







Bahasa
Halaman
Hukum


Forest Ecology and Management 339 (2015) 34–43
Contents lists available at ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/ locate/ foreco
The development of universal response functions to facilitate climate-smart regeneration of black spruce and white pine in Ontario, Canada
http://dx.doi.org/10.1016/j.foreco.2014.12.0010378-1127/Crown Copyright � 2014 Published by Elsevier B.V. All rights reserved.
⇑ Corresponding author.E-mail addresses: [email protected] (J. Yang), [email protected].
ca, [email protected] (J.H. Pedlar), [email protected], [email protected] (D.W. McKenney), [email protected] (A. Weersink).
Jing Yang a, John H. Pedlar b,⇑, Daniel W. McKenney b, Alfons Weersink a
a Dept of Food, Agricultural and Resource Economics, University of Guelph, Guelph, Ontario N1G 2W1, Canadab Canadian Forest Service, Great Lakes Forestry Centre, 1219 Queen Street E., Sault Ste. Marie, ON P6A 2E5, Canada
a r t i c l e i n f o a b s t r a c t
Article history:Received 3 September 2014Received in revised form 12 November 2014Accepted 2 December 2014
Keywords:Climate changeSeed deploymentSeed procurementUniversal response functionBlack spruceWhite pine
Climate change is expected to impact forest growth and survival as tree populations experience climatesto which they are not well adapted. However, forest regeneration efforts represent opportunities to intro-duce genetic material on the landscape that is well matched to future climates. We estimated universalresponse functions (URF) for two economically and ecologically important tree species in eastern Canada– black spruce (Picea mariana) and eastern white pine (Pinus strobus). The URF approach models treegrowth as a function of both the planting site climate (i.e., environmental effects) and seed source climate(i.e., genetic effects), allowing the growth of any seed source to be estimated at any location. Both speciesexhibited a relatively weak genetic effect, suggesting modest potential for seed movements to mitigateclimate change impacts. The optimal provenance for any given location was not local, but rather locatedin environments that were approximately 1.5 �C warmer than the planting site; this shift is consistent inboth direction and magnitude with that expected due to climate warming since the start of the industrialrevolution. Both species exhibited a strong environmental effect with clear evidence for a central climaticoptimum. As a result, climate warming is expected to enhance black spruce growth at sites with meanannual temperature colder than approximately 4.5 �C and white pine growth at sites colder than 11 �C.Thus, Ontario white pine populations may benefit from the climate changes projected to occur in theprovince over the next 50–100 years. For black spruce, populations in southern and central Ontariomay experience suboptimal climate conditions over the mid-to-end of the current century. Despite therelatively weak genetic effect, climate-smart seed movements could play a role in maintaining the pro-ductivity and genetic diversity of black spruce in these areas. While the URF approach has limitations, thiswork demonstrates its potential for informing regeneration decisions under climate change.
Crown Copyright � 2014 Published by Elsevier B.V. All rights reserved.
1. Introduction
Tree species often exhibit adaptive genetic variation along cli-matic gradients or clines (Morgenstern, 1996 and Rehfeldt et al.,1999). For example, Lu et al. (2003) reported significantly higher coldtolerance in white pine populations from northern Ontario as com-pared to those from southern Ontario. Similarly, black spruce popu-lations from northern Quebec set their buds in fall at an earlier datethan southern populations (Beaulieu et al., 2004). Such variationallows populations to survive and grow well under local climate con-ditions; however there are also instances in which regenerativematerials have shown optimal growth under climatic conditions thatdiffer from their site of origin (e.g., Thomson and Parker, 2008).
Given these strong ties to climate, it is expected that recent andanticipated climate change will have impacts on forest growth andsurvival. In the long term, species may be able to adapt and migratein response to climate change, but in the short term locally adaptedpopulations are likely to experience climates to which they are notwell adapted (Matyas, 1994, Wang et al., 2006; Leites et al., 2012and Williams and Dumroese, 2013). Therefore, seed deploymentand procurement strategies that are part of a long-term, sustain-able reforestation plan may need to be modified to ensure planta-tions are well matched to future climates (Wang et al., 2006, 2010)while minimizing current risks.
Provenance studies, in which parent material from a range ofgeographic and climatic origins (or provenances) are grown atone or more planting sites, provide valuable information onpatterns of forest genetic variation in relation to climate(Morgenstern, 1996). Here we use the term population to refer toa set of intermating individuals that live in the same area andthe terms provenance and seed source to describe the geographic

J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43 35
and climatic origin of a forest tree population (Morgenstern, 1996).The approaches used by forest geneticists to analyze and summa-rize provenance data have evolved over time. Simple responsefunctions typically employ a quadratic function to model the rela-tionship between a measured provenance characteristic (e.g.,height growth) and climate (e.g., mean annual temperature) – thusillustrating how a given population grows across a range of condi-tions (Rehfeldt et al., 1999; Wang et al., 2006 and Thomson andParker, 2008). Alternatively, a simple transfer function showshow a selection of seed sources grow at a particular site (Matyas,1994; Carter, 1996 and Rehfeldt et al., 1999). These approaches arelimited in that they summarize information for only a single seedsource or planting site. To address this limitation, transfer functionshave been pooled across planting sites to provide insights into seedmovements across the range of a species (Rehfeldt et al., 2003 andAndalo et al., 2005). However this approach assumes that all popu-lations respond similarly to climate transfers, which is often notthe case. Wang et al. (2010) developed a universal response function(URF) which models provenance growth as a function of both theplanting site climate (i.e., environmental effects) and seed source cli-mate (i.e., genetic effects). The URF model allows forest growth incurrent and future climates to be predicted for any population inany location as a function of the climates at the planting sites andprovenance sites, effectively combining the transfer function andresponse function into a single model.
Here we develop URFs for two economically and ecologicallyimportant tree species in eastern Canada – black spruce (Piceamariana) and eastern white pine (Pinus strobus). For this effort,we employ range-wide provenance data from trials establishedover several decades by various research organizations. This isone of the first studies to apply the URF approach to tree speciesin eastern North America and our findings should provide valuableinformation to forest managers regarding seed movements of theseimportant forest species under climate change.
2. Methods and materials
2.1. Description of provenance and climate data
Black spruce provenance data was obtained from remeasure-ments on a portion of the Canadian Forest Service’s (CFS) long-term
Fig. 1. Location of the black spruce planting sites (black triangle
black spruce provenance trial, which originally incorporated 202populations across 34 test sites in Canada and the United States(see Selkirk, 1974 for details). The remeasurements were carriedout in 2003 (33 years of age from seed) and involved measuringheight and DBH of all surviving trees at each test site (seeThomson et al., 2009 for details). In total, 192 provenances at 18test sites in Canada and one test site in Minnesota were measured(Fig. 1). Mean height values for each provenance at each test sitewere generously provided by Dr. William Parker at LakeheadUniversity.
White pine provenance data were obtained from two sources.The first was a provenance trial initiated by the United StatesDepartment of Agriculture (USDA) in 1955 at 13 test sites in thenortheastern United States and two in Ontario, Canada (see Kingand Nienstaedt, 1969 for details). Regional remeasurements ofthese test sites were reported at age 16 for 10 test sites in thenortheastern U.S. (Demeritt and Kettlewood, 1976); at age 16 forthree test sites in Maryland (Genys, 1983, 1987); and at age 28for two test sites in southern Ontario (Abubaker and Zsuffa,1990). The second source for white pine data was a provenancetrial initiated by several cooperating agencies in 1964 (Wrightet al., 1979). This trial included 41 seed sources from the southernAppalachian region that were planted at one or more of 11 testsites located between 37.1 and 44.5 �N and 79.7 and 95.9 �W.Height growth was measured at ages ranging from 7 to 11 yearsdepending on the test site (Wright et al., 1979). In total, informa-tion for 26 test sites and 195 provenances were obtained fromthese studies (Fig. 2).
