







Bahasa
Halaman
Hukum


The combined effect of photoperiod and temperature onegg dormancy in an island and a mainland populationof the hemlock looper, Lambdina fiscellariaJohanne Delisle1*, Lucie Royer2, Michèle Bernier-Cardou1, Eric Bauce3 & AlainLabrecque1
1Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, 1055 du P.E.P.S., PO Box 10380, Stn. Sa-
inte-Foy, Quebec, QC G1V 4C7, Canada, 2Natural Resources Canada, Canadian Forest Service, Atlantic Forestry Centre,
University Drive, PO Box 960, Corner Brook, NL A2H 6J3, Canada, and 3Departement des sciences du bois et de la foret,
Universite Laval, Pavillon Abitibi-Price, 2405 de la Terrasse, Quebec, QC G1V 0A6, Canada
Accepted: 1 September 2009
Key words: diapause duration, ecotypes, egg mass, embryonic pre-diapause, diapause, post-dia-
pause, Geometridae, Lepidoptera, time to hatch
Abstract Embryonic dormancy characteristics of the hemlock looper (HL), Lambdina fiscellaria (Guenee)
(Lepidoptera: Geometridae), an important coniferous defoliator, were investigated using eggs from
an island (Newfoundland) and a mainland (Quebec) population in eastern Canada. We determined
(1) the pre-diapause duration or time required for eggs to change color at 15 �C, under L16:D8 and
L12:D12, (2) the percent hatch and time to hatch of early-diapausing eggs that were, as of 1 Septem-
ber, either (a) incubated under each combination of three photoperiods (L16:D8, L12:D12, or
L8:D16) and three temperatures (15, 20, or 25 �C) or (b) stored outdoors prior to their monthly
incubation (October–May) under the same treatments, and (3) the diapause duration at 15 �C. The
two HL ecotypes completed their pre-diapause phase in ca. 15 days under long-day photoperiod. In
the mainland ecotype, photoperiod did not influence the pre-diapause duration. Regardless of
photoperiod and ecotype, only eggs incubated continuously at 15 �C hatched successfully after ca.
120 days. Temperature was the most important factor modulating the dormancy of eggs acclimated
outdoors, in both ecotypes. From October to December (diapause phase), percent egg hatch at 20
and 25 �C increased from low (20%) to levels similar to those obtained at 15 �C (70%). These per-
centages remained stable throughout the post-diapause phase (January–May). Time to hatch, which
was shorter at warmer temperatures, decreased from October to December at all temperatures. It
remained stable from January to March (quiescence), however, and declined thereafter. Eggs from
the island were heavier than those from the mainland and their odds of hatching were 2.3 times
higher. At 15 �C, diapause lasted ca. 90 days. Our findings indicate that eggs from the two HL eco-
types (1) undergo an obligatory diapause, (2) complete diapause without pre-exposure to cold, (3)
respond similarly to photoperiod and temperature during diapause and post-diapause, and (4) do
not hatch successfully after prolonged exposure to high temperatures.
Introduction
In response to harsh environmental conditions, insects
have developed adaptations that enable them to escape
either spatially through dispersal ⁄ migration or temporally
through dormancy (Solbreck, 1978; Tauber et al., 1986).
Diapause is an endogenously and hormonally controlled
form of dormancy characterized by an arrest of morpho-
logical development combined with a slowing of the
insect’s basal metabolism (Danks, 1987). Two types of dia-
pause have been recognized: facultative and obligatory.
The facultative diapause is induced by external cues,
mainly photoperiod and ⁄ or temperature, whereas the
obligatory diapause is not environmentally regulated but
rather genetically predetermined (Danks, 1987; Kostal,
*Correspondence: Johanne Delisle, Natural Resources Canada,
Canadian Forest Service, Laurentian Forestry Centre, 1055 du
P.E.P.S., PO Box 10380, Stn. Sainte-Foy, Quebec, QC G1V 4C7,
Canada. E-mail: [email protected]
� Her Majesty the Queen in Right of Canada, as represented by
the Minister of Natural Resources Canada Entomologia Experimentalis et Applicata 133: 232–243, 2009
232 Journal compilation � 2009 The Netherlands Entomological Society
DOI: 10.1111/j.1570-7458.2009.00931.x

2006). However, after the induction or initiation phase
(pre-diapause), the possibility still exists that the intensity
or duration of diapause of either type can be affected by
external cues (Tauber et al., 1986; Danks, 1987).
Once diapause is completed, resumption of post-
diapause development depends mainly on temperature
conditions: cold keeps the insects in a quiescent state until
spring arrives. This allows species to synchronize their life
cycle with the season (Nilssen & Tenow, 1990; Keena, 1996;
Broufas et al., 2006). The possibility, however, that photo-
period might influence the post-diapause phase is not
totally excluded, as demonstrated in a certain number of
univoltine species (Sawchyn & Church, 1973; McNeil &
Fields, 1985; Polavarapu & Seabrook, 1996). This empha-
sizes the importance of assessing the simultaneous effect of
photoperiod and temperature over the whole physiological
process, including both diapause and post-diapause phases.
Dormancy characteristics (photoperiodic response, dia-
pause intensity, and temperature requirements) can vary
greatly among individuals within a given population, and
the heritability of these traits has been demonstrated in
certain species (Du Merle, 1999; van Asch et al., 2007;
Demont & Blanckenhorn, 2008). Similarly, differences in
dormancy characteristics have also been observed between
populations of a single species located at different latitudes
(Mousseau & Roff, 1989; Du Merle, 1999; Hunter &
McNeil, 2000; Demont & Blanckenhorn, 2008). This intra-
and inter-population ability to adapt to local environmen-
tal conditions suggests that dormancy characteristics
might also differ between island and mainland popu-
lations, as has been observed for other ecological or biolog-
ical traits such as biodiversity or morphology (Gillespie &
Roderick, 2002; Joy et al., 2007).
The hemlock looper (HL), Lambdina fiscellaria
(Guenee) (Lepidoptera: Geometridae), is a univoltine
species in eastern Canada and an outbreaking pest of
coniferous forests (Rose & Lindquist, 1977), preferentially
attacking balsam fir, Abies balsamea (L.) Miller (Pinaceae)
(Carroll, 1952). Adults are active from mid-August to
October (Delisle et al., 1998) and eggs, which are laid indi-
vidually on tree trunks or branches, are exposed to harsh
weather during the winter and do not hatch until the fol-
lowing spring (Carroll, 1952). Although winter survival
represents an important component of insect population
dynamics, to our knowledge, the embryonic dormancy of
the HL has never been investigated.
We therefore undertook a series of experiments to char-
acterize HL dormancy by (1) determining the duration of
the pre-diapause phase under different photoperiods, (2)
assessing the combined effect of photoperiod and temper-
ature on percent egg hatch and time to egg hatch during
the diapause and post-diapause phases, and (3) estimating
the duration of diapause under these various experimental
conditions using eggs originating from an island (New-
foundland) and a mainland (Quebec) population (eco-
types) in northeastern Canada.
