







Bahasa
Halaman
Hukum


Sugar-inducible expression of the nucleolin-1 gene ofArabidopsis thaliana and its role in ribosome synthesis,growth and development
Hisae Kojima1, Takamasa Suzuki1, Takenori Kato1, Ken-ichi Enomoto1, Shusei Sato2, Tomohiko Kato2, Satoshi Tabata2,
Julio Saez-Vasquez3, Manuel Echeverrıa3, Tsuyoshi Nakagawa4, Sumie Ishiguro1 and Kenzo Nakamura1*
1Laboratory of Biochemistry, Department of Biological Functions and Mechanisms, Graduate School of Bioagricultural
Sciences, Nagoya University, Furo-cho, Chikusa, Nagoya 464-8601, Japan,2Kazusa DNA Research Institute, Kisarazu, Chiba 292-0818, Japan,3Laboratoire Genome et Developpement des Plantes, UMR CNRS-IRD 5096, Universite de Perpignan, 66860 Perpignan Cedex,
France, and4Research Institute of Molecular Genetics, Shimane University, Matsue, Shimane 690-8504, Japan
Received 23 July 2006; revised 21 October 2006; accepted 10 November 2006.*For correspondence (fax þ81 52 789 4094; e-mail [email protected]).
Summary
Animal and yeast nucleolin function as global regulators of ribosome synthesis, and their expression is tightly
linked to cell proliferation. Although Arabidopsis contains two genes for nucleolin, AtNuc-L1 is the
predominant if not only form of the protein found in most tissues, and GFP–AtNuc-L1 fusion proteins were
targeted to the nucleolus. Expression of AtNuc-L1 was strongly induced by sucrose or glucose but not by non-
metabolizable mannitol or 2-deoxyglucose. Sucrose also caused enhanced expression of genes for subunits of
C/D and H/ACA small nucleolar ribonucleoproteins, as well as a large number of genes for ribosomal proteins
(RPs), suggesting that carbohydrate availability regulates de novo ribosome synthesis. In sugar-starved cells,
induction of AtNuc-L1 occurred with 10 mM glucose, which seemed to be a prerequisite for resumption of
growth. Disruption of AtNuc-L1 caused an increased steady-state level of pre-rRNA relative to mature 25S
rRNA, and resulted in various phenotypes that overlap those reported for several RP gene mutants, including a
reduced growth rate, prolonged lifetime, bushy growth, pointed leaf, and defective vascular patterns and pod
development. These results suggest that the rate of ribosome synthesis in the meristem has a strong impact
not only on the growth but also the structure of plants. The AtNuc-L1 disruptant exhibited significantly
reduced sugar-induced expression of RP genes, suggesting that AtNuc-L1 is involved in the sugar-inducible
expression of RP genes.
Keywords: nucleolin, ribosome synthesis, ribosomal protein genes, snoRNP, sugar.
Introduction
In addition to growth regulators such as auxin and cytokinin,
nutrient availability is an important factor limiting cell pro-
liferation in plants. Sucrose (Suc) and glucose (Glc) induce
the expression of CycD2, CycD3 and CycD4 in Arabidopsis,
and cytokinin induction of CycD3 only occurs in the presence
of Suc (Murray et al., 1998; de Veylder et al., 1999, Riou-
Khamlichi et al., 2000). During cell division, cell proliferation
(increase in cell number) must be accompanied by cell
growth (increase in cell mass), which is primarily determined
by the protein synthetic activity of the cell, a process that uses
a great deal of energy and is tightly regulated by the
nutritional status of the cell. In yeast and animals, the protein
synthetic activity of the cell is regulated by the ‘target of
rapamycin’ (TOR) kinase-mediated pathway, which adjusts
the translational activity of pre-existing ribosomes and the
synthesis of ribosomes according to nutritional availability
(reviewed by Schmelzle and Hall, 2000; Raught et al., 2001).
The TOR gene in Arabidopsis is expressed in primary
meristem and is essential for growth (Menand et al., 2002).
The synthesis of functional ribosomes requires the
coordinated assembly of 70–80 different ribosomal proteins
(RPs) and four species of rRNA, yielding mature 40S and 60S
ª 2006 The Authors 1053Journal compilation ª 2007 Blackwell Publishing Ltd
The Plant Journal (2007) 49, 1053–1063 doi: 10.1111/j.1365-313X.2006.03016.x

ribosomal subunits. Most steps of ribosome synthesis take
place in the nucleolus, which contains many non-ribosomal
RNAs and proteins that assist in ribosome synthesis. One of
these proteins, nucleolin, plays important roles in various
steps of ribosomal synthesis, such as the transcription of
rDNA repeats, the modification and processing of pre-rRNA,
the assembly of pre-ribosomal particles, and nuclear–cyto-
plasmic transport of RPs and ribosomal subunits (reviewed
by Tuteja and Tuteja, 1998; Ginisty et al., 1999; Srivastava
and Pollard, 1999). The N-terminal part of nucleolin from
various eukaryotes contains variable numbers of acidic
stretches that are similar to those of nuclear high-mobility
group proteins. This N-terminal region interacts with non-
transcribed spacer regions in rDNA repeats and histone H1
to influence rDNA transcription. The middle of the nucleolin
sequences includes RNA-binding domains called RNA
recognition motifs (RRMs). Animal nucleolin possesses four
RRMs, whereas yeast homologs possess two. Nucleolin
interacts with the stem–loop structure of RNA through its
RRM and participates in the modification and processing of
pre-rRNA. The C-terminal part of nucleolin contains glycine-
and arginine-rich (GAR) domains that are implicated in
ribosomal assembly and nuclear import of RPs.
Nucleolin is also involved in processes other than the
ribosome synthesis. Nucleolin possesses DNA helicase
activity and interacts with replication protein A, suggesting
that it participates in DNA unwinding and replication (Kim
et al., 2005; Nasirudin et al., 2005). Nucleolin in animals
interacts with various transcription factors and nuclear
components, and is involved in the regulation of RNA
polymerase II-dependent gene expression (Masumi et al.,
2006; Huddleson et al., 2006). Remarkably, nucleolin pos-
sesses a histone chaperone activity that activates chromatin
remodeling complexes and facilitates transcription through
the nucleosomes (Angelov et al., 2006).
In animals and yeast, expression and activity of nucleolin is
coordinated with the expression of genes for rRNA and RPs,
and correlates with the proliferative activity of the cell
(Srivastava and Pollard, 1999). Like yeast nucleolin, nucleo-
lin-like proteins from alfalfa (Medicago sativa; Bogre et al.,
1996), pea (Pisum sativum; Tong et al., 1997; Reichler et al.,
2001) and Arabidopsis (Saez-Vasquez et al., 2004) possess
two RRMs, and expression of pea nucleolin cDNA in a yeast
mutant deficient in nucleolin rescues the reduced level of
rRNA and the growth rate (Reichler et al., 2001). Regulation of
expression of the nucleolin gene in plants has been studied in
only a few cases. Expression of the alfalfa gene for nucleolin,
nucMs1, occurs predominantly in meristematic tissues,
where its expression is limited to cells actively engaged in
cell division (Bogre et al., 1996). Light induces the expression
of nucleolin in alfalfa and pea, which is probably mediated by
phytochrome (Bogre et al., 1996; Tong et al., 1997).
Recent global gene expression analyses in Arabidopsis
indicate that the expression of a large number of genes
involved in protein synthesis is regulated by the carbohy-
drate and nitrogen nutritional status (Price et al., 2004; Li
et al., 2006). Our microarray analysis also indicated that
genes involved in protein synthesis are significantly
enriched among genes that are upregulated by Suc (Yoine
et al., 2006). In particular, the nucleolin gene was one of
the genes that was induced the most strongly by Suc. In the
present study, we examine the role of nucleolin in the
regulation of ribosome synthesis and in the growth and
development of plants.
Results
Nucleolin genes of Arabidopsis thaliana
Of the sugar-induced genes that we identified by micro-
array analysis, a gene encoding nucleolin (At1g48920) was
one of the most strongly induced by Suc within 6 h. This
gene encodes a protein with structural similarities to Nsr1
of Saccharomyces cerevisiae (Kondo and Inouye, 1992)
and Gar2 of Schizosaccharomyces pombe (Gulli et al.,
1995), as well as nucleolins from alfalfa (Bogre et al., 1996)
and pea (Tong et al., 1997). The Arabidopsis genome also
contains another gene for nucleolin, At3g18610. The pro-
teins encoded by At1g48920 and At3g18610 are identical
with AtNuc-L1 and AtNuc-L2, respectively, described pre-
viously (Saez-Vasquez et al., 2004). The AtNuc-L1 and
AtNuc-L2 genes are located on segment 1, a large dupli-
cated segment between chromosomes 1 and 3, respect-
ively (Arabidopsis Genome Initiative, 2000). Although both
AtNuc-L1 and AtNuc-L2 contain two RRMs, the AtNuc-L1
gene is interrupted by 14 introns (Figure 1a), whereas the
AtNuc-L2 gene contains 17 introns. AtNuc-L1 mRNA was
detected in various organs of Col plants by RT-PCR
(Figure 1b). In contrast, little AtNuc-L2 mRNA was detected
in organs other than flower buds.
