







Bahasa
Halaman
Hukum


Sph
JM
CC
Os
D
S
P
ItFs
M
Rtrsdms
Cb(
Key Words: Human fertilization failures, microtubules, FISH, human oocyte, immunocytochemistry
tipisetoasl
untsvufwpts
Rr4StS(T(Ag(RRCaDU3P8ja
aDUb
FaUC
FERTILITY AND STERILITY�VOL. 82, NO. 6, DECEMBER 2004
Copyright ©2004 American Society for Reproductive MedicinePublished by Elsevier Inc.
Printed on acid-free paper in U.S.A.
0d0
1
eceived January 7, 2004;evised and accepted May, 2004.upported by a grant from
he Portuguese Nationalcience Foundation
Fundação para a Ciência eecnologia, FCT) to J. R.-S.
POCTI/ESP/38049/2001)..A. is supported by a Ph.D.rant from FCT, Portugal
SFRH/BD/12665/2003).eprint requests: Joãoamalho-Santos, Ph.D.,enter for Neurosciencend Cell Biology,epartment of Zoology,niversity of Coimbra,004-517 Coimbra,ortugal (FAX: � 351-239-26798; E-mail:
[email protected]).Center for Neurosciencend Cell Biology,epartment of Zoology,niversity of Coimbra.Department of Maternal-etal Medicine, Geneticsnd Human Reproduction,
udip
niversity Hospitals ofoimbra.
015-0282/04/$30.00oi:10.1016/j.fertnstert.2004.5.086
654
imultaneous analysis of cytoskeletalatterns and chromosome positioning inuman fertilization failures
oão Ramalho-Santos, Ph.D.,a Alexandra Amaral,a Raquel Brito,b
ariana Freitas, M.Sc., a and Teresa Almeida Santos, M.D., Ph.D.b
enter for Neuroscience and Cell Biology, University of Coimbra; and University Hospitals of Coimbra,oimbra, Portugal
bjective: To sequentially and reliably apply both tubulin immunocytochemistry (ICC) and fluorescence initu hybridization (FISH) to human fertilization failures, thus providing a tool for a multiple analysis of arrest.
esign: Analysis of human fertilization failures at several stages of arrest.
etting: Academic and clinical institutions.
atient(s): Consenting patients undergoing assisted reproduction technologies.
ntervention(s): Failed fertilizations displaying signs of activation without pronuclear development, or withhe absence of polar body emission or cleavage 48 hours after insemination or microinjection were analyzed.ertilization failures were fixed and processed for ICC. After data was collected the same samples were thenubjected to FISH analysis using probes for chromosomes X, Y, and 18.
ain Outcome Measure(s): Simultaneous ICC and FISH data on the same sample.
esult(s): Sequential application of straightforward standard ICC and FISH techniques was not possible, ashe morphologic features had been altered, microtubular patterns were not preserved, and many samples wereendered opaque. Only chromatin at the cell surface or outside the oocyte/zygote, such as metaphase IIpindles or polar body nuclei, could be routinely probed for FISH after ICC. However, an increase inetergent-induced sample permeabilization as well as the removal of several steps usually performed for FISHade it possible to directly compare microtubular patterns and chromosome position, regardless of chromatin
tatus.
onclusion(s): Analysis of specific proteins by immunocytochemistry and of chromosome status/positioningy FISH can be carried out sequentially in human fertilization failures, irrespective of the stage of arrest.Fertil Steril� 2004;82:1654–9. ©2004 by American Society for Reproductive Medicine.)
idO
By definition, analysis of failed human fer-ilization takes place after the fact, by evaluat-ng oocytes/zygotes that failed to developroperly after IVF or intracytoplasmic spermnjection (ICSI) or were otherwise deemed un-uitable for embryo transfer. This leads to sev-ral limitations, not least of which is the facthat appropriate controls (i.e., normally devel-ping embryos) are rarely, if ever, available fornalysis. On the other hand, technical con-traints often imply that a sample that is ana-yzed using a particular approach is renderedseless for further procedures, which requireifferent preparatory steps. This is particularlymportant for two reasons. First, the intrinsic
reciousness of the material as well as the iniqueness of any given sample limits theumber of statistically significant experimentshat can be done in a given laboratory/clinicaletting, especially one without access to rele-ant animal models. Second, given that a fail-re in fertilization probably depends on severalactors (1), a multiparameter analysis, whichould probe a single oocyte/zygote for severalossible miscues, could be useful to determinehe causes of failure as well as to aid in theeparation of causes from consequences.