For white pine, height measurements needed to be standard-ized to a consistent age before using them to develop a URF. Todo this, we employed published height-age equations that weredeveloped in the same geographic region as the test site beingstandardized. For example, for each provenance and test sitereported in Abubaker and Zsuffa (1990), we substituted the heightand age values measured at 28 years of age into a height-age equa-tion for white pine in Ontario (Payandeh, 1991) and solved for siteindex. This metric indicated how well a seed source was growing ata site and identified a trajectory through time for that seed source.Once the site index for each provenance at each test site was deter-mined, that site index value was substituted into the height-ageequation and the equation was solved for 16 years of age (see
s) and provenances (gray circles) used in the current study.

Fig. 2. Location of the white pine planting sites (black triangles) and provenances (gray circles) used in the current study.
36 J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43
Appendix A for worked example). A similar approach was used forthe results reported in Wright et al. (1979), but two height-ageequations provided in Carmean et al. (1989) were employed. Spe-cifically, test sites north of 40 �N were standardized using an equa-tion for Vermont, while those south of 40 �N made use of anequation for the southern Appalachians.
We modeled tree height as a function of mean annual temper-ature (MAT) and total annual precipitation (TAP) following previ-ous studies that have identified temperature (Rehfeldt et al.,1999, Wang et al., 2010, and Leites et al., 2012) and moisture gra-dients (Andalo et al., 2005; Mitton and Duran, 2004 and Sáenz-Romero et al., 2006) as important drivers of forest genetic varia-tion. In selecting these variables, we carried out a preliminaryscreening (using single variable regressions) of 56 climate vari-ables that included both monthly and annual summaries of tem-perature and precipitation.
Climate estimates were obtained by interrogating spatial mod-els of 1991–1960 climate normals of MAT and TAP at the locationof each provenance and planting site. The climate models weredeveloped by interpolating temperature and precipitation datafrom over 12,000 stations from across Canada, Alaska and the con-tiguous United States (see McKenney et al., 2011 for details). Theinterpolations were carried out using ANUSPLIN (Hutchinson,2004), which makes use of thin plate smoothing splines to developspatially continuous climate models that incorporate latitude, lon-gitude and elevation as independent variables. Withheld errorsassociated with the MAT and TAP models are approximately 1 �Cand 20% respectively (McKenney et al., 2011). Climate change pro-jections, required to illustrate the use of the URFs under climatechange, were based on outputs from the CanESM2 general circula-tion model (GCM; Chylek et al., 2011) under the 8.5 representativeconcentration pathway (RCP; Van Vuuren et al., 2011). Raw GCMoutputs were downscaled using an approach that involved add-ing coarse-scale changes (or deltas) predicted by the GCM to
1961–1990 climate normals at climate stations across North Amer-ica (see Price et al., 2011 for details).
2.2. Estimation of Universal Response Function (URF)
Universal response functions (Wang et al., 2010) integrate theeffect of climate on populations at the planting site (traditionallymodeled using transfer functions) and the response of the popula-tion to test site climate (traditionally modeled using responsefunctions) into a single model, allowing tree growth to be pre-dicted for any seed source at any location. The model can beexpressed as:
Ysp ¼ a0 þ a1Xs þ a2X2s þ a3Xp þ a4X2
p þ a5XsXp þ esp ð1Þ
where Ysp is the tree height of population p at planting site s; Xs is avector of climate variables at planting site s; Xp is a vector of climatevariables at provenance location p; a0, a1, a2, a3, a4 and a5 areparameters to be estimated; and esp is the residual. Here the vectorsXs and Xp consist of MAT and TAP measurements at each test siteand provenance respectively. We recognize that complex experi-mental structure can be addressed using a mixed-effects model(e.g., Leites et al., 2012), but have used a fixed-effects model herefor continuity with the Wang et al. (2010) approach and to reflectthe relatively simple structure of the data (i.e., lack of subsamplingwithin blocks and populations). A URF for each species, as specifiedin (1), was estimated using SPSS (IBM Corp., 2010). Additionally,two simplified models were estimated for each species: (1) aMAT-only model (with interactions) to allow visualization of thegrowth–climate response surface, and (2) a MAT-only model (with-out interactions) to examine the relative strength of environmentalversus genetic effects.
J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43 37
2.3. Applications of universal response functions
Since the universal response function incorporates the climateof the planting site and the provenance, it is possible to separatethe environmental and genetic effects of climate. The environmen-tal effect of a given climate variable on tree height can be deter-mined by taking the first partial derivative of the estimated URFwith respect to the test site climate variable,
@Ysp
@xs¼ a1 þ 2a2xs þ a5xp ð2Þ
where a1, a2, and a5 are the estimated parameters of (1), xs is an ele-ment of Xs (MATs in this case), and xp is an element of Xp (MATp inthis case).
Similarly, the genetic effect of a specific climate variable isfound by taking the first partial derivative of the URF with respectto that provenance climate variable,
@Ysp
@xp¼ a3 þ 2a4xp þ a5xs ð3Þ
where a3 and a4 are the estimated parameters of (1).The universal response function can also be used to provide
growth estimates under current and future climate. To estimatethe growth of a local provenance under current climate, plantingsite climate is set equal to that of provenance climate (i.e., MATs = -MATp and TAPs = TAPp). To predict growth of a local seed sourceunder future climate, current climate values at the site (i.e., MATs
and TAPs) are replaced with future values at the site.Finally, URFs can help to identify optimal seed sources for a
given planting site (i.e., seed procurement) and optimal plantingsites for a given seed source (i.e., seed deployment) under climatechange. For seed procurement, this can be achieved by: (1) takingthe first-order partial derivative of the URF with respect to a signif-icant provenance climate variable (e.g., MATp), (2) substituting thefuture climate value at the site (e.g., MATs) into the equation, and(3) setting the equation equal to zero and solving for MATp. Alter-natively, for seed deployment, the partial derivative can be takenwith respect to a significant planting site climate variable (e.g.,MATs), then set to zero and solved to identify optimal planting sitesfor a given seed source.
2.4. Estimation of model errors
Standard regression diagnostics, such as F, p, and R-square wereobtained from the SPSS output. For a more rigorous test of the URF
Table 1Fitted regression coefficients for three black spruce universal response functions: a full moMAT-only model, and a MAT-only model with no interactions.
Independent variable Full model MA
Coefficient Partial R2 Coe
Intercept 8.008 7.TAPs �0.006 0.011MATs 0.672 0.240 0.TAPp 0.006 0.006MATp 0.092 0.012 0.TAPs
2 1.408E�6 0.005MATs
2 �0.062 0.041 �0.TAPp
2 �3.649E�6 0.043MATp
2 �0.021 0.001 �0.TAPs � TAPp 1.315E�6 0.003MATs �MATp 0.MATs
2 �MATp2 �0.001 0.004
MATs2 �MATp
2 0.012 0.007Model R2 0.373
Notes: the subscripts ‘‘s’’ or ‘‘p’’ denote test sites or provenances respectively. All variableinteractions were specified in the models; only interaction terms that were significant i
models, we also estimated their predictive accuracy and bias using10-fold cross validation. This approach involved the followingsteps: (1) run the model using all the data and determine the sig-nificant terms in the full model; (2) create 10 datasets that ran-domly split the full dataset into model building (90% of the fulldata) and model testing (10% of the full data) components; (3)for each of the 10 model building datasets, generate a multipleregression model using the terms that were identified as signifi-cant in step 1; (4) using each model generated in step 3, predictheight growth at the locations in the corresponding model testingdataset; (5) calculate predictive error at each location in the modeltesting datasets by subtracting the predicted from the actualheight value; and (6) calculate the overall mean and mean absolutepredictive errors.
