







Bahasa
Halaman
Hukum


www.elsevier.com/locate/yhbeh
Hormones and Behavior
Social cues and hormone levels in male Octodon degus (Rodentia): a field
test of the Challenge Hypothesis
Mauricio Soto-Gamboaa,*, Manuel Villalonb, Francisco Bozinovicc
aInstituto de Zoologıa, Facultad de Ciencias, Universidad Austral de Chile, Casilla (Box) 567, Valdivia, ChilebUnidad de Reproduccion y Desarrollo, Departamento de Ciencias Fisiologicas, Facultad de Ciencias Biologicas,
Pontificia Universidad Catolica de Chile, ChilecCenter for Advanced Studies in Ecology and Biodiversity and Departamento de Ecologıa, Facultad de Ciencias Biologicas,
Pontificia Universidad Catolica de Chile, Chile
Received 21 March 2004; revised 28 October 2004; accepted 1 November 2004
Available online 16 January 2005
Abstract
Social interactions are important factors determining and regulating individual behaviors. Testosterone has been related to agonistic
interactions, while glucocorticoids have been related to social stress, especially during interactions of dominance. We compared testosterone
and cortisol concentrations in male degus (Octodon degus, Rodentia) under laboratory conditions without male social interactions, with data
from wild males in nature. Under natural conditions, males should present higher levels of testosterone during the breeding season due to
social interactions (Challenge Hypothesis). Alternatively, intense social instability could act as a stressing environment, raising
glucocorticoids, which inhibit testosterone concentrations. Our results show a significant increase in agonistic interactions between males
during the breeding season, and disappearance of non-agonistic male interactions during this period. Hormone levels in breeding season show
nonsignificant differences between laboratory groups, but testosterone concentrations in field males were significantly higher than in
laboratory males. Testosterone levels were similar among pre-breeding and breeding periods, but in field animals the concentration was ~30%
higher than in laboratory degus. In field animals, we found two different mating strategies: resident males, with territorial behavior, and
transient males, displayed an opportunistic approach to females. Finally, cortisol presents a similar pattern in both laboratory and field
animals; pre-breeding values of cortisol are higher than during the breeding season. This suggests that social interactions in O. degus activate
a rise in testosterone, supporting the Challenge Hypothesis, and could be considered as partial support of the Social Stress Hypothesis.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Testosterone; Cortisol; Social interaction; Social stress; Social instability; Challenge Hypothesis; Octodon degus; Degu
Introduction
Social interactions are an important factor determining
and modulating individual behavior and performance. In
many cases, individual behavior is directly dependent on
relationships with other individuals that live together or in
the neighborhood (Clutton-Brock et al., 2001; Conradt and
Roper, 2000; Kappeler, 1999). These interactions include
agonistic and non-agonistic behaviors that may determine
0018-506X/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.yhbeh.2004.11.010
* Corresponding author. Fax: +56 63 221315.
E-mail address: [email protected] (M. Soto-Gamboa).
social position, dominance, and hierarchy relations between
individuals within social groups (Clarke and Faulkes, 2001;
Cote, 2000; Smale and Holekamp, 1997), and in extreme
social situations may affect individual reproductive success
(Clarke and Bennett, 2001; Clutton-Brock et al., 2001;
Raouf et al., 1997; Sherman et al., 1995; Stoehr and Hill,
2000; Zielinski and Vandenbergh, 1993). Nevertheless,
individual behavior also has an important physiological
component, especially hormonal regulation, which can
determine the successful acquisition of a social position in
the hierarchy or access to reproductive females (Alberts et
al., 1992; Buck and Barnes, 2003; Clarke and Faulkes,
1998; Crews, 1997; Randall and Bromage, 1998; Raouf et
47 (2005) 311–318

M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318312
al., 1997). In this sense, social interactions and hormonal
regulation are narrowly related and determine behavioral
patterns of the individual (Crews, 1997).
Among vertebrates, testosterone has several principal
functions in the physiology of an organism. It is
responsible for spermatogenesis and some secondary sex
characters as well as the activation of male sexual
behaviors (Blottner et al., 2000; Nelson, 2000; Wingfield
et al., 1997). During the reproductive period, testosterone
has direct effects on mating behaviors including attractive-
ness to females and on males’ interactions for access to
females (Cavigelli and Pareira, 2000; Dunlap et al., 2002;
Emerson, 1997; Goodson and Bass, 2001; Sinervo et al.,
2000; Wingfield et al., 1990, 1994). The Challenge
Hypothesis was originally proposed for bird species
(Wingfield et al., 1990), and predicts that androgen levels
and aggression are positively related, and are closely
associated with social context. An increase in male
interactions activates testosterone secretion and, conse-
quently, increases aggressive behaviors (Wingfield et al.,
1997). The degree of increase in testosterone levels and the
magnitude of agonistic response depend on the degree of
social stimulus (Cavigelli and Pareira, 2000; Wingfield et
al., 1990). Among mammals, male interactions and
testosterone regulation of aggressive behavior have
recently been studied (Cavigelli and Pareira, 2000; Lynch
et al., 2002; Muller and Wrangham, 2004; Ostner and
Kappeler, 2002). In these cases, there is a positive
correlation between male aggression and testosterone
levels during the reproductive period. In rodents, testoster-
one levels have been associated with territory defense,
reproductive group formation, social dominance, and
hierarchy formation (Cavigelli and Pareira, 2000; Clarke
and Faulkes, 1998; Faulkes and Abbott, 1997; Rogovin et
al., 2003; Wolff, 1994; Zielinski and Vandenbergh, 1993).
