







Bahasa
Halaman
Hukum


The cell wall architecture of Candida albicans wild-typecells and cell wall-defective mutants
J. C. Kapteyn,1* L. L. Hoyer,2 J. E. Hecht,2
W. H. MuÈ ller,3 A. Andel,1 A. J. Verkleij,3 M. Makarow,4,5
H. Van Den Ende1 and F. M. Klis1
1Swammerdam Institute of Life Sciences, University of
Amsterdam, Kruislaan 318, 1098 SM Amsterdam,
The Netherlands.2Department of Veterinary Pathobiology, College of
Veterinary Medicine, University of Illinois, Urbana,
IL 61802, USA.3Department of Molecular Cell Biology, University of
Utrecht, Utrecht, The Netherlands.4Institute of Biotechnology, University of Helsinki,
Helsinki, Finland.5Department of Biochemistry and Biotechnology,
Faculty of Medicine, University of Kuopio, Finland.
Summary
In Candida albicans wild-type cells, the b1,6-gluca-
nase-extractable glycosylphosphatidylinositol (GPI)-
dependent cell wall proteins (CWPs) account for
about 88% of all covalently linked CWPs. Approxi-
mately 90% of these GPI-CWPs, including Als1p and
Als3p, are attached via b1,6-glucan to b1,3-glucan.
The remaining GPI-CWPs are linked through b1,6-
glucan to chitin. The b1,6-glucanase-resistant protein
fraction is small and consists of Pir-related CWPs,
which are attached to b1,3-glucan through an alkali-
labile linkage. Immunogold labelling and Western
analysis, using an antiserum directed against Sac-
charomyces cerevisiae Pir2p/Hsp150, point to the
localization of at least two differentially expressed
Pir2 homologues in the cell wall of C. albicans. In
mnn9D and pmt1D mutant strains, which are defective
in N- and O-glycosylation of proteins respectively, we
observed enhanced chitin levels together with an
increased coupling of GPI-CWPs through b1,6-glucan
to chitin. In these cells, the level of Pir-CWPs was
slightly upregulated. A slightly increased incorpora-
tion of Pir proteins was also observed in a b1,6-
glucan-de®cient hemizygous kre6D mutant. Taken
together, these observations show that C. albicans
follows the same basic rules as S. cerevisiae in con-
structing a cell wall and indicate that a cell wall
salvage mechanism is activated when Candida cells
are confronted with cell wall weakening.
Introduction
The cell wall of the opportunistic human pathogenic fun-
gus Candida albicans is composed of complex polymers
of glucose (b1,3- and b1,6-glucan), chains of N-acetylglu-
cosamine (chitin) and cell wall mannoproteins (Shepherd,
1987; Fleet, 1991). The b-glucans are the main constitu-
ents, accounting for 50±60% by weight of the cell wall.
Together with chitin, a relatively minor component (0.6±
3%), the glucans form a rigid skeleton, responsible for
the shape of the cell and its physical strength (Shepherd,
1987). The mannoproteins represent 30±40% of the wall
and determine the cell surface properties, enabling Can-
dida albicans cells to adhere to host tissues and to immu-
nomodulate the host immune response (Chaf®n et al.,
1998). Our knowledge about the molecular organization
of the Candida cell wall is limited, but considerable pro-
gress has been made in unravelling the cell wall architec-
ture of Saccharomyces cerevisiae (Klis et al., 1997; KollaÂr
et al., 1997; Lipke and Ovalle, 1998; Kapteyn et al.,
1999a,b; Smits et al., 1999). S. cerevisiae is generally
considered to be a valuable model organism for studying
cell wall construction in Candida (Klis et al., 1997; Chaf®n
et al., 1998). Recently, a tentative model of the molecular
organization of the S. cerevisiae cell wall has been pre-
sented (Fig. 1; Lipke and Ovalle, 1998; Smits et al.,
1999). According to this cell wall model, the b1,3-glucan
molecules form a three-dimensional matrix surrounding
the entire cell. As these b1,3-glucan molecules are
branched, they may have multiple non-reducing ends.
These may function as attachment sites for cell wall pro-
teins (CWPs). Two main types of covalently linked
CWPs have been identi®ed, namely glycosylphosphatidy-
linositol (GPI)-dependent CWPs and Pir-CWPs (Kapteyn
et al., 1999b). The GPI-CWPs are attached via b1,6-
glucan to a non-reducing end of a b1,3-glucan molecule
(Fig. 1; Kapteyn et al., 1996, 1999b; KollaÂr et al., 1997).
A GPI-derived structure is responsible for the linkage of
this type of proteins to b1,6-glucan (KollaÂr et al., 1997;
Fujii et al., 1999). The Pir-related CWPs are directly
attached to b1,3-glucan through an, as yet unknown,
Molecular Microbiology (2000) 35(3), 601±611
Q 2000 Blackwell Science Ltd
Received 2 September, 1999; revised 13 October, 1999; accepted 18October, 1999.*For correspondence. E-mail [email protected]; Tel. (�31) 20 5257850; Fax (�31) 20 525 7934.

base-labile linkage (Fig. 1; Mrsa et al., 1997; Kapteyn et
al., 1999a). After cytokinesis, chitin molecules may also
attach to the non-reducing ends of the b1,3-glucan mol-
ecules and, less frequently, also to the non-reducing
ends of highly branched b1,6-glucan molecules (KollaÂr et
al., 1995, 1997), thereby strengthening the cell wall (Fig.
1; Lipke and Ovalle, 1998; Smits et al., 1999).
The cell wall structure of S. cerevisiae is highly ¯exible.
For instance, yeast cells have been shown to be able to
compensate for defects in b1,3-glucan synthesis and/or
assembly by altering the molecular organization of the
wall. This compensation is characterized by a several-
fold increase in chitin content (Popolo et al., 1997) and in
an augmented number of linkages between chitin and
b1,6-glucan, the attachment moiety for GPI-CWPs (Kap-
teyn et al., 1997; Lipke and Ovalle, 1998; Ram et al.,
1998). As a consequence of this compensatory reaction,
a 20-fold increase was observed in the level of GPI-
CWPs that were resistant to b1,3-glucanase-extraction
(Kapteyn et al., 1997). When yeast cells were confronted
with loss or faulty assembly of b1,6-glucan, resulting in
the release of GPI-CWPs into the medium, a strong
increase in the level of b1,3-glucan-bound Pir proteins
was observed (Kapteyn et al., 1999a). In addition, chitin
deposition was also massively enhanced (Kapteyn et al.,
1999a; Ketela et al., 1999). This latter phenotype is shared
by many other yeast cell wall-defective mutants (Ram et
al., 1994; Gentzsch and Tanner, 1996; Lussier et al.,
1997; Dallies et al., 1998). The observed increase in chitin
levels may be mediated by stimulation of Chs3p activity
through the putative cell wall stress sensor Mid2p (Ketela
et al., 1999). High chitin levels have also been reported for
a b1,6-glucan-de®cient Candida glabrata kre9D strain
(Nagahashi et al., 1998) and a C. albicans phr1D mutant,
which is defective in b-glucan assembly (Popolo and Vai,
1998).