Genes encoding fusion proteins with GFP at either the
C-terminus (AtNuc-L1–GFP) or N-terminus (GFP–AtNuc-L1)
of AtNuc-L1 were placed downstream of the CaMV 35S
promoter and used to generate stably transformed tobacco
BY-2 cells (Matsuoka and Nakamura, 1991). In cells expres-
sing AtNuc-L1–GFP or GFP–AtNuc-L1, strong fluorescent
GFP signals were detected in nucleoli and absent from the
cytoplasm (Figure 1c). Weak GFP fluorescence was also
detected in the nucleoplasm, where fluorescence occasion-
ally appeared as spots. In contrast, signals were absent in
nucleoli of cells expressing free GFP. In roots of Arabidopsis
plants transformed with these fusion genes, strong GFP
fluorescence appeared in nucleoli (Figure 1d).
Sugar-inducible expression of AtNuc-L1
Figure 2(a) shows the time course of changes in the level of
AtNuc-L1 mRNA after treatment of leaves of Arabidopsis Col
1054 Hisae Kojima et al.
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063
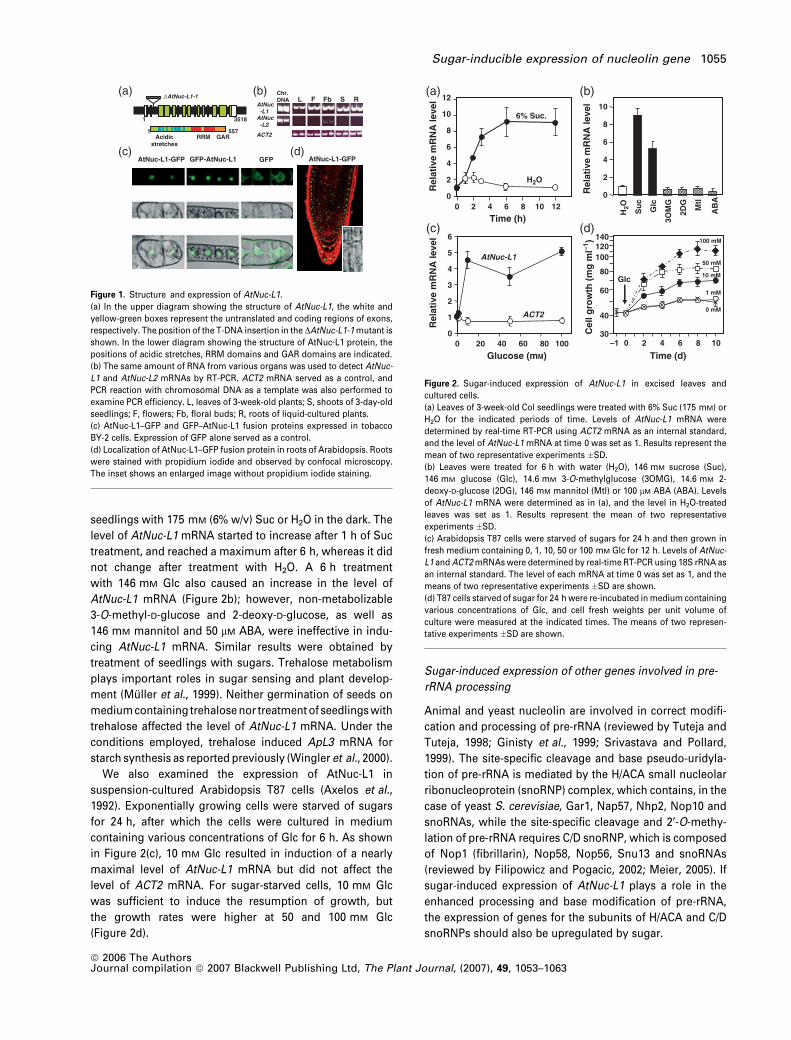
seedlings with 175 mM (6% w/v) Suc or H2O in the dark. The
level of AtNuc-L1 mRNA started to increase after 1 h of Suc
treatment, and reached a maximum after 6 h, whereas it did
not change after treatment with H2O. A 6 h treatment
with 146 mM Glc also caused an increase in the level of
AtNuc-L1 mRNA (Figure 2b); however, non-metabolizable
3-O-methyl-D-glucose and 2-deoxy-D-glucose, as well as
146 mM mannitol and 50 lM ABA, were ineffective in indu-
cing AtNuc-L1 mRNA. Similar results were obtained by
treatment of seedlings with sugars. Trehalose metabolism
plays important roles in sugar sensing and plant develop-
ment (Muller et al., 1999). Neither germination of seeds on
medium containing trehalose nor treatment of seedlings with
trehalose affected the level of AtNuc-L1 mRNA. Under the
conditions employed, trehalose induced ApL3 mRNA for
starch synthesis as reported previously (Wingler et al., 2000).
We also examined the expression of AtNuc-L1 in
suspension-cultured Arabidopsis T87 cells (Axelos et al.,
1992). Exponentially growing cells were starved of sugars
for 24 h, after which the cells were cultured in medium
containing various concentrations of Glc for 6 h. As shown
in Figure 2(c), 10 mM Glc resulted in induction of a nearly
maximal level of AtNuc-L1 mRNA but did not affect the
level of ACT2 mRNA. For sugar-starved cells, 10 mM Glc
was sufficient to induce the resumption of growth, but
the growth rates were higher at 50 and 100 mM Glc
(Figure 2d).
Sugar-induced expression of other genes involved in pre-
rRNA processing
Animal and yeast nucleolin are involved in correct modifi-
cation and processing of pre-rRNA (reviewed by Tuteja and
Tuteja, 1998; Ginisty et al., 1999; Srivastava and Pollard,
1999). The site-specific cleavage and base pseudo-uridyla-
tion of pre-rRNA is mediated by the H/ACA small nucleolar
ribonucleoprotein (snoRNP) complex, which contains, in the
case of yeast S. cerevisiae, Gar1, Nap57, Nhp2, Nop10 and
snoRNAs, while the site-specific cleavage and 2¢-O-methy-
lation of pre-rRNA requires C/D snoRNP, which is composed
of Nop1 (fibrillarin), Nop58, Nop56, Snu13 and snoRNAs
(reviewed by Filipowicz and Pogacic, 2002; Meier, 2005). If
sugar-induced expression of AtNuc-L1 plays a role in the
enhanced processing and base modification of pre-rRNA,
the expression of genes for the subunits of H/ACA and C/D
snoRNPs should also be upregulated by sugar.
Figure 1. Structure and expression of AtNuc-L1.
(a) In the upper diagram showing the structure of AtNuc-L1, the white and
yellow-green boxes represent the untranslated and coding regions of exons,
respectively. The position of the T-DNA insertion in the DAtNuc-L1-1 mutant is
shown. In the lower diagram showing the structure of AtNuc-L1 protein, the
positions of acidic stretches, RRM domains and GAR domains are indicated.
(b) The same amount of RNA from various organs was used to detect AtNuc-
L1 and AtNuc-L2 mRNAs by RT-PCR. ACT2 mRNA served as a control, and
PCR reaction with chromosomal DNA as a template was also performed to
examine PCR efficiency. L, leaves of 3-week-old plants; S, shoots of 3-day-old
seedlings; F, flowers; Fb, floral buds; R, roots of liquid-cultured plants.
(c) AtNuc-L1–GFP and GFP–AtNuc-L1 fusion proteins expressed in tobacco
BY-2 cells. Expression of GFP alone served as a control.
(d) Localization of AtNuc-L1–GFP fusion protein in roots of Arabidopsis. Roots
were stained with propidium iodide and observed by confocal microscopy.
The inset shows an enlarged image without propidium iodide staining.
Figure 2. Sugar-induced expression of AtNuc-L1 in excised leaves and
cultured cells.
(a) Leaves of 3-week-old Col seedlings were treated with 6% Suc (175 mM) or
H2O for the indicated periods of time. Levels of AtNuc-L1 mRNA were
determined by real-time RT-PCR using ACT2 mRNA as an internal standard,
and the level of AtNuc-L1 mRNA at time 0 was set as 1. Results represent the
mean of two representative experiments �SD.