In this study, we hypothesized that if photoperiod regu-
lates the embryonic dormancy of the HL in the two eco-
types, it would similarly affect their pre-diapause,
diapause, and post-diapause characteristics, as both eco-
types come from the same northern latitude. In contrast,
temperature requirements for completing the diapause
process and resuming post-diapause development may
differ between the two ecotypes, as winters are generally
milder on the island of Newfoundland than on the Cana-
dian mainland (Environment Canada, 2008). Throughout
these experiments, we have intentionally selected relatively
warm incubation temperatures to assess the potential
impact of fall conditions expected under global warming
scenarios on HL egg dormancy.
Materials and methods
Ecotype origin
The HL island ecotype came from Port Saunders, New-
foundland (50�38¢N, 57�16¢W; elevation: 90.8 m), and the
mainland ecotype came from Baie-Trinite, Quebec
(49�25¢N, 67�18¢W; elevation: 55.0 m; Figure 1A). These
two regions were selected because their similar latitudinal
position makes it possible to avoid a bias in the dormancy
study. Port Saunders is about 95 km away from the nearest
mainland shore and it is more than 400 km away from
Baie-Trinite; therefore, the two ecotypes can be considered
geographically separate from each other. Although no
information is currently available on the flight capacity of
the HL, males and females have been seen taking off and fly-
ing up about 1–3 m above the canopy, presumably to dis-
perse locally from one stand to another, as reported by
Raske & Farrell (1986). The two regions selected also have
different climates. Based on the mean monthly temperature
recorded historically (1971–2000), winters are milder on
the island, whereas spring and summer are characterized by
cooler temperatures than on the mainland; however, fall
temperatures are similar for the two regions (Figure 1B;
Environment Canada, 2008). Therefore, to better identify
potential dormancy differences between the two ecotypes,
we conducted our experiments in Quebec City (46�46¢N,
71�16¢W; elevation: 74.4 m) as it is located farther south
than Baie-Trinite and has warmer fall and spring tempera-
tures, whereas the winters are just as harsh (Figure 1A,B).
Insect rearing
Eggs obtained from HL adults collected in the fall of 1999
from the two sites were used to establish colonies that were
Egg dormancy in hemlock looper 233

subsequently propagated under semi-natural conditions
for six consecutive generations before being used in the
experiments described next. Each year, from 2000 to 2004,
eggs were placed in glass Mason jars covered with nylon
and stored from late summer (September) to the following
spring (May) in an outdoor insectary located in Quebec
City. In spring, the eggs were transferred to a controlled
growth chamber kept under standard conditions of
L16:D8, 15 ± 1 �C, and 80 ± 5% relative humidity (r.h.).
Neonate larvae were then reared on shoots of balsam fir, A.
balsamea, which were renewed every 3–4 days.
Eggs produced in the fall of 2005 were used for our
experiments. For each ecotype, eggs from at least 120
mated females were thoroughly mixed and then redistrib-
uted in batches of 25 or 50 eggs into 1-dram glass vials for
experiments under laboratory or a mix of semi-natural
and laboratory conditions, respectively. All glass vials were
covered with a foam plug to permit air circulation. During
the storage and incubation periods, variations in tempera-
ture, day length (based on the time that light intensity
exceeded 0.3 lux), and relative humidity were recorded at
15-min intervals with miniaturized data loggers (HOBO
H8 and PRO; Hoskin Scientifique, Montreal, QC,
Canada). Figure 2 shows the variation in day length and
minimum and maximum temperatures recorded daily in
both the outdoor insectary and the storage room (see next)
throughout the experimental period, from the beginning
of September to the end of May.
Mass of eggs
Prior to experiments, the mean mass of eggs from each
ecotype was estimated by weighing 20 batches of 25 freshly
laid eggs on a metric balance accurate to 0.005 mg
(Sartorius Mechatronics, R200D, Gottingen, Germany).
B
A
Figure 1 (A) Place of origin of the island and mainland ecotypes (Port Saunders and Baie-Trinite, respectively) and location of the study
site (Quebec City) in eastern Canada, and (B) average daily temperatures recorded historically (1971–2000) throughout the year. Vertical
lines span the corresponding 95% confidence intervals.
234 Delisle et al.

Experiments
Experiment 1: Pre-diapause duration. The phase that pre-
cedes the onset of diapause, called pre-diapause, is often
characterized by a change in either the color or form of the
dormant stage (Danks, 1987). In the HL, the mean dura-
tion of pre-diapause was estimated from the number of
days it took for eggs to change color after their oviposition
at 15 �C under L16:D8. A single observer used various
paint color cards (Glidden’s Coordinated Collections;
http://www.glidden.com/colors/selector) to monitor, for
each individual egg, the daily color change from green
(card #BL43) to coppery brown (card #NC11), until the
color remained stable for four consecutive days. The date
on which the final color was first observed was considered
the end of pre-diapause. Seventeen freshly laid eggs from
both the island ecotype and the mainland ecotype were
observed. An additional group of 16 eggs from the main-
land ecotype, laid and maintained at 15 �C under
L12:D12, was used to investigate the effect of a short day-
length on pre-diapause duration.
Experiment 2: Impact of photoperiod and temperature on egg
dormancy. This group of experiments was conducted
with eggs laid at 15 �C and under L16:D8 during the
2nd week of August and kept under these standard
conditions until the beginning of the experiment on 1
September.
Experiment 2a: Eggs kept continuously under laboratory con-
ditions. The combined effects of photoperiod and tem-
perature conditions during egg diapause were first
assessed in the laboratory. Two vials of 25 eggs from
each ecotype were incubated under each combination of
three photoperiods (L16:D8, L12:D12, or L8:D16) and
three temperatures (15, 20, or 25 �C). To determine
whether seasonal variations in natural day length (Fig-
ure 2A) influence the progression of diapause (Tauber &
Tauber, 1976), two additional vials of 25 eggs from each
ecotype were placed near the window of a storage room
with no other light sources, where mean monthly tem-
peratures varied between 18.6 and 21.7 �C (Figure 2B).
Thus, in total, there were 10 laboratory photoperiod-
temperature treatments · two ecotypes · two replicates
(40 vials).
Experiment 2b: Eggs acclimated to outdoor conditions for
1–8 months. The course of diapause and post-diapause
was also studied when eggs were stored under semi-natural
conditions for different periods of time prior to their incu-
bation under laboratory conditions. At the start of each
month, from October to May, two vials of 50 eggs, accli-
mated and kept in the outdoor insectary since 1 Septem-
ber, were transferred to each of the 10 incubation
treatments described for experiment 2a · two ecotypes ·two replicates · 8 months (=320 vials).
A
B
C
Figure 2 Day length (A) and minimum and maximum temperatures recorded daily from September 2005 to May 2006 in the storage room
(B) or the insectary (C). Monthly mean day lengths and temperatures are also indicated at the top of each graph.
Egg dormancy in hemlock looper 235

In experiments 2a and 2b, eggs that hatched in each vial
were counted daily throughout the incubation period to
determine (1) the total number, r, of hatched eggs per vial,
out of n, over the whole experiment period, and (2) the
time to hatch for each egg, in days, defined as the time
elapsed between incubation date and the day the egg
hatched. All vials stored in the outdoor insectary were also
examined once a week from 1 September to the end of
November, and larvae that hatched before incubation were
counted.
