







Bahasa
Halaman
Hukum


RGS9 proteins facilitate acute tolerance to mu-opioideffects
Javier GarzoÂn, Marta RodrõÂguez-DõÂaz, Almudena LoÂpez-Fando and Pilar SaÂnchez-BlaÂzquezNeurofarmacologõÂa, Instituto de NeurobiologõÂa Santiago RamoÂn y Cajal, Consejo Superior de Investigaciones Cientõ®cas,
Avenida Doctor Arce 37, E-28002 Madrid, Spain
Keywords: 125I-Tyr27-human b-endorphin, analgesia, antinociception, antisense technology, G-proteins, mouse, receptordesensitization, regulator of G-protein signalling
Abstract
This paper reports that regulators of G-protein signalling (RGS) proteins modulate the timing and amplitude of opioid signals by a
push±pull mechanism. This is achieved without noticeable changes in the binding properties of opioids, e.g. b-endorphin to mu-
opioid receptors. The expression of RGS proteins was reduced by blocking their mRNA with antisense oligodeoxynucleotides(ODN). Knock down of RGS2 or RGS3 diminished morphine and b-endorphin analgesia, whereas that of RGS9 or RGS12
enhanced this activity. In mice with impaired RGS9, but not impaired RGS2, the potency and, in particular, the duration of opioid
antinociception increased. Further, the animals did not exhibit acute tolerance generated by a single and ef®cacious dose of
morphine, nor did they develop tolerance after a daily i.c.v. injection of the opioid for 4 days. In a model of sustained morphinetreatment, the impairment of RGS9 proteins facilitated increases in the response to the delivered opioid. This was only effective
for 2±3 h after the subcutaneous implantation of an oily morphine pellet; later, tolerance developed. To reduce the impact of the
chronic morphine acting on opioid receptors, other RGS proteins presumably substitute the GTPase-activating function of RGS9on morphine-activated Ga-GTP subunits. The desensitization of mu-opioid receptors appears to be a cell membrane-limited
process facilitated by RGS9¢s sequestering of agonist-segregated Ga subunits.
Introduction
Normally, repeated administration of opioids bring about a progres-
sive decrease in their potency. This is observed after even single
doses of agonists (acute tolerance or tachyphylasis), and after long-
term treatments that lead to development of tolerance and physical
dependence on these substances. Signi®cant knowledge has been
accumulated on the adaptive alterations triggered by the chronic use
of opioids, including the gating of ion channels, the expression of
particular genes, changes in the number and function of guanine
nucleotide-binding regulatory protein (G-proteins) (Strassheim &
Malbon, 1994), marked up-regulation of the adenylyl cyclase/cAMP
system, enhancement of (cAMP)-dependent protein kinase (PKA)
(Nestler et al., 1993) and PKC activities (Narita et al., 1994), as well
as modi®cations in cell morphology (see, e.g. Nestler & Aghajanian,
1997). However, there have been fewer studies on tolerance induced
by an adequate single dose of opioids. The threshold dose necessary
to produce acute tolerance is about three to four times greater than
that required for producing analgesia (Huidobro et al., 1976). This
interesting phenomenon of acute tolerance appears within hours of
agonist administration and lasts for at least 2±3 days (Huidobro-Toro
& Way, 1978). It has been proposed that the mechanisms responsible
for acute tolerance parallel those underlying chronic morphine
tolerance, as both situations can be modulated by identical agents
(Fairbanks & Wilcox, 1997). In fact, tolerance to single doses of
opioids is accompanied by some degree of physical dependence that
is milder than that observed in the chronic morphine-dependent state
(Huidobro-Toro & Way, 1978; Bilsky et al., 1996). Pharmacological
studies have shown the possibility of regulating opioid-induced acute
tolerance (see, e.g., Fairbanks & Wilcox, 1997). However, precise
information is lacking on the role of opioid-acted transduction on the
genesis and timing of the changes responsible for acute tolerance or
tachyphylaxis.
Our knowledge of the particular elements involved in the
internalization and processing of the extracellular signals has greatly
increased in recent years. Besides receptors, G-proteins and effectors,
regulator of G-protein signalling (RGS) proteins have been described
as cooperating in the internalization of agonist-initiated messages. To
date, > 20 proteins are reported to contain RGS domains. The
majority are expressed in brain and differ in size, amino-acid
composition and distribution (Gold et al., 1997; Hepler, 1999). RGS
proteins serve as GTPase-activating proteins (GAPs) for a subunits of
heterotrimeric Gi, Go, Gz, Gt and Gq proteins (Berman & Gilman,
1998). By activating the metabolism of GTP to GDP, RGS proteins
promote the Ga-GDP state capable of binding to Gbg dimers
(Berman & Gilman, 1998; Hepler, 1999). However, during the period
which RGS proteins remain bound to Ga subunits, Ga(GDP)-Gbg re-
association is delayed. Thus, RGS proteins emerge as potential
negative regulators of G-protein signalling. The present study
addresses the role of RGS proteins in the potency of opioids, the
development of tachyphylasis to their effects, and long-term toler-
ance. With this purpose, oligodeoxynucleotides (ODNs) to mRNAs
coding for the described murine RGS proteins were given to mice,
and antinociception was taken to represent a paradigmatic opioid
Correspondence: Dr Javier GarzoÂn, as above.E-mail: [email protected]
Received 31 July 2000, revised 27 October 2000, accepted 11 December 2000
European Journal of Neuroscience, Vol. 13, pp. 801±811, 2001 ã Federation of European Neuroscience Societies

effect. The results show an important role for RGS9 proteins in the
development of acute tolerance to opioids at mu-receptors.
Materials and methods
Reduction of RGS function
Synthetic end-capped phosphorothioate ODNs were prepared by solid
phase phosphoramidite chemistry using a CODER 300 DNA
synthesizer (DUPONT; Wilmington, DE, USA) at the 1-mmol
scale. The introduction of phosphorothioate linkages was achieved
by tetraethylthiuram disulphide sulphurization. Crude ODNs were
puri®ed by conventional reverse-phase chromatography through a 5-
mm/C18 column (Spherisorb ODS-2, 150 3 4.6 mm, Symta, Madrid,
Spain) using 0.1 M triethylammonium acetate (pH 7.0) and aceto-
nitrile as the mobile phase. To reduce the synthesis of RGS proteins,
the following ODN sequences were used (PrimerSelect, DNASTAR
Inc, Madison, WI, USA):
FIG. 1. Knock down of RGS proteins andmorphine antinociception. Animals receivedeither saline, the ODN-RD or the antisenseODN into the right lateral ventricle over a 5-day schedule. On day 6, 3 nmol morphine wasi.c.v.-injected and antinociceptive activityevaluated 30 and 90 min later. The opioidagonist produced similar effects in mice thati.c.v.-received the vehicle or the random oligo,and in noninjected (naive) animals. Thus, theresponsiveness of the mice was not altered bythe experimental procedure alone. Values arethe mean 6 SEM from groups of 10 mice.*P < 0.05 vs. the control group, ANOVA,Student±Newman±Keuls test. (Lower panels)Detection of RGS9-2 and RGS2 in mousebrain. The ef®cacy and selectivity of the ODNtreatments was assessed with antibodiesdirected to RGS2 proteins, RGS9 proteins,Gb1/2 subunits, Gi2a subunits and mu-opioidreceptors. SDS-solubilized cerebral cortex,striatum and PAG were resolved by SDS±PAGE (10±20% acrylamide/2,6%bisacrylamide) and Western blotted.Immunodetection was carried out as describedin Materials and Methods. Detection of RGS9-2 mRNA using RT-PCR. Lanes contain 0.3 mgof total RNA from the mouse brain structures.