The use of tubulin immunofluorescence tonvestigate the status of the metaphase II spin-le in human oocytes is well established (2).n the other hand, fluorescence in situ hybrid-
zation (FISH) has also been an important tool

tfbdmcbhfaacmb
ncarttsitdetstb
ptTffbwar
phvtiam
pmg
B
ffvtabaih
TF
mfimi(sia
bbs0A(iwesDewe
ttX(c1Ccn
pbeD
F
o analyze chromosome status in human oocytes/fertilizationailures (3). The importance of proper microtubular assem-ly patterns during fertilization includes spindle formationuring meiosis, but also extends to pronuclear appositionediated by the sperm aster formed from the male-borne
entrosome and to the subsequent organization of the em-ryo’s mitotic apparatus (4, 5). Indeed, microtubular patternsave been used to pinpoint several stages of arrest in humanailed fertilizations (2). Such an analysis would be potenti-ted if information for chromosome status and position werevailable for the same sample, that is, if following immuno-ytochemistry (ICC) the same oocyte/zygote could be sub-itted to FISH without the original structural configuration
eing affected.
The sequential use of ICC and FISH has been described,otably employing both DNA probes and antibodies againsthromosome-associated proteins (6–9). This analysis haslso been extended to the evaluation of oocyte status, workemarkably pioneered in the mouse (10–12). The experimen-al approaches developed were used to tackle issues relatedo abnormal chromosome disjunction during meiosis, andhowed murine metaphase II spindles, with complementarynformation on chromosome position. Furthermore, observa-ions in human metaphase II spindles have also been brieflyescribed (5). It should be noted that cytoplasmic interfer-nce is not usually an issue in these experiments, given thathe cytoplasm has either been removed (chromosomepreads) or that the DNA has a peripheral, cortical, localiza-ion relative to the oocyte (metaphase II spindles, polarodies).
Simultaneous evaluation of human oocyte structures androteins by immunofluorescence is straightforward, providedhe individual protocols involve similar methodologies (13).he standard procedure, adapted to individual settings, calls
or formaldehyde/paraformaldehyde or methanol fixation,ollowed by Triton X-100 permeabilization/wash, and incu-ation of samples with the appropriate probes. However, ase describe, the most common protocols for ICC and FISH
re not very similar and can interfere with one another,endering some samples useless for analysis in the process.
Therefore, we report a simple method that can be em-loyed to analyze not only metaphase II spindles but alsouman failed fertilizations arrested at several stages of de-elopment using, in this case, both tubulin ICC for micro-ubular patterns and FISH for specific chromosome position-ng. The technique preserves the sample morphologyscertained by ICC, even after the harsher conditions nor-ally required for probe hybridization during FISH.
MATERIALS AND METHODSAll chemicals were obtained from Sigma Chemical Com-
any (St. Louis, MO) unless stated otherwise. All humanaterial was used in accordance with the appropriate ethics
uidelines provided by the University Hospitals of Coimbra. h
ERTILITY & STERILITY�
iologic MaterialThe human fertilization failures analyzed were obtained
rom the fertility clinic (University Hospitals of Coimbra)rom consenting patients undergoing fertility treatments in-olving both IVF and ICSI, following appropriate institu-ional review board guidelines. Failed fertilized oocytes werenalyzed that displayed signs of activation (second polarody extrusion) without pronuclear development or had anbsence of polar body emission or cleavage 48 hours afternsemination or microinjection. A total of 161 oocytes/faileduman fertilizations were available for this study.
ubulin Immunocytochemistry andluorescence in Situ HybridizationThe zona pellucida was removed with a short (1 to 5
inutes) incubation in acid Tyrode’s solution. Oocytes werexed for 1 hour with 2% (v/v) formaldehyde in Ferticultedium (FertiPro N.V., Beernem, Belgium) and permeabil-
zed in 5% Triton X-100 (v/v) in phosphate-buffered salinePBS; pH 7.2) for 1 to 2 hours at room temperature. Non-pecific antibody reactions were blocked by a 30-minutencubation in PBS containing 1 mg/mL of bovine serumlbumin and 100 mM of glycine.