In contrast, social interactions can also act as important
stressing agent (Rogovin et al., 2003). The physiological
cost of reproduction associated with the cost of dominance
behavior often results in a stressing situation (Ketterson and
Nolan, 1999; Sapolsky, 2002). During the breeding season,
males compete for access to females and continually express
intense aggressive or agonistic interactions. These inter-
actions determine social status and, consequently, the
reproductive success of animals. Finally, these relationships
directly increase energetic demand and glucocorticoid levels
(Goymann and Wingfield, 2004). As a consequence of these
behavioral interactions, individuals reach high levels of
glucocorticoids, via the activation of the hypothalamic–
pituitary–adrenal axis (HPA). In extreme cases (chronic
stress), glucocorticoids can produce a decrease in testoster-
one secretion by the hypothalamic–pituitary–gonadal (HPG)
axis inhibition, thus affecting an individual’s social status in
the group (Blanchard et al., 2001; Knapp and Moore, 1997).
In this sense, social interactions can produce two different
types of hormonal outcomes: (1) enhancement of testoster-
one concentrations and consequent aggressive displays, and
(2) an increase in glucocorticoids via HPA axis suppressing
the HPG axis, and diminished testosterone levels. The
objective of this work is to evaluate how social interaction
levels affect hormonal levels of males, and how these have
direct repercussions on the behavioral patterns in male
degus.
The degu, Octodon degus, is a social caviomorph rodent
endemic to central Chile. This semi-fossorial, diurnal, and
herbivorous rodent (~180 g), lives in groups, but only
presents a structured social system during the breeding
season (Soto-Gamboa, unpublished data; Vasquez, 1997).
During this period, O. degus have been observed to form
polygynous groups consisting in one male and one to three
females (Ebensperger et al., 2004; Fulk, 1976). Previous
laboratory work reported a direct relationship between
male’s aggression and testosterone levels only during the
breeding period (Farfan et al., 2001). In this study, we
compare variation of testosterone and cortisol levels in
captive males without and with low social interactions with
field animals in the pre-breeding and breeding seasons. We
hypothesized that social interactions have positive effect on
testosterone levels enhancing aggressive behavior during the
breeding season, i.e., the Challenge Hypothesis. Alterna-
tively, intense social instability could be a stressing environ-
ment, raising glucocorticoids, which ultimately inhibit
testosterone concentrations, i.e., social stress hypothesis.
These hypotheses are not mutually exclusive, but the
relative importance of variation in hormonal changes
depends on the magnitude of social interactions.
Methods
To test social interactions, we divided the experimen-
tal design into two different approaches. The field Study
includes behavioral patterns of activity and testosterone
and cortisol levels. These data were compared with
laboratory animals separated into virgin (without social
interaction) and reproductive males (only with female
social interaction).
Field study
We studied a natural population of O. degus in
Rinconada de Maipu (33831VS, 70850VW), a field station
of the Universidad de Chile, located near Santiago, in
Central Chile. In April and June–July of 2002, we captured
animals using a capture effort of 100 Sherman live traps
during 5 days. All captured animals were sexed, weighed,
and marked with a uniquely colored collar ring. For later
hormone analysis (see below), we took blood samples from
all captured males (approximately 700 Al from the
suborbital sinus with a heparinized glass pipette). To avoid
manipulation effects, the traps were checked every half
hour, following Place and Kenagy (2000), and based on
preliminary data that degus do not present significant

M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318 313
variation in testosterone and glucocorticoid levels when
animals were maintained between 0 and 2 h inside the traps
(K.S. Matt and F. Bozinovic, unpublished data). The
handling time for blood sampling was never over 30 s,
and was obtained immediately after animals were removed
from the traps. In total, we got blood samples from 9 males
in April and from 12 males in June–July.
During the 5 days after capture, we recorded behavioral
data through direct observations, consisting of 15 min of
focused male observation, registering all interactions with
other animals. In total, we registered 20 focal observations
per period, during which we identified all animals interact-
ing with the focal animal. The interaction rate was classified
using four different classes based on behavioral repertoires
previously described by Fulk (1976): (1) agonistic inter-
actions between males, including chases, mounting and
foreleg pushing; (2) non-agonistic interactions between
males, including allogrooming, nose contact (olfaction),
and group foraging; (3) agonistic interactions of males
towards females, including chases and foreleg pushing; and
(4) non-agonistic interactions of males towards females,
including allogrooming, nose contact (olfaction), and group
foraging. Rates of interaction were calculated for individual
animals and represent a percentage of the total interactions
for a particular male.
Finally, we classified males as resident or transient in
function of their capacity to defend territories and monop-
olize females. This differentiation was made in order to
compare hormone levels of animals with high (resident) and
low (transient) rates of social interactions. During breeding
period, we recognized a total of five resident males and
seven transient male (in total 12 animals).
Laboratory study
For our experimental study, we used males born in the
laboratory by parents from a native population from Lampa
(33817VS, 70853VW), near Santiago, in Central Chile.
Animals were separated from their mother at 2 months of
age (weaning), and were maintained in individual cages.
Photoperiod was maintained on a D:L 12:12-h cycle with
controlled temperature between 15 and 258C following the
night–day cycle. Rabbit food (ChampionR) pellets and
water were offered ad libitum.