802 J. GarzoÂn et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

the 16-base ODN 5¢-C*T*CTATGAAGTCGG*T*A-3¢ to nucleotides
428±443 of the RGS2 gene (ODN1),
the 16-base ODN 5¢-G*C*TGAATTCAAGGT*C*A-3¢ to nucleotides
743±758 of the RGS2 gene (ODN2),
the 16-base ODN 5¢-C*A*CAGCATTCGGTC*C*A-3¢ to nucleotides
1211±1227 of the RGS2 gene (ODN3) (U67187),
the 16-base ODN 5¢-C*T*TCATGTCCTTGG*C*T-3¢ to nucleotides
105±120 of the RGS3 gene (AA880930),
the 17-base ODN 5¢-G*G*ACTTAGTTTAGAA*G*G-3¢ to nucleo-
tides 310±326 of the RGS4 gene (Ab004315),
the 16-base ODN 5¢-C*T*GTCTGGATGAAT*T*C-3¢ to nucleotides
389±404 of the RGS5 gene (U67188),
the 17-base ODN 5¢-G*G*TCTTGTCATAACT*C*T-3¢ to nucleo-
tides 1381±1397 of the RGS7 gene (Af011360),
the 16-base ODN 5-T*G*GATCAACATAGA*C*G-3¢ (ODN1) to
nucleotides 532±547 of the RGS9 gene,
the 16-base ODN 5¢-C*T*CGAATCAGTTCG*C*T-3¢ to nucleotides
973±988 of the RGS9 gene (ODN2),
the 16-base ODN 5¢-C*G*CAGACCCACATT*T*A-3¢ to nucleotides
1234±1249 of the RGS9 gene (ODN3) (Af011358),
the 16-base ODN 5¢-C*T*ACAGGCCTCTGA*C*A-3¢ to nucleotides
165±180 of the RGS12 gene (AA855540),
the 16-base ODN 5¢-G*T*AGTGGCAGCTTG*T*A-3¢ to nucleotides
431±446 of the RGS14 gene (U70427) and
an 18-base ODN 5¢-A*T*GTCCGTCTGTCCTT*C*C-3¢ to nucleo-
tides 832±850 of the RGS16 gene (U67189).
These sequences displayed no homology to other relevant cloned
proteins (GeneBank database). Antisense ODN controls consisted of
mismatched sequences in which some bases were switched (in bold)
without altering the remaining sequence:
RGS2M1, 5¢-C*A*CTAAGATGTCCGAA-3¢;RGS2M2, 5¢-G*C*AGAATACATGGA*C*A-3¢;RGS2M3, 5¢-CACTCCATACGCTGCA-3¢; and
RGS9M1, 5¢-T*C*GATGAACTAAGT*G*C-3¢;RGS9M2, 5¢-C*T*GCAATGAGTTGC*T*C-3¢;RGS9M3, 5¢-C*G*TACACGCACTTA*T*A-3¢;
and also of a random ODN (ODN-RD) (SaÂnchez-BlaÂzquez et al.,
1995).
ODN solutions were made up in saline immediately prior to use.
Different control groups of animals were used to ascertain the
speci®city of ODN treatments. These controls include noninjected
mice (naive), those that received the vehicle (saline), and animals
injected with a random sequence (ODN-RD) or mismatched antisense
sequence (ODN-RGS#M). As no signi®cant differences were
observed in the response to the analgesic substances of naive,
vehicle-, ODN-RD- or ODN-RGS#M-injected mice (see e.g. Figs 1
and 2), only the data from the control ODNs (ODN-RD or ODN-
RGS#M) are presented in this paper.
Animals were lightly anaesthetized with ether and injections made
into the lateral ventricle with a 10-mL Hamilton syringe to a depth of
3 mm, 2 mm lateral and 2 mm caudal to the bregma. The 4-mL
content was infused at a rate of 1 mL every 5 s. The needle was
maintained for an additional 10 s. To minimize the chance of
neurotoxicity caused by repeated i.c.v. injections, an interval of 24 h
was allotted between successive administrations of the ODNs
(Chiasson et al., 1994). Each ODN treatment was performed on a
distinct group of mice according to the following 5-day schedule: on
days 1 and 2 with 1 nmol, on days 3 and 4 with 2 nmol, on day 5 with
3 nmol (SaÂnchez-BlaÂzquez et al., 1995; GarzoÂn et al. 2000).
Functional studies usually started on day 6. The possible alterations
promoted by ODN treatments on animal activity were recorded with a
Digiscan animal activity monitor system (activity cage) (Omnitech
electronics, Columbus, OH, USA). Groups of four mice that had
received an ODN following the above-described schedule were
monitored for horizontal and vertical activity for periods of 10 min.
Only ODNs that produced no signi®cative changes in these
behavioural parameters were selected for this study (ANOVA,
Student±Newman±Keuls test, P < 0.05).
To monitor any possible injury to tissue structure caused by i.c.v.
delivery of the ODNs, Nissl staining was routinely performed on
brain coronal slices from mice receiving each ODN treatment. These
slices included some of the periventricular regions responsible for the
antinociceptive effects of opioids injected via the i.c.v. route (Yaksh
et al., 1976; Rossi et al., 1994). The histological procedure is
described elsewhere (GarzoÂn et al. 2000). The ODNs produced no
alterations to the normal structure of periaqueductal grey matter
(PAG) or any other periventricular region (not shown).
Detection of RGS9 and RGS2 in mouse brain: electrophoresisand immunoblotting
At the end of ODN treatment, groups of mice were killed by
decapitation. The animals' cerebral cortices, striata and PAG were
then collected. For each determination, the structures from four mice
were pooled and SDS-solubilized P2 membranes were subsequently
resolved by SDS/polyacrylamide gel electrophoresis (PAGE) in
8 3 11 3 1.5 cm gel slabs (10±20% total acrylamide concentration/
2.6% bisacrylamide cross-linker concentration). For RGS9 immuno-
detection, 45, 60 and 80 mg protein/lane were used for striatum, PAG
and cerebral cortex determination, respectively. For RGS2 immuno-
detection, 60 mg protein/lane were used for each neural structure.
Afterwards, the separated proteins were transferred to 0.2-mm
polyvinylidene di¯uoride membranes (Bio-Rad). Polyclonal anti
RGS2 and anti RGS9 1 : 500 [Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA; SC9103 (rabbit) and SC8143 (goat),
respectively,], anti Gi2a subunits (SaÂnchez-BlaÂzquez et al., 1995),
anti Gb1/2 subunits (Lang & Costa, 1987) and anti mu-opioid
receptor (SaÂnchez-BlaÂzquez et al., 1995, 1997) 1 : 1000 were diluted
in TBS-0.05% Tween 20 (TTBS) and incubated with the transfer
membranes at 6 °C for 24 h. The SC8143 was detected with donkey
antigoat IgG horseradish peroxidase conjugate antiserum (Santa Cruz
Biotechnology, SC2020). The other polyclonal antisera were detected
with a goat antirabbit IgG (H+L) horseradish peroxidase conjugate
antiserum (BioRad; 170±6515, Madrid, Spain). Secondary antisera
were diluted 1 : 2000 in TTBS, incubated for 3 h and visualized with
the DAB substrate solution. Immunoblots were analysed by
densitometry using an instrument with re¯ectance capabilities (GS-
700 Imaging Densitometer, BioRad) and Molecular Analyst Software
(v 1.5; BioRad).