(b) Leaves were treated for 6 h with water (H2O), 146 mM sucrose (Suc),
146 mM glucose (Glc), 14.6 mM 3-O-methylglucose (3OMG), 14.6 mM 2-
deoxy-D-glucose (2DG), 146 mM mannitol (Mtl) or 100 lM ABA (ABA). Levels
of AtNuc-L1 mRNA were determined as in (a), and the level in H2O-treated
leaves was set as 1. Results represent the mean of two representative
experiments �SD.
(c) Arabidopsis T87 cells were starved of sugars for 24 h and then grown in
fresh medium containing 0, 1, 10, 50 or 100 mM Glc for 12 h. Levels of AtNuc-
L1 and ACT2 mRNAs were determined by real-time RT-PCR using 18S rRNA as
an internal standard. The level of each mRNA at time 0 was set as 1, and the
means of two representative experiments �SD are shown.
(d) T87 cells starved of sugar for 24 h were re-incubated in medium containing
various concentrations of Glc, and cell fresh weights per unit volume of
culture were measured at the indicated times. The means of two represen-
tative experiments �SD are shown.
Sugar-inducible expression of nucleolin gene 1055
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063
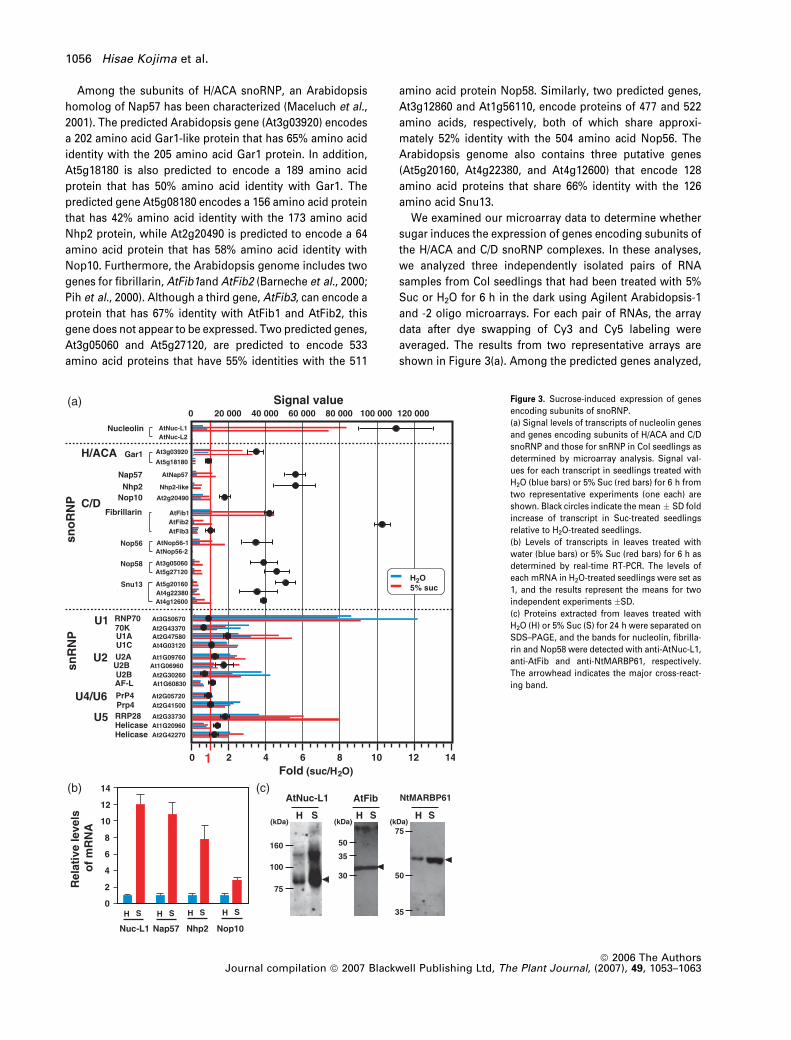
Among the subunits of H/ACA snoRNP, an Arabidopsis
homolog of Nap57 has been characterized (Maceluch et al.,
2001). The predicted Arabidopsis gene (At3g03920) encodes
a 202 amino acid Gar1-like protein that has 65% amino acid
identity with the 205 amino acid Gar1 protein. In addition,
At5g18180 is also predicted to encode a 189 amino acid
protein that has 50% amino acid identity with Gar1. The
predicted gene At5g08180 encodes a 156 amino acid protein
that has 42% amino acid identity with the 173 amino acid
Nhp2 protein, while At2g20490 is predicted to encode a 64
amino acid protein that has 58% amino acid identity with
Nop10. Furthermore, the Arabidopsis genome includes two
genes for fibrillarin, AtFib1and AtFib2 (Barneche et al., 2000;
Pih et al., 2000). Although a third gene, AtFib3, can encode a
protein that has 67% identity with AtFib1 and AtFib2, this
gene does not appear to be expressed. Two predicted genes,
At3g05060 and At5g27120, are predicted to encode 533
amino acid proteins that have 55% identities with the 511
amino acid protein Nop58. Similarly, two predicted genes,
At3g12860 and At1g56110, encode proteins of 477 and 522
amino acids, respectively, both of which share approxi-
mately 52% identity with the 504 amino acid Nop56. The
Arabidopsis genome also contains three putative genes
(At5g20160, At4g22380, and At4g12600) that encode 128
amino acid proteins that share 66% identity with the 126
amino acid Snu13.
We examined our microarray data to determine whether
sugar induces the expression of genes encoding subunits of
the H/ACA and C/D snoRNP complexes. In these analyses,
we analyzed three independently isolated pairs of RNA
samples from Col seedlings that had been treated with 5%
Suc or H2O for 6 h in the dark using Agilent Arabidopsis-1
and -2 oligo microarrays. For each pair of RNAs, the array
data after dye swapping of Cy3 and Cy5 labeling were
averaged. The results from two representative arrays are
shown in Figure 3(a). Among the predicted genes analyzed,
Figure 3. Sucrose-induced expression of genes
encoding subunits of snoRNP.
(a) Signal levels of transcripts of nucleolin genes
and genes encoding subunits of H/ACA and C/D
snoRNP and those for snRNP in Col seedlings as
determined by microarray analysis. Signal val-
ues for each transcript in seedlings treated with
H2O (blue bars) or 5% Suc (red bars) for 6 h from
two representative experiments (one each) are
shown. Black circles indicate the mean � SD fold
increase of transcript in Suc-treated seedlings
relative to H2O-treated seedlings.
(b) Levels of transcripts in leaves treated with
water (blue bars) or 5% Suc (red bars) for 6 h as
determined by real-time RT-PCR. The levels of
each mRNA in H2O-treated seedlings were set as
1, and the results represent the means for two
independent experiments �SD.
(c) Proteins extracted from leaves treated with
H2O (H) or 5% Suc (S) for 24 h were separated on
SDS–PAGE, and the bands for nucleolin, fibrilla-
rin and Nop58 were detected with anti-AtNuc-L1,
anti-AtFib and anti-NtMARBP61, respectively.
The arrowhead indicates the major cross-react-
ing band.
1056 Hisae Kojima et al.
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063

AtNuc-L2 and AtNop56-2 (At1g56110) did not give signifi-
cant signals. One of the two genes for Gar1-like protein,
At5g18180, and AtFib3 showed signals that did not vary
between Suc- and H2O-treated plants. However, all the other
genes showed Suc-induced increases in the mRNA level. In
addition to AtNuc-L1, Suc caused an approximately 10-fold
increase in the level of AtFib2 mRNA. Other genes showed
1.8–5.5-fold increases by Suc treatment. To confirm the
microarray data, we examined the levels of several mRNAs
by quantitative real-time RT-PCR. In addition to an 11-fold
induction of AtNuc-L1 mRNA by Suc, the levels of mRNAs
for AtNap57, AtNhp2 and AtNop10 were 3–10-fold higher in
seedlings treated with Suc than those treated with H2O
(Figure 3b). In contrast to genes encoding subunits of H/ACA
and C/D snoRNP complexes, Suc did not increase the mRNA
levels for the 14 putative genes encoding the subunits of U1,
U2, U4/U6 and U5 snRNP complexes, which are involved in
intron splicing of pre-mRNAs.