In this paper, we present evidence for C. albicans fol-
lowing the same rules as S. cerevisiae in constructing a
cell wall. In Candida cells, < 88% of all covalently linked
CWPs, including proteins encoded by the ALS gene
family, are immobilized in the wall as GPI modules,
whereas the remaining proteins are attached as Pir mod-
ules. At least two CWPs were identi®ed, showing signi®-
cant homology to S. cerevisiae Pir2p/Hsp150. In cell
wall-defective mutants, we observed changes in the rela-
tive frequency of GPI and Pir proteins, enhanced chitin
levels and increased cross-linking of GPI-CWPs through
b1,6-glucan to chitin. These and other data therefore
suggest that, like in S. cerevisiae, mechanisms are acti-
vated by which C. albicans is able to counteract cell wall
damage.
Results
Identi®cation of putative GPI-dependent CWPs,
including Als1p and Als3p, in cell walls of C. albicans
Fig. 1 shows the current model of the S. cerevisiae cell
wall. Our experiments were aimed at determining whether
the same model applies to C. albicans. In order to demon-
strate that C. albicans contains b1,6-glucan-linked GPI-
dependent CWPs, we analysed b1,6-glucanase digests
of SDS-extracted cell walls from C. albicans CAI4 yeast
forms grown in SC medium at 288C. Western blotting
using Concanavalin A (ConA) peroxidase revealed a
large smear and four relatively discrete mannosylated pro-
tein bands with an apparent molecular mass of 220, 170,
100 and 65 kDa respectively (Fig. 2A, lane 1). Anti-b1,6-
glucan antiserum did not recognize any of this material,
indicating that the b1,6-glucanase did not leave any
detectable b1,6-glucan epitope attached to the proteins
(data not shown).
Hoyer et al. (1998a,b, 1999) have shown the existence
of a family of cell surface proteins that are probably
involved in cell adhesion. In view of their homology to a-
agglutinin, a sexual adhesion protein, the corresponding
genes have been called agglutinin-like sequences (ALS
genes). Reprobing of the stripped blot with an anti-Alsp
antiserum (Hoyer et al., 1998b) revealed a band at
< 600 kDa (Fig. 2B, lane 1). Endoglycosidase (Endo) H
treatment of the Als protein band reduced its apparent
molecular mass by < 120 kDa and demonstrated the pre-
sence of N-linked carbohydrate (Fig. 2B, lane 2). Because
the anti-Als antiserum recognizes various Als proteins
(Hoyer et al., 1999), Northern blot analysis was performed
to determine which ALS genes were transcribed in SC-
grown yeast forms. In a ®rst screen, we used the ALS1
tandem repeat probe and the ALS5 tandem repeat
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Fig. 1. Molecular architecture of the cell wall of Saccharomycescerevisiae and Candida albicans. The arrows denote the orientationof the polysaccharides from a non-reducing end to the reducingend. It is important to realize that mature GPI-dependent CWPs,once they are incorporated in the cell wall, are believed to have losttheir lipid moiety, and that therefore they are not attached to theplasma membrane (Lu et al., 1995; Kolla r et al., 1997; Fujii et al.,1999). Note that a relatively small number of chitin chains is linkedto b1,6-glucan molecules. The model is based on data from KollaÂret al. (1995, 1997), Kapteyn et al. (1997, 1999a) and Smits et al.(1999).
602 J. C. Kapteyn et al.
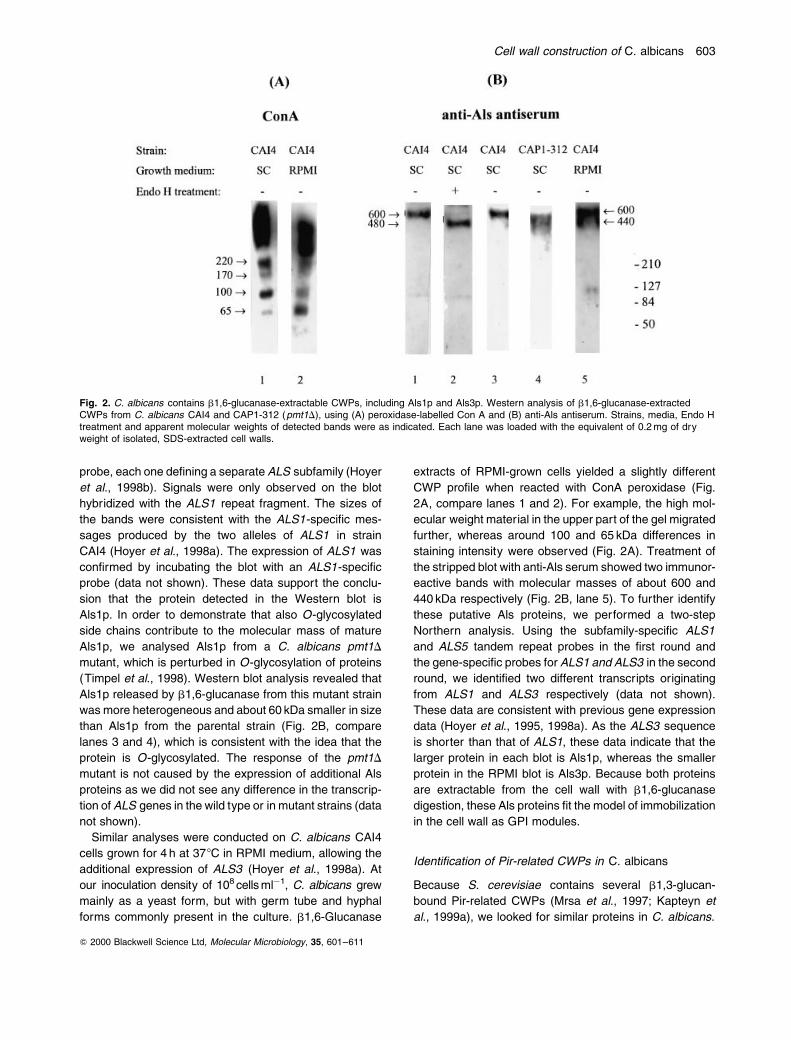
probe, each one de®ning a separate ALS subfamily (Hoyer
et al., 1998b). Signals were only observed on the blot
hybridized with the ALS1 repeat fragment. The sizes of
the bands were consistent with the ALS1-speci®c mes-
sages produced by the two alleles of ALS1 in strain
CAI4 (Hoyer et al., 1998a). The expression of ALS1 was
con®rmed by incubating the blot with an ALS1-speci®c
probe (data not shown). These data support the conclu-
sion that the protein detected in the Western blot is
Als1p. In order to demonstrate that also O-glycosylated
side chains contribute to the molecular mass of mature
Als1p, we analysed Als1p from a C. albicans pmt1D
mutant, which is perturbed in O-glycosylation of proteins
(Timpel et al., 1998). Western blot analysis revealed that
Als1p released by b1,6-glucanase from this mutant strain
was more heterogeneous and about 60 kDa smaller in size
than Als1p from the parental strain (Fig. 2B, compare
lanes 3 and 4), which is consistent with the idea that the
protein is O-glycosylated. The response of the pmt1D
mutant is not caused by the expression of additional Als
proteins as we did not see any difference in the transcrip-
tion of ALS genes in the wild type or in mutant strains (data
not shown).