RT-PCR
Total RNA was harvested from mouse brain structures using a single-
step procedure (Ultraspec RNA isolation system, Biotecx Labs,
Houston, TX, USA), based on the formation of RNA complexes with
guanidinium molecules, followed by isopropanol precipitation. The
pellet was washed in 75% ethanol, dried, resuspended in 40 mL RNA
storage solution (Ambion, Austin, TX, USA) and stored at ±80 °C
until analysis. The yield of RNA was determined using UV
spectrometry (260 nm).
Two micrograms of total striatum, PAG and cerebral cortex RNA
was reverse transcribed using the RT-PCR First Strand Synthesis Kit
(RETROscript, Ambion) with oligo (deoxythymidine) priming.
cDNA synthesis was carried out at 42 °C for 60min. PCR reactions
were performed on 3, 5 and 7 mL of RT product (equivalent to 0.3,
0.5 and 0.7 mg total RNA, respectively) in a ®nal volume of 50 mL of
RGS9 facilitates opioid tachyphylaxis 803
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

the buffer solution containing 10 mM Tris HCl (pH 8.3), 50 mM KCl,
1.5 mM MgCl2, 125 mM of each dNTP, 0.5 mM of each primer and
1 unit of SuperTaq Thermostable DNA polymerase (Ambion). cDNA
primers (AmershamPharmacia Biotech, Barcelona, Spain) directed
towards the murine RGS9 gene (AF011358) were adapted from those
described by Granneman et al. (1998) directed towards rat and human
counterparts. The sequences were: forward 5¢-GAAGCCTGTGAGG-
ACCTGAAGTACG-3¢ (sense) corresponding to base pairs 1060±
1084, and reverse 5¢-AGGAGGCAGCTCCTTTTTGAGTTG-3¢(antisense) corresponding to base pairs 1498±1521 (in order to
yield an expected band size of 462 base pairs). The RT-PCR products
were initially denatured at 95 °C for 1 min and then subjected to
cycles of denaturation (95 °C, 20 s) and annealing (65 °C, 30 s).
Ampli®cation cycles were 35 in total. After the last, extension was
performed for 7 min at 65 °C. A DNA Thermal Cycler 480 (PE
Biosystems, Foster City, CA, USA) and thin-walled 0.5 mL PCR
tubes (Ambion) were used. PCR products were electrophoresed
(34 V, 2.5 h) in a 1.5% agarose gel with PCR markers 50±2000
(Novagen, Madison, WI, USA), and then incubated in SYBR gold
solution (Molecular Probes, Poortgebouw, Netherlands) for 20 min.
FIG. 2. The in¯uence of RGS9 proteins on the time course of morphine-evoked analgesia. Animals with reduced levels of RGS9 proteins throughODN treatment were i.c.v.-injected with various doses of morphine andanalgesia was determined at various intervals by the warm water 52 °C tail-¯ick test. The effects of the opioid at 30, 60 and 90 min post injection arealso presented in dose-effect curves. Values are mean 6 SEM from groupsof 10±20 mice. *P < 0.05 vs. the group that had received the mismatchedcontrol ODN-RGS9M or the vehicle (saline).
FIG. 3. Opposite effects of RGS9 and RGS2 proteins on the time course ofb-endorphin-evoked analgesia. *,+P < 0.05 vs. the group that had receivedthe mismatched control ODN-RGS9M or ODN-RGS2M, respectively.Details as in Fig. 2.
804 J. GarzoÂn et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

DNA products were visualized with UV light (Gelprinter, T.D.I.,
Madrid, Spain).
Preparation of membranes from mouse brain
Male albino CD/1 mice (Charles River, Barcelona, Spain) weighing
22±27 g were used to provide experimental tissue. Mice that had
received i.c.v. injections of the ODNs were killed by cervical
dislocation. Brains were quickly removed and washed in ice-cold
50 mM Tris HCl, 0.32 M sucrose, pH 7.5, at 4 °C. PAGs (up to 1 mm
around the aqueduct) were collected and homogenized using a
Polytron (model PT 10±35, Kinematica, Kriens-Luzern, Switzerland)
at setting of 3 for 15 s. The homogenate was then centrifuged at
1000 g at 4 °C for 10 min (Sorvall RC5C, rotor SS-34 Newton, CT,
USA). After the pellet was discarded, the supernatant was centrifuged
at 20 000 g for 20 min. The pellet was resuspended in buffer and
centrifuged at 20 000 g for an additional 20 min. The ®nal pellet (P2)
was diluted in Tris buffer supplemented with a mixture of protease
inhibitors (0.2 mM phenylmethylsulphonyl ¯uoride, 2 mg/mL leu-
peptin and 0.5 mg/mL aprotinin) to a ®nal protein concentration of
» 2 mg/mL.
Binding studies125I-Tyr27 human b-endorphin, obtained lyophilized, was reconsti-
tuted with 100 mL of distilled water to give 0.25% BSA, 5%
lactose, 0.2% L-cysteine hydrochloride, 10 mM citric acid, and
800 KIU/mL aprotinin. Four-millitre siliconized borosilicate glass
tubes (Sigmacote from Sigma, St Louis, MO, USA) were used
(adsorption of the iodinated peptide was reduced to < 5%). To
restrict the speci®c binding of iodinated b-endorphin to only mu-
opioid receptors, saturation assays of 125I-Tyr27 human b-
endorphin, used at concentrations ranging from 0.3 pM to 3 nM,
were conducted in the presence of 300 nM N,N-diallyl-Tyr-(a-
aminoisobutyric acid)2-Phe-Leu-OH (ICI-174,864; GarzoÂn et al.,
1998). The membrane suspension reached a ®nal concentration of
» 0.2 mg/mL, 0.2% BSA, 0.01% bacitracin, incubation volume
was made up to 2 mL with 50 mM (®nal concentration) Tris HCl
buffer, pH 7.5. Non-speci®c binding was assessed in the presence
of 1 mM unlabelled b-endorphin. Samples were incubated in
triplicate at 25 °C for 90 min in a shaking incubator and ®ltered
under vacuum (Harvester M-12R, Brandel Gaitherburg, Maryland,
USA) through glass ®bre disks (Whatman GF/B, Maidstone,
England) previously immersed for 3 h in 5 mM Tris HCl/0.3%
polyethylenimine (Sigma), to minimize binding to the ®lters. The
®lters were then washed three times with 4 mL of ice-cold 5 mM
Tris HCl buffer, pH 7.5, placed in polyethylene counting vials
and counted using a LKB Compugamma CS counter (Bromma,
Sweden).