To determine whether the induction of AtNuc-L1 mRNA by
sugar is accompanied by an increase in the level of protein,
we extracted proteins from seedlings that had been treated
with 5% Suc or H2O for 24 h and analyzed them by immu-
noblotting with an anti-AtNuc-L1 antibody (Saez-Vasquez
et al., 2004). The anti-AtNuc-L1 antibody reacted with an
approximately 78 kDa polypeptide, which is larger than the
expected molecular mass of 58.8 kDa (Figure 3c). In addition,
the antibody revealed a weaker band of approximately
140 kDa. The intensities of both bands were stronger in
extracts from Suc-treated seedlings than those from H2O-
treated seedlings. We also examined levels of fibrillarin using
an anti-AtFib antibody (Saez-Vasquez et al., 2004) and those
of the putative Nop58 protein using an antibody against a
tobacco homolog of Nop58 (NtMARBP61; Fujiwara et al.,
2002). The anti-AtFib antibody detected a single band with an
apparent molecular mass of 32 kDa. Despite the induction of
AtFib1 and AtFib2 mRNAs by Suc, the intensity of this 32 kDa
band did not differ between the H2O- and Suc-treated
seedlings. The anti-NtMARBP61 antibody reacted with a
single band with an apparent molecular mass of 58 kDa that
appeared to be increased in the Suc-treated plants.
Reduced processing of rRNA precursors in a disruptant of
AtNuc-L1
We searched for T-DNA insertion lines of AtNuc-L1 in the
Kazusa T-DNA tag-line collection by PCR screening of
pooled chromosomal DNA, and identified a line in which
T-DNA was inserted in the second intron (Figure 1a). In this
DAtNuc-L1-1 mutant in the Col background, we detected
neither AtNuc-L1 mRNA by RT-PCR (Figure 4a) nor AtNuc-L1
protein by immunoblotting with an anti-AtNuc-L1 antibody
(Figure 4b).
The 45S pre-rRNA contains a 5¢ external transcribed
sequence (ETS), internal transcribed sequences 1 and 2
(ITS1 and ITS2), and a 3¢ ETS, which are removed during a
complex series of maturation steps (reviewed by Fromont-
Racine et al., 2003). To examine the effects of AtNuc-L1
deficiency on the processing of 45S pre-rRNA, we compared
the levels of corresponding RNA sequences between Col
and DAtNuc-L1-1 plants by real-time RT-PCR using primers
specific to various regions of the 45S pre-rRNA (Figure 4c).
To quantify ETS and ITS sequences, 1 lg of RNA was used
as a template for the first-strand cDNA synthesis, whereas
0.2 ng of RNA was used for quantification of mature rRNA
sequences. We normalized the signal values for each PCR
product against the values for 25S rRNA and then compared
the relative values for Col and DAtNuc-L1-1 plants. We found
that the relative levels of the 5¢ ETS, ITS1, ITS2 and 3¢ ETS
sequences were 1.7–4-fold higher in DAtNuc-L1-1 plants
than in Col plants (Figure 4c), suggesting that the steady-
state level of pre-rRNA relative to mature 25S rRNA is higher
in DAtNuc-L1-1 than in Col plants.
Growth and developmental phenotypes of DAtNuc-L1-1
plants
The DAtNuc-L1-1 plants showed various growth and devel-
opmental phenotypes. The growth of roots of DAtNuc-L1-1
Figure 4. Reduced pre-rRNA processing in DAtNuc-L1-1.
(a) Detection of AtNuc-L1 mRNA by RT-PCR in the same amounts of RNA from
Col and DAtNuc-L1-1 plants (D). Tubulin mRNA (TUA3/5) served as a control.
(b) Total proteins (15 lg) extracted from Col and DAtNuc-L1-1 (D) seedlings
were separated by SDS–PAGE and analyzed by immunoblotting with an anti-
AtNuc-L1 antibody. The arrowhead indicates the 78 kDa band (see Figure 3c).
(c) Comparison of the relative levels of the various regions of pre-rRNA in Col
and DAtNucL1-1 plants. Primer sets specific to various regions of 45S pre-
rRNA (upper panel) were used for synthesis of the first-strand cDNA and
determination of the levels of corresponding RNA sequences by real-time RT-
PCR. Signal values for each PCR product were normalized against the values
for 25S mature rRNA, and then compared between Col and DAtNuc-L1-1
plants (lower panel). Black bars represent ETS and ITS sequences, and white
bars represent 16S, 5.8S and 25S vRNAs. The values represent the
means � SD for two independent isolations of RNAs.
Sugar-inducible expression of nucleolin gene 1057
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063

germinated on vertically placed agar plates was 60–80%
slower than that of Col seedlings (Figure 5a), and the above-
ground parts of seedlings grown on agar plates were stun-
ted (Figure 5b). The DAtNuc-L1-1 plants also grew more
slowly than Col plants on vermiculite (Figure 5c). They often
showed outgrowth of organs from axillary buds (Figure 5d),
bushy growth with many stems, and a longer life duration
compared with the Col plants (Figure 5e).
The leaves of young DAtNuc-L1-1 seedlings had a narrow
pointed shape in contrast to the round shape of Col leaves
(Figure 5b), and showed abnormal vascular patterns (Fig-
ure 5f). In addition to the reduced growth rate, these
phenotypes are similar to those commonly observed for
mutants with disruptions of RP genes, such as pointed first
leaf(pfl), in which AtRPS18A (pfl1; van Lijsebettens et al.,
1994) or AtRPS13A (pfl2; Ito et al., 2000) is disrupted,
Arabidopsis minute-like 1 (aml1), in which AtRPS5A is
disrupted (Weijers et al., 2001), and short valve 1 (stv1), in
which AtRPL24B is disrupted (Nishimura et al., 2005). In
addition, pods of DAtNuc-L1-1 plants were shorter and
irregular in size and shape compared with those of Col
plants (Figure 5g). Furthermore, the stalks of the DAtNuc-L1-
1 pods were longer than those of Col pods (Figure 5g;
arrowheads). A long stalk is a characteristic feature of the
pods from the stv1 mutant as well as those from mutants of
auxin response factors ETTIN and MONOPTEROS, which are
defective in gynoecium development (Nishimura et al.,
2005). AtRPL24, encoded by STV1, is specifically involved
in translation re-initiation of polycistronic mRNAs, and
disruption of AtRPL24B affects the expression of ETTIN
and MONOPTEROS genes, which contain upstream open
reading frames in their 5¢ untranslated sequences (Nishim-
ura et al., 2005). These results suggest that DAtNuc-L1-1
plants contain a reduced amount of RPs due to defective
ribosome synthesis and suffer from a shortage of ribosomes
in proliferating cells.
Expression of RP genes in DAtNuc-L1-1 plants
Because DAtNuc-L1-1 plants seemed to contain a reduced
amount of ribosomes and RPs, we examined the expression
of RP genes in these plants. Figure 6(a) shows the scatter
plots of representative microarray data for transcript levels
of Col and DAtNuc-L1-1 seedlings treated with 5% Suc or
H2O. Similar results were obtained in three independent
experiments for Col plants and two independent experi-
ments for DAtNuc-L1-1 plants. In Arabidopsis, each of the 80
RPs are encoded by a small family of 2–7 members (Barakat
et al., 2001). In Figure 6(a), the 217 RP genes are indicated by
pink triangles. In Col plants, the mRNA levels for 85% of the
RP genes were increased more than twofold in Suc-treated
plants compared with H2O-treated plants. In contrast, the
mRNA level for none of the RP genes was increased more
than twofold by Suc in the DAtNuc-L1-1 plants (Figure 6a).
For example, the mRNAs for AtRPL34A, AtRPL27A, At-
RPL27C, AtRPS3aA and AtRPS11A were increased two to
fivefold in Col plants treated with 5% Suc compared to plants
treated with H2O, whereas there was no noticeable induction
by Suc in DAtNuc-L1-1 plants (Figure 6b-1). The signal val-
ues for RP transcripts in H2O-treated seedlings were not
noticeably different between Col and DAtNuc-L1-1plants,
suggesting that the nucleolin deficiency affected the sugar-
induced increase in RP mRNAs. On average, Suc treatment
caused a 2.8-fold increase in the levels of the 217 RP mRNAs
in Col plants but only a 1.3-fold difference in DAtNuc-L1-1
plants.
Comparison of the scatter plots for Col and DAtNuc-L1-1
plants (Figure 6a) suggests that there was a general reduc-
tion in the magnitude of sugar-responsive changes in the
mRNA level in DAtNuc-L1-1 plants. Nevertheless, expression
of typical sugar-inducible genes such as b-amylase (Atb-
Amy; Mita et al., 1995), ADP-glucose pyrophosphorylase S,
Figure 5. Phenotypes of the DAtNuc-L1-1 mutant.
(a) Length of roots of Col (hatched bar) and DAtNuc-L1-1 plants (black bar) that
had been grown on vertically placed plates containing 0% or 1% Suc for 8 or
14 days. The results represent the means for 10–20 seedlings �SD.
(b, c) Seeds of Col and DAtNuc-L1-1 were germinated and grown on plates
containing 2% Suc for 3 weeks (b) or on vermiculite for 48 days (c).
(d) Axillary organ development in DAtNuc-L1-1.