Males were randomly separated into two different
groups: virgin males and reproductive males. Animals in
the first group were constantly maintained in individual
cages, while in the second group, males were enclosed with
two or three females to establish reproductive families
during the breeding period. Reproductive groups were
utilized to evaluate the effect of male–female interactions
on circulating testosterone levels. In this group, all males
were effective as reproductive animals and in most families
at least one female was pregnant. We obtained blood
samples from a total of 30 individuals: 10 virgin males in
the pre-breeding period (April 2003), 10 virgin males in the
breeding period, and 10 reproductive males with females
during the breeding period (June–July 2003). Similar to
wild animals, for blood sampling, handling time was never
over 30 s. This protocol was used to compare non-
reproductive and reproductive values of hormones in virgin
(single) males, and in males interacting with females and to
compare hormone results between wild and laboratory
animals. In this way, we designed different levels of social
interactions: virgin (without social interactions), reproduc-
tive males (only with females interactions), wild transients
males (with low male–male interactions), and resident males
(with highest levels of social interaction, including female
and male interactions).
Steroid assays
Blood samples were transported to the laboratory in a
refrigerated cooler (not more than 8 h after being taken) and
centrifuged at 7000 rpm for 10 min. Plasma was separated
from blood cells and stored at �208C for latter hormone
assays. Testosterone and cortisol concentrations were
measured by enzyme immunoassay with reagents supplied
by the World Health Organization program for the Provision
of Matched Reagents for RIA of Hormone in Reproductive
Physiology, and following their recommended procedures
(Sufi et al., 1998). For testosterone determination, we
extracted the hormone with diethyl-ether, evaporating 50
Al of plasma, and resuspending in RIA assay buffer. For
cortisol determination, we diluted 25 Al of plasma sample in
500 Al of RIA buffer solution, and took 20 Al of this solutionfor protein denaturation to 608C. These methods had
resolution limits for testosterone of 0.12 ng/ml and 17.8
ng/ml for cortisol. Finally, all samples were analyzed in
duplicate, and the precision of the assay was evaluated by
determining the intra- and interassay coefficient of variation
(CV). Intra- and interassay CV for testosterone were 5.3%
and 9.3%, and for cortisol, 8.3% and 11.7%, respectively.
Statistical analyses
Statistical analyses were performed using parametric and
non-parametric statistical tests. To analyze behavioral
relationships, we used interaction rates expressed as a
percentage of total interactions displayed by an individual
male. We utilized a Mann–Whitney U test to evaluate
significant differences between breeding periods for each
interaction type at P b 0.05. For hormonal comparisons, we
utilized ANOVA. Normality of the data was evaluated using
the Kolmogorov–Smirnov test and homogeneity of variance
was evaluated with Levene’s test. In the seasonal compar-
ison, we utilized two-way ANOVA, using as factors
seasonality (pre-breeding and breeding) and treatment
(laboratory and field animals). During breeding season,
one-way ANOVA was performed to compare the different
classes of males. In the case of the testosterone data set, we
performed a log-transformation to meet assumptions (Zar,
M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318314
1996). An unequal Tukey Test was utilized for post hoc
analysis. All statistical analyses were performed using
Statistica 6.0 software (StatSoft, 2001). Results are pre-
sented as means F 1 standard error (SEM).
Ethical note
All experimental procedures in this work were carried
out under the approval of Pontificia Universidad Catolica de
Chile ethical committee and according to the current
Chilean law, under permit number SAG-698 of the Servicio
Agrıcola y Ganadero.
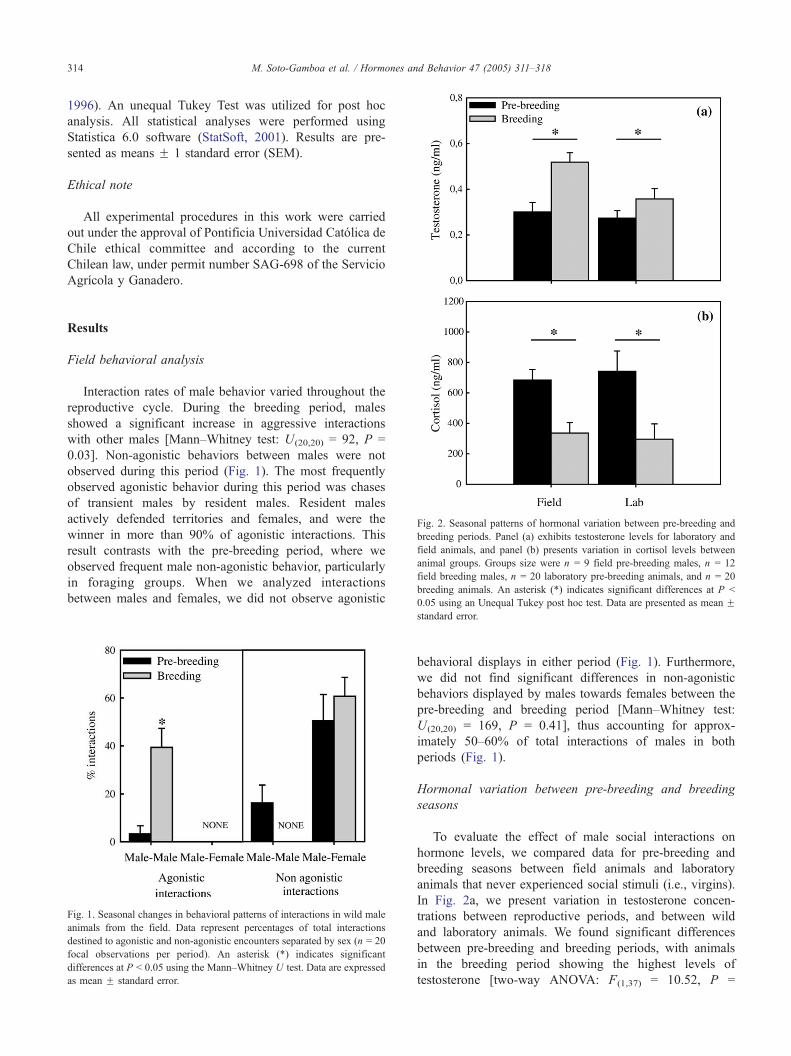
Fig. 2. Seasonal patterns of hormonal variation between pre-breeding and
breeding periods. Panel (a) exhibits testosterone levels for laboratory and
field animals, and panel (b) presents variation in cortisol levels between
animal groups. Groups size were n = 9 field pre-breeding males, n = 12
field breeding males, n = 20 laboratory pre-breeding animals, and n = 20
breeding animals. An asterisk (*) indicates significant differences at P b
