to −2128bp. This MSIR regulates NAMPT promoter activity, mRNA
expression, and STAT5 binding, which is significantly increased by 18% CS. In addition,
NAMPT promoter activity was increased by pharmacologic promoter demethylation and
inhibited by STAT5 silencing. ARDS-associated NAMPT promoter SNPs, rs59744560 (-948G/T)
and rs7789066 (-2422A/G), each significantly elevated NAMPT promoter activity in response to
18% CS in a STAT5-dependent manner. Conclusions: NAMPT is a key novel ARDS
therapeutic target and candidate gene with genetic/epigenetic transcriptional regulation in
response to excessive mechanical stress.
Page 2 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

3
INTRODUCTION
Acute respiratory distress syndrome (ARDS) is characterized by severe hypoxemia and an
unacceptable but persistently high mortality rate (~30%) (1, 2). Mechanical ventilation is a life-
saving intervention in critically ill patients with respiratory failure due to ARDS, however,
excessive mechanical ventilation also contributes directly to inflammatory lung injury, a process
known as ventilator-induced lung injury (VILI) (3, 4). Like ARDS, VILI is also associated with
augmented capillary leakage, acute inflammation, and increases in inflammatory cytokine
expression (5).
We previously utilized high-throughput functional genomic approaches with extensive
microarray-based lung gene expression profiling, in canine and murine preclinical models of
ARDS as well as human ARDS patients, to search for novel ARDS/VILI biomarkers and
therapeutic targets. These studies identified NAMPT (6), a gene encoding the cytozyme
nicotinamide phosphoribosyltransferase (NAMPT) also known as pre–B-cell colony-enhancing
factor (PBEF) or visfatin (7), as a novel biomarker. NAMPT/PBEF, originally named for its
effects on the maturation of B-cell precursors (8), was subsequently found to be secreted from
adipocytes (9). NAMPT/PBEF expression in human amniotic epithelial cell lines is up-regulated
by mechanical stress and by inflammatory cytokines (10). Our prior studies demonstrated that
NAMPT/PBEF is a direct neutrophil chemotactic factor, which synergistically exacerbated VILI-
mediated inflammatory injury in vivo by intratracheal delivery of recombinant NAMPT/PBEF (6,
11). Multiple indices of VILI-associated lung injury were reduced in heterozygous NAMPT+/-
mice including peak inspiratory pressures and gene expression pathways (e.g., NFkB signaling,
leukocyte extravasation, apoptosis, and Toll receptor pathways) (6, 11). In addition, our study in
NAMPT sequencing and subsequent genotyping identified single nucleotide polymorphisms
(SNPs) within the NAMPT promoter to be significantly associated with enhanced susceptibility
to sepsis and ARDS (6), suggesting NAMPT is an ARDS susceptibility gene. The genetic
association of NAMPT promoter SNPs with ARDS was replicated in a separate ARDS cohort
and found to be associated with the number of ventilator-free days and overall ARDS mortality
(12). Finally, novel ARDS therapeutic strategies designed to reduce NAMPT enzymatic activity
or bioavailability in preclinical models of ARDS and VILI resulted in significant VILI protection
(11, 13, 14).
These studies, highlighting the intimate involvement of NAMPT activity/expression in ARDS/VILI
susceptibility and severity, led us to explore the molecular regulation of NAMPT promoter
activity. Although the genetic and epigenetic factors involved in lung cell responses to
Page 3 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

4
mechanical stress are poorly understood, we now report significantly increased NAMPT
expression in human pulmonary artery endothelial cells (EC) exposed to 18% cyclic stretch (CS)
and in murine lungs exposed to high tidal volume mechanical ventilation. Increased NAMPT
expression evoked by 18% CS was mediated by the binding of NAMPT promoter to the
transcription factor, STAT5, resulting in increased promoter activity. STAT5 silencing attenuated
NAMPT promoter activation in human EC and NAMPT expression was also significantly
enhanced by DNA demethylation, an event significantly associated with increased STAT5
binding. Finally, two single nucleotide polymorphisms (SNPs) located in NAMPT promoter,
rs59744560 (-948G/T) and rs7789066 (-2422A/G), significantly linked to ARDS in two European
descent (ED) cohorts (Chicago, Spain), increased basal and 18% CS-induced NAMPT promoter
activation in a STAT5-dependent manner. Together, these findings suggest that NAMPT is an
important ARDS candidate gene and novel therapeutic target with genetic/epigenetic
transcriptional regulation highly influenced by excessive mechanical stress and ARDS-
associated SNPs.
MATERIALS AND METHODS
Cell Culture, siRNA Transfection, and Cyclic Stretch. Human pulmonary artery endothelial
cells (EC) were obtained from Lonza (Walkersville MD) and were cultured as described
previously (15) in the manufacturer’s recommended endothelial growth medium-2 (EGM-2).
Cells were grown at 37°C in a 5% CO2 incubator, and passages 6 to 9 were used for
experiments. Media was changed one day before experimentation. For RNA interference, On-
Target Plus siRNAs against STAT5A, STAT5B, and firefly luciferase (siCONTROL#2 used as
negative control) were obtained from Dharmacon (Lafayette CO) and transfected into EC at a
final concentration of 100nM. For cyclic stretch (CS) studies, EC were plated on Bioflex
collagen I type cell culture plates (FlexCell International, Hillsborough NC) and stimulated for 4
hours at 5% or 18% CS as previously described (16) on the FlexCell FX-5000 System (FlexCell
International), mimicking low and high tidal volume ventilation, respectively. For demethylation
studies, EC were treated with 5-aza-2’-deoxycytidine (5’-Aza) (Sigma-Aldrich, St. Louis MO) for
72 hours at indicated concentrations to inhibit DNA methyltransferase enzymes. Cells were
subsequently harvested for RNA analysis by qPCR.
Models of Ventilator-Induced Murine Lung Injury. Male C57BL/6J mice were induced VILI,
as previously described(11). Bronchoalveolar lavage (BAL) and lung tissue were collected from
Page 4 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

5
these mice and stained (IHC) with goat polyclonal anti-NAMPT/PBEF antibodies (Lampire
Biological Laboratories, Pipersville PA) as previously described (6).
Chromatin Immunoprecipitation (ChIP) and qPCR. ChIP of DNA bound to STAT protein
complexes was performed using the EZ-Magna ChIP assay (EMD Millipore, Billerica MA) in
accordance with manufacturer’s recommended procedure. STAT cross-linked protein/DNA
complexes were immunoprecipitated using STAT2, STAT3 and STAT5 antibodies (sc-476, sc-
482, sc-835, Santa Cruz Biotechnology, Santa Cruz CA). qPCR was performed using SsoFast
EvaGreen Supermix (BioRad) as per manufacturer’s protocol.
Gene Cloning, Mutagenesis, and 5’-Deletion Mutations. Gene cloning, mutagenesis and
luciferase activity assays were performed as previously described (17). Luciferase Reporter
Gene Assays. All constructs were transfected into EC, where a plasmid containing the Renilla
luciferase gene (pRL-TK) was co-transfected as a control. Transfected cells were exposed to
either static conditions or 18% CS for 4 hours, and lysed in passive lysis buffer. Luciferase
activity was measured by Dual-Luciferase Assay Kits using the GloMax-Multi Detection System
(Promega). Relative activities were expressed as the ratio of firefly luciferase in pGL3 to renilla
luciferase in pRL-TK (RLU). Five independent transfections and duplicate luciferase assays
were performed for each condition.Statistical Analysis. Meta-analysis of genotypic information
from Chicago and Spanish cohort samples were conducted assuming a fixed effects model
using a Mantel-Haenszel stratified analysis with Epidat 3.0. The ANOVA test was used for
comparison of luciferase activities among different constructs. Statistical significance was
defined at p<0.05 in both tests.
RESULTS
Excessive Mechanical Stress Increases NAMPT Expression in vivo and in vitro. Validating
our prior studies in mice exposed to excessive mechanical stress (6), we identified increased
NAMPT protein expression in BAL macrophages (cytoplasm and nucleus) (Figure 1A-B) and
pulmonary endothelium (Figure 1C-D) in VILI-exposed mice. NAMPT expression was next
examined in vitro in lung EC exposed to 5% and 18% cyclic stretch (CS), simulating low and
high tidal volume ventilation, respectively (18). Analysis of EC NAMPT mRNA levels (RT-PCR)
after exposure to 5% or 18% CS demonstrated significant increases in NAMPT transcription in
ECs exposed to 18% CS (4, 6, and 48 hrs) compared to ECs exposed to 5% CS (Figure 2A).
Page 5 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