Animals and evaluation of antinociception
Male albino mice CD-1 weighing 22±25 g were housed and used
strictly in accordance with the guidelines of the European Community
for the Care and Use of Laboratory Animals (Council Directive 86/
FIG. 4. Role of RGS9 on acute tolerance to a single dose of morphine.Saline or a priming dose of 10 nmol morphine was i.c.v.-injected to themice that had received the ODN2 to RGS9, the mismatched ODN2-RGS9Mor ODN2-RGS2M. The group treated with ODN2-RGS2 received 30 nmolmorphine instead. All groups received, 24 h later, an i.c.v. test injection of10 nmol (or 30 nmol for ODN2-RGS2 group) morphine (A), and analgesiawas evaluated after 30 min. The time course of antinociception produced by10 nmol morphine was analysed in mice that had received saline or10 nmol morphine 24 h before. This was performed in mice undergoingODN2-RGS9M or ODN2-RGS9 treatment (B). Values are mean 6 SEMfrom groups of 10±20 mice. *P < 0.05 vs. the group injected with salinebefore the second dose of the opioid.
RGS9 facilitates opioid tachyphylaxis 805
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811
609/EEC). Animals were lightly anaesthetized with ether and, to
reduce the possibility of interference from spinal events, all substances
were i.c.v.-injected as 4 mL volumes into the right lateral ventricle as
previously described (SaÂnchez-BlaÂzquez et al., 1995). The response of
the animals to nociceptive stimuli was determined in the warm water
(52 °C) tail-¯ick test. Baseline latencies ranged from 1.5 to 2.2 s.
Antinociception was expressed as a percentage of the maximum
possible effect, MPE = 100 3 (test latency ± baseline latency)/[cut-
off time(10 s) ± baseline latency]. For the different assays of the
study, either a ®xed dose or increasing doses of the opioid agonists
were given, and antinociception assessed at the interval of peak effect
[30 min after morphine and b-endorphin or, in time-course studies, at
various intervals after the injection of the opioids (Figs 2 and 3)]. In
chronic morphine assays, antinociception evoked by the opioid was
determined at several intervals after subcutaneous (s.c.) implantation
of the pellet. Statistical analysis of the results was performed by
analysis of variance (ANOVA) followed by the Student±Newman±
Keuls test (SigmaStat, SPSS Science Software, Erkrath, Germany).
The level of signi®cance was set at P < 0.05.
Evaluation of acute tolerance to morphine
A single i.c.v. injection of 10 nmol morphine was used to produce
acute tolerance. Controls were given saline instead. Analgesia was
evaluated 24 h later when the pretreatment had no effect on baseline
latencies. The opioid was i.c.v.-administered to these mice and acute
tolerance was determined by the decrease of antinociceptive potency.
Induction and assessment of tolerance upon chronic morphinetreatment
In one set of assays, the mice received a daily i.c.v. injection of
10 nmol morphine for four consecutive days, and the development of
tolerance was monitored by measuring the analgesic response
attained after each injection. In other assays, the animals were s.c.
implanted with 10 mL/kg body weight of a suspension containing
50% saline (0.9% NaCl in distilled water), 42,5% mineral oil (Sigma
#400±5), 7.5% Mannide Monooleate (Sigma #M-8546), and 0.1 g/
mL morphine base (adapted from SaÂnchez-BlaÂzquez et al., 1997).
Development of tolerance was determined by measuring the
analgesic response to the s.c.-implanted chronic opioid and to a
single i.c.v. dose of 10 nmol morphine that produced, in mice not
previously exposed to the opioid, an effect of » 80% MPE in the tail-
¯ick test.
Chemicals
(3-[125I]Iodotyrosyl27)-b-endorphin-(1-31) (human) (IM.162,
2000 Ci/mmol) was obtained from Amersham (Buckinghamshire,
England). Human b-endorphin (1±31) was purchased from
Peninsula Laboratories (San Carlos, CA, USA). Morphine sulphate
was acquired from Merck (Darmstadt, Germany), ICI-174,864
from CRB (Cambridge, UK), and leupeptin, aprotinin, bacitracin,
phenylmethylsulphonyl ¯uoride and BSA were purchased from
Sigma.
Results
Treatment with the ODNs selected for this study did not alter the
basal latencies of the animals in the tail-¯ick test. Base-line latencies
always ranged from 1.5 to 2.5 s at 52 °C. The impairment of RGS4,
RGS7, RGS9, RGS12, RGS14 or RGS16 brought about increases in
the amplitude and duration of opioid-evoked antinociception,
whereas knock down of RGS2 and RGS3 diminished these param-
eters (Fig. 1). The improvement of opioid activity was even more
pronounced when the antinociceptive activity declined, e.g. 90 min
after a single injection of morphine. The ODN to RGS5 did not
change the analgesic pro®le exhibited by this opioid. RGS9 and
RGS2 were selected to characterize these opposed actions on opioid
antinociception.
The ef®cacy and selectivity of the ODN treatments were assessed
with antibodies directed to RGS2 and RGS9 proteins. After resolving
mouse cerebral cortex, striatum and PAG by SDS-PAGE, the
antibodies recognized a protein of » 28 kDa for RGS2 (Heximer
et al., 1997) and of » 77 kDa for RGS9-2, respectively (Rahman et al.,
1999). In mice treated with the ODNs to RGS2 proteins, a signi®cant
decrease in the immunolabeling of antibodies to RGS2 was found. In
Fig. 1 the data corresponding to ODN2-RGS2 vs. ODN2-RGS2M are
presented. Percentage decreases were 55 6 5* for striatum, 44 6 4*
for cerebral cortex and 51 6 5* for PAG (Fig. 1). The three ODNs to
RGS9 proteins brought about similar decreases of RGS9-2 associated
immunoreactivity. Percentage decreases promoted by ODN2-RGS9
vs. ODN2-RGS9M on RGS9-2 immunosignals were: 40 6 4* for
striatum, 60 6 5* for cerebral cortex and 48 6 5* for PAG (values
are the mean 6 SEM from three independent experiments; values are
signi®cantly different from the ODN-mismatch group; P < 0.05,
ANOVA, Student±Newman±Keuls test). Treatment with the ODNs-
RGS2M, ODNs-RGS9M or the ODN-RD did not alter the immuno-
reactivity associated to RGS2 or RGS9-2 proteins. The ODNs directed
to RGS proteins distinct from RGS2 or RGS9 brought about no
changes in RGS2-or RGS9-2-associated immunoreactivity. The knock
down of RGS9-2 proteins did not alter the expression of other proteins
also implicated in opioid cellular signalling, e.g. Gb1/2 subunits, Gi2asubunits or the mu-opioid receptor (Fig. 1). Knock down of RGS2
proteins also lacked effect on these signalling elements (not shown).