0.05 using an Unequal Tukey post hoc test. Data are presented as mean F
Results
Field behavioral analysis
Interaction rates of male behavior varied throughout the
reproductive cycle. During the breeding period, males
showed a significant increase in aggressive interactions
with other males [Mann–Whitney test: U(20,20) = 92, P =
0.03]. Non-agonistic behaviors between males were not
observed during this period (Fig. 1). The most frequently
observed agonistic behavior during this period was chases
of transient males by resident males. Resident males
actively defended territories and females, and were the
winner in more than 90% of agonistic interactions. This
result contrasts with the pre-breeding period, where we
observed frequent male non-agonistic behavior, particularly
in foraging groups. When we analyzed interactions
between males and females, we did not observe agonistic
Fig. 1. Seasonal changes in behavioral patterns of interactions in wild male
animals from the field. Data represent percentages of total interactions
destined to agonistic and non-agonistic encounters separated by sex (n = 20
focal observations per period). An asterisk (*) indicates significant
differences at P b 0.05 using the Mann–Whitney U test. Data are expressed
as mean F standard error.
standard error.
behavioral displays in either period (Fig. 1). Furthermore,
we did not find significant differences in non-agonistic
behaviors displayed by males towards females between the
pre-breeding and breeding period [Mann–Whitney test:
U(20,20) = 169, P = 0.41], thus accounting for approx-
imately 50–60% of total interactions of males in both
periods (Fig. 1).
Hormonal variation between pre-breeding and breeding
seasons
To evaluate the effect of male social interactions on
hormone levels, we compared data for pre-breeding and
breeding seasons between field animals and laboratory
animals that never experienced social stimuli (i.e., virgins).
In Fig. 2a, we present variation in testosterone concen-
trations between reproductive periods, and between wild
and laboratory animals. We found significant differences
between pre-breeding and breeding periods, with animals
in the breeding period showing the highest levels of
testosterone [two-way ANOVA: F(1,37) = 10.52, P =

M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318 315
0.003]. On the other hand, there were no significant
differences between animal groups [two-way ANOVA:
F(1,37) = 2.79, P = 0.103] or significant interactions, i.e.,
both animal groups (field and lab) presented the same
pattern of variation in testosterone concentrations between
pre-breeding and breeding periods [two-way ANOVA:
F(1.37) = 0.501, P = 0.48]. Cortisol concentrations varied
only between pre-breeding and breeding periods (Fig. 2b),
where the pre-breeding period presented the highest
concentrations [two-way ANOVA: F(1.37) = 18.150, P =
0.0001], and there were no significant differences between
field and lab animals [two-way ANOVA: F(1,37) = 0.0852,
P = 0.772]. These results indicate that field animals have
the same cortisol concentrations patterns as laboratory
animals.
Hormone relationships during the breeding season
During the breeding season, field males are exposed to
intense social instability caused by interactions with other
males (Fig. 1), while laboratory animals never were
exposed to male interactions. Comparing the four different
classes of males, i.e., virgin laboratory males, reproductive
laboratory males, transient males, and resident males, we
Fig. 3. Hormonal levels of male degus during the breeding season. We
compare field animals separated in resident (n = 5) and transient (n = 7)
males with virgin (n = 10) and reproductive laboratory (n = 10) males.
Panel (a) presents variation in testosterone levels between groups, and panel
(b) presents variation in cortisol between groups. An asterisk (*) indicates
significant differences at P b 0.05 using ANOVA. Data are expressed as
mean F standard error.
found significant differences in testosterone levels [one-
way ANOVA F(3,38) = 5.96 P = 0.003]. Tukey post hoc
test showed that resident males had significantly higher
levels of testosterone than the other groups, and transient
males were not significantly difference than either
laboratory animal groups (Fig. 3a). Cortisol levels present
the same pattern. Exist significant differences between
male groups [one-way ANOVA F(3,28) = 3.92 P = 0.019].
Tukey post hoc test showed that resident males present
significant highest hormonal level than the other groups
(Fig. 3b).