Statistical analyses
The mean mass of eggs from the two ecotypes was com-
pared by Student’s t-test. In experiment 1, the mean dura-
tion of pre-diapause of eggs from the two ecotypes under
standard conditions (15 �C and L16:D8) and that of eggs
from the mainland ecotype under the two day lengths
(L16:D8 and L12:D12) at 15 �C were compared through
an analysis of variance and two specific, a priori contrasts.
In experiment 2b, the number (r) of eggs that hatched
out of n, in each vial, was assumed to follow a binomial
distribution with probability p of hatching and a logit link
function between p and a linear combination of the effects
of ecotype, treatment, incubation date, and their interac-
tions [Agresti, 2002; GENMOD procedure of SAS (SAS
Institute 1999)]. The proportion of eggs that hatched
within any ecotype, treatment, or incubation date was
assessed in terms of odds, p=ð1� pÞ; and any two such
odds were compared through their ratio, p1=ð1� p1Þ½ �=p2=ð1� p2Þ½ �; where indices 1 and 2 refer to the two arms
of the comparison, respectively (e.g., 1 = island ecotype
and 2 = mainland ecotype; see Agresti, 2002). Log odds
ratios and their 95% confidence limits were computed on
the logit scale and back-transformed to odds ratios by
exponentiation.
The natural logarithm of time to hatch was averaged
within each vial, each log-scale mean was weighted by
the number of hatched eggs in the vial, and the variance
of these means was analyzed with a model that depended
on ecotype, treatment, incubation date, and their inter-
actions. Inspection of the residuals revealed that the
variance component for vials should be allowed to vary
with incubation date from October to February and con-
strained to remain constant from February to May (Proc
MIXED of SAS).
Analyses of percent egg hatch and time to hatch from
experiment 2a were based on models similar to those used
for the corresponding variables in experiment 2b, except
that the incubation date factor and its interactions with
ecotype or treatment were dropped from the initial
models. The variance between vials was thus assumed
constant.
In experiment 2b, total time to hatch was obtained as
the sum of time to hatch of each egg and the number of
days between 1 September and its incubation date. A
model similar to that for time to hatch was fit to the
logarithm of total time to hatch, but the variance com-
ponent for vials was allowed to vary among all incuba-
tion dates.
The duration of diapause (x0) under a constant incuba-
tion temperature of 15 �C was estimated as a function of
selected means estimated from results of experiments 1
(duration of pre-diapause) and 2b (time to hatch and total
time to hatch). Details will be given in the ‘Results’ section.
The standard error of the estimate [SE(x0)] was approxi-
mated by the delta method taking into account the
variability of both time to hatch and duration of post-
diapause (Wolter, 1985).
All tests were conducted at the a = 0.05 level. Through-
out, back-transformed means, percentages, odds, and odds
ratios reported in the text are followed by their lower and
upper 95% confidence limits in parentheses.
Results
Mass of eggs
Eggs from the island ecotype weighed on average 5.43 mg
per group of 25 eggs (5.32–5.53), slightly more than those
of the mainland ecotype, which weighed 5.03 mg
per group of 25 eggs (4.92–5.13; t = 5.58, d.f. = 38,
P<0.001).
Experiment 1: Pre-diapause duration
Under standard conditions (L16:D8 and 15 �C), eggs from
the island ecotype completed their pre-diapause in
15.1 days (14.4–15.7) and those from the mainland eco-
type did so in 13.9 days (13.2–14.5), a significant differ-
ence of 1.2 days (0.23–2.12; t = 2.51, d.f. = 47, P = 0.02).
The duration of pre-diapause of eggs from the mainland
ecotype obtained under L12:D12 was estimated to be
14.3 days (13.6–14.9), a value that was not statistically dif-
ferent from that obtained under L16:D8 (t = 0.77,
d.f. = 47, P = 0.44).
Experiment 2: Impact of photoperiod and temperature on egg dor-mancy
Because the eggs used in experiment 2 were laid under
standard conditions (L16:D8 and 15 �C) during the 2nd
week of August, it was assumed that the majority began
their diapause at the time they were either incubated under
constant or fluctuating laboratory conditions (experiment
2a) or transferred to the insectary prior to incubation
under the same laboratory conditions at different dates
(experiment 2b).
236 Delisle et al.

Experiment 2a: Eggs kept continuously under laboratory conditions
After incubation on 1 September, observed numbers of
hatched eggs were mostly zero or nearly so at incubation
temperatures of 20 or 25 �C (Table 1). The analysis was
thus restricted to the 15 �C temperature and the model
was modified accordingly. At 15 �C, photoperiod had no
detectable effect on percent egg hatch (v2 = 5.21, d.f. = 2,
P = 0.07), nor was there any indication that the effect of
photoperiod differed among the two ecotypes (eco-
type*photoperiod: v2 = 0.50, d.f. = 2, P = 0.78). The
odds of hatching were 2.4 times (1.4–4.0) higher among
eggs from the island ecotype than among those from the
mainland ecotype (v2 = 11.71, d.f. = 1, P<0.001). Overall,
percent hatch reached 70.6% (62.0–77.9) among eggs from
the island ecotype compared with 49.8% (41.4–58.3)
among those from the mainland ecotype (Table 1).
After incubation at 15 �C on 1 September, neither pho-
toperiod nor ecotype seemed to affect time to hatch
(P>0.18 for the effects of photoperiod, ecotype, or their
interaction). For all photoperiods and ecotypes combined,
eggs hatched after a period of 120 days (115–125), from 1
September to the end of December, with a range extending
from 70 to 173 days. Given that time to hatch includes the
duration of diapause and post-diapause in this experi-
ment, diapause must last less than 120 days and, therefore,
it must have been completed before the end of December
in both HL populations.
Experiment 2b: Eggs acclimated to outdoor conditions for1–8 months
In the insectary, only six eggs from the island ecotype and
two from the mainland ecotype (out of a total of 16 000)
hatched before they were transferred to the various incuba-
tion treatments. This represents a total loss of 0.05% dis-
tributed over a number of treatments and months. These
eggs were excluded from the analyses.