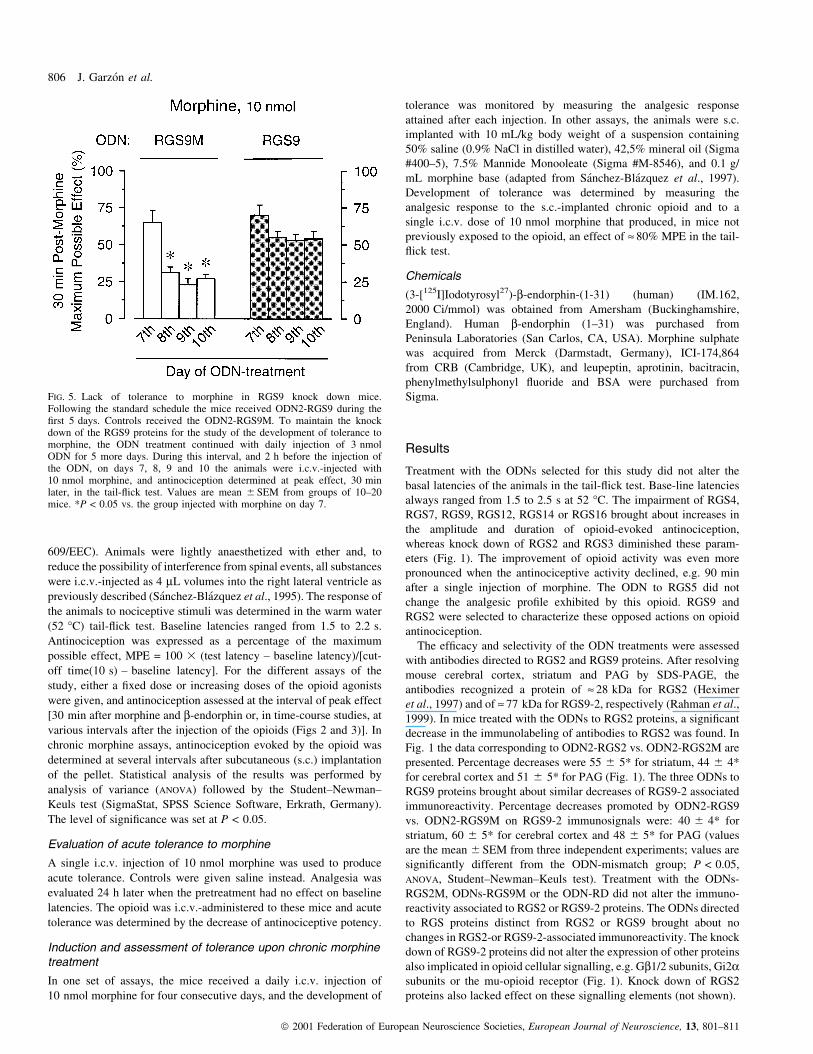
FIG. 5. Lack of tolerance to morphine in RGS9 knock down mice.Following the standard schedule the mice received ODN2-RGS9 during the®rst 5 days. Controls received the ODN2-RGS9M. To maintain the knockdown of the RGS9 proteins for the study of the development of tolerance tomorphine, the ODN treatment continued with daily injection of 3 nmolODN for 5 more days. During this interval, and 2 h before the injection ofthe ODN, on days 7, 8, 9 and 10 the animals were i.c.v.-injected with10 nmol morphine, and antinociception determined at peak effect, 30 minlater, in the tail-¯ick test. Values are mean 6 SEM from groups of 10±20mice. *P < 0.05 vs. the group injected with morphine on day 7.
806 J. GarzoÂn et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

In rat and human, the highest levels of RGS9-2 mRNA are found in
striatum and hypothalamus (Gold et al., 1997; Granneman et al.,
1998). In CD-1 mice, RT-PCR showed high signals for RGS9 mRNA
in striatum, but also low to medium levels in cerebral cortex and PAG
(Fig. 1).
In animals treated with the ODNs to RGS9, the potency of various
doses of morphine was signi®cantly enhanced over most of their
effective time course. The analgesia evoked by the opioid was
boosted and also became ef®cacious for longer postopioid intervals.
In using ODN2-RGS9, the apparent ED50s (i.c.v. morphine nmol/
mouse) and 95% con®dence limits in control mice were 3.51 (2.59±
4.71), 6.52 (4.67±9.03) and > 10 when determined 30, 60 and 90 min,
respectively, after the opioid. In RGS9 knock down mice, the
morphine ED50s at these intervals shifted to 1.10 (0.91±1.32), 2.31
(1.81±2.92) and 3.60 (2.53±5.11) nmol/mouse (Fig. 2). Similarly, in
animals treated with the ODNs to RGS9 proteins, the potency of
various doses of the opioid neuropeptide b-endorphin was signi®-
cantly enhanced over most of their effective time courses. Analgesia
was boosted and also made ef®cacious over longer postopioid
intervals. The ODNs to RGS2 proteins produced the opposite effects.
The activity of b-endorphin was diminished over the entire time
course. Results with ODN2-RGS2 and ODN2-RGS9 are shown in
Fig. 3. Impairment of RGS9 function brought about a leftward shift
of opioid dose±effect curves constructed at various postopioid
intervals, whereas de®cits of RGS2 proteins produced rightward
shifts (Figs 2 and 3).
The in¯uence of RGS2 and RGS9 on acute tolerance (tachyphy-
laxis) to morphine was also investigated. Morphine was used at
10 nmol/mouse in mice that had received ODN2 to RGS9, ODN2-
RGS9M and ODN2-RGS2M. Treatment with ODN2 to RGS2
produced marked decreases of morphine analgesic potency (Fig. 1).
Thus, in order to obtain an effect comparable to that produced by the
opioid in the other experimental groups, the dose of morphine was
raised to 30 nmol in ODN2-RGS2 mice. Mice received either saline
(control) or the opioid, and 24 h later the analgesia evoked by a second
injection of morphine was evaluated. In ODN2-RGS9M-treated mice,
antinociception reached 76 6 5% and 34 6 4% MPE (n = 12) in
control animals and in mice pretreated (±24 h) with the acute dose of
morphine. Morphine displayed no reduced activity when this assay
was performed in RGS9 knock down mice. Values of 72 6 5% and
74 6 4% MPE (n = 12) were recorded for controls and mice
pretreated with morphine, respectively (Fig. 4). In these mice, the
time course for morphine antinociception was not affected by the
previous dose of the opioid. The ODN-induced impairment of RGS2
proteins provided no protection against the acute tolerance evoked by
the single dose of 30 nmol morphine (Fig. 4).
To determine the in¯uence of RGS9 knock-down on the analgesic
effects evoked by daily injections of the opioid, the ODN treatment
was extended for 10 days. During the ®rst 5 days, treated mice
received the ODN2-RGS9 following the standard schedule, while
control animals received the ODN2-RGS9M. Afterwards, a daily
injection of 3 nmol ODN was given for 5 more days. The animals were
i.c.v.-injected with 10 nmol morphine on days 7, 8, 9 and 10. Control
mice exhibited a signi®cant decrease of morphine activity after the
second dose of the opioid given on day 8. No further decreases were
promoted by the morphine doses of days 9 and 10 (Fig. 5). RGS9
knock down mice exhibited normal responses to the consecutive doses
of morphine. Thus, acute tolerance to morphine as well as tolerance to
repeated injections of the opioid were greatly diminished in these
animals (Figs 4 and 5).