Discussion
In this study, we experimentally manipulated the
social interactions in laboratory male degus and com-
pared their hormone levels with two different classes of
wild males (resident and transient). We found that both
groups of animals, laboratory and field males, have a
seasonal variation in hormonal levels. During the pre-
breeding season, animals show high levels of cortisol but
this diminished during breeding season. Contrary, testos-
terone levels show low values during pre-breeding
season, but increase significantly during breeding season.
Previous studies have not found seasonal variation in
testosterone levels of wild males (Kenagy et al., 1999).
These authors reported very low values of testosterone,
and few animals had detectable testosterone levels. This
makes interpretation of their results difficult. There is an
obvious contradiction between results from Kenagy et al.
(1999) and our data, which show a very clear pattern of
testosterone variation between pre-reproductive and
reproductive period, but the reason for this discrepancy
remains unexplained. Probably the very low levels of
testosterone observed by Kenagy and collaborators were
nearby the limit of detection.
The high levels of testosterone found during repro-
ductive periods among field males indicate that, under
natural conditions, there exist environmental factors that
directly affect testosterone levels. We found that social
agonistic interactions between males during this period
increase testosterone, concordantly with Challenge
Hypothesis. Particularly, resident males that are more
exposed to social interaction and present the highest
levels of testosterone, but have highest levels of cortisol
too. These results propose that resident males have two
different responses to social stimuli. First, increases in
testosterone by male–male interaction, increasing behav-
ioral agonistic displays, and second, increases in cortisol
levels in response to the social stress of defending the
territory and/or females. These increases of the levels of
cortisol also might be only an effect of the increase of
the energetic demands associated with the cost of the
defense of territory and/or females (Moore and Jessop,
2003).

M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318316
The Challenge Hypothesis
The Challenge Hypothesis claims a direct relationship
between temporal patterns of male testosterone levels and
mating system strategies (Wingfield et al., 1990). In
particular, testosterone secretion in polygynous species is
postulated to be directly related to male–male interactions
(Wingfield, 1984). Moreover, it is possible to distinguish
between baseline testosterone concentrations in the non-
reproductive season (i.e., the non-breeding baseline), the
reproductive season (i.e., the reproductive baseline), and the
transient surges of testosterone secretions induced by male–
male interactions (Muller and Wrangham, 2004; Wingfield
et al., 1990). Our results represent a complete synthesis of
this pattern. O. degus are polygynous social rodents where
males compete for female access to form reproductive
groups (Fulk, 1976). Our laboratory results represent the
physiological baseline during the pre-breeding season and
the reproductive baseline during the breeding season. It is
very interesting that testosterone levels in the pre-breeding
season are not different between laboratory and field males.
This suggests that both laboratory and field degus show a
similar endogenous pattern of their reproductive cycle, and
the same non-reproductive baseline of testosterone secre-
tions. In contrast, during the reproductive period, laboratory
animals represent the reproductive baseline of testosterone
levels, and have significantly higher testosterone levels than
animals observed in the pre-breeding period. The higher
testosterone levels observed in field reproductive males
could indicate that testosterone levels that are near their
maximum physiological levels could be due to the effects of
social interactions.
We were able to differentiate between two different
classes of field males: resident males with a defensible
female group, and transient males with reduced presence in
the study area (Clutton-Brock, 1989). It is very striking that
only resident males exhibited high levels of testosterone,
and they were also the animals that most frequently
presented agonistic behaviors during the breeding season,
while transient males exhibited the basal reproductive level,
thus maintaining their potential to breed. These results may
represent two different male reproductive strategies. First, at
the beginning of the breeding season, males actively
compete to monopolize females, and then quickly form
reproductive groups. This period of social instability
induces elevated testosterone levels in group-forming males
(i.e., resident males), while the males that are not able to
form groups lose their chance to breed in this season (i.e.,
transient males). Alternatively, transient males may use an
opportunistic strategy for female accessibility at low
physiological cost. Clearly, the defensible harem has high
physiological costs associated with it, to maintain the
exclusivity of females, this includes the increased metabolic
cost of maintaining high levels of testosterone (Ketterson
and Nolan, 1999; Ketterson et al., 1996; Wingfield et al.,
1997) and possible effects on immune system suppression
(Braude et al., 1999; Casto et al., 2001; Folstad and Karter,
1992; Wedekind and Folstad, 1994). To differentiate
between these alternatives, it is necessary to take a new
approach towards the study of mating systems in degus to
determine the real reproductive output of resident versus
transient males.
Finally, hormonal differences in male mating strategies
could be associated with other factors more than social
stimuli. Age, experience, energy storage, and health are
important factors than can affect directly hormonal levels
and reproductive success (Wingfield, 1994). In O. degus, it
has been demonstrated that the testosterone levels and
testicular development decrease in function of age and older
animal increase body mass (Bustos-Obregon and Ramirez,
1997). In our results, we did not find a significant difference
in body mass between different classes of males during
breeding season and all laboratory animals were of same age
(proximate 10 months). We cannot estimate the age of wild
males, and most probably, we include different cohorts in
our analysis.
Social stress
Our results indicate that male degus presented a similar
seasonal pattern of cortisol concentrations in both laboratory
and wild groups. During the pre-breeding season, cortisol
levels were higher than during the breeding season,
presumably following an endogenous rhythm. This affirma-
tion is supported by the fact that laboratory animals lack
environmental factors that may modulate cortisol variations.