Temperature significantly influenced percent egg hatch
following incubation at the beginning of each month from
October to May, but photoperiod did not, nor did the
interaction between temperature and photoperiod (Tr, T,
P, and P*T; Table 2). Before the winter solstice, eggs
hatched in larger proportions at 15 �C than at warmer
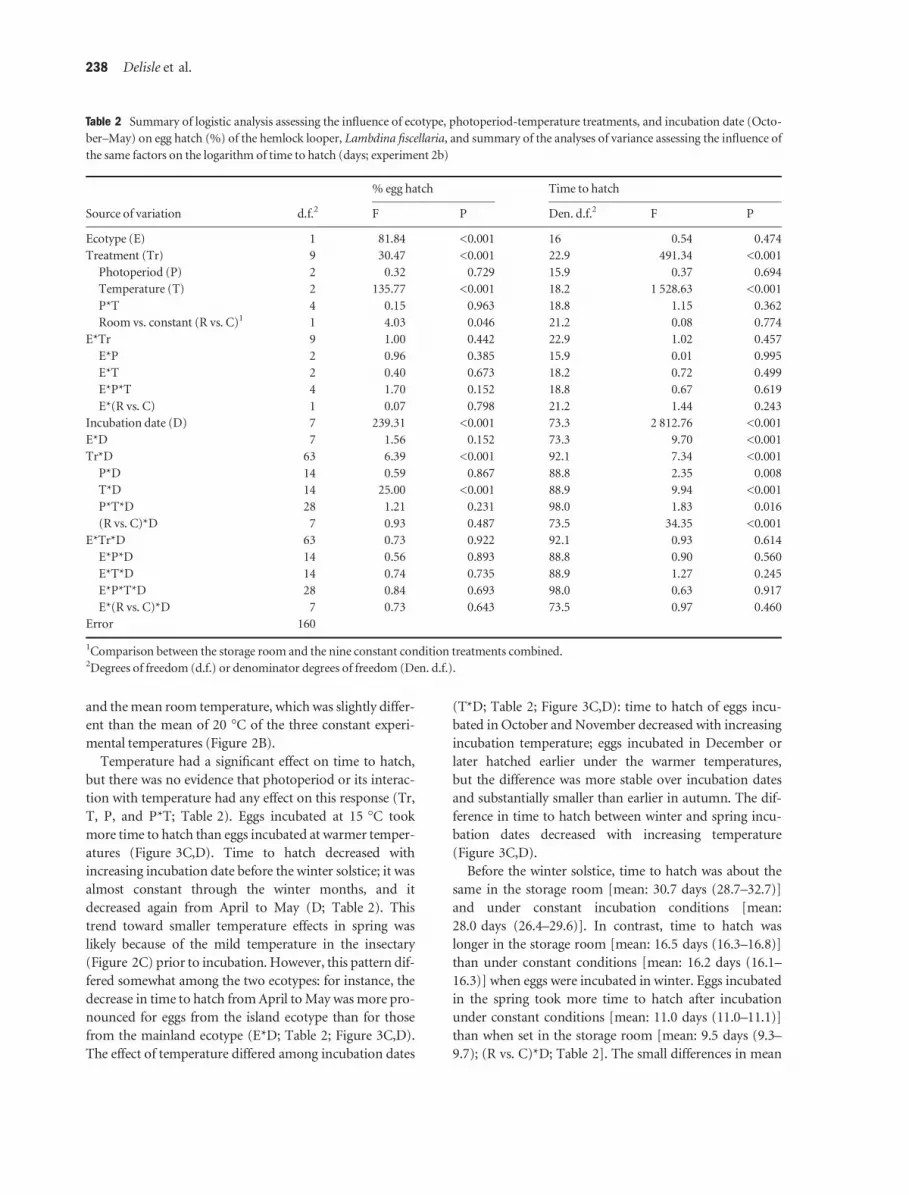
temperatures (Figure 3A,B). Furthermore, percent hatch
generally increased with incubation date, but the increase
was more pronounced under incubation temperatures of
20 or 25 �C than at 15 �C (D and T*D; Table 2). At later
incubation dates, percent hatch remained stable and high
(Figure 3A,B). The odds of hatching of eggs from the
island ecotype were 2.3 times (2.0–2.5) greater than the
corresponding odds for eggs from the mainland ecotype:
72% (71–74) of eggs from the island ecotype hatched,
whereas only 53% (51–55) from the mainland ecotype did
so (E; Table 2).
On average over incubation dates and the two ecotypes,
the odds of hatching were 1.2 times (1.0–1.4) higher under
fluctuating room conditions than under all constant con-
ditions combined (P<0.05 for R vs. C; Table 2): percent
hatch averaged 66% (63–70) in the storage room and 63%
(62–64) under constant temperature and photoperiod
conditions. There was no indication that this effect differed
among ecotypes or incubation dates [E*(R vs. C), (R vs.
C)*D, E*(R vs. C)*D; Table 2]. These small differences
were somewhat expected, given the apparently negligible
effect of photoperiod on percent hatch (Tables 1 and 2)
Table 1 Back-transformed egg hatch (%) for the island and mainland ecotypes of the hemlock looper, Lambdina fiscellaria, following incu-
bation on 1 September 2005, under constant and fluctuating (room) conditions (experiment 2a)
Photoperiod (L:D) Temperature (�C)
Island Mainland
n % egg hatch1 n % egg hatch
16:8 15 44 68.2 (52.6–80.6) 49 51.0 (36.8–65.1)
12:12 15 50 80.0 (66.1–89.2) 50 56.0 (41.6–69.5)
8:16 15 47 61.7 (46.6–74.8) 47 42.6 (28.9–57.5)
16:8 20 49 16.32 49 2.0
12:12 20 50 4.0 51 7.8
8:16 20 50 8.0 50 0.0
16:8 25 50 2.0 50 0.0
12:12 25 49 0.0 50 0.0
8:16 25 50 0.0 51 0.0
Room Room 50 12.0 50 2.0
1Lower and upper 95% confidence limits in parentheses.2Lower and upper 95% confidence limits are unavailable ; the data obtained at 20 �C, 25 �C, or room temperature were not statistically
analysed.
Egg dormancy in hemlock looper 237
and the mean room temperature, which was slightly differ-
ent than the mean of 20 �C of the three constant experi-
mental temperatures (Figure 2B).
Temperature had a significant effect on time to hatch,
but there was no evidence that photoperiod or its interac-
tion with temperature had any effect on this response (Tr,
T, P, and P*T; Table 2). Eggs incubated at 15 �C took
more time to hatch than eggs incubated at warmer temper-
atures (Figure 3C,D). Time to hatch decreased with
increasing incubation date before the winter solstice; it was
almost constant through the winter months, and it
decreased again from April to May (D; Table 2). This
trend toward smaller temperature effects in spring was
likely because of the mild temperature in the insectary
(Figure 2C) prior to incubation. However, this pattern dif-
fered somewhat among the two ecotypes: for instance, the
decrease in time to hatch from April to May was more pro-
nounced for eggs from the island ecotype than for those
from the mainland ecotype (E*D; Table 2; Figure 3C,D).
The effect of temperature differed among incubation dates
(T*D; Table 2; Figure 3C,D): time to hatch of eggs incu-
bated in October and November decreased with increasing
incubation temperature; eggs incubated in December or
later hatched earlier under the warmer temperatures,
but the difference was more stable over incubation dates
and substantially smaller than earlier in autumn. The dif-
ference in time to hatch between winter and spring incu-
bation dates decreased with increasing temperature
(Figure 3C,D).