For labeling, sheep anti-human tubulin polyclonal anti-ody (Cytoskeleton, Denver, CO) was solubilized in thelocking solution (1:200 dilution) and incubated with theamples for 1 hour at 37°C. After washing in PBS containing.1% Triton X-100 (v/v), the oocytes were labeled withlexa Fluor 488 or Alexa Fluor 568 donkey anti-sheep IgG
Molecular Probes, Eugene, OR) at a 1:200 dilution in block-ng solution for 40 to 60 minutes at 37°C, then washed againith PBS containing 0.1% Triton X-100. To stain the DNA,
ach oocyte was put in a slide with a 10-�L drop of antifadeolution with 0.1 �g/mL of 4,6-diamidino-2-phenylindole-API (Appligene, Tucson, AZ). The slides were then cov-
red with coverslips and sealed with nail polish. Oocytesere examined with a Zeiss Axiophot II microscope
quipped with a triple band pass filter.
After ICC analysis and photography, the coverslips werehen carefully removed and the oocytes, which stayed at-ached to the slides, were gently rinsed with 0.1% Triton-100 in PBS and prepared for FISH. The DNA probes
Vysis, Inc., Downers Grove, IL) used for this study wereentromeric fluorescent probes for chromosomes X, Y, and8 (CEP X Spectrum Green, CEP Y Spectrum Orange, andEP 18 Spectrum Orange), applied both individually and inombination. Fixation with acetic acid/methanol or an etha-ol dehydration series were not used.
The FISH procedure was performed according to therotocol recommended by the manufacturer for directly la-eled probes: 10 �L of the probe mixture was applied toach slide under a coverslip sealed with rubber cement. TheNA was denatured at 75°C on a hot plate for 5 minutes and
ybridization took place overnight in a humidified chamber1655
aw(IwDm
asdFscssasFmqop
swimToFwgfas
fmstcAimstvq
o
CfHstXswcmaa(ib
Ramalho-Santos. ICG and FISH in human fertilization failures. Fertil Steril 2004.
1
t 42°C. Coverslips were removed, and the slides wereashed twice: first, in 0.4� SSC solution with 0.3% Igepal
Appligene) at 75°C for 1 minute, then, with 2� SSC/0.1%gepal solution for 1 minute at room temperature. Slidesere mounted in antifade medium containing 0.1 �g/mL ofAPI (Appligene) and analyzed with a Zeiss Axiophot IIicroscope as previously noted.
RESULTSInitial attempts to first analyze and document tubulin ICC
nd then perform FISH on the same sample using the re-pective standard methodologies were largely unsuccessfulue to several factors. Some steps normally required forISH (ethanol/acetic acid fixation, ethanol dehydration)eemed to disrupt the chromatin structure or erase the mi-rotubular staining patterns detected by ICC (data nothown). On the other hand, standard permeabilization ofamples in 1% Triton X-100 for 30 minutes following form-ldehyde fixation, although suitable for ICC, rendered theamples opaque following the heating steps required forISH probe hybridization, probably due to massive cytoplas-ic protein denaturation (data not shown). As a conse-
uence, only chromatin at the cell surface could be unequiv-cally labeled with FISH probes, namely, metaphase IIlates and polar bodies as already described (7, 10–12).
We tackled these issues with two methodologicalhifts. To avoid oocyte opacity following FISH analysis,e increased the permeabilization used for ICC. Follow-
ng fixation with formaldehyde, samples were thus per-eabilized for 1 to 2 hours with 5% Triton X-100 in PBS.his was particularly effective in removing enough of theocyte cytoplasm so that the heating steps necessary forISH did not render the cell opaque. On the other hand,e found that, after ICC data had been collected for aiven sample, it was no longer necessary to performurther fixation steps for FISH analysis with methanol/cetic acid or proceed through the ethanol dehydrationeries (data not shown).
