







Bahasa
Halaman
Hukum


doi:10.1182/blood-2004-02-0474Prepublished online August 12, 2004;
and Maria GranoDavide Del Prete, Roberta Faccio, Arcangelo Liso, Silvana Capalbo, Vincenzo Liso, Alberta Zallone Silvia Colucci, Giacomina Brunetti, Rita Rizzi, Antonia Zonno, Giorgio Mori, Graziana Colaianni, interactionhuman multiple myeloma bone disease: the role of the OPG/Trail T cells support osteoclastogenesis in an in vitro model derived from
(1930 articles)Signal Transduction � (4217 articles)Neoplasia �
(5020 articles)Immunobiology � (746 articles)Apoptosis �
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
articles must include the digital object identifier (DOIs) and date of initial publication. priority; they are indexed by PubMed from initial publication. Citations to Advance online prior to final publication). Advance online articles are citable and establish publicationyet appeared in the paper journal (edited, typeset versions may be posted when available Advance online articles have been peer reviewed and accepted for publication but have not
Copyright 2011 by The American Society of Hematology; all rights reserved.Washington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

1
T CELLS SUPPORT OSTEOCLASTOGENESIS IN AN IN VITRO MODEL
DERIVED FROM HUMAN MULTIPLE MYELOMA BONE DISEASE: THE
ROLE OF THE OPG/TRAIL INTERACTION
T CELLS AND OSTEOCLASTOGENESIS IN MULTIPLE MYELOMA
Silvia Colucci, Giacomina Brunetti, Rita Rizzi*, Antonia Zonno, Giorgio Mori, Graziana Colaianni,
Davide Del Prete, Roberta Faccio, Arcangelo Liso°, Silvana Capalbo*, Vincenzo Liso*, Alberta
Zallone and Maria Grano.
Department of Human Anatomy and Histology, *Hematology Section, Department of Internal
Medicine and Public Medicine, University of Bari Medical School, Bari, Italy, °Hematology
Section, University of Foggia Medical School, Foggia, Italy.
Supported by Ministero dell’Istruzione Università e Ricerca COFIN PRIN 2001, AIRC (Italian
Foundation for Cancer Research) and Italian Space Agency (ASI), MIUR-CNR and AIL-Bari.
Corresponding author:Maria GranoDepartment of Human Anatomy and HistologyUniversity of Bari Medical SchoolPiazza Giulio Cesare, 11 70124 Bari, ITALY Phone: +39-080-5478306Fax: +39-080-5478308e-mail: [email protected]
ABSTRACT WORD COUNT: 200
TEXT WORD COUNT: 4991
SCIENTIFIC HEADING: Neoplasia
Blood First Edition Paper, prepublished online August 12, 2004; DOI 10.1182/blood-2004-02-0474
Copyright © 2004 American Society of Hematology
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

2
ABSTRACT
The development of multiple myeloma (MM) bone disease is mediated by increased number and
activity of osteoclasts (OCs). Using an in vitro osteoclastogenesis model consisting of unstimulated
and unfractionated peripheral blood mononuclear cells (PBMCs) from MM patients, we showed
that T cells support the formation of OCs with longer survival. Differently in T cell-depleted MM
PBMC cultures, exogenous macrophage-colony stimulating factor (M-CSF) and receptor activator
of nuclear factor-kappa B ligand (RANKL) were necessary to the formation of OCs, not exhibiting
however longer survival. We found up-regulated production of RANKL, osteoprotegerin (OPG)
and TNF-related apoptosis inducing ligand (TRAIL) by fresh MM T cells. Despite high OPG
levels, the persistence of osteoclastogenesis can be related to the formation of the OPG/TRAIL
complex demonstrated by immunoprecipitation experiments, and the addition of anti-TRAIL
antibody which decreases OC formation. OCs overexpressed TRAIL decoy receptor DcR2 in the
presence of MM T cells, and death receptor DR4 in T cell-depleted cultures. In addition, increased
Bcl-2/Bax ratio, following Bcl-2 upregulation, was detected in OCs generated in the presence of T
cells. Our results highlight that MM T cells support OC formation and survival, possibly involving
OPG/TRAIL interaction and unbalanced OC expression of TRAIL death and decoy receptors.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

3
INTRODUCTION
Multiple myeloma (MM) is a B cell neoplasm characterized by clonal expansion of malignant
plasma cells in the bone marrow (BM) with frequent occurrence of lytic bone disease; in about a
third of cases, MM follows the monoclonal gammapathy of undetermined significance (MGUS).1
MM lytic bone disease, causing a high morbidity, is observed in the majority of patients. It results
from an unbalanced bone turnover with enhanced resorption related to increased osteoclast (OC)
recruitment and activity and low bone formation. The interaction between MM cells and BM
microenvironment is essential for maintenance and progression of the disease process. In particular,
the reciprocal relationship between MM cells and OCs is known to be critical for the induction of
osteoclastogenesis and the activation of bone resorption1,2 as well as for the inhibition of
osteoblasts, thus preventing lesion repair.3 The linkage between immunoregulation by T cells and
bone loss is becoming more evident in MM and other bone loss-associated diseases.4-11
Osteoclastogenesis is positively or negatively regulated by a complex signaling system, that
involves the receptor activator of nuclear factor (NF)-kB (RANK), osteoprotegerin (OPG) and
receptor activator of nuclear factor (NF)-kB ligand (RANKL), all belonging to the tumor necrosis
factor (TNF) family.12 The osteoclastogenic factor RANKL is expressed by osteoblasts and stromal
cells as a membrane-bound protein, cleaved into a soluble molecule (sRANKL) by
metalloproteinases;13-15 it is also secreted by activated T cells in a primary soluble form, being a
crucial paracrine link between bone metabolism and immune system.16,17 RANKL promotes
differentiation and fusion of OC precursor cells (OCPs), and activates mature OCs to bone
resorption by binding to its specific receptor RANK.17 Early stage OCPs express c-fms, that is the
cellular receptor for macrophage-colony stimulating factor (M-CSF) necessary for OC
development.18 Further, TNF-α synergizes with RANKL and potentiates osteoclastogenesis.19
OPG, a soluble decoy receptor secreted by osteoblasts and BM stromal cells, competes with RANK
for binding to RANKL, preventing its osteoclastogenic effect.13-15,20-24 In addition, OPG can act as a
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

4
decoy receptor for TNF-related apoptosis-inducing ligand (TRAIL), exerting an anti-apoptotic
effect. 25-28
TRAIL is a cytotoxic protein inducing apoptosis mostly in tumor cells, upon binding to death-
domain containing receptors DR4 and DR5. It can activate the apoptotic pathway of MM cells by
modulating the Bcl-2 family proteins;29 among these, the ratio between Bcl-2 and Bax, anti- and
pro-apoptotic molecules respectively, is of major relevance for cell survival.30,31 TRAIL activity can
be modulated by association with two membrane-bound decoy receptors, namely DcR1 and DcR2,
which lack of functional death domains and conferring TRAIL resistance on expressing cells.
Furthermore, it has recently been shown that TRAIL may regulate OPG activity through the
OPG/TRAIL interaction, which can induce cross-regulatory mechanisms between the two
molecules.25
OC biology appears to be dominantly regulated by the RANK/RANKL/OPG axis. In particular, the
RANK/RANKL interaction is implicated in induction of osteoclastogenesis and activation of bone
resorption. The binding of soluble OPG to RANKL, by interference with the RANK/RANKL
interaction, inhibits OC differentiation and activation neutralizing the bone resorption effect. On the
other hand, OPG binding to TRAIL can be followed by both inhibition of OPG anti-
osteoclastogenic activity and blocking of TRAIL-induced apoptosis.25,32,33 A disruption of
RANKL/OPG ratio, following up-regulation of RANKL and down-regulation of OPG expression,
has been reported in MM.34,35 More recently, Giuliani et al. have suggested that T cells can
contribute to MM-induced osteoclastogenesis through up-regulated RANKL and down-regulated
IFN-γ secretion.11
On the basis of the novel paradigm for T cells as regulators of bone turnover,5 the primary aim of
our study was to investigate the potential involvement of MM T cells in osteoclastogenesis using in
vitro models consisting of unfractionated peripheral blood mononuclear cells (PBMCs) and bone
marrow mononuclear cells (BM MNCs) derived from the MM patients and the controls,
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