3. Results
3.1. Black spruce URF
For black spruce, all site (TAPs, TAPs2, MATs, MATs
2) and prove-nance (TAPp, TAPp
2, MATp and MATp2) climatic variables were statis-
tically significant along with three interaction variables (Table 1).The full model explained 37% of the total variation in 33-year oldblack spruce height, with mean annual temperature at the plantingsite accounting for 28% of the total variation. Predictive accuracy ofthe black spruce regression model was ±0.97 m (or 12% of averagetree height at age 33) with a bias of 0.13 m (Table 3).
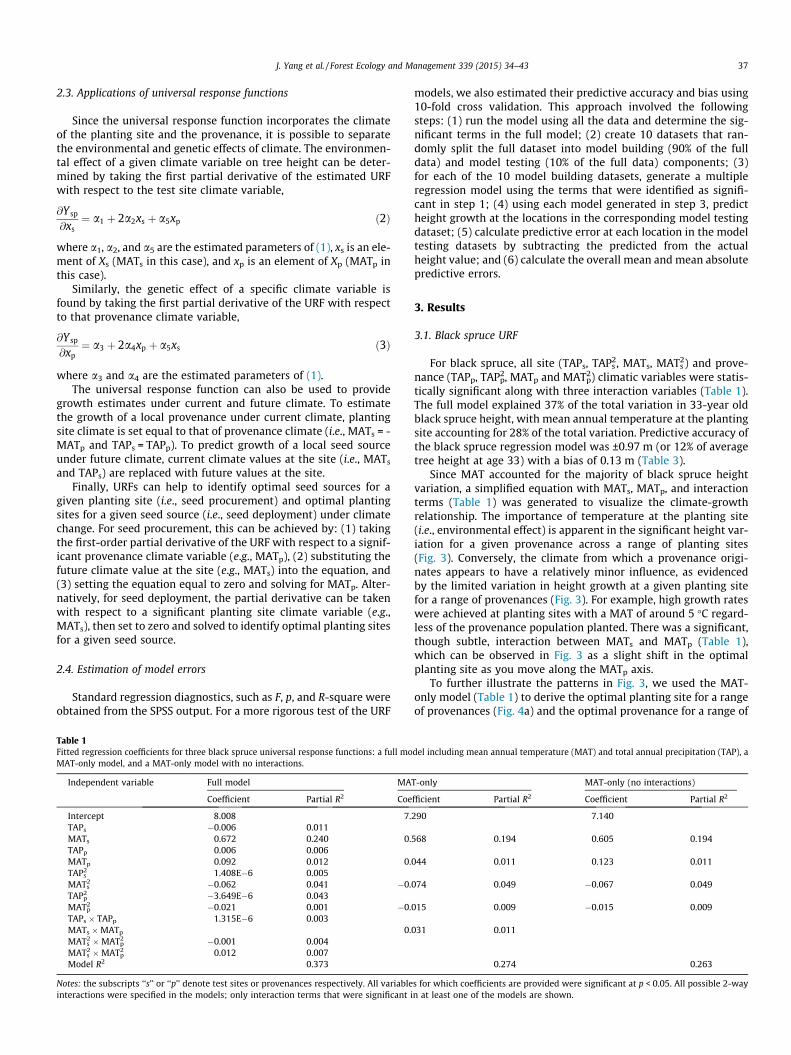
Since MAT accounted for the majority of black spruce heightvariation, a simplified equation with MATs, MATp, and interactionterms (Table 1) was generated to visualize the climate-growthrelationship. The importance of temperature at the planting site(i.e., environmental effect) is apparent in the significant height var-iation for a given provenance across a range of planting sites(Fig. 3). Conversely, the climate from which a provenance origi-nates appears to have a relatively minor influence, as evidencedby the limited variation in height growth at a given planting sitefor a range of provenances (Fig. 3). For example, high growth rateswere achieved at planting sites with a MAT of around 5 �C regard-less of the provenance population planted. There was a significant,though subtle, interaction between MATs and MATp (Table 1),which can be observed in Fig. 3 as a slight shift in the optimalplanting site as you move along the MATp axis.
To further illustrate the patterns in Fig. 3, we used the MAT-only model (Table 1) to derive the optimal planting site for a rangeof provenances (Fig. 4a) and the optimal provenance for a range of
del including mean annual temperature (MAT) and total annual precipitation (TAP), a
T-only MAT-only (no interactions)
fficient Partial R2 Coefficient Partial R2
290 7.140
568 0.194 0.605 0.194
044 0.011 0.123 0.011
074 0.049 �0.067 0.049
015 0.009 �0.015 0.009
031 0.011
0.274 0.263
s for which coefficients are provided were significant at p < 0.05. All possible 2-wayn at least one of the models are shown.
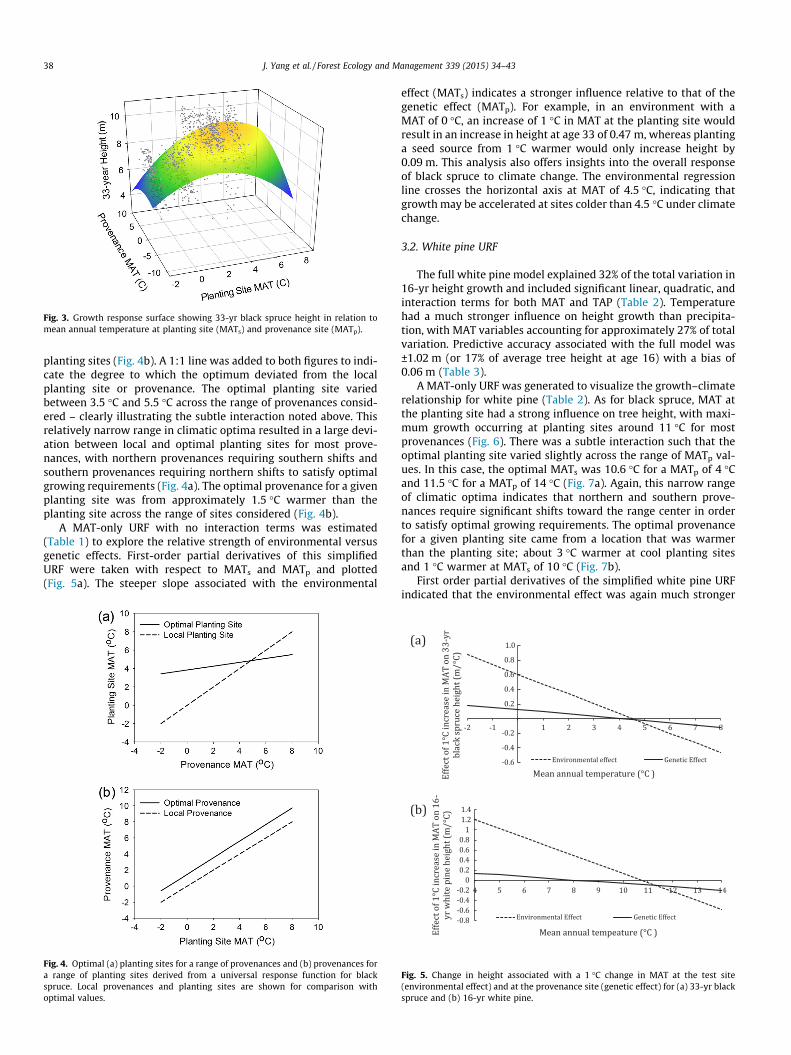
Fig. 3. Growth response surface showing 33-yr black spruce height in relation tomean annual temperature at planting site (MATs) and provenance site (MATp).
38 J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43
planting sites (Fig. 4b). A 1:1 line was added to both figures to indi-cate the degree to which the optimum deviated from the localplanting site or provenance. The optimal planting site variedbetween 3.5 �C and 5.5 �C across the range of provenances consid-ered – clearly illustrating the subtle interaction noted above. Thisrelatively narrow range in climatic optima resulted in a large devi-ation between local and optimal planting sites for most prove-nances, with northern provenances requiring southern shifts andsouthern provenances requiring northern shifts to satisfy optimalgrowing requirements (Fig. 4a). The optimal provenance for a givenplanting site was from approximately 1.5 �C warmer than theplanting site across the range of sites considered (Fig. 4b).