6
These results were supported by studies utilizing a 3kb NAMPT promoter (−3028 bp to +1 ATG)
in luciferase reporter transfected into EC and exposed to 18% CS with NAMPT promoter activity
significantly increased 1-4 hrs post exposure (Figure 2B).
Effects of 5'-Deletions on Mechanical Stress- Dependent NAMPT Promoter Activity. To
determine the core promoter sequence and regulatory elements within the NAMPT promoter,
we next analyzed NAMPT luciferase reporter promoter activation in response to 18% CS using
a nested deleted promoter with varying DNA length in ECs (Figure 3). These studies revealed
that truncation of the NAMPT promoter from −2428 to −1228 (2082 bp to 882 bp upstream of
TSS) decreased promoter activity by ~30% (Figure 3B), whereas truncation of the NAMPT
promoter from −1528 to −628 (1182 bp to 282 bp upstream of TSS) increased promoter activity
by ~50% (Figure 3B). These results suggest that the −2428 to −1228 region is potentially
regulated by factors suppressing promoter activity, while the −1228 to −328 region (882 bp
upstream and 18 bp downstream of TSS) appears essential for core NAMPT promoter activity.
ECs transfected with identical series of NAMPT promoter fragments were next exposed to 18%
CS or static conditions (4 hrs) with exposure to 18% CS increasing NAMPT promoter activity in
the presence of the −3028 to −2128 region (2682 bp to 1782 bp upstream of TSS) to 80~100%
(Figure 2B) . And these 18% CS-mediated increases were abolished by truncation of the
NAMPT promoter from −2428 to −2128 (2082 bp to 1782 bp upstream of TSS) (Figure 3B).
These data are highly suggestive of a mechanical stress-inducible region (MSIR at -2428 to -
2128) contains critical NAMPT promoter responsive elements.
NAMPT Promoter Responses to Mechanical Stretch Are Regulated by STAT5. We next
analyzed the −2428 to −2128 sequence using Genomatix (www.genomatix.de) and TESS
software (www.cbil.upenn.edu/cgi-bin/tess) and identified STAT family transcription factors as
potentially capable of binding to this genomic region (Figure 3A). To address whether specific
STAT transcription factors interacting with the putative STAT-binding sites within the NAMPT
promoter, we evaluated STAT2, STAT3, and STAT5 and performed ChIP assays in EC
exposed to 18% CS versus static condition, using two sets of primers targeting the 5’ and 3’
STAT binding sites. Compared with static cells, 18% CS significantly increased the occupancy
of STAT5 at the NAMPT promoter in EC relative to an input control at both the 5’ STAT binding
site (Figure 4A) and the 3’ STAT binding site (Figure 4B). These data suggest STAT5 is a
critical transcription factor that mediates 18% CS-induced NAMPT promoter activation.
Page 6 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

7
DNA Demethylation Increases NAMPT Transcription and STAT5 Binding to NAMPT
Promoter. To address epigenetic regulation of the NAMPT promoter in response to 18% CS,
we assessed the status of NAMPT promoter methylation. Exposure to 18% CS-induced
demethylation of the promoter region (-2000 to -1800 bp) relative to ATG, corresponding to a
region containing in silico PAX5 binding sites (Supplemental Figure 1). To verify that STAT5
binding to the NAMPT promoter is influenced by DNA demethylation, we performed ChIP
assays in 5-aza-2’-deoxycytidine (5’-Aza)- or control carrier (PBS)- exposed EC and determined
that 5’-Aza significantly increases STAT5 NAMPT promoter occupancy, as well as STAT3
(Figure 5A). Consistent with these results, 5’-Aza incubation dramatically increased NAMPT
mRNA level compared to controls (~2.5 fold) (Figure 5B), similar to the effect of 18% CS. In EC
pretreated with 5’-Aza, exposure to 18% CS did not further increase NAMPT mRNA levels (data
not shown).
Two NAMPT Promoter Variants are Associated with ARDS Susceptibility and Increase
Promoter Activity via STAT5. Our prior sequencing studies identified two NAMPT promoter
SNPs (rs9770242 and rs61330082) that were found in subsequent genotyping studies to be
highly associated with ARDS susceptibility and severity (6). Additional sequencing studies in
Chicago and Spanish ARDS cohorts identified NAMPT SNPs rs59744560 (-948G/T) and
rs7789066 (-2422A/G, cohort) as significant ARDS-associated SNPs (Tables 1 and 2). The
consistency of allele effects was evidenced by a stratified meta-analysis yielding per allele odds
ratios of 3.29 and 2.73 for -948T and -2422G, respectively. In silico studies identified SNPs -
948T and -2422G with significant potential for altering binding of two transcription factors,
glucocorticoid receptor (GR) and nuclear factor of activated T-cells (NFAT), to the NAMPT
promoter compared to -948G and -2422A, respectively. Using site-directed mutagenesis, minor
alleles of the two variants were inserted into the NAMPT promoter reporter and transfected into
ECs. Compared to -948G and -2422A, ARDS-associated alleles -948T and -2422G significantly
increased NAMPT promoter activities under static conditions. These ARDS-associated allele
promoter activities were further enhanced by 18% CS. Reductions in EC STAT5 protein levels
(using siRNA against STAT5a/b), however, attenuated the enhanced promoter response
produced by the ARDS-associated SNPs (Figure 6).
DISCUSSION
Despite improved understanding of the pathophysiology of ARDS, the underlying mechanisms
for the injurious effects of mechanical ventilation in the setting of ARDS remain unclear, and
Page 7 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