5
respectively; parallel T cell-depleted PBMC and BM MNC cultures derived from patients with MM
were also established. Our secondary aim was to assess the expression of the major mediators of
osteoclastogenesis in our culture system. The results showed that MM T cells support the formation
of OCs and their longer survival via RANKL, OPG and TRAIL.
PATIENTS, MATERIALS AND METHODS
Patients
The samples included peripheral blood (PB) and BM aspirate from 32 patients diagnosed as having
MM. The controls included PB and BM aspirate from 32 subjects with nonneoplastic disease
without any skeletal involvement, matched for age and sex. Informed consent was obtained in
keeping with institutional policies. The diagnosis and the stage of MM were established according
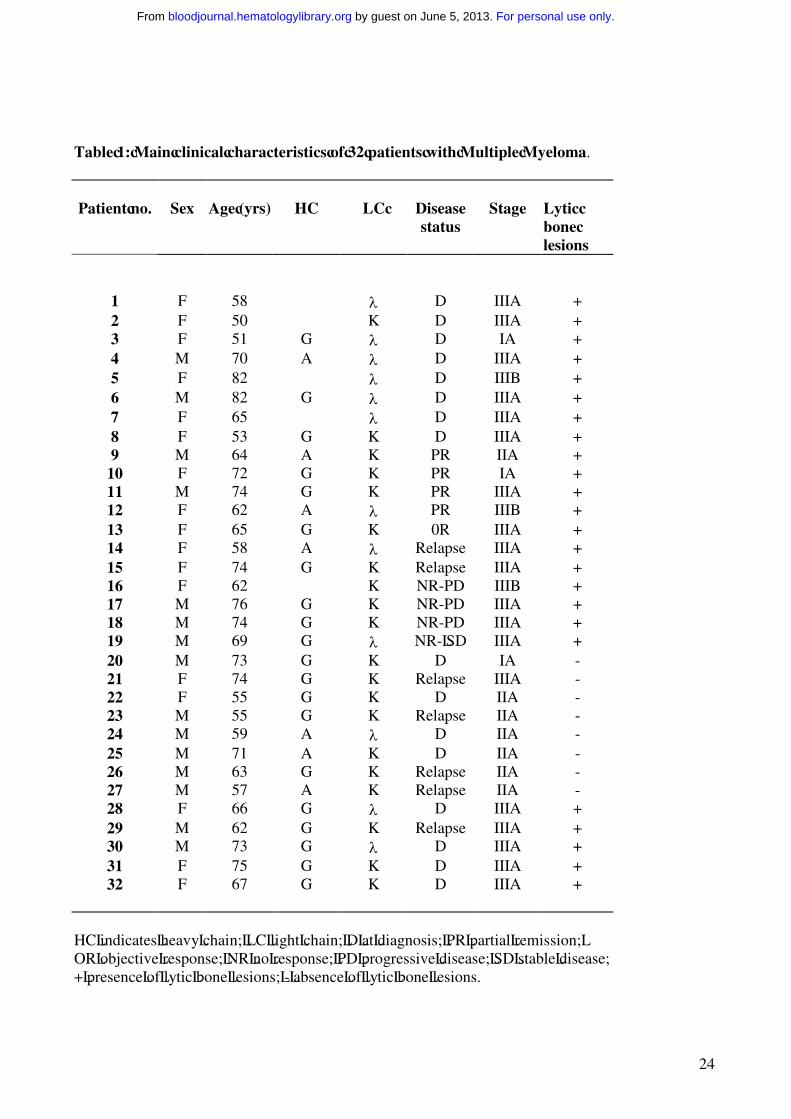
to the South West Oncology Study Group (SWOG) criteria.36,37 The main clinical characteristics of
the patients (15 males and 17 females), aged from 50 to 82 (median 65.5 ± 8.7), are listed in Table
1. Three of them had stage IA, 7 stage IIA and 21 stage III (A in 19 and B in the other 2). The M-
component was IgA in 7, IgG in 20 and only light chains in 5 patients. The light chain type was k in
20 and λ in 12 patients. Sixteen patients were included in the study at diagnosis, and the other 16 at
relapse or at progression of their disease. The latter had already undergone treatment for MM, but
they did not receive chemotherapy in the 6 months prior their study entry.
The patients were divided in two subgroups according to the presence (group a) or the absence
(group b) of osteolysis. This was documented by skeleton standard radiography and in some cases
also by nuclear magnetic resonance (NMR).
Cell cultures
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

6
OCs were obtained from unfractionated PBMCs and BM MNCs of the MM patients and the
controls as well as from T cell-depleted PBMCs and BM MNCs of the MM patients. PBMCs and
BM MNCs were isolated by centrifugation over Histopaque 1077 density gradient (Sigma Chemical
Co., St. Louis, MO, USA), diluted at 1x106cells/ml in α-Minimal Essential Medium (α-MEM) and
supplemented with 10% Fetal Bovine Serum (FBS), 100 IU/ml penicillin and 100 µg/ml
streptomycin (Gibco Limited, Uxbridge, UK). To obtain fully differentiated human OCs, the
PBMCs and BM MNCs were then cultured for about 30 days in the presence or absence of 25
ng/ml recombinant human Macrophage Colony Stimulating Factor (rh-MCSF) and increasing
amounts of RANKL, ranging from 30 to 100 ng/ml (R&D Systems Inc MN, USA). At the end of
the culture period, mature OCs were identified as tartrate-resistant acid phosphatase positive
(TRAP+) multinucleated cells (Sigma Aldrich, Milan, Italy) containing 3 or more nuclei. Their
resorbing activity was demonstrated by plating the cells on Millennium multiwell slides
(Millennium Biologix Inc., Ontario, Canada). To visualize the pits formed by the OCs, the cells
were removed by adding NaOCl to each well. The OCs were purified from their precursors and
other cell types by trypsin treatment. The mature OCs, strongly adherent to the plastic, were further
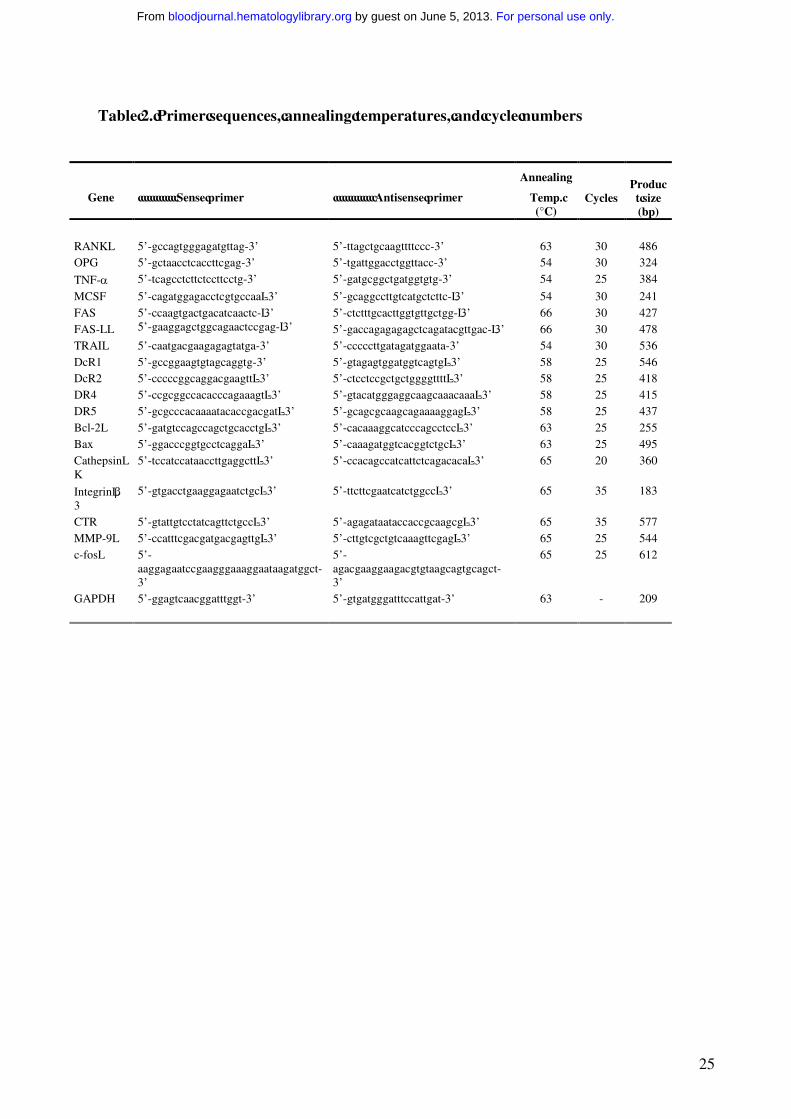
characterized by RT-PCR for the expression of specific markers such as β3 integrin, cathepsin K,
matrix metalloproteinase-9 (MMP-9), calcitonin receptor (CTR) and c-fos, utilizing the primers
reported in Table 2. The T cell depletion was performed by using anti-CD2 Ab coated
immunomagnetic Dynabeads (Dynal, Lake Success, NY, USA). Briefly, CD2+ cells were captured
from the PB or BM buffy coats, incubating 2x107 beads/ml with 5x106 cells/ml for 30 min at 4°C
on an apparatus which allows both gentle tilting and rotation. The T cell-depleted MM PBMCs and
BM MNCs were cultured with or without exogenous cytokines, in the same conditions described
above.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

7
For some experiments, PBMCs were cultured in the presence of different concentrations of RANK-
Fc (20 to 100 ng/ml) or anti-TRAIL monoclonal antibodies (mAb) (R&D Systems Inc.) (10 to 500
ng/ml) with or without RANKL at the concentration of 30 ng/ml.
Cell viability assay
Cell viability was measured by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(MTT) assay. PBMCs were cultured in 96-well tissue-culture plates in the presence or absence of T
cells. 25 ng/ml of M-CSF and 30 ng/ml of RANKL were added to the T cell-depleted cultures. At
day 25, 30 and 35, 200 µl/well of MTT 0.5 mg/ml were added, followed by 4 hour incubation at
37°C in a humidified 5% CO2 atmosphere. The reaction was stopped by the addition of 150 µl of
0.04 N HCl in absolute isopropanol. The optical density was read at 570 nm using an automatic
plate reader (550 Microplate Reader Bio-Rad Laboratories Inc., CA, USA). The results were
normalized to cells incubated under control conditions.
RNA isolation and reverse-transcriptase (RT)-PCR amplification
The OCs obtained from PBMC cultures of the MM patients and the controls were subjected to
mRNA extraction using spin columns (RNeasy, Qiagen, Hilden, Germany), according to the
manifacturer’s instructions, to detect the expression of TRAIL and its death and decoy receptors
(i.e.DR4 and DR5, DcR1 and DcR2, respectively) as well as of Bcl-2 family members (i.e.Bcl-2
and Bax). Before RNA extraction, cell cultures were trypsinized to remove early or late OC
precursors. The cells persisting in the culture following trypsin treatment were fully differentiated
OCs. RNA was also extracted from freshly prepared T cells from PBMCs and BM MNCs of MM
patients and controls to evaluate the expression of M-CSF, TNFα, RANKL, OPG and TRAIL.
Futhermore, mRNA levels of RANKL were detected in CD138 positive fresh MM cells.11 Briefly
for the first strand cDNA synthesis (SuperScript First-Strand Syntesis System for RT-PCR,
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