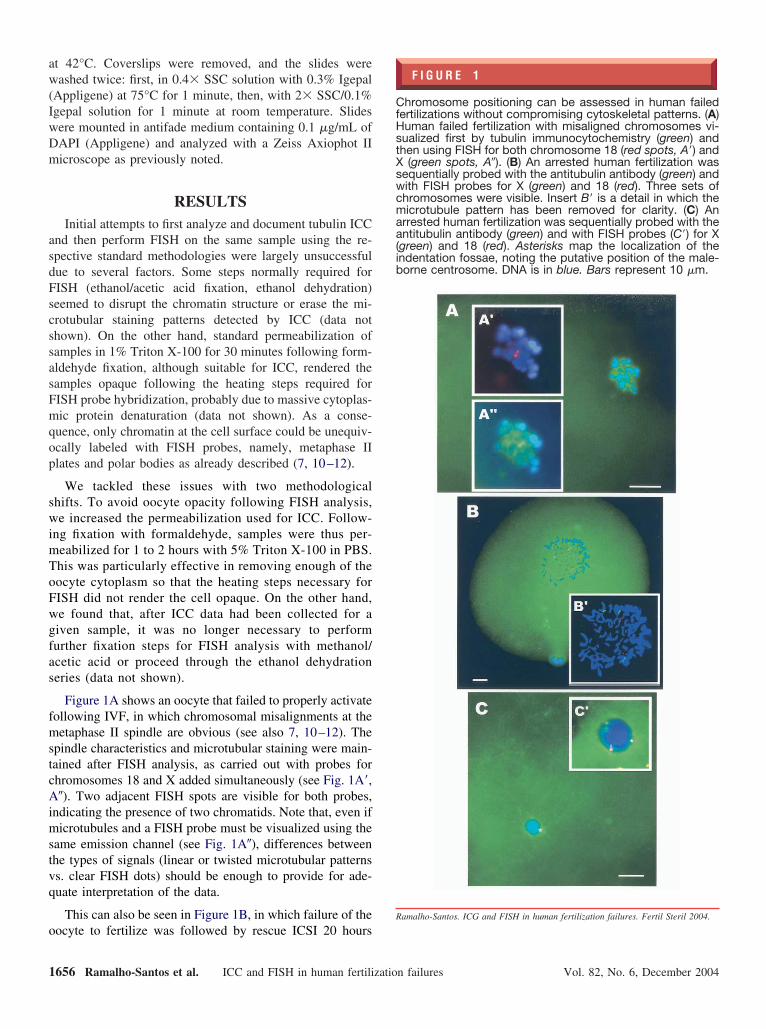
Figure 1A shows an oocyte that failed to properly activateollowing IVF, in which chromosomal misalignments at theetaphase II spindle are obvious (see also 7, 10–12). The
pindle characteristics and microtubular staining were main-ained after FISH analysis, as carried out with probes forhromosomes 18 and X added simultaneously (see Fig. 1A=,�). Two adjacent FISH spots are visible for both probes,
ndicating the presence of two chromatids. Note that, even ificrotubules and a FISH probe must be visualized using the
ame emission channel (see Fig. 1A�), differences betweenhe types of signals (linear or twisted microtubular patternss. clear FISH dots) should be enough to provide for ade-uate interpretation of the data.
This can also be seen in Figure 1B, in which failure of the
ocyte to fertilize was followed by rescue ICSI 20 hours656 Ramalho-Santos et al. ICC and FISH in human fertilizatio
F I G U R E 1
hromosome positioning can be assessed in human failedertilizations without compromising cytoskeletal patterns. (A)uman failed fertilization with misaligned chromosomes vi-ualized first by tubulin immunocytochemistry (green) andhen using FISH for both chromosome 18 (red spots, A=) and
(green spots, A�). (B) An arrested human fertilization wasequentially probed with the antitubulin antibody (green) andith FISH probes for X (green) and 18 (red). Three sets ofhromosomes were visible. Insert B= is a detail in which theicrotubule pattern has been removed for clarity. (C) An
rrested human fertilization was sequentially probed with thentitubulin antibody (green) and with FISH probes (C=) for X
green) and 18 (red). Asterisks map the localization of thendentation fossae, noting the putative position of the male-orne centrosome. DNA is in blue. Bars represent 10 �m.
n failures Vol. 82, No. 6, December 2004
anpIwa
p
famnptI
Hfhmsoc(
R
F
fter conventional insemination. The oocyte was then diag-osed in the clinic as a 3PN (possible retention of the secondolar body) and made available for analysis. Analysis byCC–FISH revealed three sets of condensed chromosomesith a very loose microtubule network that failed to organizespindle.