A MAT-only URF with no interaction terms was estimated(Table 1) to explore the relative strength of environmental versusgenetic effects. First-order partial derivatives of this simplifiedURF were taken with respect to MATs and MATp and plotted(Fig. 5a). The steeper slope associated with the environmental
Fig. 4. Optimal (a) planting sites for a range of provenances and (b) provenances fora range of planting sites derived from a universal response function for blackspruce. Local provenances and planting sites are shown for comparison withoptimal values.
effect (MATs) indicates a stronger influence relative to that of thegenetic effect (MATp). For example, in an environment with aMAT of 0 �C, an increase of 1 �C in MAT at the planting site wouldresult in an increase in height at age 33 of 0.47 m, whereas plantinga seed source from 1 �C warmer would only increase height by0.09 m. This analysis also offers insights into the overall responseof black spruce to climate change. The environmental regressionline crosses the horizontal axis at MAT of 4.5 �C, indicating thatgrowth may be accelerated at sites colder than 4.5 �C under climatechange.
3.2. White pine URF
The full white pine model explained 32% of the total variation in16-yr height growth and included significant linear, quadratic, andinteraction terms for both MAT and TAP (Table 2). Temperaturehad a much stronger influence on height growth than precipita-tion, with MAT variables accounting for approximately 27% of totalvariation. Predictive accuracy associated with the full model was±1.02 m (or 17% of average tree height at age 16) with a bias of0.06 m (Table 3).
A MAT-only URF was generated to visualize the growth–climaterelationship for white pine (Table 2). As for black spruce, MAT atthe planting site had a strong influence on tree height, with maxi-mum growth occurring at planting sites around 11 �C for mostprovenances (Fig. 6). There was a subtle interaction such that theoptimal planting site varied slightly across the range of MATp val-ues. In this case, the optimal MATs was 10.6 �C for a MATp of 4 �Cand 11.5 �C for a MATp of 14 �C (Fig. 7a). Again, this narrow rangeof climatic optima indicates that northern and southern prove-nances require significant shifts toward the range center in orderto satisfy optimal growing requirements. The optimal provenancefor a given planting site came from a location that was warmerthan the planting site; about 3 �C warmer at cool planting sitesand 1 �C warmer at MATs of 10 �C (Fig. 7b).
First order partial derivatives of the simplified white pine URFindicated that the environmental effect was again much stronger
-0.6
-0.4
-0.2
0.2
0.4
0.6
0.8
1.0
-2 -1 1 2 3 4 5 6 7 8
Eff
ect
of
1°C
in
cre
ase
in
MA
T o
n 3
3-y
r
bla
ck s
pru
ce h
eig
ht
(m/
° C)
Mean annual temperature (°C )
(a)
Environmental effect Genetic Effect
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
4 5 6 7 8 9 10 11 12 13 14
Eff
ect
of
1° C
in
cre
ase
in
MA
T o
n 1
6-
yr
wh
ite
pin
e h
eig
ht
(m/
° C)
Mean annual tempeature (°C )
(b)
Environmental Effect Genetic Effect
Fig. 5. Change in height associated with a 1 �C change in MAT at the test site(environmental effect) and at the provenance site (genetic effect) for (a) 33-yr blackspruce and (b) 16-yr white pine.
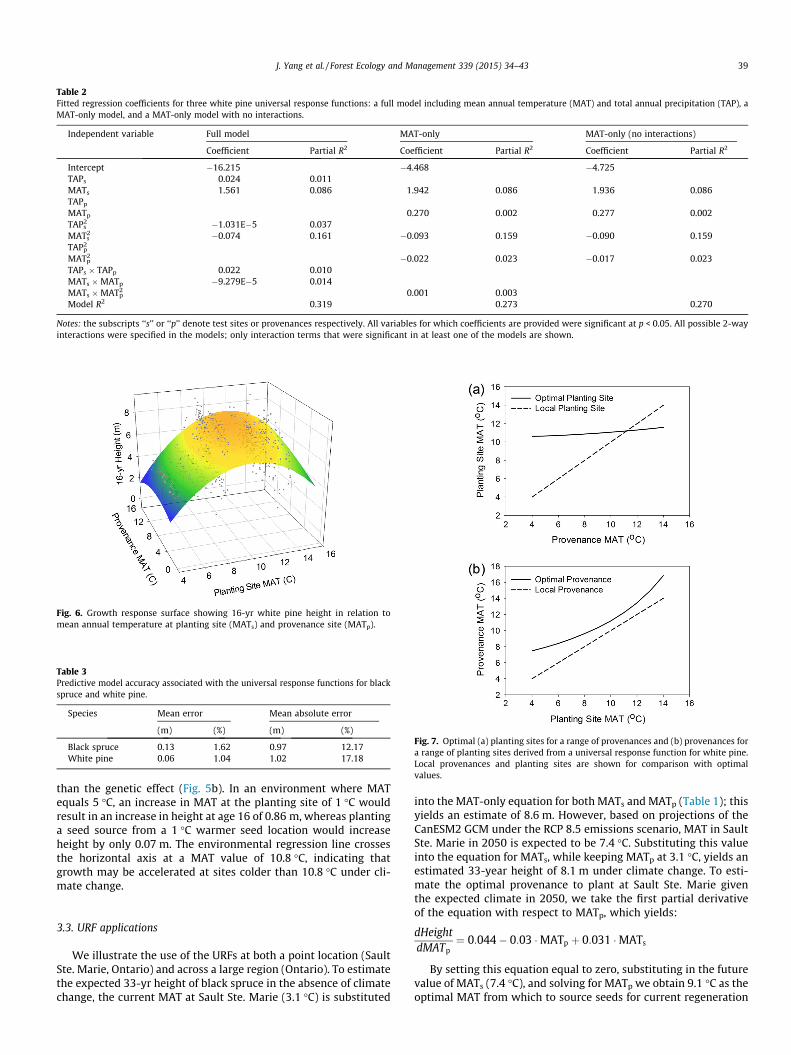
Table 2Fitted regression coefficients for three white pine universal response functions: a full model including mean annual temperature (MAT) and total annual precipitation (TAP), aMAT-only model, and a MAT-only model with no interactions.
Independent variable Full model MAT-only MAT-only (no interactions)
Coefficient Partial R2 Coefficient Partial R2 Coefficient Partial R2
Intercept �16.215 �4.468 �4.725TAPs 0.024 0.011MATs 1.561 0.086 1.942 0.086 1.936 0.086TAPp
MATp 0.270 0.002 0.277 0.002TAPs
2 �1.031E�5 0.037MATs
2 �0.074 0.161 �0.093 0.159 �0.090 0.159TAPp
2
MATp2 �0.022 0.023 �0.017 0.023
TAPs � TAPp 0.022 0.010MATs �MATp �9.279E�5 0.014MATs �MATp
2 0.001 0.003Model R2 0.319 0.273 0.270
Notes: the subscripts ‘‘s’’ or ‘‘p’’ denote test sites or provenances respectively. All variables for which coefficients are provided were significant at p < 0.05. All possible 2-wayinteractions were specified in the models; only interaction terms that were significant in at least one of the models are shown.
Table 3Predictive model accuracy associated with the universal response functions for blackspruce and white pine.
Species Mean error Mean absolute error
(m) (%) (m) (%)
Black spruce 0.13 1.62 0.97 12.17White pine 0.06 1.04 1.02 17.18
Fig. 6. Growth response surface showing 16-yr white pine height in relation tomean annual temperature at planting site (MATs) and provenance site (MATp).
Fig. 7. Optimal (a) planting sites for a range of provenances and (b) provenances fora range of planting sites derived from a universal response function for white pine.Local provenances and planting sites are shown for comparison with optimalvalues.