(e) Representative Col and DAtNuc-L1-1 plants grown for 60 and 90 days,
respectively. Bar ¼ 1 cm.
(f) Vascular patterns in young leaves of Col and DAtNuc-L1-1 plants.
(g) Flowers and pods from the top (left) to the bottom (right) of one stem of Col
and DAtNuc-L1-1 are shown. The stalks of a pod from Col and DAtNuc-L1-
1plants are indicated by open and closed arrowheads, respectively.
Bar ¼ 0.5 cm.
1058 Hisae Kojima et al.
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063
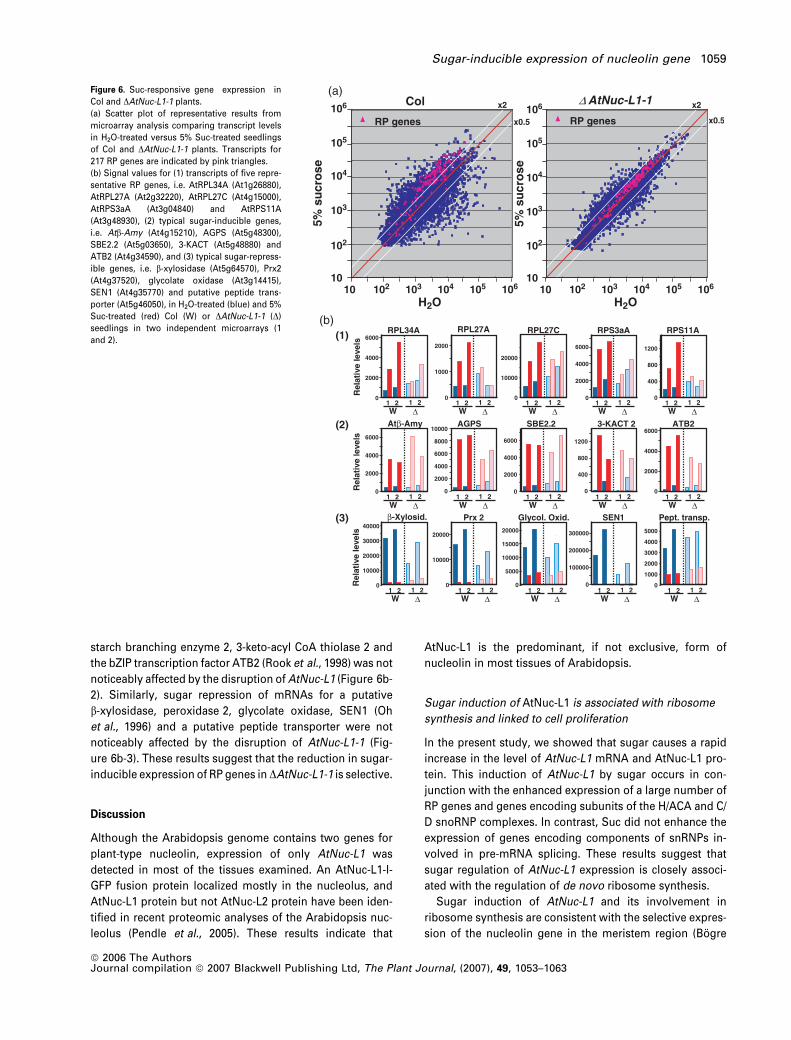
starch branching enzyme 2, 3-keto-acyl CoA thiolase 2 and
the bZIP transcription factor ATB2 (Rook et al., 1998) was not
noticeably affected by the disruption of AtNuc-L1 (Figure 6b-
2). Similarly, sugar repression of mRNAs for a putative
b-xylosidase, peroxidase 2, glycolate oxidase, SEN1 (Oh
et al., 1996) and a putative peptide transporter were not
noticeably affected by the disruption of AtNuc-L1-1 (Fig-
ure 6b-3). These results suggest that the reduction in sugar-
inducible expression of RP genes in DAtNuc-L1-1 is selective.
Discussion
Although the Arabidopsis genome contains two genes for
plant-type nucleolin, expression of only AtNuc-L1 was
detected in most of the tissues examined. An AtNuc-L1-l-
GFP fusion protein localized mostly in the nucleolus, and
AtNuc-L1 protein but not AtNuc-L2 protein have been iden-
tified in recent proteomic analyses of the Arabidopsis nuc-
leolus (Pendle et al., 2005). These results indicate that
AtNuc-L1 is the predominant, if not exclusive, form of
nucleolin in most tissues of Arabidopsis.
Sugar induction of AtNuc-L1 is associated with ribosome
synthesis and linked to cell proliferation
In the present study, we showed that sugar causes a rapid
increase in the level of AtNuc-L1 mRNA and AtNuc-L1 pro-
tein. This induction of AtNuc-L1 by sugar occurs in con-
junction with the enhanced expression of a large number of
RP genes and genes encoding subunits of the H/ACA and C/
D snoRNP complexes. In contrast, Suc did not enhance the
expression of genes encoding components of snRNPs in-
volved in pre-mRNA splicing. These results suggest that
sugar regulation of AtNuc-L1 expression is closely associ-
ated with the regulation of de novo ribosome synthesis.
Sugar induction of AtNuc-L1 and its involvement in
ribosome synthesis are consistent with the selective expres-
sion of the nucleolin gene in the meristem region (Bogre
Figure 6. Suc-responsive gene expression in
Col and DAtNuc-L1-1 plants.
(a) Scatter plot of representative results from
microarray analysis comparing transcript levels
in H2O-treated versus 5% Suc-treated seedlings
of Col and DAtNuc-L1-1 plants. Transcripts for
217 RP genes are indicated by pink triangles.
(b) Signal values for (1) transcripts of five repre-
sentative RP genes, i.e. AtRPL34A (At1g26880),
AtRPL27A (At2g32220), AtRPL27C (At4g15000),
AtRPS3aA (At3g04840) and AtRPS11A
(At3g48930), (2) typical sugar-inducible genes,
i.e. Atb-Amy (At4g15210), AGPS (At5g48300),
SBE2.2 (At5g03650), 3-KACT (At5g48880) and
ATB2 (At4g34590), and (3) typical sugar-repress-
ible genes, i.e. b-xylosidase (At5g64570), Prx2
(At4g37520), glycolate oxidase (At3g14415),
SEN1 (At4g35770) and putative peptide trans-
porter (At5g46050), in H2O-treated (blue) and 5%
Suc-treated (red) Col (W) or DAtNuc-L1-1 (D)
seedlings in two independent microarrays (1
and 2).
Sugar-inducible expression of nucleolin gene 1059
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063

et al., 1996). Expression of Arabidopsis RP genes, such as
AtRPS18A (van Lijsebettens et al., 1994) and AtRPS5A
(Weijers et al., 2001), also occurs predominantly in cells
actively engaged in cell division in the meristem. In Arabid-
opsis suspension-cultured cells that had been starved of
sugars, inclusion of 10 mM Glc in the medium was sufficient
to induce the maximum level of expression of AtNuc-L1 and
to cause the resumption of growth. On the other hand, the
growth rate increased with the concentration of Glc (up to
100 mM), suggesting that the induction of AtNuc-L1 expres-
sion by 10 mM Glc is prerequisite for the resumption of
growth rather than the expression of AtNuc-L1 being
controlled by the growth rate. Sugar induces the expression
of a variety of genes involved in nutrient storage, such as
genes for vegetative storage proteins and for the synthesis
of starch (reviewed by Koch, 1996; Rolland et al., 2006).
Sugar induction of expression of genes for reserve synthesis
generally occurs at much higher concentrations of sugars
and in quiescent cells.
Relationship of sugar regulation of ribosome synthesis with
cell division
The availability of sugars affects cell division in the meristem,
for which cell proliferation must be accompanied by cell
growth. In the early developmental stage of Vicia fabaseeds,
where cell proliferation and differentiation predominate, the
spatial distribution of Glc, rather than Suc, correlates well
with the mitotic index (Borisjuk et al., 1998). During these
stages, transported Suc is cleaved into Glc and fructose by
cell-wall-bound invertase, generating a high Glc/Suc ratio. In
the later stages of seed development, the Suc/Glc ratio is
high because of a decline in cell-wall-bound invertase, and
Suc synthase plays a predominant role in the utilization of
Suc for reserve synthesis (Borisjuk et al., 1998, 2004; ). In
Arabidopsis cells starved for sugars, the expression of CycD2
and CycD3 is induced by 10 mM Glc (Riou-Khamlichi et al.,
2000). Under similar conditions, the induction of expression
of AtNuc-L1 and other genes involved in ribosome synthesis
also occurred with 10 mM Glc. CycD2, but not CycD3, is
induced by 2-deoxy-D-glucose (2DG), suggesting that CycD2
expression is under the control of hexokinase sensor-
dependent sugar signaling, wherein hexokinase functions as
a Glc sensor (Jang et al., 1997). A recent study with Physc-
omytrella patens, however, suggested that sugar regulation
of CycD2 is more closely related to the developmental pro-
gression in response to nutrient availability than the control
of cell division (Lorenz et al., 2003). Similar to CycD3, the
expression of AtNuc-L1 and RP genes is not induced by 2DG,
suggesting that a metabolic signal derived from Glc is
required for the expression of genes related to cell division
and growth; however, in itself, the absence of a response to
2DG does not exclude the involvement of hexokinase sensor-
dependent signaling in the regulation.