More importantly, the technique could also be applied to
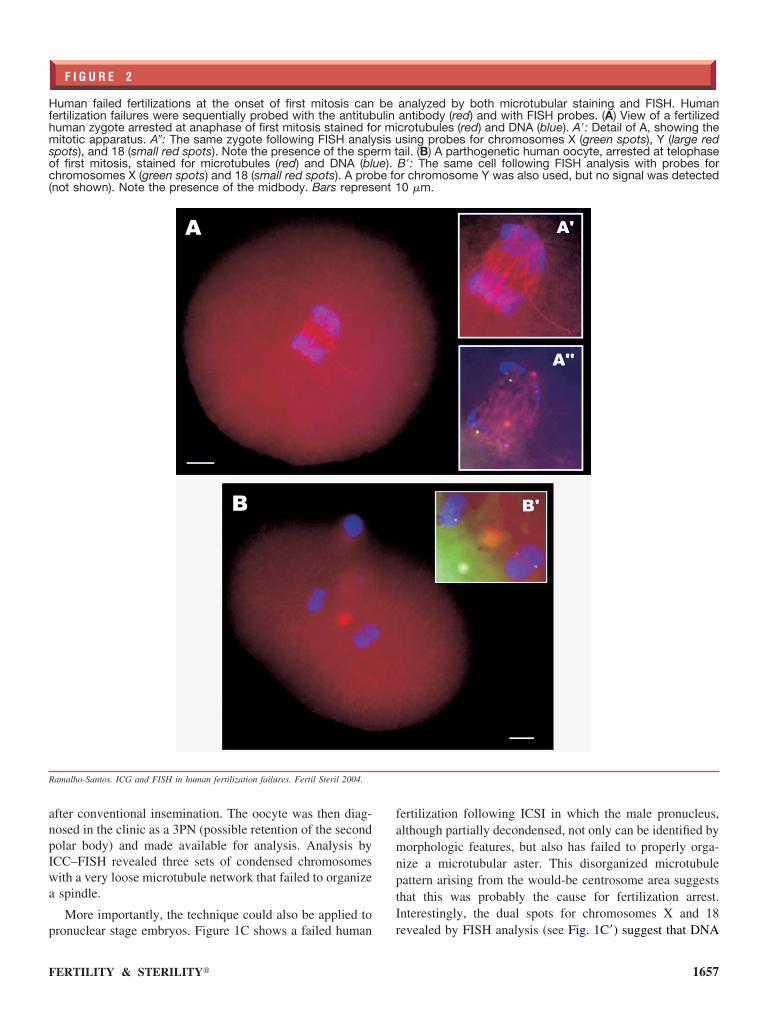
F I G U R E 2
uman failed fertilizations at the onset of first mitosis canertilization failures were sequentially probed with the antitubuman zygote arrested at anaphase of first mitosis stained foitotic apparatus. A�: The same zygote following FISH analy
pots), and 18 (small red spots). Note the presence of the spef first mitosis, stained for microtubules (red) and DNA (blhromosomes X (green spots) and 18 (small red spots). A pro
not shown). Note the presence of the midbody. Bars repres
amalho-Santos. ICG and FISH in human fertilization failures. Fertil Steril 2004.
ronuclear stage embryos. Figure 1C shows a failed human r
ERTILITY & STERILITY�
ertilization following ICSI in which the male pronucleus,lthough partially decondensed, not only can be identified byorphologic features, but also has failed to properly orga-
ize a microtubular aster. This disorganized microtubuleattern arising from the would-be centrosome area suggestshat this was probably the cause for fertilization arrest.nterestingly, the dual spots for chromosomes X and 18
analyzed by both microtubular staining and FISH. Humanantibody (red) and with FISH probes. (A) View of a fertilizedcrotubules (red) and DNA (blue). A=: Detail of A, showing thesing probes for chromosomes X (green spots), Y (large red
ail. (B) A parthogenetic human oocyte, arrested at telophaseB=: The same cell following FISH analysis with probes forr chromosome Y was also used, but no signal was detected0 �m.