Seasonal variations of glucocorticoids have been reported
for a variety of vertebrate taxa, but remain unclear for most
mammals (Romero, 2002). However, rodents exhibiting
clear patterns of seasonal variation have been reported (see
Boswell et al., 1994; Kenagy and Place, 2000; Place and
Kenagy, 2000), particularly degus (Kenagy et al., 1999). In
all of these cases, glucocorticoids decline during mating
season. What is the cause of this variation is unknown, but
the most likely explanation is that during the pre-breeding
season, food is scarce and animals must allocate more time
to feeding and storing energetic reserves for the breeding
period. This could be strictly associated with an increase of
glucocorticoids, thus stimulating foraging behavior and
energy mobilization for storage (Boswell et al., 1994; Green
et al., 1992; Romero, 2002; Sapolsky et al., 2000; Wingfield
and Kitaysky, 2002).
Field resident male degus showed different cortisol levels
in comparison to transient and laboratory groups of males.
While resident males presented the highest levels of
testosterone and cortisol, the others groups have low levels.
It is clear that social and physical stressors can cause an
increase in glucocorticoids, which can suppress male
reproduction (Boonstra and McColl, 2000), but in our
study, males exposed to increased social interactions (i.e.,
resident males) did not show hormonal signals of repro-
ductive suppression. Our results represent a transitional

M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318 317
situation in which resident males have intermediate levels of
stress, increased cortisol levels but not sufficient to inhibit
testosterone secretion. Recent studies of cooperative
breeders in the field show that dominant males can have
high levels of glucocorticoids as a result of the increased
cost of maintaining their dominance status and hierarchical
position (Boonstra and McColl, 2000; Creel, 2001; Creel et
al., 1996, 1997). Our results suggest that male degus have a
similar pattern, in which resident males experience a high
cost in terms of territorial defense and challenges with
transient animals. Finally, it is unclear what factors
determine resident or transient males, but they are affected
by age, body condition, health, or sensitivity to environ-
mental condition. These possibilities could be considered
for future approach to this problem.
In summary, our study of captive and free living male
degus has contributed new information regarding hormonal
regulation of behavior in group-living animals. In particular,
we found evidence to support the fundamental premises of
the Challenge Hypothesis, indicating that male degus are
directly affected by social interactions with other males.
However, in wild males, we observed two different
strategies (i.e., resident versus transient) for breeding males.
Both classes of males presented significant hormonal
differences possibly associated with behavioral strategies
of mate formation. These data could be partially supported
by the Social Stress Hypothesis since the high level of
cortisol in resident males did not have an effect on their
reproductive performance and testosterone levels.
Acknowledgments
We are grateful to Luis Ebensperger, Diego Urrejola, and
Marıa Jose Hurtado for field assistance. Special thanks to
John C. Wingfield and James G. Kenagy for critical
comments on the manuscript. Thanks are due to the
Universidad de Chile and Jose Daniel Garcıa (Field Station
Administrator) for access to the study area. This research
was funded by a CONICYT Doctoral Thesis Fellowship to
M. Soto-Gamboa and by FONDAP Grant 1501-001
program 1 to F. Bozinovic.
References
Alberts, S.G., Sapolsky, R.M., Altmann, J., 1992. Behavioral, endocrine,
and immunological correlates of immigration by an aggressive male
into a natural primate group. Horm. Behav. 26, 167–178.
Blanchard, R.J., McKittrick, C.R., Blanchard, D.C., 2001. Animal models
of social stress: effects on behavior and brain neurochemical systems.
Physiol. Behav. 73, 261–271.
Blottner, S., Rohleder, M., Zinke, O., Stuermer, I.W., 2000. Higher
testicular activity in laboratory gerbils compared to wild Mongolian
gerbils (Meriones unguiculatus). J. Zool. 250, 461–466.
Boonstra, R., McColl, C.J., 2000. Contrasting stress response of male
Arctic ground squirrels and red squirrels. J. Exp. Zool. 286, 390–404.
Boswell, T., Woods, S.C., Kenagy, G.J., 1994. Seasonal changes in body
mass, insulin and glucocorticoids of free-living golden-mantled ground
squirrels. Gen. Comp. Endocrinol. 96, 339–346.
Braude, S., Tang-Martinez, Z., Taylor, G.T., 1999. Stress, testosterone, and
the imunoredistribution hypothesis. Behav. Ecol. 10, 345–350.
Buck, C.L., Barnes, B.M., 2003. Androgen in free-living Arctic ground
squirrels: seasonal changes and influence of staged male–male
aggressive encounters. Horm. Behav. 43, 318–326.
Bustos-Obregon, E., Ramirez, O., 1997. Aging and testicular function in
Octodon degus. Andrologia 29, 319–326.
Casto, J.M., Nolan, V.J., Ketterson, E.D., 2001. Steroid hormones and
immune function: experimental studies in wild and captive dark-eye
Juncos. Am. Nat. 157, 408–420.
Cavigelli, S., Pareira, M., 2000. Mating season aggression and fecal
testosterone levels in male ring-tailed lemurs (Lemur catta). Horm.
Behav. 37, 246–255.
Clarke, F.M., Faulkes, C.G., 1998. Hormonal and behavioral correlates of
male dominance and reproductive status in captive colonies of the
naked mole-rat, Heterocephalus glaber. Proc. R. Soc. Lond., Ser. B
265, 1391–1399.
Clarke, F.M., Bennett, N.C., 2001. Family values: group dynamics and
social control of reproduction in African mole-rats. Trends Ecol. Evol.
16, 184–190.