In the model of sustained morphine treatment, the impairment of
RGS9 proteins facilitated increases in the response to the delivered
opioid. This was only effective for 2±3 h after s.c.-implantation of the
morphine pellet. The opposite, a decrease in morphine activity, was
observed for RGS2 knock-down mice (Fig. 6A). In these mice, the
FIG. 6. In¯uence of RGS knock down on the development of tolerance tosustained chronic morphine. Animals received the ODNs into the rightlateral ventricle following a 5-day schedule. On day 6 the mice were s.c.-implanted at time zero with the oily suspension containing morphine.Development of tolerance was monitored by measuring the analgesiaproduced by the release of the s.c.-implanted opioid (A), and by a singlei.c.v. dose of 10 nmol morphine (B). This dose produced, in mice notpreviously exposed to the opioid, an effect of » 80% MPE in the tail-¯icktest. This is indicated in the ®gure with the dashed line. Values are themean 6 SEM from groups of 15±20 mice. *,+P < 0.05 vs. the group thathad received the mismatched control ODN-RGS9M or ODN-RGS2M,respectively, ANOVA, Student±Newman±Keuls test.
RGS9 facilitates opioid tachyphylaxis 807
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

increases or decreases in the potency of i.c.v. doses of morphine were
not evident 18±24 h after commencing chronic treatment with the
opioid (Fig. 6B). Thus, the impairment of RGS9 function did not bring
about long-term increases in the response of tolerant mice to i.c.v.
morphine.
Speci®c binding of 125I-Tyr27 human b-endorphin to mu-opioidreceptors in mouse PAG membranes
Membranes from mouse PAG were used in saturation assays of 125I-
Tyr27 human b-endorphin. Binding assays were performed in the
presence of 300 nM ICI-174,864 to prevent binding of labelled b-
endorphin to delta-opioid receptors. The data were analysed with the
LIGAND programme (Munson & Rodbard, 1980) and indicated a Kd
of » 0.3 nM and a binding capacity of » 50±60 fmol/mg of membrane
protein (present work; GarzoÂn et al., 1998). Thus, neither the af®nity of
the neuropeptide to mu-opioid receptors, nor its binding capacity,
changed due to treatment with ODNs directed towards RGS2 or RGS9
proteins (Fig. 7).
Discussion
The ODNs directed to RGS2 and RGS9 proteins effectively reduced
the expression of the respective proteins, but did not alter the
presence of other proteins required for signalling via the mu-opioid
receptors, i.e. Gi2a subunits, Gb1/2 subunits. Moreover, the number
of mu-opioid receptors and the af®nity displayed towards the agonists
were also unchanged. The use of ODNs revealed that RGS2 and
RGS9 proteins modulated opioid activity by a push±pull mechanism.
Signi®cant structural and functional differences exist between RGS9
and RGS2 which might account for RGS selectivity towards Gasubunits, and for the kinetics of their interaction. RGS6, 7, 9 and 11
contain G-protein g-subunit-like domains that mediate their associ-
ation with Gb5, but not with the other Gb subunits (Snow et al., 1998;
Hepler, 1999; Zhang & Simonds, 2000). Functionally active RGS9,
but not RGS2, forms a tight complex with the long spliced variant of
Gb5 (Makino et al., 1999). RGS2 is described as selectively binding
Gqa in brain membranes, but not other Ga subunits (Gi, Go, Gs,
G12/13), and completely inhibits Gq-directed activation of phospho-
lipase Cb1 (Heximer et al., 1997). This RGS2 is unique because it is
the only RGS protein that does not interact with Gi family members
and also because its expression is dynamically responsive to neuronal
activity (Ingi et al., 1998). It has been shown that RGS2 mRNA levels
increase in response to activation of different G-protein-coupled
receptors (Burchett et al., 1999; Song et al., 1999). In the rat, the
mRNA of RGS9-2 is common in striatum and hypothalamus but is
less frequent in other brain structures such as the neocortex (Gold
et al., 1997; Granneman et al., 1998). In this species, RGS9-1 mRNA
is abundant in retina but hardly detected in brain. The present results
from CD-1 mice indicate a greater presence of the long RGS9-2
protein in striatum than in cerebral cortex or PAG. The protein in
these structures could originat from the detected RGS9 mRNA and/or
be supplied by striatal or hippocampal projections (Saper, 1985;
Boylan et al., 1986; Hoover & Strick, 1993).
The classes of G-proteins regulated by opioid receptors in the
production of analgesia are acted upon by RGS9 and RGS2 proteins,
i.e. Gi, Gz and Gq proteins (GarzoÂn et al. 2000). In their opposite role
in opioid analgesia, RGS2 and RGS9 proteins might exhibit
differences in their binding to Ga-GTP and release from Ga-GDP
(rapid for RGS2 and slow for RGS9). By increasing the availability of
certain Gbg dimers, RGS proteins would also affect opioid-regulated
effects, e.g. Gbg direct binding to inwardly rectifying K+ channels
FIG. 7. Speci®c binding of 125I-Tyr27 human b-endorphin to mu-opioidreceptors in mouse PAG membranes. The Scatchard plots show theiodinated neuropetide's binding to PAG membranes from mice i.c.v.-injected with the ODNs to RGS2 or RGS9 proteins. Increasingconcentrations of 125I-Tyr27 human b-endorphin in the presence of 300 nM
ICI-174,864 (GarzoÂn et al., 1998) were incubated with approximately0.2 mg/mL ®nal protein concentration at 25 °C for 90 min. The Kd valueswere derived using the Ligand weighted nonlinear least squares regressionprogramme (Munson & Rodbard, 1980).
808 J. GarzoÂn et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

and increasing their probability of opening (Chuang et al., 1998; Han
et al., 1999). An augmented availability of regulated G-proteins, or
prolonged activity of Ga-GTP on the effectors, would account for the
extended time course of the agonist effects.
Reports in literature, and also this work, show that tachyphylaxis is
produced by a single dose of opioids (Huidobro et al., 1976; Vaught
& Takemori, 1979; Porreca et al., 1987). This suggests that acute
tolerance initiates when opioid agonists promote the activation and
segregation of Ga-GTP monomers from Gbg dimers. Agonist-
activated receptors then switch to a nonsignalling state (silent
receptors) (Fig. 8) until they couple once more with trimeric G-
proteins. Therefore, to re-establish an adequate number of active
receptors, Ga-GDP subunits need to bind Gbg to reconstitute the
trimeric G-proteins. However, the desired restoration of G-proteins is
inef®cient. The reconstitution of trimeric G-proteins will mostly
depend on the speed at which Ga-GTP is converted to Ga-GDP by
the RGS proteins (Berman & Gilman, 1998). These GAPs bind to
active Ga-GTP subunits, competitively antagonizing the productive
interaction between effector and Ga-GTP (Tesmer et al., 1997), and
promoting the Ga-GDP form capable of binding to Gbg dimers
(Berman & Gilman, 1998; Hepler, 1999). Structural studies indicate
that Gbg and RGS do not bind simultaneously to the same Ga-GDP
(Tesmer et al., 1997) therefore Ga-GDP subunits must be released
from RGS proteins before they reconstitute the trimeric G-proteins.
Although RGS proteins bind with high af®nity to the transition state
of Ga-GTP, they also bind with lower af®nity to Ga-GTP or Ga-
GDP forms (Hunt et al., 1996; Hepler et al., 1997). Consequently,
while RGS proteins remain bound to Ga subunits, Gbg-Ga±GDP
association is delayed (BuÈnemann & Hosey, 1998; Hepler, 1999).
RGS proteins exhibiting this negative regulation of G-protein
signalling might determine the rate of desensitization of G-receptors
(Neil et al., 1997; Chuang et al., 1998; Tseng & Zhang, 1998;
DiverseÂ-Pierluissi et al., 1999; Potenza et al., 1999).