8
Invitrogen, Carlsbard, CA, USA), a RT mixture containing 1 µg total RNA, dNTPs, Oligo(dT), RT
buffer, MgCl2, DTT, RNaseOUT, SuperScript II RT, DEPC-treated water to final volume 100 µl
was prepared, according to the manifacturer’s instructions. Two µl of diluted cDNA were
transferred into a 50 µl PCR reaction mixture containing dNTPs, MgCl2, primers, autoclaved
distilled water, Platinum Taq DNA polymerase (Invitrogen).
Amplification reactions specific for the cDNAs of RANK-L, M-CSF, OPG, TNF-α, TRAIL, DcR1,
DcR2, DR4, DR5, Bcl-2, Bax and the housekeeping genes glyceraldehyde phosphate
dehydrogenase (GAPDH) were carried out using taq/polymerase (Invitrogen). The primers and RT-
PCR conditions are reported in Table 2. PCR products were analyzed by 1.5% agarose gel
electrophoresis containing 0.01% ethidium bromide, and the resulting bands were detected by a
light sensitive CCD video system (BioDocAnalyze, Whatman Biometra).
Western blot analysis
Proteins from OCs, developed in PBMC cultures of MM patients and controls, were solubilized
with lysis buffer [50 mmol/L Tris-HCl (pH 8.0), 150 mmol/L NaCl, 5 mmol/L
ethylenediaminetetraacetic acid, 1% NP40, and 1 mmol/L phenylmethyl sulfonyl fluoride]. Protein
determination was performed by BCA Protein assay Reagent Kit (Pierce Biotecnology, Rockford,
MN USA). 50 µg of cell proteins were subjected to SDS-PAGE gel and subsequently transferred to
nitrocellulose membranes (Hybond, Amersham Pharmacia, London, UK). The blots were probed
overnight at 4°C with: mouse anti-OPG, anti-β actin (Chemicon International Inc.), anti-TRAIL,
anti-RANKL mAb (R&D Systems Inc.); rabbit anti-DR4, anti-DR5, anti-DcR1, anti-DcR2
polyclonal Ab (Oncogene Research Product Inc); mouse anti-Bcl-2 and Bax mAb (Santa Cruz
Biotechnology). After incubation with appropriate peroxidase-conjugated secondary Ab, specific
reactions were revealed with the ECL detection kit and visualized on HyperfilmTM (Amersham
Pharmacia, UK). To detect the TRAIL-OPG complex, immunoprecipitation of PBMC fresh T cell
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

9
lysates and the media collected from PBMC cultures was performed using anti-TRAIL mAb or IgG.
Briefly, 150 µg of T cell lysate and 1 mg supernatant were incubated with 1µg/ml of anti-TRAIL or
IgG for 1 hour at 4°C; then, 20 µL of protein-G PLUS-agarose (Santa Cruz Biotechnology, Santa
Cruz, CA) were added and incubated with mixing overnight at 4°C. Pellets were collected by
centrifugation and washed 3 times with lysis buffer. After the final wash, the immunoprecipitated
material was recovered by boiling in sample loading buffer and separated on 10% SDS-PAGE gel.
ELISA assay
RANKL, OPG and TRAIL were detected in the media of unfractionated and T cell-depleted PBMC
cultures from the MM patients as well as from the controls by enzyme-linked immunosorbent assay
(ELISA), according to the manufacturer’s instructions. The ELISA kit for sRANKL (Biomedica,
GmbH, Wien, Austria) is designed to determine soluble uncomplexed human RANKL, whereas
those for OPG (Biomedica, GmbH, Wien, Austria) and for TRAIL (Biomol Research Laboratories
Inc., Plymouth Meeting, PA, USA), respectively, are designed to detect uncomplexed and
complexed molecules. The samples were diluted to the concentrations within the standard curve
range. The absorption was determined with an ELISA reader at 450 nm (550 Microplate Reader
Bio-Rad) and the results were expressed as mean ± SE.
Statistical analyses
Statistical analyses were performed by Student’s t-test with the Statistical Package for the Social
Sciences (spssx/pc) software (SPSS, Chicago, IL, USA).The results were considered statistically
significant for p<0.05.
RESULTS
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

10
Spontaneous osteoclastogenesis in unstimulated cultures of unfractionated PBMCs from MM
patients
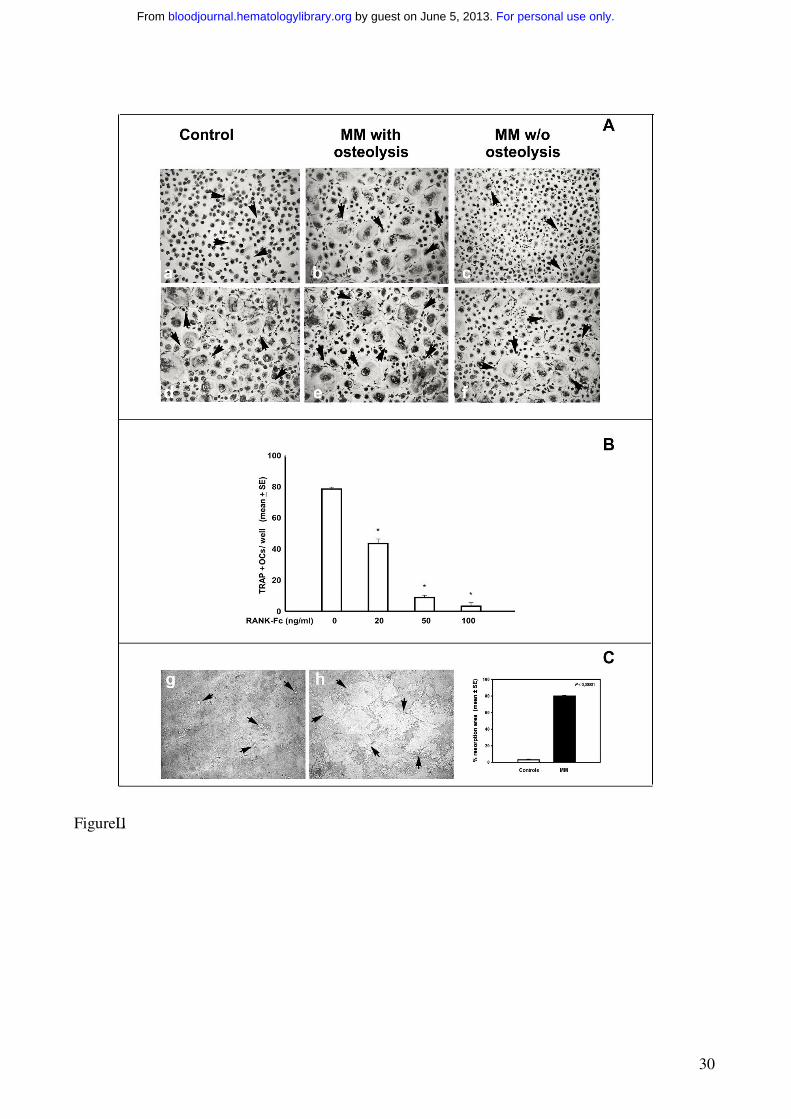
Numerous large TRAP+ OCs were identified in the unstimulated PBMC cultures from MM patients
with osteolysis (OC average number/well = 62 ± 4) (Figure 1b), whereas smaller and fewer OCs
appeared in the PBMC cultures from the MM patients without osteolysis (Figure 1c) and the
controls (Figure 1a) (OC average number/well = 6 ± 2 and 4 ± 1, respectively). Moreover, the
addition of M-CSF and RANKL to the PBMC cultures from MM patients with osteolysis did not
significantly change the OC number (OC average number/well = 70 ± 6) (Figure 1e) compared to
the parallel unstimulated cultures (Figure 1b). In particular, the addition of increasing amounts of
RANKL did not further enhance the osteoclastogenesis, thereby already maximal (data not shown).
However, the above cytokines were essential to trigger and sustain osteoclastogenesis of PBMCs
from MM patients without osteolysis (Figure 1f) and from the controls (Figure 1d) (OC average
number/well = 62 ± 3 and 58 ± 7, respectively). These results suggest the presence of
osteoclastogenic factors in the PBMC cultures derived from MM patients with osteolysis. The
experiment performed in the presence of RANK-Fc indicates RANKL as the major cytokine
involved in the spontaneous osteoclastogenesis from PBMCs of patients with MM bone disease
(Figure 1B). Moreover, the resorbing activity of the OCs obtained in unstimulated PBMC cultures
from MM patients with osteolysis was also demonstrated (Figure 1C), resulting significantly higher
than that detected in the unstimulated PBMC cultures from the controls (Figure 1h-g, respectively).
T cell-mediated osteoclastogenesis and OC survival in MM patients
The unstimulated cultures of T cell-depleted PBMCs from MM patients with osteolysis resulted in
the development of few small-sized OCs (OC average number/well = 10 ± 2) (Figure 2A). By
contrast, the addition of M-CSF and RANKL to these cultures induced the formation of numerous
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