beulinr misis urm t
ue).be foent 1
evealed by FISH analysis (see Fig. 1C=) suggest that DNA
1657
ssmn
tFyfit
sfld(2sb
toaiatf
aaIasmFttscstataas
hcgscFtbscpfaopk
Hbait(fi
R
1
ynthesis had already begun, similar to what has been de-cribed for other primate species following cytoplasmic re-oval of sperm structures after ICSI (14) but before pro-
uclear apposition.
We were also able to analyze human fertilization failureshat failed to progress beyond the first mitotic division.igure 2 shows two failed fertilizations from the same 33-ear-old patient with endometriosis undergoing IVF. Therst embryo analyzed had arrested at anaphase of first mi-
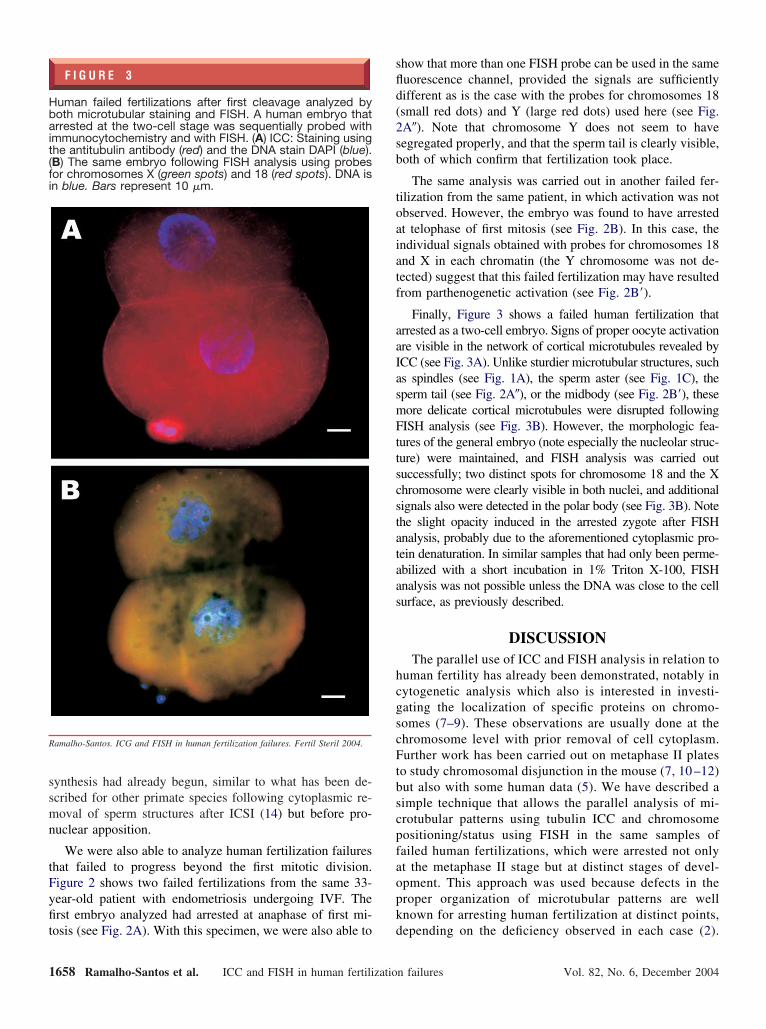
F I G U R E 3
uman failed fertilizations after first cleavage analyzed byoth microtubular staining and FISH. A human embryo thatrrested at the two-cell stage was sequentially probed with
mmunocytochemistry and with FISH. (A) ICC: Staining usinghe antitubulin antibody (red) and the DNA stain DAPI (blue).B) The same embryo following FISH analysis using probesor chromosomes X (green spots) and 18 (red spots). DNA isn blue. Bars represent 10 �m.
amalho-Santos. ICG and FISH in human fertilization failures. Fertil Steril 2004.
osis (see Fig. 2A). With this specimen, we were also able to d
658 Ramalho-Santos et al. ICC and FISH in human fertilizatio
how that more than one FISH probe can be used in the sameuorescence channel, provided the signals are sufficientlyifferent as is the case with the probes for chromosomes 18small red dots) and Y (large red dots) used here (see Fig.A�). Note that chromosome Y does not seem to haveegregated properly, and that the sperm tail is clearly visible,oth of which confirm that fertilization took place.