8
effective pharmacotherapy has not yet emerged. We previously used genomic-intensive
approaches to identify potential ARDS and VILI susceptibility candidate genes (19, 20) and
determined that NAMPT, the gene encoding the pro-inflammatory cytokine NAMPT/PBEF, as a
potential VILI candidate gene and novel biomarker in sepsis and ARDS (6, 11). NAMPT SNPs
in the promoter region were determined to confer susceptibility to sepsis-induced ARDS (6), a
finding that was further validated in an independent cohort with increased ARDS mortality (12).
However, the underlying molecular mechanism involved in NAMPT gene regulation in the
settings of ARDS and VILI remains poorly understood.
In this study, we demonstrate that excessive levels of mechanical stress (18% CS), but not
homeostatic levels (5% CS), rapidly increase NAMPT promoter activity, transcription, and
protein expression that is significantly associated with specific inducible promoter segments.
Through serial progressive 5’ to 3’ unidirectional deletions and promoter activity assays, we
detected a distal promoter or enhancer region (-3028 to -2428 bp), a proximal promoter region (-
1228 to -628 bp), and a negative regulatory region (-2428 to -1228 bp). In silico analysis also
revealed that the proximal promoter of the NAMPT gene is conserved across vertebrate species,
suggesting a common regulation mechanism of the NAMPT gene across species
(Supplemental Table 1). Our study demonstrates that the mechanical stress-inducible region
critical for NAMPT responses to 18% CS resides within the promoter region between -2400 and
-2100 bp. Although additional genetic and epigenetic mechanisms of NAMPT regulation are
also in play (21), the current studies indicate that mechanical stress-mediated increases in
NAMPT expression involve a group of transcription factors known as the STATs (signal
transducer and activator of transcription proteins). STATs are recognized as activated by
mechanical stretch (22) and we recently reported mechanical stress-mediated increased
promoter activity within the HMGB1 gene as STAT3 dependent (23). Cyclic stretch significantly
increases JAK2/STAT5 phosphorylation (24) and STAT5 translocation to the nucleus (25).
Leveraging in silico information generated by predictions from Genomatix (www.genomatix.de),
UCSC Genome Browser (genome.ucsc.edu/cgi-bin/hgGateway) (GRCh37/hg19), and previous
reports (8), we identified transcription factor-binding sites for STAT5 and determined that STAT5
exerts critical positive regulatory control in NAMPT responses to mechanical stretch. STAT5
activation is characterized by translocation into the nucleus and subsequent binding to the
corresponding DNA sequence to regulate target gene transcription, promote cell differentiation,
proliferation, and survival (26). STAT5a and STAT5b are fraternal twins of signal transduction
and transcriptional activation. STAT5 is frequently activated by cytokines and induces cytokine
Page 8 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

9
functions (IL-2, IL-3, IL-5, GM-CSF) (27) and promotes B cell development by controlling cell
survival (28). In prior and current studies, we have demonstrated involvement of the STAT
pathway in responses to excessive pathological mechanical stress and VILI development (11)
via enhanced STAT5 (and STAT3) binding to the NAMPT promoter and up-regulated NAMPT
promoter activity, confirming NAMPT’s critical role in mechanical ventilation-induced ARDS and
exacerbation of ARDS (6, 8, 10, 11).
Epigenetic mechanisms of NAMPT regulation are also involved in NAMPT promoter regulation
as 18% CS significantly increased DNA demethylation of PAX5 in silico binding sites and DNA
demethylation by 5’-Aza increased NAMPT transcription and enhanced STAT5 as well as
STAT3 recruitment to the NAMPT promoter. Since STAT3:STAT5 heterodimers are elicited by
M-CSF binding to cis-inducible element sites with high affinity to promote STAT5-dependent
genes (29), our data suggests the possibility that STAT5 may form heterodimers with STAT3 to
regulate the NAMPT promoter. During maturation of pro-B cells, STAT5 also cooperates with
PAX5 to promote distal immunoglobulin heavy-chain gene transcription (30). Under 18% CS,
increased recruitment of STAT5 could synergize with demethylation of PAX5-binding elements
to promote NAMPT transcription.
Finally, our studies identified two ARDS-associated NAMPT promoter SNPs (-2422A/G, -
948G/T) and despite study limitations, including relatively small groups of ARDS patients and
controls and lack of cross validation in the two ARDS cohorts, our in vitro experiments
confirmed robust and significant influence of these SNPs on NAMPT promoter activity in
responses to 18% CS challenge. Functionality of the -948G/T (C/A) variant is supported by
recent studies associating this NAMPT SNP with low-grade inflammation in diabetic patients
(31). Under static conditions, silencing of STAT5 failed to influence the activity of promoters
with -2422A/G or -948G/T. However, in response to 18% CS, reductions in STAT5 expression
significantly attenuated SNP-dependent enhanced NAMPT promoter activity with either NAMPT
variant. Since GR acts as a transcriptional co-activator for STAT5 and enhances STAT5-
dependent transcription (32), and STAT5 combines with NFAT to promote Foxp3 transcription
(33), the effects of the -948G/T and -2422A/G SNPs may synergize with STAT5 to significantly
enhance NAMPT transcription with the influence of these co-transcription factors during
pathological stress. Together, these studies demonstrated that STAT5 significantly regulates
NAMPT transcription under pathological mechanical stress.
Page 9 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

10
Interaction of the NAMPT promoter with inflammation-related transcription factor STAT5,
NAMPT promoter demethylation, and ARDS-associated SNPs regulate NAMPT promoter
activity, thereby contributing to VILI and ARDS development.
ACKNOWLEDGEMENTS
The authors wish to acknowledge the assistance of Lakshmi Natarajan, Michael S. Wade,
Viswanathan Natarajan, and Giovanni Infusino (University of Illinois at Chicago) and Jaideep
Moitra (Mumbai, India).
This work was supported by the NIH/NHLBI grant R01-HL73994 (JGNG)
REFERENCES
1. Matthay MA, Zimmerman GA, Esmon C, Bhattacharya J, Coller B, Doerschuk CM, Floros J, Gimbrone MA, Jr., Hoffman E, Hubmayr RD, et al. Future research directions in acute lung injury: Summary of a national heart, lung, and blood institute working group. Am J Respir Crit Care Med 2003;167(7):1027-1035. 2. Goss CH, Brower RG, Hudson LD, Rubenfeld GD. Incidence of acute lung injury in the united states. Crit Care Med 2003;31(6):1607-1611. 3. ARDSNet. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. The acute respiratory distress syndrome network. N Engl J Med 2000;342(18):1301-1308. 4. Dreyfuss D, Saumon G. Ventilator-induced lung injury: Lessons from experimental studies. Am J Respir Crit Care Med 1998;157(1):294-323. 5. Ranieri VM, Suter PM, Tortorella C, De Tullio R, Dayer JM, Brienza A, Bruno F, Slutsky AS. Effect of mechanical ventilation on inflammatory mediators in patients with acute respiratory distress syndrome: A randomized controlled trial. JAMA 1999;282(1):54-61. 6. Ye SQ, Simon BA, Maloney JP, Zambelli-Weiner A, Gao L, Grant A, Easley RB, McVerry BJ, Tuder RM, Standiford T, et al. Pre-b-cell colony-enhancing factor as a potential novel biomarker in acute lung injury. Am J Respir Crit Care Med 2005;171(4):361-370. 7. Wang T, Zhang X, Bheda P, Revollo JR, Imai S, Wolberger C. Structure of nampt/pbef/visfatin, a mammalian nad+ biosynthetic enzyme. Nat Struct Mol Biol 2006;13(7):661-662. 8. Samal B, Sun Y, Stearns G, Xie C, Suggs S, McNiece I. Cloning and characterization of the cdna encoding a novel human pre-b-cell colony-enhancing factor. Mol Cell Biol 1994;14(2):1431-1437. 9. Havel PJ. Update on adipocyte hormones: Regulation of energy balance and carbohydrate/lipid metabolism. Diabetes 2004;53 Suppl 1:S143-151. 10. Ognjanovic S, Ku TL, Bryant-Greenwood GD. Pre-b-cell colony-enhancing factor is a secreted cytokine-like protein from the human amniotic epithelium. Am J Obstet Gynecol 2005;193(1):273-282.
Page 10 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