11
large TRAP+ OCs (OC average number/well = 68 ± 5) (Figure 2B), like those observed in the
presence of T cells (Figure 1b). In the T cell-depleted and stromal cell-free BM cultures, OCs did
not form (data not shown).
By MTT assay, we found that the OCs generated in the presence of MM T cells exhibited a
significantly longer survival than those formed in MM T cell-depleted PBMCs and in the controls.
In particular, on the 30th day of culture, the viability of the OCs generated in the absence of T cells
was significantly lower than that displayed by the OCs from unfractionated MM PBMC cultures.
This result was even more evident by the 35th day of culture (Figure 3), being also confirmed by the
presence of a lower number of immunostained multinucleated TRAP+ OCs (data not shown).
Therefore, the longer survival displayed by the OCs derived from the unstimulated cultures of MM
PBMCs clearly appeared to be T cell-dependent.
Cytokine expression by fresh T cells, MM cells and culture media
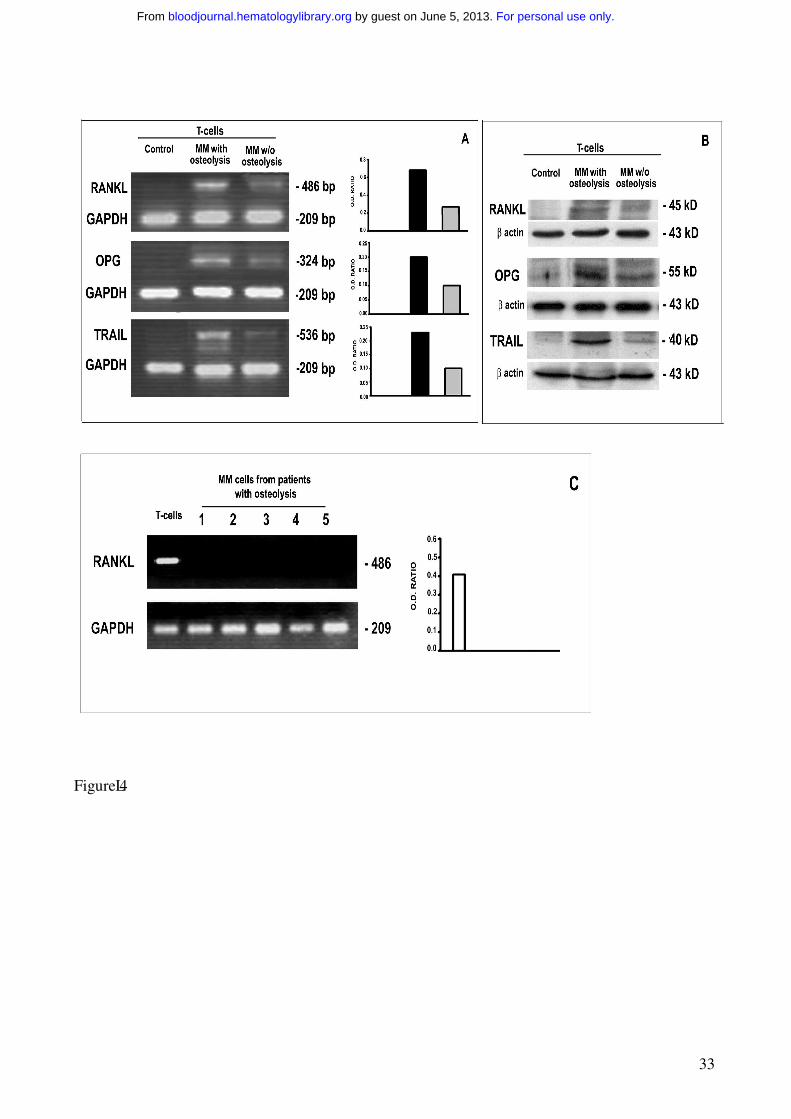
In MM patients with osteolysis, fresh T cells purified from PBMCs as well as from BM MNCs
overexpressed RANKL at mRNA and protein levels; in the fresh T cells from MM patients without
osteolysis, a very low expression of RANKL was observed, while no expression was found in the T
cells from the controls (Figure 4A-B). Furthermore, MM cells were found not expressing RANKL
at mRNA level (Figure 4C), indicating that T cells were the only source of RANKL in our system.
No difference was detected in the expression of M-CSF among MM patients with or without
osteolysis, and in the controls (data not shown). Despite the enhanced osteoclastogenesis, we found
that OPG resulted overexpressed in the fresh T cells isolated from MM patients with osteolysis;
however, it was barely detectable in those without osteolysis, and undetectable in the controls
(Figure 4 A-B). This finding could be explained by the higher levels of ambient TRAIL,
demonstrated by RT-PCR and western blotting in the fresh T cells from MM patients with
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

12
osteolysis compared to the lower levels detected in those from the patients without osteolysis, and
in the controls (Figure 4 A-B).
Based on the spontaneous osteoclastogenesis occurring in the cultures from MM patients with
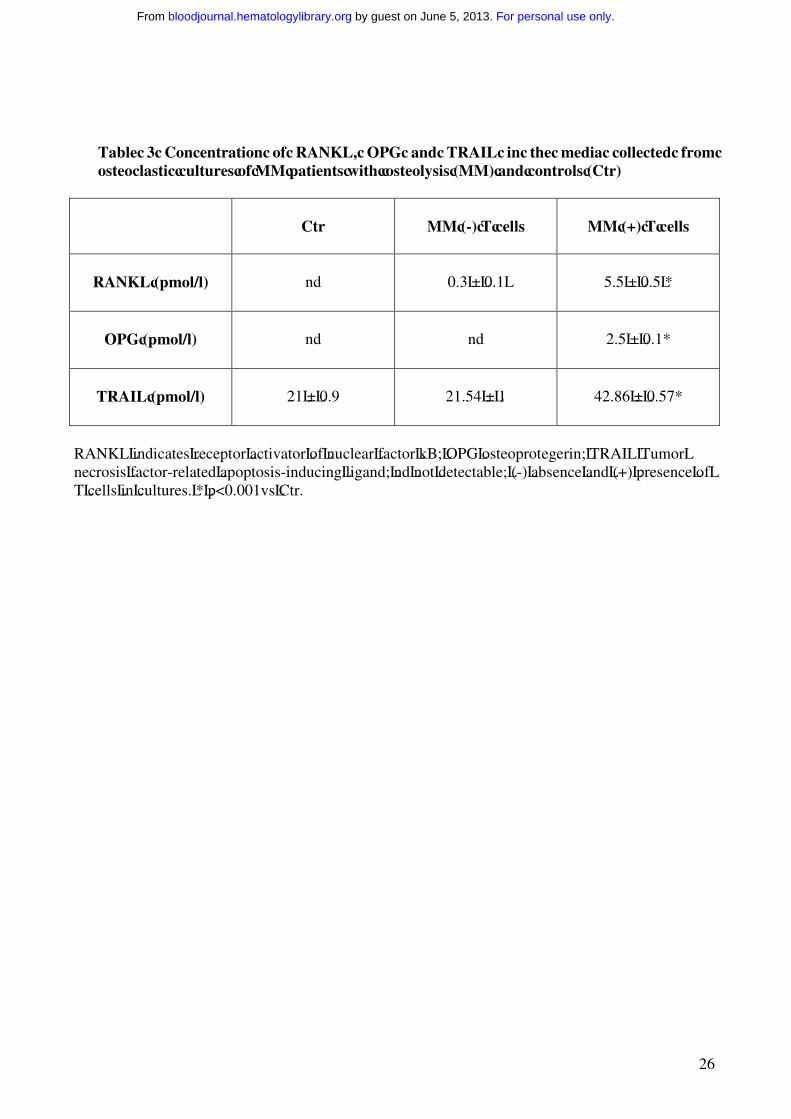
osteolysis, the expression of RANKL, OPG and TRAIL was evaluated by ELISA assay in the
media collected from unfractionated as well as T cell-depleted PBMCs of MM patients with
osteolysis, and from the controls. Detectable levels of RANKL and OPG were found only in the
media from unfractionated MM PBMC cultures, whereas these cytokines were undetectable in the
media from T cell-depleted MM PBMCs and the controls (Table 3). TRAIL levels demonstrated in
the media from unfractionated MM PBMCs were significantly higher than those found in the media
from T cell-depleted MM PBMC cultures or controls, suggesting that T cells could be the major but
not the exclusive source of this cytokine (Table 3).
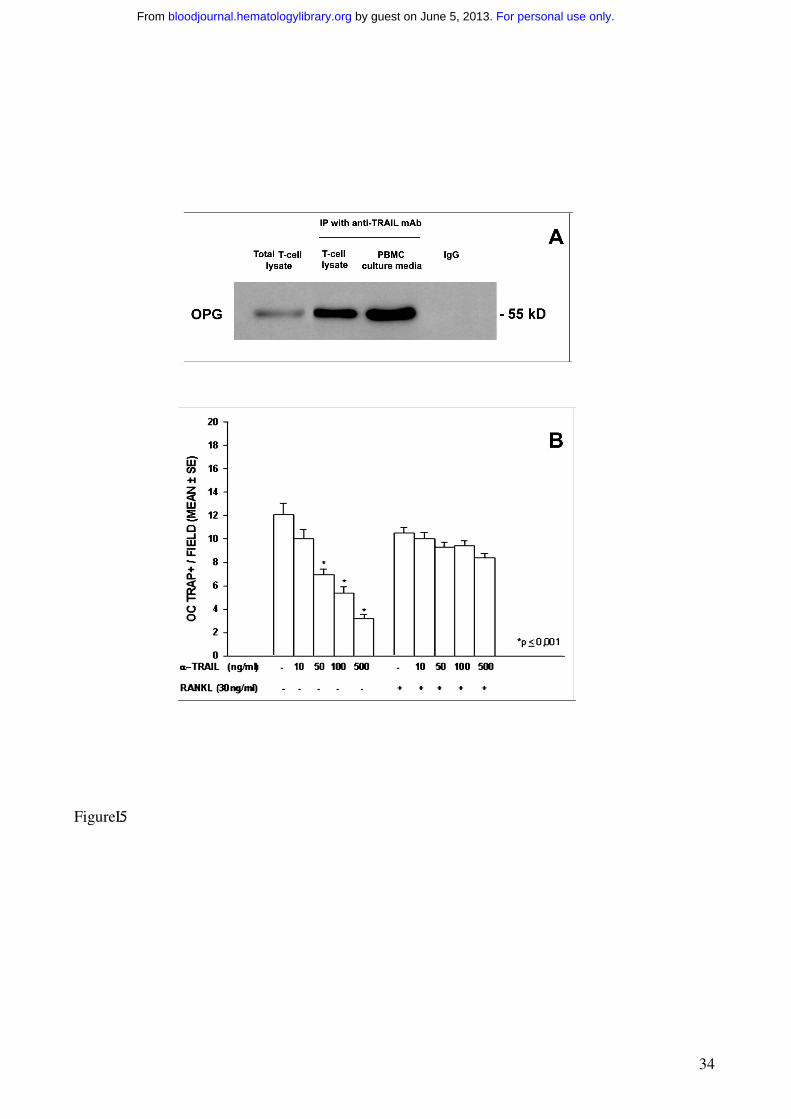
OPG/TRAIL complex in fresh T cells and culture media from PBMCs of MM patients
To assess whether TRAIL bound OPG inhibiting the anti-osteoclastogenic OPG effect, we
performed an immunoprecipitation with an anti-TRAIL mAb on PBMC fresh T cell lysates and
media collected from the PBMC cultures of MM patients with osteolysis. Our findings indicated
that TRAIL and OPG formed a complex in both T cell lysates and culture media. OPG was
identified as a band of about 55 kD, more evident in the culture media than in the T cell lysates
(Figure 5A).
In addition, the involvement of TRAIL in T cell-mediated osteoclastogenesis was further
demonstrated by the addition of different concentrations of a neutralizing anti-TRAIL mAb to the
media of PBMCs from MM patients with osteolysis. Our results showed a dose-dependent
inhibition of spontaneous osteoclastogenesis; this effect was completely abolished by the addition
of RANKL (Figure 5B).
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