The ®nding that RGS9 knock down animals did not exhibit
tachyphylaxis after a single dose of the agonists indicates that
phosphorylation (inactivation) of Ga subunits, probably by PKC
(Strassheim & Malbon, 1994), is not greatly responsible for acute
tolerance to opioids. The reduction of RGS2 function did not prevent
acute tolerance to morphine which effectively developed in RGS2
knock down mice. Several hours after commencing chronic morphine
FIG. 8. Diagrammatic model describing theassigned roles of RGS2 and RGS9 on G-protein reconstitution. RGS2 proteins, afterbinding to activated Ga-GTP subunits,promote the rapid formation and release ofGa-GDP, thus making re-constituted trimericGabg proteins available to receptor regulation.Binding of RGS9 proteins to Ga-GTP delaysthe release of the Ga-GDP form much longer,allowing the signals originating at agonist-activated receptors to decline. Depletion ofRGS2 proteins decreases opioid potency.However, impairment of RGS9 proteins bringsabout marked enhancements of opioid potencyand the duration of its effects.
RGS9 facilitates opioid tachyphylaxis 809
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

treatment, the bene®cial effects of RGS9 knock down disappeared
and tolerance occurred. To reduce the impact of the chronic morphine
acting on the opioid receptors, other RGS proteins presumably
substitute the GAP function of RGS9 upon activated Ga-GTP
subunits.
These ®ndings indicate that over the time course of an opioid effect
there is a progressive reduction in the number of receptor-regulated
G-proteins. The agonist-induced separation of activated Ga-GTP
subunits from Gbg dimers is faster than the reconstitution of the
trimeric Gabg proteins. Hence tachyphylaxis develops and helps to
control the intensity of exogenous signals that could threaten cell
survival. Acute tolerance to opioids then arises as a bene®cial
membrane-delimited process caused by the RGS-facilitated progres-
sive depletion of receptor-regulated G-proteins. During this time, the
receptors remain capable of regulating new G-proteins. However,
long-term tolerance that occurs after repeated doses of opioids is
mostly due to uncoupling of opioid receptors from the regulated
transduction.
Acknowledgements
This work was supported by funding from the `ComisioÂn Interministerial deCiencia y TecnologõÂa' (grant CICYT SAF98-0057) and the `ProgramaSectorial de PromocioÂn General del Conocimiento' (grant DGICYT PB98-0649). Marta RodrõÂguez-DõÂaz is a predoctoral fellow from CAM. AlmudenaLoÂpez-Fando is a predoctoral fellow from FPI (MEC).
Abbreviations
GAP, GTPase-activating protein; G-protein, guanine nucleotide-bindingregulatory protein; ICI-174,864, N,N-diallyl-Tyr-(a-aminoisobutyric acid)2-Phe-Leu-OH; MPE, maximum possible effect; ODN, antisense oligodeoxy-nucleotide; ODN-RD, random ODN; ODN-RGS#M, mismatched antisensesequence; PAG, periaqueductal grey matter; RGS, regulator of G-proteinsignalling protein; TTBS, TBS-0.05% Tween 20.
References
Berman, D.M. & Gilman, A.G. (1998) Mammalian RGS proteins: barbariansat the gate. J. Biol. Chem., 273, 1269±1272.
Bilsky, E.J., Bernstein, R.N., Wang, Z., SadeÂe, W. & Porreka, F. (1996)Effects of naloxone and D-Phe-Cys-Tyr-D-Trp-Arg-Thr-Pen-Thr-NH2 andthe protein kinase inhibitors H7 and H8 on acute morphine dependence andantinociceptive tolerance in mice. J. Pharmacol. Exp. Ther., 277, 484±490.
Boylan, M.K., Fisher, R.S., Hull, C.D., Buchwald, N.A. & Levine, M.S.(1986) Axonal branching of basal forebrain projections to the neocortex: adouble-labelling study in the cat. Brain Res., 375, 176±181.
BuÈnemann, M. & Hosey, M. (1998) Regulators of G protein signalling (RGS)proteins constitutively activate Gbg-gated potassium channels. J. Biol.Chem., 47, 31186±31190.
Burchett, S.A., Bannon, M.J. & Granneman, J.G. (1999) RGS mRNAexpression in rat striatum: modulation by dopamine receptors and effects ofrepeated amphetamine administration. J. Neurochem., 72, 1529±1533.
Chiasson, B.J., Armstrong, J.N., Hooper, M.L., Murphy, P.R. & Robertson,H.A. (1994) The application of antisense oligonucleotide technology to thebrain: some pitfalls. Cell. Mol. Neurobiol., 14, 507±521.
Chuang, H.H., Yu, M., January, Y.N. & January, L.Y. (1998) Evidence thatthe nucleotide exchange and hydrolysis cycle of G proteins causesdesensitization of G-protein gated inward recti®er K+ channels. Proc.Natl. Acad. Sci. USA, 95, 11727±11732.
DiverseÂ-Pierluissi, M.A., Fischer, T., Jordan, J.D., Schiff, M., Ortiz, D.F.,Farquhar, M.G. & De Vries, L. (1999) Regulators of G protein signallingproteins as determinants of the rate of desensitization of presynapticcalcium channels. J. Biol. Chem., 274, 14490±14494.
Fairbanks, C.A. & Wilcox, G.L. (1997) Acute tolerance to spinallyadministered morphine compares mechanistically with chronicallyinduced morphine tolerance. J. Pharmacol. Exp. Ther., 282, 1408±1417.
GarzoÂn, J., Castro, M. & SaÂnchez-BlaÂzquez, P. (1998) In¯uence of Gz and Gi2
transducer proteins in the af®nity of opioid agonists to m receptors. Eur. J.Neurosci., 10, 2557±2564.
GarzoÂn, J., DeAntonio, I. & SaÂnchez-BlaÂzquez, P. (2000) In vivo modulationof G-proteins and opioid receptor function by antisenseoligodeoxynucleotides. Meth. Enzymol., 314, 3±20.
Gold, S.J., Ni, Y.G., Dohlman, H.G. & Nestler, E.J. (1997) Regulators of G-protein signalling (RGS) proteins: region-speci®c expression of ninesubtypes in rat brain. J. Neurosci., 17, 8024±8037.
Granneman, J.G., Zhai, Y., Zhu, Z., Bannon, M.J., Burchett, S.A., Schmidt,C.J., Andrade, R. & Cooper, J. (1998) Molecular characterization of humanand rat RGS 9L, a novel splice variant enriched in dopamine target regions,and chromosomal localization of the RGS 9 gene. Mol. Pharmacol., 54,687±694.
Han, S.H., Cho, Y.W., Kim, C.J., Min, B.I., Rhee, J.S. & Akaike, N. (1999)Mu-opioid agonist-induced activation of G-protein-coupled inwardlyrectifying potassium current in rat periaqueductal gray neurons.Neuroscience, 90, 209±219.
Hepler, J.R. (1999) Emerging roles for RGS proteins in cell signalling. Trends.Pharmacol. Sci., 20, 376±382.
Hepler, J.R., Berman, D.M., Gilman, A.G. & Kozasa, T. (1997) RGS4 andGAIP are GTPase-activating proteins for Gq alpha and block activation ofphospholipase C beta by gamma-thio-GTP-Gq alpha. Proc. Natl. Acad. Sci.USA, 94, 428±432.