J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43 39
than the genetic effect (Fig. 5b). In an environment where MATequals 5 �C, an increase in MAT at the planting site of 1 �C wouldresult in an increase in height at age 16 of 0.86 m, whereas plantinga seed source from a 1 �C warmer seed location would increaseheight by only 0.07 m. The environmental regression line crossesthe horizontal axis at a MAT value of 10.8 �C, indicating thatgrowth may be accelerated at sites colder than 10.8 �C under cli-mate change.
3.3. URF applications
We illustrate the use of the URFs at both a point location (SaultSte. Marie, Ontario) and across a large region (Ontario). To estimatethe expected 33-yr height of black spruce in the absence of climatechange, the current MAT at Sault Ste. Marie (3.1 �C) is substituted
into the MAT-only equation for both MATs and MATp (Table 1); thisyields an estimate of 8.6 m. However, based on projections of theCanESM2 GCM under the RCP 8.5 emissions scenario, MAT in SaultSte. Marie in 2050 is expected to be 7.4 �C. Substituting this valueinto the equation for MATs, while keeping MATp at 3.1 �C, yields anestimated 33-year height of 8.1 m under climate change. To esti-mate the optimal provenance to plant at Sault Ste. Marie giventhe expected climate in 2050, we take the first partial derivativeof the equation with respect to MATp, which yields:
dHeightdMATp
¼ 0:044� 0:03 �MATp þ 0:031 �MATs
By setting this equation equal to zero, substituting in the futurevalue of MATs (7.4 �C), and solving for MATp we obtain 9.1 �C as theoptimal MAT from which to source seeds for current regeneration
40 J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43
efforts at Sault Ste. Marie given climate change. Note that theexpected height growth for this optimal seed source is 8.7 m – amodest improvement over the local seed source (8.1 m), whichsupports the relatively weak genetic effect described above.
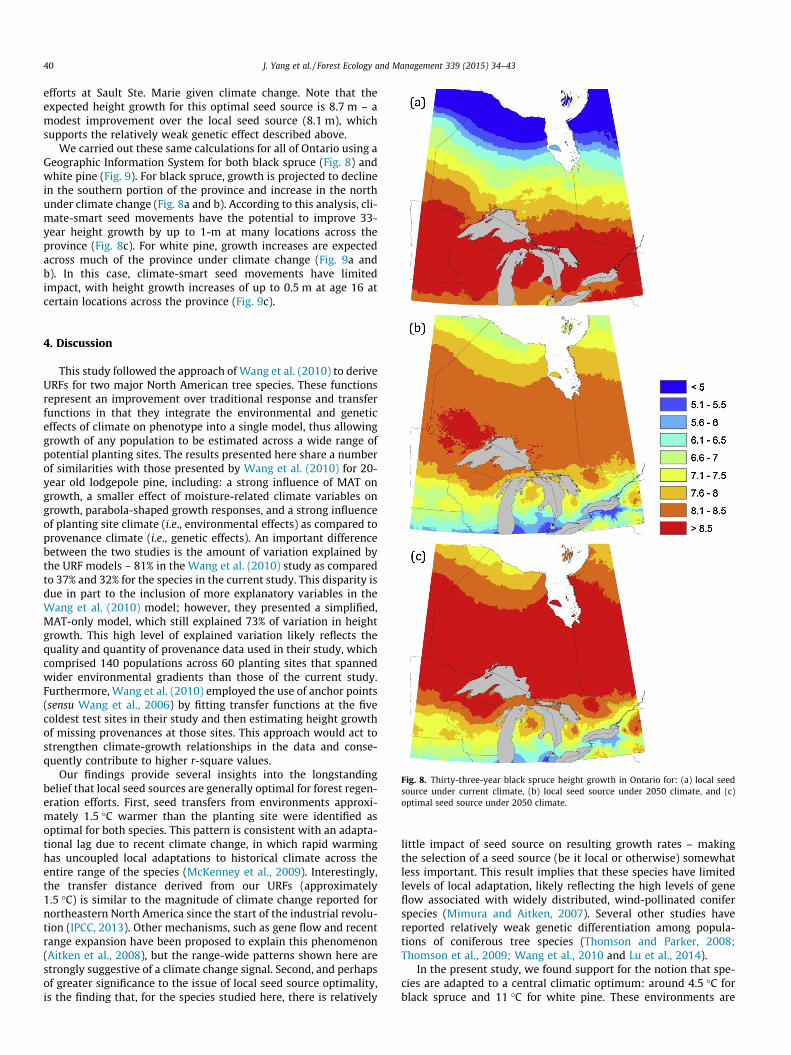
We carried out these same calculations for all of Ontario using aGeographic Information System for both black spruce (Fig. 8) andwhite pine (Fig. 9). For black spruce, growth is projected to declinein the southern portion of the province and increase in the northunder climate change (Fig. 8a and b). According to this analysis, cli-mate-smart seed movements have the potential to improve 33-year height growth by up to 1-m at many locations across theprovince (Fig. 8c). For white pine, growth increases are expectedacross much of the province under climate change (Fig. 9a andb). In this case, climate-smart seed movements have limitedimpact, with height growth increases of up to 0.5 m at age 16 atcertain locations across the province (Fig. 9c).
Fig. 8. Thirty-three-year black spruce height growth in Ontario for: (a) local seedsource under current climate, (b) local seed source under 2050 climate, and (c)optimal seed source under 2050 climate.
4. Discussion
This study followed the approach of Wang et al. (2010) to deriveURFs for two major North American tree species. These functionsrepresent an improvement over traditional response and transferfunctions in that they integrate the environmental and geneticeffects of climate on phenotype into a single model, thus allowinggrowth of any population to be estimated across a wide range ofpotential planting sites. The results presented here share a numberof similarities with those presented by Wang et al. (2010) for 20-year old lodgepole pine, including: a strong influence of MAT ongrowth, a smaller effect of moisture-related climate variables ongrowth, parabola-shaped growth responses, and a strong influenceof planting site climate (i.e., environmental effects) as compared toprovenance climate (i.e., genetic effects). An important differencebetween the two studies is the amount of variation explained bythe URF models – 81% in the Wang et al. (2010) study as comparedto 37% and 32% for the species in the current study. This disparity isdue in part to the inclusion of more explanatory variables in theWang et al. (2010) model; however, they presented a simplified,MAT-only model, which still explained 73% of variation in heightgrowth. This high level of explained variation likely reflects thequality and quantity of provenance data used in their study, whichcomprised 140 populations across 60 planting sites that spannedwider environmental gradients than those of the current study.Furthermore, Wang et al. (2010) employed the use of anchor points(sensu Wang et al., 2006) by fitting transfer functions at the fivecoldest test sites in their study and then estimating height growthof missing provenances at those sites. This approach would act tostrengthen climate-growth relationships in the data and conse-quently contribute to higher r-square values.
Our findings provide several insights into the longstandingbelief that local seed sources are generally optimal for forest regen-eration efforts. First, seed transfers from environments approxi-mately 1.5 �C warmer than the planting site were identified asoptimal for both species. This pattern is consistent with an adapta-tional lag due to recent climate change, in which rapid warminghas uncoupled local adaptations to historical climate across theentire range of the species (McKenney et al., 2009). Interestingly,the transfer distance derived from our URFs (approximately1.5 �C) is similar to the magnitude of climate change reported fornortheastern North America since the start of the industrial revolu-tion (IPCC, 2013). Other mechanisms, such as gene flow and recentrange expansion have been proposed to explain this phenomenon(Aitken et al., 2008), but the range-wide patterns shown here arestrongly suggestive of a climate change signal. Second, and perhapsof greater significance to the issue of local seed source optimality,is the finding that, for the species studied here, there is relatively
little impact of seed source on resulting growth rates – makingthe selection of a seed source (be it local or otherwise) somewhatless important. This result implies that these species have limitedlevels of local adaptation, likely reflecting the high levels of geneflow associated with widely distributed, wind-pollinated coniferspecies (Mimura and Aitken, 2007). Several other studies havereported relatively weak genetic differentiation among popula-tions of coniferous tree species (Thomson and Parker, 2008;Thomson et al., 2009; Wang et al., 2010 and Lu et al., 2014).