Similar analyses were conducted on C. albicans CAI4
cells grown for 4 h at 378C in RPMI medium, allowing the
additional expression of ALS3 (Hoyer et al., 1998a). At
our inoculation density of 108 cells mlÿ1, C. albicans grew
mainly as a yeast form, but with germ tube and hyphal
forms commonly present in the culture. b1,6-Glucanase
extracts of RPMI-grown cells yielded a slightly different
CWP pro®le when reacted with ConA peroxidase (Fig.
2A, compare lanes 1 and 2). For example, the high mol-
ecular weight material in the upper part of the gel migrated
further, whereas around 100 and 65 kDa differences in
staining intensity were observed (Fig. 2A). Treatment of
the stripped blot with anti-Als serum showed two immunor-
eactive bands with molecular masses of about 600 and
440 kDa respectively (Fig. 2B, lane 5). To further identify
these putative Als proteins, we performed a two-step
Northern analysis. Using the subfamily-speci®c ALS1
and ALS5 tandem repeat probes in the ®rst round and
the gene-speci®c probes for ALS1 and ALS3 in the second
round, we identi®ed two different transcripts originating
from ALS1 and ALS3 respectively (data not shown).
These data are consistent with previous gene expression
data (Hoyer et al., 1995, 1998a). As the ALS3 sequence
is shorter than that of ALS1, these data indicate that the
larger protein in each blot is Als1p, whereas the smaller
protein in the RPMI blot is Als3p. Because both proteins
are extractable from the cell wall with b1,6-glucanase
digestion, these Als proteins ®t the model of immobilization
in the cell wall as GPI modules.
Identi®cation of Pir-related CWPs in C. albicans
Because S. cerevisiae contains several b1,3-glucan-
bound Pir-related CWPs (Mrsa et al., 1997; Kapteyn et
al., 1999a), we looked for similar proteins in C. albicans.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Fig. 2. C. albicans contains b1,6-glucanase-extractable CWPs, including Als1p and Als3p. Western analysis of b1,6-glucanase-extractedCWPs from C. albicans CAI4 and CAP1-312 (pmt1D), using (A) peroxidase-labelled Con A and (B) anti-Als antiserum. Strains, media, Endo Htreatment and apparent molecular weights of detected bands were as indicated. Each lane was loaded with the equivalent of 0.2 mg of dryweight of isolated, SDS-extracted cell walls.
Cell wall construction of C. albicans 603

For these studies, we utilized the b1,6-glucanase-resis-
tant b1,3-glucanase-extractable CWPs from both RPMI-
and SC-grown cultures and an anti-Pir2p antiserum,
which is known to recognize Pir2p-related proteins in vari-
ous yeast species (Russo et al., 1992). Western blots of
proteins from the RPMI culture yielded a single band
with an apparent molecular weight of 235 kDa (Fig. 3,
lane 1). After Endo H treatment, the molecular mass of
the protein decreased to about 200 kDa (Fig. 3, lane 2).
Before and after Endo H treatment, the antigenic Pir2p
homologue detected was also reactive with an antiserum
raised against b1,3-glucan, indicating that the protein
was associated with a b1,3-glucan moiety (Fig. 3, lanes
6 and 7). However, no immunoreactive bands were
observed with the anti-b1,6-glucan antiserum (data not
shown). Upon very mild alkali treatment, the b1,3-glucan
epitope was lost (Fig. 3, lane 8), which was accompanied
by a drop in molecular size of the Endo H-treated Pir2
homologue by < 50 kDa (Fig. 3, lane 3). These data
strongly suggest that C. albicans incorporates at least
one Pir-related protein into its cell wall through an alkali-
sensitive linkage to b1,3-glucan. Clear labelling of the C.
albicans cell wall by immunogold electron microscopy
with the anti-Pir2p serum supports this conclusion (Fig.
4A and B). The intensity of the immunogold labelling is,
however, slightly less than that observed in cell walls of
S. cerevisiae (Fig. 4C and D). Hardly any gold labelling
was observed with preimmune serum (data not shown).
These ®ndings are therefore in accordance with earlier
data from Mormeneo et al. (1994), who were the ®rst
investigators to release proteinaceous material from Can-
dida by mild alkali extraction.
Interestingly, when cells were grown in SC medium,
pH 4.5, two immunoreactive bands were observed in the
b1,6-glucanase-resistant b1,3-glucanase-extractable wall
fraction, which upon Endo H treatment had apparent mol-
ecular weights of 200 and 320 kDa, respectively (Fig. 3,
lanes 4 and 5), suggesting the existence of more than
one Pir protein in C. albicans. Further research is required
to establish whether C. albicans indeed contains a family
of PIR genes as has been described for S. cerevisiae
(Toh-E et al., 1993; Mrsa et al., 1997). Notably, homology
searches of the S. cerevisiae PIR1, PIR2, PIR3 and PIR4/
CCW5/CCW11 genes with sequences of the Candida
genome data bank (http://candida.stanford.edu/ybc.html)
resulted in the identi®cation of a putative C. albicans PIR-
related ORF, 265178D12.yl.seq (data not shown). The
translated sequence of this ORF has 32±45% identity to,
and 48±60% similarity with, the four Pir-CWPs of S. cere-
visiae. The C. albicans PIR-related ORF is predicted to
encode a secretory polypeptide of 104 amino acids with
a putative N-terminal signal sequence of 18 amino acids
(PSORTII).
Changes in cell wall architecture of cell wall-defective
mutants
Because the cell wall of S. cerevisiae is a highly dynamic
structure, we reasoned that the C. albicans cell wall might
also have a ¯exible architecture. To explore this possibi-
lity, we utilized several C. albicans mutant strains, includ-
ing mnn9D (defective in N-glycosylation of proteins),
pmt1D (defective in O-glycosylation) and a hemizygous
kre6D strain, which has an 80% reduction in the level of
b1,6-glucan (Mio et al., 1997). As expected from earlier
reports on S. cerevisiae b1,6-glucan-de®cient kreD
mutants (Lu et al., 1995; Jiang et al., 1996; Kapteyn et
al., 1999a), the hemizygous kre6D mutant secreted large
amounts of Als proteins into the culture supernatant (Fig.
5A). Because these cells were grown in RPMI medium,
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Fig. 3. C. albicans Pir2 homologues areattached to b1,3-glucan via an alkali-sensitivelinkage. Western analysis of b1,6-glucanase-resistant b1,3-glucanase-extracted CWPsfrom C. albicans CAI4 using (A) anti-Pir2pantiserum and (B) anti-b1,3-glucan antiserum.Lanes 1±3 and 6±8, material from RPMI-grown cultures; lanes 4 and 5, material fromSC-grown cultures. Each lane was loadedwith the equivalent of 0.4 mg of dry weight ofisolated SDS-extracted cell walls. Endo Htreatments and apparent molecular weights ofdetected bands were as indicated.
604 J. C. Kapteyn et al.

the secreted immunoreactive proteins probably represent
both Als1p and Als3p. These observations provide
additional evidence that Als proteins are immobilized in
the cell wall through linkage to b1,6-glucan. No enhanced
secretion of Als proteins was observed for the pmt1 or
mnn9 null strains (data not shown).
To investigate the molecular cell wall organization of
C. albicans CAI4 and the selected mutants, we ®rst
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Fig. 4. Immunogold labelling of Pir2-related proteins in cell walls of Candida albicans. As a control, Saccharomyces cerevisiae wild-type cellsare shown. A and B, C. albicans CAI4; C and D, S. cerevisiae wild-type TR99. Hardly any signals were obtained with the preimmune serum.