13
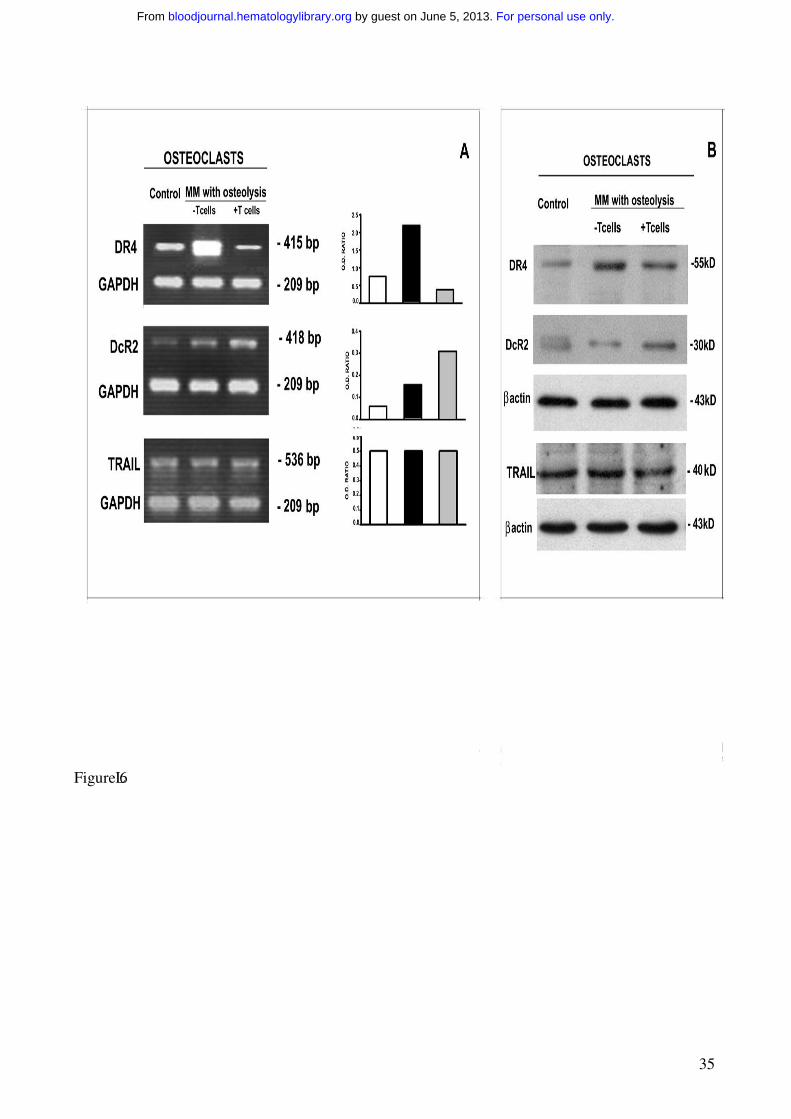
The expression of OC TRAIL receptors is under T cell-control
In the assessment of the mechanisms involved in the longer survival displayed by the OCs from
unfractionated PBMCs of MM patients with osteolysis, we evaluated the expression of death and
decoy TRAIL receptors. Despite the fact that Fas/FasL system seems to be the dominant
mechanism in OC apoptosis, we did not detect any difference in Fas receptor expression among the
OCs developed in the presence of T cells, those from T cell-depleted cultures, and the controls.
However, consistently with the literature data, the fresh T cells isolated from MM PBMCs were
found to express FasL at higher levels than those from the controls (data not shown).
Consequently, we evaluated whether T cells could regulate the expression of TRAIL receptors on
the OCs displaying longer survival. In the OCs generated from unfractionated MM PBMCs, the
analysis of TRAIL receptors demonstrated downregulation of death receptor DR4 and upregulation
of decoy receptor DcR2, at both mRNA and protein levels (Figure 6). On the contrary, DR4
resulted overexpressed and DcR2 reduced in the OCs developed from T cell-depleted cultures. In
addition, the expression of death DR5 and decoy DcR1 receptors was not modulated by T cells
(Figure 6). Thus in MM bone disease, OCs seem to be protected by TRAIL-induced apoptosis via
upregulation of its decoy receptor DcR2 and downregulation of death receptor DR4, possibly
explaining the effect of T cells on OC survival.
To support the potential regulatory role of T cells in OC survival, we evaluated the OC expression
of anti- and pro-apoptotic Bcl-2 family proteins by RT-PCR and western blotting in unfractionated
as well as T cell-depleted PBMC cultures from MM patients with osteolysis. The anti-apoptotic
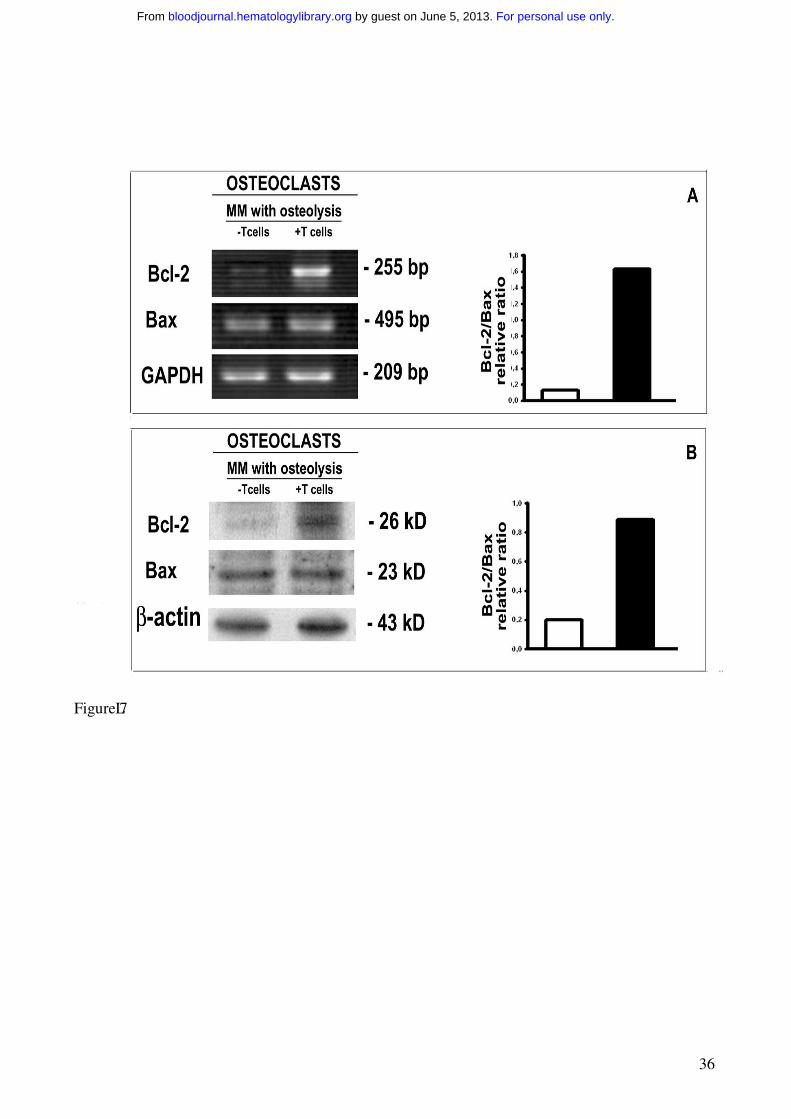
protein Bcl-2 resulted upregulated at mRNA and protein levels in the OCs generated in the presence
of T cells (Figure 7), whereas no significant change of the pro-apoptotic protein Bax was found
(Figure 7). Our data show that the T cells induce the positive modulation of Bcl-2/Bax ratio,
indicating a possible mechanism by which T cells promote the OC longer survival.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

14
DISCUSSION
A regulatory role of bone turnover played by T cells in physiological and pathological conditions is
emerging from the recent literature. In particular, T cell involvement in OC activation through
RANKL overexpression has been reported in osteoporosis,38 periodontal disease,39 adjuvant
arthritis,5 rheumatoid arthritis,40 adult T-cell leukaemia with hypercalcemia,10 and MM bone
disease.11
In the assessment of biologic mechanisms involved in the pathogenesis of MM bone disease,
Giuliani et al. had shown that myeloma cells induce an OPG/RANKL imbalance, playing a critical
role in this condition .35 Later, the same investigators suggested a possible involvement of T cells in
the OC activation through the cross-talk between RANKL and interferon-γ (IFN-γ); in particular,
they demonstrated the MM cell-induced up-regulation of RANKL and down-regulation of IFN-γ,
which is known to be a strong suppressor of osteoclastogenesis.11 Moreover, Giuliani et al.
indicated the possible implication of IL-7 in osteoclastogenesis through its stimulatory effect on the
RANKL production by T cells, and suggested a vicious loop between IL-6 and IL-7 involving the
MM cells; they detected high levels of IL-7 in the supernatants of human MM cell lines (HMLC) as
well as in BM plasma cells and sera of MM bone disease patients. 11
In line with the primary aim of our study, we used a culture system for OC development as
paradigm of in vivo bone resorption to assess the mechanism by which T cells could support the
increase in number and activity of OCs, occurring in MM bone disease. In particular, our in vitro
PBMC osteoclastogenesis model represents a good system to study the effect of T cells on
osteoclastogenesis due to the absence of stromal cells, possible source of osteoclastogenic
cytokines. In unfractionated PBMC cultures from MM bone disease patients, we observed the
formation of numerous, mature, multinucleated and bone resorbing OCs with a longer survival, that
were not evident in the same type of cultures derived from the MM patients without lytic bone
lesions and the controls. Major evidences supporting a T cell-regulation of osteoclastogenesis came
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