Before the winter solstice, time to hatch was about the
same in the storage room [mean: 30.7 days (28.7–32.7)]
and under constant incubation conditions [mean:
28.0 days (26.4–29.6)]. In contrast, time to hatch was
longer in the storage room [mean: 16.5 days (16.3–16.8)]
than under constant conditions [mean: 16.2 days (16.1–
16.3)] when eggs were incubated in winter. Eggs incubated
in the spring took more time to hatch after incubation
under constant conditions [mean: 11.0 days (11.0–11.1)]
than when set in the storage room [mean: 9.5 days (9.3–
9.7); (R vs. C)*D; Table 2]. The small differences in mean
Table 2 Summary of logistic analysis assessing the influence of ecotype, photoperiod-temperature treatments, and incubation date (Octo-
ber–May) on egg hatch (%) of the hemlock looper, Lambdina fiscellaria, and summary of the analyses of variance assessing the influence of
the same factors on the logarithm of time to hatch (days; experiment 2b)
Source of variation d.f.2
% egg hatch Time to hatch
F P Den. d.f.2 F P
Ecotype (E) 1 81.84 <0.001 16 0.54 0.474
Treatment (Tr) 9 30.47 <0.001 22.9 491.34 <0.001
Photoperiod (P) 2 0.32 0.729 15.9 0.37 0.694
Temperature (T) 2 135.77 <0.001 18.2 1 528.63 <0.001
P*T 4 0.15 0.963 18.8 1.15 0.362
Room vs. constant (R vs. C)1 1 4.03 0.046 21.2 0.08 0.774
E*Tr 9 1.00 0.442 22.9 1.02 0.457
E*P 2 0.96 0.385 15.9 0.01 0.995
E*T 2 0.40 0.673 18.2 0.72 0.499
E*P*T 4 1.70 0.152 18.8 0.67 0.619
E*(R vs. C) 1 0.07 0.798 21.2 1.44 0.243
Incubation date (D) 7 239.31 <0.001 73.3 2 812.76 <0.001
E*D 7 1.56 0.152 73.3 9.70 <0.001
Tr*D 63 6.39 <0.001 92.1 7.34 <0.001
P*D 14 0.59 0.867 88.8 2.35 0.008
T*D 14 25.00 <0.001 88.9 9.94 <0.001
P*T*D 28 1.21 0.231 98.0 1.83 0.016
(R vs. C)*D 7 0.93 0.487 73.5 34.35 <0.001
E*Tr*D 63 0.73 0.922 92.1 0.93 0.614
E*P*D 14 0.56 0.893 88.8 0.90 0.560
E*T*D 14 0.74 0.735 88.9 1.27 0.245
E*P*T*D 28 0.84 0.693 98.0 0.63 0.917
E*(R vs. C)*D 7 0.73 0.643 73.5 0.97 0.460
Error 160
1Comparison between the storage room and the nine constant condition treatments combined.2Degrees of freedom (d.f.) or denominator degrees of freedom (Den. d.f.).
238 Delisle et al.
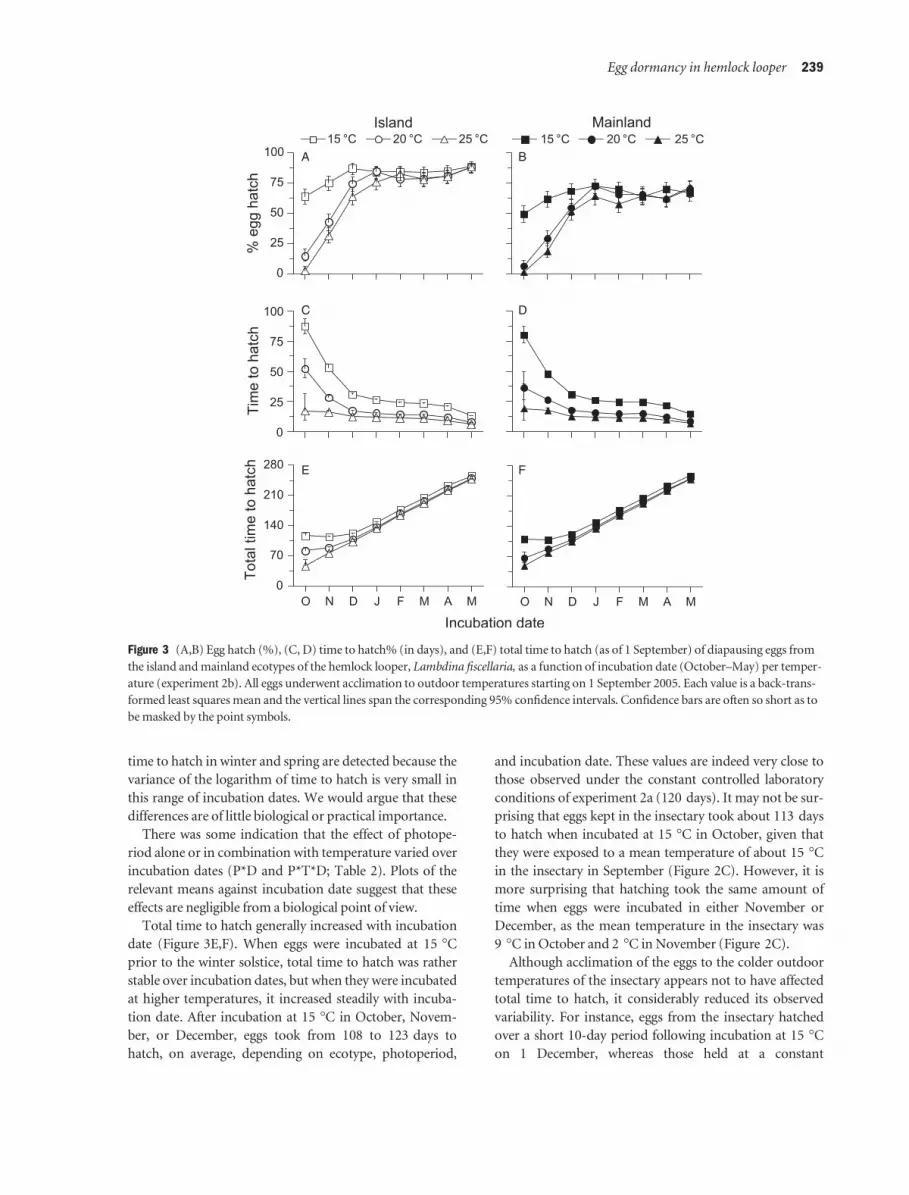
time to hatch in winter and spring are detected because the
variance of the logarithm of time to hatch is very small in
this range of incubation dates. We would argue that these
differences are of little biological or practical importance.
There was some indication that the effect of photope-
riod alone or in combination with temperature varied over
incubation dates (P*D and P*T*D; Table 2). Plots of the
relevant means against incubation date suggest that these
effects are negligible from a biological point of view.
Total time to hatch generally increased with incubation
date (Figure 3E,F). When eggs were incubated at 15 �C
prior to the winter solstice, total time to hatch was rather
stable over incubation dates, but when they were incubated
at higher temperatures, it increased steadily with incuba-
tion date. After incubation at 15 �C in October, Novem-
ber, or December, eggs took from 108 to 123 days to
hatch, on average, depending on ecotype, photoperiod,
and incubation date. These values are indeed very close to
those observed under the constant controlled laboratory
conditions of experiment 2a (120 days). It may not be sur-
prising that eggs kept in the insectary took about 113 days
to hatch when incubated at 15 �C in October, given that
they were exposed to a mean temperature of about 15 �C
in the insectary in September (Figure 2C). However, it is
more surprising that hatching took the same amount of
time when eggs were incubated in either November or
December, as the mean temperature in the insectary was
9 �C in October and 2 �C in November (Figure 2C).
Although acclimation of the eggs to the colder outdoor
temperatures of the insectary appears not to have affected
total time to hatch, it considerably reduced its observed
variability. For instance, eggs from the insectary hatched
over a short 10-day period following incubation at 15 �C
on 1 December, whereas those held at a constant
A B
C D
E F
Figure 3 (A,B) Egg hatch (%), (C, D) time to hatch% (in days), and (E,F) total time to hatch (as of 1 September) of diapausing eggs from
the island and mainland ecotypes of the hemlock looper, Lambdina fiscellaria, as a function of incubation date (October–May) per temper-
ature (experiment 2b). All eggs underwent acclimation to outdoor temperatures starting on 1 September 2005. Each value is a back-trans-
formed least squares mean and the vertical lines span the corresponding 95% confidence intervals. Confidence bars are often so short as to
be masked by the point symbols.