11
11. Hong SB, Huang Y, Moreno-Vinasco L, Sammani S, Moitra J, Barnard JW, Ma SF, Mirzapoiazova T, Evenoski C, Reeves RR, et al. Essential role of pre-b-cell colony enhancing factor in ventilator-induced lung injury. Am J Respir Crit Care Med 2008;178(6):605-617. 12. Bajwa EK, Yu CL, Gong MN, Thompson BT, Christiani DC. Pre-b-cell colony-enhancing factor gene polymorphisms and risk of acute respiratory distress syndrome. Crit Care Med 2007;35(5):1290-1295. 13. Camp SM, Ceco E, Evenoski CL, Danilov SM, Zhou T, Chiang ET, Moreno-Vinasco L, Mapes B, Zhao J, Gursoy G, et al. Unique toll-like receptor 4 activation by nampt/pbef induces nfκb signaling and inflammatory lung injury. J Clin Invest (submitted) 2014. 14. Moreno-Vinasco L, Quijada H, Sammani S, Siegler J, Letsiou E, Deaton R, Saadat L, Zaidi RS, Messana J, Gann PH, et al. Nicotinamide phosphoribosyltransferase inhibitor is a novel therapeutic candidate in murine models of inflammatory lung injury. Am J Respir Cell Mol Biol 2014; [Mar 3 Epub ahead of print]. 15. Dudek SM, Jacobson JR, Chiang ET, Birukov KG, Wang P, Zhan X, Garcia JG. Pulmonary endothelial cell barrier enhancement by sphingosine 1-phosphate: Roles for cortactin and myosin light chain kinase. J Biol Chem 2004;279(23):24692-24700. 16. Birukov KG, Jacobson JR, Flores AA, Ye SQ, Birukova AA, Verin AD, Garcia JG. Magnitude-dependent regulation of pulmonary endothelial cell barrier function by cyclic stretch. Am J Physiol Lung Cell Mol Physiol 2003;285(4):L785-797. 17. Sun X, Ma SF, Wade MS, Flores C, Pino-Yanes M, Moitra J, Ober C, Kittles R, Husain AN, Ford JG, et al. Functional variants of the sphingosine-1-phosphate receptor 1 gene associate with asthma susceptibility. J Allergy Clin Immunol 2010;126(2):241-249, 249 e241-243. 18. Birukova AA, Moldobaeva N, Xing J, Birukov KG. Magnitude-dependent effects of cyclic stretch on hgf- and vegf-induced pulmonary endothelial remodeling and barrier regulation. Am J Physiol Lung Cell Mol Physiol 2008;295(4):L612-623. 19. Simon BA, Easley RB, Grigoryev DN, Ma SF, Ye SQ, Lavoie T, Tuder RM, Garcia JG. Microarray analysis of regional cellular responses to local mechanical stress in acute lung injury. Am J Physiol Lung Cell Mol Physiol 2006;291(5):L851-861. 20. Grigoryev DN, Ma SF, Irizarry RA, Ye SQ, Quackenbush J, Garcia JG. Orthologous gene-expression profiling in multi-species models: Search for candidate genes. Genome Biol 2004;5(5):R34. 21. Adyshev DM, Elangovan VR, Moldobaeva N, Mapes B, Sun X, Garcia JG. Mechanical stress induces pre-b-cell colony-enhancing factor/nampt expression via epigenetic regulation by mir-374a and mir-568 in human lung endothelium. Am J Respir Cell Mol Biol 2014;50(2):409-418. 22. Mammoto A, Mammoto T, Ingber DE. Mechanosensitive mechanisms in transcriptional regulation. J Cell Sci 2012;125(Pt 13):3061-3073. 23. Wolfson RK, Mapes B, Garcia JG. Excessive mechanical stress increases hmgb1 expression in human lung microvascular endothelial cells via stat3. Microvasc Res 2013; [Dec 24 Epub ahead of print]. 24. Honsho S, Nishikawa S, Amano K, Zen K, Adachi Y, Kishita E, Matsui A, Katsume A, Yamaguchi S, Nishikawa K, et al. Pressure-mediated hypertrophy and mechanical stretch induces il-1 release and subsequent igf-1 generation to maintain compensative hypertrophy by affecting akt and jnk pathways. Circ Res 2009;105(11):1149-1158. 25. Penta K, Sawyer ST. Erythropoietin induces the tyrosine phosphorylation, nuclear translocation, and DNA binding of stat1 and stat5 in erythroid cells. J Biol Chem 1995;270(52):31282-31287. 26. Reich NC. Stats get their move on. JAKSTAT 2013;2(4):e27080.
Page 11 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

12
27. Teglund S, McKay C, Schuetz E, van Deursen JM, Stravopodis D, Wang D, Brown M, Bodner S, Grosveld G, Ihle JN. Stat5a and stat5b proteins have essential and nonessential, or redundant, roles in cytokine responses. Cell 1998;93(5):841-850. 28. Malin S, McManus S, Cobaleda C, Novatchkova M, Delogu A, Bouillet P, Strasser A, Busslinger M. Role of stat5 in controlling cell survival and immunoglobulin gene recombination during pro-b cell development. Nat Immunol 2009;11(2):171-179. 29. Novak U, Mui A, Miyajima A, Paradiso L. Formation of stat5-containing DNA binding complexes in response to colony-stimulating factor-1 and platelet-derived growth factor. J Biol Chem 1996;271(31):18350-18354. 30. Bertolino E, Reddy K, Medina KL, Parganas E, Ihle J, Singh H. Regulation of interleukin 7-dependent immunoglobulin heavy-chain variable gene rearrangements by transcription factor stat5. Nat Immunol 2005;6(8):836-843. 31. Zhang YY, Gottardo L, Thompson R, Powers C, Nolan D, Duffy J, Marescotti MC, Avogaro A, Doria A. A visfatin promoter polymorphism is associated with low-grade inflammation and type 2 diabetes. Obesity (Silver Spring) 2006;14(12):2119-2126. 32. Stocklin E, Wissler M, Gouilleux F, Groner B. Functional interactions between stat5 and the glucocorticoid receptor. Nature 1996;383(6602):726-728. 33. Hermann-Kleiter N, Baier G. Nfat pulls the strings during cd4+ t helper cell effector functions. Blood 2010;115(15):2989-2997. 34. Leander K, Gigante B, Silveira A, Vikstrom M, Hamsten A, Hogberg J. Nampt (visfatin) and akt1 genetic variants associate with myocardial infarction. Clin Chim Acta 2012;413(7-8):727-732. 35. Wade MS, Ma SF, Christie JD, Flores C, Garcia JG. Pbef gene polymorphisms are associated with acute lung injury in us- and spanish based cohorts. Am J Respir Crit Care Med 2010;181; 2010:A1022. 36. Ma SF, Xie L, Pino-Yanes M, Sammani S, Wade MS, Letsiou E, Siegler J, Wang T, Infusino G, Kittles RA, et al. Type 2 deiodinase and host responses of sepsis and acute lung injury. Am J Respir Cell Mol Biol 2011;45(6):1203-1211. 37. Flores C, Ma SF, Maresso K, Wade MS, Villar J, Garcia JG. Il6 gene-wide haplotype is associated with susceptibility to acute lung injury. Transl Res 2008;152(1):11-17. 38. Bernard GR, Artigas A, Brigham KL, Carlet J, Falke K, Hudson L, Lamy M, Legall JR, Morris A, Spragg R. The american-european consensus conference on ards. Definitions, mechanisms, relevant outcomes, and clinical trial coordination. Am J Respir Crit Care Med 1994;149(3 Pt 1):818-824. 39. Bone RC, Balk RA, Cerra FB, Dellinger RP, Fein AM, Knaus WA, Schein RM, Sibbald WJ. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The accp/sccm consensus conference committee. American college of chest physicians/society of critical care medicine. Chest 1992;101(6):1644-1655.
Page 12 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