15
from the parallel MM T cell-depleted PBMC cultures, where the addition of exogenous M-CSF and
RANKL was necessary to the OC formation, suggesting that T cells alone could provide the above
cytokines in our system. Moreover, OCs obtained in T cell-depleted PBMC cultures were not
protected from apoptosis, indicating that T cells could also have a role in promoting the longer OC
survival. Thus for the first time, our findings demonstrate T cell-dependent osteoclastogenesis from
human MM PBMCs, and OCs exhibiting a longer survival only in the presence of MM T cells.
Based on the knowledge that OCs form normally in BM, we demonstrated a T cell-regulation of
osteoclastogenesis in the BM cultures from MM patients, whereas in the T cell-depleted and
stromal cell-free system OCs did not form. These results are consistent with the recent literature
data, showing the contribution of MM BM T cells to MM induced osteolysis through RANKL
overexpression.11
According to our secondary aim, we evaluated the cytokines possibly involved in OC formation and
survival, demonstrating that RANKL, OPG and TRAIL were overexpressed by MM PB fresh T
cells at both mRNA and protein levels and were also detectable in the media collected from
unfractionated PBMC cultures. Interestingly, the levels of these cytokines were higher in PB fresh T
cells from patients with MM bone disease compared to those from patients without clinical
evidence of lytic bone lesions, and resulted undetectable in the controls. The above findings show
that MM T cells could be the only source of these molecules in our system except for TRAIL,
whose significantly higher release in the media from MM PBMC cultures shows that it is produced
by both T cells and OCs, as we demonstrated by RT-PCR and western blot analysis of OC lysates.
A confirmation of RANKL production by MM T cells came from the exogenous RANKL necessary
to obtain mature OCs not displaying, however, longer survival in the absence of T cells.
Furthermore in our system, the involvement of RANKL as major osteoclastogenic cytokine is
confirmed by the inhibitory effect exerted by RANK-Fc on spontaneous osteoclastogenesis in
unstimulated PBMC cultures from the MM bone disease patients.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

16
The OPG overexpression we detected by human MM T cells had been previously reported only in
murine T cells.7 Based on RANKL and OPG effects on bone remodeling, consisting of RANKL
stimulation and OPG inhibition of osteoclastogenesis, we can argue that the persistence of
osteoclastogenesis despite the high levels of OPG in our system could be explained by the OPG
binding to TRAIL, blocking OPG anti-osteoclastogenic activity. Although the affinity of OPG for
RANKL is higher than that for TRAIL, the OPG/TRAIL interaction could be favoured by the
elevated TRAIL levels detected in our culture media. This possibility is supported by the detection
of OPG/TRAIL complex in T cell lysates as well as the culture media of PBMCs from MM bone
disease patients, that we demonstrated by immunoprecipitation with an anti-TRAIL mAb.
Moreover, the neutralizing anti-TRAIL Ab caused a dose-dependent inhibition of spontaneous
osteoclastogenesis, that recurred after the addition of exogenous RANKL. This findings are
consistent with the literature data, showing that TRAIL blocks the anti-osteoclastogenic effect of
OPG in mouse.25 We cannot exclude that the OPG/TRAIL interaction also blocks the apoptotosis-
inducing TRAIL activity. Moreover the overexpression of OPG, resulting in a protective cell
strategy against apoptosis in the presence of high TRAIL levels, is of major relevance as indicated
by in vitro studies prospecting OPG as a survival factor for human prostate cancer cells32 and
human MM cells.33 The high levels of OPG detected in our system were consistent with some
literature reports on bone loss-associated diseases such as postmenopausal osteoporosis,41
seropositive rheumatoid arthritis42 and prostate cancer bone metastases.32 Otherwise, low OPG
levels in BM microenvironment as well as in BM plasma and sera were detected in patients with
MM bone disease.34,35,43-45 These findings were explained with MM cell-induced impairment of
OPG production by osteoblast/stromal cells. 34,35 Later on, a further mechanism consisting of
binding, internalization and/or degradation of OPG by MM cells was provided, as supported by
both in vitro uptake of I-OPG and demonstration of plasma cells containing OPG in BM biopsies
from MM patients.44 The absence of MM cells in our system could explain the OPG
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

17
overexpression, that we detected in the fresh T cells purified from MM bone disease patients. In
addition, the high OPG levels that we demonstrated in the culture media of PBMCs from the same
patients could be related to the ELISA kit recognizing both free and bound forms of OPG.
The longer survival, exhibited exclusively by the OCs generated in the presence of MM T cells,
prompted a T cell-control of OC survival. In these OCs, we demonstrated up-regulated decoy
receptor DcR2 and down-regulated death receptor DR4 expression, whereas DR4 was found
overexpressed and DcR2 at low levels on the OCs from T cell-depleted cultures. Our data indicate
that T cells can regulate OC survival by modulating their surface expression of TRAIL death and
decoy receptors, whose ratio in turn results critical for OC sensitivity to TRAIL-mediated
apoptosis. This is consistent with the evidence of OC apoptosis in the T cell-depleted cultures,
where the OC DNA fragmentation following the TRAIL addition was observed (manuscript in
preparation); however, the mechanism by which T cells control OC survival is not yet defined.
Furthermore, we detected comparable Fas receptor levels in the OCs from both unfractionated and
T cell-depleted MM PBMC cultures, whereas FasL was overexpressed by MM fresh T cells,
suggesting that OC Fas receptor expression was not regulated by T cells in our system. Therefore,
we cannot exclude the possibility that Fas/FasL pathway is activated in our system leading to the
normal life-span of OCs generated from T cell-depleted cultures. Finally, the mechanism by which
T cells control OC survival seems to involve the positive modulation of Bcl-2/Bax ratio through the
up-regulation of Bcl-2, detected in the OCs generated in the presence of MM T cells. In other
systems, it has been demonstrated that the Bcl-2/Bax ratio is important to determine the fate of the
cells toward apoptosis or survival.30,31 Several mechanisms could be involved in the modification of
this Bcl-2/Bax ratio, but they are still unknown in the OCs. A possible modulator could be TRAIL,
recently described in a variety of normal cells32,46-48 and currently being investigated in our
laboratory in relation with its effect on OC apoptosis in normal conditions.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

18
In conclusion, our results highlight the T cell-regulation of OC formation and survival through
RANKL, OPG and TRAIL overexpression, and provide the basis for a potential role of the novel
OPG/TRAIL interaction in the biology of MM bone disease.
ACKNOWLEDGMENTS
We thank Prof. D. Ribatti for helpful discussions, and A. Grano for expert technical support.
REFENCENCES
1. Barillé-Nion S, Bataille R. New insights in myeloma-induced osteolysis. Leuk Lymphoma.
2003;44:1463-1467.
2. Epstein J, Yaccoby S. Consequences of interactions between the bone marrow stroma and
myeloma. Hematol. J. 2003;4:310-314.
3. Tian E, Zhan F, Walker R, Rasmussen E, Ma Y, Barlogie B, Shaughnessy D. The role of
the wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple
myeloma. N Engl J Med. 2003;349:2483-2494.
4. Kong YY, Yoshida H, Sarosi I, et al. OPGL is a key regulator of osteoclastogenesis,
lymphocyte development and lymph-node organogenesis. Nature. 1999;397:315-323.
5. Kong YY, Feige U, Sarosi I et al. Activated T cells regulate bone loss and joint destruction
in adjuvant arthritis through osteoprotegerin ligand. Nature. 1999;402:304-309.
6. Takayanagi H, Ogasawara K, Hida S, et al. T cell-mediated regulation of
osteoclastogenesis by signalling cross-talk between RANKL and IFN-gamma. Nature.
2000;408:600-605.
7. Choi Y, Woo KM, Ko SH, et al. Osteoclastogenesis is enhanced by activated B cells but
suppressed by activated CD8(+) T cells. Eur J Immunol. 2001;31:2179-2188.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

19
8. Weitzmann MN, Cenci S, Rifas L, Haug J Dispersio J, Pacifici R. T cell activation induces
human osteoclast formation via receptor activator of nuclear factor kappaB ligand-
dependent and- independent mechanisms. J Bone Miner Res. 2001;6:328-337.
9. Takayanagi H, Kim S, Taniguchi T. Signaling crosstalk between RANKL and interferons
in osteoclast differentiation. Arthritis Res. 2002;4: 227-232.
10. Nosaka K, Miyamoto T, Sakai T, Mitsuya H, Suda T, Matsuoka M. Mechanism of
hypercalcemia in adult T-cell leukemia: overexpression of receptor activator of nuclear
factor kappaB ligand on adult T-cell leukemia cells. Blood.2002;99:634-640.
11. Giuliani N, Colla S, Sala R, et al. Human myeloma cells stimulate the receptor activator of
nuclear factor-kappa B ligand (RANKL) in T lymphocytes: a potential role in multiple
myeloma bone disease. Blood. 2002;100:4615-4621.
12. Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and activation. Nature.
2003;423:337-342.
13. Yasuda H, Shima N, Nakagawa N, et al. Osteoclast differentiation factor is a ligand for
osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL.
Proc. Natl. Acad. Sci. USA. 1998;95:3597-3602.
14. Lacey DL, Timms E, Tan HL, et al. Osteoprotegerin ligand is a cytokine that regulates
osteoclast differentiation and activation. Cell. 1998;93:165-176.
15. Lum, L., Wong, B. R., Josien, R., et al. Evidence for a role of a tumor necrosis factor-
alpha (TNF-alpha)-converting enzyme-like protease in shedding of TRANCE, a TNF
family member involved in osteoclastogenesis and dendritic cell survival. J. Biol. Chem.
1999;274:13613-13618.
16. Hofbauer LC, Neubauer A, Heufelder AE. Receptor activator of nuclear factor-kappaB
ligand and osteoprotegerin: potential implications for the pathogenesis and treatment of
malignant bone diseases. Cancer. 2001;92:460-470.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