Deficiency of AtNuc-L1 leads to a shortage of ribosomes in
dividing cells
Although Nsr1 of S. cerevisiae (Kondo and Inouye, 1992)
and Gar2 of S. pombe (Gulli et al., 1995) are not essential for
cell viability, their null mutants show growth defects. The
nsr1and gar2 mutants accumulate 35S pre-rRNA and have
reduced steady-state levels of the 40S ribosomal subunit,
which most likely causes the defective growth. Expression of
pea nucleolin cDNA in the nsr1 mutant rescues the reduced
amount of large subunit rRNA and the reduced growth rate
(Reichler et al., 2001). We found that the DAtNuc-L1-1 null
mutant showed an increased steady-state level of pre-rRNA
relative to mature 25S rRNA compared to Col. This differ-
ence could be due to delayed pre-rRNA processing or the
production of abnormal transcripts of pre-rRNA that are not
processed promptly. It is suggested that ribosome synthesis
is reduced in the mutant.
The idea that DAtNuc-L1-1 plants have a shortage of
ribosomes is supported by its phenotypes. In addition to
reduced growth rate, DAtNuc-L1-1 plants exhibited various
developmental phenotypes that overlap with those of pre-
viously reported Arabidopsis mutants defective in specific
RP genes (van Lijsebettens et al., 1994; Ito et al., 2000;
Weijers et al., 2001; Nishimura et al., 2005), including poin-
ted leaves, abnormal vascular patterning, and defective
gynoecium development. Thus, the synthesis of ribosomes
might be limited in both DAtNuc-L1-1 and RP mutants.
The DAtNuc-L1-1 plants showed reduced growth rate and
lived longer than Col plants. The reduced growth rate of
DAtNuc-L1-1 plants is probably due to reduced cell division
as a result of a shortage of ribosomes, and therefore an
inability to meet the demands for active protein synthesis.
The DAtNuc-L1-1 plants showed outgrowth of axillary
organs and bushy growth. A reduced rate of organ devel-
opment may alter the distribution of photoassimilates
among meristems of DAtNuc-L1-1 plants. These results
suggest that the rate of ribosome synthesis in the meristem
has a strong impact on the growth and the structure and
architecture of plants.
AtNuc-L1 deficiency affects gene expression pattern
The sugar-enhanced expression of RP genes was severely
diminished in DAtNuc-L1-1 plants compared with Col plants.
Although we observed a general diminution in the sugar-
induced changes of the transcript levels in DAtNuc-L1-1
plants, expression of typical sugar-inducible or -repressible
genes, e.g. Atb-Amy (Mita et al., 1995), ATB2 (Rook et al.,
1998) or SEN1 (Oh et al., 1996), was not noticeably affected.
The nucleolin deficiency seems to selectively affect the ability
of sugar to induce the expression of RP genes because the
signal values for transcripts in H2O-treated seedlings were
not noticeably different between Col and DAtNuc-L1-1plants.
1060 Hisae Kojima et al.
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063

Because sugar does not affect the stability of several RP
mRNAs (K. Enomoto, H. Kojima and K. Nakamura,
unpublished results), the reduced ability of sugar to activate
RP genes in DAtNuc-L1-1 is probably due to effects on tran-
scription. Most sugar-induced changes in transcript levels in
Arabidopsis require de novo protein synthesis (Price et al.,
2004). Although a reduction in protein synthesis may explain
the general reduction in the magnitude of sugar-induced
changes in transcript levels in DAtNuc-L1-1, it does not
account for the selective effect of nucleolin deficiency on the
sugar-inducible expression of RP genes.
Nucleolin binds to spacer regions in rDNA and histone H1
through its N-terminal acidic stretches, and participates in
transcription of rDNA by RNA polymerase I (Ginisty et al.,
1999; Srivastava and Pollard, 1999). Nuclear factor D purified
from the inflorescence of cauliflower (Brassica oleracea)
binds to rDNA and contains nucleolin-like protein, fibrillarin
and snoRNAs, suggesting that snoRNP may link rDNA
transcription and pre-rRNA maturation (Saez-Vasquez et al.,
2004). In addition, nucleolin in animals interacts with various
transcription factors and nuclear components (Ginisty et al.,
1999; Srivastava and Pollard, 1999), and is required for
transcriptional regulation of several genes by RNA polym-
erase II (Masumi et al., 2006; Huddleson et al., 2006). Animal
nucleolin has been shown to exhibit histone chaperone
activity and enhances the remodeling of nucleosomes by
SWI/SNF and ATP-dependent chromatin-assembly factor
(Angelov et al., 2006). In particular, nucleolin promotes the
remodeling of nucleosomes that contain histone variant,
which are otherwise resistant to remodeling, and it facili-
tates passage of RNA polymerase II through nucleosomes.
In yeast, the TOR pathway regulates expression of rDNA and
RP genes by chromatin-mediated mechanisms in response
to nutrient availability (Rohde and Cardenas, 2003; Tsang
et al., 2003). It seems worth examining the possibility that
AtNuc-L1 regulates the transcription of RP genes through a
chromatin-mediated mechanism.
Experimental procedures
Plant materials and treatment with sugars
Seeds of Arabidopsis thaliana (L.) Heynh. (ecotype Col-0) weresurface-sterilized, kept at 4�C for 3 days in sterile water, and sown on0.3% gellan gum plates containing Murashige and Skoog medium(pH 5.7), 100 mg l)1 myo-inositol, 10 mg l)1 thiamine-HCl, 1 mg l)1
nicotinic acid, 1 mg l)1 pyridoxine HCl and 2% w/v Suc. Plates wereincubated in a growth chamber at 22�C under continuous light of65 lmol m)2 sec)1. Mature leaves of the 3-week-old plants wereexcised with a sharp razor blade, and the cut edges of petioles wereimmersed in a sterile solution of sugar or water and incubated at22�C in the dark (Mita et al., 1995). Plants were also grown on ver-miculite at 22�C under continuous light. Plants were watered withHoagland’s nutrient solution every week and water as needed.
A primary PCR screen for a T-DNA insertion mutant of AtNuc-L1was performed on pooled chromosomal DNA from approximately
20 000 individual lines of a collection of Arabidopsis T-DNAinsertion lines (Kazusa DNA Research Institute, Kazusa, Japan)using gene-specific primers and T-DNA border primers. For geno-typing of DAtNuc-L1-1, we carried out genomic PCR with a gene-specific PCR primer and a T-DNA right border primer (Table S1).Seeds of the homozygous T-DNA insertion line that had been back-crossed to Col twice were used for further analyses.
Arabidopsis suspension-cultured cell line T87 derived from Col(Axelos et al., 1992), was obtained from the RIKEN Plant Cell Bank(Yokohama, Japan) and grown in GB5 medium (Yamada et al.,2004), pH 5.7, containing 3.3 g l)1 of Gamborg’s B5 salt mixture(Wako www.wako-chem.co.jp), 1 ml l)1 of diluted Gamborg’s vita-min solution (Sigma-Aldrich, http://www.sigmaaldrich.com/),0.5 g l)1 of 2-morpholinoethanesulfonic acid monohydrate, 1 lM
1-naphthalene acetic acid and 1.5% Suc (w/v). The 6-day-old cellswere collected by centrifugation, and washed three times with freshmedium without Suc. Cells were starved of sugars for 24 h,resuspended in fresh medium containing 0, 1, 10, 50 or 100 mM
Glc, and grown for 6 h before isolating RNA.
Localization of GFP fusion proteins of AtNuc-L1
The full-length cDNA for AtNuc-L1 was obtained by RT-PCR usingthe primer set shown in Table S1. The cDNA was cloned intopGWB5 or pGWB6 vectors containing the coding sequence for sGFPby GatewayTM cloning technology (Invitrogen, http://www.invitro-gen.com/) to produce binary Ti plasmids carrying genes encodingAtNuc-L1–GFP and GFP–AtNuc-L1 fusion proteins under the controlof the CaMV 35S promoter, respectively. These plasmids were usedto transform tobacco BY-2 cells as described previously (Matsuokaand Nakamura, 1991). Several independent transformed calli werebrought into suspension culture, and GFP fluorescence was ob-served using an FV500 confocal fluorescence microscope (Olym-pus, http://www.olympus-global.com/). The same Agrobacteriumstrains were used to transform Arabidopsis plants, and roots of T2
plants from several independent transformed lines were stainedwith propidium iodide and observed for GFP fluorescence by con-focal fluorescence microscopy.