Clarke, F.M., Faulkes, C.G., 2001. Intracolony aggression in the
eusocial naked mole-rat, Heterocephalus glaber. Anim. Behav. 61,
311–324.
Clutton-Brock, T.H., 1989. Mammalian mating systems. Proc. R. Soc.
Lond., Ser. B 236, 339–372.
Clutton-Brock, T.H., Brotherton, P.N.M., Russell, A.F., O’Riain, M.J.,
Gaynor, D., Kansky, R., Griffin, A., Manser, M., Sharpe, L., McIlrath,
G.M., Small, T., Moss, A., Monfort, S., 2001. Cooperation, control, and
concession in meerkat groups. Science 291, 478–481.
Conradt, L., Roper, T.J., 2000. Activity synchrony and social cohesion: a
fission–fusion model. Proc. R. Soc. Lond., Ser. B. 267, 2213–2218.
Cote, S.D., 2000. Dominance hierarchies in female mountain goats:
stability, aggressiveness and determinants of rank. Behaviour 137,
1541–1566.
Creel, S., 2001. Social dominance and stress hormones. Trends Ecol. Evol.
16, 491–497.
Creel, S., Creel, M.N., Monfort, S.L., 1996. Social stress and dominance.
Nature 379, 212.
Creel, S., Creel, N., Mills, M., Monfort, S., 1997. Rank and reproduction in
cooperatively breeding African wild dogs: behavioral and endocrine
correlates. Behav. Ecol. 8, 198–306.
Crews, D., 1997. Species diversity and the evolution of behavioral
controlling mechanisms. Ann. N. Y. Acad. Sci. 807, 1–21.
Dunlap, K.D., Pelczar, P.L., Knapp, R., 2002. Social interactions and
cortisol treatment increase the production of aggressive electrocommu-
nication signals in male electric fish, Apteronotus leptorhynchus. Horm.
Behav. 42, 97–108.
Ebensperger, L.A., Hurtado, M.J., Soto-Gamboa, M., Lacey, E., Chang,
A.T., 2004. Communal nesting and kinship in degus (Octodon degus).
Naturwissenschaften 91, 391–395.
Emerson, S.B., 1997. Testis size variation in frogs: testing the alternatives.
Behav. Ecol. Sociobiol. 41, 227–235.
Farfan, G., Ahumada, P., Larach, O., Cecchi, C., Sabat, P., Vasquez, R.M.,
2001. Relaciones entre conductas agresivas y niveles de testosterona
plasmatica en Octodon degus. I Reunion Binacional de Ecologıa, XX
Reunion Argentina de Ecologıa, X Reunion de la Sociedad de Ecologıa
de Chile, p. 265.
Faulkes, C.G., Abbott, D.H., 1997. The physiology of a reproductive
dictatorship: regulation of male and female reproduction by a single
breeding female in colonies of naked mole-rats. In: Solomon, N.G.,
French, J.A. (Eds.), Cooperative Breeding in Mammals. Cambridge
Univ. Press, Cambridge, pp. 302–334.
Folstad, I., Karter, J., 1992. Parasites, bright males, and the immunocom-
petence handicap. Am. Nat. 139, 603–622.

M. Soto-Gamboa et al. / Hormones and Behavior 47 (2005) 311–318318
Fulk, G.W., 1976. Notes on the activity, reproduction, and social behavior
of Octodon degus. J. Mammal. 57, 495–505.
Goymann, W., Wingfield, J.C., 2004. Allostatic load, social status and
stress hormones: the costs of social status matter. Anim. Behav. 67,
591–602.
Goodson, J.L., Bass, A.H., 2001. Social behavior functions and related
anatomical characteristics of vasotocin/vasopressin systems in verte-
brates. Brain Res. Rev. 35, 246–265.
Green, P.K., Wilkinson, C.W., Woods, S.C., 1992. Intraventricular cortico-
sterone increases the rate of body weight gain in underweight
adrenalectomized rats. Endocrinology 269, 269–275.
Kappeler, P.M., 1999. Primate socioecology: new insights from males.
Naturwisswnschaften 85, 18–29.
Kenagy, G.J., Place, N.J., 2000. Seasonal changes in plasma glucocorticoste-
roids of free-living female yellow-pine chipmunks: effects of reproduc-
tion and capture and handling. Gen. Comp. Endocrinol. 117, 189–199.
Kenagy, G.J., Place, N.J., Veloso, C., 1999. Relation of glucocorticoids and
testosterone to the annual cycle of free-living degus in semiarid central
Chile. Gen. Comp. Endocrinol. 115, 236–243.
Ketterson, E.D., Nolan, V.J., 1999. Adaptation, exaptation, and constraint: a
hormonal perspective. Am. Nat. 154, s4–s25.
Ketterson, E.D., Nolan, V.J., Cawthorn, M.J., Parker, P.G., Ziegenfus, C.,
1996. Phenotypic engineering: usinghormones to explore themechanistic
and functional bases of phenotypic variation in nature. Ibis 138, 70–86.
Knapp, R., Moore, M.C., 1997. Male morphs in tree lizards have different
testosterone responses to elevated levels of corticosterone. Gen. Comp.
Endocrinol. 107, 273–279.
Lynch, J.W., Ziegler, T.E., Strier, K.B., 2002. Individual and seasonal
variation in fecal testosterone and cortisol levels of wild male tufted
capuchin monkeys, Cebus apella nifritus. Horm. Behav. 41, 275–287.