20
17. Theill LE, Boyle WJ, Penninger JM. RANK-L and RANK: T cell, bone loss, and
mammalian evolution. Ann Rev Immunol 2002;20:795-823.
18. Arai F, Miyamoto T, Ohneda O., et al. Commitment and Differentiation of osteoclast
precursor cells by the sequential expression of c-Fms and receptor activator of nuclear
factor kB (RANKL) receptors.J Exp Med 1999;190:1741-1754.
19. Fuller K, Murphy C, Kirstein B, Fox SW, Chambers TJ. TNFα potently activates
osteoclasts, through a direct action independent of and strongly synergistic with RANKL.
Endocrinology.2002;143:1108-1118.
20. Anderson, D. M., Maraskovsky, E., Billingsley, W. L., et al. A homologue of the TNF
receptor and its ligand enhance T-cell growth and dendritic-cell function. Nature.
1997;390:175-179.
21. Wong BR, Rho J, Arron J, et al. TRANCE is a novel ligand of the tumor necrosis factor
receptor family that activates c-Jun N-terminal kinase in T cells. J. Biol. Chem.
1997;272:25190-25194.
22. Simonet WS, Lacey DL, Dunstan CR, W, et al. Osteoprotegerin: a novel secreted protein
involved in the regulation of bone density.Cell. 1997;89:309-319.
23. Hsu H, Lacey DL, Dunstan CR, et al. Tumor necrosis factor receptor family member
RANK mediates osteoclast differentiation and activation induced by osteoprotegerin
ligand. Proc Natl Acad Sci U S A.1999;96:3540-3545.
24. Li J, Sarosi I, Yan XQ, et al. RANK is the intrinsic hematopoietic cell surface receptor that
controls osteoclastogenesis and regulation of bone mass and calcium metabolism. Proc
Natl Acad Sci U S A. 2000; 97:1566-1571.
25. Emery JG, McDonnell P, Burke MB, et al. Osteoprotegerin is a receptor for the cytotoxic
ligand TRAIL. J Biol Chem. 1998;273:14363-14367.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

21
26. Pan G, O’Rourke K, Chinnaiyan AM, et al. The receptor for the cytotoxic ligand TRAIL.
Science. 1997;276:111-113.
27. Pan G, Ni J, Wei Y-F, Yu G-l, Gentz R, Dixit VM. An antagonist decoy receptor and a
death domain-containing receptor for TRAIL. Science. 1997;277:815-818.
28. Sheridan JP, Marsters SA, Pitti RM, et al. Control of TRAIL-induced apoptosis by a
family of signaling and decoy receptors. Science. 1997;277:818-821.
29. Chen Q, Gong B, Mahmoud-Ahmed AS, et al. Apo2L/TRAIL and Bcl-2-related proteins
regulate type I interferon-induced apoptosis in multiple myeloma. Blood. 2001;98:2183-
2192.
30. Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo with a conserved
homolog, Bax, that accelerates programmed cell death. Cell. 1993;74:609-619.
31. Adams JM, Cory S. The Bcl-2 protein family: arbiters of cell survival. Science.
1998;281:1322-1326.
32. Holen I, Croucher PI, Hamdy FC, Eaton CL. Osteoprotegerin is a survival factor for
human prostate cancer cell. Cancer Res. 2002;62:1619-1623.
33. Shipman CM, Croucher PI. Osteoprotegerin is a soluble decoy receptor for tumor necrosis
factor-related apoptosis-inducing ligand/Apo2 ligand and can function as a paracrine
survival factor for human myeloma cells. Cancer Res. 2003;63:912-916.
34. Pearse RN, Sordillo EM, Yaccoby S, et al. Multiple myeloma disrupts the TRANCE/
osteoprotegerin cytokine axis to trigger bone destruction and promote tumor progression.
Proc Natl Acad Sci U S A. 2001;98:11581-11586.
35. Giuliani N, Bataille R, Mancini C, Lazzaretti M, Barille S. Myeloma cells induce
imbalance in the osteoprotegerin/osteoprotegerin ligand system in the human bone marrow
environment. Blood. 2001;98:3527-3533.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

22
36. Durie BGM, Salmon SE. A clinical staging system for multiple myeloma. Correlation of
measured myeloma cell mass with presenting clinical features, response to treatment and
survival. Cancer; 1975;36:842-854
37. Durie BGM, Salmon SE. Multiple myeloma, macroglobulinemia and monoclonal
gammopathies. In: Hoffbrand AV, Brain MC, Jirsh J eds. Recent advances in Hematology.
New York, NY: Churchill Livingstone; 1977:243-261.
38. Roggia C, Gao Y, Cenci S, et al. Up-regulation of TNF-producing T cells in the bone
marrow: a key mechanism by which estrogen deficiency induces bone loss in vivo. Proc
Nat Acad Sci U S A. 2001;98:13960-13965.
39. Teng YT, Nguyen H, Gao X, et al. Functional human T-cell immunity and osteoprotegerin
ligand control alveolar bone destruction in periodontal infection. J Clin Invest.
2000;106:59-67.
40. Kotake S, Udagawa N, Hakoda M, et al. Activated human T cells directly induce
osteoclastogenesis from human monocytes: possible role of T cells in bone destruction in
rheumatoid arthritis patients. Arthritis Rheum. 2001;44:1003-1012.
41. Yano K, Tsuda E, Washida N, et al. Immunological characterization of circulatin
osteoprotegerin/osteoclastogenesis inhibitory factor: increase serum concentrations in
postmenopausal women with osteoporosis. J Bone Min Res 1999;14:518-527.
42. Feuerhem AJ, Borset M, Seidel C, et al. Elevated levels of osteoprotegerin (OPG) and
hepatocyte growth factor (HGF) in rheumatoid arthritis. Scand J Rheumatol. 2001;30:229-
234.
43. Seidel C, Hjertner O, Abildgaard N, et al. Nordic Myeloma Study Group. Serum
osteoprotegerin levels are reduced in patients with multiple myeloma with lytic bone
disease. Blood. 2001;98:2269-2271.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

23
44. Standal T, Seidel C, Hjertner O, et al. Osteoprotegerin is bound, internalized, and degraded
by multiple myeloma cells. Blood. 2002;100:3002-3007.
45. Sezer O, Heider U, Zavrski I, Kuhne CA, Hofbauer LC. RANK ligand and osteoprotegerin
in myeloma bone disease. Blood. 2003;101:2094-2098.
46. Martin-Villalba A, Herr I, Jeremias I, et al. CD95 ligand (Fas-L/APO-1L) and tumor
necrosis factor-related apoptosis-inducing ligand mediate ischemia-induced apoptosis in
neurons. J Neurosci. 1999;19:3809-3817.
47. Jo M, Kim TH, Seol DW, et al. Apoptosis induced in normal human hepatocytes by tumor
necrosis factor-related apoptosis-inducing ligand. Nat Med. 2000;6:564-567.
48. Simon AK, Williams O, Mongkolsapaya J, et al. Tumor necrosis factor-related apoptosis-
inducing ligand in T cell development: sensitivity of human thymocytes. Proc Natl Acad
Sci U S A. 2001; 98: 5158-5163.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
24
Table 1: Main clinical characteristics of 32 patients with Multiple Myeloma.
Patient no. Sex Age (yrs) HC LC Diseasestatus
Stage Lytic bone lesions
1 F 58 λ D IIIA +2 F 50 K D IIIA +3 F 51 G λ D IA +4 M 70 A λ D IIIA +5 F 82 λ D IIIB +6 M 82 G λ D IIIA +7 F 65 λ D IIIA +8 F 53 G K D IIIA +9 M 64 A K PR IIA +
10 F 72 G K PR IA +11 M 74 G K PR IIIA +12 F 62 A λ PR IIIB +13 F 65 G K 0R IIIA +14 F 58 A λ Relapse IIIA +15 F 74 G K Relapse IIIA +16 F 62 K NR-PD IIIB +17 M 76 G K NR-PD IIIA +18 M 74 G K NR-PD IIIA +19 M 69 G λ NR- SD IIIA +20 M 73 G K D IA -21 F 74 G K Relapse IIIA -22 F 55 G K D IIA -23 M 55 G K Relapse IIA -24 M 59 A λ D IIA -25 M 71 A K D IIA -26 M 63 G K Relapse IIA -27 M 57 A K Relapse IIA -28 F 66 G λ D IIIA +29 M 62 G K Relapse IIIA +30 M 73 G λ D IIIA +31 F 75 G K D IIIA +32 F 67 G K D IIIA +
HC indicates heavy chain; LC light chain; D at diagnosis; PR partial remission; OR objective response; NR no response; PD progressive disease; SD stable disease;+ presence of lytic bone lesions; - absence of lytic bone lesions.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
25
Table 2. Primer sequences, annealing temperatures, and cycle numbers
Gene Sense primer Antisense primer
Annealing
Temp. (°C)
CyclesProduct size(bp)
RANKL 5’-gccagtgggagatgttag-3’ 5’-ttagctgcaagttttccc-3’ 63 30 486OPG 5’-gctaacctcaccttcgag-3’ 5’-tgattggacctggttacc-3’ 54 30 324
TNF-α 5’-tcagcctcttctccttcctg-3’ 5’-gatgcggctgatggtgtg-3’ 54 25 384
MCSF 5’-cagatggagacctcgtgccaa -3’ 5’-gcaggccttgtcatgctcttc- 3’ 54 30 241FAS 5’-ccaagtgactgacatcaactc- 3’ 5’-ctctttgcacttggtgttgctgg- 3’ 66 30 427FAS-L 5’-gaaggagctggcagaactccgag- 3’ 5’-gaccagagagagctcagatacgttgac- 3’ 66 30 478TRAIL 5’-caatgacgaagagagtatga-3’ 5’-cccccttgatagatggaata-3’ 54 30 536DcR1 5’-gccggaagtgtagcaggtg-3’ 5’-gtagagtggatggtcagtg -3’ 58 25 546DcR2 5’-cccccggcaggacgaagtt -3’ 5’-ctcctccgctgctggggtttt -3’ 58 25 418DR4 5’-ccgcggccacacccagaaagt -3’ 5’-gtacatgggaggcaagcaaacaaa -3’ 58 25 415DR5 5’-gcgcccacaaaatacaccgacgat -3’ 5’-gcagcgcaagcagaaaaggag -3’ 58 25 437Bcl-2 5’-gatgtccagccagctgcacctg -3’ 5’-cacaaaggcatcccagcctcc -3’ 63 25 255Bax 5’-ggacccggtgcctcagga -3’ 5’-caaagatggtcacggtctgc -3’ 63 25 495Cathepsin K
5’-tccatccataaccttgaggctt -3’ 5’-ccacagccatcattctcagacaca -3’ 65 20 360
Integrin β3
5’-gtgacctgaaggagaatctgc -3’ 5’-ttcttcgaatcatctggcc -3’ 65 35 183
CTR 5’-gtattgtcctatcagttctgcc -3’ 5’-agagataataccaccgcaagcg -3’ 65 35 577MMP-9 5’-ccatttcgacgatgacgagttg -3’ 5’-cttgtcgctgtcaaagttcgag -3’ 65 25 544c-fos 5’-
aaggagaatccgaagggaaaggaataagatggct-3’
5’-agacgaaggaagacgtgtaagcagtgcagct-3’
65 25 612
GAPDH 5’-ggagtcaacggatttggt-3’ 5’-gtgatgggatttccattgat-3’ 63 - 209
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
26
Table 3 Concentration of RANKL, OPG and TRAIL in the media collected from osteoclastic cultures of MM patients with osteolysis (MM) and controls (Ctr)
Ctr MM (-) T cells MM (+) T cells
RANKL (pmol/l) nd 0.3 ± 0.1 5.5 ± 0.5 *
OPG (pmol/l) nd nd 2.5 ± 0.1*
TRAIL (pmol/l) 21 ± 0.9 21.54 ± 1 42.86 ± 0.57*
RANKL indicates receptor activator of nuclear factor kB; OPG osteoprotegerin; TRAIL Tumor necrosis factor-related apoptosis-inducing ligand; nd not detectable; (-) absence and (+) presence of T cells in cultures. * p<0.001vs Ctr.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