In the present study, we found support for the notion that spe-cies are adapted to a central climatic optimum: around 4.5 �C forblack spruce and 11 �C for white pine. These environments are
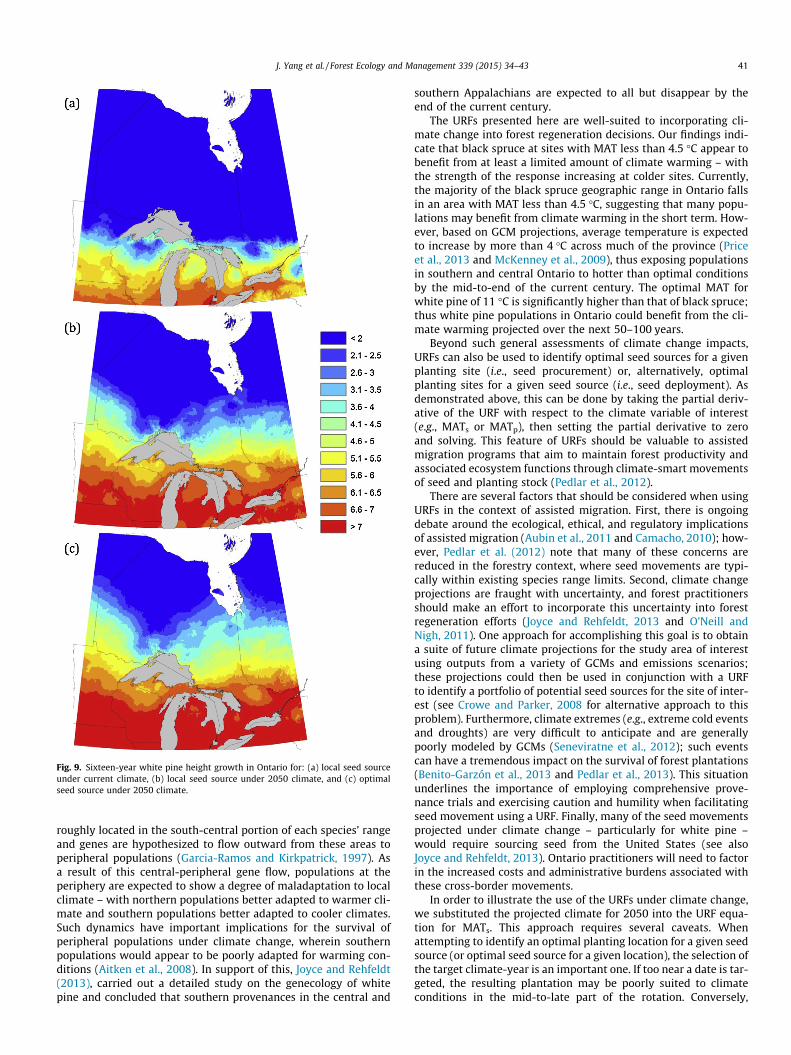
Fig. 9. Sixteen-year white pine height growth in Ontario for: (a) local seed sourceunder current climate, (b) local seed source under 2050 climate, and (c) optimalseed source under 2050 climate.
J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43 41
roughly located in the south-central portion of each species’ rangeand genes are hypothesized to flow outward from these areas toperipheral populations (Garcia-Ramos and Kirkpatrick, 1997). Asa result of this central-peripheral gene flow, populations at theperiphery are expected to show a degree of maladaptation to localclimate – with northern populations better adapted to warmer cli-mate and southern populations better adapted to cooler climates.Such dynamics have important implications for the survival ofperipheral populations under climate change, wherein southernpopulations would appear to be poorly adapted for warming con-ditions (Aitken et al., 2008). In support of this, Joyce and Rehfeldt(2013), carried out a detailed study on the genecology of whitepine and concluded that southern provenances in the central and
southern Appalachians are expected to all but disappear by theend of the current century.
The URFs presented here are well-suited to incorporating cli-mate change into forest regeneration decisions. Our findings indi-cate that black spruce at sites with MAT less than 4.5 �C appear tobenefit from at least a limited amount of climate warming – withthe strength of the response increasing at colder sites. Currently,the majority of the black spruce geographic range in Ontario fallsin an area with MAT less than 4.5 �C, suggesting that many popu-lations may benefit from climate warming in the short term. How-ever, based on GCM projections, average temperature is expectedto increase by more than 4 �C across much of the province (Priceet al., 2013 and McKenney et al., 2009), thus exposing populationsin southern and central Ontario to hotter than optimal conditionsby the mid-to-end of the current century. The optimal MAT forwhite pine of 11 �C is significantly higher than that of black spruce;thus white pine populations in Ontario could benefit from the cli-mate warming projected over the next 50–100 years.
Beyond such general assessments of climate change impacts,URFs can also be used to identify optimal seed sources for a givenplanting site (i.e., seed procurement) or, alternatively, optimalplanting sites for a given seed source (i.e., seed deployment). Asdemonstrated above, this can be done by taking the partial deriv-ative of the URF with respect to the climate variable of interest(e.g., MATs or MATp), then setting the partial derivative to zeroand solving. This feature of URFs should be valuable to assistedmigration programs that aim to maintain forest productivity andassociated ecosystem functions through climate-smart movementsof seed and planting stock (Pedlar et al., 2012).
There are several factors that should be considered when usingURFs in the context of assisted migration. First, there is ongoingdebate around the ecological, ethical, and regulatory implicationsof assisted migration (Aubin et al., 2011 and Camacho, 2010); how-ever, Pedlar et al. (2012) note that many of these concerns arereduced in the forestry context, where seed movements are typi-cally within existing species range limits. Second, climate changeprojections are fraught with uncertainty, and forest practitionersshould make an effort to incorporate this uncertainty into forestregeneration efforts (Joyce and Rehfeldt, 2013 and O’Neill andNigh, 2011). One approach for accomplishing this goal is to obtaina suite of future climate projections for the study area of interestusing outputs from a variety of GCMs and emissions scenarios;these projections could then be used in conjunction with a URFto identify a portfolio of potential seed sources for the site of inter-est (see Crowe and Parker, 2008 for alternative approach to thisproblem). Furthermore, climate extremes (e.g., extreme cold eventsand droughts) are very difficult to anticipate and are generallypoorly modeled by GCMs (Seneviratne et al., 2012); such eventscan have a tremendous impact on the survival of forest plantations(Benito-Garzón et al., 2013 and Pedlar et al., 2013). This situationunderlines the importance of employing comprehensive prove-nance trials and exercising caution and humility when facilitatingseed movement using a URF. Finally, many of the seed movementsprojected under climate change – particularly for white pine –would require sourcing seed from the United States (see alsoJoyce and Rehfeldt, 2013). Ontario practitioners will need to factorin the increased costs and administrative burdens associated withthese cross-border movements.
In order to illustrate the use of the URFs under climate change,we substituted the projected climate for 2050 into the URF equa-tion for MATs. This approach requires several caveats. Whenattempting to identify an optimal planting location for a given seedsource (or optimal seed source for a given location), the selection ofthe target climate-year is an important one. If too near a date is tar-geted, the resulting plantation may be poorly suited to climateconditions in the mid-to-late part of the rotation. Conversely,

42 J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43
targeting a date too far in the future may result in poor growth andsurvival in the early stages of plantation development. A targetdate of approximately 1/3 of rotation length has been suggestedas a reasonable balance between these competing concerns(Ukrainetz et al., 2011). Alternatively, and preferably, the URFscan be used in conjunction with annual climate projections to sim-ulate growth through time (see O’Neill and Nigh, 2011 for aworked example). This approach has the advantage of incorporat-ing the gradual changes in climate that will occur over the courseof this century; however, employing this method was beyond thescope of the current work, for which we focused on URF develop-ment and basic application.