Isolation of RNA, RT-PCR and real-time RT-PCR
Total RNA was isolated from plants or tissues using an RNeasy PlantMini Kit (Qiagen, http://www.qiagen.com) and dissolved in ribo-nuclease-free water. For detection of mRNAs, first-strand cDNA wassynthesized from 2 lg of total RNA using oligo(dT)20 primers andSuperscript III (Invitrogen) and diluted with four volumes of water.PCR was performed in a 25 ll mixture containing 2 ll of the dilutedcDNA solution and 0.4 lM of each primer. The PCR reaction cycleswere as follows: denaturation at 95�C for 30 sec, annealing at 60�C for30 sec, and extension at 72�C for 30 sec. The number of cycles wasoptimized for each mRNA. For quantitative real-time RT-PCR, PCRwas performed with iQ SYBR Green Supermix using an iCycler iQ(Bio-Rad, http://www.bio-rad.com/), and the comparative thresholdcycle method was used to determine the relative levels of mRNAs,with ACT2 mRNA as an internal reference. The primer sets for AtNuc-L1, ACT2, AtNap57, AtNhp2 and AtNop10 are listed in Table S1.
To examine the processing of pre-rRNA, total RNA was isolatedfrom3-week-oldColandDAtNuc-L1-1 plants.Forquantification of25S,18S and 5.8S rRNA, cDNAs were synthesized from 200 pg of total RNAwith specific primers in a 20 ll reaction, whereas 1 lg of total RNAwas used for the synthesis of cDNA with specific primers for quanti-fication of ETS and ITS sequences of pre-rRNA. Quantitative real-timeRT-PCR was carried out using the primer sets listed in Table S1.
Sugar-inducible expression of nucleolin gene 1061
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063

Oligo microarray analysis
Total RNAs used for microarray analyses were prepared usingTrizol reagent (Invitrogen www.invitrogen.com) and subsequentlypurified using an RNeasy Plant Mini Kit (Qiagen). Cy3- andCy5-labeled cDNA probes were synthesized and hybridized to theAgilent Arabidopsis-1 and -2 oligo microarrays (Agilent Technol-ogies www.home.agilent.com) according to the manufacturer’sinstructions. The microarray analysis was performed with two orthree independently isolated RNA samples and assessed in eachexperiment by dye swapping as described previously (Yoine et al.,2006).
Extraction of proteins and immunological detection
Proteins were extracted from 3-week-old seedlings that had beentreated with 5% Suc or H2O for 24 h. SDS–PAGE was carried out inan 8% acrylamide gel. The proteins were transferred from the gel toa poly(vinylidene difluoride) membrane (Immobilon; Milliporewww.millipore.com), and the antigen on the membrane wasdetected with primary antibodies, followed by horseradish peroxi-dase-coupled protein A and chemiluminescence reagents (ECL kit;GE Healthcare www.gehealthcare.com). Antibodies raised againstrecombinant AtNuc-L1 (AtNuc-L1) and AtFib1 (Saez-Vasquez et al.,2004) were used, and the antibody against recombinantNtMARBP61 (Fujiwara et al., 2002) was a generous gift from DrMasayoshi Maeshima of Nagoya University.
Acknowledgements
We thank S. Ukai and T. Kawai for technical assistance, M. Mae-shima of Nagoya University for anti-NtMARBP61, and K. Shinozakiof RIKEN for T87 cells. This work was supported in part by theResearch for the Future program of the Japan Society for the Pro-motion of Science (grant number 00L01603) and the 21st CenturyCOE program from the Ministry of Education, Science, Sports andCulture of Japan to K.N.
Supplementary Material
The following supplementary material is available for this articleonline:Table S1 List of primersThis material is available as part of the online article from http://www.blackwell-synergy.com.
References
Angelov, D., Bondarenko, V.A., Almagro, S. et al. (2006) Nucleolin isa histone chaperone with FACT-like activity and assists remode-ling of nucleosomes. EMBO J. 25, 1669–1679.
Arabidopsis Genome Initiative (2000) Analysis of the genomesequence of the flowering plant Arabidopsis thaliana. Nature,408, 796–815.
Axelos, M., Curie, C., Mazzolini, L., Bardet, C. and Lescure, B. (1992)A protocol for transient gene expression in Arabidopsis thali-anaprotoplasts isolated from cell suspension cultures. PlantPhysiol. Biochem. 30, 123–128.
Barakat, A., Szick-Miranda, K., Chang, I.F., Guyot, R., Blanc, G.,
Cooke, R., Delseny, M. and Bailey-Serres, J. (2001) The organ-ization of cytoplasmic ribosomal protein genes in the Arabidopsisgenome. Plant Physiol. 127, 398–415.
Barneche, F., Steinmetz, F. and Echeverria, M. (2000) Fibrillaringenes encode both a conserved nucleolar protein and a novelsmall nucleolar RNA involved in ribosomal RNA methylation inArabidopsis thaliana. J. Biol. Chem. 275, 27212–27220.
Bogre, L., Jonak, C., Mink, M. et al. (1996) Developmental and cellcycle regulation of alfalfa nucMs1, a plant homolog of the yeastNsr1 and mammalian nucleolin. Plant Cell, 8, 417–428.
Borisjuk, L., Walenta, S., Weber, H., Mueller-Klieser, W. and Wobus,
U. (1998) High-resolution histographical mapping of glucoseconcentrations in developing cotyledons of Vicia faba in relationto mitotic activity and storage processes: glucose as a possibledevelopmental trigger. Plant J. 15, 583–591.
Borisjuk, L., Rolletschek, H., Radchuk, R., Weschke, W., Wobus, U.
and Weber, H. (2004) Seed development and differentiation: arole for metabolic regulation. Plant Biol. 6, 375–386.
de Veylder, L., de Almeida Engler, J., Burssens, S., Manevski, A.,
Lescure, B., Van Montagu, M., Engler, G. and Inze, D. (1999)A new D-type cyclin of Arabidopsis thaliana expressed duringlateral root primordial formation. Planta, 208, 453–462.
Filipowicz, W. and Pogacic, V. (2002) Biogenesis of small nucleolarribonucleoproteins. Curr. Opin. Cell Biol. 14, 319–327.
Fromont-Racine, M., Senger, B., Saveanu, C. and Fasiolo, F. (2003)Ribosome assembly in eukaryotes. Gene, 313, 17–42.
Fujiwara, S., Matsuda, N., Sato, T., Sonobe, S. and Maeshima, M.
(2002) Molecular properties of a matrix attachment region-bind-ing protein located in the nucleoli of tobacco cells. Plant CellPhysiol. 43, 1558–1567.
Ginisty, H., Sicard, H., Roger, B. and Bouvet, P. (1999) Structure andfunctions of nucleolin. J. Cell Sci. 12, 761–772.
Gulli, M.P., Girard, J.P., Zabetakis, D., Lapeyre, B., Melese, T. and
Caizergues-Ferrer, M. (1995) gar2 is a nucleolar protein fromSchizosaccharomyces pombe required for 18S rRNA and 40Sribosomal subunit accumulation. Nucleic Acids Res. 23, 1912–1918.
Huddleson, J.P., Ahmad, N. and Lingrel, J.B. (2006) Upregulation ofthe KLF2 transcription factor by fluid shear stress requiresnucleolin. J. Biol. Chem. 281, 15121–15128.
Ito, T., Kim, G.T. and Shinozaki, K. (2000) Disruption of an Arabid-opsis cytoplasmic ribosomal protein S13-homologous gene bytransposon mutagenesis causes aberrant growth and develop-ment. Plant J. 22, 257–264.
Jang, J.C., Leon, P., Zhou, L. and Sheen, J. (1997) Hexokinase as asugar sensor in higher plants. Plant Cell, 9, 5–19.
Kim, K., Dimitrova, D.D., Carta, K.M., Saxena, A., Daras, M. and
Borowiec, J.A. (2005) Novel checkpoint response to genotoxicstress mediated by nucleolin–replication protein A complex for-mation. Mol. Cell. Biol. 25, 2463–2474.
Koch, K.E. (1996) Carbohydrate-modulated gene expression inplants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 509–540.
Kondo, K. and Inouye, M. (1992) Yeast NSR1 protein that hasstructural similarity to mammalian nucleolin is involved in pre-rRNA processing. J. Biol. Chem. 267, 16252–16258.