Heximer, S.P., Watson, N., Linder, M.E., Blumer, K.J. & Hepler, J.R. (1997)RGS2/GOS8 is a selective inhibitor of Gqa function. Proc. Natl. Acad. Sci.USA, 94, 14389±14393.
Hoover, J.E. & Strick, P.L. (1993) Multiple output channels in the basalganglia. Science, 259, 819±821.
Huidobro, F., Huidobro-Toro, J.P. & Way, E.L. (1976) Studies on tolerancedevelopment to single doses of morphine in mice. J. Pharmacol. Exp. Ther.,198, 318±326.
Huidobro-Toro, J.P. & Way, E.L. (1978) Single-dose tolerance toantinociception, and physical dependence on b-endorphin in mice. Eur. J.Pharmacol., 52, 179±189.
Hunt, T.W., Fields, T.A., Casey, P.J. & Peralta, E.G. (1996) RGS10 is aselective activator of G alpha i GTPase activity. Nature, 383, 175±177.
Ingi, T., Krumins, A.M., Chidiac, P., Brothers, G.M., Chung, S., Snow, B.E.,Barnes, C.A., Lanahan, A.A., Siderovski, D.P., Ross, E.M., Gilman, A.G. &Worley, P.F. (1998) Dynamic regulation of RGS2 suggests a novelmechanism in G-protein signalling and neuronal plasticity. J. Neurosci., 18,7178±7188.
Lang, J. & Costa, T. (1987) Antisera against the 3±17 sequence of rat Gairecognize only a 40 kDa G-protein in brain. Biochem. Biophys. Res.Commun., 148, 838±848.
Makino, E.R., Handy, J.W., Li, T. & Arshavsky, V.Y. (1999) The GTPaseactivating factor for transducin in rod photoreceptors is the complexbetween RGS9 and type 5 G protein b subunit. Proc. Natl. Acad. Sci. USA,96, 1947±1952.
Munson, P.J. & Rodbard, D. (1980) LIGAND: a versatile computerizedapproach for characterization of ligand-binding systems. Anal. Biochem.,107, 220±239.
Narita, M., Makimura, M., Feng, Y., Hoskins, B. & Ho, I.K. (1994) In¯uenceof chronic morphine treatment on protein kinase C activity: comparisonwith butorphanol and implication for opioid tolerance. Brain Res., 650,175±179.
Neil, J.D., Wayne-Duck, L., Sellers, J.C., Musgrove, L.C., Scheschonka, A.,Druey, K.M. & Kehrl, J.H. (1997) Potential role for a RGS3 in GnRHstimulated desensitization. Endocrinology, 138, 843±846.
Nestler, E.J. & Aghajanian, G.K. (1997) Molecular and cellular basis ofaddiction. Science, 278, 58±63.
Nestler, E.J., Hope, B.T. & Widnell, K.L. (1993) Chronic morphine treatmentincreases cyclic AMP-dependent protein kinase activity in the rat locuscoeruleus. Mol. Pharmacol., 33, 127±132.
Porreca, F., Heyman, J.S., Mosberg, H.I., Omnaas, J.R. & Vaught, J.L. (1987)Role of mu and delta receptors in the supraspinal and spinal analgesiceffects of [D-Pen2, D-Pen5] enkephalin in the mouse. J. Pharmacol. Exp.Ther., 241, 393±400.
Potenza, M.N., Gold, S.J., Roby-Shemkowitz, A., Lerner, M.R. & Nestler, E.J.(1999) Effects of regulators of G-protein-signalling proteins on thefunctional response of the m-opioid receptor in a melanophore-basedassay. J. Pharmacol. Exp. Ther., 291, 482±491.
Rahman, Z., Gold, S.J., Potenza, M.N., Cowan, C.W., Ni, Y.G., He, W.,Wensel, T.G. & Nestler, E.J. (1999) Cloning and characterization of RGS9-2: a striatal-enriched alternatively spliced product of the RGS9 gene. J.Neurosci., 19, 2016±2026.
810 J. GarzoÂn et al.
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811

Rossi, G.C., Pasternak, G.W. & Bodnar, R.J. (1994) m and d opioid synergybetween the periaqueductal gray and the rostroventral medulla. Brain Res.,665, 85±93.
SaÂnchez-BlaÂzquez, P., GarcõÂa-EspanÄa, A. & GarzoÂn, J. (1995) In vivoinjection of oligodeoxynucleotides to Ga subunits and supraspinal analgesiaevoked by mu and delta opioid agonists. J. Pharmacol. Exp. Ther., 275,1590±1596.
SaÂnchez-BlaÂzquez, P., GarcõÂa-EspanÄa, A. & GarzoÂn, J. (1997) Antisenseoligodeoxynucleotides to opioid Mu and DELTA receptors reducedmorphine dependence in mice: role of DELTA2 opioid receptors. J.Pharmacol. Exp. Ther., 280, 1423±1431.
Saper, C.B. (1985) Organization of cerebral cortical afferent systems in the rat.II. Hypothalamocortical projections. J. Comp Neurol., 237, 21±46.
Snow, B.E., Krumins, A.M., Brothers, G.M., Lee, S.-F., Wall, M.A., Chung,S., Mangion, J., Arya, S., Gilman, A.G. & Siderovski, D.P. (1998) A Gprotein g subunit-like domain shared between RGS11 and other RGSproteins speci®es binding to Gb5 subunits. Proc. Natl. Acad. Sci. USA, 95,13307±13312.
Song, L., De Sarno, P. & Jope, R.S. (1999) Muscarinic receptor stimulationincreases regulators of G-protein signalling 2 mRNA levels through a
protein kinase C-dependent mechanism. J. Biol. Chem., 274,29689±29693.
Strassheim, D. & Malbon, C.C. (1994) Phosphorylation of Gia2 attenuatesinhibitory adenylyl cyclase in neuroblastoma/glioma hybrid (NG-108±15)cells. J. Biol. Chem., 269, 14307±11413.
Tesmer, J.J., Berman, D.M., Gilman, A.G. & Sprang, S.R. (1997) Structure ofRGS4 bound to AlF4-activated G (i alpha 1): stabilization of the transitionstate for GTP hydrolysis. Cell, 89, 251±261.
Tseng, C.C. & Zhang, X.-Y. (1998) Role of regulator of G protein signallingin desensitization of the glucose-dependent insulinotropic peptide receptor.Endocrinology, 139, 4470±4475.
Vaught, J.L. & Takemori, A.E. (1979) Differential effects of leucine andmethionine enkephalin on morphine-induced analgesia, acute tolerance anddependence. J. Pharmacol. Exp. Ther., 208, 86±90.
Yaksh, T.L., Yeung, J.C. & Rudy, T.A. (1976) Systematic examination in therat of brain sites sensitive to the direct application of morphine: observationof differential effects within the periaqueductal gray. Brain Res., 114, 83±103.
Zhang, J.-H. & Simonds, W.F. (2000) Copuri®cation of brain G-protein b5with RGS6 and RGS7. J. Neurosci., 20, 1±5.
RGS9 facilitates opioid tachyphylaxis 811
ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 13, 801±811
Top Related
Copyright © 2022 FDOKUMEN