The same analysis was carried out in another failed fer-ilization from the same patient, in which activation was notbserved. However, the embryo was found to have arrestedt telophase of first mitosis (see Fig. 2B). In this case, thendividual signals obtained with probes for chromosomes 18nd X in each chromatin (the Y chromosome was not de-ected) suggest that this failed fertilization may have resultedrom parthenogenetic activation (see Fig. 2B=).
Finally, Figure 3 shows a failed human fertilization thatrrested as a two-cell embryo. Signs of proper oocyte activationre visible in the network of cortical microtubules revealed byCC (see Fig. 3A). Unlike sturdier microtubular structures, suchs spindles (see Fig. 1A), the sperm aster (see Fig. 1C), theperm tail (see Fig. 2A�), or the midbody (see Fig. 2B=), theseore delicate cortical microtubules were disrupted followingISH analysis (see Fig. 3B). However, the morphologic fea-
ures of the general embryo (note especially the nucleolar struc-ure) were maintained, and FISH analysis was carried outuccessfully; two distinct spots for chromosome 18 and the Xhromosome were clearly visible in both nuclei, and additionalignals also were detected in the polar body (see Fig. 3B). Notehe slight opacity induced in the arrested zygote after FISHnalysis, probably due to the aforementioned cytoplasmic pro-ein denaturation. In similar samples that had only been perme-bilized with a short incubation in 1% Triton X-100, FISHnalysis was not possible unless the DNA was close to the cellurface, as previously described.
DISCUSSIONThe parallel use of ICC and FISH analysis in relation to
uman fertility has already been demonstrated, notably inytogenetic analysis which also is interested in investi-ating the localization of specific proteins on chromo-omes (7–9). These observations are usually done at thehromosome level with prior removal of cell cytoplasm.urther work has been carried out on metaphase II plates
o study chromosomal disjunction in the mouse (7, 10 –12)ut also with some human data (5). We have described aimple technique that allows the parallel analysis of mi-rotubular patterns using tubulin ICC and chromosomeositioning/status using FISH in the same samples ofailed human fertilizations, which were arrested not onlyt the metaphase II stage but at distinct stages of devel-pment. This approach was used because defects in theroper organization of microtubular patterns are wellnown for arresting human fertilization at distinct points,
epending on the deficiency observed in each case (2).n failures Vol. 82, No. 6, December 2004

Tmaavgatgpb
olcpsdaowlwipaptoatcacicmsmb
rb
Aad(p
R
1
1
1
1
1
F
he methodology may be useful for example, in deter-ining which specific chromosomes are missorted or mis-
ligned in oocytes or zygotes from specific patients, inssessing whether arrest took place before or after acti-ation of DNA synthesis in the pronuclei, or in distin-uishing failed fertilization from failed parthenogeneticctivation. More importantly the optimized protocol re-ains the structural morphology of the sample withoutross distortions and can be used regardless of chromatinosition in the oocyte; that is, the DNA does not need toe close to the cell surface.