Cell wall construction of C. albicans 605

measured chitin levels in SC-grown cells. The glycosyla-
tion-de®cient pmt1 and mnn9 null mutants showed a
two- to threefold increase in the level of chitin. Consistent
with an essential role of chitin in strengthening the wall, the
pmt1 and mnn9 null strains were found to be more sensi-
tive to exochitinase and nikkomycin Z, a competitive inhibi-
tor of chitin synthases (Choi et al., 1994; Gaughran et al.,
1994), than the parental strain (data not shown). The chitin
level in the hemizygous kre6D strain was unaffected
(Table 1) and consistent with this the hemizygous kre6D
strain was as sensitive to nikkomycin Z as the parental
strain (data not shown). Lack of altered chitin levels in
the hemizygous kre6D strain was unexpected because
similar mutations in S. cerevisiae haploid strains result in
increased chitin content (Dallies et al., 1998; Kapteyn et
al., 1999a; Ketela et al., 1999). Next, we investigated
whether the increased chitin levels in pmt1 and mnn9
null mutants were accompanied by an increase in the
amount of b1,3-glucanase-resistant GPI-CWPs because
of an enhanced coupling of these proteins through b1,6-
glucan to chitin. In wild-type cells and in the hemizygous
kre6D strain, about 8±9% of the total radiolabelled
CWPs were resistant to b1,3-glucanase extraction, whereas
this percentage was increased to 21% and 33% in the
pmt1 and the mnn9 null mutants respectively (Table 1).
Subsequent treatment with exochitinase resulted in
the solubilization of almost all b1,3-glucanase-resistant
proteins (Table 1). Consistent with these results, Western
analyses demonstrated that the b1,3-glucanase-resistant
exochitinase-soluble mnn9D (data not shown) and
pmt1D wall fractions contained higher levels of b1,6-gluco-
sylated ConA-reactive GPI-CWPs (Fig. 6A and B), includ-
ing Als1p (Fig. 6C). Notably, the ConA-reactive CWP
bands released from the pmt1D strain were generally
smaller and more heterogeneous than those derived
from the parental strain (Fig. 6A).
We also determined the level of b1,6-glucanase-resistant
b1,3-glucanase-extractable Pir proteins in the strain CAI4
and in the selected C. albicans mutants. In CAI4, about
12% of the CWPs were resistant to b1,6-glucanase diges-
tion, a value that is slightly lower than that observed in S.
cerevisiae wild-type cells (Kapteyn et al., 1999a). Similar
amounts of b1,6-glucanase-resistant CWPs were only
slightly enhanced in the mutant strains (Table 1). This result
was con®rmed using Western analyses, which showed that
the b1,6-glucanase-resistant b1,3-glucanase extract from
both the glycosylation mutants (data not shown) and the
hemizygous kre6D strain contained moderately increased
levels of the putative Pir2 homologue compared with the cor-
responding wild-type fraction (Fig. 5B).
Discussion
In this study, we investigated whether the structure of the
C. albicans cell wall resembles the molecular organization
of the S. cerevisiae cell wall (Fig. 1; Lipke and Ovalle,
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Fig. 5. b1,6-Glucan-de®cient hemizygous C.albicans kre6D strain is affected in theretention of GPI-CWPs and shows anincreased incorporation of Pir2 homologues inthe cell wall. Western analysis of (A) proteinsreleased into the medium, using the anti-Alsantiserum (both lanes were loaded with theequivalent of 2 ml of culture supernatant fromcells grown to an OD600 of 2.0 (3 ´ 107
cells mlÿ1) and (B) b1,6-glucanase-resistantb1,3-glucanase-extracted CWPs, using theanti-Pir2p antiserum (all four lanes wereloaded with the equivalent of 0.4 mg of dryweight of isolated SDS-extracted cell walls).Strains, molecular weight marker (210 kDa)and Endo H treatments are as indicated.
Table 1. Chitin content and extractability of SDS-resistant 14C-labelled CWPs by sequential cell wall digestions with different hydrolytic enzymes(comparison of Candida albicans cells and cell wall-defective mutants grown in SC medium at 288C).
Solubilized (%) by
b1,3-Glucanase Exochitinase b1,6-Glucanase b1,3-GlucanaseStrain Chitin content (%) (first digestion) (second digestion) (first digestion) (second digestion)
CAI-4 2.4 91.2 99.7 88.0 99.2Dmnn9 /Dmnn9 6.2 67.7 97.1 84.4 98.5Dpmt1 /Dpmt1 4.7 79.1 99.8 84.5 98.0KRE6 /Dkre6 2.3 91.8 99.9 84.2 98.2
606 J. C. Kapteyn et al.

1998; Smits et al., 1999). We found many similarities
between the two cell wall structures. As in S. cerevisiae,
the C. albicans cell wall contains GPI-CWP modules that
are linked to b1,6-glucan (Fig. 2). C. albicans also con-
tains Pir-CWP modules that are linked through an alkali-
labile linkage to b1,3-glucan and are resistant to b1,6-
glucanase treatment (Figs 3 and 4). We demonstrated
immunologically the existence of at least two Pir-CWPs.
It was also found that, like S. cerevisiae, C. albicans is
able to alter its cell wall structure in response to cell wall
damage or weakening (Table 1; Figs 5 and 6). A signi®-
cant difference between the two cell wall structures, how-
ever, is that the relative amount of b1,3-glucanase-
resistant CWPs in C. albicans is approximately fourfold
higher than that in S. cerevisiae wild-type cells (Table 1;
Kapteyn et al., 1997, 1999a). As both organisms have
similar chitin levels, this suggests that there are more lin-
kages between chitin and b1,6-glucan in C. albicans
than in S. cerevisiae. The two- to threefold increased chitin
deposition in cell walls of the glycosylation-de®cient C.
albicans strains led to a proportional increase in the
amount of b1,3-glucanase-resistant CWPs, suggesting
that the additional chitin molecules synthesized under
these cell wall stress conditions are coupled to b1,6-
glucan rather than to b1,3-glucan.
As well as de®ning the basic architecture of the C. albi-
cans cell wall, our studies also contributed to a greater
understanding of the structure and localization of Als pro-
teins. The work presented here leads to the following con-
clusions about these proteins: they are linked to b1,6-
glucan and they are N- and O-glycosylated. Amino acid
sequences predicted from the ALS genes are consistent
with these conclusions. Each Als protein encodes a hydro-
phobic C terminus with characteristics of a GPI-anchor-
addition site (Hoyer et al., 1998b). Although our data do
not precisely identify an Als protein intermediate with an
attached GPI moiety, they serve to emphasize the paral-
lels between what is known for S. cerevisiae GPI-CWPs
and the C. albicans Als family proteins. Als proteins are
linked in the C. albicans cell wall in a manner consistent
with proteins that carry a transiently linked GPI moiety.
N-Glycosylation of Als proteins is probable due to the pre-
sence of consensus N-glycosylation sites, predominantly
in the tandem repeat and C-terminal domain regions of
each protein (Hoyer et al., 1995, 1998a,b). O-Glycosyla-
tion of Als proteins is assumed because of the serine/
threonine-rich content of the tandem repeat and C-term-
inal domains (Hoyer et al., 1995, 1998a,b).