Egg dormancy in hemlock looper 239

incubation temperature of 15 �C since 1 September
hatched over a period of 49 days for eggs from the island
ecotype and 31 days for those from the mainland ecotype.
Estimation of diapause duration at 15 �C
Results from experiments 1, 2a, and 2b suggest that eggs
laid under standard conditions (L16:D8 and 15 �C) go
through three developmental phases: a pre-diapause,
which lasts about 2 weeks, diapause itself, and post-dia-
pause and ⁄ or quiescence. The stability of total time to
hatch (ca. 112 days) and hatching success (>70 or 50%
depending on ecotype) after incubation at 15 �C in
October, November, and December in experiment 2b
(Figure 3A,B,E,F) and their consistency with the corre-
sponding responses of eggs incubated at the same tempera-
ture on 1 September in experiment 2a (120 days) suggest
that diapause of HL eggs is of fixed duration under these
conditions. Moreover, the decrease in mean time to hatch
with increasing incubation date before the winter solstice
followed by its levelling off to a practically constant value
after incubation in January, February, and March indicates
that diapause was completed by the end of December, and
eggs had entered a quiescent state, ready to hatch when
environmental conditions became favourable. Time to
hatch at 15 �C was thus used to estimate diapause dura-
tion as follows.
First, it was assumed that 1 September marked the
beginning of diapause for eggs from both ecotypes. Sec-
ond, the duration of post-diapause was estimated from
mean time to hatch after incubation at 15 �C in January,
February, and March as 24.5 days (24.2–24.8) for the
island ecotype and 25.0 days (24.7–25.4) for the mainland
ecotype. Finally, for each ecotype, the latter value was sub-
tracted from mean total time to hatch after incubation in
October, November, and December at 15 �C to yield a dia-
pause duration estimate of 94 days (93–96) for the island
ecotype and 90 days (88–92) for the mainland ecotype.
Under the conditions of the insectary, which approximate
natural conditions, diapause of eggs from both ecotypes
must have been completed between 30 November and 4
December.
Discussion
In both HL ecotypes, the change observed in egg color,
which marks the onset of diapause, was induced even
though the embryos and their mothers (during both their
immature and adult stages) were maintained under a
L16:D8 photoperiod. Furthermore, eggs of the mainland
ecotype took the same number of days to enter diapause
under either short- or long-day photoperiods. Together,
these results suggest that the embryonic diapause of the
HL is not photoperiodically regulated but rather geneti-
cally pre-programmed, as expected for insects that
undergo an obligatory diapause. This type of diapause
may explain why HL populations are univoltine in eastern
Canada.
Differences observed in pre-diapause, diapause, and
post-diapause duration between eggs of the island and
mainland ecotypes were biologically negligible, as they
averaged 1–4 days. Nonetheless, percent egg hatch was
consistently higher in the island ecotype compared with
the mainland ecotype in all experiments. If egg size is a
good indicator of tolerance to environmental stress (Fox,
1997), the higher odds of hatching among eggs from the
island ecotype compared with those for the mainland eco-
type may be attributable to the greater egg mass of the for-
mer. Given the milder winters, the later onset of the
growing season and the cooler spring temperatures of the
island environment (Figure 1B), eggs of the island ecotype
must remain quiescent for a longer time period than eggs
of the mainland ecotype. This may increase the risk of pre-
mature death possibly associated with desiccation or
depletion of metabolic reserves (Sota & Mogi, 1992). The
greater egg mass may provide eggs from the island ecotype
with an adaptive advantage over those of the mainland
ecotype, much more so than the small differences observed
in the duration of the different phases of dormancy. If the
duration of post-diapause quiescence is a challenge for egg
survival, egg size should vary with the duration of win-
ter ⁄ spring conditions, and this should be reflected in an
increasing south-to-north trend in egg size (Armbruster
et al., 2001). Recent work by Berthiaume (2007) showing
an increase in the size of HL eggs with latitude in eastern
Canada supports this hypothesis. However, no doubt that
several HL populations, located at different latitudes,
should also be investigated to lend credence to the positive
relationship observed between percent egg hatch and egg
mass from the two ecotypes studied.
In HL eggs maintained under laboratory conditions, the
course of diapause was observed only at 15 �C, as evi-
denced by the higher percent egg hatch at this temperature
compared with percent hatch at 20 or 25 �C or at fluctuat-
ing temperatures in the storage room. As the percent egg
hatch obtained in an additional experiment conducted at
10 �C (data not shown) was comparable with that
observed at 15 �C, the latter appears to approximate the
upper temperature limit for the progression of diapause in
the HL, as demonstrated for the embryonic diapause of
other species (Andrewartha, 1952; Tauber et al., 1990).
Therefore, it seems that temperatures close to or below
0 �C, are not a prerequisite for diapause completion in a
number of species inhabiting temperate regions (reviewed
by Hodek & Hodkova, 1988).
240 Delisle et al.

The increasingly short time to hatch observed as the sea-
son advanced (Figure 3C,D) has often been linked to a
decrease in diapause intensity (Tauber et al., 1986; Danks,
1987). However, if we consider total time to hatch (dura-
tion of egg storage in the insectary plus time to hatch since
incubation), as recommended by Hodek & Hodkova
(1988), HL eggs took about 115 days to hatch at 15 �C.
Thus, diapause intensity remained stable over the pre-
solstice period in the HL, and the passage of time alone
leads to diapause completion after about 3 months, in
both ecotypes. This consistency in total time to hatch over
incubation dates prior to the winter solstice also shows
that egg acclimation to outdoor conditions did not affect
the course of diapause, as shown in diapausing larvae of
Busseola fusca (Fuller), a noctuid species with an obligatory
diapause (Kfir, 1991).
Total time to hatch of eggs kept continuously at 15 �C
or stored in the outdoor insectary prior to their incubation
in early fall exhibited considerable intra-population vari-
ability (range: 70–173 days for eggs from the island eco-
type and 90–164 days for those from the mainland
ecotype after incubation on 1 September). This variability
declined sharply with increasing incubation temperature
(e.g., after incubation at 25 �C in October, total time to
hatch ranged from 38 to 69 days for eggs from the main-
land ecotype and from 46 to 52 days for those from the
mainland). We also noted that mean total time to egg
hatch decreased with increasing incubation temperatures
after incubation in October and November (Figure 3E,F).
Rather than accelerating the diapause process, the shorter
total time to hatch observed at high temperatures in early
fall suggests that only the eggs that were the furthest along
in diapause or the most resistant eggs survived these sub-
optimal conditions.
In September, daily temperatures in the outdoor insec-
tary ranged from 5 to 29 �C (Figure 2C). Despite the large
temperature fluctuations, more than half the eggs hatched
after incubation at 15 �C the following month (Fig-
ure 3A,B), suggesting that HL eggs can withstand short
periods of suboptimal temperatures at the beginning of
diapause. Interestingly, Fantinou et al. (1998) showed that
mature diapausing larvae of Sesamia nonagrioides
(Lefebvre) could more easily tolerate high temperatures
when their exposure to these temperatures was of short
duration and intermittent rather than continuous.