27
FIGURE LEGENDS
Figure 1. OCs generated from human MM PBMCs.
(A) OCs were obtained from unfractionated PBMCs of MM patients with or without osteolysis (b-
e- c-f) and controls (a-d). Numerous and large-sized OCs (arrows) developed in the unstimulated
cultures from MM patients with osteolysis (b), whereas rare and small-sized OCs were observed in
the cultures from MM patients without osteolysis (c) and controls (a). No significant increase in OC
formation was observed in MM PBMCs from the patients with osteolysis by exogenous M-CSF and
RANKL (e), whereas these cytokines were essential to trigger the OC formation in MM patients
without osteolysis (f) and controls (d). Multinucleated (>3 nuclei per cell) and TRAP+ cells were
identified as OCs. The arrows point to the OCs (magnification 200 X). (B) The inhibition of
RANKL by RANK-Fc prevented in a dose-dependent manner the OC formation in unstimulated
and unfractionated PBMC cultures from MM patients. (C) Photomicrographs of the pits formed on
Millennium Osteologic slides by the OCs. Numerous and large resorption areas (arrows) were
observed in the MM bone disease samples (h) respect to the few and small pits detected in the
controls (g). The percentage of mineral surface resorbed by the OCs was quantified with an image
analyzer. The data reported in graph corresponde to the mean ± SE (i).
Figure 2. T cells mediate osteoclastogenesis in human MM PBMCs.
Osteoclastogenesis occurred in T cell-depleted MM PBMCs cultured in the absence (A) or in the
presence (B) of M-CSF and RANKL. Small-sized OCs developed in the absence of M-CSF and
RANKL (A), whereas exogenous cytokines triggered the formation of large-sized OCs (B).
Multinucleated and TRAP+ cells were identified as OCs. The arrows point to the OCs
(magnification 200 X).
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

28
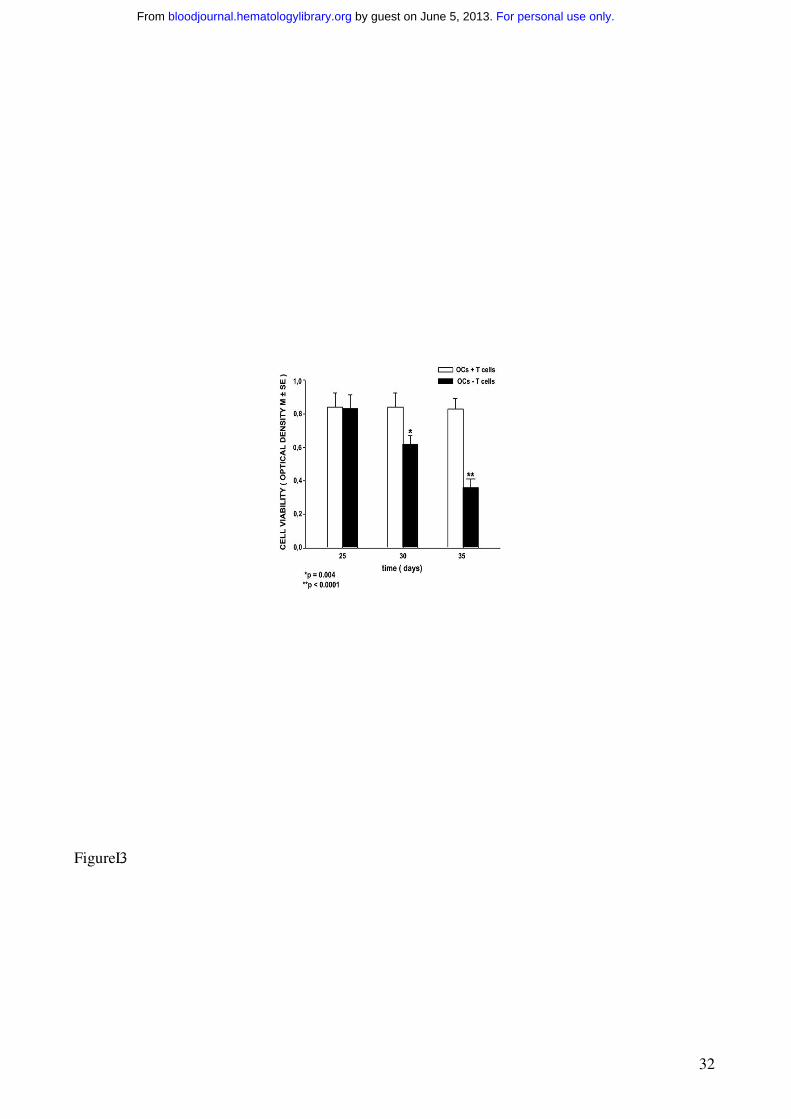
Figure 3. T cells increase cell viability of the OCs from human MM PBMCs.
Cell viability was measured by MTT assay. MM PBMCs from patients with osteolysis were
cultured in the presence (OCs + T cells) or in the absence (OCs - T cells) of T cells for 25, 30 and
35 days. M-CSF and RANKL were added to the T cell-depleted cultures in order to allow OC
formation. The viability of the OCs generated in the presence of T cells without addition of
exogenous cytokines was significantly higher than that of the OCs formed in the absence of T cells.
Figure 4. Cytokine expression by fresh T and MM cells.
Fresh T cells purified from PBMCs of MM patients with or without osteolysis as well as from
controls were analyzed for RANKL, OPG and TRAIL expression, respectively, by RT-PCR (A) and
western-blotting (B). MM cells purified from BM of MM patients with osteolysis were analyzed for
RANKL expression by RT-PCR (C).
The results show the overexpression of RANKL, OPG and TRAIL by fresh T cells from MM
patients with osteolysis compared to those without osteolysis and controls (A-B). MM cells were
found not expressing RANKL (C). The intensity of the bands obtained by RT-PCR was quantified
by densitometry (histograms) and normalized to GAPDH.
Figure 5. OPG/TRAIL complex in fresh T cells and culture media from PBMCs of MM
patients, and anti-TRAIL mAb effect on osteoclastogenesis.
TRAIL and OPG were coimmunoprecititated by a mAb against TRAIL in PBMC fresh T cell
lysates and PBMC culture media. Mouse IgG were utilized as control, and a total T cell lysate was
also loaded. OPG was identified as 55 kD band (A).
PBMCs from MM patients were cultured in the presence of an anti-TRAIL mAb at different
concentrations ± RANKL. The anti-TRAIL mAb, added to the culture media in the absence of
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

29
exogenous RANKL, significantly reduced osteoclastogenesis (B). Further, the addition of RANKL
rescued the anti-TRAIL mAb inhibitory effect. The graph represents the mean values ± SE of a
representative experiment.
Figure 6. OC expression of TRAIL and its receptors.
OC expression of TRAIL and its death (DR4) and decoy (DcR2) receptors was analyzed by RT-
PCR (A) and western blotting (B). Fully differentiated OCs were obtained in unfractionated as well
as T cell-depleted MM PBMC cultures (the latter following the addition of M-CSF and RANKL).
The OCs generated in the absence of T cells overexpressed TRAIL DR4 receptor at both mRNA
(A) and protein levels (B). On the contrary, the OCs from T cell-depleted cultures overexpressed
DcR2 at both mRNA (A) and protein levels (B). No significant difference was found in TRAIL
expression. The intensity of the bands was quantified by densitometry (histograms) and normalized
to GAPDH.
Figure 7. OC expression of Bcl-2 and Bax
The analysis of Bcl-2 and Bax expression by OCs, developed in unfractionated as well as T cell-
depleted PBMC cultures derived from MM patients with osteolysis, was performed by RT-PCR (A)
and western blotting (B). T cell-depleted cultures were stimulated with exogenous M-CSF and
RANKL. The intensity of the bands was quantified by densitometry. The graphs represent the mean
OD of Bcl-2/Bax ratio normalized to the OD of GAPDH or β-actin of a representative experiment
for RT-PCR and western blotting, respectively. Bcl-2 is highly expressed in the OCs obtained in the
presence of T cells.
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
30
Figure 1
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom

31
Figure 2
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
32
Figure 3
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
33
Figure 4
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
34
Figure 5
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
35
Figure 6
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
36
Figure 7
For personal use only. by guest on June 5, 2013. bloodjournal.hematologylibrary.orgFrom
Top Related
Copyright © 2022 FDOKUMEN