13
Table 1. Association of ARDS risk in relation to NAMPT −2422 and −948 polymorphisms in an ARDS population
Polymorphism Genotype Odds Ratio Confidence Interval P Value
-2422 T/C TT Referent -- --
CC 2.73 1.18 - 6.30 0.015
-948 G/T GG Referent -- --
TT 3.29 1.07 - 10.12 0.03
Table 2. Summary of NAMPT SNPs significantly associated with phenotypes of various diseases
Positiona Locationb SNP_ID Alleles African American Associated Phenotypesc
European American Associated Phenotypesc
Spanish Associated Phenotypec
100287367 -2422T>C rs7789066 T/C - - ARDS
100286480 -1535C>T (-1543C>T)
rs61330082 C/T ARDS ARDS, CAD -
100285946 -1001G>T rs9770242 G/T - ARDS -
100285893 -948G>T rs59744560 G/T(C/A) - ARDS, DM2 -
100285368 -423A>G rs1319501 A/G - Risk of MI -
a. Position in chromosome 1 according to NCBI build 36 b. Reference sequence is NM_005746.2 c. References for associated phenotypes: (6, 31, 34, 35).
FIGURE LEGENDS
Figure 1. Exposure to high tidal volume ventilation significantly increases murine NAMPT
protein expression in BAL leukocytes and lung endothelium. Compared to spontaneously
breathing mice (A), protein expression of NAMPT was increased in bronchoalveolar lavage
(BAL) leukocytes obtained from VILI-exposed mice (B). Protein expression of NAMPT was also
increased in pulmonary endothelium from VILI-exposed mice (D) compared to spontaneous
breathing mice (C).
Figure 2. Excessive mechanical stress increases NAMPT expression in vitro. (A) mRNA
levels, as measured by RT-PCR, showed a significant increase in NAMPT transcription in EC
exposed to 18% CS, while significant changes were not observed in EC exposed to 5% CS (*
p<0.05 vs. 5% CS). (B) 18% CS significantly increased full length NAMPT promoter activity in
EC (* p<0.05 vs. 0h).
Figure 3. Functional analysis of NAMPT promoter activity following EC exposure to 18%
CS. A NAMPT promoter region sensitive to 18% CS was identified using a series of nested
Page 13 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

14
deletion constructs in conjunction with luciferase reporter activity assays. Each experiment was
repeated 4 times and the mean ± SD is shown in each bar graph (* p<0.05 vs. static).
Figure 4. 18% CS significantly increases STAT5 binding to the NAMPT promoter. qPCR of
ChIP was performed for STAT2, STAT3 and STAT5. Primers for the 5’- (A) and 3’- (B) STAT
binding sites on the NAMPT promoter were designed to capture in silico transcription factor-
binding motifs that were specific to STAT family transcription factors. The sites of these motifs
were located in the mechanical stress-inducible region. STAT occupancy was normalized to the
change in qPCR product of static input fractions and 18% CS input fractions (* p<0.05 vs. input).
Figure 5. Demethylation significantly increases NAMPT transcription and recruitment of
STAT5. (A) 5’-Aza significantly increased mRNA levels of NAMPT (* p<0.05 vs. 0 µM). (B)
NAMPT promoter demethylation by 5’-Aza significantly increased STAT5 and STAT3 binding to
NAMPT promoter (* p<0.05 vs. Control).
Figure 6. Influence of mechanical stress, STAT5 and ARDS-associated variants on
NAMPT promoter activities. Activities of the NAMPT promoter containing the recently
identified risk alleles for ARDS in Chicago or Spanish ARDS cohorts (-948T or -2422G,
respectively) were assessed. NAMPT promoter activity was significantly increased in NAMPT
constructs containing either -948T or -2422G upon response to 18% CS, compared to the
“wildtype” NAMPT promoter constructs. Reductions in EC STAT5 protein levels (using siRNA
against STAT5a/b compared to negative control siRNA) (inset), partially or fully abolished the
effects of -948T and -2422G on promoter activities in response to 18% CS (* p<0.01 vs. static, #
p<0.05 vs. wildtype, + p<0.05 vs. 18% CS).
Page 14 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

Figure 1. Exposure to high tidal volume ventilation significantly increases murine NAMPT protein expression in BAL leukocytes and lung endothelium. Compared to spontaneously breathing mice (A), protein expression of NAMPT was increased in bronchoalveolar lavage (BAL) leukocytes obtained from VILI-exposed mice (B).
Protein expression of NAMPT was also increased in pulmonary endothelium from VILI-exposed mice (D) compared to spontaneous breathing mice (C).
152x138mm (300 x 300 DPI)
Page 15 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

Figure 2
A.
0.5
NAM
PT m
RNA
1Time (hr)
*
0
1.0
2.0
3.0
1.5
0.5
2.5
0
5
7
B.
Static 1 3218% CS (hr)
4
1
*
(Fol
d Ch
ange
)
2 4 6 12 24 48
*
*5% CS18% CS
4
6
32
NAM
PT P
rom
oter
Activ
ity (R
LU)
**
*
Page 16 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