Although a clear picture is beginning to develop as to
how CWPs are immobilized in the cell wall of C. albicans
and S. cerevisiae, the function of most of these proteins
is unknown (Chaf®n et al., 1998; Kapteyn et al., 1999b;
Smits et al., 1999). Proteins of the C. albicans Als family
may constitute an exception, however, as heterologous
expression of ALA1/ALS5 and ALS1 in S. cerevisiae con-
fers adherence to human cells (Gaur and Klotz, 1997; Fu
et al., 1998), suggesting that ALS genes may encode
adhesins. Recently, Hwp1p, another GPI-dependent
CWP of candidal germ tubes and hyphae, was demon-
strated to mediate covalent attachment of Candida cells
to human epithelial tissue, as a substrate for human
transglutaminase activity (Staab et al., 1999). In yeast,
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Fig. 6. A C. albicans pmt1 null mutant showing an increased coupling of GPI-CWPs to chitin through b1,6-glucan. Western analysis of b1,3-glucanase-resistant exochitinase-extracted CWPs from CAI4 and pmt1D cells grown in SC medium, using (A) peroxidase-labelled Con A, (B)anti-b1,6-glucan antiserum, and (C) anti-Als antiserum. Each lane was loaded with the equivalent of 0.5 mg of dry weight of isolated SDS-extracted cell walls. Strains, b1,6-glucanase treatments and molecular weight markers are as indicated.
Cell wall construction of C. albicans 607

Pir-CWPs confer resistance to the plant antifungal protein
osmotin (Yun et al., 1997). Sequential disruption of the
PIR-related genes CCW5, PIR2/CCW7, PIR1/CCW6
and PIR3/CCW8 led to reduction in growth, swollen and
irregular cell morphology and an increased susceptibility
to calco¯uor white (Mrsa and Tanner, 1999). In addition,
the triple and quadruple deletants showed a diminished
mating ability (Mrsa and Tanner, 1999). Expression of
the best-studied Pir protein Pir2p is induced by a variety
of stress conditions, including heat shock, nitrogen starva-
tion, oxidative stress, aluminium and cell wall damage
owing to reduced levels of b1,6-glucan (Russo et al.,
1992, 1993; Ezaki et al., 1998; Kapteyn et al., 1999a).
However, a clear function has been ascribed neither to
Pir2p/Hsp150 nor to any of the other Pir proteins. Like
S. cerevisiae Pir2p/Hsp150, the Candida Pir2-related pro-
tein seems to be expressed at a higher level under con-
ditions of cell wall weakening (Fig. 5). Whether the
expression of C. albicans Pir proteins is differentially regu-
lated and whether these proteins play a role in the patho-
genicity of C. albicans is currently being explored.
Experimental procedures
C. albicans strains and growth conditions
The Candida strains used are CAI4 (ura3D::imm434/ura3D::imm434), CAP1-312 (pmt1D::hisG-URA3-hisG/pmt1D::hisG in CAI4), SSCA-2 (mnn9D::hisG-URA3-hisG/mnn9D::hisG in CAI4) and 211 (KRE6/kre6D::hisG-URA3-hisG in CAI4). Cells were grown at 288C in synthetic complete(SC) medium, containing 0.17% (w/v) yeast nitrogen basewithout amino acids and ammonium sulphate (Difco), 2%(w/v) glucose, 0.5% (w/v) casein hydrolysate, 0.5% (w/v)ammonium sulphate, 0.25% (w/v) succinate and 0.01% (w/v) uridine at pH 4.5. In some experiments, cells were precul-tured overnight in SC medium at 288C, washed, resuspendedand grown for a further 4 h at 378C in RPMI-1640 medium(pH 7.2), which contains 2% (w/v) glucose, various inorganicsalts and amino acids, as de®ned by the manufacturer (GibcoBRL), with an initial cell density of 108 cells mlÿ1.
Immunogold labelling of Pir2-related proteins
Cells grown in SC medium were taken up by capillary action insingle dry cellulose capillary tubes of about 5±10 mm inlength (Hohenberg et al., 1994). After ®lling, the capillarytube was submerged in 1-hexadecane and cut into piecesabout 2.5 mm long. The 2.5 mm tubes with cells were sealedat their ends by pressing with a blunt razor blade. Fourtubes were sandwiched in aluminium specimen holders.Thereafter, this sandwich was inserted in the holder of aLeica high-pressure freezer. After high-pressure freezingwith a Leica EM HPF, the sandwich was put into liquid nitro-gen (LN2). The two specimen holders were separated underLN2. The capillary tubes still attached to one of the specimenaluminium holders were freed from adhering 1-hexadecane
under LN2 by gently scraping with a ®ne needle. Subse-quently, the capillary tubes attached to the specimen holderwere transferred to a screw-capped vial, which contained fro-zen substitution medium consisting of 0.3% uranyl acetateand 0.01% glutaraldehyde in anhydrous methanol and a min-iature transfer basket (Hohenberg et al., 1994). This vial wasplaced in the substitution apparatus CS-auto (Sitte et al.,1985) at ÿ908C for 38 h. Subsequently, the temperaturewas raised to ÿ358C at a speed of 78C hÿ1. The freeze-sub-stitution medium was exchanged for methanol. The speci-men holders were removed from the vial. The capillarytubes were in®ltrated with an increasing concentration ofLowicryl HM20 (Carlemalm et al., 1982): 25% for 1 h; 50%for 1 h; 70% for 1 h and overnight; then at ÿ208C 100% for2´ 2 h; 100% over the weekend; freshly made 100% for2 h. Thereafter, the capillary tubes were embedded in100% Lowicryl HM20. Polymerization was carried out atÿ208C for 42 h with a UV source attachment (Sitte et al.,1985). After 1 day of curing under UV light at room tempera-ture, 80 nm sections were cut with a Diatome diamond knifeand mounted on 1.1% pioloform-coated (Pioloform resin2295/0; Polaron), carbon-evaporated 150 mesh nickelgrids. After embedment, labelling was performed on sectionsof cells to locate Pir2-related proteins, using rabbit serum1660 recognizing Pir2p/Hsp150 (1:500) (Russo et al.,1992). The antigen-antibody complexes were visualized withgoat anti-rabbit antibodies (1 : 20) conjugated with 10-nmgold particles (Aurion).
Nucleic acid blots
Total RNA extraction, formaldehyde gel electrophoresis andNorthern blotting were performed to analyse ALS geneexpression (Hoyer et al., 1998a,1998b). Northern blots were®rst probed with an 875 bp Kpn I fragment of pLH21 thatencodes only ALS1 tandem repeat sequences (Hoyer et al.,1995) and a PCR-ampli®ed fragment that includes sequencesfrom the ALS5 tandem repeat region (Hoyer et al., 1998b).These probes each speci®cally recognize one of the two sub-families of the ALS gene family: the ALS1 repeat probe frag-ment recognizes ALS1, ALS2, ALS3 and ALS4, whereas theALS5 repeat probe fragment recognizes ALS5, ALS6 andALS7 (Hoyer et al., 1998b; L. L. Hoyer, unpublished). Basedon the signals observed with the initial screening with the tan-dem repeat probes, positive signals were precisely identi®edusing ALS gene-speci®c probes. In this case, an ALS1-speci-®c Xba I±HindII fragment from the 38 end of ALS1 (Hoyer etal., 1995) and a PCR-ampli®ed ALS3-speci®c fragment fromthe 38 end of the coding region (Hoyer et al., 1998a) wereused. This procedure allows the correct identi®cation of theseven known ALS genes.