There are a number of limitations to the current study that war-rant further discussion. First, Wang et al. (2010) point out thatURFs require extensive provenance data to fully define a growthsurface over range-wide climate conditions. While the provenancedatasets used here provide extremely valuable genecology data,they cover relatively limited environmental gradients comparedto those contained in the dataset used by Wang et al. (2010). Asa result, caution should be employed when applying the URFs toconditions that lie outside the gradients contained in the prove-nance data – MAT of approximately �2 �C to 8 �C for black spruceand 4–14 �C for white pine. Second, the URFs presented here arebased on average climate values – mean annual temperature andmean annual precipitation. While the effectiveness of these vari-ables for predicting tree growth is well documented in the litera-ture (Rehfeldt et al., 1999; Wang et al., 2006, 2010 and O’Neillet al., 2008), they may not capture all of the important climaticconstraints on plant growth). Finally, non-climatic factors (e.g.,topographic information, soil factors) were not incorporated intoour URFs, but clearly play an important role in determining siteproductivity (Wang et al., 2010 and Joyce and Rehfeldt, 2013). Ascurrently constructed, the URFs can be used to identify generalareas for locating optimal seed sources or planting sites, but prac-titioners are responsible for matching fine scale site conditionswith species’ ecological requirements.
5. Conclusions
Local seeds have traditionally been used for artificial forestregeneration, because they are assumed to be well adapted to localenvironmental conditions. However, this approach to seed selec-tion may no longer be optimal under rapid climate change and,in fact, our models suggest that existing populations may alreadybe adapted to temperatures about 1.5 �C cooler than those cur-rently found at locations across the ranges of both species. Giventhat more than 50,000 hectares of forest are planted annually inOntario alone as part of industrial forestry operations, a climate-smart seed selection strategy offers considerable potential formaintaining healthy and productive future forests. The universalresponse function, as presented here, is a tool that may assist forestmanagers in making such decisions.
Our findings indicate that climate warming may enhance blackspruce growth at sites colder than 4.5 �C and white pine growth atsites colder than 11 �C. This means that white pine in Ontario maybenefit from the climate changes projected to occur in the provinceover the next 50–100 years. In this case, limited movements ofseed from U.S. provenances could be used to diversify Ontarioplantations and help conserve southern genotypes that may be atrisk under climate change.
For black spruce, many Ontario populations may experiencesuboptimal climate conditions over the mid-to-end of the currentcentury. This is particularly concerning for populations in thesouthern and central boreal regions, which represent importantcommercial stocks, but appear to be poorly adapted to climate
warming. In this case, climate-smart seed movements could playa role in maintaining the productivity and genetic diversity of blackspruce in Ontario. While the simple universal response function inthe current study has limitations, it demonstrates the potential thisapproach has for assisting seed movements under climate change.Further testing and modification of these URFs will be necessary asmore data becomes available on species’ responses to climatechange.
Acknowledgements
We thank William H. Parker at Lakehead University and Pen-gxin Lu, Bill Parker, and Steve Colombo at the Ontario ForestResearch Institute for kindly sharing provenance data. Kevin Law-rence, Glenn Lawrence, and Kathy Campbell provided valuabletechnical support on various aspects of this project. Pierre Bernier,Glenn Fox, Brady Deaton, and two anonymous reviewers providedcomments that helped to improve the manuscript.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.foreco.2014.12.001.
References
Abubaker, H.I., Zsuffa, L., 1990. Provenance variation in eastern White Pine (Pinusstrobus L.): 28th-year results from two southern Ontario plantations. In:Proceedings of a Symposium on White Pine Provenances and Breeding IUFROMontréal, pp. 69–77.
Aitken, S.N., Yeaman, S., Holliday, J.A., Wang, T., Curtis-McLane, S., 2008. Adaptation,migration or extirpation: climate change outcomes for tree population. Evol.Appl. 1 (1), 95–111.
Andalo, C., Beaulieu, J., Bousquet, J., 2005. The impact of climate change on growthof local white spruce populations in Quebec, Canada. Forest Ecol. Manage. 205(1), 169–182.
Aubin, I., Garbe, C.M., Colombo, S., Drever, C.R., McKenney, D.W., Messier, C., Pedlar,J., Saner, M.A., Venier, L., Wellstead, A.M., Winder, R., Witten, E., Ste-Marie, C.,2011. Why we disagree about assisted migration 1: ethical implications of a keydebate regarding the future of Canada’s forests. Forest Chron. 87 (6), 755–765.
Beaulieu, J., Perron, M., Bousquet, J., 2004. Multivariate patterns of adaptive geneticvariation and seed source transfer in Picea mariana. Can. J. Forest Res. 34 (3),531–545.
Benito-Garzón, M., Ha-Duong, M., Frascaria-Lacoste, N., Fernández- Maniarrés, J.F.,2013. Extreme climate variability should be considered in forestry assistedmigration. BioScience 63 (5), 317–318.
Camacho, A.E., 2010. Assisted migration: redefining nature and natural resource lawunder climate change. Yale J. Regul. 27 (2), 171–255.
Carmean, W.H., Hahn, J.T., Jacobs, R.D., 1989. Site index curves for forest tree speciesin the eastern United States. General Technical Report NC-128. U.S. Dept. ofAgriculture, Forest Service, North Central Forest Experiment Station, St. Paul,MN.
Carter, K.K., 1996. Provenance tests as indicators of growth response to climatechange in 10 north temperate tree species. Can. J. Forest Res. 26 (6), 1089–1095.
Chylek, P., Li, J., Dubey, M.K., Wang, M., Lesins, G., 2011. Observed and modelsimulated 20th century arctic temperature variability: Canadian earth systemmodel CanESM2. Atmos. Chem. Phys. Discuss. 11 (8), 22893–22907.
Crowe, K.A., Parker, W.H., 2008. Using portfolio theory to guide reforestation andrestoration under climate change scenarios. Climatic Change 89 (3), 355–370.
Demeritt Jr., M.E., Kettlewood, H.C, 1976. Eastern white pine seed source variationin the northeastern United States: 16 year results. USDA Forest ServiceGeneticist and Forestry Technician Report. Northeastern Center ForestryExperiment Station.
Garcia-Ramos, G., Kirkpatrick, M., 1997. Genetic models of adaptation and gene flowin peripheral populations. Evolution 51 (1), 21–28.
Genys, J.B., 1983. Growth rates of different white pines studied in Maryland’sAppalachian region. Castanea 48 (4), 300–304.
Genys, J.B., 1987. Provenance variation among different populations of Pinusstrobus from Canada and the United States. Can. J. Forest Res. 17 (3), 228–235.
Hutchinson, M.F., 2004. ANUSPLIN Version 4.37, Australian National University,Canberra, Australia. <http://fennerschool.anu.edu.au/research/products/anusplin> (assessed 15.01.13).
IPCC 2013. Summary for policymakers. In: Stocker, T.F., Qin, D., Plattner, G.-K.,Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M.(Eds.), Climate Change 2013: The Physical Science Basis. Contribution ofWorking Group I to the Fifth Assessment Report of the Intergovernmental

J. Yang et al. / Forest Ecology and Management 339 (2015) 34–43 43
Panel on Climate Change. Cambridge University Press, Cambridge, UnitedKingdom and New York, NY, USA.
Joyce, D.G., Rehfeldt, G.E., 2013. Climatic niche, ecological genetics, and impact ofclimate change on eastern white pine (Pinus strobus L.): guidelines for landmanagers. Forest Ecol. Manage. 295 (1), 173–192.
King, J.P., Nienstaedt, H., 1969. Variation in eastern white pine seed sources plantedin the Lake States. Silvae Genet. 18 (3), 83–86.