Figure 3
A.
Static18% CS
B.
NAMPT Promoter Activity (RLU)0 10 3020 40 50 60
-1528
-328
pGL3
-1228
-928
-628
*
-2128
-1828
**-3028
-2728
-2428
Mechanical stress-
inducible region (MSIR)
Mechanical stress-inducible region (MSIR)
ATG
rs59744560(-948G/T)
STAT5-3028 TSS(-346)
+1
BA
rs7789066(-2422A/G)
Page 17 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society
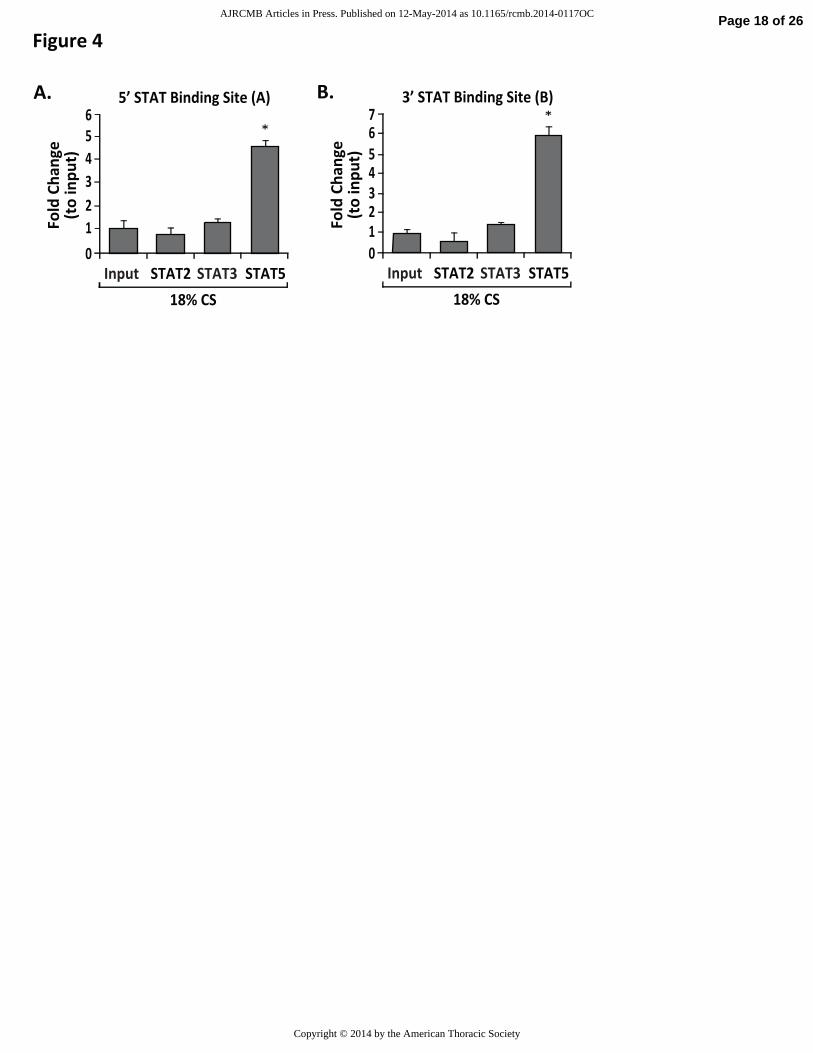
Figure 4
A.
0
56
Input STAT2 STAT5STAT318% CS
1
*
432
Fold
Cha
nge
(to
inpu
t)5’ STAT Binding Site (A) B.
0
5
7
Input STAT2 STAT5STAT318% CS
1
*
4
6
32
Fold
Cha
nge
(to
inpu
t)
3’ STAT Binding Site (B)
Page 18 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

A.
0
3
4
STAT3 STAT5
1
*
2
Fold
Cha
nge
(to
inpu
t)
Control5’-Aza
*
Figure 5
B.
0
2
3
0 0.1 51 5’-Aza (uM)
1
*
NAM
PT m
RNA
Fold
Cha
nge
*
**
10
Page 19 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

Static18% CS
Figure 6
NAMPT Promoter Activity (RLU)
*
+
0 20 6040 80
+
*
+
100 120
#
*#
siControlsiSTAT5
siControlsiSTAT5
siControlsiSTAT5
siControlsiSTAT5
siControlsiSTAT5
siControlsiSTAT5
WT
(-242
2A/-
948G
)SN
P(-2
422G
)SN
P(-9
48T)
siControl siSTAT5
STAT5
Actin
Page 20 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

1
ONLINE SUPPLEMENTAL MATERIALS
SUPPLEMENTAL MATERIALS AND METHODS
Models of Ventilator-Induced Murine Lung Injury. All animal procedures were approved by
the University of Illinois at Chicago Institutional Animal Care and Use Committee (IACUC). Mice
were maintained in standard laboratory conditions (14/10 hr light-dark cycle) and with full
access to food and water ad libitum. Male C57BL/6J mice were anesthetized with an
intraperitoneal mix of ketamine (100 mg/kg) and xylazine (5 mg/kg), exposed to ventilation for 4
hours (room air; tidal volume, 40 ml/kg, 65 breath/min, 0 cm H2O positive end-expiratory
pressure), as previously described, to induce VILI (11). In addition to the high tidal ventilation
group (VILI) (n=4–6 for all groups), spontaneously breathing (SB) mice served as a control
group. After the 4-hour ventilation, mice were euthanized with ketamine and xylazine followed
by cervical dislocation. Bronchoalveolar lavage (BAL) and lung tissue were collected from these
mice and stained (IHC) with goat polyclonal anti-NAMPT/PBEF antibodies (Lampire Biological
Laboratories, Pipersville PA) as previously described (6).
Chromatin Immunoprecipitation (ChIP) and qPCR. Cross-linked protein/DNA was sheared
using three pulses (fifteen seconds each) in a VirSonic 60 sonicator (SP Scientific/VirTis,
Warminster PA) and qPCR of DNA performed in a CFX384/C1000 Thermal Cycler (BioRad,
Hercules CA). Two STAT transcription factor-binding sites in the NAMPT promoter were
identified in silico with Genomatix software (Genomatix Software GmbH, Munich, Germany).
Primers were designed to capture these two regions using forward-BM5 (5’ –GCAGCAT
AGAACAACGGTCA) and reverse-BM7 (5’ -TTTTTGTCCTTAATTTGGTGTTTTT) yielding a
product of 131 base pairs that captures the proximal STAT binding site, and forward-BM6 (5’–
GCCCGGCCTAAACAAATTAT) and reverse-BM8 (5’–AAAAACACCAAATTAA GGACAAA)
yielding a product of 117 base pairs that captures the distal STAT binding site. PCR was
performed using SsoFast EvaGreen Supermix (BioRad) as per manufacturer’s protocol. An
initial quantification was derived using the 2-∆∆Ct method from both stretched and unstretched
(static) cell lysate that had been immunoprecipitated using STAT2, STAT3, and STAT5
antibodies, as well as rabbit IgG antibody (values not shown) as a negative control. As a
measure of the overall change in quantity of the PCR target in stretched versus static conditions
the ratio of stretched 2-∆∆Ct versus the static 2-∆∆Ct was calculated for each fraction (input
fraction, STAT2 IP, STAT3 IP, and STAT5 IP). Finally each change in quantity was then
Page 21 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

2
normalized to the change in quantity of the input fraction. Errors were expressed as a standard
error of the mean.
Gene Cloning, Mutagenesis, and 5’-Deletion Mutations. 3028 bp DNA fragments of the
NAMPT promoter region (−3028 bp to +1 ATG) were amplified by PCR using the human
genomic DNA as template. Amplicons were modified by site-directed mutagenesis to generate
fragments containing minor alleles at these loci. All allele-containing fragments were fused to a
pGL3-basic reporter vector (Promega, Madison WI). To further study the putative cis-elements,
a series of mutant plasmids were constructed sparing the transcription start site (TSS) by PCR
amplification with primers. The DNA fragments containing the truncated region were inserted
into the XhoI and MluI sites of pGL3, and the relevant regions of the final constructs were
confirmed by sequencing.
Association Study of Populations and Demographics. DNA samples from a previously
described Chicago cohort (36) included samples from 71 European descent (ED) and 34 African
descent (AD) subjects with severe sepsis-associated ARDS, as well as 186 ED and 185 AD
healthy unrelated controls. DNA samples from a second previously described Spanish cohort
(37) consisted of 96 population-based controls and 80 severe sepsis-associated ARDS cases
collected from post-surgical and intensive care units. Both Chicago and Spanish cohort subjects
met sepsis-associated acute lung injury definitions as proposed by the American–European
Criteria (38) and the Society of Critical Care Medicine Consensus statements (39). All above
studies were approved by the University of Illinois at Chicago IRB and Ethics Committees of
Hospital Universitario N. S. de Candelaria (Spain), and written informed consents were obtained
from all participants.
Genotyping for Association Studies. Genotyping of Chicago samples was conducted using
the iPLEX GoldTM Platform (Sequenom, San Diego CA) and TaqManTM allelic discrimination
assays (Applied Biosystems, Foster City CA). Briefly, iPLEX™ assays were subjected to
MALDI-TOF mass spectrometry and individual SNP genotype calls were automatically
generated using Sequenom TYPER 3.4™ software. TaqMan genotyping was performed using a
7900HT Fast Real-Time PCR System (Applied Biosystems) with automated calls generated
using the SDS software based on discriminating plots (95% confidence). Approximately 10% of
the samples were genotyped in duplicate to monitor genotyping quality. Genotyping was blind to
Page 22 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