In contrast, percent egg hatch following acclimation to
cold temperatures for 1–8 months and incubation at high
temperatures (‡20 �C) increased sharply from October to
January, and then remained approximately constant until
May. The increase in percent egg hatch up to the winter
solstice suggests that temperature requirements for dia-
pause completion may change over time (Braune, 1973;
Tauber et al., 1986) or, more likely, that eggs with a more
advanced diapause at incubation time are more tolerant to
suboptimal temperatures during incubation. The relatively
high percent egg hatch observed after January in both eco-
types suggests that acclimation of the eggs to the colder
outdoor temperatures probably prevented the rapid deple-
tion of their energy reserves. However, this is not the only
benefit of prolonged exposure to cold temperature: this
acclimation also temporarily suspends their post-diapause
development (quiescence), thus favoring synchronization
of hatching in spring (Morden & Waldbauer, 1980; Polav-
arapu & Seabrook, 1996).
Throughout the post-diapause phase, HL eggs did not
respond to photoperiod as is the rule for several insect spe-
cies (Tauber et al., 1986, 1990; Danks, 1987). However, in
the European skipper, Thymelicus lineola (Ochs) (McNeil
& Fields, 1985), the blueberry leaftier, Croecia curvalana
(Kearfott) (Polavarapu & Seabrook, 1996), and three spe-
cies of Odonata (Lestes disjunctus Selys, Lestes unguiculatus
Hagen, and Lestes dryas Kirby; Sawchyn & Church, 1973),
percent hatch of post-diapausing eggs decreased with
decreasing photoperiod and this effect was more pro-
nounced at cool than at high temperatures. According to
McNeil & Fields (1985), this photoperiodic response may
represent a safeguard mechanism against temperature that
reduces death associated with precocious hatchings either
in the fall (Lestes) or spring (T. lineola and C. curvalana).
Because all of these species overwinter as either fully devel-
oped larvae within the chorion (T. lineola and C. curval-
ana) or as partially developed embryos with visible
eyespots (the three Lestes species), this sensitivity to photo-
period appears to depend on the presence of a well-
developed nervous system. In contrast, L. congener, the
only species among Lestes that overwinters as an undiffer-
entiated embryo, does not respond to photoperiod during
the post-diapause phase, as also demonstrated for the HL.
Therefore, it would not be unrealistic to speculate that HL
eggs enter diapause at an early stage of embryogenesis, such
as the germ band formation that precedes blastokinesis.
As resumption of embryogenesis in the HL is not photo-
periodically regulated, the only information needed to
develop a predictive model of the hatching of young larvae
in spring is the accumulated degree days above a certain
temperature threshold that remains to be determined for
this species. Given that diapause is completed well before
the winter solstice, it would be fairly easy, once the temper-
ature threshold is known, to determine the date on which
degree-day accumulation should begin, a step that would
increase the accuracy and reliability of the predictive
model (Tauber & Tauber, 1976; Levine, 1986).
It is clear that cool temperatures (£15 �C) were required
for the progression of diapause in the HL. Our study has
Egg dormancy in hemlock looper 241

identified two periods during which HL eggs would be espe-
cially vulnerable to the climatic variations associated with
global warming: the beginning of diapause and post-
diapause quiescence. The longer growing season predicted
by models of global warming (Easterling et al., 2000) could
increasetheriskofmortalityamongeggsthatwillbeexposed
to high temperatures over a longer period, as shown in the
spruce budworm, Choristoneura fumiferana Clemens (Han
& Bauce,1997). However, evenifembryo mortalitydoes not
occur, exposure of the eggs to warm fall temperatures could
have long-term repercussions on cold resistance (Han &
Bauce, 1998) or other developmental phases (Denlinger,
2002). Furthermore, as HL embryos are ready to resume
their morphogenetic development in late fall, winter thaws,
which may become more frequent with global warming
(Easterling et al., 2000), could lead to the anticipatory
resumption of embryogenesis in the HL. Should this phe-
nomenon be accompanied by an increase in the supercool-
ing point (Sullivan, 1965; Hanec, 1966; Nilssen & Tenow,
1990), the survival of HL embryos would depend on their
ability totoleraterepeatedfreeze-thawcycles.
Given these considerations, an intensification of the glo-
bal warming trend could lead to a northward shift of the
HL geographic range, a phenomenon already observed in
other species (Parmesan & Yohe, 2003). It would be, how-
ever, premature to predict the extinction of the HL in the
southern part of Canada as the observed phenotypic varia-
tion may be sufficient to allow a fraction of the population
to survive and adapt to the new local temperature condi-
tions. It is consequently imperative to increase monitoring
of HL populations in new areas that are at high risk of out-
breaks and to verify whether existing tools are still reliable
for predicting the insect’s phenology, particularly the
hatching of young larvae in spring. The tools could be made
more effective by incorporating newly acquired knowledge
of HL dormancy characteristics in eastern Canada.
Acknowledgements
The authors thank M. Marcotte, G. Hamel, and K. McCar-
thy for their technical assistance. They also acknowledge
Drs D. Quiring, J.N. McNeil, and two anonymous review-
ers for their helpful comments on an earlier version of the
manuscript and Ms. I. Lamarre for editorial assistance. This
research was supported by the Canadian Forest Service of
Natural Resources Canada and a Collaborative Research
and Development Grant from NSERC, CRDPJ 335085.
References
Agresti A (2002) Categorical Data Analysis. John Wiley & Sons,
New York, NY, USA.
Andrewartha HG (1952) Diapause in relation to the ecology of
insects. Biological Reviews 27: 50–107.
Armbruster P, Bradshaw WE, Ruegg K & Holzapfel CM (2001)
Geographic variation and the evolution of reproductive alloca-
tion in the pitcher-plant mosquito, Wyeomyia smithii. Evolu-
tion 55: 439–444.
van Asch M, van Tienderen PH, Holleman LJM & Visser ME
(2007) Predicting adaptation of phenology in response to cli-
mate change, an insect herbivore example. Global Change
Biology 13: 1596–1604.
Berthiaume R (2007) Ecologie Evolutive des Populations
d’Arpenteuse de la Pruche. PhD Thesis, Universite Laval,
Quebec, QC, Canada.
Braune HJ (1973) The role of temperature in controlling
obligatory diapause. Effects of Temperature on Ectothermic
Organisms. Ecological Implication and Mechanisms of
Compensation (ed. by W. Wieser), pp. 233–238. Springer-
Verlag, New York, NY, USA.
Broufas GD, Pappas ML & Koveo DS (2006) Effect of cold expo-
sure and photoperiod on diapause termination of the preda-
tory mite Euseius finlandicus (Acari: Phytoseiidae).
Physiological Ecology 35: 1216–1221.
Carroll WJ (1952) The Biology and External Morphology of the
Hemlock Looper, Lambdina fiscellaria (Guenee) in Newfound-
land (Lepidoptera: Geometridae). MSc Thesis, McGill Univer-
sity, Montreal, QC, Canada.
Danks HV (1987) Insect Dormancy: An Ecological Perspective.