Isolation of cell wall proteins
Cells were harvested, washed and homogenized as has beendescribed before (Montijn et al., 1994; Kapteyn et al., 1995).Subsequently, cell walls were isolated and washed exten-sively with 1 M NaCl. Isolated cell walls were boiled twice inthe presence of SDS, EDTA and b-mercaptoethanol to solubi-lize the non-covalently linked CWPs and to remove any
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
608 J. C. Kapteyn et al.

contaminants derived from the cytosol and/or plasma mem-brane (Mrsa et al., 1997; Klis et al., 1998). SDS-extractedcell walls were washed and treated with recombinant Tricho-derma harzianum endo-b1,6-glucanase [0.8 U gÿ1 (wetweight of cell walls)], and puri®ed according to the proceduredescribed by Bom et al. (1998) to isolate the GPI-CWPs (Kap-teyn et al., 1996). Subsequently, b1,6-glucanase-treated cellwalls were digested with Quantazyme (Quantum Biotechnol-ogies) [1500 U gÿ1 (wet weight of cell walls)], a recombinantb1,3-glucanase (Kapteyn et al., 1995), to release the b1,6-glucanase-resistant Pir-CWPs. In some experiments, Quan-tazyme-digested cell walls were further digested with exochi-tinase [0.3 U gÿ1 (wet weight of cell walls)] as has beendescribed previously (KollaÂr et al., 1995). All enzymatic diges-tions were stopped by adding SDS at a ®nal concentration of0.4% (w/v) and heating for 3 min at 1008C.
The ef®cacy of the various hydrolytic enzymes to solubilizeCWPs was studied by extracting isolated walls from cells thathad been radiolabelled with [14C]-protein hydrolysate accord-ing to the procedure described by Kapteyn et al. (1997). Inbrief, cells were precultured in SC medium to OD600� 1.0(equivalent to 107 cells mlÿ1). Cells were collected, washedand resuspended in 100 ml of labelling medium, containing0.17% (w/v) yeast nitrogen base without amino acids orammonium sulphate (Difco), 2% (w/v) glucose, 0.05% (w/v)casein hydrolysate, 0.5% (w/v) ammonium sulphate, 0.25%(w/v) succinate and 0.001% (w/v) uracil with an initial densityof 107 cells mlÿ1, and were incubated for 15 min at 288C. Sub-sequently, [14C]-protein hydrolysate (0.46 Mbq; Amersham)was added, and the cultures were grown for 3.5 h at 288C.Labelled cells were collected and homogenized as indicatedabove. Cell walls were isolated, SDS extracted, rinsed anddigested with hydrolytic enzymes as described above. Proteincontents of the different fractions were calculated as percen-tages of total label incorporated into SDS-extracted walls.
Enzymatic and chemical treatments of CWPs
b1,6-Glucanase-resistant CWPs released by Quantazymewere treated very mildly with alkali according to Kapteyn etal. (1999a). In brief, Quantazyme digests were dialysed,freeze-dried and incubated for 4 h at 48C in 15 mM NaOH.The incubation was terminated by addition of neutralizingamounts of acetic acid, and alkali-treated CWPs were precipi-tated by addition of nine volumes of ice-cold methanol. After2 h on ice, the sample was centrifuged at 14 000 g for15 min, and the pellet was taken up in sample buffer asdescribed by Laemmli (1970). Part of b1,6-glucanase-released and Quantazyme-released CWPs were treatedwith Endo-N-acetylglucosaminidase H (Endo H) (BoehringerMannheim). CWPs dissolved in 50 ml of distilled water, con-taining 0.4% (w/v) SDS and 40 mM b-mercaptoethanol,were heated for 5 min at 1008C and were mixed with 50 mMsodium acetate, pH 5.5, containing 2.5 mM EDTA (150 ml).De-N-glycosylation was performed by adding 100 mU ofEndo H. After incubating at 378C for 17 h, Endo H-treatedCWPs were precipitated by adding nine volumes of ice-coldmethanol and were prepared for SDS±PAGE as describedabove. In some experiments, Quantazyme-resistant exochiti-nase-solubilized CWPs were digested with b1,6-glucanaseaccording to the procedure described by Kapteyn et al. (1996).
Analysis of cell wall proteins
CWPs were separated by electrophoresis using linear 2.2±20% polyacrylamide gels and were electrophoretically trans-ferred onto an Immobilon polyvinylidene di¯uoride (PVDF)membrane (Montijn et al., 1994). CWPs were visualized byprobing the membranes with peroxidase-labelled Concanava-lin A (ConA; 1 mg mlÿ1) in PBS, containing 3% (w/v) BSA,2.5 mM CaCl2 and 2.5 mM MnCl2 (Klis et al., 1998). Westernimmunoblot analyses were performed with af®nity-puri®edpolyclonal antisera speci®cally directed against b1,6-glucan,and b1,3-glucan as described previously (Kapteyn et al.,1997; Klis et al., 1998). Blots were also probed with an anti-serum raised against yeast Pir2p (Russo et al., 1992) accord-ing to Kapteyn et al. (1999a). Furthermore, we probed thePVDF ®lters with an anti-Als antiserum (Hoyer et al.,1998a). In this case, a serum dilution of 1:5000 in PBS, con-taining 5% (w/v) non-fat milk powder, was used, and beforethe blocking step the membranes were treated for 30 minwith 50 mM periodic acid and 100 mM sodium acetate(pH 4.5) to enhance the speci®city of the antiserum. In allcases, binding of antibodies was assessed with goat anti-rabbit IgG peroxidase (Pierce) at a dilution of 1:10 000 inPBS/5% (w/v) milk powder. The blots were visualized usingECL Western blotting detection reagents (Amersham)according to the manufacturer's instructions.
Determination of chitin content in cell walls
SDS-extracted cell walls (about 4 mg dry weight) were hydro-lysed in 6 N HCl (1 ml) at 1008C for 17 h. After evaporation at658C, the samples were taken up in water (1 ml). A 0.1 mlvolume of solution A (1.5 N Na2CO3 in 4% acetylacetone)was added to 0.1 ml of sample. The mixture was incubatedat 1008C for 20 min, and 0.7 ml of 96% ethanol was added.One hour after adding 0.1 ml of solution B (1.6 g of p-dimethy-laminobenzaldehyde in 30 ml of concentrated HCl and 30 mlof ethanol), the absorbance at 520 nm was measured andcompared with a standard curve of 0±200 mg of glucosamine(Tracey, 1956; Popolo et al., 1997).