3
case-control status and ethnic background of the samples. A follow-up study of the two SNPs in
the Spanish samples was performed using TaqManTM allelic discrimination assays (Applied
Biosystems) with the same protocols and equipment as described above.
Bisulfite Modification and Sequencing of NAMPT Promoter. To evaluate the epigenetic
regulation of the NAMPT promoter through DNA methylation in response to 18% CS, genomic
DNA was isolated from static-, 5% CS- and 18% CS-exposed EC. The DNA was bisulfite
converted (Zymo Research, Irvine CA) and the NAMPT promoter was amplified using primers
specific for bisulfite-converted DNA utilizing a nested PCR approach. The amplified products
were verified on an agarose gel and extracted as per manufacturer’s instructions (Omega Biotek,
Norcross, GA). The extracted DNA was then cloned into pGEM-T Easy vectors (Promega) and
DNA from several clones sequenced to determine the methylation status of CpG sites at single
nucleotide resolution.
SUPPLEMENTAL TABLE
Supplemental Table 1. Alignment of NAMPT 5’UTR in Six Vertebrates Species Sequence
Human CTCGGGCCGGAGGACAGGGGCCG-CGCGCCGCGAGCTCCCTGGCGCGGCTGCGAGGAAGGAGAAAAATGAG
Cow CTCGGGCGGGAGCGTGGGGGCCG-CGCGCCGCGAACTGCCCGGCGCGGCTGCGAGGAAGGAGAAAAATGGG
Mouse CTCGGGCCGGAGC---GGAGACGCCGCGCCACGAGCTGCCCGGCGCGGCTGCGAGCAAGGAGAAAAATGGG
Rat CTCGGGCCAGAGC---GGAGGCGCCGCGCCACGAGCTGCCCGGCGCGGCTGCGAGCAAGGAGAAAAATGGG
Chimp CTCGGGCCGAAGGACAGGGGCCG-CGCGCCGCGAGCTCCCTGGCGCGGTTGCGAGGAAGGAGAAAAATGAG
Human CTTCACCGCGCTCCGTTGCTTAAGTCACTGCTCGGTCGGC---GGA
Cow CTTCACCGCGCTCCGTTGCTTAAGTCACCGATCGGTCGGAGGG---
Mouse CTTCGCTGCGCTCCGTTGCTTAAGTCACCGCTCGCTCGCAGGG---
Rat CTTCGCTGCGCTCCGTTGCTTAAGTCACCGCTCGCTCGCAGGG---
Chimp CTTCACCGCGCTCCGTTGCTTAAGTCACTGCTCGGTCGGCGGAGGA
Human GGAGGGGGAGAGGGGGAAACGGAGAGA-GGGGAGGGGTCAGAGGAGGGCGGGC
Chimp GGAGGGGGAGAGGGGGAAACGGAGAGA-GGGGAGGGGTCAGAGGAGGGCGGGC
Rat -GGGCGGGAGAGCAGGGAACCGAGCAC-GGGGAGGGGCC--GGGAGGGCGGG-
Mouse -GGGCGGGAGAGCAGGGAACGGAGCGC-GGGGAGGGGCC--GGGAGGGCGGG-
Cow GGGTAAGAGGAGGGGGAAAGGGAGCGAGTAGGAGGGGCCAGAGGAGGGCGGTC
Human CCGGGAGCCGTGACGCGGCGCGGGTGACGGCTGCGGCGGCT-----------------
Chimp CCGGGAGCCGTGACGCGGCGCGGGTGACGGCTGCGGCGGCT-----------------
Rat --CAGGTCCGTGACGCGGCG------------GCGGAGGAA-----------------
Mouse --CAGGTCCGTGACGCGGCG------------GTGGAGGCT-----------------
Cow CCGGGAGCCGTGACGCGGCGAGGGAA------GCGGCGGCGGTGGAGCGGCGGCGGAG
Human ----CGCGTGCT-CGCAGTCTGGGAGCT-CTGGCGGACTCCCCACCTCGGTTCCCCCG
Cow GCGGAGCGCGCCGGCAATCTAGGGACTCCCGGCGGCCTCCTCACCCCGGCT-GCCCGC
Mouse ----AGCGCGCTCCCCTGCCCAGGCGCT-CCGGCGGCTTTCCCG--GCGGCTCCGCGC
Rat ----AGCGCGCTACACGACCCAGGCGCT-CCGGCGGCTTTCCCG--GCGGCTCCGCGC
Chimp ----CGCGTGCT-CGCAGTCTGGGAGCT-CTAGCGGACTCCCCACCTCGGTTCCCCCG
Human CCTTCACCCCGTCACCCTCCGGGGGCCGAGAAAGGGCGGGGCGCGGCAGCgcgctgcgcagtgcgc
Cow CCCCG-TCCCATCGCTCCGCCGGGGACGCGA-GGGGCGGGGTGAGGCCGCgcg-tgcgcagatagc
Mouse CCC---GCCCATCCC-------GGG--GACAAGGGGCGGGGCACGGCTGCgcg-tgcgcagcgcag
Rat CCC---GCCCATCCC-------GGG--GATAAGGGGCGGGGCACGGCTGCgcg-tgcgcaacgcag
Chimp CCTTCACCCCGTCACCCTCGGGGGGCCGAGAAAGGGCGGGGCGCGGCAGCgcg-tgcgcagtgcgc
Page 23 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