Biological Survey of Canada (Terrestrial Arthropods), Ottawa,
ON, Canada.
Delisle J, West RJ & Bowers WW (1998) The relative performance
of pheromone and light traps in monitoring the seasonal activity
of both sexes of the eastern hemlock looper, Lambdina fiscellaria
fiscellaria. Entomologia Experimentalis et Applicata 89: 87–98.
Demont M & Blanckenhorn WU (2008) Genetic differentiation in
diapause response along a latitudinal cline in European yellow
dung fly populations. Ecological Entomology 33: 197–201.
Denlinger DL (2002) Regulation of diapause. Annual Review of
Entomology 47: 93–122.
Du Merle P (1999) Egg development and diapause: ecophysiolog-
ical and genetic basis of phenological polymorphism and adap-
tation to varied hosts in the green oak tortrix, Tortrix viridana
L. (Lepidoptera: Tortricidae). Journal of Insect Physiology 45:
599–611.
Easterling DR, Meehl GA, Parmesan C, Changnon SA, Karl TR &
Mearns LO (2000) Climate extremes: observations, modeling,
and impacts. Science 289: 2068–2074.
Environment Canada (2008) Canadian Climate Normals or Aver-
ages 1971–2000. http://climate.weatheroffice.ec.gc.ca/climate_
normals/index_e.html (accessed 27 March 2009).
Fantinou AA, Tsitsipis JA & Karandinos MG (1998) Diapause ter-
mination in Sesamia nonagrioides (Lepidoptera: Noctuidae)
under laboratory and field conditions. Environmental Ento-
mology 27: 53–58.
Fox CW (1997) Egg-size manipulations in the seed beetle Stator
limbatus: consequences for progeny growth. Canadian Journal
of Zoology 75: 1465–1473.
242 Delisle et al.

Gillespie RG & Roderick GK (2002) Arthropods on islands: colo-
nization, speciation, and conservation. Annual Review of
Entomology 47: 595–632.
Han EN & Bauce E (1997) Effects of early temperature exposure
on diapause development of spruce budworm (Lepidoptera:
Tortricidae). Environmental Entomology 26: 307–310.
Han EN & Bauce E (1998) Timing of diapause initiation, meta-
bolic changes and overwintering survival of the spruce bud-
worm, Choristoneura fumiferana. Ecological Entomology 23:
160–167.
Hanec WM (1966) Cold-hardiness in the forest tent caterpillar,
Malacosoma disstria Hubner (Lasiocampidae: Lepidoptera).
Journal of Insect Physiology 12: 1443–1449.
Hodek I & Hodkova M (1988) Multiple role of temperature dur-
ing insect diapause: a review. Entomologia Experimentalis
et Applicata 49: 153–165.
Hunter MD & McNeil JN (2000) Geographic and parental influ-
ences on diapause by a polyphagous insect herbivore. Agricul-
tural and Forest Entomology 2: 49–55.
Joy DA, Craig DA & Conn JE (2007) Genetic variation tracks eco-
logical segregation in Pacific island black flies. Heredity 99:
452–459.
Keena MA (1996) Comparison of the hatch of Lymantria dispar
(Lepidoptera: Lymantriidae) eggs from Russia and the United
States after exposure to different temperatures and durations
of low temperature. Annals of the Entomological Society of
America 89: 564–572.
Kfir R (1991) Duration of diapause in the stem borers, Busseola
fusca and Chilo partellus. Entomologia Experimentalis et Ap-
plicata 61: 265–270.
Kostal V (2006) Eco-physiological phases of insect diapause.
Journal of Insect Physiology 52: 113–127.
Levine E (1986) Termination of diapause and postdiapause devel-
opment in eggs of the stalk borer (Lepidoptera: Noctuidae).
Environmental Entomology 15: 403–408.
McNeil JN & Fields PG (1985) Seasonal diapause development
and diapause termination in the European skipper, Thymelicus
lineola (Ochs.). Journal of Insect Physiology 31: 467–470.
Morden RD & Waldbauer GP (1980) Diapause and its termina-
tion in the psychid moth, Thyridopteryx ephemeraeformis.
Entomologia Experimentalis et Applicata 28: 322–333.
Mousseau TA & Roff DA (1989) Adaptation to seasonality in a
cricket: patterns of phenotypic and genotypic variation in body
size and diapause expression along a cline in season length.
Evolution 43: 1483–1496.
Nilssen A & Tenow O (1990) Diapause, embryo growth and
supercooling capacity of Epirrita autumnata eggs from north-
ern Fennoscandia. Entomologia Experimentalis et Applicata
57: 39–55.
Parmesan C & Yohe G (2003) A globally coherent fingerprint of
climate change impacts across natural systems. Nature 421:
37–42.
Polavarapu S & Seabrook WD (1996) Seasonal diapause develop-
ment, effects of temperature and photoperiod on post-dia-
pause egg development, and validation of degree-day model
predicting larval eclosion of blueberry leaftier, Croesia curval-
ana (Kearfott) (Lepidoptera: Tortricidae). The Canadian
Entomologist 128: 187–198.
Raske AG & Farrell JE (1986) Aspects of hemlock looper moth
behaviour. Newfoundland Forest Research Centre. File Report
Study 2717: 1985–1986A.
Rose AH & Lindquist OH (1977) Insects of Eastern Spruces, Fir
and Hemlock. Forest Technical Report 23, Canadian Forest
Service, Ottawa, ON, Canada.
SAS Institute (1999) SAS ⁄ STAT� User’s Guide, Version 8. Cary,
NC, USA.
Sawchyn WW & Church NS (1973) The effects of temperature
and photoperiod on diapause development in the eggs of four
species of Lestes (Odonata: Zygoptera). Canadian Journal of
Zoology 51: 1257–1265.
Solbreck C (1978) Migration, diapause, and direct development
as alternative life histories in a seed bug, Neacoryphus bicrucis.
Evolution of Insect Migration and Diapause (ed. by H Dingle),
pp. 196–217. Springler-Verlag, Berlin, Germany.
Sota T & Mogi M (1992) Interspecific variation in desiccation
survival time of Aedes (Stegomyia) mosquito eggs is correlated
with habitat and egg size. Oecologia 90: 353–358.
Sullivan CR (1965) Laboratory and field investigations on the
ability of eggs of the European pine sawfly, Neodiprion sertifer
(Geoffroy) to withstand low winter temperatures. The Cana-
dian Entomologist 97: 978–993.
Tauber MJ & Tauber CA (1976) Insect seasonality: diapause
maintenance, termination, and postdiapause development.
Annual Review of Entomology 21: 81–107.
Tauber MJ, Tauber CA & Masaki S (1986) Seasonal Adaptations
of Insects. Oxford University Press, New York, NY, USA.
Tauber MJ, Tauber CA, Ruberson JR, Tauber AJ & Abrahamson
LP (1990) Dormancy in Lymantria dispar (Lepidoptera:
Lymantriidae): analysis of photoperiodic and thermal
responses. Annals of the Entomological Society of America 83:
494–503.
Wolter KM (1985) Introduction to Variance Estimation.
Springer, New York, NY, USA.
Egg dormancy in hemlock looper 243
Top Related
Copyright © 2022 FDOKUMEN