Acknowledgements
We thank Isaac Bom (Unilever Research Laboratories, Vlaar-dingen, The Netherlands) and Enrico Cabib (Laboratory ofBiochemistry and Genetics, NIDDKD, MD, USA) for givingsamples of pure endo-b1,6-glucanase and exochitinaserespectively. We are grateful to Susan Southard (Center forCancer Research, MIT, MA, USA) and Hisafumi Yamada-Okabe (Nippon Roche Research Center, Kamakura, Japan)for gifts of the C. albicans strains SSCA-2 and 211 respec-tively. We thank George Livi and Megan McLaughlin (SmithK-line Beecham Pharmaceuticals) for donating the anti-Alsantiserum. We also thank Joachim Ernst (Institut fuÈr Mikrobio-logie, Heinrich-Heine-UniversitaÈ t DuÈsseldorf, Germany) fordonating C. albicans strain CAP1-312 and for critical readingof the manuscript. Piet de Groot is thanked for his help withthe homology searches of S. cerevisiae PIR genes with sequ-ences of the Candida genome data bank. This work was®nancially supported by the Netherlands Technology Founda-tion (STW) and was co-ordinated by the Earth Life Sciences
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Cell wall construction of C. albicans 609

Foundation (ALW). L.L.H. was supported by Public HealthService Grant AI39441.
References
Bom, I.J., Dielbandhoesing, S.K., Harvey, K.N., Oomes,S.J.C.M., Klis, F.M., and Brul, S. (1998) A new tool forstudying the molecular architecture of the fungal cell wall:one-step puri®cation of recombinant Trichoderma b-(1,6)-glucanase expressed in Pichia pastoris. Biochim Bio-phys Acta 1425: 419±424.
Carlemalm, E., Gavavito, R.M., and Villiger, W. (1982) Resindevelopment for electron microscopy and an analysis ofembedding at low temperature. J Microscopy 126: 123±143.
Chaf®n, W.L., Lopez-Ribot, J.L., Casanova, M., Gozalbo, D.,and Martinez, J.P. (1998) Cell wall and secreted proteins ofCandida albicans: identi®cation, function, and expression.Microbiol Mol Biol Rev 62: 130±180.
Choi, W.J., Santos, B., Duran, A., and Cabib, E. (1994) Areyeast chitin synthases regulated at the transcriptional orpost-translational level? Mol Cell Biol 14: 7685±7694.
Dallies, N., FrancËois, J., and Paquet, V. (1998) A new methodfor quantitative determination of polysaccharides in theyeast cell wall. Application to the cell-wall-defectivemutants of Saccharomyces cerevisiae. Yeast 14: 1297±1306.
Ezaki, B., Gardner, R.C., Ezaki, Y., Kondo, H., and Matsu-moto, H. (1998) Protective roles of two aluminium (Al)-induced genes, HSP150 and SED1 of Saccharomycescerevisiae, in Al and oxidative stresses. FEMS MicrobiolLett 159: 99±105.
Fleet, G.H. (1991) Cell walls. In The Yeasts, vol. 4. Rose,A.H., and Harrison, J.S. (eds). London: Academic Press,pp. 199±277.
Fu, Y., Rieg, G., Fonzi, W.A., Belanger, P.H., Edwards, J.E.,and Filler, S.G. (1998) Expression of the Candida albicansgene ALS1 in Saccharomyces cerevisiae induces adher-ence to endothelial and epithelial cells. Infect Immun 66:1783±1786.
Fujii, T., Shimoi, H., and Iimura, Y. (1999) Structure of theglucan-binding sugar chain of Tip1p, a cell wall protein ofSaccharomyces cerevisiae. Biochim Biophys Acta 1427:133±144.
Gaughran, J.P., Lai, M.H., Kirsch, D.R., and Silverman, S.J.(1994) Nikkomycin Z is a speci®c inhibitor of Saccharo-myces cerevisiae chitin synthase isozyme Chs3 in vitroand in vivo. J Bacteriol 176: 5857±5860.
Gaur, N.K., and Klotz, S.A. (1997) Expression, cloning, andcharacterization of a Candida albicans gene, ALA1, thatconfers adherence properties upon Saccharomyces cere-visiae for extracellular matrix proteins. Infect Immun 65:5289±5294.
Gentzsch, M., and Tanner, W. (1996) The PMT gene family:protein O-glycosylation in Saccharomyces cerevisiae isvital. EMBO J 15: 5752±5759.
Hohenberg, H., Mannweiter, K., and MuÈller, M. (1994) High-pressure freezing of cell suspensions in cellulose capillarytubes. J Microscopy 175: 34±43.
Hoyer, L.L., Scherer, S., Shatzman, A.R., and Livi, G.P.(1995) Candida albicans ALS1: domains related to a
Saccharomyces cerevisiae sexual agglutinin separatedby a repeating motif. Mol Microbiol 15: 39±54.
Hoyer, L.L., Payne, T.L., Bell, M., Myers, A.M., and Scherer,S. (1998a) Candida albicans ALS3 and insights into thenature of the ALS gene family. Curr Genet 33: 451±459.
Hoyer, L.L., Payne, T.L., and Hecht, J.E. (1998b) Identi®ca-tion of Candida albicans ALS2 and ALS4 and localizationof Als proteins to the fungal cell surface. J Bacteriol 180:5334±5343.
Hoyer, L.L., Clevenger, J., Hecht, J.E., Ehrhart, E.J., andPoulet, F.M. (1999) Detection of Als proteins on the cellwall of Candida albicans in murine tissues. Inf Immun67: 4251±4255.
Jiang, B., Sheraton, J., Ram, A.F.J., Dijkgraaf, G.J.P., Klis,F.M., and Bussey, H. (1996) CWH41 encodes a novelendoplasmic reticulum membrane N-glycoprotein involvedin b1,6-glucan assembly. J Bacteriol 178: 1162±1171.
Kapteyn, J.C., Montijn, R.C., Dijkgraaf, G.J.P., Van DenEnde, H., and Klis, F.M. (1995) Covalent association ofb1,3-glucan with b1,6±glucosylated mannoproteins in cellwalls of Candida albicans. J Bacteriol 177: 3788±3792.
Kapteyn, J.C., Montijn, R.C., De Vink, E., La Cruz, J., Llobell,A., Douwes, J.E., et al. (1996) Retention of Saccharomycescerevisiae cell wall proteins through a phosphodiester-linkedb1,3-/b1,6±glucan heteropolymer. Glycobiology 6: 337±345.
Kapteyn, J.C., Ram, A.F.J., Groos, E.M., KollaÂr, R., Montijn,R.C., Van Den Ende, H., et al. (1997) Altered extent ofcross-linking of b1,6-glucosylated mannoproteins to chitinin Saccharomyces cerevisiae mutants with reduced cellwall b1,3-glucan content. J Bacteriol 179: 6279±6284.
Kapteyn, J.C., Van Egmond, P., Sievi, E., Van Den Ende, H.,Makarow, M., and Klis, F.M. (1999a) The contribution ofthe O-glycosylated protein Pir2p/Hsp150 to the construc-tion of the yeast cell wall in wild-type cells and b1,6-glu-can-de®cient mutants. Mol Microbiol 31: 1835±1844.
Kapteyn, J.C., Van Den Ende, H., and Klis, F.M. (1999b) Thecontribution of cell wall proteins to the organization of theyeast cell wall. Biochim Biophys Acta 1426: 373±383.
Ketela, T., Green, R., and Bussey, H. (1999) Saccharomycescerevisiae Mid2p is a potential cell wall stress sensor andupstream activator of the PKC1-MPK1 cell integrity path-way. J Bacteriol 181: 3330±3340.