4
Human ggaggcggggcg-gg--gaggaggacgtgatgcacgcgctcttcctcccagac
Cow gggggcggggt------gggaaggacgtgaggcacgcgcctttctc-ggagac
Rat --gggcggggggttgccgaggaggacgtgaggcacgcgctctgcctcggcggc
Mouse --gggcgggggg-gactgaggaggacgtgaggcacgcgctctgcctcagctgc
Chimp ggaggcggggtg-gg--gaggaggacgtgatgcacgcgctcttcctcccagac
Human gccagctctgggaagctggaggcagcggggcagccccggcgcg-tgacccgggcgcttacctaag
Cow gcccaccccgggaagccggaggctgaggggccgccccggc-ccgtgacccctgagcacgcttggc
Rat gcccgccccaggc--ctaaagccg--acagtcaccccagcgcg-tgacctgggcgcctgcctctg
Mouse gcccgccccgggc--ctaaagccg--accgtcaccccagcgcg-tgacctgggcgcctgcctctg
Chimp gccagctccgggaagctggaggcggcggggcaaccccggcgcg-tgactcgggcgcttacctaag
Human ttcgagttccc ggcacgggcgcggg------agggcggggcctg------ga-ggg-------ggcgttc
Cow tttcatttccc ggccc-tacgctgcggggtgggggcggggccgg------gagggg-------ggcgttc
Rat ------------------------a------ggggcgtggtttgatataggg-gcgt ggcttgggcgttc
Mouse ------------------------a------ggggcgtggtctgatgcaggg-gcgt ggctt-ggcgttc
Chimp ttcgagttccc ggcacgggcgcggg------agggcggggcctg------ga-ggg-------ggcgttc
Human ccagctttgccagtgccacgaggagccggttcgcccgccccgcctgggaccttcc----g
Chimp caagcttcgccagtgccacgaggagccggttcacccgccccgcctgggaccttcc----g
Mouse ccagcctcgcccctcctgccgggagtctctttgcccgccccaacccggaccttcctcttg
Rat ccagcctcgcccctcctgtcgtgaacctctttgcccgccccagcccggaccttcctcttg
Cow ccagactcgccactcctacgattagcctgttcgcccgccccg-ctggcaccttcc---ta
Human tcc-tacccagtcctggccggttttctgggtcctcctgaagtcacgccacccggctagggggcgagga
Cow tccttacccagtcctggcccgttttctgagtcctcctgaagtcacgccact-ggctgggggctgagga
Rat tcc-tacccagtccctgctcgtcctgagagtcttgttggagtcacgccacctagctaaggcc-aagga
Mouse tcc-tacccagtccctgctcgtcctgagagtcttgttggagtcacgccacc-agctaaggcc-aagga
Chimp tcc-tacccagtcctggccggttttctgggtcctcctgaagtcacgccacccggctagggggcgagga
Human gcctcctactgcccatcttcccgt-ccaccacgcgcag ttactc
Cow gcctccctctgtccgtctccccgt-ccccctcgcgcag ttcctc
Rat gccaccca---------ctcttag-ttcccgcacgcgc tgaatt
Mouse gccacccactgtcgt--ctccctg-ttcccgcacgcgc tgaatt
Chimp gcctcctactgcccatcttccagt-acaccacgcgcag ttactc
Human acctttgtctccggcctggattaaggatccagcctttcgcctccatccctcttgtccc
Chimp acctttgtctccggcctggattaatgatccagcctttcgcctccatccctcttgtccc
Rat acctttgtctttcgtttggaccgtggatggatcctttggtcaccatccgtcttgtcga
Mouse acctttgtctcccgcttggacgatggatggaatctttggtccccatccgtcttgtgga
Cow acctttgtctctggctcagattcaggatcgagccttttgccttcatccctcgtgtctc
Human tcctgatcctcctgaccctgtctttaagatcccaggagctgcggtgaggagtgagg
Chimp tcctgatcctcctgaccctgtctttaagatcccaggagctgcggtgaggagtgagg
Mouse ttccgatcctctagattttcc-tttaaaatcagaggagctgct--------tgcgg
Rat tcctgatcctctagattttct-tttgaaataagaggagctgct--------tgaag
Cow tcctggtcctcctaaccgt--ctttaaaatctggagcgctgcgctgaggaacaagg
Human -ctgaggggcccctttcatctgatgcagcgactccgctttcctccggcggctct
Cow gctgaggggtttgcttaacctgactcatcggctccactttcctc---cacc---
Rat tctgag--------ttcccctggtgcacggccttgctttcctcc---ctgctct
Mouse tctgag--------ttgccctggcgcgtggacttgcttttctcc---gcgctca
Chimp -ctgaggggcccctttcatctgacgcagcgactccgctttcctccggcggctct
Human gtctatggctgagctctttgatcctttgagagatggtttgacttttcccga
Chimp gtctatggctgagctctttgatcctttgaggaatggtttgacttttcccga
Mouse gtctgtagctgagctctttgatcctttgaaggctg----acttttcctgga
Rat gtctgtagctgagctctttgatcctttgaagactg----acttttcctgga
Cow -tctgtgactgagctctttgatcctttga-ggatgttttgacttttcccga
Human gca---------aagagcctgcgttgaaaag-cgggggtggaattc
Chimp gca---------aagagcctgcgttgaaaag-cgggggtggaattc
Mouse gaa---------aagcctttgcgtctt-agg-gaaaggttaaattc
Rat gaaagtcttaacagtcctttgcaccttgagg-ggaaggttaaattc
Cow gaa---------aatagcctgcacccaaattcgggtgttgaattc
Human agtcctcacagataatgaggggacaagacctaattgaaccgagtattgccgg
Chimp agtcctcacagataatgaggggacaagacctaattgaaccgagtattgccgg
Mouse aaccctctcagacaatgtagggacaagaccaaattgaatctaatattgttgg
Page 24 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

5
Rat aaccctctccgacaatgaaaggacaagacc-aattgaatccagtattgttgg
Cow agtcccgacagacaatcagaggataagacccagttgagtctagtttgccgag
Human gaaggaaaaggcaacgggccaagcctttgacagggtgcgacactgactttta--------
Chimp gaaggaaaaggcaacgggccaagcctttgacagggtgcgaaactgactttta--------
Mouse gaagtaaaaggcatacagccaagtct----tatacagtggatctgttttttggttttttg
Rat gaagtaaaaggcatccagccaagtct----tagacagaggatctgtt-------------
Cow gagtaattggcaacgggccacagtctttgacagggtgcgagactgacttgtg--------
Human tcatc-gttatagtctttaaatcct-gggaaacgagttg gcaacccca
Chimp tcatc-gttatagtctttaaatcct-gggaaacgagttg gcaacccca
Mouse ttttc-ctttaaatcattaaatcgtgggggaaagagcta gtaacctcc
Rat ttttc-ctttaaatcattaaatcct-ggggaaagagcta gcaaccttc
Cow tcatc-gctgtaatcgctcaatctt-gggaaacgggttg gcaaactgg
* Alignment of NAMPT from -1 from ATG (start of CDS) to -1058 from ATG (NM_005746)
(http://genome.ucsc.edu).
SUPPLEMENTAL FIGURE LEGEND
Supplemental Figure 1. Mechanical stress-induced demethylation of the NAMPT
promoter. Genomic DNA from EC that were unstretched (static), cyclic stretched at
physiological levels (5%), or cyclic stretched at injurious levels (18%) were bisulfite converted.
PCR was performed with primers specific for the CpG island in the mechano-sensitive region of
the NAMPT promoter with the converted DNA as a template. PCR products purified from the gel
were cloned into pGEM-T easy vector and multiple clones were sequenced to identify
differentially methylated CpG residue in the mechano-sensitive region of the NAMPT promoter.
Each column represents a unique CpG site while each row represents a different clone. Black
dots and white dots represent methylated and unmethylated CpG sites, respectively. At site 15,
18% CS induces demethylation compared with static or 5% CS EC.
Page 25 of 26 AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society

STATIC 5% CS 18% CS
MSIR
CpG Island CpG Island
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Supplemental Figure 1 Page 26 of 26
AJRCMB Articles in Press. Published on 12-May-2014 as 10.1165/rcmb.2014-0117OC
Copyright © 2014 by the American Thoracic Society
Top Related
Copyright © 2022 FDOKUMEN