Simultaneous detection of several types of molecules,rganelles, or structures in a human oocyte or any otheriving cell is straightforward (13), provided that severalonditions are fulfilled. For example, the appropriaterobes must be available, and they must not overlap inpecificity or in the emitted signals. The final hurdle isetermining if the methodology used for different probes/nalysis is compatible. For example, some probes cannly be used to label live cells, while others work bestith fixed material. In this particular case, the most chal-
enging aspect was avoiding cytoplasmic interference,hich would render the samples opaque after FISH. This
ssue was resolved using increased detergent-mediatedermeabilization. It is therefore important to note thatpplying this technique to compare the localization ofarticular proteins/cellular structures may be limited byhe nature of the protein/structure in question. If it is lostr altered following extensive permeabilization, for ex-mple, the results may be inconclusive or may not reflecthe accurate localization/concentration of a given mole-ule of interest. This is particularly important in thenalysis of human oocytes or arrested zygotes, given thatell size and the abundance of concentrated cytoplasm cannterfere with analysis if they are not adequately pro-essed. A balance must be struck between the two (orore) methodologic approaches that need to be applied,
imultaneously or sequentially. Therefore, the protocolust be optimized for each particular analysis, with care
eing taken that a given sample is not irreversibly dis-
ERTILITY & STERILITY�
upted at any stage, thus rendering direct comparisonsetween techniques difficult.
cknowledgments: The authors thank Dr. Ana Barbosa, Conceição Dias,nd Paula Henriques (University Hospitals of Coimbra) for many helpfuliscussions and technical assistance, and Prof. Agostinho Almeida SantosUniversity Hospitals of Coimbra) for continuous encouragement and sup-ort. We also thank Dr. H. Beard for thoughtful advice.
eferences1. Trounson AO, Gosden RG, eds. Biology and pathology of the oocyte:
its role in fertility and reproductive medicine Cambridge, UK: Cam-bridge University Press, 2003.
2. Asch R, Simerly C, Ord T, Ord VA, Schatten G. The stages at whichhuman fertilization arrests: microtubule and chromosome configura-tions in inseminated oocytes which failed to complete fertilization anddevelopment in humans. Hum Reprod 1995;10:1897–906.
3. Coonen E, Hopman AHN, Geraedts JPM, Remaekers CS. Applicationof in-situ hybridization techniques to study human preimplantationembryos: a review. Hum Reprod Update 1998;4:135–52.
4. Schatten G. The centrosome and its mode of inheritance: the reductionof the centrosome during gametogenesis and its restoration duringfertilization. Dev Biol 1994;165:299–335.
5. Hassold T, Hunt P. To err (meiotically) is human: the genesis of humananeuploidy. Nat Rev Genet 2001;2:280–91.
6. Barlow AL, Tease C, Hulten MA. Meiotic chromosome pairing in fetaloocytes of trisomy 21 human females. Cytogenet Genome Res 2002;96:45–51.
7. Hodges CA, LeMaire-Adkins R, Hunt PA. Coordinating the segrega-tion of sister chromatids during the first meiotic division: evidence forsexual dimorphism. J Cell Sci 2001;114:2417–26.
8. Hodges CA, Hunt PA. Simultaneous analysis of chromosomes andchromosome-associated proteins in mammalian oocytes and embryos.Chromosoma 2002;111:165–9.
9. Tease C, Hartshorne GM, Hulten MA. Patterns of meiotic recombina-tion in human fetal oocytes. Am J Hum Genet 2002;70:1469–79.
0. Hunt P, LeMaire R, Embury P, Sheean L, Mroz K. Analysis ofchromosome behavior in intact mammalian oocytes: monitoring thesegregation of a univalent chromosome during female meiosis. HumMol Genet 1995;4:2007–12.
1. LeMaire-Adkins R, Radke K, Hunt PA. Lack of checkpoint control atthe metaphase/anaphase transition: a mechanism of meiotic nondisjunc-tion in mammalian females. J Cell Biol 1997;139:1611–9.
2. LeMaire-Adkins R, Hunt PA. Nonrandom segregation of the mouseunivalent X chromosome: evidence of spindle-mediated meiotic drive.Genetics 2000;156:775–83.
3. Simerly C, Moreno RD, Ramalho-Santos J, Hewitson L, Schatten G.Confocal imaging of structural molecules in mammalian gametes. In:Gardner DK, Lane M, eds. A laboratory guide of the mammalianembryo. Oxford, UK: Oxford University Press, 2003.
4. Ramalho-Santos J, Sutovsky P, Simerly C, Oko R, Wessel G, HewitsonL, et al. ICSI choreography: fate of sperm structures after monospermic
rhesus ICSI and first cell cycle implications. Hum Reprod 2000;15:2610–20.1659
Top Related
Copyright © 2022 FDOKUMEN