Klis, F.M., Caro, L.H.P., Vossen, J.H., Kapteyn, J.C., Ram,A.F.J., Montijn, R.C., et al. (1997) Identi®cation and char-acterization of a major building block in the cell wall of Sac-charomyces cerevisiae. Biochem Soc Trans 25: 856±860.
Klis, F.M., Ram, A.F.J., Montijn, R.C., Kapteyn, J.C., Caro,L.H.P., Vossen, J.H., et al. (1998) Posttranslational modi®ca-tions of secretory proteins. Methods Microbiol 26: 223±238.
KollaÂr, R., PetraÂkovaÂ, E., Ashwell, G., Robbins, P.W., andCabib, E. (1995) Architecture of the yeast cell wall: the link-age between chitin and b (1,3)-glucan. J Biol Chem 270:1170±1178.
KollaÂr, R., Reinhold, B.B., PetraÂkovaÂ, E., Yeh, H.J.C., Ash-well, G., Drgonova , J., et al. (1997) Architecture of theyeast cell wall: b(1,6)-glucan interconnects mannoproteins,b(1,3)-glucan, and chitin. J Biol Chem 272: 17762±17775.
Laemmli, U.K. (1970) Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature227: 680±685.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
610 J. C. Kapteyn et al.

Lipke, P.N., and Ovalle, R. (1998) Cell wall architecture inyeast: new structure and new challenges. J Bacteriol180: 3735±3740.
Lu, C.F., Montijn, R.C., Brown, J.L., Klis, F.M., Kurjan, J.,Bussey, H., and Lipke, P.N. (1995) Glycosyl phosphatidyli-nositol-dependent cross-linking of a-agglutinin and b1,6-glucan in the Saccharomyces cerevisiae cell wall. J CellBiol 128: 333±340.
Lussier, M., White, A.-M., Sheraton, J., Di Paolo, T., Tread-well, J., Southard, S.B., et al. (1997) Large scale identi®ca-tion of genes involved in cell surface biosynthesis andarchitecture in Saccharomyces cerevisiae. Genetics 147:435±450.
Mio, T., Yamada-Okabe, T., Yabe, T., Nakajima, T., Arisawa,M., and Yamada-Okabe, H. (1997) Isolation of the Candidaalbicans homologs of Saccharomyces cerevisiae KRE6and SKN1 : expression and physiological function. J Bac-teriol 179: 2363±2372.
Montijn, R.C., Van Rinsum, J., Van Schagen, F.A., and Klis,F.M. (1994) Glucomannoproteins in the cell wall of Sac-charomyces cerevisiae contain a novel type of carbohy-drate side chain. J Biol Chem 269: 19338±19342.
Mormeneo, S., Marcilla, A., Iranzo, M., and Sentandreu, R.(1994) Structural mannoproteins released by b-eliminationfrom Candida albicans cell walls. FEMS Microbiol Lett 123:131±136.
Mrsa, V., and Tanner, W. (1999) Role of NaOH-extractablecell wall proteins Ccw5p, Ccw6p, Ccw7p and Ccw8p(members of the Pir protein family) in stability of the Sac-charomyces cerevisiae cell wall. Yeast 15: 813±820.
Mrsa, V., Seidl, T., Gentzsch, M., and Tanner, W. (1997)Speci®c labelling of cell wall proteins by biotinylation. Iden-ti®cation of four covalently linked O-mannosylated proteinsof Saccharomyces cerevisiae. Yeast 13: 1145±1154.
Nagahashi, S., Lussier, M., and Bussey, H. (1998) Isolationof Candida glabrata homologs of the Saccharomyces cere-visiae KRE9 and KNH1 genes and their involvement in cellwall b-1,6-glucan synthesis. J Bacteriol 180: 5020±5029.
Popolo, L., and Vai, M. (1998) Defects in assembly of theextracellular matrix are responsible for altered morphogen-esis of a Candida albicans phr1 mutant. J Bacteriol 180:163±166.
Popolo, L., Gilardelli, D., Bonfante, P., and Vai, M. (1997)Increase in chitin as an essential response to defects inassembly of cell wall polymers in the ggp1D mutant of Sac-charomyces cerevisiae. J Bacteriol 179: 463±469.
Ram, A.F.J., Wolters, A., Ten Hoopen, R., and Klis, F.M.(1994) A new approach for isolating cell wall mutants in
Saccharomyces cerevisiae by screening for hypersensitiv-ity to Calco¯uor White. Yeast 10: 1019±1030.
Ram, A.F.J., Kapteyn, J.C., Montijn, R.C., Caro, L.H.P.,Douwes, J.E., Baginsky, W., et al. (1998) Loss of theplasma membrane-bound protein Gas1p in Saccharo-myces cerevisiae results in the release of b1,3-glucaninto the medium and induces a compensation mechanismto ensure cell wall integrity. J Bacteriol 180: 1418±1424.
Russo, P., Kalkinen, N., Sareneva, H., Paakkola, J., andMakarow, M. (1992) A heat shock gene from Saccharo-myces cerevisiae encoding a secretory protein. Proc NatlAcad Sci USA 89: 3671±3675.
Russo, P., Simonen, M., Uimari, A., Teesalu, T., andMakarow, M. (1993) Dual regulation by heat and nutrientstress of the yeast HSP150 gene encoding a secretory gly-coprotein. Mol Gen Genet 239: 273±280.
Shepherd, M.G. (1987) Cell envelope of Candida albicans.Crit Rev Microbiol 15: 7±25.
Sitte, H., Neuman, K., and Edelman, L. (1985) Cryo-®xationand cryo-substitution for routine work in transmission elec-tron microscopy. In Science of Biological Specimen Pre-paration. MuÈller, M., Becker, R.P., Boyde, A., andWolosewick, J.J. (eds). Chicago, IL: SEM Inc. AMFO'Hare, pp. 103±118.
Smits, G.J., Kapteyn, J.C., Van Den Ende, H., and Klis, F.M.(1999) Cell wall dynamics in yeast. Curr Opin Microbiol 2:348±352.
Staab, J.F., Bradway, S.D., Fidel, P.L., and Sundstrom, P.(1999) Adhesive and mammalian transglutaminase sub-strate properties of Candida albicans Hwp1. Science283: 1535±1538.
Timpel, C., Strahl-Bolsinger, S., Ziegelbauer, K., and Ernst,J.F. (1998) Multiple functions of Pmt1p-mediated proteinO-mannosylation in the fungal pathogen Candida albicans.J Biol Chem 273: 20837±20846.
Toh E.A., Yasunaga, S., Nisogi, H., Tanaka, K., Oguchi, T.,and Matsui, Y. (1993) Three yeast genes, PIR1, PIR2,and PIR3, containing internal tandem repeats, are relatedto each other, and PIR1 and PIR2 are required for toler-ance to heat shock. Yeast 9: 481±494.
Tracey, M.V. (1956) Chitin. In Modern Methods of Plant Ana-lysis, vol. II. Peach, P., and Tracey, M.V. (eds). Berlin:Springer-Verlag, pp. 264±274.
Yun, D.J., Zhao, Y., Pardo, J.M., Narasimhan, M.L.,Damsz, B., Lee, H., et al. (1997) Stress proteins on theyeast cell surface determine resistance to osmotin, aplant antifungal protein. Proc Natl Acad Sci USA 94:7082±7087.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 601±611
Cell wall construction of C. albicans 611
Top Related
Copyright © 2022 FDOKUMEN