Li, Y., Lee, K.H., Walsh, S., Smith, C., Hadingham, S., Sorefan, K.,
Cawley, G. and Bevan, M.W. (2006) Establishing glucose- andABA regulated transcription networks in Arabidopsisby microar-ray analysis and promoter classification using a Relevance VectorMachine. Genome Res. 16, 414–427.
Lorenz, S., Tintelnot, S., Reski, R. and Decker, E.L. (2003)Cyclin D-knockout uncouples developmental progression fromsugar availability. Plant Mol. Biol. 53, 227–236.
Maceluch, J., Kmieciak, M. and Szweykowska-Kulinska, Z. (2001)Cloning and characterization of Arabidopsis thaliana AtNAP57 – ahomolog of yeast pseudouridine synthase Cbf5p. Acta Bio-chim.Pol. 48, 699–709.
1062 Hisae Kojima et al.
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063

Masumi, A., Fukazawa, H., Shimazu, T., Yoshida, M., Ozato, K.,
Komuro, K. and Yamaguchi, K. (2006) Nucleolin is involved ininterferon regulatory factor-2-dependent transcriptional activa-tion. Oncogene, 25, 5113–5124.
Matsuoka, K. and Nakamura, K. (1991) Propeptide of a precursor toa plant vacuolar protein required for vacuolar targeting. Proc. NatlAcad. Sci. USA, 88, 834–838.
Meier, U.T. (2005) The many facets of H/ACA ribonucleoproteins.Chromosoma, 114, 1–14.
Menand, B., Desnos, T., Nussaume, L., Berger, F., Bouchez, D.,
Meyer, C. and Robaglia, C. (2002) Expression and disruption ofthe Arabidopsis TOR (target of rapamycin) gene. Proc. Natl Acad.Sci. USA, 99, 6422–6427.
Mita, S., Suzuki-Fujii, K. and Nakamura, K. (1995) Sugar-inducibleexpression of a gene for b-amylase in Arabidopsis thaliana. PlantPhysiol. 107, 895–904.
Muller, J., Wiemken, A. and Aeschbacher, R. (1999) Trehalosemetabolism in sugar sensing and plant development. Plant Sci.147, 37–47.
Murray, J.A.H., Freeman, D., Greenwood, J. et al. (1998) PlantD cyclins and retinoblastoma protein homologues. In Plant CellDivision (Francis, D., Dudits, D. and Inze, D., eds). London: Port-land Press, pp. 99–127.
Nasirudin, K.M., Ehtesham, N.Z., Tuteja, R., Sopory, S.K. and
Tuteja, N. (2005) The Gly-Arg-rich C-terminal domain of peanucleolin is a DNA helicase that catalytically translocates in the5¢- to 3¢-direction. Arch. Biochem. Biophys. 434, 306–315.
Nishimura, T., Wada, T., Yamamoto, K.T. and Okada, K. (2005) TheArabidopsis STV1 protein, responsible for translation reinitiation,is required for auxin-mediated gynoecium patterning. Plant Cell,17, 2940–2953.
Oh, S.A., Lee, S.Y., Chung, I.K., Lee, C.H. and Nam, H.G. (1996) Asenescence-associated gene of Arabidopsis thaliana is distinc-tively regulated during natural and artificially induced leaf sen-escence. Plant Mol. Biol. 30, 739–754.
Pendle, A.F., Clark, G.P., Boon, R., Lewandowska, D., Lam, Y.W.,
Andersen, J., Mann, M., Lamond, A.I., Brown, J.W. and Shaw,
P.J. (2005) Proteomic analysis of the Arabidopsis nucleolus sug-gests novel nucleolar functions. Mol. Biol. Cell, 16, 260–269.
Pih, K.T., Yi, M.J., Liang, Y.S., Shin, B.J., Cho, M.J., Hwang, I. and
Son, D. (2000) Molecular cloning and targeting of a fibrillarinhomolog from Arabidopsis. Plant Physiol. 123, 51–58.
Price, J., Laxmi, A., St. Martin, S.K. and Jang, J.-C. (2004) Globaltranscription profiling reveals multiple sugar signal transductionmechanisms in Arabidopsis. Plant Cell, 16, 2128–2150.
Raught, B., Gingras, A.-C. and Sonenberg, N. (2001) The target ofrapamycin (TOR) proteins. Proc. Natl Acad. Sci. USA, 98, 7037–7044.
Reichler, S.A., Balk, J., Brown, M.E., Woodruff, K., Clark, G.B. and
Roux, S.J. (2001) Light differentially regulates cell division andthe mRNA abundance of pea nucleolin during de-etiolation. PlantPhysiol. 125, 339–350.
Riou-Khamlichi, C., Menges, M., Healy, J.M.S. and Murray, J.A.H.
(2000) Sugar control of the plant cell cycle: differential regulation
of Arabidopsis D-type cyclin gene expression. Mol. Cell. Biol. 20,4513–4521.
Rohde, J.R. and Cardenas, M.E. (2003) The Tor pathway regulatesgene expression by linking nutrient sensing to histone acetyla-tion. Mol. Cell. Biol. 23, 629–635.
Rolland, F., Baena-Gonzalez, E. and Sheen, J. (2006) Sugar sensingand signaling in plants: conserved and novel mechanisms. Annu.Rev. Plant Biol. 57, 675–709.
Rook, F., Weisbeek, P. and Smeekens, S. (1998) The lightregulatedArabidopsis bZIP transcription factor gene ATB2 encodes a pro-tein with an unusually long leucine zipper domain. Plant Mol.Biol. 37, 171–178.
Saez-Vasquez, J., Caparros-Ruiz, D., Barneche, F. and Echeverrıa,
M. (2004) A plant snoRNP complex containing snoRNAs, fibrilla-rin, and nucleolin-like proteins is competent for both rRNA genebinding and pre-rRNA processing in vitro. Mol. Cell. Biol. 24,7284–7297.
Schmelzle, T. and Hall, M.N. (2000) TOR, a central controller of cellgrowth. Cell, 103, 253–262.
Srivastava, M. and Pollard, H.B. (1999) Molecular dissection ofnucleolin’s role in growth and cell proliferation: new insights.FASEB J. 13, 1911–1922.
Tong, C.G., Reichler, S., Blumenthal, S., Balk, J., Hsieh, H.L. and
Roux, S.J. (1997) Light regulation of the abundance of mRNAencoding a nucleolin-like protein localized in the nucleoli of peanuclei. Plant Physiol. 114, 643–652.
Tsang, C.K., Bertram, P.G., Ai, W., Drenan, R. and Zheng, X.F.S.
(2003) Chromatin-mediated regulation of nucleolar structure andRNA polymerase I localization by TOR. EMBO J. 22, 6045–6056.
Tuteja, R. and Tuteja, N. (1998) Nucleolin: a multifunctional majornucleolar phosphoprotein. Crit. Rev. Biochem. Mol. Biol. 33, 407–436.
van Lijsebettens, M., Vanderhaeghen, R., de Block, M., Bauw, G.,
Villarroel, R. and van Montagu, M. (1994) An S18 ribosomalprotein gene copy at the Arabidopsis PFL locus affects plantdevelopment by its specific expression in meristems. EMBO J. 13,3378–3388.
Weijers, D., Franke-van Dijk, M., Vencken, R.J., Quint, A., Hooykaas,
P. and Offringa, R. (2001) An Arabidopsis Minute-like phenotypecaused by a semi-dominant mutation in a RIBOSOMAL PROTEINS5 gene. Development, 128, 4289–4299.
Wingler, A., Fritzius, T., Wiemken, A., Boller, T. and Aeschbacher,
R.A. (2000) Trehalose induces the ADP-glucose pyrophosphory-lase gene, ApL3, and starch synthesis in Arabidopsis. PlantPhysiol. 124, 105–114.
Yamada, H., Koizumi, N., Nakamichi, N., Kiba, T., Yamashino, T. and
Mizuno, T. (2004) Rapid response of Arabidopsis T87 culturedcells to cytokinin through His-to-Asp phosphorelay signal trans-duction. Biosci. Biotechnol. Biochem. 68, 1966–1976.
Yoine, M., Ohto, M., Onai, K., Mita, S. and Nakamura, K. (2006)The lba1 mutation of UPF1 RNA helicase involved in nonsense-mediated mRNA decay causes pleiotropic phenotypic chan-ges and altered sugar signaling in Arabidopsis. Plant J. 47, 49–62.
Sugar-inducible expression of nucleolin gene 1063
ª 2006 The AuthorsJournal compilation ª 2007 Blackwell Publishing Ltd, The Plant Journal, (2007), 49, 1053–1063
Top Related
Copyright © 2022 FDOKUMEN