Moore, I.T., Jessop, T.S., 2003. Stress, reproduction, and adrenocortical
modulation in amphibians and reptiles. Horm. Behav. 43, 39–47.
Muller, M.N., Wrangham, R.W., 2004. Dominance, aggression and
testosterone in wild chimpanzees: a test of the dChallenge hypothesisT.Anim. Behav. 67, 113–123.
Nelson, R.J., 2000. An Introduction to Behavioral Endocrinology. Sinauer
Associates Inc., Suderland, Massachusetts.
Ostner, J., Kappeler, P.M., 2002. Seasonal variation and social correlates of
androgen excretion in male redfronted lemurs (Eulemur fulvus rufus).
Behav. Ecol. Sociobiol. 52, 485–495.
Place, N.J., Kenagy, G.J., 2000. Seasonal changes in plasma testosterone
and glucocorticoids in free-living male yellow-pine chipmunks and
response to capture and handling. J. Comp. Physiol., B 170, 245–251.
Randall, C.F., Bromage, N.R., 1998. Photoperiod history determines
the reproductive response of rainbow trout to changes in daylength.
J. Comp. Physiol., A 183, 651–660.
Raouf, S.A., Parker, P.G., Ketterson, E.D., Nolan, V.J., Ziegenfus, C., 1997.
Testosterone affects reproductive success by influencing extra-pair
fertilizations in male dark-eyed juncos. Proc. R. Soc. Lond., Ser. B 264,
1599–1603.
Rogovin, K., Randall, J.A., Kolosova, I., Moshkin, M., 2003. Social
correlates of stress in adult males of the great gerbil, Rhombomys
opimus, in years of high and low population densities. Horm. Behav.
43, 132–139.
Romero, L.M., 2002. Seasonal changes in plasma glucocorticoid
concentrations in free-living vertebrates. Gen. Comp. Endocrinol.
128, 1–24.
Sapolsky, R.M., 2002. Endocrinology of the stress-response. In:
Becker, J.B., Breedlove, S.M., Crews, D., McCarthy, M.M. (Eds.),
Behavioral Endocrinology. Massachusetts Institute of Technology,
Cambridge, pp. 409–449.
Sapolsky, R.M., Romero, L.M., Munk, A.U., 2000. How do glucocorticoids
influence stress responses? Integrating permissive, suppressive, stim-
ulatory, and preparative actions. Endocr. Rev. 21, 55–89.
Sherman, P.W., Lacey, E.A., Reeve, H.K., Keller, L., 1995. The eusociality
continuum. Behav. Ecol. 6, 102–108.
Sinervo, B., Miles, D.B., Frankino, W.A., Klukowski, M., DeNardo, D.F.,
2000. Testosterone, endurance, and Darwinian fitness: natural selection
and sexual selection on the physiological bases of alternative male
behaviors in side-Blotched lizards. Horm. Behav. 38, 222–233.
Smale, L., Holekamp, K.E., 1997. Sexually dimorphic dispersal in
mammals: patterns, causes, and consequences. Adv. Study Behav. 26,
181–250.
StatSoft, 2001. STATISTICA (data analysis software system). Version 6.
http://www.statsoft.com.
Stoehr, A.M., Hill, G.E., 2000. Testosterone and the allocation of
reproductive effort in male house finches (Carpodacus mexicanus).
Behav. Ecol. Sociobiol. 48, 407–411.
Sufi, S., Donaldson, A., Jeffcoate, S., 1998. WHO matched reagent
progamme. Steroid Radioimmunoassy Method Manual. ONU, London,
pp. 1–65.
Vasquez, R.M., 1997. Vigilance and social foraging in Octodon degus
(Rodentia: Octodontidae) in central Chile. Rev. Chil. Hist. Nat. 70,
557–563.
Wedekind, C., Folstad, I., 1994. Adaptative and nonadaptative immuno-
suppression by sex hormones? Am. Nat. 143, 938.
Wingfield, J.C., 1984. Androgens and mating systems: testosterone-induced
polygyny in normally monogamous birds. Auk 101, 665–671.
Wingfield, J.C., Kitaysky, A.S., 2002. Endocrine responses to unpredictable
environmental events: stress or anti-stress hormones? Integr. Comp.
Biol. 42, 600–609.
Wingfield, J.C., Hegner, R.E., Dufty Jr., A.M., Ball, G.F., 1990. The
bChallenge hypothesisQ: theoretical implications for patterns of testos-
terone secretion, mating systems, and breeding strategies. Am. Nat. 136,
829–846.
Wingfield, J.C., Whaling, C.S., Marler, P., 1994. Communication in
vertebrates aggression and reproduction: the role of hormones. In:
Knobil, E., Neill, J.D. (Eds.), The Physiology of Reproduction. Raven
Press, Ltd., New York, pp. 303–342.
Wingfield, J.C., Jacobs, J., Hillgarth, N., 1997. Ecological constraints and
the evolution of hormone–behavior interrelationships. Ann. N. Y. Acad.
Sci. 807, 22–41.
Wolff, J.O., 1994. Reproductive success of solitary and communally nesting
white-footed mice and deer mice. Behav. Ecol. 5, 206–209.
Zar, J.H., 1996. Bioestatistical Analysis. Prentice Hall, New Jersey.
Zielinski, W.J., Vandenbergh, J.G., 1993. Testosterone and competitive
ability in male house mice, Mus musculus: laboratory and field studies.
Anim. Behav. 45, 873–891.
Top Related
Copyright © 2022 FDOKUMEN