Leites, L.P., Robinson, A.P., Rehfeldt, G.E., Marshall, J.D., Crookston, N.L., 2012.Height-growth response to climate changes differs among populations ofDouglas-fir: a novel analysis of historic data. Ecol. Appl. 22 (1), 154–165.
Lu, P., Joyce, D.G., Sinclair, R.W., 2003. Geographic variation in cold hardiness amongeastern white pine (Pinus strobus L.) provenances in Ontario. Forest Ecol.Manage. 178 (3), 329–340.
Lu, P., Parker, W.H., Cherry, M., Colombo, S., Parker, W.C., Man, R., Roubal, N., 2014.Survival and growth patterns of white spruce (Picea glauca [Moench] Voss)rangewide provenances and their implications for climate change adaptation.Ecol. Evol. 4 (12), 2360–2374.
Matyas, C., 1994. Modeling climate change effects with provenance test data. TreePhysiol. 14 (7), 797–804.
McKenney, D.W., Pedlar, J.H., O’Neill, G.A., 2009. Climate change and forest seedzones: past trends, future prospects and challenges to ponder. Forestry Chron.85 (2), 258–266.
McKenney, D.W., Hutchinson, M.F., Papadopol, P., Lawrence, K., Pedlar, J.H.,Campbell, K., Owen, T., 2011. Customized spatial climate models for NorthAmerica. Bull. Am. Meteorol. Soc. 92 (12), 1611–1622.
Mitton, J.B., Duran, K.L., 2004. Genetic variation in pinon pine, Pinus edulis,associated with summer precipitation. Mol. Ecol. 13 (5), 1259–1264.
Mimura, M., Aitken, S.N., 2007. Adaptive gradients and isolation-by-distance withpostglacial migration in Picea sitchensis. Heredity 99 (2), 22–24.
Morgenstern, E.K., 1996. Geographic Variation in Forest Trees: Genetic Basis andApplication of Knowledge in Silviculture. University of British Colombia Press,Vancouver, BC.
O’Neill, G.A., Nigh, G., 2011. Linking population genetics and tree height growthmodels to predict impacts of climate change on forest production. GlobalChange Biol. 17 (10), 3208–3217.
O’Neill, G.A., Hamann, A., Wang, T., 2008. Accounting for population variationImproves estimates of the impact of climate change on species’ growth anddistribution. J. Appl. Ecol. 45 (4), 1040–1049.
Payandeh, B., 1991. Plonski’s (metric) yield tables formulated. Forestry Chron. 67(5), 545–546.
Pedlar, J.H., McKenney, D.W., Aubin, I., Beardmore, T., Beaulieu, J., Iverson, L., O’Neill,G.A., Catherine, S.W., Ste-Marie, C., 2012. Placing forestry in the assistedmigration debate. BioScience 62 (9), 835–842.
Pedlar, J.H., McKenney, D.W., Aubin, I., Winder, R.S., Ste-Marie, C., O’Neill, G.A., 2013.Extreme climate variability should be considered in forestry assisted migration:a reply. BioScience 63 (5), 317–318.
Price, D.T., Alfaro, R.I., Brown, K.J., Flannigan, M.D., Fleming, R.A., Hogg, E.H.,Girardin, M.P., Lakusta, T., Johnston, M., McKenney, D.W., Pedlar, J.H., Stratton,T., Sturrock, R.N., Thompson, I.D., Trofymow, J.A., Venier, L.A., 2013. Anticipatingthe consequences of climate change for Canada’s boreal forest ecosystem.Environ. Rev. 21 (4), 322–365.
Price, D.T., McKenney, D.W., Joyce, L.A., Siltanen, R.M., Papadopol, P., Lawrence, K.,2011. High resolution interpolation of IPCC AR4 GCM climate scenarios for
Canada. Natural Resources Canada, Canadian Forest Service, Northern ForestryCenter, Edmonton, Alberta. Inf. Rep. NOR-X-421.
Rehfeldt, G.E., Ying, C.C., Spittlehouse, D.L., Hamilton Jr., D.A., 1999. Geneticresponses to climate in Pinus contorta: niche breadth, climate change, andreforestation. Ecol. Monogr. 69 (3), 375–407.
Rehfeldt, G.E., Tchebakova, N.M, Milyutin, L.I., Parfenova, E.E., Wykoff, W.R.,Kouzmina, N.A., 2003. Assessing population responses to climate in Pinussylvestris and Larix spp. of Eurasia with climate-transfer models. Eurasian J.Forest Res. 6 (2), 83–98.
Sáenz-Romero, C., Guzmán-Reyna, R.R., Rehfeldt, G.E., 2006. Altitudinal geneticvariation among< i> Pinus oocarpa</i> populations in Michoacán, Mexico:Implications for seed zoning, conservation, tree breeding and global warming.Forest Ecol. Manage. 229 (1), 340–350.
Selkirk, W.H., 1974. Origin of Provenances in the Cooperative, Range-wide BlackSpruce Study. Petawawa Forest Experiment Station, Chalk River, Ontario.
Seneviratne, S.I., Nicholls, N., Easterling, D., et al., 2012. Changes in climate extremesand their impacts on the natural physical environment. In: Field, C.B., Barros, V.,Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J.,Plattner, G.K., Allen, S.K., Tignor, M., Midgley, P.M. (Eds.), Managing the Risks ofExtreme Events and Disasters to Advance Climate Change Adaptation. A SpecialReport of Working Groups I and II of the Intergovernmental Panel on ClimateChange (IPCC), Cambridge, UK, and New York, NY: Cambridge University Press,pp. 109–230.
IBM Corp., 2010. IBM SPSS Statistics for Windows, Version 19.0. IBM Corp., Armonk,NY.
Thomson, A.M., Parker, W.H., 2008. Boreal forest provenance tests used to predictoptimal growth and response to climate change. 1. Jack pine. Can. J. Forest Res.38 (1), 157–170.
Thomson, A.M., Riddell, C.L., Parker, W.H., 2009. Boreal forest provenance tests usedto predict optimal growth and response to climate change: 2. Black spruce. Can.J. Forest Res. 39 (1), 143–153.
Ukrainetz, N.K., O’Neill, G.A., Jaquish, B., 2011. Comparison of fixed and focal pointseed transfer systems for reforestation and assisted migration: a case study forinterior spruce in British Columbia. Can. J. Forest Res. 41 (7), 1452–1464.
Van Vuuren, D.P., Edmonds, J., Kainuma, M., Riahi, K., Thomson, A., Hibbard, K.,Hurtt, G.C., Kram, T., Krey, V., Lamarque, J., Masui, T., Meinshausen, M.,Nakicenovic, N., Smith, S.J., Rose, S.K., 2011. The representative concentrationpathways: an overview. Climatic Change 109 (1–2), 5–31.
Wang, T., Hamann, A., Yanchuk, A., O’Neill, G.A., Aitken, S.N., 2006. Use of responsefunctions in selecting lodgepole pine populations for future climate. GlobalChange Biol. 12 (12), 2404–2416.
Wang, T., O’Neill, G.A., Aitken, S.N., 2010. Integrating environmental and geneticeffects to predict responses of tree populations to climate. Ecol. Appl. 20 (1),153–163.
Williams, M.I., Dumroese, R.K., 2013. Preparing for climate change: forestry andassisted migration. J. Forest. 111 (4), 287–297.
Wright, J.W., Amiel, R.J., Cech, F.C., Kriebel, H.B., Jokela, J.J., Lemmien, W.A.,Matheson, A.C., Meritt, C., Read, R.A., Roth, P., Thor, E., Thulin, I.J., 1979.Performance of eastern white pine from the southern Appalachians in easternUnited States, New Zealand and Australia. In: Proceedings of the 26thNortheastern Forest Tree Improvement Conference, July 25–26, 1978.Contributed papers, Northeastern Forest Experimental Station, pp. 203–217.
Top Related
Copyright © 2022 FDOKUMEN

